diplomarbeit andrea fuchs - welcome to...
TRANSCRIPT

Carl von Ossietzky
Universität Oldenburg
Diplomstudiengang Biologie
DIPLOMARBEIT
Wirkung des Ballastwasser-Biozids Acrolein auf verschiedene pathogene
Bakterienstämme in Meer-, Brack- und Süßwasser
vorgelegt von: Andrea Fuchs
Betreuender Gutachter: Dr. Gunnar Gerdts
Zweiter Gutachter: Prof. Dr. Meinhard Simon
Oldenburg, den 19.12.2008

Inhaltsverzeichnis
Inhaltsverzeichnis
Abbildungsverzeichnis.................................................................................................... 3
Tabellenverzeichnis......................................................................................................... 6
Zusammenfassung........................................................................................................... 7
1 Einleitung ................................................................................................................... 9
1.1 Allgemeines...................................................................................................... 9
1.2 Ballastwasser-Behandlung ............................................................................. 10
1.3 Bedeutung der untersuchten pathogenen Mikroorganismen .......................... 11
1.4 Zielsetzung ..................................................................................................... 14
2 Material und Methoden .......................................................................................... 15
2.1 Bakterienstämme ................................................................................................. 15
2.1.1 Kultivierung............................................................................................ 15
2.1.2 Wachstumskurven................................................................................... 16
2.1.3 Einstellung der Optischen Dichte (OD).................................................. 16
2.2 Flüssigkeiten und Lösungen........................................................................... 16
2.2.1 TTC (2,3,5-Triphenyltetrazoliumchlorid) .............................................. 16
2.2.2 Acrolein C3H4O ...................................................................................... 18
2.2.3 Probenwasser .......................................................................................... 19
2.3 Versuchsaufbau .............................................................................................. 19
2.3.1 ScanMIC-Methode .................................................................................19
2.3.2 Acrolein-Versuch.................................................................................... 21
2.4 Material .......................................................................................................... 23
2.4.1 Technische Geräte .................................................................................. 23
2.4.2 Verbrauchsmaterial................................................................................. 24
2.4.3 Chemikalien............................................................................................ 25
2.4.4 Hergestellte Medien und Lösungen ........................................................ 25
3 Ergebnisse................................................................................................................. 27
3.1 Phänotyp der Bakterienstämme......................................................................27
3.1.1 Kultivierung............................................................................................ 27
3.1.2 ScanMIC-Mikrotiterplatten .................................................................... 28
3.2 Wachstum der untersuchten Isolate................................................................ 30

Inhaltsverzeichnis
3.2.1 Einteilung in Gruppen............................................................................. 30
3.2.2 Wachstum in Medium............................................................................. 31
3.2.3 Wachstum der Isolate in Meer-, Brack- und Süßwasser......................... 33
3.3 Wachstum der Isolate mit Acrolein................................................................ 35
3.3.1 Wirkung der Acrolein-Konzentrationen 0.01 und 0.1 mg/L auf die
untersuchten Isolate ................................................................................ 37
3.3.2 Wirkung der Acrolein-Konzentration 1 mg/L auf die untersuchten Isolate
................................................................................................................ 39
3.3.3 Acrolein-Konzentrationen 5, 10 und 15 mg/L........................................ 45
4 Diskussion................................................................................................................. 50
4.1 Bakterienstämme............................................................................................ 50
4.1.1 Wachstum der Isolate in Medium ........................................................... 51
4.1.2 Probenwasser .......................................................................................... 54
4.1.3 Einteilung der untersuchten Bakterienstämme in Gruppen und Wachstum
der Isolate in Meer-, Brack- und Süßwasser........................................... 54
4.2 Wachstum mit verschiedenen Acrolein-Konzentrationen .............................. 57
4.2.1 Acrolein-Konzentration 0.01 und 0.1 mg/L ........................................... 57
4.2.2 Wirkung der Acrolein-Konzentrationen 1-15 mg/L auf die untersuchten
Isolate...................................................................................................... 58
4.3 Vergleich mit anderen Bioziden die in der Ballastwasser-Behandlung
eingesetzt werden........................................................................................... 61
4.4 Acrolein als Biozid für Ballastwasserbehandlung.......................................... 65
4.5 Methoden-Kritik ............................................................................................. 66
5 Ausblick .................................................................................................................... 69
Literaturliste.................................................................................................................. 70
Anhang.......................................................................................................................... 76
Abkürzungsverzeichnis................................................................................................. 86
Danksagung................................................................................................................... 88
Erklärung....................................................................................................................... 90

Abbildungsverzeichnis
Abbildungsverzeichnis
Abbildung 1 A und B: A zeigt die Strukturformeln und Reduktion von 2,3,5-
Triphenyltetrazoliumchlorid zu Formazan, B zeigt eine Mikrotiterplatte
mit Bakteriensuspension und TTC vor (links) und nach (rechts) 24 h
Inkubation……………………………………………………………......13
Abbildung 2: Strukturformel und Gefahrenklassen „leichtentzündlich“, „giftig“ und
„umweltgefährlich“ von Acrolein............................................................. 18
Abbildung 3: Schema zum Versuchsaufbau. ................................................................. 21
Abbildung 4: Inokulation der Mikrotiterplatten aus den Probenflaschen. Jeweils drei
Kavitäten werden aus einer Probenflasche inokuliert. Pro sieben Flaschen
werden zwei Mikrotiterplatten belegt. Die Zahlen auf den Probenflaschen
und über den Mikrotiterplatten geben die verwendeten Acrolein-
Konzentrationen an, die pro Flasche in je drei Kavitäten überimpft
wurden. ..................................................................................................... 22
Abbildung 5 a-f: Kolonien der kultivierten Bakterien auf Agarplatten.
a) Escherichia coli DSM 30083, b) Enterococcus faecalis DSM 20478,
c) Pseudomonas aeruginosa DSM 50071, d) Serratia liquefaciens
DSM 4478, e) Vibrio parahaemolyticus DSM 2172, f) Umweltisolat dB2
.................................................................................................................. 27
Abbildung 6 a-b: Koloniebildung von V. vulnificus auf Agarplatten. a) V. vulnificus
DSM 10143, b) V. vulnificus DSM 11507................................................ 28
Abbildung 7 a-f: Färbung der Kavitäten in den Mikrotiterplatten. a) E. coli,
b) E. faecalis, c) P. aeruginosa, d) S. liquefaciens, e) V. parahaemolyticus,
f) Umweltisolat dB2 .................................................................................. 29
Abbildung 8 a-b: Färbung der beiden V. vulnificus-Stämme in Mikrotiterplatten.
a) V. vulnificus DSM 10143, b) V. vulnificus DSM 11507....................... 30
Abbildung 9: Wachstumskurven der pathogenen Bakterienstämme bei 30° C ............. 31
Abbildung 10: Wachstum des Umweltisolates d B2 bei 18°C und bei 30°C in ZoBell-
Medium..................................................................................................... 32

Abbildungsverzeichnis
Abbildung 11 a-c: Zellzahlen der untersuchten Bakterien ohne Acroleinzugabe von
Tag 0 bis Tag 5 im Probenwasser. Die Zellzahlen entsprechen den
Mittelwerten aus jeder Gruppe. a) Meerwasser, b) Brackwasser,
c) Süßwasser ............................................................................................. 33
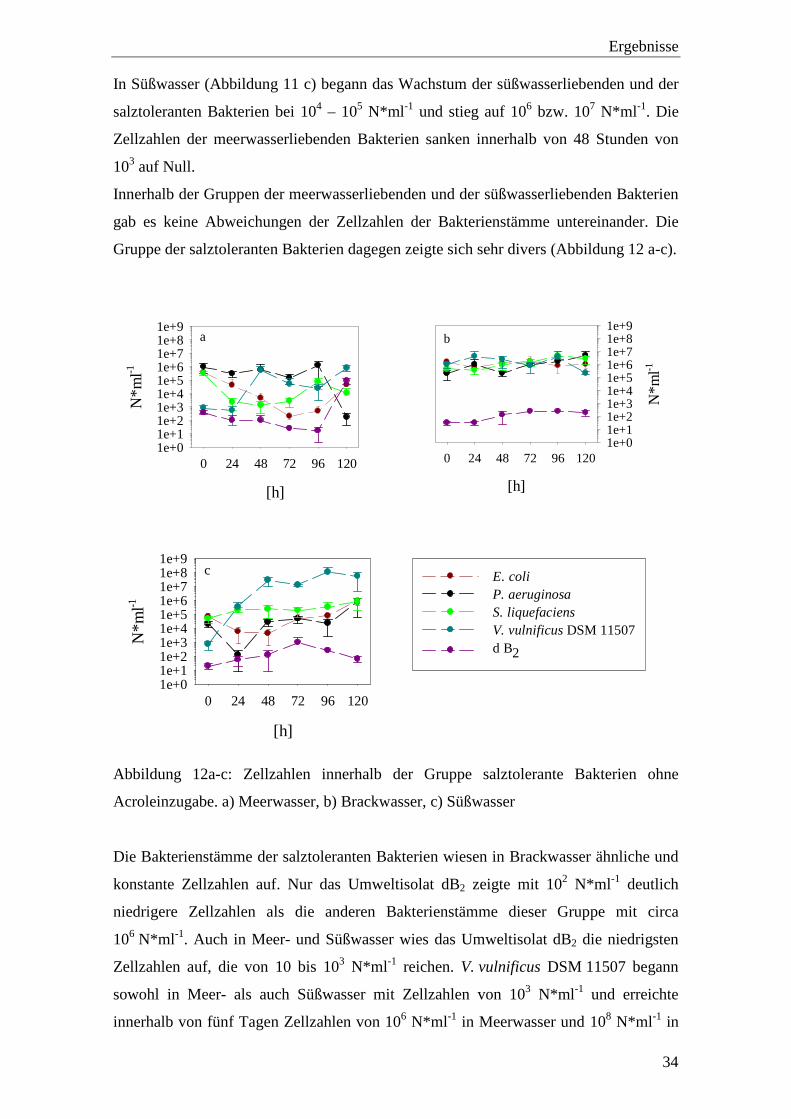
Abbildung 12a-c: Zellzahlen innerhalb der Gruppe salztolerante Bakterien ohne
Acroleinzugabe. a) Meerwasser, b) Brackwasser, c) Süßwasser.............. 34
Abbildung 13: Wachstum der zwei Stämme V. vulnificus ohne Acrolein-Zugabe über
fünf Tage in Meer-, Brack- und Süßwasser. a) V. vulnificus DSM 10143,
b) V. vulnificus DSM 11507 ..................................................................... 35
Abbildung 14 a-c: Wirkung niedriger Acrolein-Konzentrationen nach fünf Tagen auf
die Zellzahlen der untersuchten Bakterien. a) Meerwasser,
b) Brackwasser, c) Süßwasser .................................................................. 37
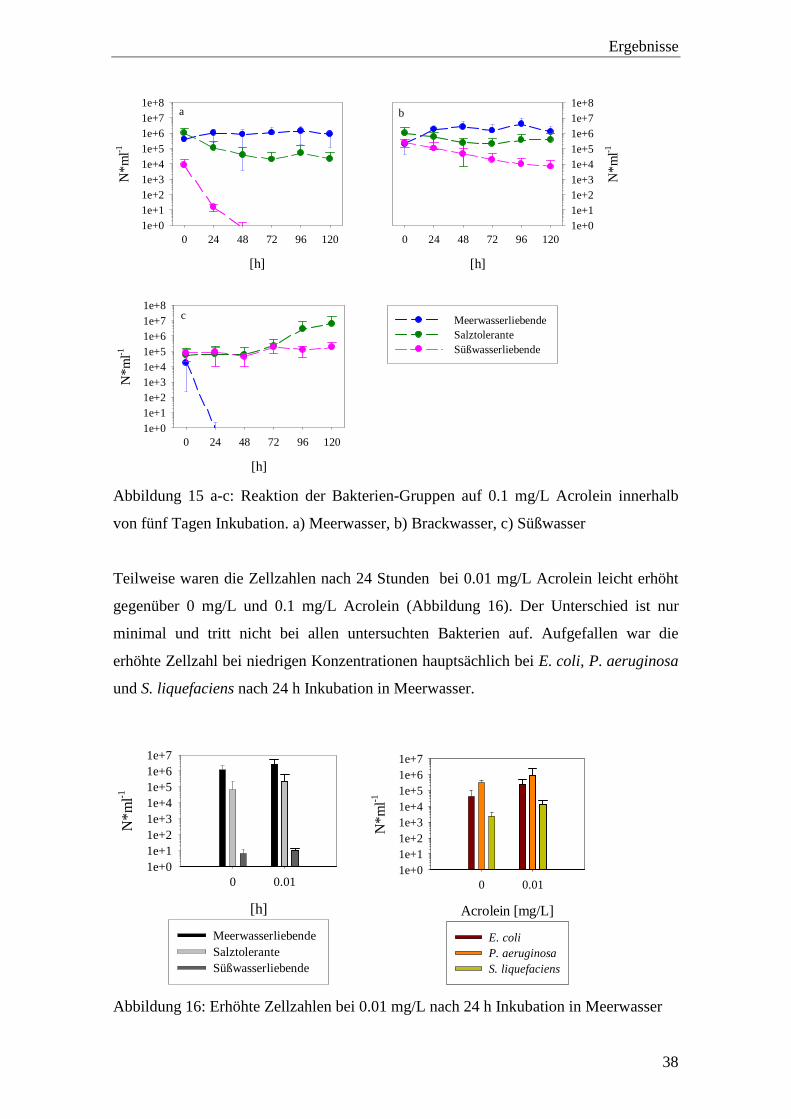
Abbildung 15 a-c: Reaktion der Bakterien-Gruppen auf 0.1 mg/L Acrolein innerhalb
von fünf Tagen Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser
.................................................................................................................. 38
Abbildung 16: Erhöhte Zellzahlen bei 0.01 mg/L nach 24 h Inkubation in Meerwasser
.................................................................................................................. 38
Abbildung 17: V. vulnificus DSM 10143 (a) und V. vulnificus DSM 11507 (b) mit
0.1 mg/L Acrolein.....................................................................................39
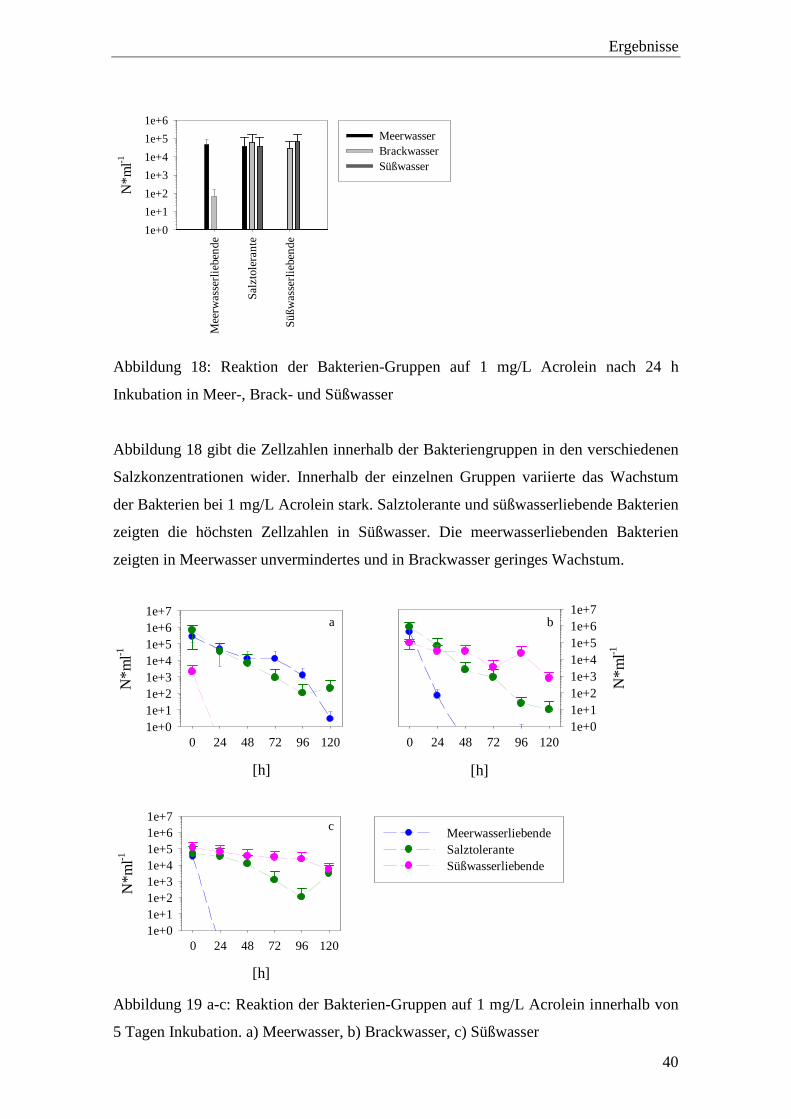
Abbildung 18: Reaktion der Bakterien-Gruppen auf 1 mg/L Acrolein nach 24 h
Inkubation in Meer-, Brack- und Süßwasser ............................................ 40
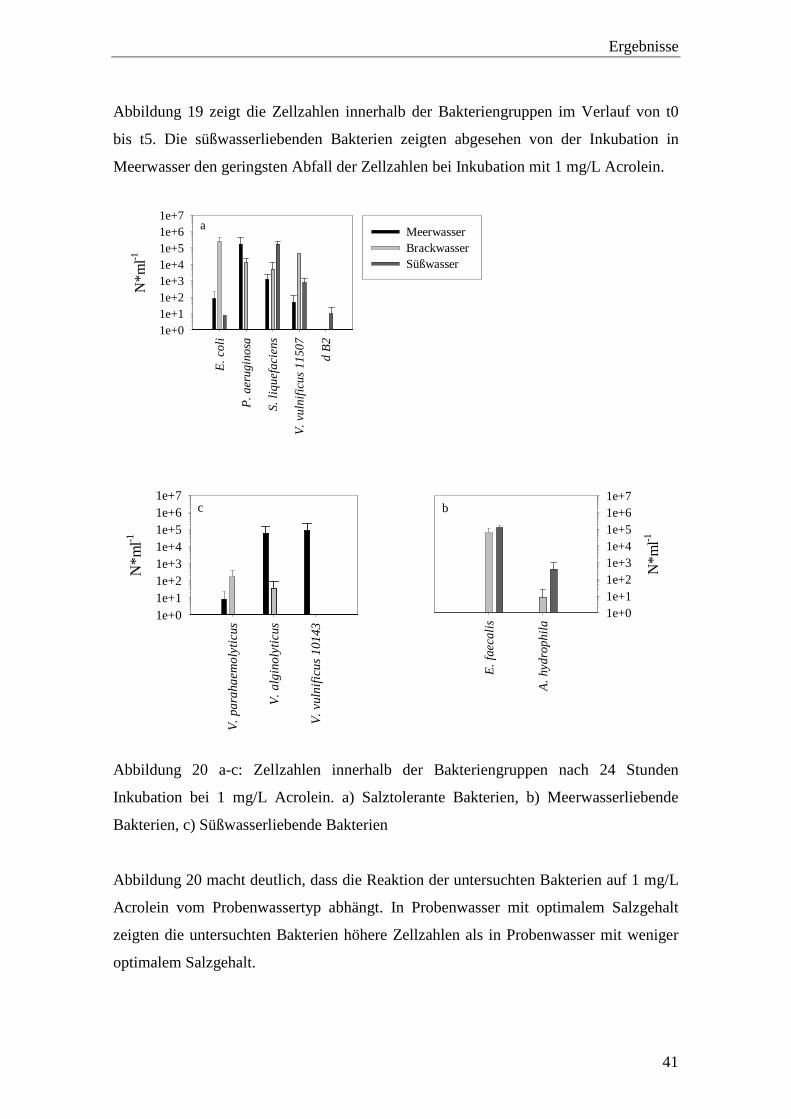
Abbildung 19 a-c: Reaktion der Bakterien-Gruppen auf 1 mg/L Acrolein innerhalb von
5 Tagen Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser........ 40
Abbildung 20 a-c: Zellzahlen innerhalb der Bakteriengruppen nach 24 Stunden
Inkubation bei 1 mg/L Acrolein. a) Salztolerante Bakterien, b)
Meerwasserliebende Bakterien, c) Süßwasserliebende Bakterien……….41
Abbildung 22 a-c: Wachstum der meerwasserliebenden Bakterien bei 1 mg/L Acrolein
von t0 bis t5. a) Meerwasser, b) Brackwasser, c) Süßwasser ................... 43
Abbildung 23 a-c: Wachstum der süßwasserliebenden Bakterien bei 1 mg/L Acrolein
von t0 bis t5. a) Meerwasser, b) Brackwasser, c) Süßwasser ................... 44

Abbildungsverzeichnis
Abbildung 24: Vergleich der Zellzahlen der beiden Stämme V. vulnificus untereinander
nach drei Tagen Inkubation bei 0.1, 1 und 5 mg/L Acrolein.................... 44
Abbildung 25 a-c: Zellzahlen der Bakteriengruppen bei hohen Acrolein-Konzentration
nach 24 h Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser ..... 45
Abbildung 26 a-c: Wirkung von 5 mg/L Acrolein auf die Bakteriengruppen.
a) Salztolerante Bakterien, b) Meerwasserliebende Bakterien,
c) Süßwasserliebende Bakterien ............................................................... 46
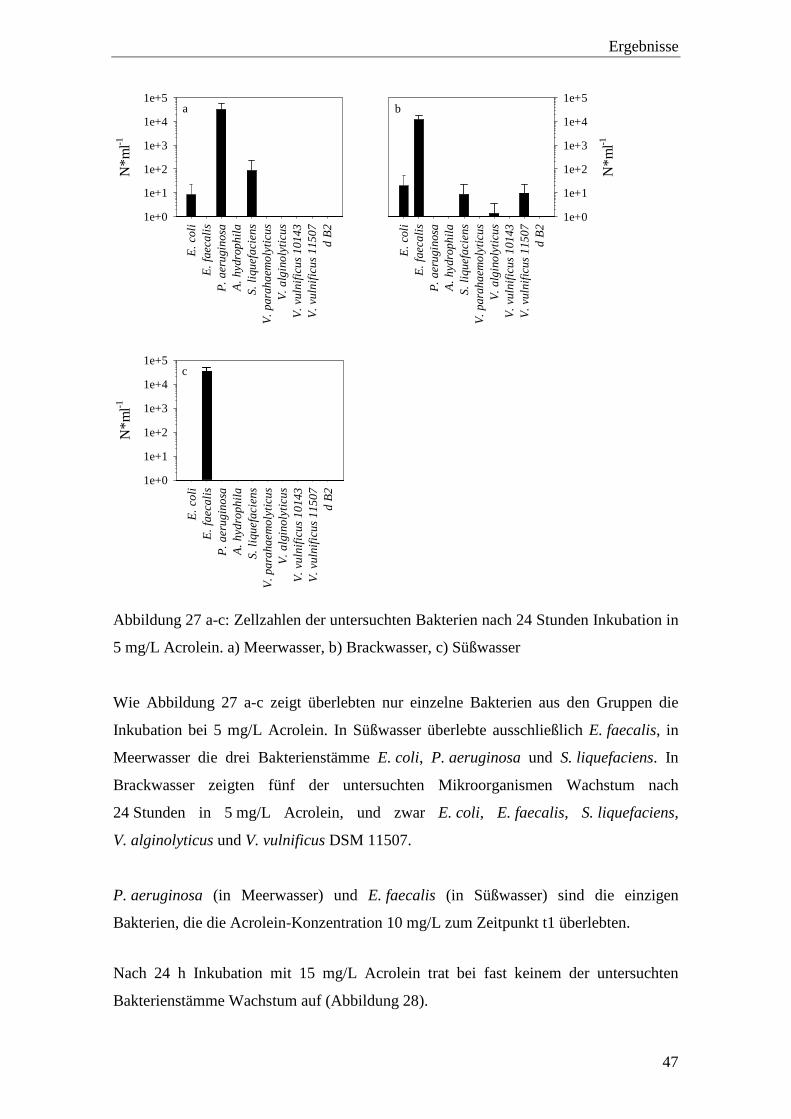
Abbildung 27 a-c: Zellzahlen der untersuchten Bakterien nach 24 Stunden Inkubation in
5 mg/L Acrolein. a) Meerwasser, b) Brackwasser, c) Süßwasser ............ 47
Abbildung 28: Wirkung von 15 mg/L Acrolein auf die Bakteriengruppen innerhalb von
5 Tagen. .................................................................................................... 48
Abbildung 29: P. aeruginosa nach 24 h in Meerwasser zeigt Wachstum bei allen
untersuchten Acrolein-Konzentrationen ................................................... 48
Abbildung 30: Minimale Acrolein-Konzentrationen, die nach 24 Stunden zur Abtötung
der untersuchten Bakterien führten........................................................... 49
Abbildung 31: Minimale Acrolein-Konzentrationen, die nach fünf Tagen zur Abtötung
der untersuchten Bakterien führten........................................................... 49
Abbildung 32: Minimale Konzentration an Bioziden, um das Wachstum von E. coli und
V. alginolyticus nach 48 h zu verhindern.................................................. 62
Abbildung 33: Mindestkonzentrationen an Bioziden, um das Wachstum von E. coli
nach 48 h Inkubation in Seewasser zu verhindern.................................... 63
Abbildung 34: Minimal notwendige Konzentrationen an Biozid, um das Wachstum
verschiedener Vibrionen zu verhindern.................................................... 64
Abbildung 35: Beispiel für eine belegte Mikrotiterplatte mit unterschiedlicher Anzahl
gefärbter Kavitäten ...................................................................................67

Tabellenverzeichnis
Tabellenverzeichnis
Tabelle 1: Einige Symptome, die die untersuchten pathogenen Bakterien beim
Menschen auslösen können (Dworkin et al, 2006) ....................................... 13
Tabelle 2: Auswirkung pathogener Bakterien auf See- und Süßwasserfische (Austin, B.
& Austin D., 1999) ........................................................................................ 13
Tabelle 3: Kultivierung der untersuchten Bakterien ...................................................... 15
Tabelle 4: Toxische Wirkung von Acrolein (Ghilarducci & Tjeerdema, 1995; Turner&
Erickson, 2003; Louder & McCoy, 1962). C3H4O (test) = Acrolein-
Konzentrationen, die in dieser Arbeit untersucht wurden, C3H4O (Literatur) =
Acrolein-Konzentrationen mit bekannter Wirkung....................................... 18
Tabelle 5: Salzgehalt und Probenahmeort für Meer-, Brack- und Süßwasser ............... 19
Tabelle 6: Darstellung von je einem Isolat pro Gruppe nach 24 h Inkubation in Meer-,
Brack- und Süßwasser. Angaben in N*ml-1. ................................................. 36
Tabelle 7: Beweglichkeit, Koloniemorphologie und Pelletbildung mit TTC-Färbung bei
den untersuchten Isolaten ........................Fehler! Textmarke nicht definiert.
Tabelle 8: Biozid-Mindestkonzentration nach 48 h Inkubation der Bakterien in
filtriertem Seewasser (25°C), bei der nach 24 h Inkubation in Medium (37°C)
kein Wachstum mehr auftritt (Gregg & Hallegraeff, 2007)........................... 61
Tabelle 9: Zellzahlen nach MPN-Tabelle (McCrady, 1915) ......................................... 67
Tabelle 10: Mittelwerte und Standardabweichungen auf der Basis von Zellzahlen aus
3er-Parallelen gefärbter Kavitäten in Mikrotiterplatten ................................ 68
Tabelle A 1: Zusammenfassung der gewonnenen Zellzahlen der Isolate aus den
Mikrotiterplatten-Versuchen mit und ohne Acrolein von t0-t5……………..76

Zusammenfassung
7
Zusammenfassung
Diese Diplomarbeit beschäftigt sich mit der Frage, welche Wirkung das Biozid
Acrolein auf verschiedene pathogene Bakterien hat. Vor dem Hintergrund der IMO-
Richtlinie 125 [53] wurde im Ballastwasser-Übereinkommen von 2004 fest gelegt, dass
Schiffe ihr Ballastwasser vorbehandeln müssen, bevor es ausgepumpt werden darf.
Diese Anordnung beruht auf der Tatsache, dass durch Ballastwasser jedes Jahr
Millionen von Organismen verschleppt werden und die heimische Flora und Fauna der
Zielhäfen aus dem Gleichgewicht bringen.
Eine Möglichkeit der Ballastwasser-Behandlung ist das Abtöten von Organismen durch
Biozide. Acrolein ist ein aquatisches Biozid, das bereits für die Behandlung von
Ballastwasser in Erwägung gezogen wurde (Baker Petrolite, 2001). Die hohe
Reaktivität von Acrolein birgt den Vorteil, nur geringe Mengen zur Behandlung
einsetzen zu müssen. In dieser Arbeit wurde die Wirkung der sechs Acrolein-
Konzentrationen 0.01, 0.1, 1, 5, 10 und 15 mg/L auf zehn verschiedene
Bakterienstämme untersucht. Es wurden neun pathogene Bakterienstämme untersucht,
der zehnte Bakterienstamm wurde aus einer Ballastwasser-Behandlungsanlage isoliert.
Zu den pathogenen Bakterien gehören Enterococcus faecalis, Escherichia coli, Serratia
liquefaciens, Pseudomonas aeruginosa, Aeromonas hydrophila, Vibrio
parahaemolyticus, Vibrio alginolyticus und zwei Stämme Vibrio vulnificus.
Basierend auf der ScanMIC-Methode (Rahman et al, 2003) wurden Mikrotiterplatten
mit Bakterienflüssigkultur, Acrolein und Probenwasser unterschiedlichen Salzgehaltes
belegt, mit dem Farbstoff TTC inkubiert und anschließend mit einem Flachbett-Scanner
eingescant. Die Auswertung erfolgte visuell und mit Hilfe der MPN-Tabelle nach
McCrady (1915).
Die visuelle Auswertung der untersuchten TTC-gefärbten Isolate in den
Mikrotiterplatten zeigte vielfältige morphologische Unterschiede auf. Der
Zusammenhang zwischen Beweglichkeit und TTC-Phänotyp eines Isolats ist nicht
eindeutig.
Die Isolate reagierten mit sehr unterschiedlichem Wachstum auf die Inkubation in
unterschiedlichen Salzkonzentrationen. Sie wurden daraufhin in die Gruppen
„salztolerante Bakterien“, „meerwasserliebende Bakterien“ und „süßwasserliebende
Bakterien“ eingeteilt.

Zusammenfassung
8
Die Behandlung mit Acrolein ergab, dass mindestens 5 mg/L Acrolein erforderlich sind,
um innerhalb von fünf Tagen Inkubation alle untersuchten Bakterienstämme in allen
untersuchten Salzkonzentrationen abzutöten. Innerhalb von 24 Stunden konnten
vereinzelt sogar hohe Acrolein-Konzentrationen überlebt werden. 1 mg/L Acrolein ist
eine Grenzkonzentration, die bei einigen Isolaten keine Reaktion hervor rief und bei
anderen Isolaten innerhalb von fünf Tagen zur Abtötung führte. Der Salzgehalt des
Probenwassers spielte dabei eine wichtige Rolle. Die Reaktion der Isolate auf niedrige
Acrolein-Konzentrationen unterschied sich je nach Salzgehalt des Probenwassers.
Unter den untersuchten Isolaten überlebten P. aeruginosa und E. faecalis die höchsten
Acrolein-Konzentrationen über die längste Zeit. Das Umweltisolat,
V. parahaemolyticus, V. alginolyticus und A. hydrophila reagierten am empfindlichsten
auf die Behandlung mit Acrolein.

