Download - 0811125 DISS-7 4



Bibliografische Informationen der Deutschen Bibliothek
Die Deutsche Bibliothek verzeichnet diese Publikation in der Deutschen Nationalbibliografie; Detaillierte bibliografische Daten sind im Internet über http://dnb.ddb.de abrufbar.
1. Auflage 2008
© 2008 by Verlag: Deutsche Veterinärmedizinische Gesellschaft Service GmbH, Gießen Printed in Germany
ISBN 978-3-939902-84-3
Verlag: DVG Service GmbH Friedrichstraße 17
35392 Gießen 0641/24466
[email protected] www.dvg.net

Tierärztliche Hochschule Hannover
Anatomisches Institut
Detailed analysis of immunoprivileged tissues in
skin appendages
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
- Doctor medicinae veterinariae -
(Dr. med. vet.)
Vorgelegt von
Katja Christina Meyer
(Hamburg)
Hannover 2008

2
Wissenschaftliche Betreuung: Univ. Prof. Dr. rer. nat. habil. Wilfried Meyer
Stiftung Tierärztliche Hochschule Hannover
Anatomisches Institut
Histologie und Embryologie
Bischofsholer Damm 15
30173 Hannover
Univ. Prof. Dr. med. habil. Ralf Paus
Universität zu Lübeck
Klinik für Dermatologie, Allergologie und Venerologie
Ratzeburger Allee 160
23538 Lübeck
1. Gutachter: Univ. Prof. Dr. rer. nat. habil. Wilfried Meyer
2. Gutachter: Univ. Prof. Dr. med. vet. habil. Marion Hewicker-Trautwein
Tag der mündlichen Prüfung: 18. November 2008

In liebevollem Gedenken an meinen Vater


TABLE OF CONTENTS
Table of contents
Table of contents _________________________________________________________ 5
Abbreviations ____________________________________________________________ 9
Figures _________________________________________________________________ 12
Tables __________________________________________________________________ 16
1 INTRODUCTION _____________________________________________________ 17
2 LITERATURE ________________________________________________________ 20
2.1 A short synthesis of hair follicle biology ________________________________ 20
2.1.1 Hair follicle morphogenesis___________________________________________________ 22
2.1.2 Functional anatomy of the hair follicle __________________________________________ 24
2.1.3 Hair follicle cycle____________________________________________________________ 31
2.2 Sinus hair follicle biology: overview ____________________________________ 35
2.2.1 Sinus hair follicle morphogenesis______________________________________________ 36
2.2.2 Functional anatomy of the sinus hair follicle_____________________________________ 38
1.1.1 Sinus hair follicle cycle_______________________________________________________ 42
2.3 Murine nail apparatus: overview________________________________________ 44
2.3.1 Nail morphogenesis _________________________________________________________ 45
2.3.2 Functional anatomy of the nail ________________________________________________ 46
2.3.3 Growth of the nail ___________________________________________________________ 50
2.4 Immunological background for the current study ________________________ 51
2.4.1 Innate immune system_______________________________________________________ 51
2.4.2 Acquired immune system ____________________________________________________ 52
2.4.3 Immune privilege: Definition and basic characteristics ____________________________ 53
2.5 Immune privilege in skin appendages___________________________________ 56
2.5.1 The anagen hair bulb as an immunoprivileged site_______________________________ 59
2.5.2 Downregulation of MHC class Ia and NK cells___________________________________ 61
2.5.3 Infantile human nail matrix is a site of relative immune privilege ___________________ 64
2.6 The immune privilege collapse model of AA pathogenesis _______________ 67
Collapse and restoration of IP in the anagen hair bulb in vitro __________________________ 69
2.7 The human bulge region as a site of relative immune privilege? __________ 71

TABLE OF CONTENTS
2.8 Bulge IP collapse and the pathogenesis of PCA _________________________ 73
2.9 Function of immunoprivileged sites ____________________________________ 75
2.10 Immune privilege markers _____________________________________________ 77
2.10.1 MHC class I molecules ____________________________________________________ 78
2.10.2 β2-microglobulin__________________________________________________________ 81
2.10.3 MHC class II molecules ___________________________________________________ 82
2.10.4 MHC class Ib molecules ___________________________________________________ 84
2.10.5 CD4+ and CD8+ T cells ___________________________________________________ 85
2.10.6 α-MSH and ACTH ________________________________________________________ 86
2.10.7 TGF-β __________________________________________________________________ 89
2.10.8 MIF_____________________________________________________________________ 91
2.10.9 IDO_____________________________________________________________________ 92
2.10.10 CD200 __________________________________________________________________ 95
2.10.11 Mast cells _______________________________________________________________ 96
2.10.12 ICAM-1 ________________________________________________________________ 100
2.10.13 β-defensin 2 ____________________________________________________________ 101
2.11 K(D)PT – a candidate as hair growth modulator and IP restorer in anagen hair
bulbs? _____________________________________________________________________ 103
2.12 Questions addressed in this study ____________________________________ 105
2.13 Experimental design _________________________________________________ 105
3 Materials and methods ______________________________________________ 107
3.1 Tissue collection_____________________________________________________ 107
3.1.1 Human tissue collection_____________________________________________________ 107
3.1.2 Murine tissue collection _____________________________________________________ 108
3.2 Human hair follicle isolation __________________________________________ 110
3.3 Human HF organ culture______________________________________________ 111
3.4 Full thickness human scalp skin organ culture _________________________ 114
3.5 Histological stainings ________________________________________________ 115
3.5.1 Hematoxylin-Eosin staining__________________________________________________ 115
3.5.2 Trichromatic staining _______________________________________________________ 116
3.5.3 Toluidine blue staining ______________________________________________________ 116
3.5.4 Leder`s esterase staining ___________________________________________________ 117

TABLE OF CONTENTS
3.5.5 Ki-67 / TUNEL_____________________________________________________________ 118
3.6 Immunohistochemistry _______________________________________________ 120
3.6.1 Primary antibodies _________________________________________________________ 120
3.6.2 Avidin Biotin Complex-Peroxidase____________________________________________ 121
3.6.3 EnVision®-alkaline phosphatase _____________________________________________ 123
3.6.4 Immunofluorescence _______________________________________________________ 123
3.6.5 Tyramide signal amplification (TSA) __________________________________________ 124
3.7 Histomorphometry ___________________________________________________ 125
3.7.1 Assessment of hair cycle stages _____________________________________________ 125
3.7.2 Assessment of proliferating matrix keratinocytes _______________________________ 125
3.7.3 Assessment of immunostaining intensity ______________________________________ 126
3.7.4 Assessment of MHC class I IR in isolated HFs _________________________________ 127
3.7.5 Assessment of mast cells ___________________________________________________ 128
3.7.6 Assessment of MHC class II, CD4, CD54, CD11b, mast cells and c-kit positive cells 128
3.7.7 Microscopical equipment____________________________________________________ 129
3.7.8 Statistical analysis _________________________________________________________ 129
4 Results ____________________________________________________________ 131
4.1 Immune privilege and the human hair follicle bulge _____________________ 131
4.1.1 Demonstration of MHC class Ia and β2-microglobulin expression on CD200+ cells __ 131
4.1.2 Demonstration of MHC class II+ cells in the bulge ______________________________ 137
4.1.3 Demonstration of HLA-E expression in CD200+ bulge cells ______________________ 140
4.1.4 Locally generated immunosuppressants complement the bulge IP ________________ 141
4.1.5 Demonstration of macrophage migration inhibitory factor and indoleamine-2,3-
dioxygenase______________________________________________________________________ 144
4.1.6 Influence of IFN-γ on ectopic MHC class I protein expression in the bulge ORS_____ 149
4.2 Influence of the α-MSH related tripeptide K(D)PT on human hair follicle
biology in situ under pro-inflammatory conditions ____________________________ 151
4.2.1 Influence of K(D)PT on IFN-γ induced MHC class I and II protein expression _______ 151
4.2.2 Reaction of K(D)PT on the IFN-γ induced upregulation of total mast cell numbers and
mast cell degranulation ____________________________________________________________ 155
4.2.3 Influence of K(D)PT on the hair cycle _________________________________________ 157
4.2.4 Influence of K(D)PT on hair matrix keratinocyte proliferation and apoptosis ________ 158
4.3 Immune privilege and murine sinus hair follicles _______________________ 159
4.3.1 Demonstration of MHC class I molecules______________________________________ 159
4.3.2 Demonstration of MHC class II molecules _____________________________________ 161

TABLE OF CONTENTS
4.3.3 Demonstration of CD4+ T cells ______________________________________________ 163
4.3.4 Demonstration of CD11b molecules __________________________________________ 165
4.3.5 Demonstration of TGF-β1 molecules__________________________________________ 167
4.3.6 Demonstration of mast cells _________________________________________________ 169
4.4 Immune privilege and the murine mouse nail apparatus_________________ 171
4.4.1 Demonstration of MHC class I molecules______________________________________ 171
4.4.2 Demonstration of MHC class II molecules _____________________________________ 173
4.4.3 Demonstration of CD4+ T cells ______________________________________________ 175
4.4.4 Demonstration of TGF-β1 molecules__________________________________________ 176
4.4.5 Demonstration of CD54 molecules ___________________________________________ 177
4.4.6 Demonstration of mast cells _________________________________________________ 178
4.4.7 Demonstration of β-defensin 2 _______________________________________________ 180
5 Discussion _________________________________________________________ 181
5.1 Introductory remarks _________________________________________________ 181
5.2 Methods employed ___________________________________________________ 182
5.3 Immune privilege in the human hair follicle bulge, murine nail and sinus hair
follicle _____________________________________________________________________ 186
5.3.1 The human HF bulge _______________________________________________________ 186
5.3.2 The murine sinus hair follicle and nail apparatus________________________________ 193
5.3.3 Comparison of the human HF bulge, murine sinus hair follicle and nail apparatus ___ 200
5.4 Effects of K(D)PT on the hair follicle immune system ___________________ 202
5.5 Conclusions _________________________________________________________ 204
5.6 Perspectives_________________________________________________________ 205
6 Summary___________________________________________________________ 207
7 Zusammenfassung _________________________________________________ 209
8 References _________________________________________________________ 212
9 Annex______________________________________________________________ 257

ABBREVIATIONS
Abbreviations
AA Alopecia areata
α-MSH Alpha-melanocyte stimulating hormone
ABC Avidin-biotin complex
AMP Antimicrobial peptides
AP Alkaline phosphatases
APC Antigen presenting cell
APM Arrector pili muscle
BM Basement membrane
ECM Extracellular matrix
eSC Epithelial stem cell
CAP Cationic antimicrobial peptide
CD Cluster of differentiation
CDLE Chronic discoid lupus erythematosus
CGRP Calcitonine-gene related peptide
CK Cytokeratin
Col Collagen
CTS Connective tissue sheath
DAB 3,3’-diaminobenzidine
DAPI 4’,6-diamidin-2’-phenylindol-dihydrochlorid
DC Dendritic cell
DP Dermal papilla
DTH Delayed type hyperpsensitivity
EAE Experimental autoimmune encephalitis
FGF Fibroblast growth factor
Fig Figure
FITC Fluorescein isothiocyanate
FT Follicular trochanter
GFP Green fluorescent protein

ABBREVIATIONS
h hour
HF
HLA
HM
Hair follicle
Human leucocyte antigen
Hair matrix
HS Hair shaft
ICAM Intraepithelial cellular adhesion molecule
IDO 2,3 indoleamine-dioxygenase
IFN-γ Interferon-gamma
IP Immune privilege
IR Immunoreactivity
IRS Inner root sheath
ITIM Immunoreceptor tyrosine inhibitory motif
KIR Killer cell immunoglobulin-like receptor
Kit CD117
LC Langerhans cell
MBP Myelin basic protein
MC Melanocortin
MC-R Melanocortin receptor
MHC Major histocompatibility complex
MICA MHC class I chain-related A
min Minutes
MIF Macrophage migration inhibitory factor
Mitf Microphthalmia-associated transcription factor
MK Matrix keratinocyte
mSC Mesenchymal stem cell
NaCl Sodium chloride
NaOH Sodium hydroxide
NFκκκκB Nuclear factor of kappa light polypeptide gene enhancer in B-cells
NK Natural killer cell
ORS Outer root sheath

ABBREVIATIONS
PAMP Pathogen-associated molecular patterns
PPR Pattern recognition receptors
PBS Phosphate buffered saline
PCA Primary cicatricial alopecia
PICS Perifollicular inflammatory cell clusters
PNF Proximal nail fold
PNM Proximal nail matrix
POD Programmed organ deletion
POMC Pro-opiomelanocortin
RER Rough endoplasmatic reticulum
SEM Standard error of the mean
SG Sebaceous gland
SW Sweat gland
Tab Table
TAP Transporter in antigen presentation
TBS Tris buffered saline
TCR T cell receptor
TGF-β Transforming growth factor β
TLR Toll like receptor
TSA Tyramide signal amplification
VIP Vasointestinal peptide