Einleitung
9
1 Einleitung
1.1 Allgemeines
Schiffsverkehr gewinnt für den Welthandel zunehmend an Bedeutung. Etwa 90% aller
Waren werden inzwischen über Schiffe transportiert. Infolgedessen nehmen auch die
Invasionen aquatischer, an Schiffe assoziierter Organismen in neue Gebiete zu.
Es gibt natürliche Verbreitungsbarrieren, wie Landbarrieren, Salzgehalt und
Temperaturunterschiede, die die Verbreitung von Organismen erschweren. Vom
Mensch werden diese Barrieren durchbrochen. Durch Schiffsverkehr, die Öffnung von
Kanälen, die Verbreiterung von Flüssen und durch den Import für Aqua- und
Aquarienkultur kommt es zur Überschreitung natürlicher Grenzen und zur
Verschleppung von Organismen in zuvor unzugängliche Gebiete (Lenz et al, 1996).
Die zwei wesentlichen anthropogenen Vektoren für die Verschleppung von Arten sind
Aquakultur und Schiffsverkehr. Der Schiffsverkehr bietet mehrere Möglichkeiten zur
Verbreitung von Organismen, z.B. den Bewuchs der Außenhaut oder die Aufnahme von
Ballast. Schon seit Jahrhunderten wird Ballast aufgenommen, um für eine größere
Stabilität von Schiffen zu sorgen. Als Ballast wurde früher Sand und Kies geladen, seit
der Entwicklung von Schiffen aus Stahl vor etwa 140 Jahren verwendet man
Ballastwasser (Lenz et al, 1996; Reise et al, 1999). Pro Jahr werden etwa 12 Milliarden
Tonnen Ballastwasser weltweit transportiert. Ballastwasser ist für circa 25% der
Invasion von Arten verantwortlich (Bax et al, 2003). Speziell an der deutschen
Nordseeküste werden 13,4 Mio. Tonnen Ballastwasser im Jahr gelenzt, und darüber 2,7
Mio. Organismen am Tag aus außereuropäischen Regionen frei gesetzt (Nehring et al,
2004; Lenz et al, 1996)
Die Bedrohung, die von eingeschleppten Arten ausgeht, ist in der Bevölkerung
weitestgehend unbekannt. Über Ballastwasser werden zahlreiche Organismen
verschleppt, u.a. Bakterien, Algen, Zooplankton, Fische und die Eier, Sporen, Samen,
Cysten und Larven aquatischer Pflanzen und Tiere (Faimali et al, 2006). Die meisten
verschleppten Organismen können sich nicht in der neuen Umgebung etablieren, und
von den etablierten Organismen sind die meisten harmlos und nicht invasiv (Bax et al,

Einleitung
10
2003). Als „invasive Arten“ definierten Kolar & Lodge (2001) nicht-indigene Arten, die
sich nach ihrer Einführung ausbreiten und auch abundant werden. Die wenigen
invasiven Organismen die eingeschleppt werden reichen aber aus, massive Schäden zu
verursachen. Sie können heimische Organismen verdrängen, dadurch die Nahrungskette
stören, Habitate verändern und zum Zusammenbruch heimischer Fisch- und
Muschelpopulationen führen. Die Rippenqualle Mnepiopsis leidyi und die Chinesische
Wollhandkrabbe Eriocheir sinensis sind bereits dafür bekannt, dass sie im Schwarzen
Meer bzw. in Europa und an der US-Westküste zum Kollaps der Küstenfischerei
führen. Invasive Mikroorganismen, wie beispielsweise Cholera-Erreger und toxische
Dinoflagellaten, können zu gesundheitlichen Schäden beim Menschen führen (Drake et
al, 2006; Bax et al, 2003).
1.2 Ballastwasser-Behandlung
Um die Verschleppung von Organismen über Ballastwasser einzuschränken wurde
2004 von der International Maritime Organisation (IMO) ein Übereinkommen
verabschiedet das vorsieht, dass Ballastwasser vor der Abgabe vorbehandelt werden
muss. Es müssen sowohl Tests an Bord eines Schiffs als auch Tests an Land
durchgeführt werden. In diesen Tests muss das Ballastwasser aufgenommen, gelagert,
behandelt und wieder abgegeben werden. Bei den Tests an Land darf das Ballastwasser
nur über Pumpen aufgenommen und abgegeben werden und die Lagerung des
Ballastwassers muss mindestens über fünf Tage erfolgen. Die Tests sollen in den
Salinitätsbereichen > 32 PSU, 3-32 PSU und < 3 PSU durchgeführt werden (MEPC 125
[53], 2004).
Es gibt bisher verschiedene Methoden der Ballastwasserbehandlung. Man unterscheidet
zwischen mechanischen, physikalischen und chemischen Methoden. Oft werden
Kombinationen aus mechanischen mit physikalischen oder chemischen Behandlungen
verwendet. Beispielsweise trennen Filter oder Hydrozyklone das Ballastwasser
mechanisch von Partikeln, anschließend erfolgt eine Behandlung mit Hitze, UV-
Bestrahlung oder Chemikalien, um im Wasser enthaltene Organismen und
Mikroorganismen abzutöten. Geeignete Chemikalien sind z. B. Chlor, Chlorderivate,
Biozide und Ozon (Hochhaus & Mehrkens, 2007). Es gibt bereits mehrere anorganische
und organische Biozide, die für eine Behandlung von Ballastwasser untersucht wurden.

Einleitung
11
Als Ballastwasser-Management-System vom Bundesamt für Seeschifffahrt und
Hydrographie (BSH) zugelassen wurde bisher nur SEDNA®, unter Verwendung der
aktiven Substanz PERACLEAN® Ocean (www.bsh.de).
In dieser Arbeit wird die Wirkung von Acrolein auf pathogene Bakterien untersucht.
Acrolein findet bisher Verwendung u.a. als aquatisches Biozid, als Mikrobiozid in
Kühltürmen und bei Ölbohrungen (Turner & Erickson, 2003). Von Baker Petrolite
(2001) wurde der Einsatz von Acrolein als Ballastwasser-Biozid untersucht. Bekannt ist
Acrolein außerdem als Kampfstoff aus dem Ersten Weltkrieg. Es ist ein sehr giftiges,
hochreaktives und umweltgefährliches Aldehyd (Ghilarducci & Tjeerdema, 1995). Die
Toxizität von Acrolein äußert sich in der Zerstörung der Zellmembranintegrität und der
Affinität für Thiolgruppen. Als Folge werden vitale Enzymsysteme beeinträchtigt, z. B.
die oxidative Phosphorylierung und die Synthese von RNA, DNA und Proteinen.
(Ghilarducci & Tjeerdema, 1995)
Acrolein verflüchtigt sich in Wasser. Neben dem primären Hydrolyseprodukt 3-
Hydroxypropanal entstehen unter anderem Acrylsäure, Allylalkohol, Propionsäure,
Propanal und 3-Hydroxypropansäure (Smith et al, 1995). Besonders auf aquatische
Organismen wirkt es hoch toxisch, die hohe Wasserlöslichkeit lässt jedoch eine niedrige
Bioakkumulation erwarten. Die Reaktivität ist temperaturabhängig (Ghilarducci &
Tjeerdema, 1995).
1.3 Bedeutung der untersuchten pathogenen Mikroorganismen
In den 1990ern wurde belegt, dass über Ballastwasser der Cholera-Erreger V. cholerae
verbreitet wurde und mehr als 10.000 Menschen in Peru das Leben kostete (Kolar &
Lodge, 2001, Drake et al, 2006). Marine pathogene Bakterien verursachen
Lebensmittelvergiftungen und Wundinfektionen beim Menschen und zahlreiche
Fischkrankheiten. Trotzdem gibt es bisher kaum Untersuchungen, die sich mit der
Verschleppung von pathogenen Bakterien über Ballastwasser oder der Wirkung von
Ballastwasserbehandlung auf pathogene Bakterien befassen.
Für diese Arbeit wurden neun verschiedene pathogene Bakterien ausgewählt, die in
Meer-, Brack- und Süßwasser vorkommen. Zu den untersuchten Pathogenen gehören

Einleitung
12
V. parahaemolyticus, V. alginolyticus, V. vulnificus, A. hydrophila, S. liquefaciens,
E. coli, E. faecalis und P. aeruginosa. Außerdem wurde noch das Umweltisolat dB2 aus
einer Pilot-Ballastwasser-Behandlungsanlage isoliert.
In Tabelle 1 sind einige der Krankheiten aufgeführt, die von den in dieser Arbeit
untersuchten Pathogenen verursacht werden können. Bei E. coli, E. faecalis und
P. aeruginosa handelt es sich um Darmbakterien und Indexkeime für verunreinigte
Badegewässer und Trinkwasser (www.dgfnb.de; www.umweltbundesamt.de). E. coli
gehört zu den meldepflichtigen Krankheitserregern (www.rki.de).
Die pathogenen Bakterien A. hydrophila und S. liquefaciens führen nur selten zu
Infektionen beim Menschen. Sie gehören zu häufigeren Krankheitserregern bei Fischen
(Tabelle 2).
Die Vibrionen sind typische Meer- und Brackwasserorganismen. Beim Menschen kann
eine Wundinfektion mit V. vulnificus, z. B. über den Kontakt von offenen Wunden mit
Meerwasser, zu einer schweren Sepsis mit einer Mortalität von bis zu 60% führen.
Außerdem ist diese Art in den USA verantwortlich für 95% aller Todesopfer durch
Verzehr von Meeresfrüchten (Linkous & Oliver, 1999). V. parahaemolyticus gehört zu
den Erregern von Gastritis nach dem Verzehr von Schalen- und Krebstieren und hat
damit besonders für Japans Esskultur große Bedeutung (Madigan et al, 2001).
Außerdem haben V. alginolyticus und V. vulnificus Bedeutung für die Pathogenese von
Fischen (Tabelle 2).
Einleitung
13
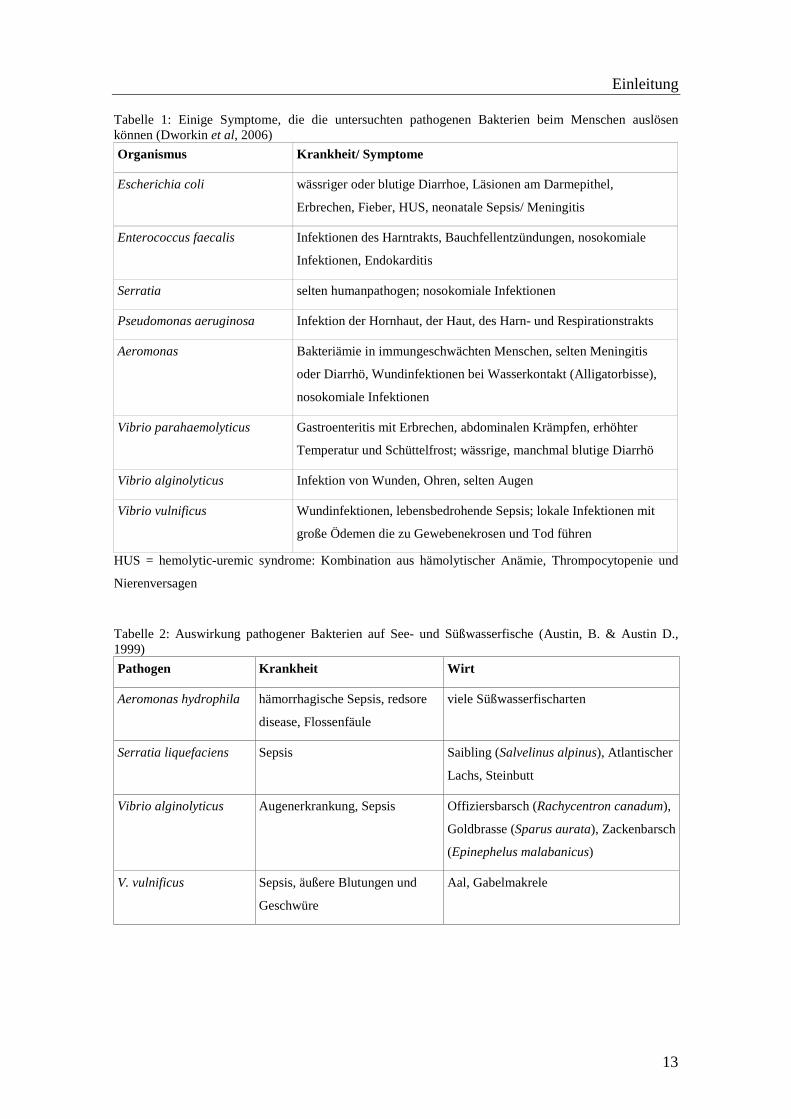
Tabelle 1: Einige Symptome, die die untersuchten pathogenen Bakterien beim Menschen auslösen können (Dworkin et al, 2006)
Organismus Krankheit/ Symptome
Escherichia coli wässriger oder blutige Diarrhoe, Läsionen am Darmepithel,
Erbrechen, Fieber, HUS, neonatale Sepsis/ Meningitis
Enterococcus faecalis Infektionen des Harntrakts, Bauchfellentzündungen, nosokomiale
Infektionen, Endokarditis
Serratia selten humanpathogen; nosokomiale Infektionen
Pseudomonas aeruginosa Infektion der Hornhaut, der Haut, des Harn- und Respirationstrakts
Aeromonas Bakteriämie in immungeschwächten Menschen, selten Meningitis
oder Diarrhö, Wundinfektionen bei Wasserkontakt (Alligatorbisse),
nosokomiale Infektionen
Vibrio parahaemolyticus Gastroenteritis mit Erbrechen, abdominalen Krämpfen, erhöhter
Temperatur und Schüttelfrost; wässrige, manchmal blutige Diarrhö
Vibrio alginolyticus Infektion von Wunden, Ohren, selten Augen
Vibrio vulnificus Wundinfektionen, lebensbedrohende Sepsis; lokale Infektionen mit
große Ödemen die zu Gewebenekrosen und Tod führen
HUS = hemolytic-uremic syndrome: Kombination aus hämolytischer Anämie, Thrompocytopenie und
Nierenversagen
Tabelle 2: Auswirkung pathogener Bakterien auf See- und Süßwasserfische (Austin, B. & Austin D., 1999)
Pathogen Krankheit Wirt
Aeromonas hydrophila hämorrhagische Sepsis, redsore
disease, Flossenfäule
viele Süßwasserfischarten
Serratia liquefaciens Sepsis Saibling (Salvelinus alpinus), Atlantischer
Lachs, Steinbutt
Vibrio alginolyticus Augenerkrankung, Sepsis Offiziersbarsch (Rachycentron canadum),
Goldbrasse (Sparus aurata), Zackenbarsch
(Epinephelus malabanicus)
V. vulnificus Sepsis, äußere Blutungen und
Geschwüre
Aal, Gabelmakrele

Einleitung
14
1.4 Zielsetzung
Vor dem Hintergrund der Ballastwasser-Behandlung wurde in dieser Arbeit die
Wirkung des Biozids Acrolein auf verschiedene pathogene Bakterien untersucht. Ziel
dieser Arbeit ist es, heraus zu finden, welche Konzentration an Acrolein nötig ist, um
verschiedene pathogene Bakterien innerhalb von fünf Tagen Inkubation in Meer-,
Brack- und Süßwasser abzutöten. Die Untersuchungen in Meer-, Brack- und Süßwasser
sollen zeigen, ob sich die Reaktion der Bakterien auf Acrolein bei unterschiedlichem
Salzgehalt ändert. Außerdem soll bewertet werden, ob die angewandte Methode für
Untersuchungen zu dieser Fragestellung geeignet war. Für die Untersuchungen wurde
die ScanMIC-Methode (Rahman et al, 2003) angewendet, gekoppelt mit einer MPN-
Auswertung (McCrady, 1915).

Material & Methoden
15
2 Material und Methoden
2.1 Bakterienstämme
Für die Untersuchungen wurden verschiedene pathogene Bakterienstämme verwendet:
Escherichia coli DSM 30083, Pseudomonas aeruginosa DSM 50071,
Aeromonas hydrophila DSM 30188, Serratia liquefaciens DSM 4487,
Enterococcus faecalis DSM 20478, Vibrio parahaemolyticus DSM 2172,
Vibrio alginolyticus DSM 2171, Vibrio vulnificus DSM 10143 und Vibrio vulnificus
DSM 11507. Zusätzlich wurde ein Umweltisolat untersucht, das aus einer Pilot-
Ballastwasser-Behandlungsanlage isoliert wurde, dB2. Die untersuchten Bakterien
decken ein breites Spektrum an Lebensräumen, Salztoleranz und pathogener Wirkung
ab. Die pathogenen Bakterienstämme wurden von der Deutschen Sammlung von
Mikroorganismen und Zellkulturen GmbH (DSMZ) erworben (www.dsmz.de). Bis auf
E. faecalis gehören alle untersuchten pathogenen Stämme den Gammaproteobacteria an.
E. faecalis ist den Bacilli zuzurechnen (www.ncbi.nlm.nih.gov).
2.1.1 Kultivierung
Die Isolate wurden auf ihrem jeweiligen Optimalmedium kultiviert (Tabelle 3). Sie
wurden circa alle zehn Tage auf frische Agarplatten überimpft und ein bis zwei Tage bei
ca. 30°C, das Umweltisolat bei 18°C, inkubiert. Sobald die Isolate Kolonien gebildet
hatten wurden die Agarplatten mit Parafilm verschlossen und bis zur nächsten
Überimpfung bei 4°C im Kühlschrank gelagert.
Tabelle 3: Kultivierung der untersuchten Bakterien
Mikroorganismus Medium Temperatur
Escherichia coli DSM 30083 M1 37°C
Pseudomonas aeruginosa DSM 50071 M1 30°C
Aeromonas hydrophila DSM 30188 M1 30°C
Serratia liquefaciens DSM 4487 M1 30°C
Enterococcus faecalis DSM 20478 M53 37°C

Material & Methoden
16
Vibrio parahaemolyticus DSM 2172 M2216E 30°C
Vibrio alginolyticus DSM 2171 M2216E 28-37°C
Vibrio vulnificus DSM 10143 M2216E 30°C
Vibrio vulnificus DSM 11507 M220 28°C
dB2 10 ml M2216E 18°C
M1 = Nutrient Agar; M53 = Corynebacterium Agar; M2216 E = ZoBell-Agar; M220 =
Caso-Agar
2.1.2 Wachstumskurven
Für alle Isolate wurden Wachstumskurven bei 30°C, für die Umweltisolate bei 18°C,
aufgenommen. Die Optische Dichte (OD) wurde bei einer Wellenlänge von 600 nm in
einem Eppendorf-Photometer alle 30 Minuten gemessen. In drei Parallelen wurden
jeweils 85 µl der Isolate in einer Küvette gevortext und gemessen.
2.1.3 Einstellung der Optischen Dichte (OD)
Um zu Beginn der Inkubationsexperimente eine ähnliche Anzahl an Bakterien
anzuimpfen wurden die Flüssigkulturen mit dem Photometer gemessen und jeweils auf
eine OD (600 nm) von 0.1 verdünnt. Die Verdünnung erfolgte mit dem jeweiligen
Verdünnungswasser (siehe Kapitel 2.4.5).
2.2 Flüssigkeiten und Lösungen
2.2.1 TTC (2,3,5-Triphenyltetrazoliumchlorid)
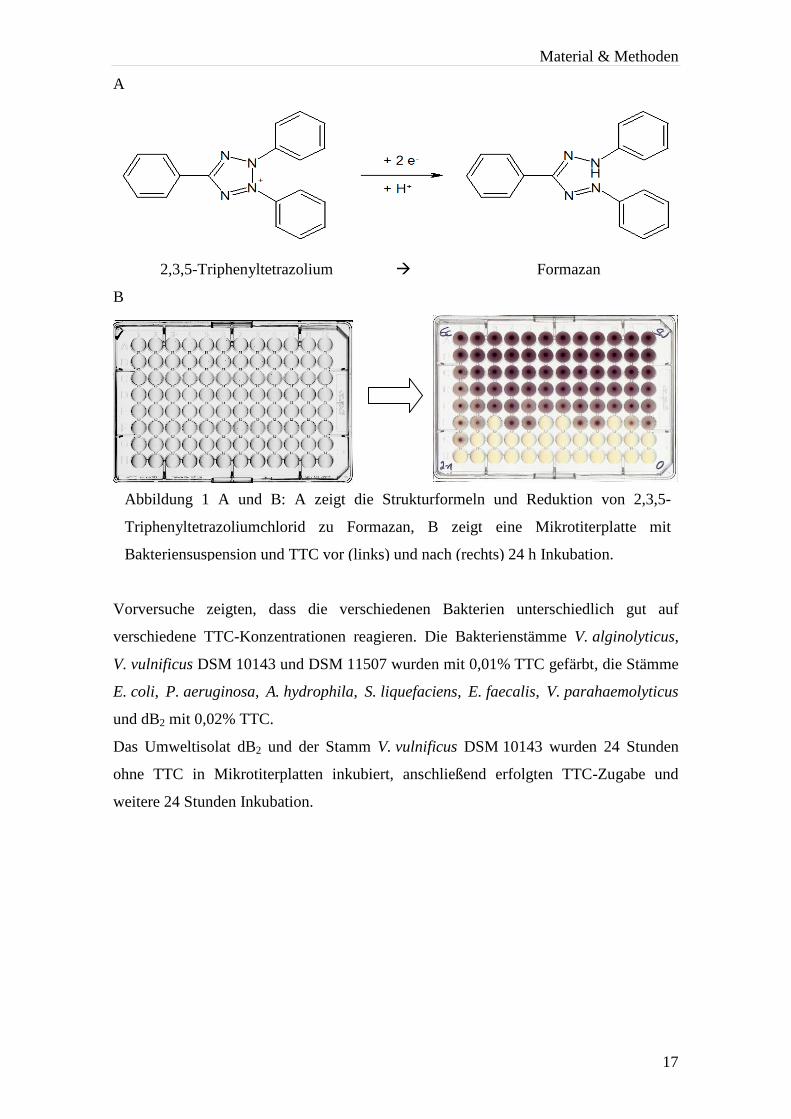
2,3,5-Triphenyltetrazoliumchlorid (TTC) dient als Farbstoff, um lebende Bakterien für
das menschliche Auge sichtbar zu machen. Bei funktionierender Atmungskette wird das
farblose Tetrazoliumsalz durch mitochondriale Dehydrogenasen zum rötlichen
Formazan (Abb. 1) reduziert.
Material & Methoden
17
A
2,3,5-Triphenyltetrazolium � Formazan
B
Vorversuche zeigten, dass die verschiedenen Bakterien unterschiedlich gut auf
verschiedene TTC-Konzentrationen reagieren. Die Bakterienstämme V. alginolyticus,
V. vulnificus DSM 10143 und DSM 11507 wurden mit 0,01% TTC gefärbt, die Stämme
E. coli, P. aeruginosa, A. hydrophila, S. liquefaciens, E. faecalis, V. parahaemolyticus
und dB2 mit 0,02% TTC.
Das Umweltisolat dB2 und der Stamm V. vulnificus DSM 10143 wurden 24 Stunden
ohne TTC in Mikrotiterplatten inkubiert, anschließend erfolgten TTC-Zugabe und
weitere 24 Stunden Inkubation.
Abbildung 1 A und B: A zeigt die Strukturformeln und Reduktion von 2,3,5-
Triphenyltetrazoliumchlorid zu Formazan, B zeigt eine Mikrotiterplatte mit
Bakteriensuspension und TTC vor (links) und nach (rechts) 24 h Inkubation.

Material & Methoden
18
2.2.2 Acrolein C3H4O
Abbildung 2: Strukturformel und Gefahrenklassen „leichtentzündlich“, „giftig“ und
„umweltgefährlich“ von Acrolein
Als potentielles Mittel zur Ballastwasserbekämpfung wurde in dieser Arbeit Acrolein
(2-Propenal) untersucht. Es ist ein sehr giftiges, hochreaktives und umweltgefährliches
Aldehyd. Die farblose, leicht entzündliche Flüssigkeit bildet mit Luft
explosionsgefährliche Gemische. Der Geruch ist beißend (Ghilarducci & Tjeerdema,
1995).
In dieser Arbeit wurde die Wirkung sechs verschiedener Acrolein-Konzentrationen auf
verschiedene Bakterienstämme untersucht. In Tabelle 4 werden diese Acrolein-
Konzentrationen mit Acroleinwerten verglichen, deren gesundheitliche Auswirkungen
in der Literatur beschrieben wurden.
Tabelle 4: Toxische Wirkung von Acrolein (Ghilarducci & Tjeerdema, 1995; Turner&
Erickson, 2003; Louder & McCoy, 1962). C3H4O (test) = Acrolein-Konzentrationen,
die in dieser Arbeit untersucht wurden, C3H4O (Literatur) = Acrolein-Konzentrationen
mit bekannter Wirkung
C3H4O
(test)
C3H4O
(Literatur)
Zeit Wirkung (Mensch) Wirkung (Fisch)
0,01 mg/L < 0,031
mg/L
96 h LC50 Regenbogenforelle
(Oncorhynchus mykiss)
0,1 mg/L 0,1 mg/m3 5 min Augenirritation LC50/EC50 Blauer
Sonnenbarsch (Lepomis
macrochirus)
1 mg/L 1 mg/m3 5 min Tränenfluss
5 mg/L 4,14 mg/m3 1 min Leichte Augenirritation
3-4 min übermäßiger
Tränenfluss, praktisch
intolerabel

Material & Methoden
19
10 mg/L 11,5 mg/m3 30 min neurotoxische
Symptome (IDLH1)
15 mg/L > 23 mg/m3 tödlich nach kurzer Zeit
1 IDLH = Immediately Dangerous to Life and Health
2.2.3 Probenwasser
Die Inkubationsversuche wurden jeweils mit Meerwasser, Süßwasser und Brackwasser
durchgeführt. Das jeweilige Wasser wurde durch 10 µm-Gaze filtriert und autoklaviert.
Die Lagerung erfolgte bei 4-6°C.
Das Meerwasser wurde auf Helgoland am Süd-Ost-Hafen geholt, das Süßwasser stammt
aus der Elbe, nahe dem Anleger Hoopte. Das Brackwasser wurde mit dem
Forschungsschiff Uthörn an der Kugelbake bei Wilhelmshaven entnommen.
Der Salzgehalt (Tabelle 5:) des hier verwendeten Wassers wurde von der AG
Biologische Ozeanographie bestimmt.
Tabelle 5: Salzgehalt und Probenahmeort für Meer-, Brack- und Süßwasser Bezeichnung Ort der Wasserentnahme Salzgehalt [PSU]
Meerwasser Helgoland, Süd-Ost-Hafen 35.31
Brackwasser Cuxhaven, Kugelbake 16.00
Süßwasser Elbeanleger Hoopte 0.27
2.3 Versuchsaufbau
2.3.1 ScanMIC-Methode
Für die Versuche in dieser Arbeit wurde die ScanMIC-Methode angewendet. ScanMIC
steht für scanner-assisted colorimetric MIC-method (MIC = minimal inhibition
concentration) (Rahman et al, 2003). Die ScanMIC-Methode sieht vor, dass
durchsichtige 96-well Mikrotiterplatten mit Medium, TTC und Bakteriensuspension
versetzt werden, und nach 24 h Inkubation mit einem Flachbett-Scanner eingescannt
werden. Man verwendet Mikrotiterplatten mit Rundboden-Vertiefungen für diese
Methode, da sich so ein Pellet am Boden bilden kann.

Material & Methoden
20
Als Scan-Programm wurde EPSON Scan verwendet. Im Professional Mode wurden die
Mikrotiterplatten mit 24 bit Farbtiefe und einer Auflösung von 600 dpi gescant.
Eine Übernachtkultur jedes Isolates wurde mit Verdünnungswasser auf eine OD von 0,1
verdünnt. In 96-well-Rundboden-Mikrotiterplatten wurden pro Kavität je 20 µl
verdünnter Bakteriensuspension auf 180 µl TTC-Medium gegeben. Das TTC-Medium
bestand aus dem jeweiligen Optimalmedium der Isolate und der jeweils erforderlichen
Konzentration an TTC-Lösung (siehe Punkt 2.2.1). Die Mikrotiterplatten wurden bei
30°C 24 h inkubiert, mit Ausnahme des Umweltisolats dB2, das bei 18°C inkubiert
wurde.
Die Auswertung der gescannten Mikrotiterplatten erfolgte anhand der MPN-Tabelle
(MPN = most probable number) nach McCrady (1915). Diese Tabelle ermöglicht es,
anhand von Parallelen und Verdünnungsstufen die wahrscheinliche Keimzellzahl pro
Milliliter zu ermitteln.
Material & Methoden
21
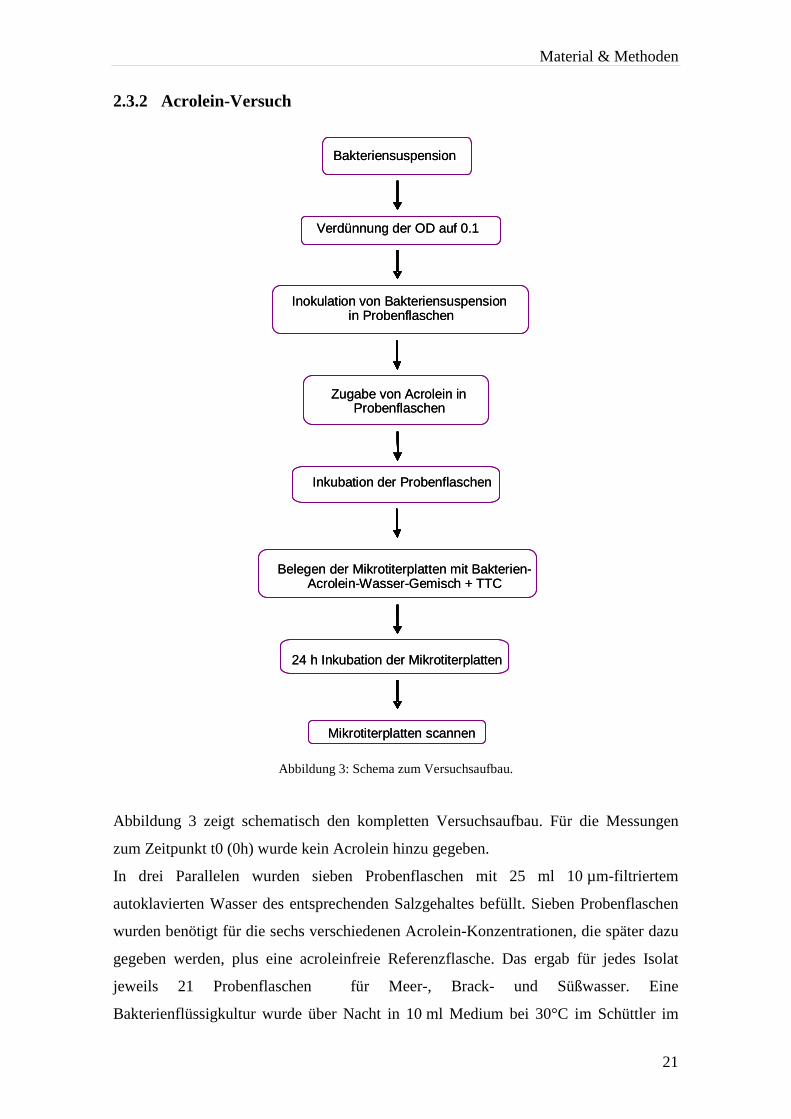
2.3.2 Acrolein-Versuch
Abbildung 3: Schema zum Versuchsaufbau.
Abbildung 3 zeigt schematisch den kompletten Versuchsaufbau. Für die Messungen
zum Zeitpunkt t0 (0h) wurde kein Acrolein hinzu gegeben.
In drei Parallelen wurden sieben Probenflaschen mit 25 ml 10 µm-filtriertem
autoklavierten Wasser des entsprechenden Salzgehaltes befüllt. Sieben Probenflaschen
wurden benötigt für die sechs verschiedenen Acrolein-Konzentrationen, die später dazu
gegeben werden, plus eine acroleinfreie Referenzflasche. Das ergab für jedes Isolat
jeweils 21 Probenflaschen für Meer-, Brack- und Süßwasser. Eine
Bakterienflüssigkultur wurde über Nacht in 10 ml Medium bei 30°C im Schüttler im
Bakteriensuspension
Verdünnung der OD auf 0.1
Inokulation von Bakteriensuspension in Probenflaschen
24 h Inkubation der Mikrotiterplatten
Mikrotiterplatten scannen
Zugabe von Acrolein inProbenflaschen
Belegen der Mikrotiterplatten mit Bakterien-Acrolein-Wasser-Gemisch + TTC
Inkubation der Probenflaschen
Bakteriensuspension
Verdünnung der OD auf 0.1
Inokulation von Bakteriensuspension in Probenflaschen
24 h Inkubation der Mikrotiterplatten
Mikrotiterplatten scannen
Zugabe von Acrolein inProbenflaschen
Belegen der Mikrotiterplatten mit Bakterien-Acrolein-Wasser-Gemisch + TTC
Inkubation der Probenflaschen

Material & Methoden
22
Dunkeln inkubiert. Es erfolgte die Verdünnung der Übernachtkultur auf eine OD von
0,1. In jede Probenflasche wurden 250 µl Bakteriensuspension der OD 0,1 zugegeben.
In je 1 ml Probenwasser wurden die sechs verschiedenen Acrolein-Konzentrationen
hergestellt und auf die Probenflaschen verteilt: Jeweils eine Konzentrationsstufe wurde
in drei parallele Probenflaschen gegeben. Die Probenflaschen wurden bei 20°C
inkubiert. Aus den inkubierten Probenflaschen wurden alle 24 h neue Mikrotiterplatten
belegt.
Für jeden Bakterienstamm wurden täglich sechs Mikrotiterplatten belegt. In jede dieser
Mikrotiterplatten wurden je 180 µl TTC-Medium und je 20 µl des Bakterien-Acrolein-
Wasser-Gemischs pro Kavität inokuliert (Abbildung 4): Aus jeder Probenflasche
wurden drei parallele Kavitäten inokuliert. Dementsprechend passten auf zwei
Mikrotiterplatten sieben Proben und ein Blank, das ergab bei 21 Probenflaschen sechs
Mikrotiterplatten. Die Beimpfung von drei parallelen Kavitäten wurde für die
Auswertung mithilfe der MPN-Tabelle benötigt.
Abbildung 4: Inokulation der Mikrotiterplatten aus den Probenflaschen. Jeweils drei
Kavitäten werden aus einer Probenflasche inokuliert. Pro sieben Flaschen werden zwei
Mikrotiterplatten belegt. Die Zahlen auf den Probenflaschen und über den
Mikrotiterplatten geben die verwendeten Acrolein-Konzentrationen an, die pro Flasche
in je drei Kavitäten überimpft wurden.
Die Mikrotiterplatten wurden mit Deckeln verschlossen 24 Stunden bei 20°C inkubiert.
Nach 24 Stunden wurden die Mikrotiterplatten auf einem Flachbett-Scanner eingelesen.
0 0.01 0.1 1
0 0.01 0.1 1 5 10 15
5 10 15 neg
Verteilung auf Mikroplatten