FIGURES
Figures
Fig. 2.1 Three dimensional diagram of the mammalian skin (FUCHS 2007)........ 20
Fig. 2.2 Terminal human HF in anagen VI ............................................................ 25
Fig. 2.3 The human HF bulge ............................................................................... 26
Fig. 2.4 HF: Keratinocyte lineages and structure.................................................. 27
Fig. 2.5 Schematic drawing of a hair bulb............................................................. 28
Fig. 2.6 Morphology of human HFs in different hair cycle stages. ........................ 32
Fig. 2.7 The hair follicle cycle. .............................................................................. 33
Fig. 2.8 Diagram of active vibrissal follicle in adult mouse.................................... 36
Fig. 2.9 Diagram of stages 1-8 in development of vibrissal follicles in mouse. ..... 37
Fig. 2.10 Murine vibrissal follicles of the snout. ...................................................... 40
Fig. 2.11 Musculature of vibrissal follicles............................................................... 42
Fig. 2.12 Mouse vibrissal follicle cycle.................................................................... 43
Fig. 2.13 Main characteristics of the nail in different animals and humans............. 46
Fig. 2.14 Schematic longitudinal section of mouse hair follicle and nail unit........... 47
Fig. 2.15 Murine nail apparatus. ............................................................................. 48
Fig. 2.16 Diagram of innate and adaptive immune system..................................... 53
Fig. 2.17 Billingham’s experiment: Survival of epidermal melanocyte allotransplants
in the host anagen HF. ............................................................................ 58
Fig. 2.18 The anagen hair bulb............................................................................... 59
Fig. 2.19 Distribution of peri- and intrafollicular CD4+, CD8+ T cells and CD1a+
cells. ........................................................................................................ 60
Fig. 2.20 Activation of NK cell activity..................................................................... 62
Fig. 2.21 The similarity of anatomical structure between HF and nail..................... 64
Fig. 2.22 Distribution of HLA-ABC, CD4+ and CD8+ T cells in the murine nail
apparatus (ITO et al. 2008b)................................................................... 65
Fig. 2.23 ´Immune privilege collapse model` of alopecia areata pathogenesis....... 67
Fig. 2.24 The human HF bulge region .................................................................... 72
Fig. 2.25 Hypothesized model of the pathogenesis of cicatricial alopecia .............. 74
Fig. 2.26 The cycle-dependency of murine HF MHC class I antigen ...................... 80

FIGURES
Fig. 2.27 The expression of MHC class I pathway molecules on the murine anagen
HF............................................................................................................ 82
Fig. 2.28 The different roles of α-MSH as immunomodulator (LUGER et al. 2000) 87
Fig. 2.29 Molecular mechanisms of IDO-induced immunosuppression .................. 94
Fig. 2.30 Stimulatory and immunosuppressive functions of mast cells. .................. 98
Fig. 2.31 Biosynthesis of POMC peptides an natural melanocortins. ................... 104
Fig. 3.1 Human scalp skin specimen .................................................................. 107
Fig. 3.2 solation of human HFs........................................................................... 111
Fig. 3.3 Hair follicles in a 24-well plate ............................................................... 112
Fig. 3.4 Isolated hair follicles .............................................................................. 112
Fig. 3.5 Experimental design of the HF organ culture......................................... 113
Fig. 3.6 Full thickness human scalp skin punch biopsies in 6-well multi well plate
........................................................................................................................ 114
Fig. 3.7 Auber’s line marked in the human HF.................................................... 126
Fig. 3.8 Reference areas for the quantitative analysis of MHC class I IR ........... 127
Fig. 4.1 IR is downregulated in the human bulge region for MHC class I ........... 132
Fig. 4.2 Schematic drawing of MHC class I IR pattern ....................................... 133
Fig. 4.3 Quantitative immunohistochemistry for MHC class I.............................. 133
Fig. 4.4 MHC class I IR is downregulated in CD200+ bulge ORS cells .............. 134
Fig. 4.5 Schematic drawing of CD200 IR pattern................................................ 135
Fig. 4.6 IR in the human hair follicle for β2-microglobulin.................................... 136
Fig. 4.7 Schematic drawing of β2-microglobulin IR pattern ................................. 136
Fig. 4.8 Quantitative immunohistochemistry for β2-microglobulin ....................... 137
Fig. 4.9 IR in the human bulge region for MHC class II ...................................... 138
Fig. 4.10 MHC class II+ cells in the human hair follicle and human bulge region . 139
Fig. 4.11 Schematic drawing of MHC class II IR pattern ...................................... 139
Fig. 4.12 HLA-E expression by CD200+ cells in the human bulge, double
Immunofluorescence ............................................................................. 140
Fig. 4.13 Schematic drawing of MHC class II IR pattern ...................................... 140
Fig. 4.14 IR in the human bulge region for α-MSH ............................................... 141
Fig. 4.15 TGF-β2 is expressed in the bulge ORS................................................. 142

FIGURES
Fig. 4.16 Schematic drawing of α-MSH IR pattern................................................ 143
Fig. 4.17 Schematic drawing of TGF-β2 IR pattern .............................................. 143
Fig. 4.18 IR is upregulated in the human bulge region for MIF............................. 144
Fig. 4.19 Quantitative immunohistochemistry for MIF........................................... 145
Fig. 4.20 IR is upregulated in the human bulge region for IDO............................. 146
Fig. 4.21 IR in different parts of the proximal ORS for IDO................................... 147
Fig. 4.22 Quantitative immunohistochemistry for IDO .......................................... 147
Fig. 4.23 Schematic drawings of MIF and IDO IR pattern .................................... 148
Fig. 4.24 Influence of IFN-γ on MHC class I expression of human HFs in full
thickness organ culture ......................................................................... 149
Fig. 4.25 Quantitative analysis of MHC class I staining intensity in the bulge....... 150
Fig. 4.26 Isolated HFs stained for MHC class I..................................................... 151
Fig. 4.27 Quantitative immunohistochemistry for MHC class I in isolated treated HFs
........................................................................................................................ 152
Fig. 4.28 Quantitative analysis of the mean number of MHC class II+ cells ......... 153
Fig. 4.29 Staining intensity of MHC class II+ cells ................................................ 154
Fig. 4.30 Staining for mast cells............................................................................ 155
Fig. 4.31 Quantitative analysis of the number of mast cells and their degranulation
ratio ....................................................................................................... 156
Fig. 4.32 Influence of K(D)PT on the hair cycle stage .......................................... 157
Fig. 4.33 Ki-67 / TUNEL staining on human HFs.................................................. 158
Fig. 4.34 IR in the murine vibrissal follicle for MHC class I ................................... 159
Fig. 4.35 Quantitative immunohistochemistry for MHC class I.............................. 160
Fig. 4.36 Schematic drawing of MHC class I IR pattern ....................................... 160
Fig. 4.37 Schematic drawing of MHC class II IR pattern ...................................... 161
Fig. 4.38 MHC class II positive cells in the murine vibrissal follicle....................... 162
Fig. 4.39 IR in the murine vibrissal follicle for CD4+ T cells.................................. 163
Fig. 4.40 Quantitative immunohistochemistry for CD4.......................................... 164
Fig. 4.41 Schematic drawing of CD4 pattern ........................................................ 164
Fig. 4.42 IR in the murine vibrissal follicle for CD11b ........................................... 165
Fig. 4.43 Quantitative immunohistochemistry for CD11b...................................... 166

FIGURES
Fig. 4.44 Schematic drawing of CD11b IR pattern................................................ 166
Fig. 4.45 IR in the murine vibrissal follicle for TGF-β1 on cryosections ................ 167
Fig. 4.46 IR in the murine vibrissal follicle for TGF-β1 on Bouin fixated specimens
........................................................................................................................ 168
Fig. 4.47 Schematic drawing of TGF-β1 pattern................................................... 168
Fig. 4.48 Demonstration of mast cells in the murine vibrissal follicle .................... 169
Fig. 4.49 Quantitative immunohistochemistry for mast cells ................................. 170
Fig. 4.50 Schematic drawing of mast cell expression pattern............................... 170
Fig. 4.51 IR in the murine nail apparatus for MHC class I .................................... 171
Fig. 4.52 Quantitative immunohistochemistry for MHC class I.............................. 172
Fig. 4.53 Schematic drawing of MHC class I expression pattern.......................... 172
Fig. 4.54 IR in the murine nail apparatus for MHC class II ................................... 173
Fig. 4.55 Quantitative immunohistochemistry for MHC class II............................. 174
Fig. 4.56 Schematic drawing of MHC class II expression pattern......................... 174
Fig. 4.57 IR in the murine nail apparatus for CD4+ T cells ................................... 175
Fig. 4.58 Schematic drawing of CD4 expression pattern...................................... 175
Fig. 4.59 IR in the murine nail apparatus for TGF-β1 ........................................... 176
Fig. 4.60 Schematic drawing of TGF-β1 expression pattern................................. 176
Fig. 4.61 IR in the murine nail apparatus for CD54............................................... 177
Fig. 4.62 Schematic drawing of CD54 expression pattern.................................... 177
Fig. 4.63 IR in the murine nail apparatus for mast cells........................................ 178
Fig. 4.64 Quantitative immunohistochemistry for MHC class I.............................. 179
Fig. 4.65 Schematic drawing of mast cell expression pattern............................... 179
Fig. 4.66 IR in the murine nail apparatus for β-defensin 2 .................................... 180
Fig. 4.67 Schematic drawing of β-defensin 2 expression pattern ......................... 180

TABLES
Tables
Tab. 2.1 Basic data on human HFs ....................................................................... 21
Tab. 2.2 HF morphogenesis in mice...................................................................... 23
Tab. 2.3 Glossary of anatomical and trichology terms........................................... 30
Tab. 2.4 Human nail apparatus – basic data ......................................................... 49
Tab. 3.1 Primary antibodies and secondary detection systems........................... 120
Tab. 3.2 Primary anti-mouse antibodies and secondary detection systems ........ 121

INTRODUCTION
17
1 INTRODUCTION
Immunologically ´privileged` sites or well-defined tissue compartments such as
present in the eye, brain, fetus, brain, and testes, have been fascinating objects of
research since the 80s and 90s. Immune privilege (IP) describes immunosuppressive
mechanisms that inhibit antigen presentation, and subsequent immune responses, in
a particular anatomical site. Immune privilege is thought to protect vulnerable tissues
with poor regenerative potential from excessive tissue damage caused by an
unrestricted immune response (MEDAWAR 1948; BILLINGHAM and SILVERS 1971;
STREILEIN 1993).
In skin appendages it has been shown previously that the proximal anagen bulb of
hair follicles (HF) (PAUS et al. 1999b, 2005; CHRISTOPH et al. 2000; ITO et al.
2004, 2007; GILHAR and KALISH 2006; GILHAR et al. 2007) and the human
proximal nail matrix (ITO et al. 2005c) are prominent sites of relative IP. In addition,
recent gene and protein expression data (MORRIS et al. 2004; TUMBAR et al. 2004;
COTSARELIS 2006b) have raised the possibility that follicular IP may not be limited
to the anagen hair bulb, but also to the epithelial stem cell region in the outer root
sheath (ORS) of HFs, termed the bulge region. So far, nothing is known about the
murine nail apparatus or vibrissal follicle.
The role of IP in the HF is currently unknown. Interestingly, collapse of IP in the
anagen hair bulb is thought to be central to the pathogenesis of the organ-specific
autoimmune condition, alopecia areata (PAUS et al. 2003). Therefore, it is possible
that IP evolved to reduce the risk of autoimmune hair-loss developing in an individual
creature, where loss of hair could threaten the ongoing survival of that individual
(PAUS et al. 2003). Since protection of bulge epithelial stem cells from immune
destruction is essential for preserving the regenerative and cycling capacity of HFs
(PAUS and COTSARELIS 1999; COTSARELIS 2006a; TIEDE et al. 2007a), it would
make sense if the bulge region also had established a relative IP. Convincing
treatment and management of alopecia areata and other hair loss disorders are still

INTRODUCTION
18
missing. Therefore, it is of paramount importance, firstly, to better characterize
immunoprivileged sites and secondly, to create substances and find mechanisms to
maintain and to restore IP.
Although limited gene expression data from isolated, human and mouse bulge-
derived cells suggested the existence of a second area of intrafollicular IP
(COTSARELIS 2006a), convincing protein evidence for this is still missing. Since IP
is a phenomenon that is based on functional protein expression patterns of entire
tissue compartments, not on gene expression patterns of individual cells in culture
(PAUS et al. 2005; NIEDERKORN 2006) we aimed in the current study to generate
protein evidence in situ [i.e. immunoreactivity (IR) evidence] that would support or
refute the hypothesis of bulge IP in human HFs. For this purpose, both routine and
increased-sensitivity immunohistochemical staining techniques were employed, and
the corresponding IR patterns were evaluated by quantitative immunohistochemistry.
This was complemented with histochemical and histomorphometric assessments. In
order to obtain functional evidence, we performed full thickness human scalp skin
organ cultures to investigate whether interferon-γ (IFN-γ), a key inducer of IP
collapse in hair bulbs, has a similar effect on the putative bulge IP.
In addition, we have evaluated a novel, synthetic α-MSH -related tripeptide [K(D)PT],
which is currently examined in different pre-clinical assay systems as a potential new
immunosuppressant with multiple clinical indication, including the possibility that it
may restore IP collapse. Therefore, the effects of K(D)PT on HF IP and biology were
examined in microdissected, organ-cultured human scalp HFs (PHILPOTT et al.
1990; BODO et al. 2007, 2008; VAN BEEK et al. 2008) under pro-inflammatory
conditions [(i.e. addition of interferon-gamma to the medium (ITO et al. 2004)].
Moreover, as an additional contribution to the charting of ´white spots` on the map of
cutaneous immunobiology, we have utilized this opportunity to clarify the IP status of
two other important skin appendages in the mammals, namely mouse sinus hair
follicles and the murine nail/claw apparatus, since these appendages still remain to

INTRODUCTION
19
be carefully characterized with respect to their IP status. The immunological
characterization of murine sinus hair follicles and nail apparatus and their IP status is
of great importance: Although vibrissal follicles differ substantially e.g. from pelage
and human terminal scalp HFs in their cycling characteristics, architecture and
innervation (DAVIDSON and HARDY 1952; MEYER 1999), they are frequently
employed in basic hair research and in drug screening assays – despite the fact that
their immunology remains largely obscure. Even less is known about the murine nail
apparatus from an immunological point of view. This may reflect the lamentable
general lack of interest in the life sciences community in something as supposedly
´profane` as nails, although many new mouse mutants display nail abnormalities and
although nail disorders in humans quite often coincide with other dermatological
disorders (CYGAN et al. 1997; AHMAD et al. 1998; GODWIN and CAPECCHI 1998;
VOLLRATH et al. 1998; KAWAKAMI et al. 2000; MECKLENBURG et al. 2004, 2005;
MOOKHERJEE et al. 2006; NAKAMURA and ISHIKAWA 2008).
In the following, after a short introduction into relevant essentials of HF, sinus hair
follicle and nail apparatus biology, the current state of research on immune privilege
in general will briefly be summarized, and key open questions will be delineated.
Subsequently, we critically discuss which relevant immune privilege markers have
been proposed in the past, with emphasis on markers that may be relevant for the
immune privilege in human HFs, and succinctly explain relevant background
information on those markers that were selected for study in the current context.
The experimental work for this thesis was performed in close collaboration with the
Department of Dermatology (Prof. Dr. R. Paus), University of Lübeck, and was
integrated into an ongoing, industry research project that exploited human HFs as an
innovative and instructive screening tool for identifying promising new drug
candidates (here: K(D)PT).

LITERATURE
20
2 LITERATURE
2.1 A short synthesis of hair follicle biology
The skin and its appendages have many different tasks to fulfill. The hair, which is
the main product of the HF, has several functions, i.e. protection against
environmental traumata, thermoregulation, social communication, mimicry and to act
as a container for sequestering and excreting unwanted compounds (PAUS and
COTSARELIS 1999; STENN and PAUS 2001). The importance of every function
depends on the mammalian species, possibly its domestication level, and the
environment. The HF is one of the most complex micro organs of the mammalian
body and the only organ, which permanently and lifelong regenerates through the so-
called HF cycle. The cycle length varies species-specific and location-specific, e.g., it
lasts years in humans and weeks in mice. But the developmental and cycle stages as
well as the basic transformations of the HF underlie the same pattern in human and
murine skin (KLIGMAN 1959) (Fig. 2.1).
Fig. 2.1 Three dimensional diagram of the mammalian skin (FUCHS 2007)

LITERATURE
21
Tab. 2.1 Basic data on human HFs Modified after PAUS et al. 2007
Total number ~ 5 000 000 (mostly vellus!) Number of scalp hair follicles
~ 100 000 Blondes: + 20% Redheads: - 20%
Average density (scalp) terminal + vellus
1135/cm2 (newborn), 615/cm2 (20-30 years), 485/cm2 (30-50 years), 435/cm2 60-80 (years) Asian: lower density; terminal only: ca 250/cm2; bald scalp (45-70 years): 330/cm2, highest density: cheek + forehead!
Hair embryology
Development progresses at fixed intervals (274-350mm) in cephalocaudal direction, first visible in eyebrow, upper lip and chin region (9th week)
Hair cycle distribution (term. scalp hair)
Anagen: 85-90%, Telogen: 10-15 %, Catagen: < 1%
Duration of hair cycle phases (terminal scalp hair)
Anagen: 2-5 years, Catagen: 2-3 weeks, Telogen: 3 months + location-specific differences (terminal moustache: 4-14 weeks, terminal arms: 6-12 weeks, terminal legs: 19-26 weeks, vellus: 6-12 weeks; premature anagen induction induced by plucking or telogen hair shafts (depilation); estrogens prolong anagen; thyroxine promotes growth, corticosteroids retard anagen onset
Number of lifetime cycles
~10-20
Physiological hair shedding rate (scalp)
~100-200/day (substantial interindividual and seasonal variations)
Hair shaft production rate (scalp)
~35mm/day, 1cm/month; hair production is not influenced by cutting/shaving; estrogens reduce hair growth rate; androgens increase hair growth rate and hair diameter in androgen-dependent sites (beard)
Hair shaft ∅ and length
Vellus: <0.03 mm; 1-2 mm Terminal: >0.06 mm; 1mm
Hair shaft structures Cuticle (outside), cortex, medulla (centre), cuticle maintains hair fibre integrity; cortex contains bulk of hair keratins and keratin-associated proteins; hair fibre strength is largely due to disulfide bonding; medulla consists of loosely connected trichocytes with large intercellular air spaces; provides insulation
Hair graying (canities)
Generally commences in the third decade of life on the temples, spreading later to crown and occiput; by the age of 50 years, 50% or the population has at least 50% grey hair
Hair patterns Pubic hair: horizontal (90% of women, 20% of men), acuminate (10% of women, 50% of men); diffuse chest hair: normally grows only in men, after puberty (until 6th decade); axillary: appears about 2 years after first pubic hairs; trichoglyphics: single, clockwise parietal present in 95% of individuals
Outer root sheath (ORS)
Outermost sheath of HF keratinocytes, merges distally into the basal layer of the epidermis and proximally into the hair bulb
Sebaceous gland (SG)
Glandular structure close to the insertion of the APM with holocrine function, lipid-filled sebocytes
Terminal hair Large, usually pigmented and medullated hair Vibrissae = Sinus hair follicle
Special sensory HFs with unique anatomy and biology, found in different regions of animal skin, but not in humans; largest and most densely innervated HFs with special sinusoid blood supply; first HFs to develop

LITERATURE
22
2.1.1 Hair follicle morphogenesis
HF morphogenesis is influenced and governed by a plethora of growth factors,
growth factor antagonists, adhesion molecules and intracellular signal transduction
components (BOTCHKAREV and PAUS 2003). At defined time points during fetal
(humans) and perinatal (rodents) skin development, HF morphogenesis begins from
small epithelial placodes (hair germs) in the epidermis above a mesenchymal
condensation (Fig. 2.2). In the following a rapid progress to the generation of
multicylindric, mature pilosebaceous units (vellus = primary HF sebaceous gland) in
the hominids, including the humans, or the HF complex (primary HF, apocrine tubular
gland, sebaceous gland) in the other mammalian groups occurs. These epidermal
keratinocytes are stimulated to commit HF specific differentiation, and the
mesenchymal cells, forming the dermal papilla (DP), send each other signals to
achieve progression to the next developmental stage. Thus, the epidermal pegs grow
downward into the dermis as a solid column of proliferating cells to enclose dermal
papillary cells and to construct the hair bulb. The hair bulb is the location, where rapid
proliferation and differentiation of the keratinocytes occurs. In the following, six
distinct cell compartments are formed: medulla, cortex and cuticle of the HS, the
cuticle and the Huxley and Henle layers of the inner root sheath (IRS). The latter
separates the HS from the ORS, which forms the external concentric layer of
epithelial cells in the HF (SENGEL 1976).

LITERATURE
23
Tab. 2.2 HF morphogenesis in mice Stage 0: Accumulation of nuclei (pre-germ), stage 1: Epidermal thickening (hair peg), stage 2: Forming of a broad column with concentrically arranged keratinocytes, stage 3: DP at the proximal end of the hair peg, stage 4: Henle’s layer of the IRS develops as a cone-shaped structure above the DP, stage 5: elongation of the IRS halfway up (hair cone), stage 6: HF reached the deep hypodermis (subcutis) and hair canal is visible, stage 7: the tip of the HS leaves the IRS and enters the hair canal, stage 8: HF acquires its maximal length and reaches the hypodermal muscle layer (panniculus carnosus) (PAUS et al. 1999b)
Eight defined gradual steps of HF morphogenesis can be distinguished, which
underlie different mechanisms and factors. In stage 0, epidermal nuclei accumulate
and form the so-called pregerm (PINKUS 1958), which develops in stage 1 into a
circumscribed epidermal thickening of enlarged keratinocytes in the basal layer of the
epidermis, termed the hair peg (DRY 1926). Forming of a broad column with
concentrically arranged epithelial keratinocytes around the follicular axis ends stage
2. At the proximal end of the column fibroblasts condensate and form the DP (stage
3). The hair peg elongates and the IRS starts to develop as a cone-shaped structure
(stage 4, 5), which in pigmented skin is visible by its melanin formation (PAUS et al.
1999b). In stage 6 the HF reaches the deep hypodermis and a hair canal is formed.
This is followed by the entering of the HS into the hair canal (stage 7). By reaching

LITERATURE
24
the hypodermal muscle layer (panniculus carnosus), if present (stage 6) the HF has
its maximal length and its prominent HS emerges through the epidermis (stage 7).
This event determines the end of morphogenesis and the onset of the first hair cycle.
First recognizable cyclic changes of HF activity start when the HF enters a stage of
physiological apoptosis-driven involution (catagen) (STRAILE et al. 1961;
DEPLEWSKI and ROSENFIELD 2000; STENN and PAUS 2001).
This course of morphogenesis occurs in humans but also in sparsely and densely
haired mammals, like pigs or mice (Fig. 2.2); the latter species are often used as a
model for the human skin (MEYER 1986, 2009; MEYER and GOERGEN 1986;
PAUS et al. 1999b)
2.1.2 Functional anatomy of the hair follicle
The human skin contains about 5 million HFs, of which mostly are vellus HFs.
Thereof are 100.000 HFs prominently displayed on the scalp (plus those of
eyelashes and eyebrows) (DAWBER 1997; PAUS and PEKER 2003; PAUS and
FOITZIK 2004). HFs can be divided into three different types: lanugo, vellus and
terminal (primary) HFs. Although the different types of HFs follow the same
construction principles of functional bioarchitecture, they display some structural and
pigmentary differences (DAWBER 1997; PAUS and COTSARELIS 1999). Most of
the HFs in the skin are of the vellus type. In contrary, the human scalp skin is
basically covered with terminal HFs only.
Every mature anagen scalp HF displays the shape of an inverted wine glass into
whose calyx an onion-like structure, the follicular dermal papilla is located. The
architecture of the HF is constructed on the need or key function as a fibre production
facility, whose outwards-growing hair shaft has to be carefully protected on its way up
to the skin surface. Interaction with the surrounding dermis would provoke infection
and therefore has to be prevented. The directional growth is based on guiding
structures and slippage planes: Terminally differentiated keratinocytes form a
LITERATURE
25
hardened inner cylinder (i.e. IRS) and guide the central hair shaft. In addition, the
companion layer of the ORS functions as a slippage plane, and facilitates the
outgrowing of the hair shaft together with the IRS (STENN and PAUS 2001;
LANGBEIN et al. 2002; PAUS and PEKER 2003).
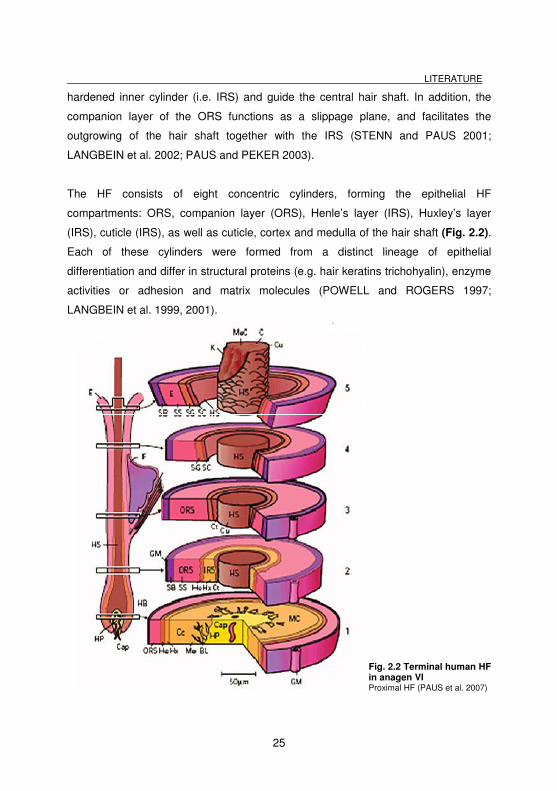
The HF consists of eight concentric cylinders, forming the epithelial HF
compartments: ORS, companion layer (ORS), Henle’s layer (IRS), Huxley’s layer
(IRS), cuticle (IRS), as well as cuticle, cortex and medulla of the hair shaft (Fig. 2.2).
Each of these cylinders were formed from a distinct lineage of epithelial
differentiation and differ in structural proteins (e.g. hair keratins trichohyalin), enzyme
activities or adhesion and matrix molecules (POWELL and ROGERS 1997;
LANGBEIN et al. 1999, 2001).
Fig. 2.2 Terminal human HF in anagen VI Proximal HF (PAUS et al. 2007)
LITERATURE
26
Progeny of eSCs generate the compartments of ORS, IRS, hair matrix, and HS.
These slow-cycling, ´label retaining` eHFSC are present throughout the entire lifetime
of the HF, and are vital as a major site of eHFSCs to the regeneration and cycling
capacity of the HF (COTSARELIS et al. 1990, 1999; LYLE et al. 1999; TUMBAR et
al. 2004; OHYAMA et al. 2006; TIEDE et al. 2007a, 2007b). These eSCs reside in an
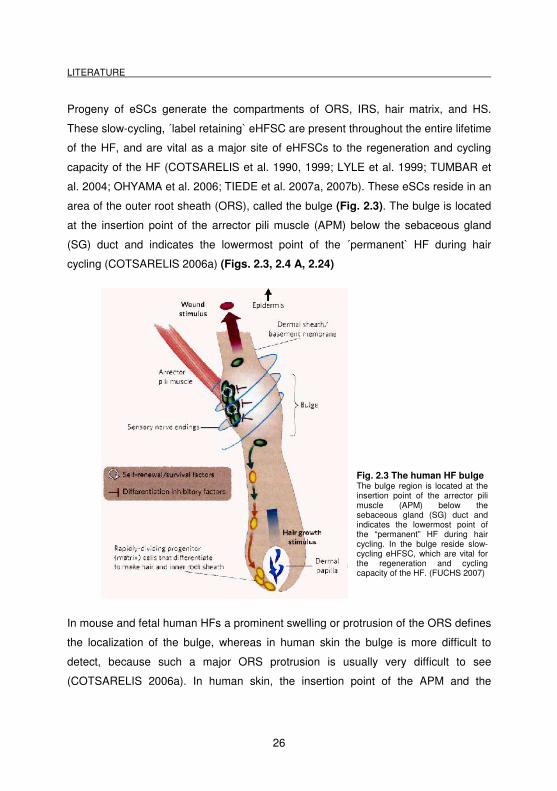
area of the outer root sheath (ORS), called the bulge (Fig. 2.3). The bulge is located
at the insertion point of the arrector pili muscle (APM) below the sebaceous gland
(SG) duct and indicates the lowermost point of the ´permanent` HF during hair
cycling (COTSARELIS 2006a) (Figs. 2.3, 2.4 A, 2.24)
Fig. 2.3 The human HF bulge The bulge region is located at the insertion point of the arrector pili muscle (APM) below the sebaceous gland (SG) duct and indicates the lowermost point of the “permanent” HF during hair cycling. In the bulge reside slow-cycling eHFSC, which are vital for the regeneration and cycling capacity of the HF. (FUCHS 2007)
In mouse and fetal human HFs a prominent swelling or protrusion of the ORS defines
the localization of the bulge, whereas in human skin the bulge is more difficult to
detect, because such a major ORS protrusion is usually very difficult to see
(COTSARELIS 2006a). In human skin, the insertion point of the APM and the