Material & Methoden
23
Im Fall des Umweltisolats erfolgte die Inkubation der Probenflaschen und der
Mikrotiterplatten bei 18°C.
Die Wirkung von Acrolein wurde von t1 (24 h) bis t5 (120 h) untersucht. Der Versuch
wurde jeweils mit Meerwasser, Brackwasser und Süßwasser durchgeführt.
2.4 Material
2.4.1 Technische Geräte Funktion Hersteller
Abzug Wesemann Laboreinrichtungen
Köttermann Labortechnik
Autoklav H+P
Bunsenbrenner ADG
Inkubationsschrank EWALD Innovationstechnik
GmbH
Rubarth&Co.
Kühlschrank Brillant
Kulturraum Linde
Magnetrührer Janke & Kunkel KG, IKA Werk
Notebook Compaq
Photometer Eppendorf
pH-Meter Knick
Pipetten Gilson, Abimed
Socorex
Eppendorf
Pumpe KNF Neuberger
Reinstwasseranlage Millipore GmbH
Scanner Epson
Schüttler Heidolph
Kippschüttler Kroll Laborbedarf
Sterilbank EHRET Reinraumtechnik
USB-Stick Hama
Vortex Bender&Hobein AG
Waagen Sartorius

Material & Methoden
24
Zentrifuge Eppendorf AG
2.4.2 Verbrauchsmaterial Funktion Hersteller
Bechergläser Schott
Vitlab
Desinfektionslösung Bode
Einwegpipetten Falcon; Becton Dickinson
Labware
Erlenmeyerkolben Brand
Filter, 0,2 µm Sartorius AG
Filter, 10 µm Gaze Sefar
Filtergestelle Nalgene
Glasflaschen Schott
Pyrex (Ribby)
Handcreme Bode
LabodermneoLab GmbH
Handschuhe rotiprotect; Carl Roth GmbH&Co.
Impfschlingen Sarstedt Aktiengesellschaft & Co.
Kautex-Kanister, 30 l Omnilab
Küvetten Brand Plastibrand®
Messzylinder Nalgene
Mikrotiterplatten Greiner
Mikrotiterplatten-Deckel Greiner
Müllbeutel Brand Plastibrand
Sarstedt Aktiengesellschaft & Co.
Reagent Reservoirs Eppendorf
Parafilm Carl Roth
American Can Company
Petrischalen Greiner
Pipettenspitzen Sarstedt Aktiengesellschaft & Co.
Brand Plastibrand®
Pipettierhilfe Bel-Art Products®

Material & Methoden
25
Brand
Probenflaschen United Glass
Ständer Nalgene
2.4.3 Chemikalien Funktion Hersteller
Acrolein Fluka
Agar
Bacto Beef Extract DIFCO Laboratories
Bacto Peptone DIFCO Laboratories
Bacto® Yeast Extract DIFCO Laboratories
BactoTM Tryptic Soy Broth
- Soybean-Casein Digest Medium
D(+)-Glucose-Monohydrat Merck
Eisen(III)-phosphat, geglüht (annähernd FePO4)
Natriumchlorid Merck
2,3,5-Triphenyltetrazoliumchlorid Merck
Tryptone Peptone - Pancreatic Digest of Casein DIFCO
2.4.4 Hergestellte Medien und Lösungen
Alle hergestellten Medien und Lösungen wurden vor dem Gebrauch autoklaviert.
Medium 1: Nutrient Agar (pH 7,0)
Peptone 5 g
Beef extract 3 g
Agar, ggf. 12 g
Aqua dest. 1000 ml
Medium 53: Corynebacterium Agar (pH 7,2 - 7,4)
Casein peptone, tryptic digest 10 g
Hefeextrakt 5 g

Material & Methoden
26
Glucose 5 g
NaCl 5 g
Agar, ggf. 12 g
Aqua dest. 1000 ml
Medium 2216E: ZoBell Agar (pH 7,6)
Peptone 5 g
Hefe 1 g
FePO4 0,01 g
Agar, ggf. 12 g
Aqua dest. 250 ml
Seewasser 750 ml
Medium 220: Caso Agar (pH 7,3)
Casein peptone 15 g
Tryptic Soy Broth – Soybean-Casein Digest 5 g
NaCl 5 g
Agar, ggf. 15 g
Aqua dest. 1000 ml
1%iges TTC
2,3,5-Triphenyletrazoliumchlorid 0,1 g
Aqua dest. 10 ml
Verdünnungswasser
Medium Verdünnungswasser
Nutrient Broth Millipore-Wasser 100 ml
Corynebacterium Medium Millipore-Wasser 100 ml
Caso Medium Millipore-Wasser 100 ml
ZoBell Medium Seewasser
Millipore-Wasser
75
25
ml
ml
Ergebnisse
3 Ergebnisse
3.1 Phänotyp der Bakterienstämme
Die untersuchten Bakterienstämme zeigten sowohl in kultivierter Form auf Agarplatten,
als auch in den Mikrotiterplatten morphologische Unterschiede.
3.1.1 Kultivierung
Wie bereits beschrieben (Punkt 2.1.1) wurden die untersuchten Bakterien auf
Agarplatten kultiviert und regelmäßig überimpft. Form und Wuchsverhalten erwiesen
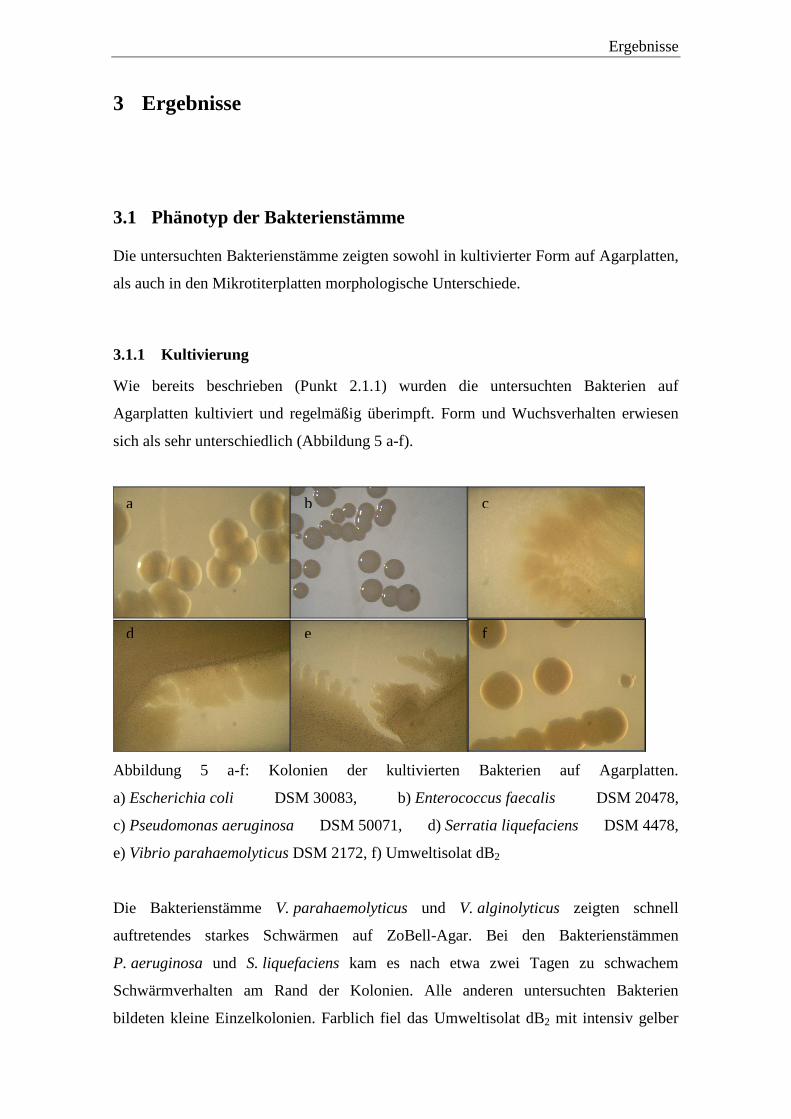
sich als sehr unterschiedlich (Abbildung 5 a-f).
Abbildung 5 a-f: Kolonien der kultivierten Bakterien auf Agarplatten.
a) Escherichia coli DSM 30083, b) Enterococcus faecalis DSM 20478,
c) Pseudomonas aeruginosa DSM 50071, d) Serratia liquefaciens DSM 4478,
e) Vibrio parahaemolyticus DSM 2172, f) Umweltisolat dB2
Die Bakterienstämme V. parahaemolyticus und V. alginolyticus zeigten schnell
auftretendes starkes Schwärmen auf ZoBell-Agar. Bei den Bakterienstämmen
P. aeruginosa und S. liquefaciens kam es nach etwa zwei Tagen zu schwachem
Schwärmverhalten am Rand der Kolonien. Alle anderen untersuchten Bakterien
bildeten kleine Einzelkolonien. Farblich fiel das Umweltisolat dB2 mit intensiv gelber
a b c
d e f
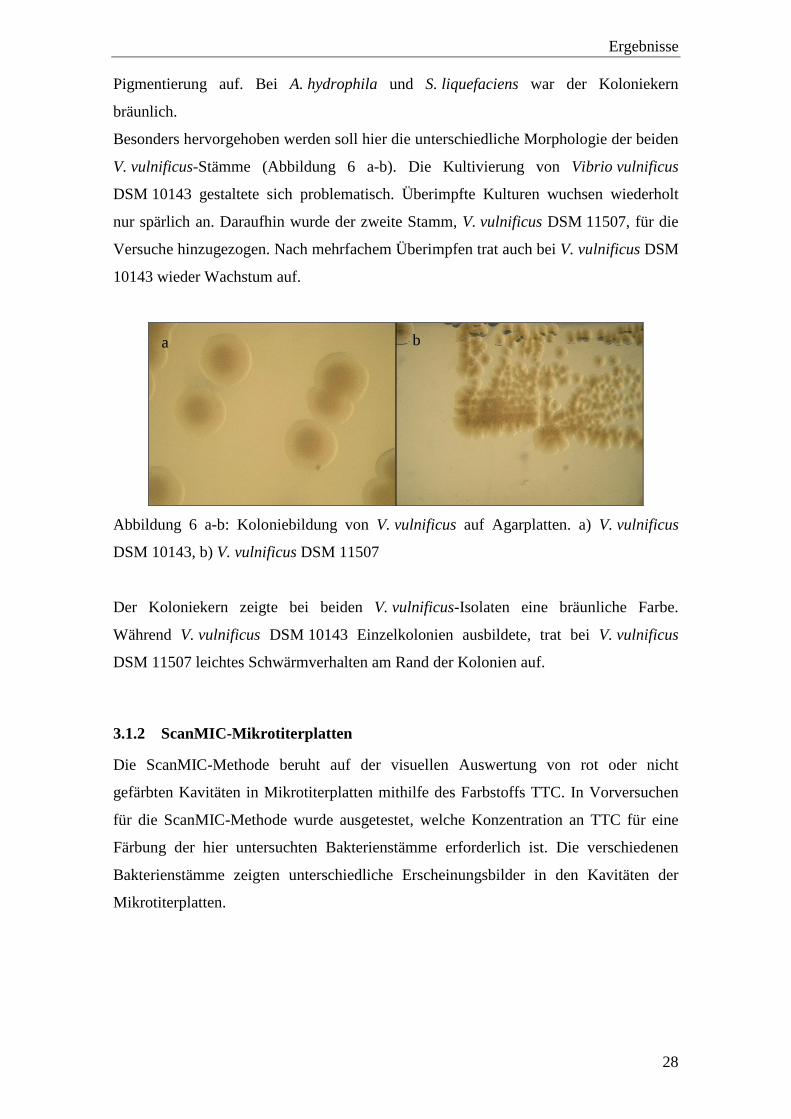
Ergebnisse
28
Pigmentierung auf. Bei A. hydrophila und S. liquefaciens war der Koloniekern
bräunlich.
Besonders hervorgehoben werden soll hier die unterschiedliche Morphologie der beiden
V. vulnificus-Stämme (Abbildung 6 a-b). Die Kultivierung von Vibrio vulnificus
DSM 10143 gestaltete sich problematisch. Überimpfte Kulturen wuchsen wiederholt
nur spärlich an. Daraufhin wurde der zweite Stamm, V. vulnificus DSM 11507, für die
Versuche hinzugezogen. Nach mehrfachem Überimpfen trat auch bei V. vulnificus DSM
10143 wieder Wachstum auf.
Abbildung 6 a-b: Koloniebildung von V. vulnificus auf Agarplatten. a) V. vulnificus
DSM 10143, b) V. vulnificus DSM 11507
Der Koloniekern zeigte bei beiden V. vulnificus-Isolaten eine bräunliche Farbe.
Während V. vulnificus DSM 10143 Einzelkolonien ausbildete, trat bei V. vulnificus
DSM 11507 leichtes Schwärmverhalten am Rand der Kolonien auf.
3.1.2 ScanMIC-Mikrotiterplatten
Die ScanMIC-Methode beruht auf der visuellen Auswertung von rot oder nicht
gefärbten Kavitäten in Mikrotiterplatten mithilfe des Farbstoffs TTC. In Vorversuchen
für die ScanMIC-Methode wurde ausgetestet, welche Konzentration an TTC für eine
Färbung der hier untersuchten Bakterienstämme erforderlich ist. Die verschiedenen
Bakterienstämme zeigten unterschiedliche Erscheinungsbilder in den Kavitäten der
Mikrotiterplatten.
a b

Ergebnisse
29
Abbildung 7 a-f: Färbung der Kavitäten in den Mikrotiterplatten. a) E. coli,
b) E. faecalis, c) P. aeruginosa, d) S. liquefaciens, e) V. parahaemolyticus,
f) Umweltisolat dB2
E. coli, A. hydrophila, V. parahaemolyticus und V. alginolyticus bildeten zentrale
scharfe Pellets in rot gefärbtem Medium aus (Abbildung 7 a+e).
E. faecalis unterschied sich am deutlichsten von den anderen untersuchten Bakterien, es
bildete neben dem Pellet wolkenartige rot gefärbte Partikelhaufen mit ungefärbten
Bereiche am Rand aus (Abbildung 7 b). Es kam zu keiner Rotfärbung des umgebenden
Mediums.
Bei P. aeruginosa (Abbildung 7 c) traten in geringen Verdünnungsstufen Pellets
umgeben von rotem Medium auf, bei steigender Verdünnung bildeten sich statt Pellets
rot gefärbte Partikel. Diese Partikel erschienen aber nicht wie bei E. faecalis als
zentraler Haufen sondern flächig innerhalb der Kavität verteilt. Bei Auftreten der
Partikel kam es zu keiner Rotfärbung des umgebenden Mediums.
In den Kavitäten mit S. liquefaciens traten neben einem zentralen Pellet rot gefärbte
Partikel auf, die eher am Rand des Kavitäten zu finden waren (Abbildung 7 d). Es kam
bei diesem Bakterium nicht zur Ausbildung gefärbter Partikel ohne ein vorhandenes
Pellet.
Das Umweltisolat dB2 bildete ebenfalls rot gefärbte Partikel neben einem zentralen
Pellet aus (Abbildung 7 f). In niedrigerer Verdünnung traten statt einem Pellet nur
gefärbte Partikel auf. Es kam in keiner Verdünnungsstufe zu einer Rotfärbung des
umgebenden Mediums.
a b c
d e f

Ergebnisse
30
Die beiden V. vulnificus-Stämme zeigten in den Mikrotiterplatten unterschiedliche
Färbungen der Kavitäten.
Abbildung 8 a-b: Färbung der beiden V. vulnificus-Stämme in Mikrotiterplatten.
a) V. vulnificus DSM 10143, b) V. vulnificus DSM 11507
Bei der Färbung von V. vulnificus DSM 10143 mit TTC kam es zur Bildung scharf
abgegrenzter zentraler roter Pellets mit einer Rotfärbung des umgebenden Mediums
(Abbildung 8 a).
V. vulnificus DSM 11507 zeigte zwei verschiedene Erscheinungsbilder. Größtenteils
erschienen in den Kavitäten scharfe rote Pellets im rot gefärbtem Medium, doch ab der
Acrolein-Konzentration 1 mg/L, oder bei großer Verdünnungsstufe, erschienen zentrale
Pellets, meistens mit rot gefärbten Partikeln darum herum, ohne eine Rotfärbung des
Mediums (Abbildung 8 b).
3.2 Wachstum der untersuchten Isolate
Mit der Hilfe von Wachstumskurven wurde ermittelt, wie lange die zu untersuchenden
Bakterien inkubiert werden müssen, um die stationäre Phase zu erreichen. Kurz nach
Erreichen dieser Phase wurden sie im Folgenden in den Versuchsaufbau überimpft. Des
Weiteren wurde die Zellzahl ermittelt, die die verschiedenen Bakterienstämme während
ihrer Inkubation in Meer-, Brack- und Süßwasser über fünf Tage hinweg erreichten. Die
Ermittlung der Zellzahl erfolgte mithilfe einer MPN-Tabelle nach McCrady (1915).
3.2.1 Einteilung in Gruppen
Bei den in dieser Diplomarbeit verwendeten Bakterienstämmen handelt es sich um
Organismen mit unterschiedlichen Anpassungsfähigkeiten an die verschiedenen Salz-
und Nährstoffbedingungen des Probenwassers. Der Übersichtlichkeit halber wurden die
a b
Ergebnisse
31
untersuchten Bakterien in drei Gruppen aufgeteilt: Meerwasserliebende Bakterien,
süßwasserliebende Bakterien und salztolerante Bakterien.
1. „Meerwasserliebende Bakterien“: In dieser Gruppe befinden sich Bakterien, die
in Meerwasser am Besten und in Süßwasser Wasser schlecht oder überhaupt
nicht wuchsen: V. parahaemolyticus, V. alginolyticus und V. vulnificus
DSM 10143
2. „Süßwasserliebende Bakterien“: Diese Gruppe fasst die zwei Bakterienstämme
zusammen, die in Meerwasser schnelles Absterben und in Süßwasser sehr gutes
Wachstum zeigten: E. faecalis und A. hydrophila
3. „Salztolerante Bakterien“: In dieser sehr diversen Gruppe sind diejenigen
Bakterien eingeordnet, die sowohl in Meer-, Brack- als auch Süßwasser gut
wuchsen: E. coli, P. aeruginosa, S. liquefaciens, V. vulnificus DSM 11507 und
das Umweltisolat dB2. Innerhalb dieser Gruppe zeigten die Bakterienstämme
trotzdem unterschiedliche Neigungen zu Meer- oder Süßwasser.
Um Zellzahlen für diese Bakteriengruppen darstellen zu können wurden Mittelwerte
verwendet. Die Mittelwerte wurden aus den Zellzahlen der jeweiligen Gruppen-
Bakterien gewonnen.
3.2.2 Wachstum in Medium
Die Wachstumskurven der untersuchten Bakterienstämme wurden im jeweiligen
Optimalmedium durch stündliche Messungen der Optischen Dichte bei einer
Wellenlänge von 600 nm erstellt.
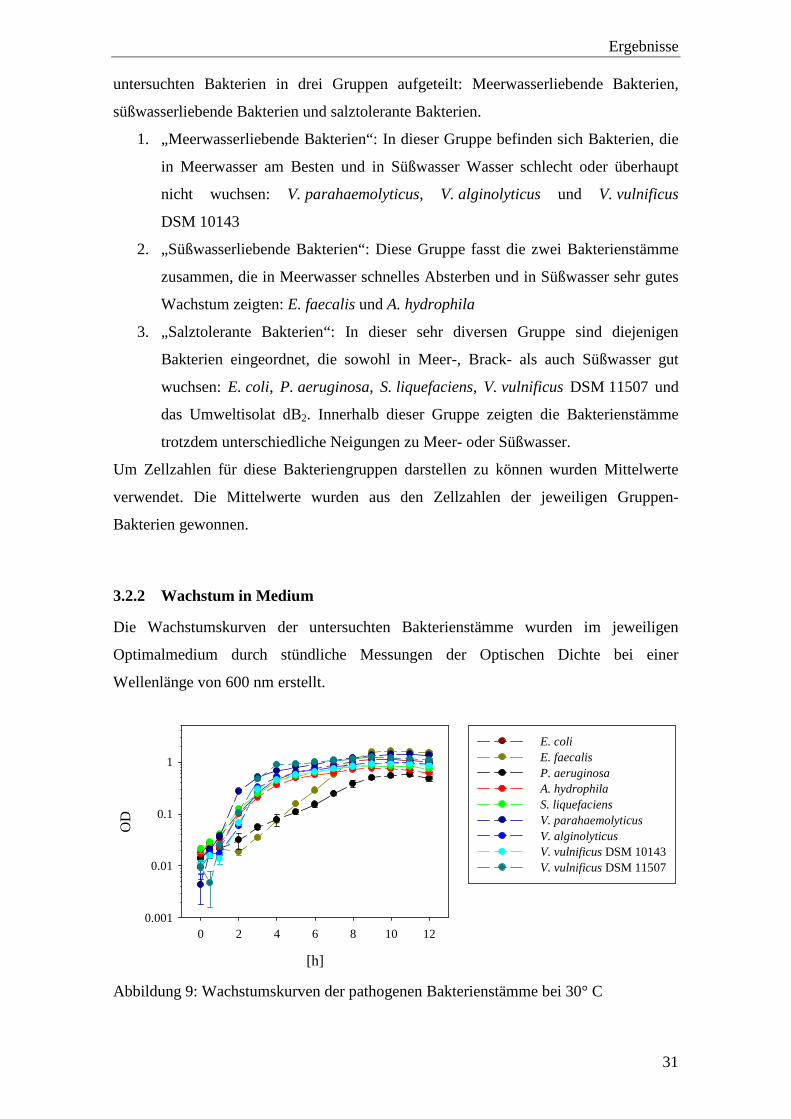
[h]
0 2 4 6 8 10 12
OD
0.001
0.01
0.1
1
E. coliE. faecalisP. aeruginosaA. hydrophilaS. liquefaciensV. parahaemolyticusV. alginolyticusV. vulnificus DSM 10143V. vulnificus DSM 11507
Abbildung 9: Wachstumskurven der pathogenen Bakterienstämme bei 30° C
Ergebnisse
32
Wie Abbildung 9 zeigt gingen die pathogenen Bakterienstämme nach einer Stunde in
die exponentielle Phase über. Der Großteil dieser Bakterien, mit Ausnahme von
P. aeruginosa und E. faecalis, wechselte nach vier bis fünf Stunden von der
exponentiellen in die stationäre Phase.
Bei P. aeruginosa verlief das Wachstum in einer viel flacheren Kurve als bei den
übrigen untersuchten Bakterien. Die Optische Dichte war außerdem niedriger als die der
anderen Bakterien. Die stationäre Phase wurde von P. aeruginosa nach etwa zehn
Stunden erreicht.
E. faecalis hatte eine längere lag-Phase und ging erst nach zwei Stunden in die
exponentielle Phase über, erreichte aber nach neun Stunden eine höhere Optische
Dichte als alle anderen untersuchten Bakterien.
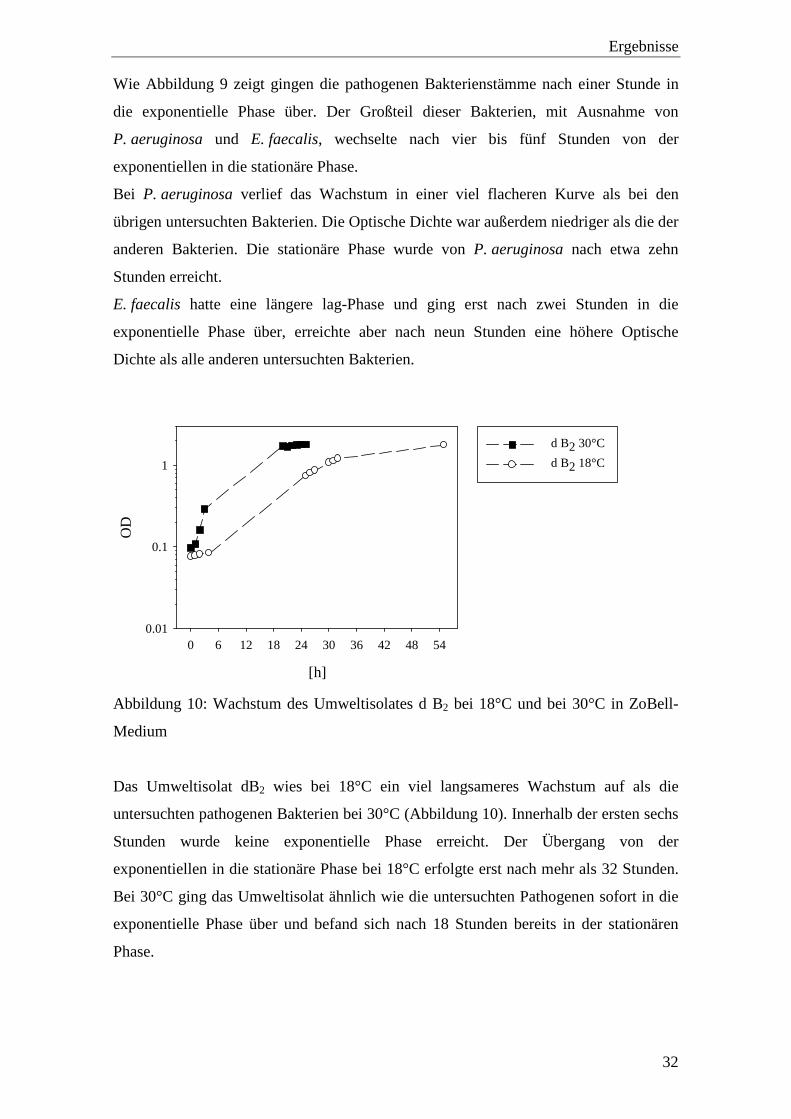
[h]
0 6 12 18 24 30 36 42 48 54
OD
0.01
0.1
1
d B2 30°C
d B2 18°C
Abbildung 10: Wachstum des Umweltisolates d B2 bei 18°C und bei 30°C in ZoBell-
Medium
Das Umweltisolat dB2 wies bei 18°C ein viel langsameres Wachstum auf als die
untersuchten pathogenen Bakterien bei 30°C (Abbildung 10). Innerhalb der ersten sechs
Stunden wurde keine exponentielle Phase erreicht. Der Übergang von der
exponentiellen in die stationäre Phase bei 18°C erfolgte erst nach mehr als 32 Stunden.
Bei 30°C ging das Umweltisolat ähnlich wie die untersuchten Pathogenen sofort in die
exponentielle Phase über und befand sich nach 18 Stunden bereits in der stationären
Phase.
Ergebnisse
33
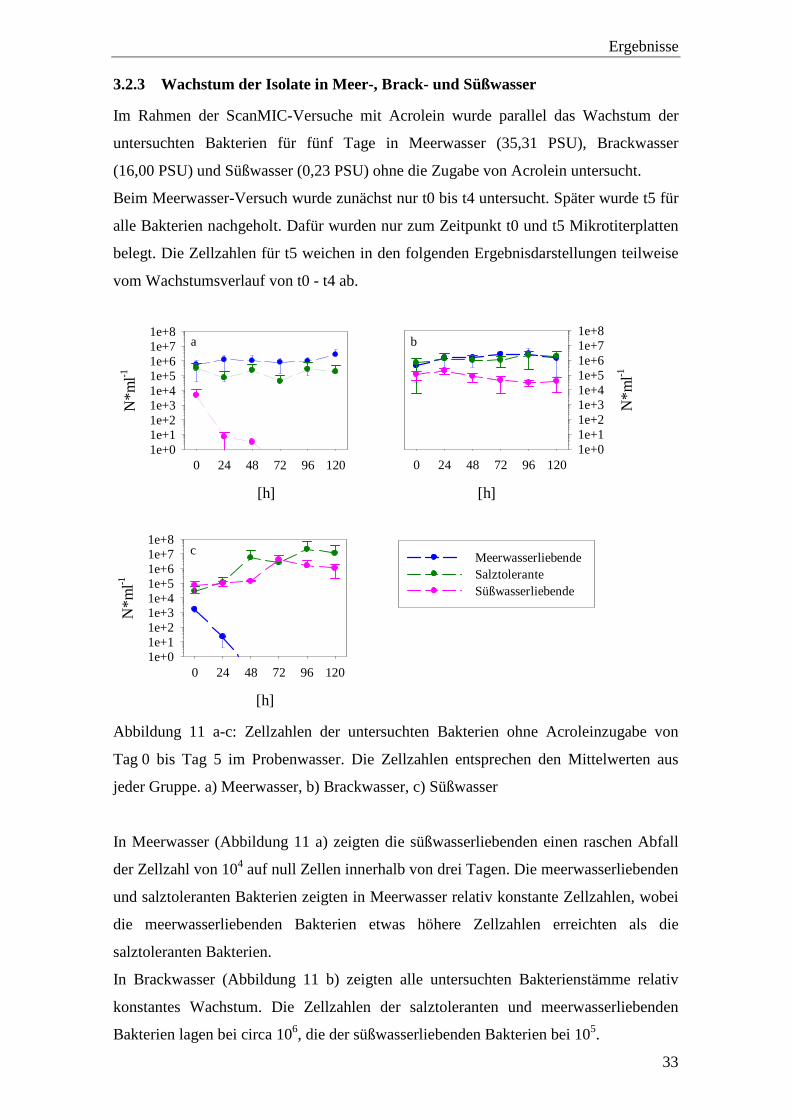
3.2.3 Wachstum der Isolate in Meer-, Brack- und Süßwasser
Im Rahmen der ScanMIC-Versuche mit Acrolein wurde parallel das Wachstum der
untersuchten Bakterien für fünf Tage in Meerwasser (35,31 PSU), Brackwasser
(16,00 PSU) und Süßwasser (0,23 PSU) ohne die Zugabe von Acrolein untersucht.
Beim Meerwasser-Versuch wurde zunächst nur t0 bis t4 untersucht. Später wurde t5 für
alle Bakterien nachgeholt. Dafür wurden nur zum Zeitpunkt t0 und t5 Mikrotiterplatten
belegt. Die Zellzahlen für t5 weichen in den folgenden Ergebnisdarstellungen teilweise
vom Wachstumsverlauf von t0 - t4 ab.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
Meerwasserliebende Salztolerante Süßwasserliebende
Abbildung 11 a-c: Zellzahlen der untersuchten Bakterien ohne Acroleinzugabe von
Tag 0 bis Tag 5 im Probenwasser. Die Zellzahlen entsprechen den Mittelwerten aus
jeder Gruppe. a) Meerwasser, b) Brackwasser, c) Süßwasser
In Meerwasser (Abbildung 11 a) zeigten die süßwasserliebenden einen raschen Abfall
der Zellzahl von 104 auf null Zellen innerhalb von drei Tagen. Die meerwasserliebenden
und salztoleranten Bakterien zeigten in Meerwasser relativ konstante Zellzahlen, wobei
die meerwasserliebenden Bakterien etwas höhere Zellzahlen erreichten als die
salztoleranten Bakterien.
In Brackwasser (Abbildung 11 b) zeigten alle untersuchten Bakterienstämme relativ
konstantes Wachstum. Die Zellzahlen der salztoleranten und meerwasserliebenden
Bakterien lagen bei circa 106, die der süßwasserliebenden Bakterien bei 105.
Ergebnisse
34
In Süßwasser (Abbildung 11 c) begann das Wachstum der süßwasserliebenden und der
salztoleranten Bakterien bei 104 – 105 N*ml -1 und stieg auf 106 bzw. 107 N*ml -1. Die
Zellzahlen der meerwasserliebenden Bakterien sanken innerhalb von 48 Stunden von
103 auf Null.
Innerhalb der Gruppen der meerwasserliebenden und der süßwasserliebenden Bakterien
gab es keine Abweichungen der Zellzahlen der Bakterienstämme untereinander. Die
Gruppe der salztoleranten Bakterien dagegen zeigte sich sehr divers (Abbildung 12 a-c).
Abbildung 12a-c: Zellzahlen innerhalb der Gruppe salztolerante Bakterien ohne
Acroleinzugabe. a) Meerwasser, b) Brackwasser, c) Süßwasser
Die Bakterienstämme der salztoleranten Bakterien wiesen in Brackwasser ähnliche und
konstante Zellzahlen auf. Nur das Umweltisolat dB2 zeigte mit 102 N*ml -1 deutlich
niedrigere Zellzahlen als die anderen Bakterienstämme dieser Gruppe mit circa
106 N*ml -1. Auch in Meer- und Süßwasser wies das Umweltisolat dB2 die niedrigsten
Zellzahlen auf, die von 10 bis 103 N*ml -1 reichen. V. vulnificus DSM 11507 begann
sowohl in Meer- als auch Süßwasser mit Zellzahlen von 103 N*ml -1 und erreichte
innerhalb von fünf Tagen Zellzahlen von 106 N*ml -1 in Meerwasser und 108 N*ml -1 in
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+81e+9
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+81e+9
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+81e+9
E. coliP. aeruginosaS. liquefaciensV. vulnificus DSM 11507d B2
Ergebnisse
35
Süßwasser. E. coli zeigte in Meer- und in Süßwasser zunächst eine Verringerung der
Zellzahlen, nach drei bis vier Tagen stiegen die Zellzahlen wieder auf den
ursprünglichen Wert von etwa 106 N*ml -1 an. S. liquefaciens zeigte in Süßwasser,
P. aeruginosa in Meerwasser sehr konstantes Wachstum bei circa 106 N*ml -1.
Umgekehrt schwankten die Zellzahlen bei S. liquefaciens in Meerwasser und bei
P. aeruginosa in Süßwasser, bewegten sich dennoch auf hohem Niveau.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+81e+9
MeerwasserBrackwasserSüßwasser
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+81e+9
MeerwasserBrackwasserSüßwasser
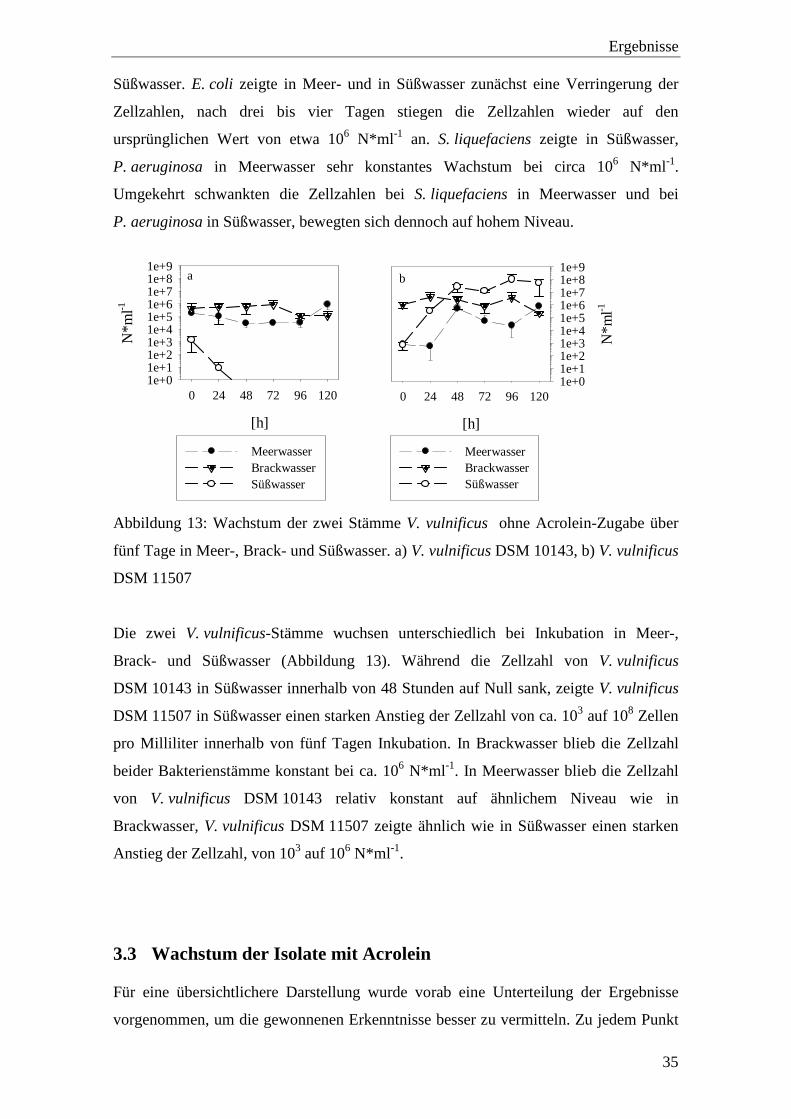
Abbildung 13: Wachstum der zwei Stämme V. vulnificus ohne Acrolein-Zugabe über
fünf Tage in Meer-, Brack- und Süßwasser. a) V. vulnificus DSM 10143, b) V. vulnificus
DSM 11507
Die zwei V. vulnificus-Stämme wuchsen unterschiedlich bei Inkubation in Meer-,
Brack- und Süßwasser (Abbildung 13). Während die Zellzahl von V. vulnificus
DSM 10143 in Süßwasser innerhalb von 48 Stunden auf Null sank, zeigte V. vulnificus
DSM 11507 in Süßwasser einen starken Anstieg der Zellzahl von ca. 103 auf 108 Zellen
pro Milliliter innerhalb von fünf Tagen Inkubation. In Brackwasser blieb die Zellzahl
beider Bakterienstämme konstant bei ca. 106 N*ml -1. In Meerwasser blieb die Zellzahl
von V. vulnificus DSM 10143 relativ konstant auf ähnlichem Niveau wie in
Brackwasser, V. vulnificus DSM 11507 zeigte ähnlich wie in Süßwasser einen starken
Anstieg der Zellzahl, von 103 auf 106 N*ml -1.
3.3 Wachstum der Isolate mit Acrolein
Für eine übersichtlichere Darstellung wurde vorab eine Unterteilung der Ergebnisse
vorgenommen, um die gewonnenen Erkenntnisse besser zu vermitteln. Zu jedem Punkt
Ergebnisse
36
wurden zunächst die Ergebnisse der Bakteriengruppen untereinander verglichen,
anschließend die Ergebnisse innerhalb der Bakteriengruppen dargestellt.
Im Hauptversuch wurden verschiedene Konzentrationen an Acrolein eingesetzt. Im
Folgenden wird zum einen die Wirkung der niedrigen Konzentrationen 0.01 und
0.1 mg/L Acrolein und zum anderen die Wirkung der hohen Konzentrationen 5-
15 mg/L Acrolein dargestellt. Die Acrolein-Konzentration 1 mg/L war der Grenzwert,
ab dem eine biozide Wirkung zu erkennen war, die aber nicht sofort zum Tod der
untersuchten Bakterien führte.
Die meerwasserliebenden Bakterien zeigten kein oder nur minimales Wachstum in
Süßwasser, ebenso zeigten die süßwasserliebenden Bakterien kein oder nur minimales
Wachstum in Meerwasser. Das gilt für alle untersuchten Acrolein-Konzentrationen, wie
Tabelle 6 verdeutlicht, und wird nicht mehr explizit erwähnt.
Tabelle 6: Darstellung von je einem Isolat pro Gruppe nach 24 h Inkubation in Meer-,
Brack- und Süßwasser. Angaben in N*ml-1.
Isolat Acrolein-
Konzentration
Meerwasser Brackwasser Süßwasser
E. coli 0 41333 600000 5667
0.01 241667 1450000 13500
0.1 73333 1100000 600
1 86 253333 8
5 8 20 0
10 0 0 0
15 0 0 0
V. parahaemolyticus 0 1066667 2500000 41
0.01 1916667 2366667 1
0.1 1866667 3366667 0
1 8 168 0
5 0 0 0
10 0 0 0
15 0 0 0
E. faecalis 0 11 135000 67500
0.01 13 88333 280000
0.1 20 203333 147500
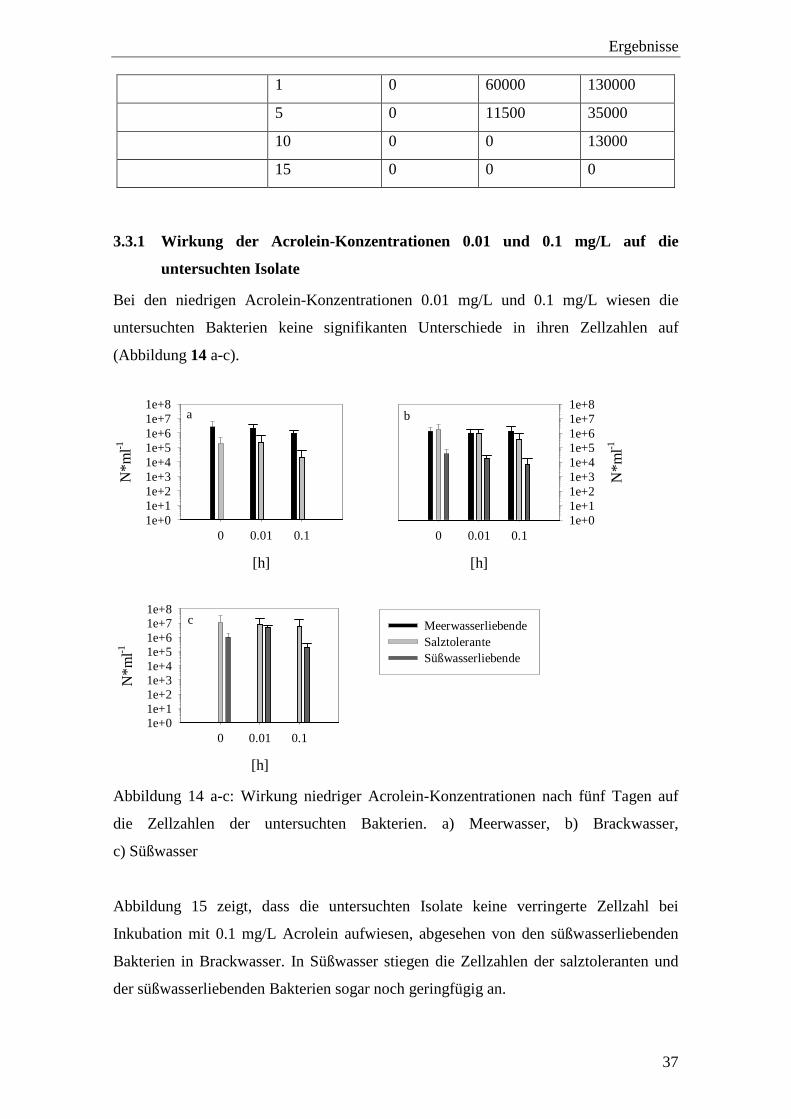
Ergebnisse
37
1 0 60000 130000
5 0 11500 35000
10 0 0 13000
15 0 0 0
3.3.1 Wirkung der Acrolein-Konzentrationen 0.01 und 0.1 mg/L auf die
untersuchten Isolate
Bei den niedrigen Acrolein-Konzentrationen 0.01 mg/L und 0.1 mg/L wiesen die
untersuchten Bakterien keine signifikanten Unterschiede in ihren Zellzahlen auf
(Abbildung 14 a-c).
a
[h]
0 0.01 0.1
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
b
[h]
0 0.01 0.1
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
c
[h]
0 0.01 0.1
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
Meerwasserliebende Salztolerante Süßwasserliebende
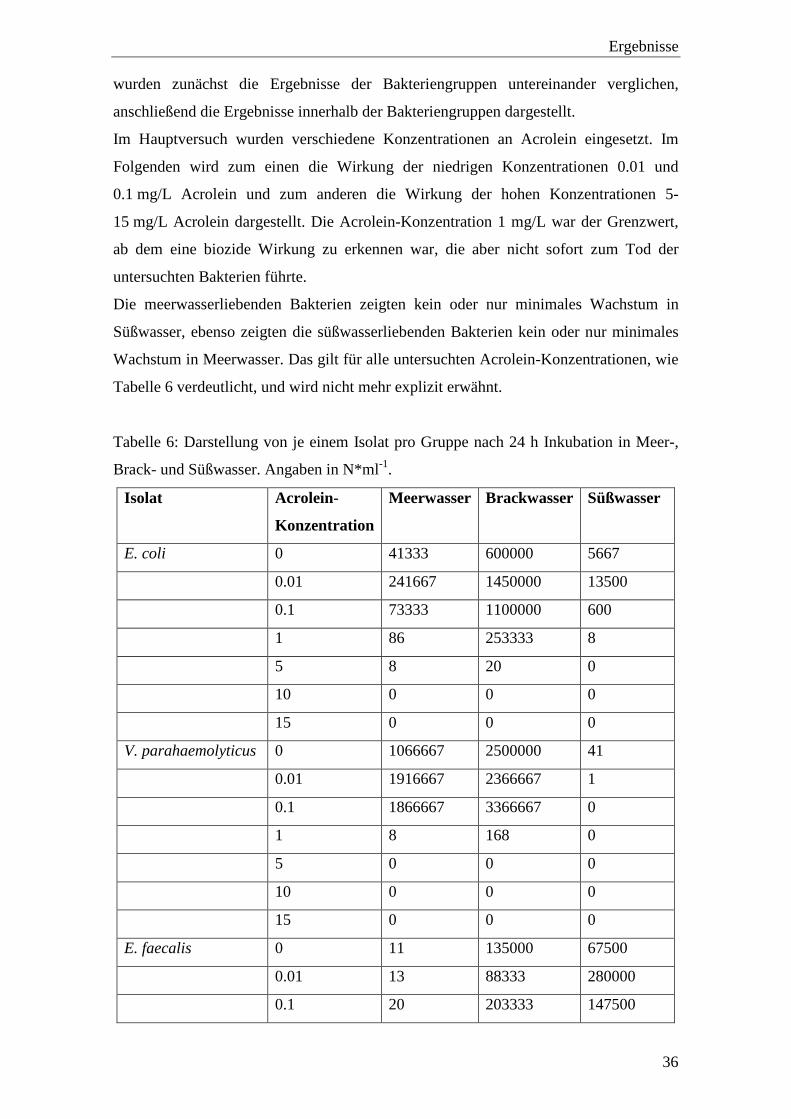
Abbildung 14 a-c: Wirkung niedriger Acrolein-Konzentrationen nach fünf Tagen auf
die Zellzahlen der untersuchten Bakterien. a) Meerwasser, b) Brackwasser,
c) Süßwasser
Abbildung 15 zeigt, dass die untersuchten Isolate keine verringerte Zellzahl bei
Inkubation mit 0.1 mg/L Acrolein aufwiesen, abgesehen von den süßwasserliebenden
Bakterien in Brackwasser. In Süßwasser stiegen die Zellzahlen der salztoleranten und
der süßwasserliebenden Bakterien sogar noch geringfügig an.
Ergebnisse
38
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
Meerwasserliebende Salztolerante Süßwasserliebende
Abbildung 15 a-c: Reaktion der Bakterien-Gruppen auf 0.1 mg/L Acrolein innerhalb
von fünf Tagen Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser
Teilweise waren die Zellzahlen nach 24 Stunden bei 0.01 mg/L Acrolein leicht erhöht
gegenüber 0 mg/L und 0.1 mg/L Acrolein (Abbildung 16). Der Unterschied ist nur
minimal und tritt nicht bei allen untersuchten Bakterien auf. Aufgefallen war die
erhöhte Zellzahl bei niedrigen Konzentrationen hauptsächlich bei E. coli, P. aeruginosa
und S. liquefaciens nach 24 h Inkubation in Meerwasser.
[h]
0 0.01
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
Meerwasserliebende Salztolerante Süßwasserliebende
Acrolein [mg/L]
0 0.01
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
E. coliP. aeruginosaS. liquefaciens
Abbildung 16: Erhöhte Zellzahlen bei 0.01 mg/L nach 24 h Inkubation in Meerwasser