LITERATURE
27
recently found characteristic structure, the so-called ´follicular trochanter` can provide
a useful histological demarcation of the human bulge (TIEDE et al. 2007a).
These eSCs, from which the ORS is generated by transient amplifying cells
(COTSARELIS et al. 1990), reside permanently in the bulge region, while those
transient amplifying cells have been postulated to arise from a second population of
stem cells that have become deposited in the secondary hair germ (PANTELEYEV et
al. 2001; BLANPAIN et al. 2004; CHRISTIANO 2004) (Fig. 2.4 A) These cells
construct IRS, hair matrix and HS. However, until now convincing proof for the latter
concept is missing. In genetically engineered mice, bulge stem cells have been
demonstrated to generate all epithelial cells lineages, including IRS, hair matrix, and
HS (TAYLOR et al. 2000; OSHIMA et al. 2001; BLANPAIN et al. 2004).
Fig. 2.4 HF: Keratinocyte lineages and structure Keratinocyte lineages in the HF (A) (PAUS et al. 2007), structure of the human HF (B) (WHITING 2004)

LITERATURE
28
The anagen hair bulb (Fig. 2.5) is one of the key compartments of the HF and
constitutes the actual HS factory, in which one of the most rapidly proliferating cell
populations are found in any mammalian tissue: the keratinocytes of the one-layered
hair matrix. In the precortical hair matrix, which is situated above the DP, these cells
initiate their terminal differentiation to trichocytes. Later on, they receive
melanosomes from the melanocytes of the HF pigmentary unit for HS pigmentation.
During further process of differentiation into HS cuticle, cortex and medulla, the
keratinocytes express a defined set of keratins (SLOMINSKI and PAUS 1993a;
TOBIN and PAUS 2001). Matrix cells, medulla, cortex, IRS and ORS represent
ectodermal derivatives, whereas the DP, the CTS and the hyaline membrane,
separating the CTS from the ORS, are derivatives of the neural crest.
Fig. 2.5 Schematic drawing of a hair bulb DP=dermal papilla, CTS, connenctive tissue sheath, bORS=basal layer of the ORS, cl=companion layer, He=Henle layer, Hu=Huxley layer, icu=cuticle of the IRS, Cu=hair shaft cuticle, ma/co=hair shaft medulla and cortex, gp=germinative pool

LITERATURE
29
The diameter and volume of the DP determines the number of specialized fibroblasts
and is an indicator for its secretory power for the release of ´papilla morphogens`
(JAHODA and REYNOLDS 1996; PAUS et al. 1999b). Thus the larger the DP is the
bigger the HF and the diameter of the hair shaft. If a DP is destroyed, it can be fully
reconstituted from the proximal connective tissue sheath (CTS) of the HF (JAHODA
1992; JAHODA and REYNOLDS 1996; REYNOLDS et al. 1999), which harbors
mSCs (LAKO et al. 2002; JAHODA 2003). The exchange (so-called trafficking) of
fibroblasts between DP and the proximal CTS occurs during each telogen-anagen-
catagen transformation and results in substantial changes in DP volume and cell
content (TOBIN et al. 2003).
The angle of the hair shaft is dependent on the action of the arrector pili muscle. The
muscle is under adrenergic control, and thus involuntarily contracts in situations of
sudden stress, anxiety or anger, ´making one’s hair stand-up` (PAUS and PEKER
2003). However, in humans this capacity became less important compared to
animals. In human scalp skin, a single APM structure is shared by all the follicles
within the so-called follicular unit, a defined group of 2–4 terminal and 1–2 vellus
(HEADINGTON 1984), joining bulky cords of muscle fibres at one pole of the
follicular unit at the upper isthmus level (POBLET et al. 2002).
The follicular innervation system is responsible for the recognition and signaling of
sensitive tactile stimuli (e.g. hair shaft movements caused by wind, insects, stroking).
In addition, the follicular neural plexus may also have important trophic and
regulatory functions by the release of neurotransmitters, neuropeptides and
neurotrophins (BOTCHKAREV et al. 1997, 1998a, 1998b, 1999; PAUS et al. 1997;
PETERS et al. 1999, 2002a). The bulge and isthmus region of human HFs contain a
particularly dense network of sensory and autonomic nerves, as well as numerous
Merkel cell complexes in human HFs (but not in murine pelage HFs) (UHR 1984;
(BOTCHKAREV et al. 1997a; PAUS et al. 1997; PETERS et al. 2002).
The vasculature is similar to the innervation very densely and basket-like located
around the HF. It arises from the dermal and hypodermal vascular plexus and is

LITERATURE
30
formed by arterioles, capillaries and venules with numerous shunts. This perfusion
system sheathes the entire follicle, weaving through its CTS, and even inserts into
the DP of terminal HFs, in humans but also in all other mammals (MECKLENBURG
et al. 2000; YANO et al. 2001). That ensures, that all key regions of the HF have
abundant access to all essential factors and that metabolic products can be removed.
Tab. 2.3 Glossary of anatomical and trichology terms (modified after PAUS and PEKER 2003; MUELLER-ROEVER et al. 2001)
Bulb Prominent onion-shaped thickening on the proximal end of the HF, relatively undifferentiated matrix cells, melanocytes and cells from the proximal ORS
Bulge Convex extension of the distal part of the ORS, near the epidermis, location of epithelial HF stem cells and insertion of the APM
Club hair Resting HS with a hollow brush of keratinized keratinocytes on the proximal end, tightly attached to the cortical cells of the hair cortex
Connective tissue sheath (CTS)
Part of the dermal connective tissue, tightly attached to the outer side of HF, composed of fibroblasts, macrophages and connective tissue
Dermal papilla (DP) = Follicular papilla
Mesodermal part of the HF, closely packed mesenchymal cells, framed by the bulb matrix during anagen
Epithelial strand Column of epithelial cells between the germ capsule and the compact DP, laterally demarcated by the thickened glassy membrane
Follicular pigmentary unit
Melanin-producing HF melanocytes located above and around the upper on-third of the DP, transfer of eu-or pheomelanosomes to differentiating HF keratinocytes in the precortical matrix; goes largely into apoptosis during each catagen phase, regenerated from melanocyte stem cells
Secondary germ capsule = Secondary hair germ
Bag-like structure of glycogen-free cells of distal ORS, surrounding the club hair
Hair shaft (HS) Terminally differentiated HF keratinocytes (trichocytes), the HS is divided into cuticle, cortex and medulla
Hyaline membrane = Vitreous membrane = Glassy membrane
Outermost noncellular part of the HF, basal lamina and two layers of orthogonally arranged collagen fibres, separates ORS from CTS
Isthmus Middle portion of the HF extending from the sebaceous duct to the insertion of APM (bulge region)
Inner root sheath (IRS) Multilayered structure composed of terminally differentiated HF keratinocytes surrounded by the ORS, surrounds the hair up to the hair canal
Lanugo hair Fine hair on the fetal body, shed in utero or during the first weeks of life Outer root sheath (ORS)
Outermost sheath of HF keratinocytes, merges distally into the basal layer of the epidermis and proximally into the hair bulb
Sebaceous gland (SG) Glandular structure close to the insertion of the APM with holocrine function, lipid-filled sebocytes
Terminal hair Large, usually pigmented and medullated hair Vibrissae = Sinus hair follicle
Special sensory HFs with unique anatomy and biology, found in different regions of animal skin, but not in humans; largest and most densely innervated HFs with special sinusoid blood supply; first HFs to develop

LITERATURE
31
2.1.3 Hair follicle cycle
The HF is one of the few micro organs of the body that undergoes lifelong cycling.
HF cycling describes the morphological evidence of rhythmically re-occurring growth,
regression and tissue re-modeling events in this complex neuroectodermal-
mesodermal interaction system (PAUS and FOITZIK 2004). Originally, HF cycling is
synchronized in mammals in accordance to seasonal changes in habitant or
procreational activities (STENN and PAUS 2001). In mice, pelage HF cycling occurs
in a wave-like synchronous pattern starting from neck to tail (MUELLER-ROEVER et
al. 2001). In humans, the synchronized follicular cycling is lost after one year of life
and is replaced by a random or mosaic pattern of asynchonized hair cycling
(WHITING 2004). Such pattern type is also observed in domesticated mammals kept
under indoor conditions (MEYER et al. 1980, 2009a). The purpose for asynchronous
HF cycling in humans is not fully investigated but may include cleaning of the skin
surface of debris and parasites and excretion of chemicals by encapsulation within
trichocytes (STENN and PAUS 2001). In addition, HF cycling might serve as
regulator of paracrine or even endocrine secretion of hormones and growth
modulators produced within the follicle and secreted into the skin and / or circulation
(PAUS and COTSARELIS 1999).
The cyclic transformations from phases of rapid growth (anagen), via apoptosis-
driven regression (catagen) to relative quiescence (telogen) (DRY 1926), are
characterized by regression and proliferation activity and are influenced by numerous
of factors (e.g. growth factors, cytokines, hormones, neuropeptides) (for review
(STENN et al. 1996; PAUS and COTSARELIS 1999). 85 to 90% of all scalp HFs are
within anagen stage, which lasts for 2-6 years. The duration of hair growth
determines the length of the HF. Catagen lasts for a few weeks and is replaced by 2
to 4 months of telogen phase. Scalp HFs grow approximately between 0.3 and 0.5
mm per day, which is determined by the proliferation and differentiation of the matrix
keratinocytes (MKs) (DAWBER 1997; STENN and PAUS 2001; PAUS et al. 2007).
LITERATURE
32
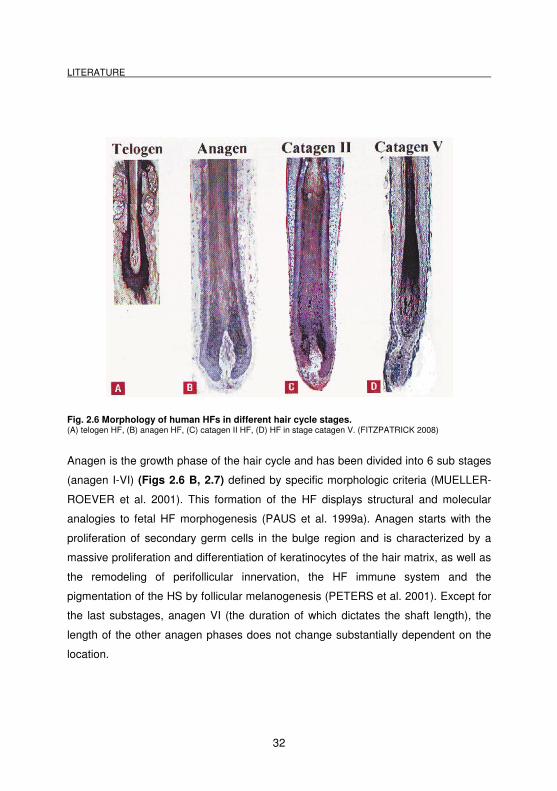
Fig. 2.6 Morphology of human HFs in different hair cycle stages. (A) telogen HF, (B) anagen HF, (C) catagen II HF, (D) HF in stage catagen V. (FITZPATRICK 2008)
Anagen is the growth phase of the hair cycle and has been divided into 6 sub stages
(anagen I-VI) (Figs 2.6 B, 2.7) defined by specific morphologic criteria (MUELLER-
ROEVER et al. 2001). This formation of the HF displays structural and molecular
analogies to fetal HF morphogenesis (PAUS et al. 1999a). Anagen starts with the
proliferation of secondary germ cells in the bulge region and is characterized by a
massive proliferation and differentiation of keratinocytes of the hair matrix, as well as
the remodeling of perifollicular innervation, the HF immune system and the
pigmentation of the HS by follicular melanogenesis (PETERS et al. 2001). Except for
the last substages, anagen VI (the duration of which dictates the shaft length), the
length of the other anagen phases does not change substantially dependent on the
location.
LITERATURE
33
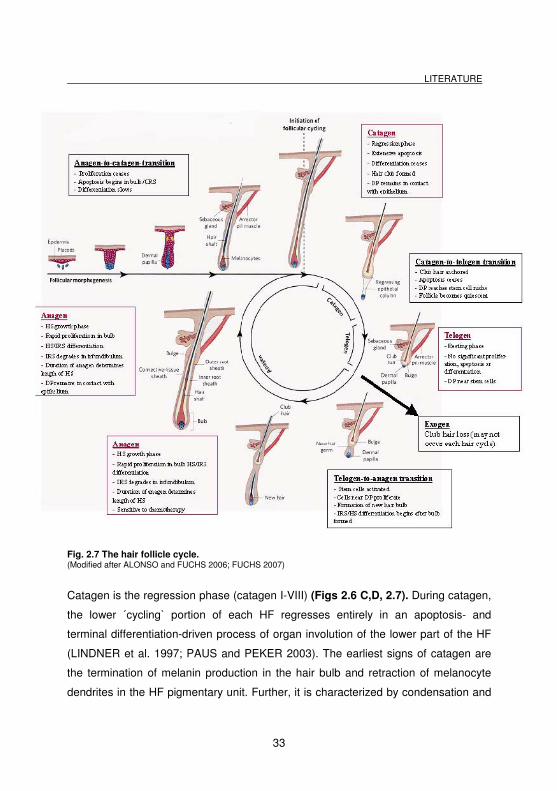
Fig. 2.7 The hair follicle cycle. (Modified after ALONSO and FUCHS 2006; FUCHS 2007)
Catagen is the regression phase (catagen I-VIII) (Figs 2.6 C,D, 2.7). During catagen,
the lower ´cycling` portion of each HF regresses entirely in an apoptosis- and
terminal differentiation-driven process of organ involution of the lower part of the HF
(LINDNER et al. 1997; PAUS and PEKER 2003). The earliest signs of catagen are
the termination of melanin production in the hair bulb and retraction of melanocyte
dendrites in the HF pigmentary unit. Further, it is characterized by condensation and

LITERATURE
34
upward movement of the DP (TOBIN et al. 1998), leaving an angiofibrotic strand or
stela indicating the former position of the anagen root (WHITING 2004).
At the end of this involution process, the HF enters into the resting phase, the so-
called telogen (Figs 2.6 A, 2.7). The resting club hair is situated at the bulge level
where the APM inserts into the HF. The telogen bulb is non-pigmented and has no
IRS. In telogen, the HF is characterized by relative quiescence. However, telogen is
considered to be much more important than the term ´resting` implies, since the
epithelial remnants of the telogen HF (distal ORS, secondary hair germ, bulge) are
engaged, e.g. in substantial biochemical activity and some degree of proliferation
(PAUS and COTSARELIS 1999; MUELLER-ROEVER et al. 2001).
In addition, during the course of cycling, substantially remodeling of both the HFs
innervation and vasculature occurs (BOTCHKAREV et al. 1997a; MECKLENBURG
et al. 2000; YANO et al. 2001). The HF transition between distinct stages of
development and postnatal cyclic regeneration is governed by a bidirectional signal
exchange between follicular keratinocytes and fibroblasts of the follicular DP, which
is supposed to be the control centre of follicle growth, initiating and terminating
anagen. Many molecular key regulators that had been involved in the regulation of
HF development are also recruited for the control of cycling. Just to point out one is
TGF-β, which induces catagen development (LITTLE et al. 1994). The HF
development is due to DP fibroblasts and its contact to hair MKs (JAHODA and
REYNOLDS 1996), which signals act on the eSCs of the follicle to initiate anagen
(bulge activation hypothesis). The stem cells are supposed to generate rapidly
dividing transient amplifying cells that migrate towards the DP for constructing a new
hair bulb (LAVKER et al. 1993).

LITERATURE
35
2.2 Sinus hair follicle biology: overview
Sinus hair follicles belong to the evolutionary oldest tactile sense organs of the
mammalian skin that have such a central role that they are the first developing hair
type in embryonal stage and even do exist in congenital hairlessness in mice
(HALATA 1993; MEYER 1999; MEYER and ROEHRS 1986). Sinus hair follicles are
also known as whiskers, vibrissae, vibrissal follicles, feelers or tactile, sensory or
sinus hairs and were firstly described as ´large stiff hairs (BLAND-SUTTON 1887;
BEDDARD 1902) that are pre-eminently sensory` and which differ from all other
types of hair through the presence of erectile tissue in their follicles (BOTEZAT 1897;
DANFORTH 1925b) (Fig. 2.8). Vibrissal follicles are highly sensitive
mechanoreceptive complexes that receive pressure and contact stimuli and initiate
behavior-relevant reactions through central nervous regulation. Vibrissal follicles are
important for the cognition of the environment, for social contacts between animals,
food intake. Loss of vibrissal follicles can immediately to loss of orientation and
subsequent erratic behavior (MEYER 1999).

LITERATURE
36
Fig. 2.8 Diagram of active vibrissal follicle in adult mouse. (DAVIDSON and HARDY 1952)
2.2.1 Sinus hair follicle morphogenesis
As in the human HF, morphogenesis of vibrissal HFs is governed by a series of
different events and is marked by a high degree of order and pattern in time and
space (DANFORTH 1925a; GRUNEBERG 1943; YAMAKADO and YOHRO 1979;
VAN EXAN and HARDY 1980): The development is more rapid than in pelage
follicles. In the 12-day embryo, the epidermal plugs of the first vibrissal follicles
appear and hairs emerge 5-6 days later. In contrast, pelage HFs start develop in the
14 day embryo and require longer to emerge (DAVIDSON and HARDY 1952).
In general, the differentiation of vibrissal follicles is almost the same as that of pelage
HFs (HARDY 1949, 1951) (Fig. 2.9). It starts with an epidermal downgrowth (stage

LITERATURE
37
1), formation of a pre-papillae (stage 2) and DP (stage 3) (ZIETZSCHMANN 1920)
are formed. In stage 4 a hollow cone develops by the hardening of certain cells from
the hair matrix, which then gives rise to the IRS. In stage 5 a hair canal is formed with
the condensation of cells, with strongly basophilic nuclei, on an elevation or cap of
epidermis above the follicle, followed by the occurrence of a keratinized hair shaft in
inside the cone (stage 6). This formation of a canal by keratinization from the
epidermal surface downwards distinguishes the vibrissal follicles from the pelage
HFs, in which keratinization begins as a blind cavity within the deeper layers of the
epidermis at the level of the stratum spinosum (HARDY 1949), and in which no cap
of epidermal cells and no condensation of basophilic nuclei occurs. At stage 6a this
condensation reaches the tip of the hair cone, whereas at stage 6b a canal is formed
within this area by the keratinization of cells in the centre. The hair shaft has pierced
the IRS at stage 7. Morphogenesis ends with the emerging of the hair shaft from the
skin (stage 8) and the forming of blood capillaries of the DP (DAVIDSON and
HARDY 1952).
Fig. 2.9 Diagram of stages 1-8 in development of vibrissal follicles in mouse. Stage 0: no follicles, stage 1: epidermal downgrowth and follicle plugs, stage 2: pre-papillae, stage 3: papillae, stage 4: hair cones develops by the hardening of certain hair matrix cells, stage 5: hair canal is formed, stage 6a: hair formation, stage 6b: opening of hair canals, stage 7: hair shafts in hair canals, stage 8: hairs emerged (DAVIDSON and HARDY 1952)

LITERATURE
38
Vibrissal follicles are much larger and stouter than those of pelage. The epidermal
plug is from the beginning on surrounded by a dermal sheath, in which few isolated
blood cells are visible at stage 8. While the follicle is growing and the DP is formed,
the follicle gets the characteristic hourglass shaped (Fig. 2.9). At stage 3 the
characteristic thickening of the ORS becomes evident, and by stage 6 the superior
and inferior swellings can be distinguished. In the newborn mouse the lower blood
sinus is differentiated with a well-developed fibrous wall and connective tissue
trabeculae, filled with blood cells. Three days later the upper sinus and ringwulst are
completely differentiated. After birth no new vibrissal follicle is added to those already
regularly arranged in rows, whereas pelage HFs continue to appear until day 5 till 8
after birth (DAVIDSON and HARDY 1952).
Later on, new four substages of vibrissa follicle development which occurred prior to
stage 1 of DAVIDSON and HARDY (1952) were described (VAN EXAN and HARDY
1980). In addition, it was found that vibrissal pattern formation is likely to be a
complex process relying on the interaction of cells and tissues (comparable to normal
HFs), rather than on unidirectional instructions from neurons to other cell types
(WRENN and WESSELLS 1984).
2.2.2 Functional anatomy of the sinus hair follicle
The major mystacial vibrissae of the mouse and rat are arranged on the snout (from
the nose to the cheek) in five ´horizontal` (rostrocaudal) rows and one ´vertical`
(dorsoventral) row which lies just caudal to the horizontal rows. Within each row a
characteristic anterior-posterior size gradient is observed: The largest vibrissae are
being located near the cheek and the smallest near the nose (OLIVER 1966b). The
location of the vibrissae follicles and the numbers in the major groups are
predetermined and constant (DANFORTH 1925a; GRUNEBERG 1943; DUN and
FRASER 1958; YAMAKADO and YOHRO 1979), except in a few mutants
(YAMAKADO and YOHRO 1979). It was also found that vibrissal follicles grew
synchronously within the same margin (IBRAHIM and WRIGHT 1975). Within this