Ergebnisse
39
Der Vergleich der beiden Stämme V. vulnificus DSM 10143 und DSM 11507 bei
Inkubation mit 0.1 mg/L Acrolein in Meer-, Brack- und Süßwasser (Abbildung 17)
zeigt erneut, wie unterschiedlich beide Stämme auf die verschiedenen
Salzkonzentrationen reagieren. In Brackwasser sinken die Zellzahlen von V. vulnificus
DSM 10143 vier Tage lang und steigen an Tag 5 wieder an. In Meerwasser zeigte das
Isolat keine Reaktion auf die Inkubation mit 0.1 mg/L Acrolein. V. vulnificus DSM
11507 zeigte keine Reaktion auf die Inkubation mit 0.1 mg/L Acrolein.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
MeerwasserBrackwasserSüßwasser
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+71e+8
MeerwasserBrackwasserSüßwasser
Abbildung 17: V. vulnificus DSM 10143 (a) und V. vulnificus DSM 11507 (b) mit
0.1 mg/L Acrolein
3.3.2 Wirkung der Acrolein-Konzentration 1 mg/L auf die untersuchten Isolate
Ab 1 mg/L Acrolein sanken schon nach 24 Stunden die Zellzahlen der drei
Bakteriengruppen. Im Verlauf von fünf Tagen Inkubation kam es bei den meisten der
untersuchten Bakterien zu einem stetigen Rückgang der Zellzahlen.
Ergebnisse
40
Mee
rwa
sser
liebe
nde
Sa
lzto
lera
nte
Süß
wa
sser
liebe
nde
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
1e+6Meerwasser Brackwasser Süßwasser
Abbildung 18: Reaktion der Bakterien-Gruppen auf 1 mg/L Acrolein nach 24 h
Inkubation in Meer-, Brack- und Süßwasser
Abbildung 18 gibt die Zellzahlen innerhalb der Bakteriengruppen in den verschiedenen
Salzkonzentrationen wider. Innerhalb der einzelnen Gruppen variierte das Wachstum
der Bakterien bei 1 mg/L Acrolein stark. Salztolerante und süßwasserliebende Bakterien
zeigten die höchsten Zellzahlen in Süßwasser. Die meerwasserliebenden Bakterien
zeigten in Meerwasser unvermindertes und in Brackwasser geringes Wachstum.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
Meerwasserliebende Salztolerante Süßwasserliebende
Abbildung 19 a-c: Reaktion der Bakterien-Gruppen auf 1 mg/L Acrolein innerhalb von
5 Tagen Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser
Ergebnisse
41
Abbildung 19 zeigt die Zellzahlen innerhalb der Bakteriengruppen im Verlauf von t0
bis t5. Die süßwasserliebenden Bakterien zeigten abgesehen von der Inkubation in
Meerwasser den geringsten Abfall der Zellzahlen bei Inkubation mit 1 mg/L Acrolein.
Abbildung 20 a-c: Zellzahlen innerhalb der Bakteriengruppen nach 24 Stunden
Inkubation bei 1 mg/L Acrolein. a) Salztolerante Bakterien, b) Meerwasserliebende
Bakterien, c) Süßwasserliebende Bakterien
Abbildung 20 macht deutlich, dass die Reaktion der untersuchten Bakterien auf 1 mg/L
Acrolein vom Probenwassertyp abhängt. In Probenwasser mit optimalem Salzgehalt
zeigten die untersuchten Bakterien höhere Zellzahlen als in Probenwasser mit weniger
optimalem Salzgehalt.
aE
. co
li
P.
ae
rug
ino
sa
S.
liqu
efa
cie
ns
V.
vuln
ificu
s 1
15
07
d B
2
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
Meerwasser Brackwasser Süßwasser
c
V.
para
ha
em
oly
ticus
V.
alg
ino
lytic
us
V.
vuln
ificu
s 1
01
43
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
E.
fae
calis
A.
hyd
rop
hila
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
Ergebnisse
42
Während der fünftägigen Inkubation sanken die Zellzahlen einiger der untersuchten
Bakterien weiter ab, während andere konstant unbeeinträchtig blieben (Abbildung 21 –
Abbildung 23).
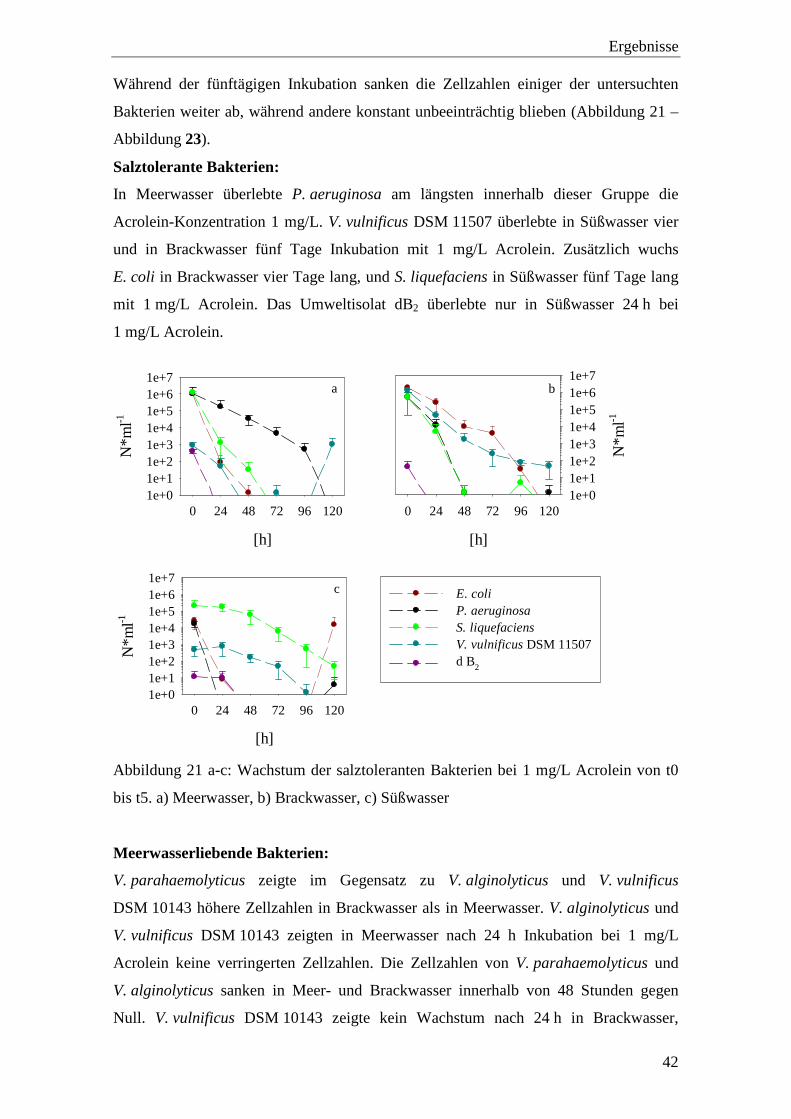
Salztolerante Bakterien:
In Meerwasser überlebte P. aeruginosa am längsten innerhalb dieser Gruppe die
Acrolein-Konzentration 1 mg/L. V. vulnificus DSM 11507 überlebte in Süßwasser vier
und in Brackwasser fünf Tage Inkubation mit 1 mg/L Acrolein. Zusätzlich wuchs
E. coli in Brackwasser vier Tage lang, und S. liquefaciens in Süßwasser fünf Tage lang
mit 1 mg/L Acrolein. Das Umweltisolat dB2 überlebte nur in Süßwasser 24 h bei
1 mg/L Acrolein.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
E. coliP. aeruginosaS. liquefaciensV. vulnificus DSM 11507d B2
Abbildung 21 a-c: Wachstum der salztoleranten Bakterien bei 1 mg/L Acrolein von t0
bis t5. a) Meerwasser, b) Brackwasser, c) Süßwasser
Meerwasserliebende Bakterien:
V. parahaemolyticus zeigte im Gegensatz zu V. alginolyticus und V. vulnificus
DSM 10143 höhere Zellzahlen in Brackwasser als in Meerwasser. V. alginolyticus und
V. vulnificus DSM 10143 zeigten in Meerwasser nach 24 h Inkubation bei 1 mg/L
Acrolein keine verringerten Zellzahlen. Die Zellzahlen von V. parahaemolyticus und
V. alginolyticus sanken in Meer- und Brackwasser innerhalb von 48 Stunden gegen
Null. V. vulnificus DSM 10143 zeigte kein Wachstum nach 24 h in Brackwasser,
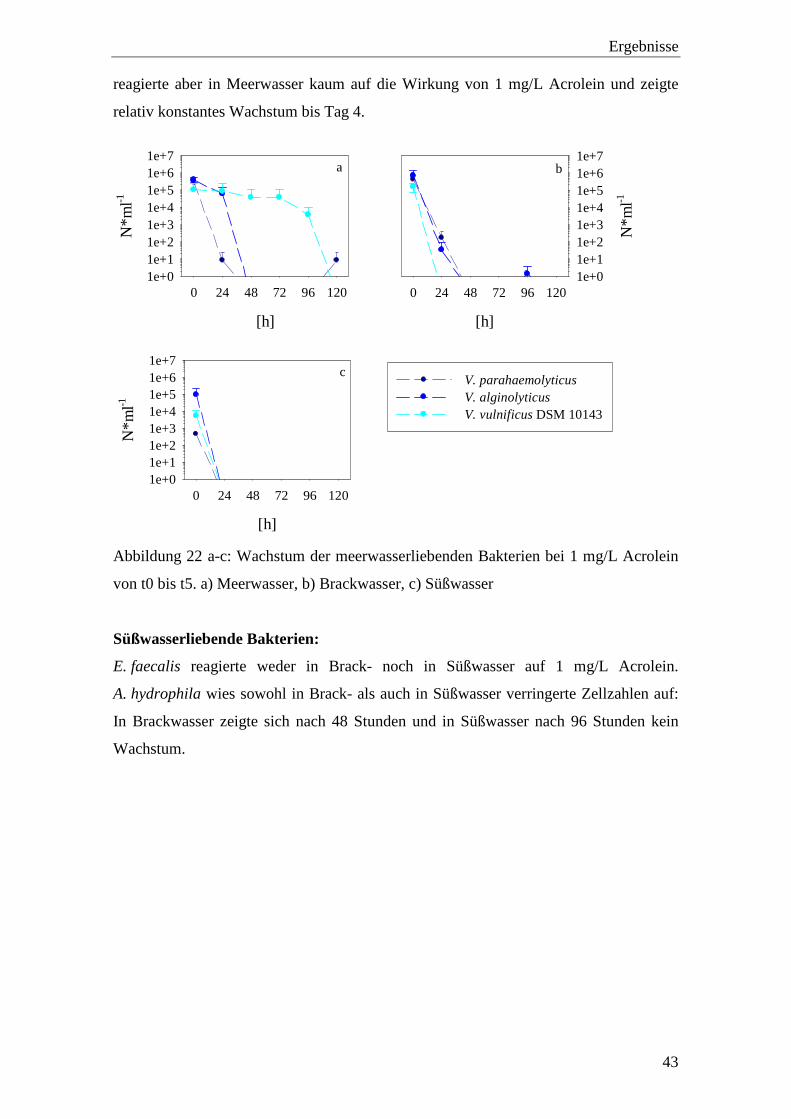
Ergebnisse
43
reagierte aber in Meerwasser kaum auf die Wirkung von 1 mg/L Acrolein und zeigte
relativ konstantes Wachstum bis Tag 4.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
V. parahaemolyticusV. alginolyticusV. vulnificus DSM 10143
Abbildung 22 a-c: Wachstum der meerwasserliebenden Bakterien bei 1 mg/L Acrolein
von t0 bis t5. a) Meerwasser, b) Brackwasser, c) Süßwasser
Süßwasserliebende Bakterien:
E. faecalis reagierte weder in Brack- noch in Süßwasser auf 1 mg/L Acrolein.
A. hydrophila wies sowohl in Brack- als auch in Süßwasser verringerte Zellzahlen auf:
In Brackwasser zeigte sich nach 48 Stunden und in Süßwasser nach 96 Stunden kein
Wachstum.
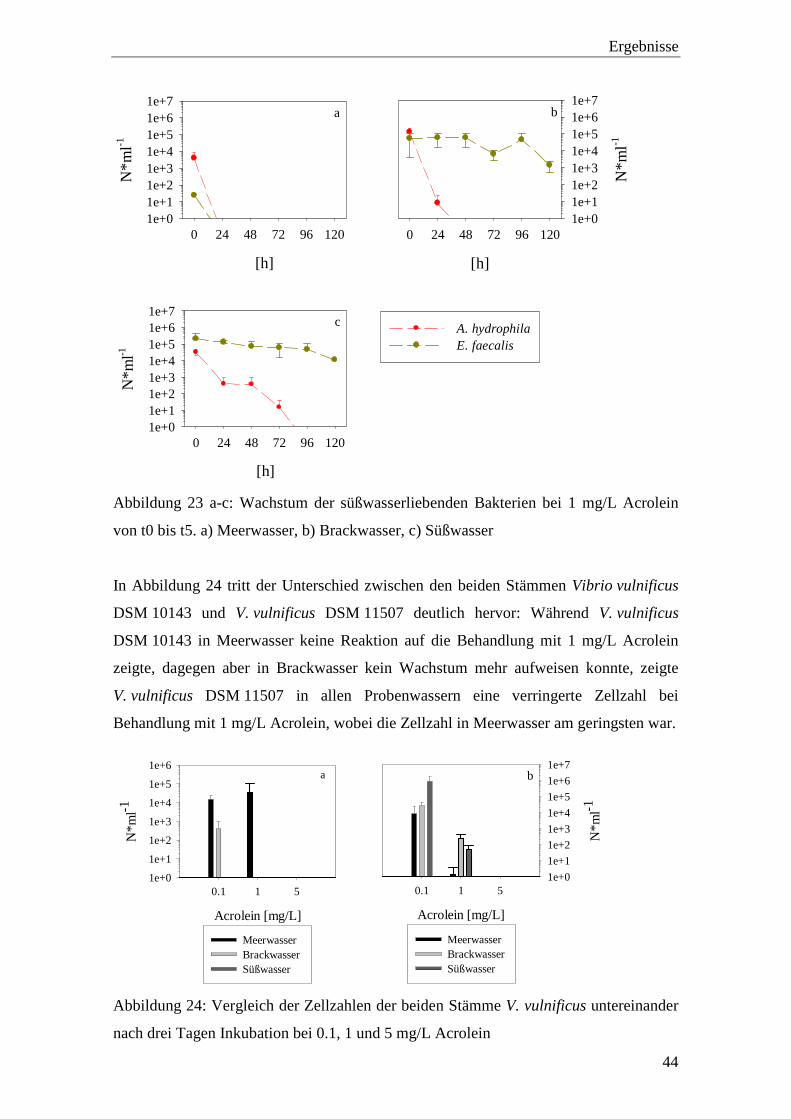
Ergebnisse
44
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
A. hydrophilaE. faecalis
Abbildung 23 a-c: Wachstum der süßwasserliebenden Bakterien bei 1 mg/L Acrolein
von t0 bis t5. a) Meerwasser, b) Brackwasser, c) Süßwasser
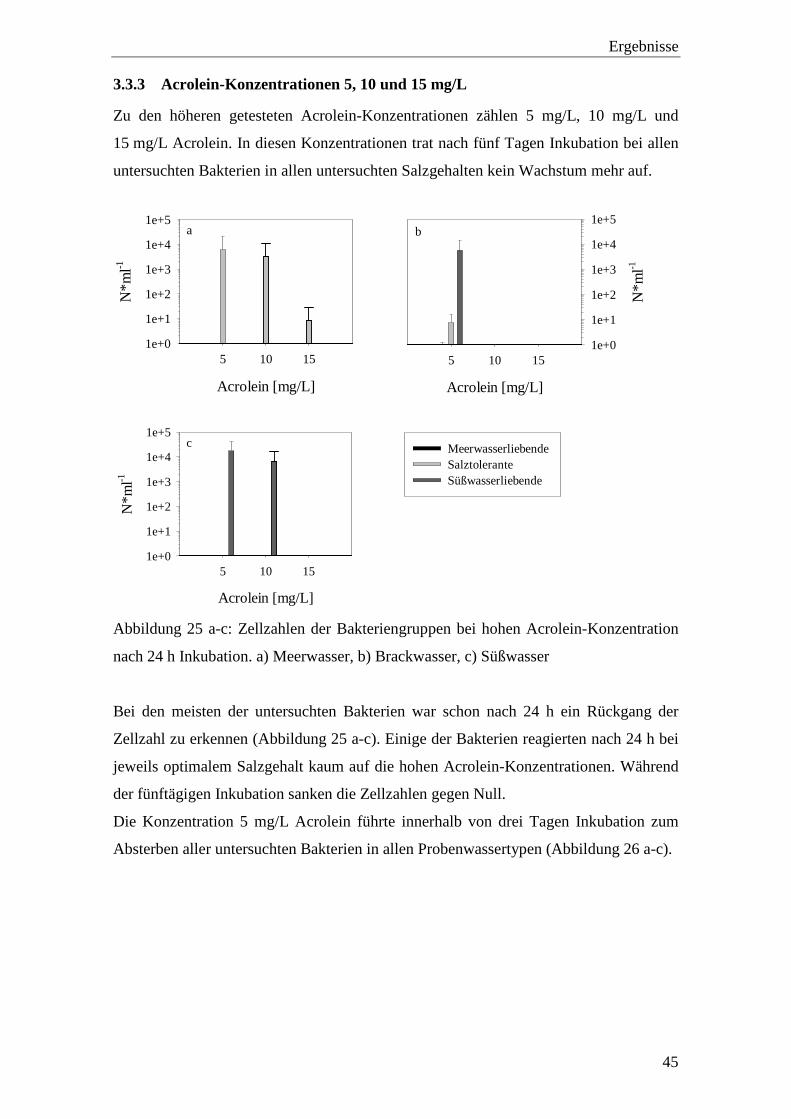
In Abbildung 24 tritt der Unterschied zwischen den beiden Stämmen Vibrio vulnificus
DSM 10143 und V. vulnificus DSM 11507 deutlich hervor: Während V. vulnificus
DSM 10143 in Meerwasser keine Reaktion auf die Behandlung mit 1 mg/L Acrolein
zeigte, dagegen aber in Brackwasser kein Wachstum mehr aufweisen konnte, zeigte
V. vulnificus DSM 11507 in allen Probenwassern eine verringerte Zellzahl bei
Behandlung mit 1 mg/L Acrolein, wobei die Zellzahl in Meerwasser am geringsten war.
a
Acrolein [mg/L]
0.1 1 5
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
1e+6
Meerwasser Brackwasser Süßwasser
b
Acrolein [mg/L]
0.1 1 5
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
1e+6
1e+7
Meerwasser Brackwasser Süßwasser
Abbildung 24: Vergleich der Zellzahlen der beiden Stämme V. vulnificus untereinander
nach drei Tagen Inkubation bei 0.1, 1 und 5 mg/L Acrolein
Ergebnisse
45
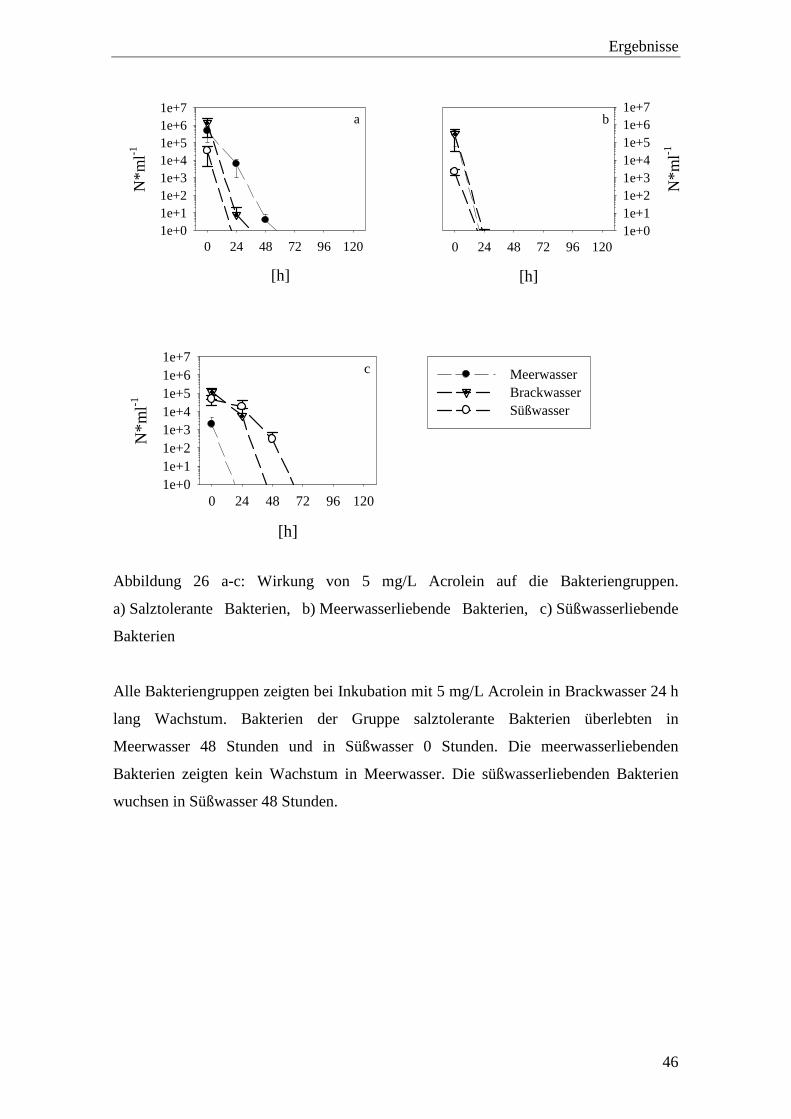
3.3.3 Acrolein-Konzentrationen 5, 10 und 15 mg/L
Zu den höheren getesteten Acrolein-Konzentrationen zählen 5 mg/L, 10 mg/L und
15 mg/L Acrolein. In diesen Konzentrationen trat nach fünf Tagen Inkubation bei allen
untersuchten Bakterien in allen untersuchten Salzgehalten kein Wachstum mehr auf.
a
Acrolein [mg/L]
5 10 15
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5b
Acrolein [mg/L]
5 10 15
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
c
Acrolein [mg/L]
5 10 15
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5Meerwasserliebende Salztolerante Süßwasserliebende
Abbildung 25 a-c: Zellzahlen der Bakteriengruppen bei hohen Acrolein-Konzentration
nach 24 h Inkubation. a) Meerwasser, b) Brackwasser, c) Süßwasser
Bei den meisten der untersuchten Bakterien war schon nach 24 h ein Rückgang der
Zellzahl zu erkennen (Abbildung 25 a-c). Einige der Bakterien reagierten nach 24 h bei
jeweils optimalem Salzgehalt kaum auf die hohen Acrolein-Konzentrationen. Während
der fünftägigen Inkubation sanken die Zellzahlen gegen Null.
Die Konzentration 5 mg/L Acrolein führte innerhalb von drei Tagen Inkubation zum
Absterben aller untersuchten Bakterien in allen Probenwassertypen (Abbildung 26 a-c).
Ergebnisse
46
Abbildung 26 a-c: Wirkung von 5 mg/L Acrolein auf die Bakteriengruppen.
a) Salztolerante Bakterien, b) Meerwasserliebende Bakterien, c) Süßwasserliebende
Bakterien
Alle Bakteriengruppen zeigten bei Inkubation mit 5 mg/L Acrolein in Brackwasser 24 h
lang Wachstum. Bakterien der Gruppe salztolerante Bakterien überlebten in
Meerwasser 48 Stunden und in Süßwasser 0 Stunden. Die meerwasserliebenden
Bakterien zeigten kein Wachstum in Meerwasser. Die süßwasserliebenden Bakterien
wuchsen in Süßwasser 48 Stunden.
a
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
b
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
c
[h]
0 24 48 72 96 120
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
MeerwasserBrackwasserSüßwasser
Ergebnisse
47
a
E.
coli
E.
fae
calis
P.
ae
rugin
osa
A.
hyd
rop
hila
S.
liqu
efa
cie
ns
V.
para
ha
em
oly
ticu
sV
. a
lgin
oly
ticu
sV
. vu
lnifi
cus
101
43
V.
vuln
ificu
s 1
15
07
d B
2
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5b
E.
coli
E.
fae
calis
P.
ae
rugin
osa
A.
hyd
rop
hila
S.
liqu
efa
cie
ns
V.
para
ha
em
oly
ticu
sV
. a
lgin
oly
ticu
sV
. vu
lnifi
cus
101
43
V.
vuln
ificu
s 1
15
07
d B
2
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
c
E.
coli
E.
fae
calis
P.
ae
rug
ino
saA
. h
ydro
ph
ilaS
. liq
ue
faci
en
sV
. p
ara
ha
em
oly
ticu
sV
. a
lgin
oly
ticu
sV
. vu
lnifi
cus
10
14
3V
. vu
lnifi
cus
11
50
7d
B2
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
Abbildung 27 a-c: Zellzahlen der untersuchten Bakterien nach 24 Stunden Inkubation in
5 mg/L Acrolein. a) Meerwasser, b) Brackwasser, c) Süßwasser
Wie Abbildung 27 a-c zeigt überlebten nur einzelne Bakterien aus den Gruppen die
Inkubation bei 5 mg/L Acrolein. In Süßwasser überlebte ausschließlich E. faecalis, in
Meerwasser die drei Bakterienstämme E. coli, P. aeruginosa und S. liquefaciens. In
Brackwasser zeigten fünf der untersuchten Mikroorganismen Wachstum nach
24 Stunden in 5 mg/L Acrolein, und zwar E. coli, E. faecalis, S. liquefaciens,
V. alginolyticus und V. vulnificus DSM 11507.
P. aeruginosa (in Meerwasser) und E. faecalis (in Süßwasser) sind die einzigen
Bakterien, die die Acrolein-Konzentration 10 mg/L zum Zeitpunkt t1 überlebten.
Nach 24 h Inkubation mit 15 mg/L Acrolein trat bei fast keinem der untersuchten
Bakterienstämme Wachstum auf (Abbildung 28).