LITERATURE
39
group of vibrissal follicles it is considered that the bigger follicles (so-called
´macrovibrissae`) have especially the function as a ´distance decoder`, whereas
smaller ´microvibrissae` are critically involved in object recognition (BRECHT et al.
1997).
Vibrissal follicles are similar in their histological structure compared to pelage HFs:
They consist of the same concentric cylinders, forming the epithelial HF
compartments: ORS, companion layer (CL), Henle’s layer (IRS), Huxley’s layer (IRS),
cuticle (IRS), as well as cuticle, cortex and medulla of the hair shaft (see Tab. 2.1).
On the other hand, there are many structural features which distinguish vibrissal
follicles from pelage HFs (DAVIDSON and HARDY 1952) (Fig. 2.10):
Sinus hair follicles are highly innervated sensory organs (VINCENT 1913;
WINKELMANN 1959), which are represented in the somatosensory cortex of the
brain by their cortical ´barrels` (WOOLSEY and VAN DER LOOS 1970). Each
vibrissal follicle is supplied by a large deep vibrissal nerve arising from the infraorbital
nerve (DORFL 1982), which enters the capsule in the lower third of the follicle
(MAROTTE et al. 1992). In addition, the vibrissal follicle is innervated by several
smaller superficial nerves (DAVIDSON and HARDY 1952) which arise from the
dermal plexus. The innervation is particularly dense in the larger caudal vibrissal with
about 200 myelinated and 100 unmyelinated fibres in each deep nerve (WAITE and
CRAGG 1982; WELKER and VAN DER LOOS 1986; CRISSMAN et al. 1991). These
afferents supply a variety of sensory receptors: Merkel cells in the epidermal rete
ridge collar at the mouth of the follicle and in the external root sheath at the level of
the ring sinus; longitudinal lanceolate endings in the mesenchymal sheath at the level
of the ring sinus; Ruffini and terminals and free nerve endings in the inner conical
body and in the cavernous sinus; and a few free nerve endings in the DP (ANDRES
1966; RENEHAN and MUNGER 1986; RICE et al. 1986; MUNGER 1991). Further it
was found, that for example Merkel cells contain receptors and transporter for
serotonin, substance P, calcitonin gene relating protein, vasointestinal peptide and
met-encephalin in rats (TACHIBANA et al. 2005; TACHIBANA and NAWA 2005).
LITERATURE
40
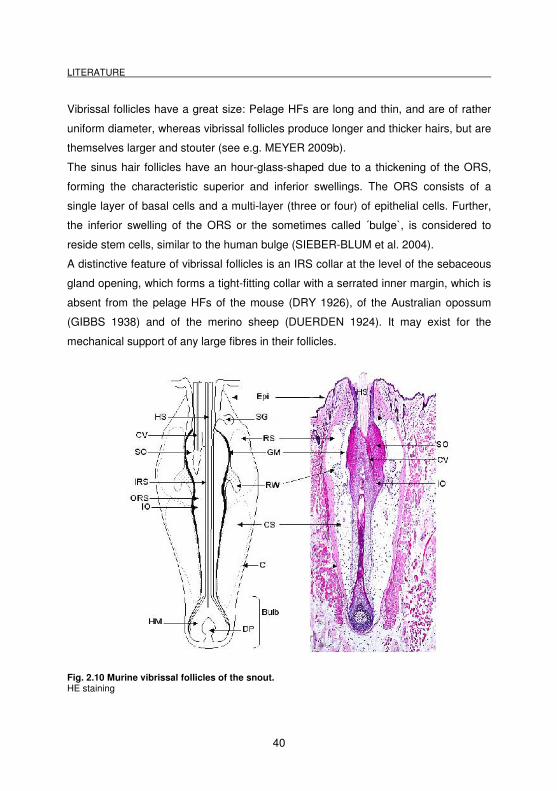
Vibrissal follicles have a great size: Pelage HFs are long and thin, and are of rather
uniform diameter, whereas vibrissal follicles produce longer and thicker hairs, but are
themselves larger and stouter (see e.g. MEYER 2009b).
The sinus hair follicles have an hour-glass-shaped due to a thickening of the ORS,
forming the characteristic superior and inferior swellings. The ORS consists of a
single layer of basal cells and a multi-layer (three or four) of epithelial cells. Further,
the inferior swelling of the ORS or the sometimes called ´bulge`, is considered to
reside stem cells, similar to the human bulge (SIEBER-BLUM et al. 2004).
A distinctive feature of vibrissal follicles is an IRS collar at the level of the sebaceous
gland opening, which forms a tight-fitting collar with a serrated inner margin, which is
absent from the pelage HFs of the mouse (DRY 1926), of the Australian opossum
(GIBBS 1938) and of the merino sheep (DUERDEN 1924). It may exist for the
mechanical support of any large fibres in their follicles.
Fig. 2.10 Murine vibrissal follicles of the snout. HE staining

LITERATURE
41
The medulla of vibrissal follicles has cells and air spaces resembling a honeycomb,
and is probably of the intermediate type (HAUSMANN 1924), whereas in pelage the
medulla cells are arranged in regular rows. The glassy membrane (basal membrane)
separates the epithelial ORS of the follicle from the mesenchymal sheath, and is also
passed by nerves and contains Merkel cells.
Vibrissal follicles are surrounded by a thick dermal sheath from the very early
beginning of morphogenesis, within the blood sinuses lie. All vibrissal follicles
possess blood sinuses and abundant nerve endings lying within the dermal sheath,
which are characteristic of all tactile HFs, but absent in pelage hair follicles. The
lower or venous blood sinus surrounds the lower portion of the follicle with connective
tissue bands within the cavity. The more superficial upper or Ring sinus has no
trabeculae, but contains the so-called ´ringwulst`. This ringwulst is attached to the
dermal sheath adjacent to the follicle wall, forming a collar surrounding the follicle. It
consists of connective tissue and is penetrated by nerve branches and blood
capillaries. Vibrissal DPs contain a blood supply in contrast to murine pelage DP and
are more richly innervated than those of the pelage.
In mice, each vibrissal and pelage HF consists of only one small sebaceous gland,
and lacks sweat glands. Vibrissal follicles have no smooth muscle corresponding to
the arrector pili muscle. Instead, the vibrissal HF has a group of larger striated
muscle fibres (so-called intrinsic muscles), which are solely attached to the dermal
sheath without any bony attachment (DOERFL 1982). Each of these follicular
muscles connects two adjacent follicles of the same row (Fig. 2.11).

LITERATURE
42
Fig. 2.11 Musculature of vibrissal follicles (A) Schematic drawing of two neighbouring follicles in the same row. R: rostral, C: caudal, B: fibrous band connecting the lower parts of the follicles, P: plate, the band between the rostral and caudal faces of two adjacent follicles, L: longitudinal muscular band formed by fibres of m. levator labii sup. and m. maxillolabialis, N: follicular nerve accompanied by an artery. (B) vertical section of two neighbouring vibrissal follicles. Masson-Goldner staining. Bar (B): 100µm.
1.1.1 Sinus hair follicle cycle
Similar to other human HFs and murine pelage HFs, vibrissal follicles show a
rhythmical and characteristic cycling. However, important differences exist, since
they do not dramatically undergo the extensive shortening seen during catagen in
pelage HFs and more importantly, their catagen and particularly telogen phases are
abbreviated (YOUNG and OLIVER 1976). For that reason, a different terminology
was outlined by YOUNG and OLIVER (1976), compared to the more detailed
classification used for pelage cycle (CHASE 1954).
Thus, hair cycle stages of the vibrissal follicles are divided into two distinct phases
(growth, regression), classified further into eight categories: pro-anagen (PA), very
early anagen (VEA), early anagen (EA), mid anagen (MA), late anagen (LA), early
catagen (EC), mid catagen (MC) and late catagen (LC) by their morphology of hair
bulbs and relative length of hair shafts (YOUNG and OLIVER 1976; ROBINSON et
al. 1997) (Fig. 2.12). Important to note is, that in contrast to pelage or human HFs,
termination of hair growth in the previous hair cycle and initiation of hair regeneration
in the following cycle partially overlaps. Thus, telogen is not obvious; otherwise the
sensory function of the tactile organ would be interrupted.

LITERATURE
43
Anagen follicles are categorized by the ratio of hair length of the growing hair shaft to
the fully grown club hair formed in the previous hair cycle; for early anagen the ratio
is less than one-quarter, for mid anagen between one-quarter and two-thirds, and for
late anagen over two-thirds.
Fig. 2.12 Mouse vibrissal follicle cycle (A) Diagram of vibrissal follicle growth cycle. (modified after (YOUNG and OLIVER 1976). (B)The bluish colour detects alkaline phosphatase activity during the hair cycle in the dermal papilla. PA: pro-anagen, VEA: very early anagen, EA: early anagen, MA: mid anagen, LA: late anagen, EC: early catagen, MC: mid catagen, LC: late catagen; Alcalic Phosphatase IR is labeled in blue (IIDA et al. 2007b)
In catagen, follicles have only one hair shaft (Fig. 2.12 A), because the old one
already sheds off from the follicle. The bulbs start to decrease their volume due to the
ceasing of proliferation of hair matrix cells and the upward movement of the hair shaft
base (YOUNG and OLIVER 1976) and become more longitudinally stretched and
thin in accordance with the upward movement of the club hair (Fig. 2.12 B).
Similar to human HFs, the underlying mechanisms guiding the vibrissal follicle
through the cycle have not yet been fully characterized. However, epithelial-
mesenchymal interactions are also considered to be important in hair growth
regulation (COHEN 1965; KOLLAR 1970; LINK et al. 1990; IIDA et al. 2007a). The

LITERATURE
44
cycle time was shown to be dependent on the size of the follicle and dermal papilla:
smaller vibrissal follicles have a shorter cycle time than that of the larger vibrissal
follicles. Relevant age and sex-dependent differences have not been demonstrated
(IBRAHIM and WRIGHT 1975), which confirmed the generally agreed hypothesis,
that vibrissal follicles are outside hormonal control in contrast to other HFs. It had
been demonstrated, that the complete removal of the DP ceases follicle growth
(OLIVER 1966b), and resumes after implantation of cultured dermal papilla cells
(OLIVER 1966a; JAHODA et al. 1984; HORNE et al. 1986; PISANSARAKIT and
MOORE 1986; JAHODA 1992; MCELWEE et al. 2003). Other studies also
suggested that the DP activity contributes to hair cycle changes, e.g. through
expressing different levels of alkaline phosphatases (IIDA et al. 2007b), fibronectin,
laminin or type IV collagen (JAHODA et al. 1992). Independently from normal cycling,
plucking of vibrissal follicles at any time during the cycle resulted in the induction of a
new hair (JOHNSON and EBLING 1964; IBRAHIM and WRIGHT 1978b, 1978a).
2.3 Murine nail apparatus: overview
The nail or claw unit comprises the distal-most, dorsal structure of vertebrate limbs.
The main purpose of the nail apparatus is to provide a protective covering, known as
the nail plate, over the dorsal aspect of each distal digit of the hands and feet
(ACHTEN 1968; RUNNE and ORFANOS 1981; JIARAVUTHISAN et al. 2007). In
addition, to protect the fingertips from traumatic injury, the nail plate also applies a
pressure that opposes the volar side of the terminal phalanx, which contributes to the
enhances sensory discriminatory ability of the fingertips (DE BERKER et al. 2001).
Besides this, claws or nails have applications like scratching and grooming and can
be utilized as a means of defense or attack. In higher primates including humans,
nails have developed in conjunction with the acquisition of manual dexterity
(DAWBER 1980), and are modified to become a cosmetic accessory and
occasionally are capable of conveying information about social standing of an
individual (MURDAN 2002); other mammals do not possess such flattened claws
(DAWBER 1980). Nails are considered to have developed from claws which are

LITERATURE
45
similar keratinized structures at the ends of the distal phalanx of many animals
(CLARK 1936), although important ultrastructural and biochemical differences are
present (THORNDYKE 1966).
2.3.1 Nail morphogenesis
Nail morphogenesis is governed by multiple different signals and develops from an
ingrowth of the epidermis into the dermis (PAUS and PEKER 2003) which gives rise
to a plate formed by fully keratinized (onychocytes), the nail plate. This visible part of
the nail apparatus functions as a protection of the tips of digital phalanges. The nail
apparatus is formed from an invagination of the primitive epidermis on the dorsum of
the terminal phalanges (DAWBER 1980), namely, the same primitive epidermis that
gives rise to hair, sweat glands and the stratum corneum (PAUS and PEKER 2003).
Its developmental time course is very constant and predictable so that fingernails in
the neonate have been exploited as useful parameters for estimating the gestational
age at term in human (BALAKRISHNAN and PURI 1973; PARKIN et al. 1976) and
farm animals. This first epidermal thickening appears during the ninth week of
gestation in human and by the 13th week the nail bed and nail fold possess a
granular layer with keratohyalin granules which disappear when the hard nail plate is
formed (LEWIS 1954). In mice, the nail primordium is first seen at embryonic day 15
in all but the anterior-most digit (thumb) of the mouse forelimb (KAUFMAN 1992).
The nail primordium of this vestigial digit is formed a few days later. At the level of its
proximal transverse groove, an epidermal duplication grows and forms the primordial
matrix (RUNNE and ORFANOS 1981). Before this matrix begins definitive nail
production, a thin rootless fore nail arises from the subsequent ´sole horn` area.
While this fore nail covers the nail bed, the primordial matrix differentiates and
produces the nail plate (ZAIAS 1963). The protruding nail finally pushes through the
eponychium and covers the entire nail bed. Initially, matrix and nail bed keratinize
similarly to the normal epidermis with the formation of keratohyalin; in both areas
typical parakeratotic keratinization follows at a later stage (RUNNE and ORFANOS
1981). Although the same principles of neuroectodermal-mesodermal interactions

LITERATURE
46
described for HF development may also be relevant for nail morphogenesis, only little
is known exactly. Currently, a limited number clues to the molecular controls of nail
growth does exist (PAUS et al. 2007).
2.3.2 Functional anatomy of the nail
In animals the nail is more important as a gripping device or scratching tool,
compared to humans, and shows differences in the anatomical structure (Fig. 2.13).
Sections of the nail in vertebrate groups, including primates and humans, reveal, that
the thickness of the sole horn varies, but that the nail plate is consistent (ZIEGLER,
1954).
Fig. 2.13 Main characteristics of the nail in different animals and humans. (ACHTEN 1968)
Like the HF, the nail unit is composed of keratinized epithelium and supporting
mesenchyme. The epithelial compartment can be divided anatomically into several
compartments contiguous with the epidermis (Fig. 2.14) (BADEN and KVEDAR
1993): The nail plate as a fully cornified structure and the four highly differentiated
epithelial structures: the proximal nail fold (PNF), the nail matrix, the nail bed, and the

LITERATURE
47
hyponychium (ZAIAS 1963). The intimate anatomic relationship between nails and
phalanges determines the shape and curvature of the nails, and is also responsible
for the common occurrence of bone alterations in nail diseases, and vice versa
(PAUS et al. 2007). In contrast to the skin, the nail apparatus lacks of hypodermal
tissue and pilosebaceous units or HF complexes, respectively (PAUS and PEKER
2003). The dermis consists of a fibrocollagenous network, with bundles of collagen
radiating into the periostium of distal phalangeal bones and is situated between the
nail plate and the bone (TOSTI and PIRACCHINI 1999). Because of this special
region with the underlying bone, bacterial and other infections in this region with a
rapid soft tissue spread can cause osteomyelitis of the distal phalanx and increase
the risk of peripheral neuropathy (PAUS et al. 2007).
Fig. 2.14 Schematic longitudinal section of mouse hair follicle and nail unit (A) Proliferating cells reside in the lower bulb region of the hair follicle (the matrix). Their descendents differentiate into distinct epithelial cell types arranged in concentric rings, such as outer root sheath (ORS), inner root sheath (IRS), and hair shaft (cuticle, cortex, and medulla). The matrix surrounds a group of specialized mesenchymal cells called the dermal papilla (DP), which are the source of signals required for hair follicle development. Activated, endogenous Notch1 is detected in the nuclei (red) of the matrix not adjacent to the DP and of the precursors of cortical cells. (B) The nail unit is composed of four types of epithelial cells. The matrix contains proliferating cells that undergo terminal differentiation in the keratogenous zone, producing the nail plate. The second epithelial cell types constitute the nail bed that underlies the nail plate and spans from the matrix to hyponychium. The hyponychium is the third cell type, underlying the free end of the nail plate. The last epithelial cell type is the nail fold, which is the skin enveloping the nail plate. The dorsal and ventral epidermis of the digit is indicated in orange. The bone is marked in gray. Dermal cells reside in areas marked in white. Activated Notch1 is detected in the nuclei (red) of cells in the dorsal matrix. Note that similarities in color between the hair follicle and nail do not imply that these cell types are homologous. (LIN and KOPAN 2003)
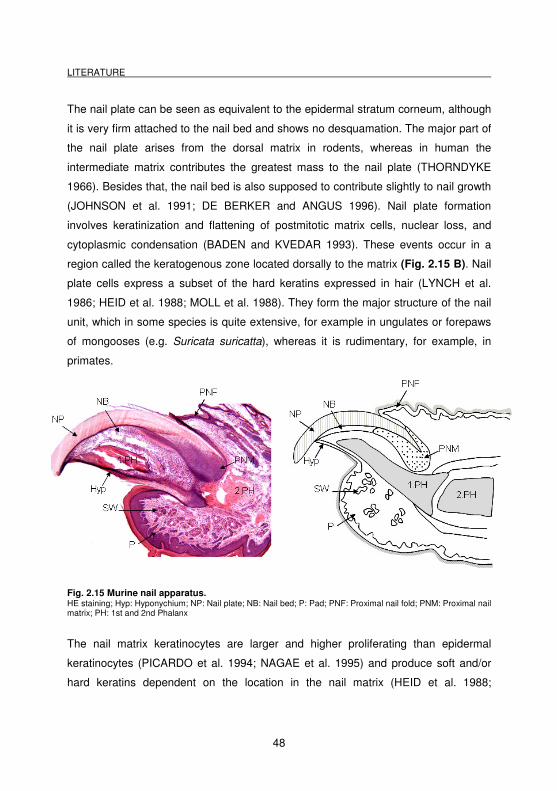
LITERATURE
48
The nail plate can be seen as equivalent to the epidermal stratum corneum, although
it is very firm attached to the nail bed and shows no desquamation. The major part of
the nail plate arises from the dorsal matrix in rodents, whereas in human the
intermediate matrix contributes the greatest mass to the nail plate (THORNDYKE
1966). Besides that, the nail bed is also supposed to contribute slightly to nail growth
(JOHNSON et al. 1991; DE BERKER and ANGUS 1996). Nail plate formation
involves keratinization and flattening of postmitotic matrix cells, nuclear loss, and
cytoplasmic condensation (BADEN and KVEDAR 1993). These events occur in a
region called the keratogenous zone located dorsally to the matrix (Fig. 2.15 B). Nail
plate cells express a subset of the hard keratins expressed in hair (LYNCH et al.
1986; HEID et al. 1988; MOLL et al. 1988). They form the major structure of the nail
unit, which in some species is quite extensive, for example in ungulates or forepaws
of mongooses (e.g. Suricata suricatta), whereas it is rudimentary, for example, in
primates.
Fig. 2.15 Murine nail apparatus. HE staining; Hyp: Hyponychium; NP: Nail plate; NB: Nail bed; P: Pad; PNF: Proximal nail fold; PNM: Proximal nail matrix; PH: 1st and 2nd Phalanx
The nail matrix keratinocytes are larger and higher proliferating than epidermal
keratinocytes (PICARDO et al. 1994; NAGAE et al. 1995) and produce soft and/or
hard keratins dependent on the location in the nail matrix (HEID et al. 1988;

LITERATURE
49
KITAHARA and OGAWA 1993, 1994, 1997). The proliferation in the proximal matrix
is, like in the HF, not restricted to cells in contact with the basement membrane (LIN
and KOPAN 2003). The nail matrix harbors melanocytes in a high density (HIGASHI
1968; HIGASHI and SAITO 1969; TOSTI et al. 1994), which are located
predominantly in the distal matrix and in suprabasal layers in contrast to epidermal
melanocytes located in the entire basal layer. In addition, Merkel cells are also found
in human nail matrix, although more in fetal than in adult nails (MOLL and MOLL
1993).
The nail fold is a specialized epidermal transition zone followed by the matrix. Distal
to the matrix the nail bed is located, a zone containing mitotically inactive cells
(ZAIAS and ALVAREZ 1968; BADEN and KVEDAR 1993). The hyponychium
connects the nail bed with the ventral epidermis of the digit. In addition, the nail
apparatus displays an abundant vascular supply with anastomoses. A network of
three systems (superficial, proximal, distal arcade) supply different parts of the nail
apparatus und thus, guarantee independent sources of blood supply for the nail
matrix, supporting normal nail growth even in the presence of temporary perfusion
deficits (FLECKMAN et al. 1997).
Tab. 2.4 Human nail apparatus – basic data (PAUS et al. 2007)
Nail plate Tightly packed cornified cells (onychocytes), arranged in lamella pattern (weakly eosin+, strongly acid-fast+. Soöver staom reveals three horizontal layers; the largest nail plate is on the first toe (covers about 50% of the digit); fingernails have a longer longitudinal axis compared to the transverse axis, while toenails usually present a greater transverse axis
Nail matrix Thick epithelium situated above the middle part of the distal phalanx of the digits; keratinizes wihtout a granular layer and gernerates the bulb of the nail plate; contains many melanocytes (up to 300/mm2) in African Americans), predominantly in suprabasal layers
Nail bed Thin epithelium consisting of 2-5 cell layers with abrupt keratinization without a granular layer; sparse or no melanocytes; contribution to nail plate fomration debated (nail bed keratinocytes supposedly contribute about on-fifth of the ventral nail plate’s thickness and mass)
Proximal nail fold Epithelium with a granular layer; its dorsal part corresponds to the skin of the dorsum of the digit and contains sweat glands is devoid of pilosebaceous units; densely innervated; the ventral part is thinner and continues into the nail matrix epithelium
Hyponychium An epithelium equivalent to the volar skin containing a granular and a thick conified layer; the hyponychial dermal-epidermal junctioin is papillary as in normal skin
Nail pigmentation Banded pigentation (longitudinal melanoncyhia) is common I Asians or African-Americans (seen in 77% of African-Americans by the age of 20, nearly 100% by the age of 50 years); rare in Caucasians

LITERATURE
50
2.3.3 Growth of the nail
In contrast to the HF, which cycles between growth, regression and resting phases,
the nail grows continuously and during adulthood (RUNNE and ORFANOS 1981).
The special arrangement of the matrix cells and the eponychium and nail bed
determine the direction of growth. The major part of nail plate formation arises from
the dorsal matrix in rodents, whereas in human the intermediate matrix contributes
the greatest mass to the nail plate (THORNDYKE 1966). Besides that, the nail bed
contributes slightly to nail growth (JOHNSON et al. 1991; DE BERKER and ANGUS
1996). The growth rate is determined by the turnover of the matrix cells; the
thickness of the nail, however, by the number of dividing matrix cells. In cases where
a part of the matrix is damaged by trauma, the nail grows upwards in an uncontrolled
manner (RUNNE 1980). On the other hand, digit tip regeneration (e.g. following
amputation at levels distal to the nail matrix) has been reported in non-primate
mammals (BORGENS 1982; MULLER et al. 1999), supporting the importance of the
nail matrix.
Variation in homeostatic control may contribute to some of the many nail
morphologies among vertebrates. Disruption of a similar balance in the epidermis
may be responsible for skin diseases, such as psoriasis. Alteration in nail
homeostasis could also result in longer, shorter, or abnormal nail plates. For
example, the HoxC13 and the nuclear repressor protein hairless (the Rhino allele,
hrrh) are independently involved in the program controlling homeostasis and nail
morphology. In the absence of either protein, long, curled, and continuously growing
nails are formed (AHMAD et al. 1998; GODWIN and CAPECCHI 1998;
MOOKHERJEE et al. 2006). Several other mutations in mice have been described,
which affect dorso/ventral patterning and disturb nail development [e.g. loss of
Wnt7a, or FOXN1, mutations in En1, LMX1B (CYGAN et al. 1997; VOLLRATH et al.
1998; KAWAKAMI et al. 2000; MECKLENBURG et al. 2004)]. In addition, several
other aspects, i.e. physiological variations like differences in the growth of toenails

LITERATURE
51
and fingernails, age or season or pathological aspects like diseases or drugs play an
important role.
2.4 Immunological background for the current study
In the following, a short overview of the two main components of normal immune
system should be given in order to better understand the special condition of
´immune privilege`, which differs from normal immune regulation.
2.4.1 Innate immune system
The innate system is thought to constitute an evolutionarily older defense strategy,
and is the dominant immune system found in plants, fungi, insects, and in primitive
multicellular organisms (JANEWAY et al. 2001). It comprises the cells and
mechanisms that defend the host from infection by other organisms, in a non-specific
manner, often but not only in the early phase. Innate immunity is present in all
individuals at all times, shows no increase in the reaction strength after repeated
exposure to a given pathogen and discriminates between groups of similar
pathogens (ALBERTS et al. 2002; JANEWAY et al. 2005). This means that, unlike
the adaptive immune system, no long-lasting or protective immunity to the host is
constituted. The main functions of the innate immune system in vertebrates include:
a) Recruitment of immune cells (i.e. neutrophils and leukocytes) to sites of
inflammation by the release of chemical factors (chemokine and cytokine interaction).
b) Activation of complement cascades and formation of antibody complexes to clear
pathogens or mark them for destruction by other cells (opsonization, cytolysis,
neutralization). c) Activation of the adaptive immune system through antigen
presentation (MHC class II), d) identification and removal of foreign substances
through phagocytosis by leukocytes.
Natural killer (NK) cells, mast cells, eosinophils, basophils; and the phagocytic cells
including macrophages, neutrophils and dendritic cells, contribute to the immune

LITERATURE
52
system by identifying and eliminating pathogens that might cause infection. These
cells are activated through different mechanisms, i.e., through important pattern-
recognition receptors (PRRs), expressed on macrophages and dendritic cells that
recognize pathogen-associated molecular patterns (PAMPs), Toll-like receptors,
activating receptors (PRRs) and inhibitory receptors (KIRs) for NK cells and the
production of antimicrobial peptides (GOLDSBY et al. 2003; PLAYFAIR and CHAIN
2006).
2.4.2 Acquired immune system
The adaptive immune system is the response of antigen-specific lymphocytes to
antigen, and provides the vertebrate immune system with the ability to recognize and
remember specific pathogens (development of an immunological memory). It is
composed of highly specialized, systemic cells and processes. The responses are
generated by clonal selection of lymphocytes in contrary to the innate immune
system (JANEWAY et al. 2005). Adaptive immunity is activated in vertebrates when a
pathogen evades the innate immune system and generates a threshold level of
antigen (JANEWAY et al. 2005). The major functions of the adaptive immune system
include: a) Distinction between ´self` and ´non-self`antigens, during the process of
antigen presentation (through MHC class I and class II molecules and interaction with
T cells), b) elimination of pathogens or pathogen infected cells, c) The development
of an immunological memory, to immediately eliminate a pathogenic antigen in case
of reoccurence/reinfection.
The cells of the adaptive immune system are lymphocytes, like B cells and T cells. B-
cells are important in the humoral immune response, interacting with T helper cells
and producing antibodies, whereas T cells are involved in cell-mediated immune
responses (GOLDSBY et al. 2003; JANEWAY et al. 2005).