Ergebnisse
48
[h]
0 24 48 72 96 120
N*m
l-1
1e+0
1e+1
1e+2
1e+3
1e+4
1e+5
1e+6
1e+7Meerwasserliebende (Meerwasser)Salztolerante (Meerwasser)Süßwasserliebende (Meerwasser)Meerwasserliebende (Brackwasser)Salztolerante (Brackwasser)Süßwasserliebende (Brackwasser)Meerwasserliebende (Süßwasser)Salztolerante (Süßwasser)Süßwasserliebende (Süßwasser)
Abbildung 28: Wirkung von 15 mg/L Acrolein auf die Bakteriengruppen innerhalb von
5 Tagen.
P. aeruginosa überlebte nach 24 h in Meerwasser sogar die höchste Konzentration von
15 mg/L (Abbildung 29).
Acrolein [mg/L]
0 0.010.1 1 5 10 15
N*m
l-1
1e+01e+11e+21e+31e+41e+51e+61e+7
P. aeruginosa
Abbildung 29: P. aeruginosa nach 24 h in Meerwasser zeigte Wachstum bei allen
untersuchten Acrolein-Konzentrationen
Die Wirkung der getesteten Acrolein-Konzentration auf die einzelnen Bakterien unter
Berücksichtigung der unterschiedlichen Salzgehalte wurde in Abbildung 30 und
Abbildung 31 zusammen gefasst. In diesen Abbildungen wird gezeigt, welche
Konzentration bei welchem Salzgehalt zu keinem Wachstum der untersuchten Bakterien
führte.
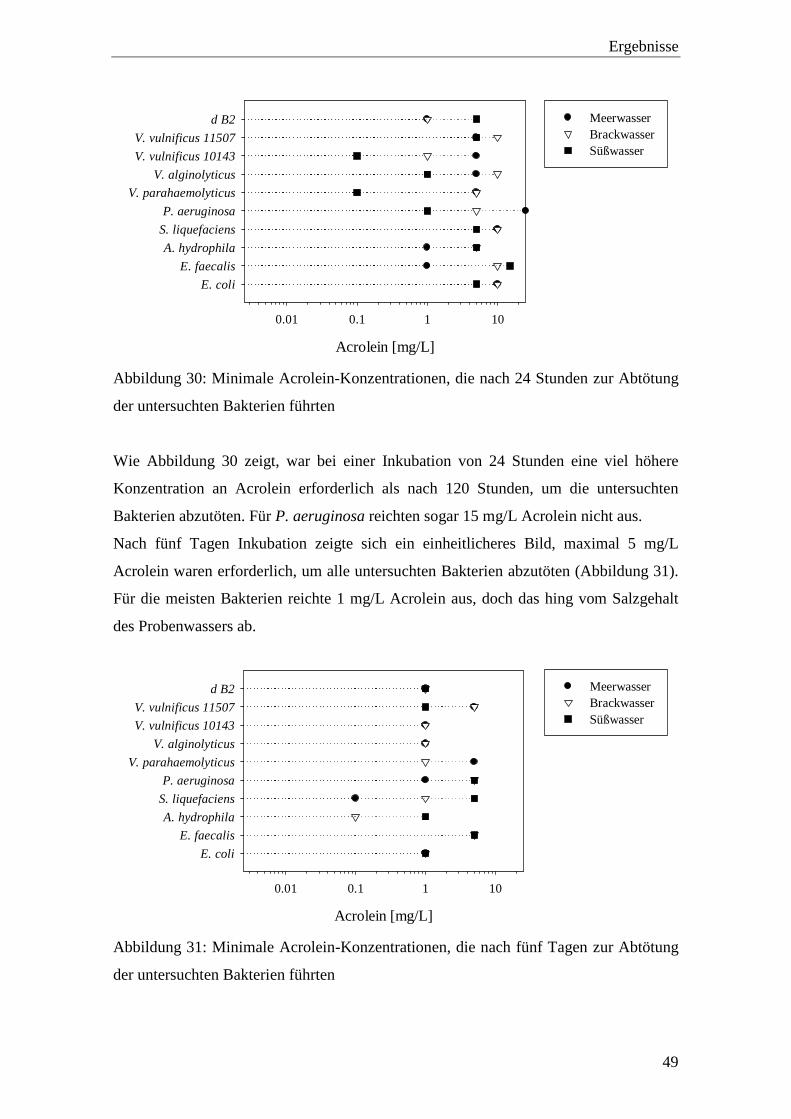
Ergebnisse
49
Acrolein [mg/L]
0.01 0.1 1 10
E. coli
E. faecalis
A. hydrophila
S. liquefaciens
P. aeruginosa
V. parahaemolyticus
V. alginolyticus
V. vulnificus 10143
V. vulnificus 11507
d B2 MeerwasserBrackwasser Süßwasser
Abbildung 30: Minimale Acrolein-Konzentrationen, die nach 24 Stunden zur Abtötung
der untersuchten Bakterien führten
Wie Abbildung 30 zeigt, war bei einer Inkubation von 24 Stunden eine viel höhere
Konzentration an Acrolein erforderlich als nach 120 Stunden, um die untersuchten
Bakterien abzutöten. Für P. aeruginosa reichten sogar 15 mg/L Acrolein nicht aus.
Nach fünf Tagen Inkubation zeigte sich ein einheitlicheres Bild, maximal 5 mg/L
Acrolein waren erforderlich, um alle untersuchten Bakterien abzutöten (Abbildung 31).
Für die meisten Bakterien reichte 1 mg/L Acrolein aus, doch das hing vom Salzgehalt
des Probenwassers ab.
Acrolein [mg/L]
0.01 0.1 1 10
E. coli
E. faecalis
A. hydrophila
S. liquefaciens
P. aeruginosa
V. parahaemolyticus
V. alginolyticus
V. vulnificus 10143
V. vulnificus 11507
d B2 Meerwasser Brackwasser Süßwasser
Abbildung 31: Minimale Acrolein-Konzentrationen, die nach fünf Tagen zur Abtötung
der untersuchten Bakterien führten

Diskussion
50
4 Diskussion
Hintergrund und Motivation dieser Diplomarbeit ist die Problematik der Verschleppung
von Organismen über das Ballastwasser in Schiffen. Besonders Mikroorganismen sind
in Ballastwasser in hoher Abundanz vorhanden und stellen ein Risiko für die
Verbreitung von Krankheitserregern, die in Küstengewässern vorkommen, dar
(Aridgides et al, 2003; Ruiz et al, 2000). Die Richtlinien der IMO verlangen, dass bei
der Abgabe von Ballastwasser pro 100 ml weniger als 1 cfu (colony forming unit)
Vibrio cholerae, weniger als 100 cfu intestinale Enterokokken und weniger als 250 cfu
Escherichia coli vorhanden sein dürfen. In dieser Arbeit wurde zwar nicht V. cholerae
untersucht, dafür aber drei Arten der gleichen Familie, die ebenfalls große
humanpathogene Bedeutung haben. Als Beispiel für intestinale Enterokokken wurde
E. faecalis untersucht. Ein Stamm von E. coli wurde hier ebenfalls untersucht.
Eine der führenden Methoden um die Einführung gebietsfremder Organismen über
Ballastwasser in Schiffen zu verhindern ist, Ballastwasser auf hoher See auszutauschen.
Mit dieser Methode können 90-95% des Ballastwassers ausgetauscht werden (Bai et al,
2004). Zusätzlich hilft die Behandlung des Ballastwassers mit Chemikalien, den Eintrag
von Organismen zu verhindern. In dieser Arbeit wurde untersucht, welche Wirkung das
Biozid Acrolein auf verschiedene pathogene Bakterien hat.
In diesem Abschnitt der Diplomarbeit werden die gewonnen Ergebnisse diskutiert.
Dabei werden zunächst die Ergebnisse ohne Acrolein-Beteiligung dargestellt und
anschließend die Wirkung von Acrolein auf die untersuchten Isolate diskutiert.
Anschließend wird die Wirkung von Acrolein mit der Wirkung anderer Ballastwasser-
Biozide auf Bakterien verglichen. Zuletzt wird die angewandte Methode evaluiert.
4.1 Bakterienstämme
Mit den erworbenen Isolaten wurden im Vorfeld zu den Acrolein-Versuchen
Vorversuche durchgeführt. In diesen Vorversuchen wurde überprüft, welche
Wachstumsgeschwindigkeit die untersuchten Isolate in ihrem Optimalmedium besitzen,

Diskussion
51
welche Koloniemorphologien bei Kultivierung auf Agarplatten auftreten, wie die
Isolate auf die Färbung mit TTC reagieren und wie sich das Wachstum der Isolate bei
Inkubation in Meer-, Brack- und Süßwasser entwickelt.
4.1.1 Wachstum der Isolate in Medium
In ihrem jeweiligen Optimalmedium zeigten fast alle untersuchten pathogenen
Bakterien einen sofortigen Übergang ihrer Wachstumskurve in die exponentielle Phase.
Unter den untersuchten Isolaten wuchsen die Vibrionen am schnellsten und
P. aeruginosa am langsamsten. Das Umweltisolat dB2 zeigte langsames Wachstum bei
18°C. Bei 30°C zeigte das Umweltisolat eine ähnliche Wachstumskurve wie die
pathogenen Bakterien.
Es besteht ein Zusammenhang zwischen der Morphologie der Bakterienkolonien in
Kultur und dem Phänotyp der Bakterien in den Kavitäten von Mikrotiterplatten nach
erfolgter TTC-Färbung.
TTC ist ein Tetrazoliumsalz, das durch Elektronenaufnahme aus oxidativen
Enzymsystemen zu Formazan reduziert wird. Das Formazan wird in Granula in den
Zellen eingelagert und färbt die Zellen rot (Beloti et al, 1999; Gabrielson et al, 2002).
TTC wurde schon vor mehr als hundert Jahren entwickelt (Tsukatani et al, 2008) und
wird seit den 1940ern als Wachstumsindikator eingesetzt (Gabrielson et al, 2002).
Im Ergebnisteil wurden Kolonie- und Pellet-Morphologien der untersuchten Isolate
beschrieben (siehe Punkt 3.1). In Tabelle 7 wurden Beweglichkeit, Koloniemorphologie
und Pelletbildung der untersuchten Isolate vergleichend nebeneinander dargestellt.
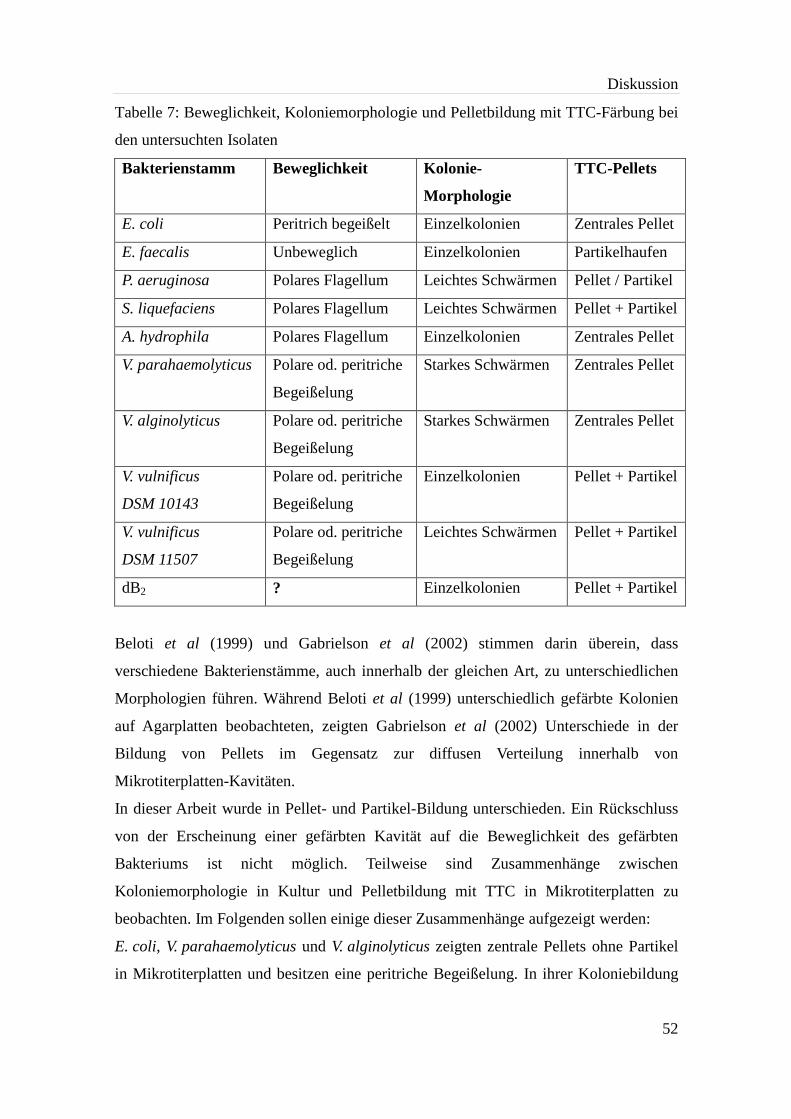
Diskussion
52
Tabelle 7: Beweglichkeit, Koloniemorphologie und Pelletbildung mit TTC-Färbung bei
den untersuchten Isolaten
Bakterienstamm Beweglichkeit Kolonie-
Morphologie
TTC-Pellets
E. coli Peritrich begeißelt Einzelkolonien Zentrales Pellet
E. faecalis Unbeweglich Einzelkolonien Partikelhaufen
P. aeruginosa Polares Flagellum Leichtes Schwärmen Pellet / Partikel
S. liquefaciens Polares Flagellum Leichtes Schwärmen Pellet + Partikel
A. hydrophila Polares Flagellum Einzelkolonien Zentrales Pellet
V. parahaemolyticus Polare od. peritriche
Begeißelung
Starkes Schwärmen Zentrales Pellet
V. alginolyticus Polare od. peritriche
Begeißelung
Starkes Schwärmen Zentrales Pellet
V. vulnificus
DSM 10143
Polare od. peritriche
Begeißelung
Einzelkolonien Pellet + Partikel
V. vulnificus
DSM 11507
Polare od. peritriche
Begeißelung
Leichtes Schwärmen Pellet + Partikel
dB2 ? Einzelkolonien Pellet + Partikel
Beloti et al (1999) und Gabrielson et al (2002) stimmen darin überein, dass
verschiedene Bakterienstämme, auch innerhalb der gleichen Art, zu unterschiedlichen
Morphologien führen. Während Beloti et al (1999) unterschiedlich gefärbte Kolonien
auf Agarplatten beobachteten, zeigten Gabrielson et al (2002) Unterschiede in der
Bildung von Pellets im Gegensatz zur diffusen Verteilung innerhalb von
Mikrotiterplatten-Kavitäten.
In dieser Arbeit wurde in Pellet- und Partikel-Bildung unterschieden. Ein Rückschluss
von der Erscheinung einer gefärbten Kavität auf die Beweglichkeit des gefärbten
Bakteriums ist nicht möglich. Teilweise sind Zusammenhänge zwischen
Koloniemorphologie in Kultur und Pelletbildung mit TTC in Mikrotiterplatten zu
beobachten. Im Folgenden sollen einige dieser Zusammenhänge aufgezeigt werden:
E. coli, V. parahaemolyticus und V. alginolyticus zeigten zentrale Pellets ohne Partikel
in Mikrotiterplatten und besitzen eine peritriche Begeißelung. In ihrer Koloniebildung

Diskussion
53
auf Agarplatten unterschieden sich die Einzelkolonien von E. coli von den stark
schwärmenden Vibrionen V. parahaemolyticus und V. alginolyticus.
P. aeruginosa, S. liquefaciens und A. hydrophila besitzen ein polares Flagellum. In
Koloniemorphologie und Pelletbildung in Mikrotiterplatten unterschieden sich
P. aeruginosa und S. liquefaciens von A. hydrophila. Die Isolate P. aeruginosa und
S. liquefaciens ähnelten sich in der Erscheinung sowohl auf Agarplatten als auch in
Mikrotiterplatten. Das Isolat A. hydrophila ähnelte in seiner Koloniemorphologie und
Pelletbildung eher E. coli.
Die Vibrionen V. vulnificus DSM 10143 und DSM 11507 unterschieden sich stark von
V. parahaemolyticus und V. alginolyticus und bildeten Partikel neben den zentralen
Pellets in den Mikrotiterplatten aus. Auf Agarplatten zeigte V. vulnificus DSM 10143
kein und V. vulnificus DSM 11507 nur leichtes Schwärmen.
Das unbewegliche Isolat E. faecalis bildete in Kultur kleine Einzelkolonien und mit
TTC diffuse Partikelhaufen, die sich vom Erscheinungsbild der anderen untersuchten
Isolate mit TTC unterschieden.
Die Beweglichkeit des Umweltisolats dB2 ist nicht bekannt. Mit TTC zeigt es eine
ähnliche Pellet- und Partikel-Bildung wie P. aeruginosa, doch bei der Kultivierung war
keinerlei Schwärmverhalten zu beobachten.
Die Untersuchungen von Gabrielson et al (2002) zeigen Mikrotiterplatten u.a. mit
Isolaten von A. hydrophila, E. coli, E. faecalis und V. alginolyticus, die mit 0.01% TTC
gefärbt wurden. Ein Vergleich anhand der dargestellten Mikrotiterplatten-Abbildungen
mit den in dieser Arbeit gewonnenen Abbildungen von gefärbten Isolaten in
Mikrotiterplatten-Kavitäten ist schwierig: Die Mikrotiterplatten wurden von Gabrielson
et al (2002) im reflective-Modus gescannt, sodass Lichtspiegelungen das Bild stören.
Außerdem sind keine vergrößerten Darstellungen der Kavitäten vorhanden. Dennoch
sind für diese Isolate ähnliche Pelletbildungen zu erkennen wie in dieser Arbeit.
A. hydrophila und E. coli scheinen klar abgegrenzte zentrale Pellets gebildet zu haben.
Die Pellets von A. hydrophila sind bei niedrigem chemischen Gradienten mit gefärbtem
Medium umgeben. E. faecalis scheint Partikelhaufen zu bilden, zeigt aber bei höherem
chemischen Gradient kleine zentrale Pellets. Derartige Pellets waren in dieser Arbeit
nur in Meerwasser bei E. faecalis zu beobachten, wo ohnehin kaum Wachstum

Diskussion
54
vorhanden war. V. alginolyticus bildet offenbar unscharf abgegrenzte zentrale Pellets,
die von gefärbtem Medium umgeben sind. In dieser Arbeit konnte ebenfalls vereinzelt
beobachtet werden, dass sich ein Iris-artiger gefärbter Kreis um das zentrale Pellet
herum bildete.
4.1.2 Probenwasser
Das Probenwasser wurde an drei für die Deutsche Bucht repräsentativen Bereichen
genommen: Einem Hochseehafen bei Helgoland, einem Hafengebiet im Ästuarbereich
Cuxhaven, und einem Süßwasserhafen in der weiteren Umgebung des internationalen
Hafens von Hamburg. Die Salz- und Nährstoffbedingungen des Probenwassers
entsprechen somit den natürlichen Zuständen, auf die durch Ballastwasseraustausch
verschleppte Organismen treffen würden.
Das Probenwasser aus Meer-, Brack- und Süßwasser unterschied sich hauptsächlich im
Salzgehalt. Da die untersuchten Bakterienstämme aus unterschiedlichen Lebensräumen
stammen, unterscheiden sie sich in ihrer Fähigkeit, sich an stark salzhaltige bzw. stark
salzarme Bedingungen anzupassen. Dementsprechend bildete in dieser Arbeit der
Salzgehalt neben der Acrolein-Wirkung den wichtigsten limitierenden Faktor auf die
Zellzahl der Bakterien. Der Salzgehalt für Meer-, Brack- und Süßwasser sollte bei 35,
15 und weniger als 1 PSU liegen. Das Ergebnis der Salzanalyse zeigte, dass die
Erwartungen bestätigt wurden.
4.1.3 Einteilung der untersuchten Bakterienstämme in Gruppen und
Wachstum der Isolate in Meer-, Brack- und Süßwasser
Das Wachstum der untersuchten Isolate in Meer-, Brack- und Süßwasser war die
Grundlage für die Aufteilung in verschiedene Bakterien-Gruppen. Die Aufteilung der
Bakterien in die Gruppen meerwasserliebende, süßwasserliebende und salztolerante
Bakterien wurde schon im Ergebnisteil beschrieben (Punkt 3.2.1).
Wachstum der salztoleranten Bakterien:
Zur Gruppe der salztoleranten Bakterien zählen E. coli, P. aeruginosa, S. liquefaciens,
V. vulnificus DSM 11507 und das Umweltisolat dB2. Diese fünf Bakterien zeichneten

Diskussion
55
sich dadurch aus, dass sie in Meer-, Brack- und Süßwasser gutes Wachstum zeigten.
Die optimale Salzkonzentration lag für diese Bakterien im Brackwasser-Milieu,
während Süßwasser- und Meerwasser-Zustände ebenfalls toleriert wurden.
E. coli und S. liquefaciens zählen zu den Enterobacteriaceae (Welch, 2006; Grimont &
Grimont, 2006) und kommen eher im Süßwasser vor. In dieser Arbeit zeigten E. coli
und S. liquefaciens die höchsten Zellzahlen in Brackwasser, gefolgt von Süßwasser und
Meerwasser.
E. coli bewohnt primär den Gastrointestinaltrakt von Menschen und Tieren und kommt
nur sekundär in Boden und Wasser vor, als Folge fäkaler Verschmutzung (Welch, 2006).
Laut Hood & Ness (1981) wächst E. coli in Meer- und Brackwasser nicht gut, dafür
aber in marinen Sedimenten.
Die Gattung Serratia ist salztolerant und wächst bei relativ niedrigen Temperaturen.
Bakterien dieser Gattung benötigen keine Wachstumszusätze zu Minimalmedium
(Grimont & Grimont, 2006). Die niedrigen Ansprüche an die Umweltbedingungen
spiegeln sich auch im Wachstum des in dieser Arbeit untersuchten Stammes von
S. liquefaciens wider, da S. liquefaciens sowohl in Meer-, Brack- und Süßwasser gutes
Wachstum zeigte.
P. aeruginosa ist ein sehr anpassungsfähiges Bakterium (Yahr & Parsek, 2006) und
zeigte in dieser Arbeit sowohl in Meer- und Brack- als auch in Süßwasser gutes
Wachstum. P. aeruginosa ist das einzige Isolat der salztoleranten Bakterien mit einer
höheren Zellzahl in Meer- als in Süßwasser.
P. aeruginosa ist ein typisches Boden- und Wasser-Bakterium. Fortbewegung kann
durch schwimmen, zucken und schwärmen erfolgen: In aquatischer Umgebung
schwimmt es mit einem einzelnen polaren Flagellum, auf festen Oberflächen zuckt es
mit Hilfe von Typ IV-Pili oder schwärmt mit Hilfe des Flagellums in Kombination mit
den Typ IV-Pili. Es kann Biofilme ausbilden. (Yahr & Parsek, 2006)
Der aus einem Ballastwassertank isolierte Bakterienstamm dB2 wuchs in jeglicher Form
von salzarmem oder salzreichem Wasser.

Diskussion
56
Wachstum der meerwasserliebenden Bakterien:
Vibrionen sind salztolerante Organismen, die in Meer- und Brackwasser vorkommen.
V. parahaemolyticus und V. alginolyticus sind dabei typische Meerwasser-Organismen.
Dementsprechend zeigten V. parahaemolyticus und V. alginolyticus in dieser Arbeit
hohes und konstantes Wachstum in Meer- und Brackwasser.
V. vulnificus DSM 10143 zeigte geringfügig besseres Wachstum in Brack- als in
Meerwasser. Kaspar & Tamplin (1993) wiesen nach, dass der optimale Salzgehalt für
V. vulnificus im Brackwasserbereich, unter 25 ppt, liegt. Ungewöhnlicherweise zeigte
V. vulnificus DSM 11507 das höchste Wachstum in Süßwasser. Über Vibrio vulnificus
DSM 11507 konnte keine Literatur gefunden werden. Die Angaben der DSMZ weisen
lediglich darauf hin, dass es sich bei diesem Bakterium um einen aus klinischem
Umfeld isolierten Keim handelt. Um sicher zu gehen, dass es sich bei dem untersuchten
Isolat tatsächlich um V. vulnificus handelt wurde die DNA zur Sanger-Sequenzierung
(Primer 63f, 1387r und 341f) an die Firma Qiagen geschickt. Über das Programm
BLAST wurde die Sequenz von V. vulnificus DSM 11507 mit anderen V. vulnificus-
Stämmen der NCBI-Datenbank verglichen und wies eine 100%ige Übereinstimmung
auf. Der Umstand, dass V. vulnificus DSM 11507 sich sowohl an salzreiches als auch an
salzarmes Wasser innerhalb von 48 Stunden sehr gut anpassen kann, steigert das
Gefahrenpotential dieses Bakterienstammes umso mehr.
Wachstum der süßwasserliebenden Bakterien:
E. faecalis und A. hydrophila zeigten optimales Wachstum in Süßwasser. Der Umstand,
dass beide Organismen auch in Brackwasser ein gutes und konstantes Wachstum
aufwiesen zeigt, dass beide Bakterienstämme bis zu einer bestimmten Grenze Salz
tolerieren können. In Meerwasser trat kein Wachstum auf. Laut Pianetti et al (2005) ist
A. hydrophila ein Süßwasserbakterium, das über längere Zeiträume in nährstoffarmen
Habitaten überleben kann und in Form von Biofilmen oder in einem VBNC- (viable but
not culturable) Zustand überdauert. Maalej et al (2004) wiesen nach, dass A. hydrophila
bei den Temperaturen 5°C und 23°C bis zu drei bis fünf Wochen in steril-filtriertem
Seewasser überleben kann. Für E. faecalis zeigten Hartke et al (2001) Überleben in
sterilem Meerwasser. Dennoch trat in dieser Arbeit für beide Bakterienstämme kein
Wachstum in Meerwasser auf.

Diskussion
57
4.2 Wachstum mit verschiedenen Acrolein-Konzentrationen
In dieser Arbeit wurde die Wirkung der sechs Acrolein-Konzentrationen 0.01, 0.1, 1, 5,
10 und 15 mg/L auf die zuvor beschriebenen Isolate untersucht. Aufgrund der
gewonnen Ergebnisse wird die Wirkung der Acrolein-Konzentrationen im Folgenden in
zwei Abschnitte getrennt, und zwar in die kaum wirksamen Konzentrationen 0.01 und
0.1 mg/L und die wirksamen Konzentrationen 1, 5, 10 und 15 mg/L.
Die LC50-Werte von Fischen liegen im Bereich niedriger Acrolein-Konzentrationen, die
beim Mensch kaum zu Reaktionen führen. Besonders auf aquatische Organismen wirkt
es hoch toxisch und beeinträchtigt sie bereits bei einer Konzentration von 11,4 µg/L.
EC50 (effect concentration for 50% population) und LC50 (lethal concentration for 50%
population) liegen für Bakterien, Algen, Crustaceen und Fische zwischen 0,02 –
2,5 mg/L (Ghilarducci & Tjeerdema, 1995).
Da die meerwasserliebenden Bakterien in Süßwasser kein Wachstum zeigte, und
genauso die süßwasserliebenden Bakterien in Meerwasser, kann für diese Gruppen in
diesen Salzkonzentrationen keine Aussage über die Wirkung der Acrolein-
Konzentrationen gemacht werden. Dementsprechend wird im Folgenden nicht extra für
jede Acrolein-Konzentration erwähnt, dass die meerwasserliebenden Bakterien in
Süßwasser und die süßwasserliebenden Bakterien in Meerwasser kein Wachstum
zeigten.
4.2.1 Acrolein-Konzentration 0.01 und 0.1 mg/L
Die niedrigen Acrolein-Konzentrationen 0.01 und 0.1 mg/L führten zu keinem
Rückgang der Zellzahlen bei den untersuchten Isolaten, außer bei A. hydrophila.
A. hydrophila zeigt bei 0.1 mg/L Acrolein nach 96 h Inkubation in Brackwasser kein
Wachstum mehr.
Bei einigen Bakterien konnte beobachtet werden, dass die Zellzahlen bei Inkubation mit
niedrigen Acrolein-Konzentrationen vergleichsweise höher war als bei inkubierten
Bakterien ohne Acrolein-Zugabe. Der Unterschied ist oft nur gering und tritt
ausschließlich bei Versuchen in Meerwasser innerhalb der ersten 24 Stunden auf. Bei
folgenden Bakterien wurde ein Anstieg der Zellzahl bei niedrigen Konzentrationen
beobachtet: E. coli, P. aeruginosa, S. liquefaciens und dB2. Möglicherweise sind diese

Diskussion
58
Isolate in der Lage, Acrolein in niedriger Konzentration zu verwerten. Eine weitere
Erklärung wäre, dass es sich um eine Schutzreaktion handelt und die Bakterien bei
Auftritt toxischer Komponenten ihren Stoffwechsel steigern um das eigene Überleben
zu garantieren. Für Enterobakterien wurde bereits gezeigt, dass sie ein rpoS-Gen
besitzen, das unter Stressbedingungen wie z.B. in der Umgebung von Seewasser eine
Anti-Stress-Antwort auslösen kann, die zu Anpassungserscheinungen und demzufolge
zu besserem Überleben führt (Trousselier et al, 1998).
4.2.2 Wirkung der Acrolein-Konzentrationen 1-15 mg/L auf die
untersuchten Isolate
Die Reaktion der einzelnen untersuchten Isolate auf 1 mg/L Acrolein war sehr divers
und lässt keine allgemeinen Aussagen zu. Bereits nach 24 Stunden konnte eine biozide
Wirkung von 1 mg/L Acrolein auf die untersuchten Isolate festgestellt werden. Der
biozide Effekt war stark davon abhängig, bei welchem Salzgehalt die Inkubation
erfolgte. Die höheren Acrolein-Konzentrationen 5, 10 und 15 mg/L führten innerhalb
von fünf Tagen Inkubation zu letalen Auswirkungen bei allen untersuchten Bakterien.
Alle Versuche ergaben einstimmig, dass bei Inkubation mit 5 – 15 mg/L Acrolein nach
fünf Tagen kein Wachstum möglich ist.
Die Wiederholung der Versuche für das Wachstum nach fünf Tagen Inkubation zeigte,
dass die Ergebnisse für 1 mg/L Acrolein von Versuchsansatz zu Versuchsansatz
abweichen können. Offenbar hängt die Reaktion auf 1 mg/L Acrolein von mehr
Faktoren als nur Salzgehalt, Temperatur und Inkubationszeit ab. Zu den Bakterien, die
im zweiten Versuchsansatz unerwartet Wachstum an Tag 5 zeigten zählen E. coli in
Süßwasser, P. aeruginosa in Brack- und Süßwasser, V. vulnificus DSM 11507 in
Meerwasser und V. parahaemolyticus in Meerwasser.
Im Zeitverlauf von 0 h bis 120 h war zu sehen, dass die toxische Wirkung von 1mg/L
Acrolein stetig zunahm. Laut Smith et al (2007) soll die Halbwertszeit von Acrolein
zwischen 15 und 60 h liegen. Die verstärkte toxische Wirkung über die Zeit bedeutet
entweder, dass die Wirkung von Acrolein erst mit der Zeit verstärkt wird, oder dass die
Abbauprodukte von Acrolein ebenfalls toxische Wirkung haben.

Diskussion
59
E. faecalis zeigte als einziger untersuchter Bakterienstamm keine Reaktion auf die
Behandlung mit 1 mg/L Acrolein. In 5 mg/L Acrolein überlebten nur zwei
Bakterienstämme 48 Stunden lang, nämlich P. aeruginosa in Meer- und Brackwasser
und E. faecalis in Brack- und Süßwasser. P. aeruginosa überlebte in 1 mg/L Acrolein
keine 120 h Inkubation, zeigte aber sogar in 15 mg/L Acrolein als einziges untersuchtes
Isolat 24 h lang Wachstum in Meerwasser.
Offenbar besitzen diese beiden Bakterienstämme Eigenschaften, die ihnen eine größere
Resistenz gegenüber Acrolein verleihen als anderen Bakterienstämmen. P. aeruginosa
verfügt über eine große Auswahl an Stoffwechselwegen und Pathogenitätsfaktoren. Es
ist in der Lage, unter verschiedensten Umweltbedingungen zu überleben, zu überdauern
und sich anzupassen. Es kann mehr als 80 organische Komponenten als Energie- und
Kohlenstoffquelle nutzen (Yahr & Parsek, 2006). Es ist also möglich, dass
P. aeruginosa in der Lage ist, kurzzeitig auch Acrolein zu verwerten. Außerdem besitzt
P. aeruginosa eine hohe Resistenz gegen Biozide. Ein hoher Gehalt an Mg2+ in der
Außenmembran führt zur Ausbildung starker LPS-LPS-Bindungen. Zudem lässt die
geringe Größe der Porine nur in beschränktem Maße Diffusionen zu (McDonnell &
Russel, 1999). Es gibt Pseudomonas-Arten die in der Lage sind, Phenole und Aldehyde
zu inaktivieren. P. aeruginosa besitzt ein Mex-Efflux-System, das grundsätzlich eine
sehr hohe Resistenz gegen Biozide ermöglicht (Beumer et al, 2000; Poole, 2002).
Als grampositives Bakterium sollte E. faecalis eigentlich empfindlicher auf Biozide
reagieren als die übrigen untersuchten Isolate. In erster Linie greifen Biozide die
Zellwand und Komponenten der Außenmembran an. Da gramnegative Bakterien eine
äußere Membran besitzen die grampositive Bakterien nicht haben, reagieren
grampositive Bakterien für gewöhnlich empfindlicher auf Biozide. Es gibt jedoch
Ausnahmen, z.B. hat Chlor eine größere biozide Wirkung auf P. aeruginosa als auf
Staphylococcus aureus. Eine weitere Möglichkeit für eine Resistenz ist das Bilden von
Biofilmen, doch auch das trifft nicht auf E. faecalis zu. Resistenzmechanismen
grampositiver Bakterien können die Glycocalyx oder Mucoexopolysaccharide sein
(Beumer et al, 2000).

Diskussion
60
V. parahaemolyticus, V. alginolyticus und das Umweltisolat dB2 reagierten am
empfindlichsten auf die Behandlung mit Acrolein. Keines dieser drei Isolate überlebte
in 1-15 mg/L Acrolein mehr als 24 h in Meer-, Brack- und Süßwasser. Die Inkubation in
5 mg/L Acrolein überlebte von diesen drei Isolaten nur V. alginolyticus 24 Stunden lang
in Brackwasser. Da die Zellzahlen des Umweltisolats dB2 unter den
Versuchsbedingungen auch ohne Acrolein niedrig waren, führte ein Abfall der
Zellzahlen schnell dazu, dass kein Wachstum mehr auftrat.
Ähnlich empfindlich reagierte A. hydrophila auf Acrolein. Mit 1 mg/L Acrolein zeigte
dieses Isolat noch 72 h lang Wachstum in Süßwasser, doch mit 5-15 mg/L Acrolein trat
schon nach 24 h kein Wachstum mehr auf. A. hydrophila wurde einst zu den
Vibrionaceae dazu gezählt.
Die beiden V. vulnificus-Stämme DSM 10143 und DSM 11507 reagierten weniger
sensitiv auf die Behandlung mit Acrolein als die Vibrionen V. parahaemolyticus und
V. alginolyticus. Beide Isolate unterschieden sich stark voneinander bezüglich
Koloniemorphologie und Salztoleranz. Obwohl V. vulnificus DSM 11507 ohne Acrolein
die höchsten Zellzahlen in Süßwasser zeigte überlebte es die Inkubation mit Acrolein
am längsten in Brackwasser. In Brackwasser zeigte V. vulnificus DSM 11507 fünf Tage
lang Wachstum mit 1 mg/L Acrolein und 24 Stunden lang Wachstum mit 5 mg/L
Acrolein. Wie schon zuvor erwähnt liegen die bevorzugten Salzkonzentrationen von
V. vulnificus im Brackwasserbereich. Ohne Acrolein hatte V. vulnificus DSM 10143 das
höchste Wachstum in Brackwasser gezeigt, doch mit 1 mg/L Acrolein wuchs das Isolat
nur in Meerwasser und zeigte darin keine Reaktion auf das Acrolein. Mit 5 mg/L
Acrolein kam es in Meerwasser zu minimalem Wachstum nach 24 h.
Tendenziell scheinen Vibrionaceae und A. hydrophila empfindlicher auf Acrolein zu
reagieren als Enterobacteriaceae. E. faecalis, E. coli und S. liquefaciens zur Familie der
Enterobacteriaceae. Die beiden Isolate E. coli und S. liquefaciens wuchsen mit 5 mg/L
Acrolein 24 Stunden lang in Meer- und Brackwasser. S. liquefaciens zeigte mit 1 mg/L
Acrolein 120 h lang Wachstum in Süßwasser. Beide Isolate zeigten erhöhte Zellzahlen
bei Inkubation in Meerwasser mit 0.01 mg/L Acrolein. Offenbar sind beide Isolate in
der Lage, Acrolein in geringen Mengen zu verwerten.

Diskussion
61
4.3 Vergleich mit anderen Bioziden die in der Ballastwasser-
Behandlung eingesetzt werden
Es wurden bereits einige Biozide auf ihre Wirkung auf pathogene Bakterien getestet.
Gregg & Hallegraeff (2007) untersuchten die drei Biozide Peraclean® Ocean,
SeaKleen® und Vibrex®. Das Biozid Peraclean® Ocean basiert auf Peroxyessigsäure,
Vibrex® besteht aus Chlordioxid und SeaKleen® enthält Menadion. Die Wirkung dieser
Biozide wurde auf die Bakterien Escherichia coli, Vibrio alginolyticus, Listeria innocua
und Staphylococcus aureus getestet. Bei L. innocua und S. aureus handelt es sich um
grampositive Bakterien. Chlordioxid wurde schon in anderen Untersuchungen auf seine
Wirkung auf die Bakterien Vibrio parahaemolyticus, Vibrio harveyi und Vibrio sp.
getestet. In Tabelle 8 wurde zusammen getragen, welche Mindestkonzentrationen an
Biozid in den verschiedenen Untersuchungen notwendig waren, um verschiedene
pathogenen Bakterien abzutöten.
Tabelle 8: Biozid-Mindestkonzentration nach 48 h Inkubation der Bakterien in
filtriertem Seewasser (25°C), bei der nach 24 h Inkubation in Medium (37°C) kein
Wachstum mehr auftritt (Gregg & Hallegraeff, 2007)
Bacteria Peraclean® Ocean (in ppm) Seakleen® (in ppm)
E. coli 250 200
V. alginolyticus 125 100
L. innocua 125 100
S. aureus 125 100
Die Autoren fanden heraus, dass Vibrex® schon in der geringen Konzentration 15 ppm
effektiv gegen Bakterien wirksam war. Niedrigere Konzentrationen wurden nicht
untersucht. Weiterhin stellten sie fest, dass sowohl bei SeaKleen® als auch bei
Peraclean® Ocean die doppelte Konzentration an Biozid notwendig war, um das
Wachstum von E. coli zu verhindern, als für V. alginolyticus, L. innocua und S. aureus.
Gegen Acrolein zeigte sich E. coli ebenfalls resistenter als V. alginolyticus (Abbildung
32).
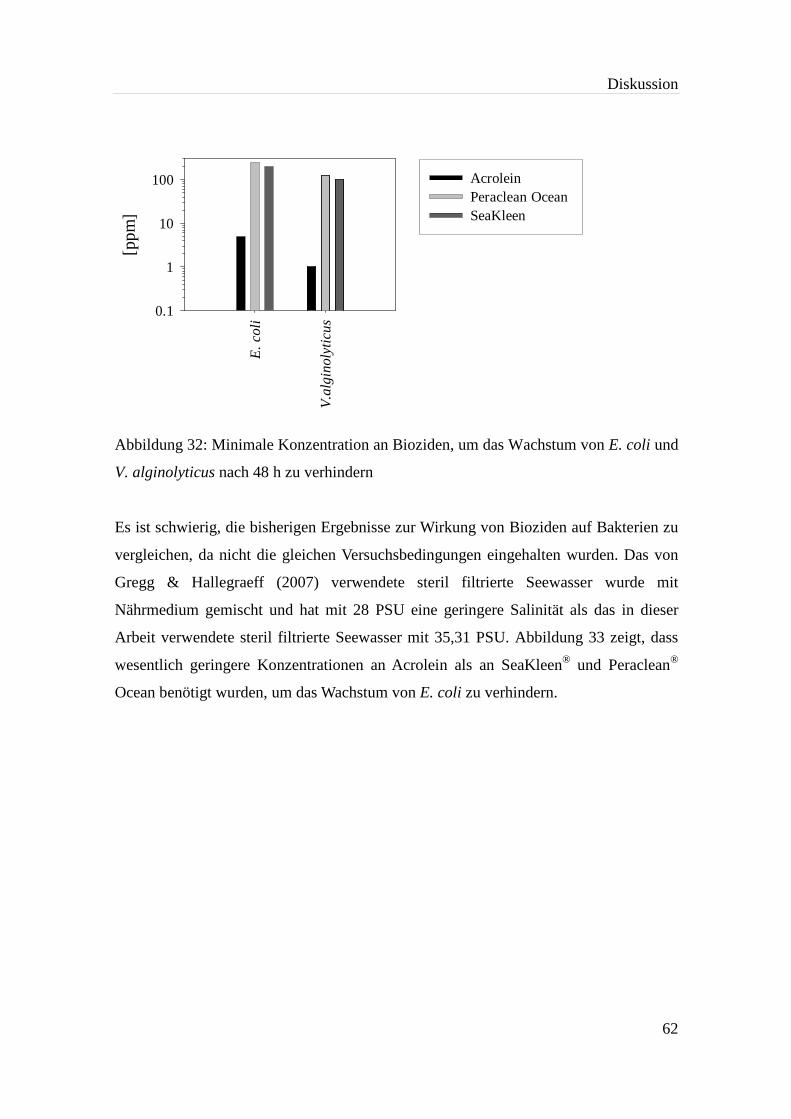
Diskussion
62
E.
coli
V.a
lgin
oly
ticu
s
[pp
m]
0.1
1
10
100 Acrolein Peraclean Ocean SeaKleen
Abbildung 32: Minimale Konzentration an Bioziden, um das Wachstum von E. coli und
V. alginolyticus nach 48 h zu verhindern
Es ist schwierig, die bisherigen Ergebnisse zur Wirkung von Bioziden auf Bakterien zu
vergleichen, da nicht die gleichen Versuchsbedingungen eingehalten wurden. Das von
Gregg & Hallegraeff (2007) verwendete steril filtrierte Seewasser wurde mit
Nährmedium gemischt und hat mit 28 PSU eine geringere Salinität als das in dieser
Arbeit verwendete steril filtrierte Seewasser mit 35,31 PSU. Abbildung 33 zeigt, dass
wesentlich geringere Konzentrationen an Acrolein als an SeaKleen® und Peraclean®
Ocean benötigt wurden, um das Wachstum von E. coli zu verhindern.
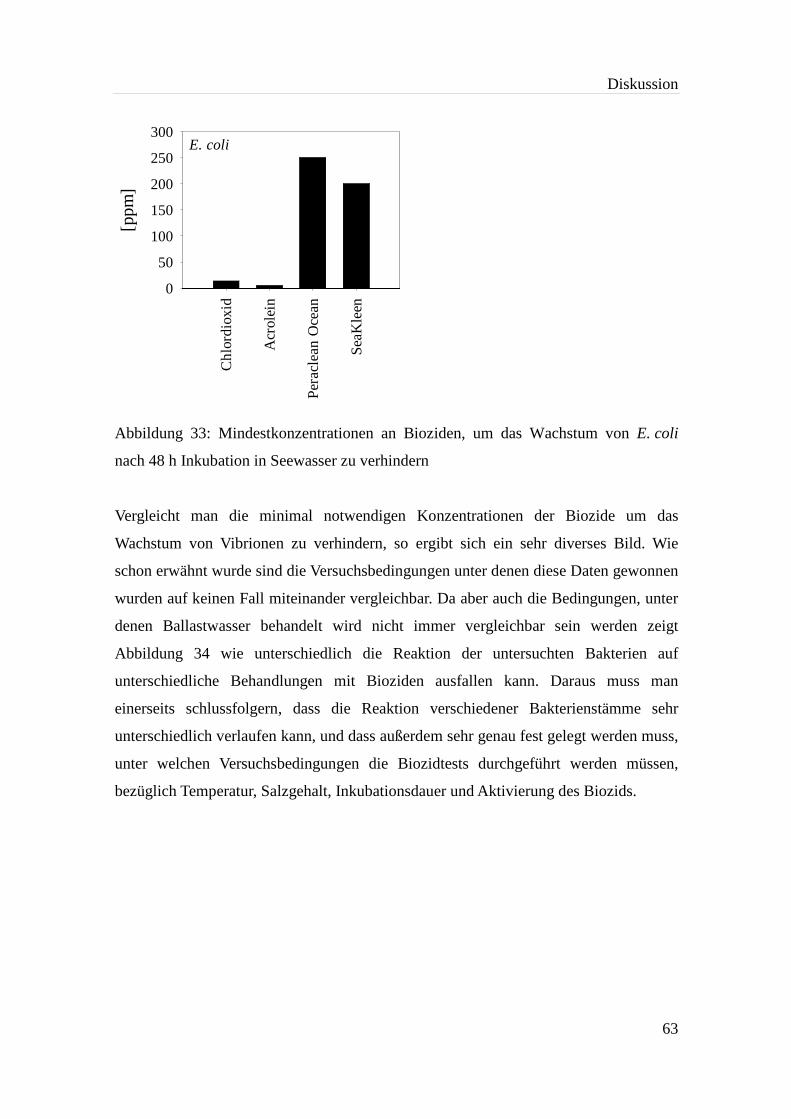
Diskussion
63
E. coli
Ch
lord
ioxi
d
Acr
olei
n
Per
acl
ean
Oce
an
Sea
Kle
en
[ppm
]
0
50
100
150
200
250
300
Abbildung 33: Mindestkonzentrationen an Bioziden, um das Wachstum von E. coli
nach 48 h Inkubation in Seewasser zu verhindern
Vergleicht man die minimal notwendigen Konzentrationen der Biozide um das
Wachstum von Vibrionen zu verhindern, so ergibt sich ein sehr diverses Bild. Wie
schon erwähnt wurde sind die Versuchsbedingungen unter denen diese Daten gewonnen
wurden auf keinen Fall miteinander vergleichbar. Da aber auch die Bedingungen, unter
denen Ballastwasser behandelt wird nicht immer vergleichbar sein werden zeigt
Abbildung 34 wie unterschiedlich die Reaktion der untersuchten Bakterien auf
unterschiedliche Behandlungen mit Bioziden ausfallen kann. Daraus muss man
einerseits schlussfolgern, dass die Reaktion verschiedener Bakterienstämme sehr
unterschiedlich verlaufen kann, und dass außerdem sehr genau fest gelegt werden muss,
unter welchen Versuchsbedingungen die Biozidtests durchgeführt werden müssen,
bezüglich Temperatur, Salzgehalt, Inkubationsdauer und Aktivierung des Biozids.
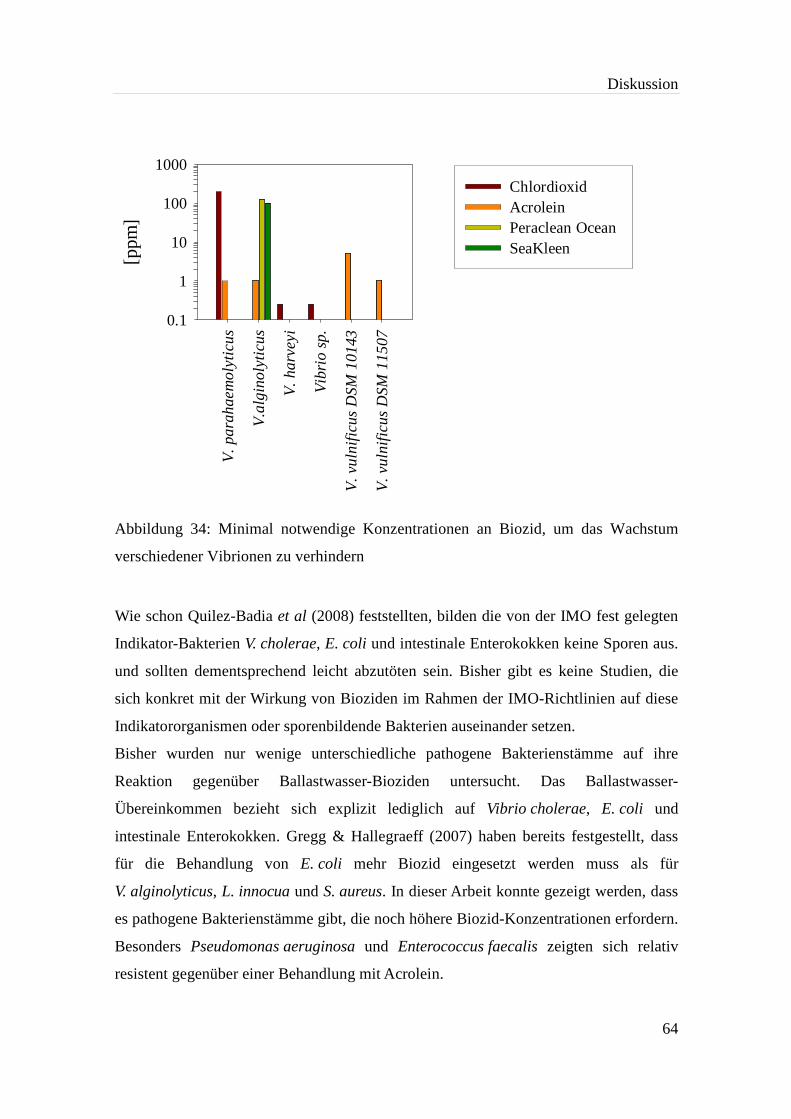
Diskussion
64
V.
pa
rah
ae
mo
lytic
us
V.a
lgin
oly
ticu
s
V.
ha
rve
yi
Vib
rio
sp
.
V.
vuln
ificu
s D
SM
10
143
V.
vuln
ificu
s D
SM
11
507
[ppm
]
0.1
1
10
100
1000Chlordioxid Acrolein Peraclean Ocean SeaKleen
Abbildung 34: Minimal notwendige Konzentrationen an Biozid, um das Wachstum
verschiedener Vibrionen zu verhindern
Wie schon Quilez-Badia et al (2008) feststellten, bilden die von der IMO fest gelegten
Indikator-Bakterien V. cholerae, E. coli und intestinale Enterokokken keine Sporen aus.
und sollten dementsprechend leicht abzutöten sein. Bisher gibt es keine Studien, die
sich konkret mit der Wirkung von Bioziden im Rahmen der IMO-Richtlinien auf diese
Indikatororganismen oder sporenbildende Bakterien auseinander setzen.
Bisher wurden nur wenige unterschiedliche pathogene Bakterienstämme auf ihre
Reaktion gegenüber Ballastwasser-Bioziden untersucht. Das Ballastwasser-
Übereinkommen bezieht sich explizit lediglich auf Vibrio cholerae, E. coli und
intestinale Enterokokken. Gregg & Hallegraeff (2007) haben bereits festgestellt, dass
für die Behandlung von E. coli mehr Biozid eingesetzt werden muss als für
V. alginolyticus, L. innocua und S. aureus. In dieser Arbeit konnte gezeigt werden, dass
es pathogene Bakterienstämme gibt, die noch höhere Biozid-Konzentrationen erfordern.
Besonders Pseudomonas aeruginosa und Enterococcus faecalis zeigten sich relativ
resistent gegenüber einer Behandlung mit Acrolein.

Diskussion
65
4.4 Acrolein als Biozid für Ballastwasserbehandlung
Ob Acrolein für die Behandlung von Ballastwasser geeignet ist, hängt von vielen
verschiedenen Faktoren ab und kann im Rahmen dieser Arbeit nicht abschließend oder
umfassend genug beurteilt werden.
Ausgehend von den in dieser Arbeit gewonnenen Ergebnissen wäre eine Konzentration
von 5 mg/L Acrolein ausreichend, um innerhalb von 5 Tagen Inkubation bei 20° C alle
untersuchten Bakterienstämme abzutöten. Diese Konzentration ist im Vergleich zu
anderen Ballastwasser-Bioziden sehr niedrig und hat den Vorteil, dass geringere
Mengen an Biozid transportiert und eingesetzt werden müssten.
Bedenklich ist das hohe Gefahrenpotential von Acrolein. Die Handhabung von Acrolein
als Ballastwasser-Biozid beinhaltet ein Gesundheits- und Lebensrisiko für die
Mannschaft an Bord. Beim Menschen führt Acrolein bei Kontakt zu Irritationen der
Augen, bei Inhalation zu Irritationen der Atemwege und bei oraler Aufnahme zu
Erbrechen, Völlegefühl und Diarrhöen. Da das Arbeiten auf hoher See stark von den
Umweltbedingungen beeinflusst wird besteht immer ein Unfallrisiko, das zu schweren
ökologischen Folgen führen würde. Im Falle eines Unfalls in einem Hafengebiet wäre
eine Umweltkatastrophe zu befürchten.
Die in dieser Arbeit durchgeführten Versuche haben gezeigt, dass bei einer
Konzentration von 1 mg/L Acrolein die biozide Wirkung innerhalb der Inkubationszeit
von fünf Tagen zunimmt. Es ist unbedingt notwendig zu überprüfen, nach welcher
Zeitspanne bei welcher Temperatur Acrolein hydrolysiert wird, da selbst niedrige
Konzentrationen an Acrolein ausreichen, um zu negativen Auswirkungen bei Fischen zu
führen. Es sind weitere Untersuchungen zu empfehlen, die sich mit der Wirkung der
Hydrolyseprodukte auf marine Organismen beschäftigen.

Diskussion
66
4.5 Methoden-Kritik
Im Rahmen dieser Arbeit war zu bewerten, inwiefern die ScanMIC-Methoden in
Kombination mit der MPN-Auswertung geeignet ist, um die Wirkung des
Ballastwasser-Biozids Acrolein auf verschiedene pathogene Bakterien zu ermitteln.
Am wichtigsten für die Verwendung von TTC ist die Wahl der richtigen Konzentration.
Zu hohe Konzentrationen an TTC können einen toxischen Effekt auf das Wachstum der
Bakterien haben, zu niedrige Konzentrationen reichen ggf. nicht aus, um eine Färbung
hervor zu rufen (Rahman et al, 2003; Beloti et al, 1999). Nicht alle Bakterien
reduzieren TTC zu Formazan. Beloti et al (1999) haben festgestellt, dass 95% der
Bakterien, die TTC nicht reduziert haben und Gram-gefärbt wurden, zu den
grampositiven Bakterien gehörten. Es gibt mehrere Faktoren, z.B. pH, Temperatur und
Licht, die die Reduktion von TTC beeinflussen.
Die von Rahman et al (2003) entwickelte ScanMIC-Methode ist unkompliziert und
erfordert nur wenige Geräte und Hilfsmittel. Es sind einige Vorversuche nötig um fest
zu stellen, welche TTC-Konzentration eine Färbung bei den untersuchten Bakterien
hervorruft. Die Zeit, die diese Vorversuche in Anspruch nehmen, hängt von der Anzahl,
der Herkunft und der Wachstumsgeschwindigkeit der untersuchten Bakterien ab. Für
die Bakterien, die für diese Arbeit über die DSMZ erworben wurden, war die Färbung
mit TTC gut geeignet. Positiv ist zu bewerten, dass man innerhalb von 24 h Ergebnisse
erhält und mehrere Biozid-Konzentrationen und mehrere Isolate gleichzeitig testen
kann.
Die Auswertung der gefärbten Kavitäten gestaltete sich schwierig, da einzelne Partikel
oft nicht von Luftblasen oder Fusseln zu unterscheiden sind. Genauso schwierig war zu
erkennen, ob diese kleinen Partikel rötlich oder ungefärbt sind. Dadurch ist die visuelle
Auswertung sehr subjektiv und personenabhängig. Wie schon zuvor erwähnt war die
Färbung der Bakterien in den Kavitäten der Mikrotiterplatten sehr unterschiedlich. Die
Auswertung über MPN-Tabellen berücksichtigt diese Unterschiede nicht. Große
zentrale gefärbte Pellets in einer rötlich gefärbten Kavität werden mit ungefärbten

Diskussion
67
Kavitäten mit einem einzelnen roten Partikel gleichgesetzt und führen in der
Auswertung zur gleichen Zellzahl.
Es ist nicht zu erkennen, ob es sich bei einzelnen rot gefärbten Kavitäten um den zu
untersuchenden Bakterienstamm oder um eine Kontaminante handelt, wenn die
Kontaminante mit TTC das gleiche Erscheinungsbild zeigt. Eine Negativkontrolle zeigt
nur, ob das Medium kontaminiert ist.
Unter Stressbedingungen gehen Bakterienstämme häufig in einen VBNC-Zustand
(viable but non-culturable) über. Zu diesen Stressbedingungen gehören beispielsweise
Temperatur und Salzgehalt (Kaspar & Tamplin, 1993; Trousselier et al, 1998; Rozen &
Belkin, 2001). Es ist unklar, inwiefern TTC Bakterienzellen im VBNC-Zustand anfärbt.
Ein großer Kritikpunkt ist die hohe Standardabweichung in den Ergebnissen. An einem
Beispiel kann dies demonstriert werden:
Abbildung 35 zeigt mögliche Kavitäten-Färbungen in einer Mikrotiterplatte. Die Menge
an gefärbten Kavitäten unterscheidet sich zwischen 1-3 nur geringfügig. Die daraus
resultierenden Zellzahlen nach MPN-Tabelle (McCrady, 1915) sind in Tabelle 9
widergegeben.
Abbildung 35: Beispiel für eine belegte Mikrotiterplatte mit unterschiedlicher Anzahl gefärbter Kavitäten
Tabelle 9: Zellzahlen nach MPN-Tabelle (McCrady, 1915) 1 2 3 4
N*ml -1 2500 1100 450 150
1 2 3 4
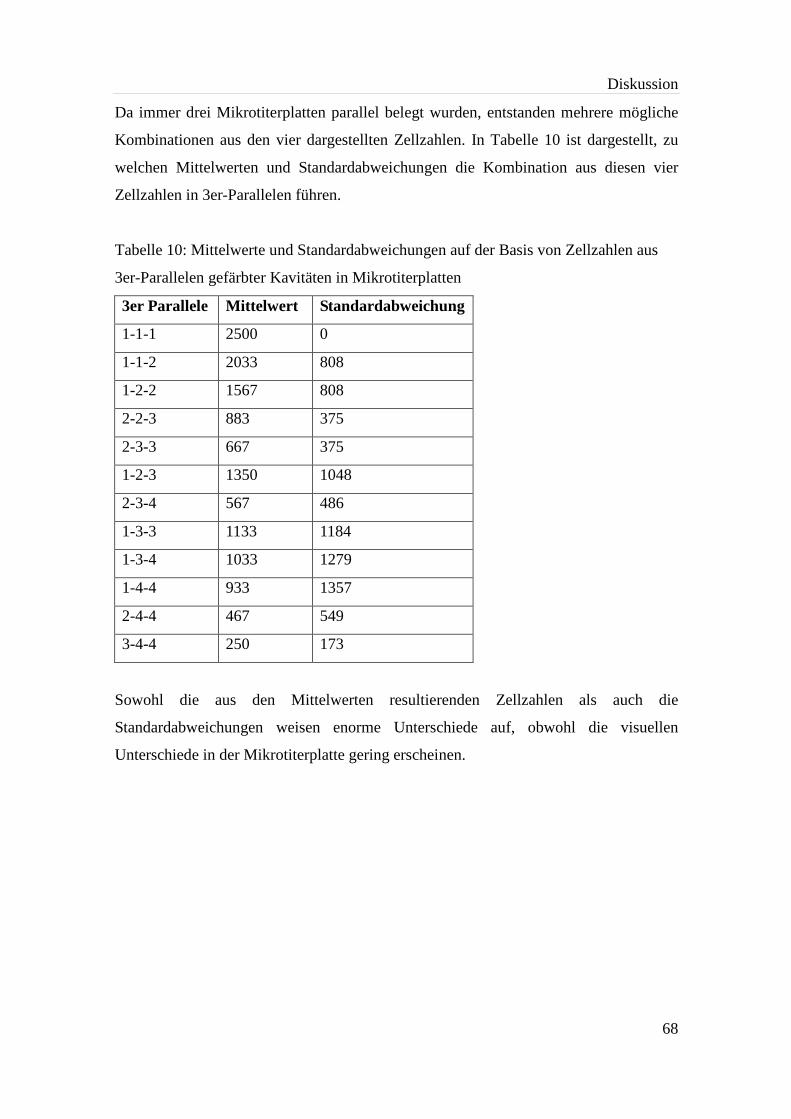
Diskussion
68
Da immer drei Mikrotiterplatten parallel belegt wurden, entstanden mehrere mögliche
Kombinationen aus den vier dargestellten Zellzahlen. In Tabelle 10 ist dargestellt, zu
welchen Mittelwerten und Standardabweichungen die Kombination aus diesen vier
Zellzahlen in 3er-Parallelen führen.
Tabelle 10: Mittelwerte und Standardabweichungen auf der Basis von Zellzahlen aus
3er-Parallelen gefärbter Kavitäten in Mikrotiterplatten
3er Parallele Mittelwert Standardabweichung
1-1-1 2500 0
1-1-2 2033 808
1-2-2 1567 808
2-2-3 883 375
2-3-3 667 375
1-2-3 1350 1048
2-3-4 567 486
1-3-3 1133 1184
1-3-4 1033 1279
1-4-4 933 1357
2-4-4 467 549
3-4-4 250 173
Sowohl die aus den Mittelwerten resultierenden Zellzahlen als auch die
Standardabweichungen weisen enorme Unterschiede auf, obwohl die visuellen
Unterschiede in der Mikrotiterplatte gering erscheinen.

Ausblick
69
5 Ausblick
Es wurde in dieser Arbeit gezeigt, dass Acrolein in niedrigen Konzentrationen eine
biozide Wirkung auf einige pathogene Bakterienstämme hat. Um den IMO-Richtlinien
zu genügen muss zusätzlich die Wirkung von Acrolein auf Vibrio cholerae und weitere
intestinale Enterokokken untersucht werden.
Studien die sich mit der Wirkung von Bioziden auf Bakterien beschäftigen setzen sich
bisher nur vereinzelt mit den im Rahmen der IMO-Richtlinien festgelegten
Indikatororganismen V. cholerae, E. coli und intestinale Enterokokken auseinander.
Interessant zu untersuchen wäre außerdem, ob sporenbildende Bakterienstämme die
Wirkung von Bioziden überleben können.
Untersuchungen auf mikrobieller Ebene zum Thema Ballastwasser müssen noch
verstärkt durchgeführt werden. Es gibt bisher erst wenige Nachweise dafür, dass
pathogene Bakterien über die Verschleppung durch Ballastwasser tatsächlich zu
vermehrten Erkrankungen führen. Dazu müsste erst untersucht werden, in welchem
Ausmaß überhaupt pathogene Bakterien regelmäßig in Ballastwasser transportiert
werden. Anschließend wäre beispielsweise ein Vergleich mit der Pathogenese von
Wildfischen und Fischen in Aquakultur sinnvoll.

Literaturliste
70
Literaturliste
Aridgides L. J., Doblin M. A., Berke T., Dobbs F. C., Matson D. O., Drake L.A.,
2003. Multiplex PCR allows simultaneous detection of pathogens in ships’ ballast
water. Marine Bulletin 48, 1096-1101
Austin B. & Austin D. A.: Bacterial Fish Pathogens. Netherlands: Springer-Verlag;
2007
Bai X., Zhang Z., Bai M., Yang B., Mindi B., 2004. Killing invasive species of ship’s
ballast water in 20t/h system using hydroxyl radicals. Plasma Chemistry and Plasma
Processing, Vol. 25, No. 1, 41-54
Bax N., Williamson A., Aguero M., Gonzalez E., Geeves W., 2003. Marine invasive
alien species: a threat to global biodiversity. Marine Policy 27, 313-323
Beloti V., Barros M. A. F., de Freitas J. C., Nero L. A., de Souza J. A., Santana E.
H. W., Franco D. G. M., 1999. Frequency of 2,3,5-Triphenyltetrazolium chloride
(TTC) non-reducing bacteria in pasteurized milk. Revista de Microbiologia 30, 137-140
Beumer R., Bloomfield S. F., Exner M., Fara G. M., Nath K. J., Scott E., 2000.
Microbial Resistance and Biocides. International Scientific Forum on Home Hygiene
Review, 1-42
Devriese L., Baele M., Butaye P.: The Genus Enterococcus: Taxonomy. In: The
Prokaryotes. Edited by Dworkin M.. Berlin, Springer-Verlag; 2006: 4, 163-174
Drake L. A., Doblin M. A., Dobbs F. C., 2006. Potential microbial bioinvasions via
ships’ ballast water, sediment, and biofilm. Marine Pollution Bulletin 55, 333-341

Literaturliste
71
Faimali M., Garaventa F., Chelossi E., Piazza V., Saracino O. D., Rubino F.,
Mariottini G. L., Pane L. , 2006. A new photodegradable molecule as a low impact
ballast water biocide: efficacy screening on marine organisms from different trophic
levels. Marine Biology 149, 7-16
Farmer III J. J.: The Family Vibrionaceae. In: The Prokaryotes. Edited by Dworkin
M. Berlin, Springer-Verlag; 2006: 6, 495-507
Farmer III J. J., Arduino M. J., Hickman-Brenner F. W.: The Genera Aeromonas
and Plesiomonas. In: The Prokaryotes. Edited by Dworkin M. Berlin, Springer-Verlag;
2006: 6, 508-563
Farmer III J. J. & Hickman-Brenner F. W. : The Genera Vibrio and Photobacterium.
In: The Prokaryotes. Edited by Dworkin M. Berlin, Springer-Verlag; 2006: 6, 508-563
Gabrielson J., Hart M., Jarelöv A., Kühn I., McKenzie D., Möllby R., 2002.
Evaluation of redox indicators and the use of digital scanners and spectrophotometer for
quantification of microbial growth in microplates. Journal of Microbiological Methods
50, 63-73
Gabrielson J., Kühn I., Colque-Navarro P., Hart M., Iversen A., McKenzie D.,
Möllby R. , 2002. Microplate-based microbial assay for risk assessment and (eco)toxic
fingerprinting of chemicals. Analytica Chimica Acta 485, 121-130
Ghilarducci D. P. & Tjeerdema R. S., 1995. Fate and effects of Acrolein. Reviews of
Environmental Contamination and Toxicology, Vol. 144, 95-147
Gollasch S., 1997. Removal of Barriers to the Effective Implementation of Ballast
Water Control and Management Measures in Developing Countries. IMO-Report, 1-
135

Literaturliste
72
Gregg M. D. & Hallegraeff G. M., 2007. Efficacy of three commercially available
ballast water biocides against vegetative microalgae, dinoflagellate cysts and bacteria.
Harmful Algae 6, 567-584
Grimont F. & Grimont P. A. D. : The Genus Serratia. In: The Prokaryotes. Edited by
Dworkin M. Berlin, Springer-Verlag; 2006: 6, 219-244
Hartke A., Lemarinier S., Pichereau V., Auffray Y., 2001. Survival of Enterococcus
faecalis in seawater microcosms is limited in the presence of bacterivorous
zooflagellates. Current Microbiology, Vol. 44, 329-335
Hochhaus K.-H. & Mehrkens C., 2007. Ballastwasseraufbereitung – eine Übersicht.
Schiff & Hafen Nr. 3, 66-71
Hood M. A. & Ness G. E., 1981. Survival of Vibrio cholerae and Escherichia coli in
Estuarine Waters and Sediments. Applied and Environmental Microbiology, Vol. 43,
No. 3, 578-584
International Maritime Organisation , 2005. Guidelines for Approval of Ballast
Water Management Systems (G8). MEPC [53], 24, Add. 1
Kaspar C. W. & Tamplin M. L. , 1993. Effects of temperature and salinity on the
survival of Vibrio vulnificus in seawater and shellfish. Applied and Environmental
Microbiology, Vol. 59, No. 8, 2425-2429
Kolar C. S. & Lodge D. M., 2001. Progress in invasion biology: predicting invaders.
TRENDS in Ecology & Evolution Vol. 16 No. 4, 199-204
Lavoie D. M., Smith L. D., Ruiz G. M., 1999. The Potential for Intracoastal Transfer
of Non-indigenous Species in the Ballast Water of Ships. Estuarine, Coastal and Shelf
Science 48, 551-564

Literaturliste
73
Lenz J., Andres H.-G., Gollasch S., Dammer M., 1996. Einschleppung fremder
Organismen in Nord- und Ostsee: Untersuchungen zumökologischen Gefahrenpotential
durch den Schiffsverkehr. Umweltforschungsplan des Bundesministers für Umwelt,
Naturschutz und Reaktorsicherheit, Forschungsbericht 102 04 250
Linkous D. A. & Oliver J. D. , 1999. Pathogenesis of Vibrio vulnificus. FEMS
Microbiology Letters 174, 207-214
Louder D. E. & McCoy E. G., 1962. Preliminary investigations of the use of aqualin
for collecting fishes. Proc. Ann. 16th Conf. Southeast. Assoc., Game & Fish
Commissioners, 240-242
Maalej S., Denis M., Dukan S., 2004. Temperature and growth-phase effects on
Aeromonas hydrophila survival in natural seawater microcosms: role of protein
synthesis and nucleic acid content on viable but temporarily nonculturable response.
Microbiology 150, 181-187
Madigan, M. T., Martinko, J. M., Parker, J.: Brock Mikrobiologie. Heidelberg:
Spektrum Akademischer Verlag GmbH; 2001
McCrady M. H. , 1915. The numerical interpretation of fermentation-tube results.
Journal of Infectious Diseases 17, 183-212
McDonnell G. & Russel A. D., 1999. Antiseptics and Desinfectants: Activity, Action,
and Resistance. Clinical Microbiology Reviews, Vol. 12, No. 1, 147-179
Meyer-Reil, L. A.: Mikrobiologie des Meeres. Wien: Facultas Verlags- und
Buchhandels AG; 2005

Literaturliste
74
Nehring S., Meyer T., Beyer K., Reimers H.-C., 2004. EU-Wasserrahmenrichtlinie –
Anthropogene Umweltauswirkungen im Pilotgebiet “Küstengewässer Elbe”.
Mitteilungen der Deutschen Gesellschaft für Meeresforschung, Heft 1, 3-8
Pianetti A., Falcioni T., Bruscolini F., Sabatini L., Sisti E., Papa S., 2005.
Determination of the viability of Aeromonas hydrophila in different types of water by
flow cytometry, and comparison with classical methods. Applied and Environmental
Microbiology, Vol. 71, No. 12, 7948-7954
Poole K., 2002. Mechanisms of bacterial biocide and antibiotic resistance. Journal of
Applied Microbiology Symposium Suppl. 92, 55S- 64S
Quilez-Badia G., McCollin T., Josefsen K. D., Vourdachas A., Gill M. E., Mesbahi
E., Frid C. L. J., 2008. Marine Pollution Bulletin 56, 127-135
Rahman Mokhlasur, Kühn I., Rahman Motiur, Olsson-Liljequist B., Möllby R.,
2003. Evaluation of a scanner-assisted colorimetric MIC method for susceptibility
testing of gram-negative fermentative bacteria. Applied and Environmental
Microbiology, Vol. 70, No. 4, 2398-2403
Reise K., Gollasch S., Wolff W. J., 1999. Introduced marine species of the North Sea
coasts. Helgoländer Meeresuntersuchungen 52, 219-234
Rozen Y. & Belkin S., 2001. Survival of enteric bacteria in seawater. FEMS
Microbiology Reviews 25, 513-529
Ruiz G. M., Rawlings T. K., Dobbs F. C., Drake L. A., Mullady T., Huq A.,
Colwell R. R., 2000. Global spread of microorganisms by ships. Nature, Vol. 408, 49

Literaturliste
75
Smith A. M., Mao J., Doane R. A., Kovacs Jr. M. F., 2007. Metabolic fate of [14C]
Acrolein under aerobic and anaerobic aquatic conditions. J. Agric. Food Chem. 43,
2497-2503
Trousselier M., Bonnefont J.-L., Courties C., Derrien A., Dupray E., Gauthier M.,
Gourmelon M., Joux F., Lebaron P., Martin Y., Pommepuy M., 1998. Responses of
enteric bacteria to environmental stresses in seawater. Oceanologica Acta, Vol. 21, No.
6, 965-981
Tsukatani T., Suenaga H., Higuchi T., Akao T., Ishiyama M., Ezoe K., Matsumoto
K., 2008. Colorimetric cell proliferation assay for microorganisms in microtiter plate
using water-soluble tetrazolium salts. Journal of Microbiological Methods 75, 109-116
Turner L. & Erickson W. , 2003. Acrolein. Analysis of risks from the aquatic
herbicide use in irrigation supply canals to eleven evolutionarily significant units of
pacific salmon and steelhead. Office of Pesticide Programs, 1-49
Welch R. A.: The Genus Escherichia. In: The Prokaryotes. Edited by Dworkin M.
Berlin, Springer-Verlag; 2006: 6, 60-71
Yahr T. L. & Parsek M. R. : Pseudomonas aeruginosa. In: The Prokaryotes. Edited by
Dworkin M. Berlin, Springer-Verlag; 2006: 6, 704-713
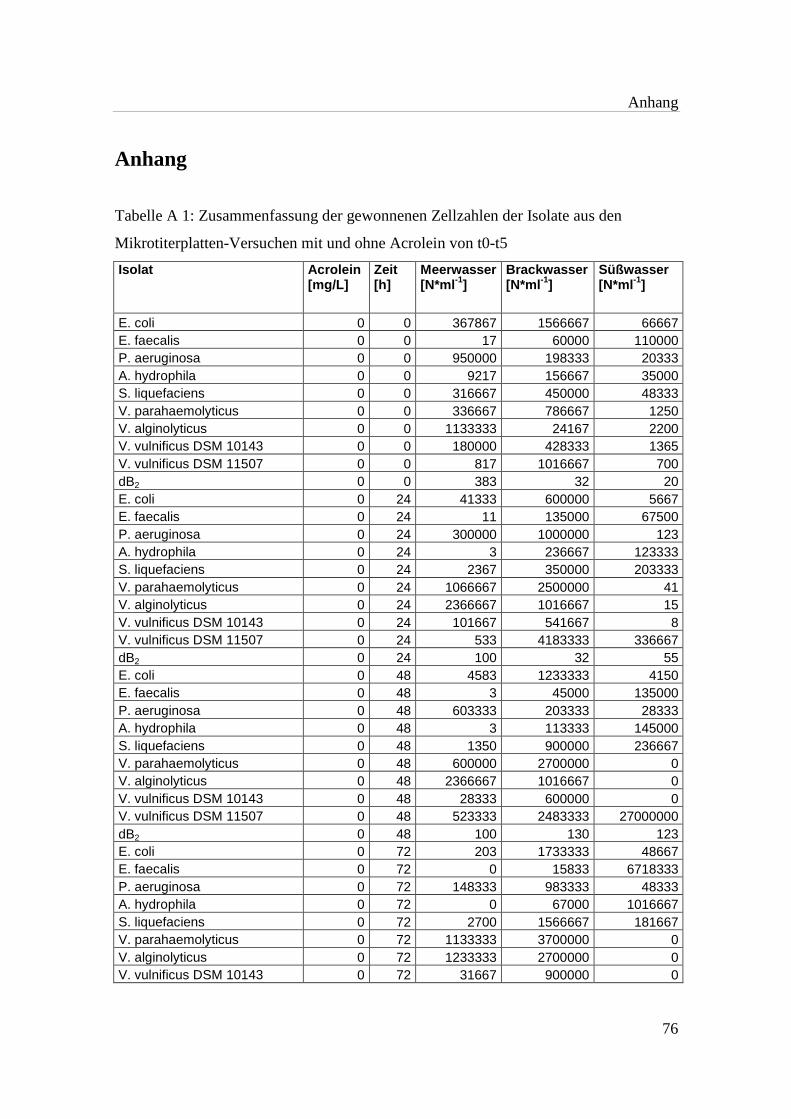
Anhang
76
Anhang
Tabelle A 1: Zusammenfassung der gewonnenen Zellzahlen der Isolate aus den
Mikrotiterplatten-Versuchen mit und ohne Acrolein von t0-t5
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
E. coli 0 0 367867 1566667 66667 E. faecalis 0 0 17 60000 110000 P. aeruginosa 0 0 950000 198333 20333 A. hydrophila 0 0 9217 156667 35000 S. liquefaciens 0 0 316667 450000 48333 V. parahaemolyticus 0 0 336667 786667 1250 V. alginolyticus 0 0 1133333 24167 2200 V. vulnificus DSM 10143 0 0 180000 428333 1365 V. vulnificus DSM 11507 0 0 817 1016667 700 dB2 0 0 383 32 20 E. coli 0 24 41333 600000 5667 E. faecalis 0 24 11 135000 67500 P. aeruginosa 0 24 300000 1000000 123 A. hydrophila 0 24 3 236667 123333 S. liquefaciens 0 24 2367 350000 203333 V. parahaemolyticus 0 24 1066667 2500000 41 V. alginolyticus 0 24 2366667 1016667 15 V. vulnificus DSM 10143 0 24 101667 541667 8 V. vulnificus DSM 11507 0 24 533 4183333 336667 dB2 0 24 100 32 55 E. coli 0 48 4583 1233333 4150 E. faecalis 0 48 3 45000 135000 P. aeruginosa 0 48 603333 203333 28333 A. hydrophila 0 48 3 113333 145000 S. liquefaciens 0 48 1350 900000 236667 V. parahaemolyticus 0 48 600000 2700000 0 V. alginolyticus 0 48 2366667 1016667 0 V. vulnificus DSM 10143 0 48 28333 600000 0 V. vulnificus DSM 11507 0 48 523333 2483333 27000000 dB2 0 48 100 130 123 E. coli 0 72 203 1733333 48667 E. faecalis 0 72 0 15833 6718333 P. aeruginosa 0 72 148333 983333 48333 A. hydrophila 0 72 0 67000 1016667 S. liquefaciens 0 72 2700 1566667 181667 V. parahaemolyticus 0 72 1133333 3700000 0 V. alginolyticus 0 72 1233333 2700000 0 V. vulnificus DSM 10143 0 72 31667 900000 0
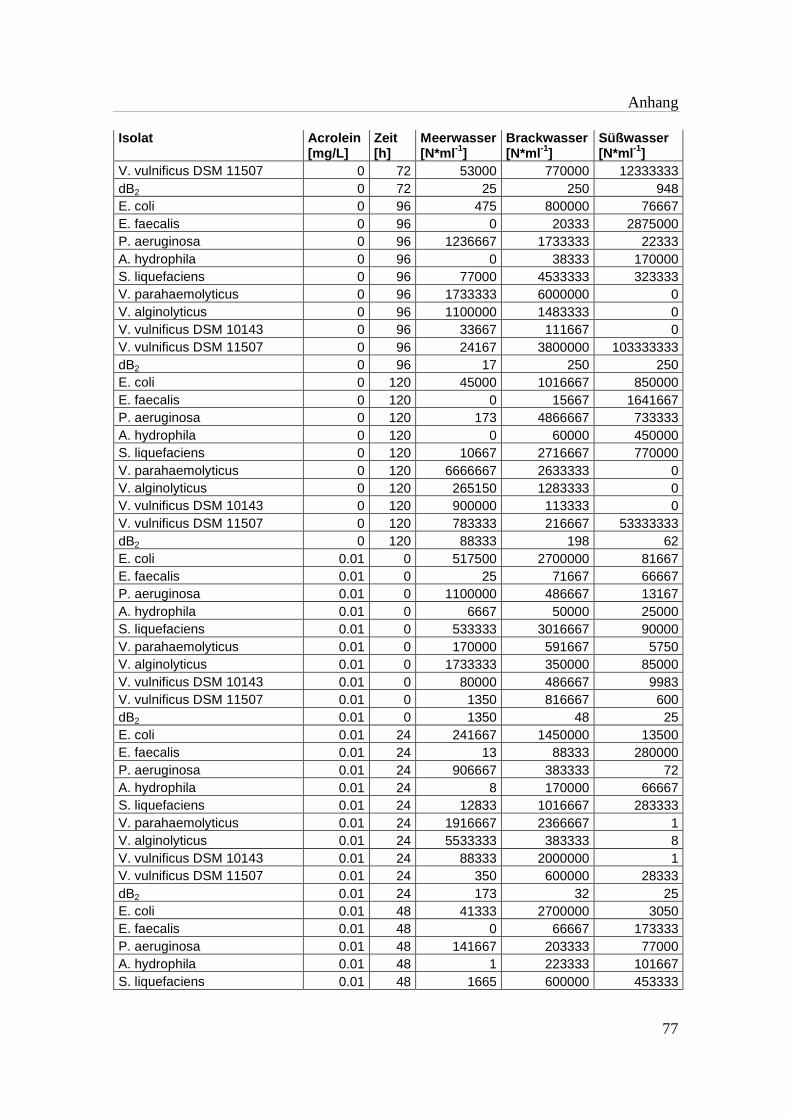
Anhang
77
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
V. vulnificus DSM 11507 0 72 53000 770000 12333333 dB2 0 72 25 250 948 E. coli 0 96 475 800000 76667 E. faecalis 0 96 0 20333 2875000 P. aeruginosa 0 96 1236667 1733333 22333 A. hydrophila 0 96 0 38333 170000 S. liquefaciens 0 96 77000 4533333 323333 V. parahaemolyticus 0 96 1733333 6000000 0 V. alginolyticus 0 96 1100000 1483333 0 V. vulnificus DSM 10143 0 96 33667 111667 0 V. vulnificus DSM 11507 0 96 24167 3800000 103333333 dB2 0 96 17 250 250 E. coli 0 120 45000 1016667 850000 E. faecalis 0 120 0 15667 1641667 P. aeruginosa 0 120 173 4866667 733333 A. hydrophila 0 120 0 60000 450000 S. liquefaciens 0 120 10667 2716667 770000 V. parahaemolyticus 0 120 6666667 2633333 0 V. alginolyticus 0 120 265150 1283333 0 V. vulnificus DSM 10143 0 120 900000 113333 0 V. vulnificus DSM 11507 0 120 783333 216667 53333333 dB2 0 120 88333 198 62 E. coli 0.01 0 517500 2700000 81667 E. faecalis 0.01 0 25 71667 66667 P. aeruginosa 0.01 0 1100000 486667 13167 A. hydrophila 0.01 0 6667 50000 25000 S. liquefaciens 0.01 0 533333 3016667 90000 V. parahaemolyticus 0.01 0 170000 591667 5750 V. alginolyticus 0.01 0 1733333 350000 85000 V. vulnificus DSM 10143 0.01 0 80000 486667 9983 V. vulnificus DSM 11507 0.01 0 1350 816667 600 dB2 0.01 0 1350 48 25 E. coli 0.01 24 241667 1450000 13500 E. faecalis 0.01 24 13 88333 280000 P. aeruginosa 0.01 24 906667 383333 72 A. hydrophila 0.01 24 8 170000 66667 S. liquefaciens 0.01 24 12833 1016667 283333 V. parahaemolyticus 0.01 24 1916667 2366667 1 V. alginolyticus 0.01 24 5533333 383333 8 V. vulnificus DSM 10143 0.01 24 88333 2000000 1 V. vulnificus DSM 11507 0.01 24 350 600000 28333 dB2 0.01 24 173 32 25 E. coli 0.01 48 41333 2700000 3050 E. faecalis 0.01 48 0 66667 173333 P. aeruginosa 0.01 48 141667 203333 77000 A. hydrophila 0.01 48 1 223333 101667 S. liquefaciens 0.01 48 1665 600000 453333
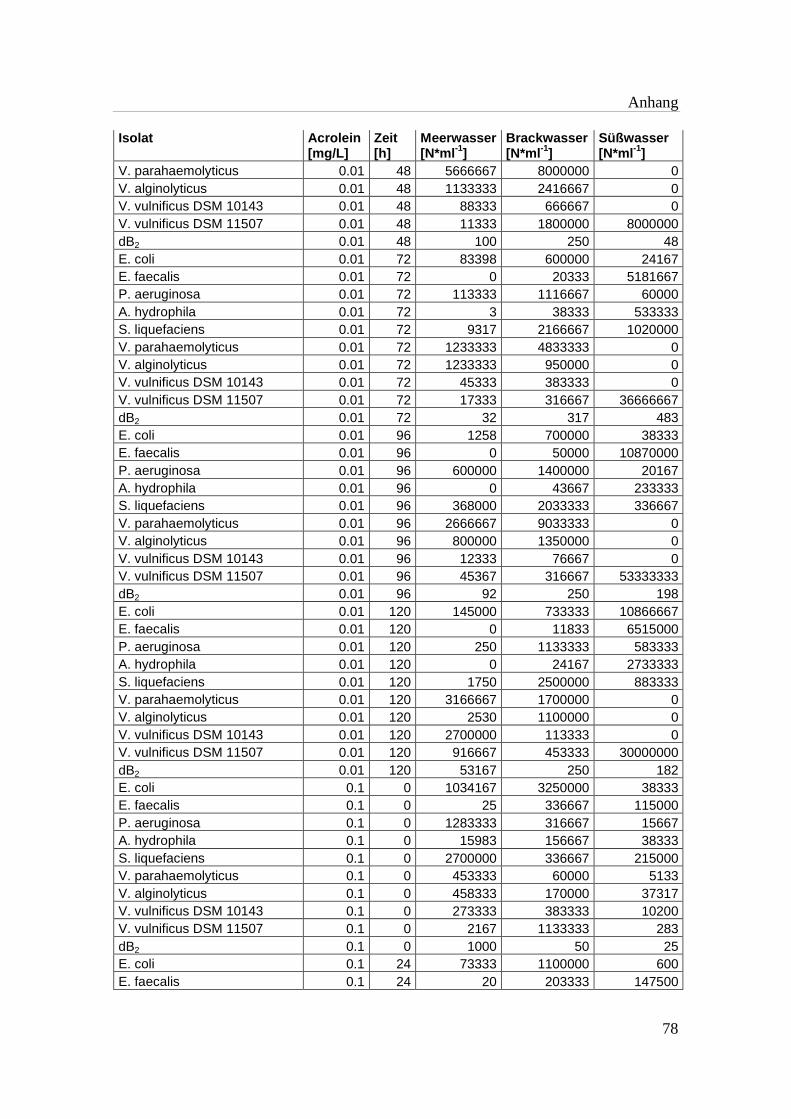
Anhang
78
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
V. parahaemolyticus 0.01 48 5666667 8000000 0 V. alginolyticus 0.01 48 1133333 2416667 0 V. vulnificus DSM 10143 0.01 48 88333 666667 0 V. vulnificus DSM 11507 0.01 48 11333 1800000 8000000 dB2 0.01 48 100 250 48 E. coli 0.01 72 83398 600000 24167 E. faecalis 0.01 72 0 20333 5181667 P. aeruginosa 0.01 72 113333 1116667 60000 A. hydrophila 0.01 72 3 38333 533333 S. liquefaciens 0.01 72 9317 2166667 1020000 V. parahaemolyticus 0.01 72 1233333 4833333 0 V. alginolyticus 0.01 72 1233333 950000 0 V. vulnificus DSM 10143 0.01 72 45333 383333 0 V. vulnificus DSM 11507 0.01 72 17333 316667 36666667 dB2 0.01 72 32 317 483 E. coli 0.01 96 1258 700000 38333 E. faecalis 0.01 96 0 50000 10870000 P. aeruginosa 0.01 96 600000 1400000 20167 A. hydrophila 0.01 96 0 43667 233333 S. liquefaciens 0.01 96 368000 2033333 336667 V. parahaemolyticus 0.01 96 2666667 9033333 0 V. alginolyticus 0.01 96 800000 1350000 0 V. vulnificus DSM 10143 0.01 96 12333 76667 0 V. vulnificus DSM 11507 0.01 96 45367 316667 53333333 dB2 0.01 96 92 250 198 E. coli 0.01 120 145000 733333 10866667 E. faecalis 0.01 120 0 11833 6515000 P. aeruginosa 0.01 120 250 1133333 583333 A. hydrophila 0.01 120 0 24167 2733333 S. liquefaciens 0.01 120 1750 2500000 883333 V. parahaemolyticus 0.01 120 3166667 1700000 0 V. alginolyticus 0.01 120 2530 1100000 0 V. vulnificus DSM 10143 0.01 120 2700000 113333 0 V. vulnificus DSM 11507 0.01 120 916667 453333 30000000 dB2 0.01 120 53167 250 182 E. coli 0.1 0 1034167 3250000 38333 E. faecalis 0.1 0 25 336667 115000 P. aeruginosa 0.1 0 1283333 316667 15667 A. hydrophila 0.1 0 15983 156667 38333 S. liquefaciens 0.1 0 2700000 336667 215000 V. parahaemolyticus 0.1 0 453333 60000 5133 V. alginolyticus 0.1 0 458333 170000 37317 V. vulnificus DSM 10143 0.1 0 273333 383333 10200 V. vulnificus DSM 11507 0.1 0 2167 1133333 283 dB2 0.1 0 1000 50 25 E. coli 0.1 24 73333 1100000 600 E. faecalis 0.1 24 20 203333 147500
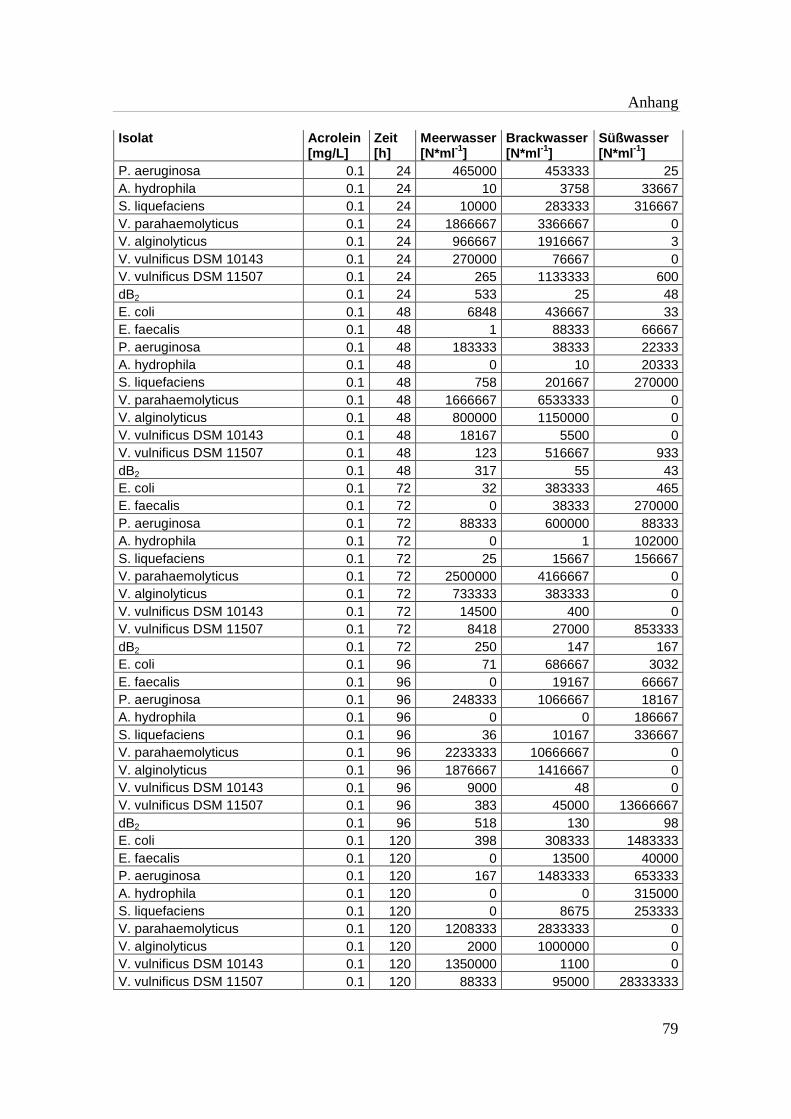
Anhang
79
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
P. aeruginosa 0.1 24 465000 453333 25 A. hydrophila 0.1 24 10 3758 33667 S. liquefaciens 0.1 24 10000 283333 316667 V. parahaemolyticus 0.1 24 1866667 3366667 0 V. alginolyticus 0.1 24 966667 1916667 3 V. vulnificus DSM 10143 0.1 24 270000 76667 0 V. vulnificus DSM 11507 0.1 24 265 1133333 600 dB2 0.1 24 533 25 48 E. coli 0.1 48 6848 436667 33 E. faecalis 0.1 48 1 88333 66667 P. aeruginosa 0.1 48 183333 38333 22333 A. hydrophila 0.1 48 0 10 20333 S. liquefaciens 0.1 48 758 201667 270000 V. parahaemolyticus 0.1 48 1666667 6533333 0 V. alginolyticus 0.1 48 800000 1150000 0 V. vulnificus DSM 10143 0.1 48 18167 5500 0 V. vulnificus DSM 11507 0.1 48 123 516667 933 dB2 0.1 48 317 55 43 E. coli 0.1 72 32 383333 465 E. faecalis 0.1 72 0 38333 270000 P. aeruginosa 0.1 72 88333 600000 88333 A. hydrophila 0.1 72 0 1 102000 S. liquefaciens 0.1 72 25 15667 156667 V. parahaemolyticus 0.1 72 2500000 4166667 0 V. alginolyticus 0.1 72 733333 383333 0 V. vulnificus DSM 10143 0.1 72 14500 400 0 V. vulnificus DSM 11507 0.1 72 8418 27000 853333 dB2 0.1 72 250 147 167 E. coli 0.1 96 71 686667 3032 E. faecalis 0.1 96 0 19167 66667 P. aeruginosa 0.1 96 248333 1066667 18167 A. hydrophila 0.1 96 0 0 186667 S. liquefaciens 0.1 96 36 10167 336667 V. parahaemolyticus 0.1 96 2233333 10666667 0 V. alginolyticus 0.1 96 1876667 1416667 0 V. vulnificus DSM 10143 0.1 96 9000 48 0 V. vulnificus DSM 11507 0.1 96 383 45000 13666667 dB2 0.1 96 518 130 98 E. coli 0.1 120 398 308333 1483333 E. faecalis 0.1 120 0 13500 40000 P. aeruginosa 0.1 120 167 1483333 653333 A. hydrophila 0.1 120 0 0 315000 S. liquefaciens 0.1 120 0 8675 253333 V. parahaemolyticus 0.1 120 1208333 2833333 0 V. alginolyticus 0.1 120 2000 1000000 0 V. vulnificus DSM 10143 0.1 120 1350000 1100 0 V. vulnificus DSM 11507 0.1 120 88333 95000 28333333
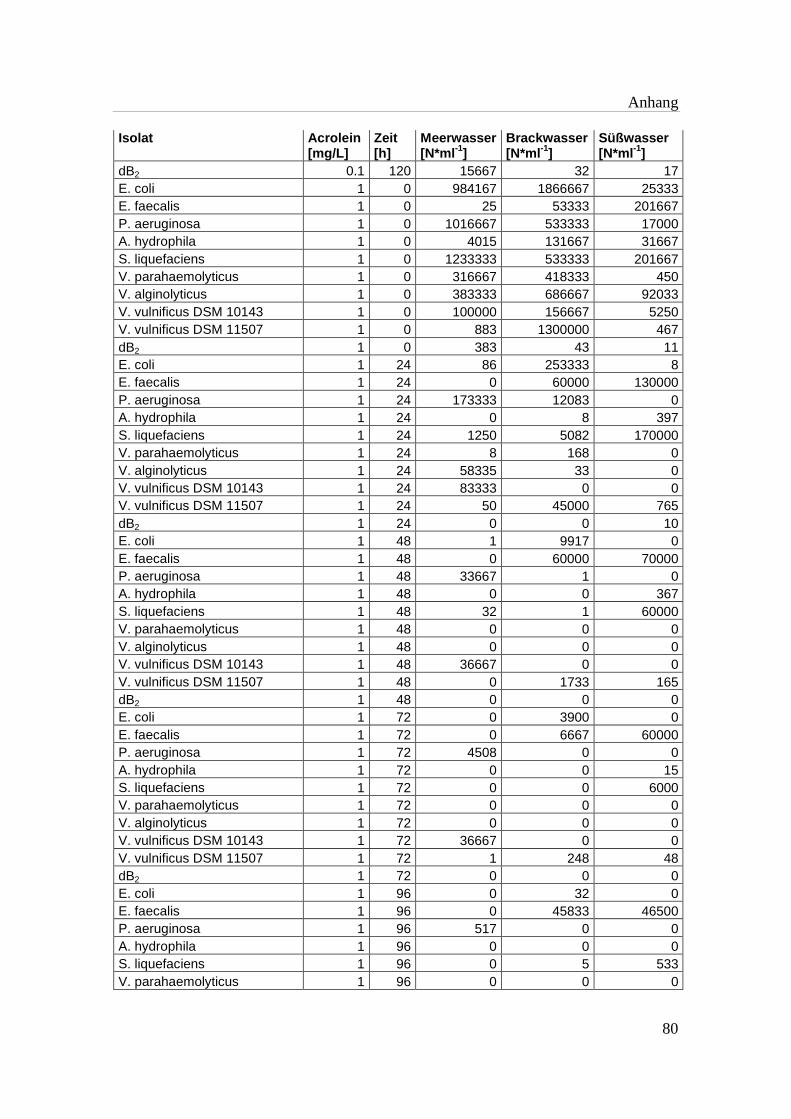
Anhang
80
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
dB2 0.1 120 15667 32 17 E. coli 1 0 984167 1866667 25333 E. faecalis 1 0 25 53333 201667 P. aeruginosa 1 0 1016667 533333 17000 A. hydrophila 1 0 4015 131667 31667 S. liquefaciens 1 0 1233333 533333 201667 V. parahaemolyticus 1 0 316667 418333 450 V. alginolyticus 1 0 383333 686667 92033 V. vulnificus DSM 10143 1 0 100000 156667 5250 V. vulnificus DSM 11507 1 0 883 1300000 467 dB2 1 0 383 43 11 E. coli 1 24 86 253333 8 E. faecalis 1 24 0 60000 130000 P. aeruginosa 1 24 173333 12083 0 A. hydrophila 1 24 0 8 397 S. liquefaciens 1 24 1250 5082 170000 V. parahaemolyticus 1 24 8 168 0 V. alginolyticus 1 24 58335 33 0 V. vulnificus DSM 10143 1 24 83333 0 0 V. vulnificus DSM 11507 1 24 50 45000 765 dB2 1 24 0 0 10 E. coli 1 48 1 9917 0 E. faecalis 1 48 0 60000 70000 P. aeruginosa 1 48 33667 1 0 A. hydrophila 1 48 0 0 367 S. liquefaciens 1 48 32 1 60000 V. parahaemolyticus 1 48 0 0 0 V. alginolyticus 1 48 0 0 0 V. vulnificus DSM 10143 1 48 36667 0 0 V. vulnificus DSM 11507 1 48 0 1733 165 dB2 1 48 0 0 0 E. coli 1 72 0 3900 0 E. faecalis 1 72 0 6667 60000 P. aeruginosa 1 72 4508 0 0 A. hydrophila 1 72 0 0 15 S. liquefaciens 1 72 0 0 6000 V. parahaemolyticus 1 72 0 0 0 V. alginolyticus 1 72 0 0 0 V. vulnificus DSM 10143 1 72 36667 0 0 V. vulnificus DSM 11507 1 72 1 248 48 dB2 1 72 0 0 0 E. coli 1 96 0 32 0 E. faecalis 1 96 0 45833 46500 P. aeruginosa 1 96 517 0 0 A. hydrophila 1 96 0 0 0 S. liquefaciens 1 96 0 5 533 V. parahaemolyticus 1 96 0 0 0
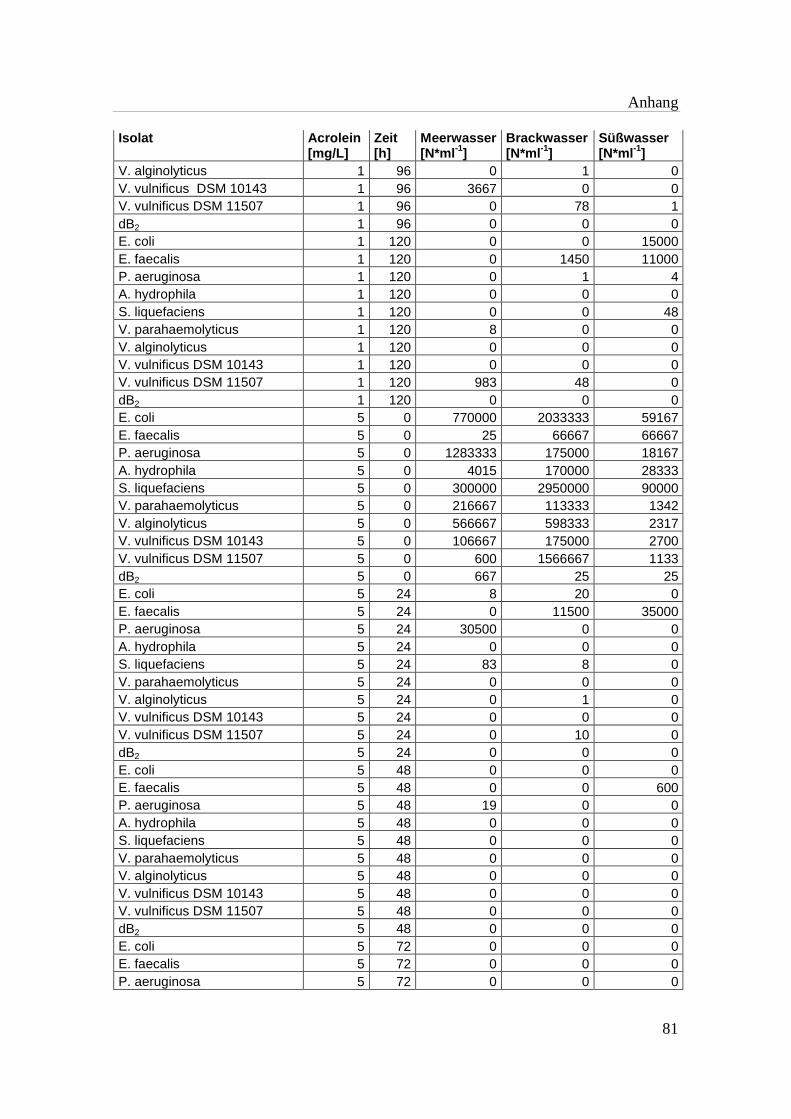
Anhang
81
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
V. alginolyticus 1 96 0 1 0 V. vulnificus DSM 10143 1 96 3667 0 0 V. vulnificus DSM 11507 1 96 0 78 1 dB2 1 96 0 0 0 E. coli 1 120 0 0 15000 E. faecalis 1 120 0 1450 11000 P. aeruginosa 1 120 0 1 4 A. hydrophila 1 120 0 0 0 S. liquefaciens 1 120 0 0 48 V. parahaemolyticus 1 120 8 0 0 V. alginolyticus 1 120 0 0 0 V. vulnificus DSM 10143 1 120 0 0 0 V. vulnificus DSM 11507 1 120 983 48 0 dB2 1 120 0 0 0 E. coli 5 0 770000 2033333 59167 E. faecalis 5 0 25 66667 66667 P. aeruginosa 5 0 1283333 175000 18167 A. hydrophila 5 0 4015 170000 28333 S. liquefaciens 5 0 300000 2950000 90000 V. parahaemolyticus 5 0 216667 113333 1342 V. alginolyticus 5 0 566667 598333 2317 V. vulnificus DSM 10143 5 0 106667 175000 2700 V. vulnificus DSM 11507 5 0 600 1566667 1133 dB2 5 0 667 25 25 E. coli 5 24 8 20 0 E. faecalis 5 24 0 11500 35000 P. aeruginosa 5 24 30500 0 0 A. hydrophila 5 24 0 0 0 S. liquefaciens 5 24 83 8 0 V. parahaemolyticus 5 24 0 0 0 V. alginolyticus 5 24 0 1 0 V. vulnificus DSM 10143 5 24 0 0 0 V. vulnificus DSM 11507 5 24 0 10 0 dB2 5 24 0 0 0 E. coli 5 48 0 0 0 E. faecalis 5 48 0 0 600 P. aeruginosa 5 48 19 0 0 A. hydrophila 5 48 0 0 0 S. liquefaciens 5 48 0 0 0 V. parahaemolyticus 5 48 0 0 0 V. alginolyticus 5 48 0 0 0 V. vulnificus DSM 10143 5 48 0 0 0 V. vulnificus DSM 11507 5 48 0 0 0 dB2 5 48 0 0 0 E. coli 5 72 0 0 0 E. faecalis 5 72 0 0 0 P. aeruginosa 5 72 0 0 0
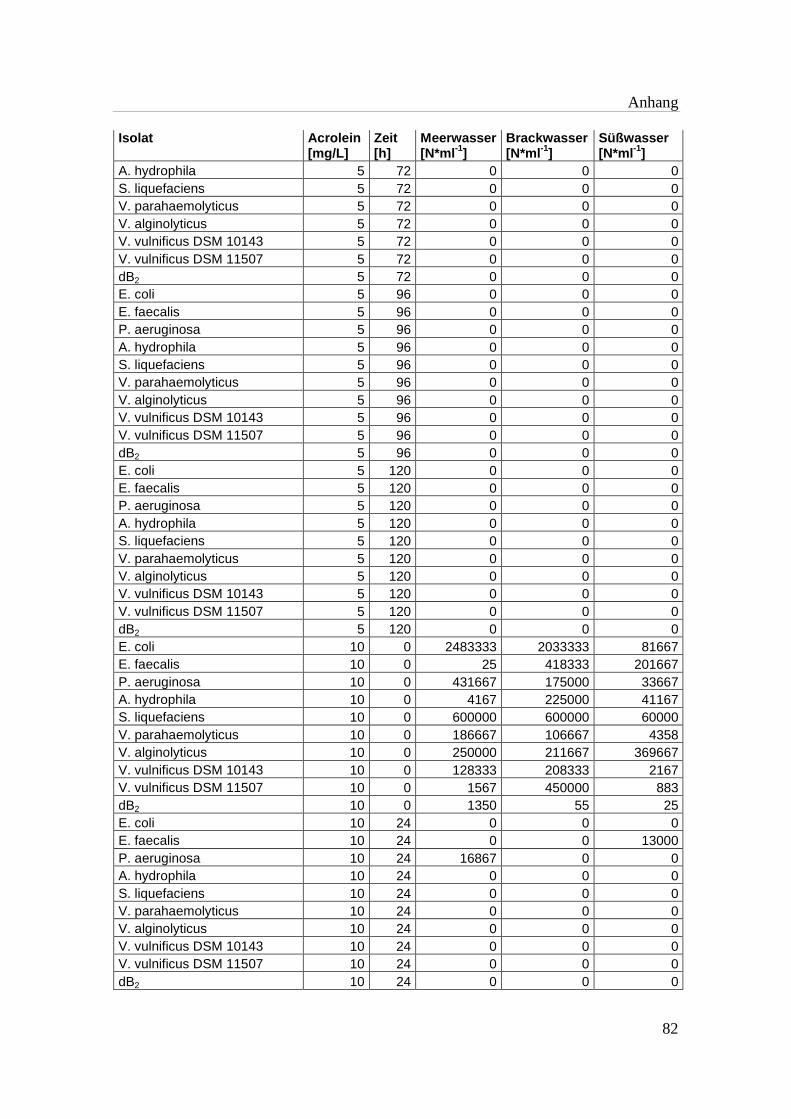
Anhang
82
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
A. hydrophila 5 72 0 0 0 S. liquefaciens 5 72 0 0 0 V. parahaemolyticus 5 72 0 0 0 V. alginolyticus 5 72 0 0 0 V. vulnificus DSM 10143 5 72 0 0 0 V. vulnificus DSM 11507 5 72 0 0 0 dB2 5 72 0 0 0 E. coli 5 96 0 0 0 E. faecalis 5 96 0 0 0 P. aeruginosa 5 96 0 0 0 A. hydrophila 5 96 0 0 0 S. liquefaciens 5 96 0 0 0 V. parahaemolyticus 5 96 0 0 0 V. alginolyticus 5 96 0 0 0 V. vulnificus DSM 10143 5 96 0 0 0 V. vulnificus DSM 11507 5 96 0 0 0 dB2 5 96 0 0 0 E. coli 5 120 0 0 0 E. faecalis 5 120 0 0 0 P. aeruginosa 5 120 0 0 0 A. hydrophila 5 120 0 0 0 S. liquefaciens 5 120 0 0 0 V. parahaemolyticus 5 120 0 0 0 V. alginolyticus 5 120 0 0 0 V. vulnificus DSM 10143 5 120 0 0 0 V. vulnificus DSM 11507 5 120 0 0 0 dB2 5 120 0 0 0 E. coli 10 0 2483333 2033333 81667 E. faecalis 10 0 25 418333 201667 P. aeruginosa 10 0 431667 175000 33667 A. hydrophila 10 0 4167 225000 41167 S. liquefaciens 10 0 600000 600000 60000 V. parahaemolyticus 10 0 186667 106667 4358 V. alginolyticus 10 0 250000 211667 369667 V. vulnificus DSM 10143 10 0 128333 208333 2167 V. vulnificus DSM 11507 10 0 1567 450000 883 dB2 10 0 1350 55 25 E. coli 10 24 0 0 0 E. faecalis 10 24 0 0 13000 P. aeruginosa 10 24 16867 0 0 A. hydrophila 10 24 0 0 0 S. liquefaciens 10 24 0 0 0 V. parahaemolyticus 10 24 0 0 0 V. alginolyticus 10 24 0 0 0 V. vulnificus DSM 10143 10 24 0 0 0 V. vulnificus DSM 11507 10 24 0 0 0 dB2 10 24 0 0 0
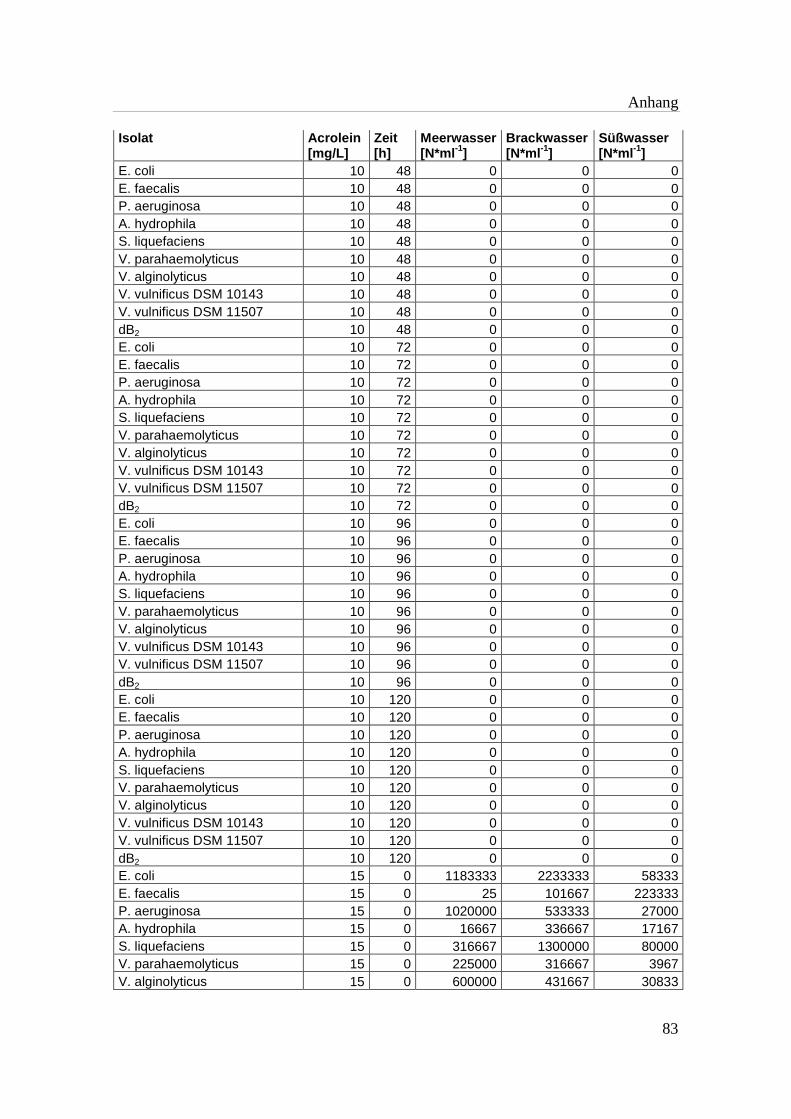
Anhang
83
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
E. coli 10 48 0 0 0 E. faecalis 10 48 0 0 0 P. aeruginosa 10 48 0 0 0 A. hydrophila 10 48 0 0 0 S. liquefaciens 10 48 0 0 0 V. parahaemolyticus 10 48 0 0 0 V. alginolyticus 10 48 0 0 0 V. vulnificus DSM 10143 10 48 0 0 0 V. vulnificus DSM 11507 10 48 0 0 0 dB2 10 48 0 0 0 E. coli 10 72 0 0 0 E. faecalis 10 72 0 0 0 P. aeruginosa 10 72 0 0 0 A. hydrophila 10 72 0 0 0 S. liquefaciens 10 72 0 0 0 V. parahaemolyticus 10 72 0 0 0 V. alginolyticus 10 72 0 0 0 V. vulnificus DSM 10143 10 72 0 0 0 V. vulnificus DSM 11507 10 72 0 0 0 dB2 10 72 0 0 0 E. coli 10 96 0 0 0 E. faecalis 10 96 0 0 0 P. aeruginosa 10 96 0 0 0 A. hydrophila 10 96 0 0 0 S. liquefaciens 10 96 0 0 0 V. parahaemolyticus 10 96 0 0 0 V. alginolyticus 10 96 0 0 0 V. vulnificus DSM 10143 10 96 0 0 0 V. vulnificus DSM 11507 10 96 0 0 0 dB2 10 96 0 0 0 E. coli 10 120 0 0 0 E. faecalis 10 120 0 0 0 P. aeruginosa 10 120 0 0 0 A. hydrophila 10 120 0 0 0 S. liquefaciens 10 120 0 0 0 V. parahaemolyticus 10 120 0 0 0 V. alginolyticus 10 120 0 0 0 V. vulnificus DSM 10143 10 120 0 0 0 V. vulnificus DSM 11507 10 120 0 0 0 dB2 10 120 0 0 0 E. coli 15 0 1183333 2233333 58333 E. faecalis 15 0 25 101667 223333 P. aeruginosa 15 0 1020000 533333 27000 A. hydrophila 15 0 16667 336667 17167 S. liquefaciens 15 0 316667 1300000 80000 V. parahaemolyticus 15 0 225000 316667 3967 V. alginolyticus 15 0 600000 431667 30833
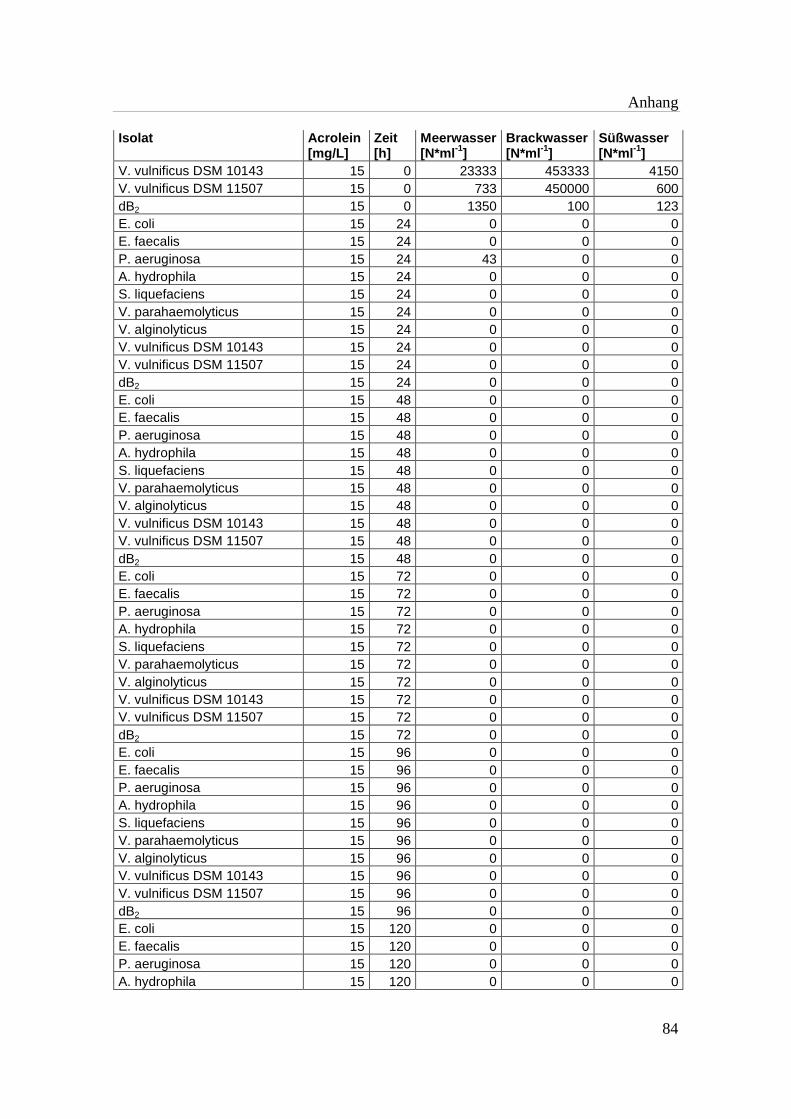
Anhang
84
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
V. vulnificus DSM 10143 15 0 23333 453333 4150 V. vulnificus DSM 11507 15 0 733 450000 600 dB2 15 0 1350 100 123 E. coli 15 24 0 0 0 E. faecalis 15 24 0 0 0 P. aeruginosa 15 24 43 0 0 A. hydrophila 15 24 0 0 0 S. liquefaciens 15 24 0 0 0 V. parahaemolyticus 15 24 0 0 0 V. alginolyticus 15 24 0 0 0 V. vulnificus DSM 10143 15 24 0 0 0 V. vulnificus DSM 11507 15 24 0 0 0 dB2 15 24 0 0 0 E. coli 15 48 0 0 0 E. faecalis 15 48 0 0 0 P. aeruginosa 15 48 0 0 0 A. hydrophila 15 48 0 0 0 S. liquefaciens 15 48 0 0 0 V. parahaemolyticus 15 48 0 0 0 V. alginolyticus 15 48 0 0 0 V. vulnificus DSM 10143 15 48 0 0 0 V. vulnificus DSM 11507 15 48 0 0 0 dB2 15 48 0 0 0 E. coli 15 72 0 0 0 E. faecalis 15 72 0 0 0 P. aeruginosa 15 72 0 0 0 A. hydrophila 15 72 0 0 0 S. liquefaciens 15 72 0 0 0 V. parahaemolyticus 15 72 0 0 0 V. alginolyticus 15 72 0 0 0 V. vulnificus DSM 10143 15 72 0 0 0 V. vulnificus DSM 11507 15 72 0 0 0 dB2 15 72 0 0 0 E. coli 15 96 0 0 0 E. faecalis 15 96 0 0 0 P. aeruginosa 15 96 0 0 0 A. hydrophila 15 96 0 0 0 S. liquefaciens 15 96 0 0 0 V. parahaemolyticus 15 96 0 0 0 V. alginolyticus 15 96 0 0 0 V. vulnificus DSM 10143 15 96 0 0 0 V. vulnificus DSM 11507 15 96 0 0 0 dB2 15 96 0 0 0 E. coli 15 120 0 0 0 E. faecalis 15 120 0 0 0 P. aeruginosa 15 120 0 0 0 A. hydrophila 15 120 0 0 0
Anhang
85
Isolat Acrolein [mg/L]
Zeit [h]
Meerwasser [N*ml -1]
Brackwasser [N*ml -1]
Süßwasser [N*ml -1]
S. liquefaciens 15 120 0 0 0 V. parahaemolyticus 15 120 0 0 0 V. alginolyticus 15 120 0 0 0 V. vulnificus DSM 10143 15 120 0 0 0 V. vulnificus DSM 11507 15 120 0 0 0 dB2 15 120 0 0 0

Abkürzungsverzeichnis
86
Abkürzungsverzeichnis
°C Grad Celsius
Abb. Abbildung
BSH Bundesamt für Seeschifffahrt und Hydrographie
Bzw beziehungsweise
cfu colony forming units
DSMZ Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH
ggf gegebenenfalls
h Stunden
HUS hemolytic uremic syndrome
IDLH immediately dangerous to life and health
IMO International Maritime Organisation
mg/L Milligramm pro Liter
Mio. Millionen
µm Mikrometer
N*ml -1 Zellen pro Milliliter
nm Nanometer
OD Optische Dichte
ppm parts per million
ppt parts per trillion
PSU practical salinity units
t0 – t5 Zeitpunkt 0 (= 0 Stunden) bis Zeitpunkt 5 (= 120 Stunden)
Tab. Tabelle

Abkürzungsverzeichnis
87
TTC 2,3,5-Triphenyltetrazoliumchlorid
u. a. unter anderem
VBNC viable but not culturable
z.B. zum Beispiel

Danksagung
88
Danksagung
An dieser Stelle möchte ich all den Diplomanden, Doktoranden, Praktikanten,
Haustechnikern, Aquarianern und allen anderen, die ich näher kennenlernen durfte,
ganz herzlich danken, für eine wunderbar abwechslungsreiche Zeit, und dass sie es
immer mit mir ausgehalten haben!
Der erste Dank gilt Gunnar Gerdts und Antje Wichels, für die Betreuung, für die
Themenstellung, für „meine“ Sterilbank, für Lob, Kritik und Korrektur.
Danke schön an Herrn Meinhard Simon, der als Professor der Uni Oldenburg meine
Arbeit betreute.
Für Korrekturen beim Schreibprozess danke ich außerdem Vroni Kuscha, Sonja
Oberbeckmann und Christina Gebühr recht herzlich. Außerdem danke ich Sonja für
Hilfe wenn ich Fragen hatte und bei der Suche nach dem roten Faden.
Vielen Dank an Kristine Carstens von der AG Biologische Ozeanographie für die
Bestimmung der Salzkonzentrationen meiner Wasserproben.
Vielen Dank an Judith Ficker und Simone Gwosdz für die Extraktion, Vervielfältigung
und Sequenzierung der DNA von V. vulnificus DSM 11507.
Vielen Dank an meine AG, für die gemeinsame Zeit, für Hilfe, Feedback und
Mittagspausen, besonders an die „Diplomanden und anderen Protozoen“ in Büro C-38.
Unbezahlbar war vor allem die private Unterstützung in all der Zeit:
Das dickste Dankeschön gilt Simone Gwosdz. Für Spitzen stecken, überstandenen
Inselkoller, für zahlreiches “Käffchen trinken”, für Hilfe in Not, für gemeinsames Leid
und gemeinsame Freud, dafür dass ich zum Ende hin nicht alleine dem
sozialinkompetenten Bananarama mit Verlust des Artikulationsvermögens verfallen
musste, für die richtige Würze im drögen Alltag.
Für eine schöne und sehr abwechslungsreiche Zeit danke ich meinen zahlreichen
unterschiedlichen Mitbewohnern im “Haggi”. Für überstandene Küchen-Krisen, für
viele unterhaltsame und lustige Abende im Wohnzimmer (und im Krebs) und für die
netten Leute die ich dort kennen lernen durfte. Für drei Monate im Kasernen-Haus

Danksagung
89
„Blakfesk“ danke ich Maike Hoffmann für eine ruhige Zeit und Jan Sdrenka für die
lauten aber sehr unterhaltsamen Phasen zwischendurch, auf die ich nicht hätte
verzichten wollen.
Ganz besonderer Dank gilt meinen Eltern und meinem Bruder Christian, fürs “immer
für mich da sein”, für Heimat-feeling, für mehr oder weniger Verständnis für alles und
für die finanzielle Unterstützung, ohne die mein Studium nie so wunderschön und lange
hätte werden können.
Tausend Dank an meine Freunde „auf dem Festland“, ganz besonders Steffi Feldmann
und Katrin Noack, die mich jederzeit beraten und unterstützen, und meinem
Oldenburger Mitbewohner Patrick Zägel, besonders fürs Blumen-gießen-gegen-
Whiskey-Import.

Erklärung
90
Erklärung
Hiermit versichere ich, dass ich diese Arbeit selbständig verfasst und keine anderen als
die angegebenen Hilfsmittel und Quellen benutzt habe.
Helgoland, den 09.12.2008
Andrea Fuchs