LITERATURE
53
Fig. 2.16 Diagram of innate and adaptive immune system (PLAYFAIR and CHAIN 2006)
2.4.3 Immune privilege: Definition and basic characteristics
The concept of immune privilege was deduced from the acceptance of allogeneic
tumor implants in defined anatomical locations, such as the eye and the brain
(GREENE 1947). Already in the nineteenth century, van Dooremaal (1873) and Zahn
(1884) noted a prolonged survival of human tumor cells and fetal cartilage xenografts
in the anterior chamber of eyes of rabbits (NIEDERKORN 2003). For the historical
perspective, however, the starting point are studies of Medawar transplanting normal
tissues into such sites (MEDAWAR 1945, 1948; BILLINGHAM et al. 1954). Because
these transplanted tissues survived longer than anticipated or were accepted
indefinitely (MUNN and MUNN 2006), the term ´immunological privilege` was

LITERATURE
54
primarily developed to illustrate transplant (e.g. corneal allograft) survival in these
sites (BILLINGHAM and BOSWELL 1953). Thus, the term immune privilege (IP)
included, originally, that a given tissue environment that hosts an allotransplant
relatively protects the transplanted cells from rejection by the host immune system as
a consequence of antigen sequestration (HEAD and BILLINGHAM 1985; BRENT
1997). Historically, it was thought that mainly passive mechanisms, notably physical
and physiologic barriers that maintained local segregation between tissues and
immune cells and antibodies, contributed most for localized immune privilege.
However, with the recognition that the establishment of IP in the fetomaternal
placentar unit is vital for avoiding fetal rejection (MELLOR and MUNN 2000;
EHRLEBACHER 2001) and that ocular IP is indispensable for normal eye function
(NIEDERKORN 2002), this narrow definition of IP was extended to tissue sites where
the local establishment of an immunosuppressive/tolerogenic environment exerts
biologically important functions to suppress a cytotoxic immune attack on the cells
and antigens harbored within these sites (PAUS et al. 2003; ITO et al. 2008b).
Correspondingly, the collapse of eye IP may result in autoimmune uveitis that is
potentially blinding ocular inflammatory diseases (ITO et al. 2008b).
Today, the term IP has become associated with a small list of defined anatomical
compartments that are protected by IP from excessive inflammatory activity and from
the normal immune system, such as the anterior chamber of the eye, the
fetotrophoblast, the testis, the central nervous system behind the blood-brain barrier,
the anagen hair follicle epithelium, the proximal nail matrix, and the hamster cheek
pouch, (NIEDERKORN 2002); (HEAD and BILLINGHAM 1985; STREILEIN 1993;
BRENT 1997; JANEWAY et al. 2001; PAUS et al. 2003, 2005; ITO et al. 2005c).
Collapse of IP is now appreciated to play a major role in different autoimmune
diseases such as multiple sclerosis (BRUNO et al. 2002), mumps orchitis (FILIPPINI
et al. 2001), and autoimmune chronic active hepatitis (LOBO-YEO et al. 1990;
FILIPPINI et al. 2001).

LITERATURE
55
In general, IP is established and maintained, e.g., by (STREILEIN 1993; BRENT
1997; MELLOR and MUNN 2000; FUZZI et al. 2002; NIEDERKORN 2002; PAUS et
al. 2005):
� Absence of lymphatics.
� Establishment of extracellular matrix barriers to hinder immune cell trafficking,
such as blood-retinal barrier, blood-brain barrier.
� Downregulation or absence of classical MHC class I expression, which
sequesters (auto)antigens in tissue sites and hinders their presentation to
CD8+ T cells with a matching T cell receptor.
� Expression of non-classical MHC class I molecules (such as the MHC class Ib
molecules HLA-G in humans and Qa-2 in mice or HLA-E as a ligand of the NK
cell inhibitory receptor CD94/NKG2A. The interaction of HLA-E with this
receptor for example may result in inhibition of NK cell- and cytotoxic T cell-
dependent lysis (GAO et al. 1997; BRAUD et al. 1998; PACASOVA et al.
1999; TRIPATHI et al. 2006).
� Functional impairment of antigen-presenting cells, for example, by
downregulation of MHC class II expression.
� Local production of potent immunosuppressants such as TGF-β1, TGF-β2, IL-
10, and α-MSH, macrophage migration inhibitory factor (MIF) and other
factors.
� Expression of Fas Ligand (FasL) in order to delete autoreactive, Fas-
expressing T cells.
In addition to these mechanisms of IP maintenance, new conditions and factors have
been explored that may also contribute to the generation and maintenance of relative
IP, such as IDO, MIF and others which will be discussed later.
IP is a relative, not an absolute, state and only some of these mechanisms, the
composition and relative importance of which vary between tissue sites, may be
present in a recognized IP compartment (HOBBS et al. 2002; JANEWAY et al. 2005).
Thus, it is an ongoing debate, which and how many mechanisms have to be existent

LITERATURE
56
to call a tissue site immunoprivileged. For example, even though fetal cells
expressing paternal antigens can leave the fetomaternal placentar unit and do evoke
a maternal response (TAFURI et al. 1995), and even though paternal DNA is found in
the lesions of one of the classical dermatoses associated with pregnancy, fetal
rejection is usually not a consequence. This suggests that the critical issue is which a
certain extent a state of tolerance to the alloantigen-bearing cells is established, and
with which quality immunotolerance is maintained throughout the persistence of
these antigens.
2.5 Immune privilege in skin appendages
Skin with its appendages is the largest organ of the mammalian body and is
continuously facing exposure to various kinds of antigens, including viruses, bacteria,
fungi and parasites as well as a large variety of chemical substances and allergens.
The skin immune system responses to most of these agents and has to find the
balance between proper immune defenses, the limitation of damaging defenses,
maintenance of immune tolerance against autoantigens and avoidance of deleterious
autoimmunity and destruction. At least two skin epithelia are now recognized to meet
key characteristics of IP sites, i.e. the anagen hair bulb of the human HF and the
proximal nail matrix (ITO et al 2005c; PAUS et al. 2005). This fact is noteworthy
because the mammalian skin and its IP sites have so far been largely ignored by
mainstream IP. Therefore, one can at best sketch outline of the potential significance
of skin immune privilege, drawing primarily upon the limited available information that
has been published so far on the IP of HF and nail.
HFs are one of the defining skin features of mammals. The HFs undergo life-long
cycles of growth (anagen), regression (catagen) and relative quiescence (telogen),
the so-called HF cycle, which result from complex bi-directional interactions between
the HF epithelium and its specialized, inductive mesenchyme. During anagen
development, a new hair shaft (HS) factory - the anagen hair bulb - is reconstructed
and the HF recapitulates in part key morphological events of its own morphogenesis.

LITERATURE
57
Then the HFs generate pigmented keratin fibres systems (hair shafts), which require
precisely coordinated interactions between hair follicle keratinocytes, melanocytes
and fibroblasts (PAUS and PEKER 2003). Since melanocyte-associated antigens
frequently are the target of cutaneous autoimmunity (e.g. in the context of vitiligo,
halo nevi or malignant melanoma regression) (COTSARELIS and MILLAR 2001), it is
reasonable to expect that HF melanocytes and their pigmentary activities may also
be targeted by autoimmune responses (ITO et al. 2008b). Indeed, it is now believed
that HF melanocyte autoantigens play a key role as potential immune targets in one
of the most frequent autoimmune diseases of man, alopecia areata, where, e.g., the
presence of autoantibodies against melanocyte-associated autoantibodies has long
been appreciated (GILHAR et al. 2005, 2007; PAUS et al. 2005; GILHAR and
KALISH 2006).
More than three decades ago, Billingham had already reported, that HFs provide a
special milieu which enables transplanted allogeneic melanocytes to escape
detection and elimination by the host immune system. In Billingham’s experiments
genetically incompatible homografts of black skin epidermis were transplanted onto
white skin beds (BILLINGHAM 1971) (Fig. 2.17). The result was that the donor
epidermis immediately lost its pigmentation, reflecting rejection of the foreign
melanocytes. However, shortly thereafter, black hair shafts began to pierce the (now
white) epidermis, which could only be explained by the possibility, that at least some
epidermal donor melanocyte allotransplants must have survived or escaped rejection
in the host hair bulbs, and that they had resumed their transfer of melanosomes to
precortical hair matrix keratinocytes (BILLINGHAM and SILVERS 1971; GILHAR and
KALISH 2006).

LITERATURE
58
Fig. 2.17 Billingham’s experiment: Survival of epidermal melanocyte allotransplants in the host anagen HF. When black ear skin epidermis is transplanted onto skin bed of genetically incompatible white guinea pigs, the transplant rapidly loses its pigmentation. This indicates that the allogeneic melanocytes are rejected. But surprisingly, shortly therafter, some black hair shafts pierce the (now white) epidermis – indicating that at least some donor melanocytes have escaped the host immune system and have begun to transfer melanosomes to precortical hair matrix keratinocytes with the result of generating black hair shafts (BILLINGHAM 1971; BILLINGHAM and SILVERS 1971).
Despite these exciting early findings indicating that the anagen hair bulb is a site of
IP, the concept of HF IP still awaits systematic exploration by mainstream
immunology (ITO et al. 2008b) and functional evidence. This has become even more
urgent, since the proximal human nail matrix (ITO et al. 2005c) recently has also
been found to be a site of relative IP, and since the gene and protein profiling data
(MORRIS et al. 2004; TUMBAR et al. 2004; GILHAR et al. 2005; OHYAMA et al.
2006) suggest that the bulge region is a strong candidate as intracutaneous
immunoprivileged site.
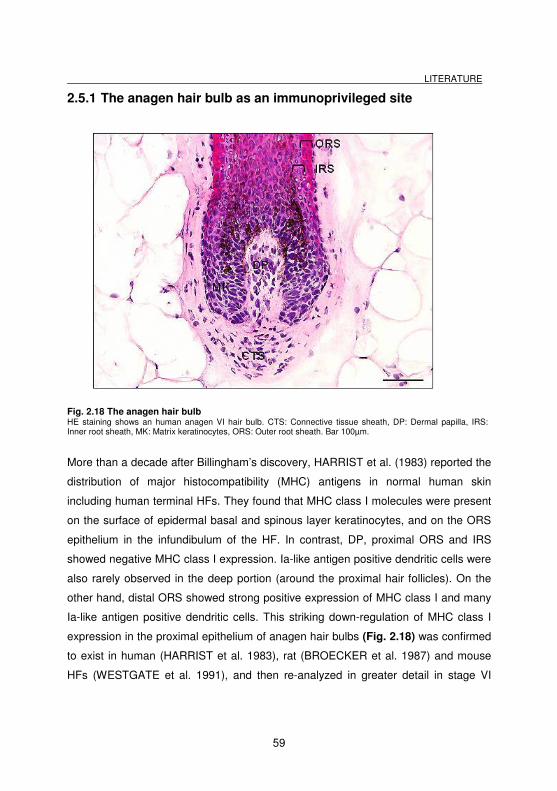
LITERATURE
59
2.5.1 The anagen hair bulb as an immunoprivileged site
Fig. 2.18 The anagen hair bulb HE staining shows an human anagen VI hair bulb. CTS: Connective tissue sheath, DP: Dermal papilla, IRS: Inner root sheath, MK: Matrix keratinocytes, ORS: Outer root sheath. Bar 100µm.
More than a decade after Billingham’s discovery, HARRIST et al. (1983) reported the
distribution of major histocompatibility (MHC) antigens in normal human skin
including human terminal HFs. They found that MHC class I molecules were present
on the surface of epidermal basal and spinous layer keratinocytes, and on the ORS
epithelium in the infundibulum of the HF. In contrast, DP, proximal ORS and IRS
showed negative MHC class I expression. Ia-like antigen positive dendritic cells were
also rarely observed in the deep portion (around the proximal hair follicles). On the
other hand, distal ORS showed strong positive expression of MHC class I and many
Ia-like antigen positive dendritic cells. This striking down-regulation of MHC class I
expression in the proximal epithelium of anagen hair bulbs (Fig. 2.18) was confirmed
to exist in human (HARRIST et al. 1983), rat (BROECKER et al. 1987) and mouse
HFs (WESTGATE et al. 1991), and then re-analyzed in greater detail in stage VI

LITERATURE
60
human anagen scalp HFs (PAUS et al. 1994b). In these studies, the question
whether the human hair follicle bulge, i.e. the area of stem cell, is also relatively
immunoprivileged has been left uncontrolled so far.
Besides MHC class I negativity, additional features of HF biology strongly supported
the concept that anagen hair bulbs in all mammalian species belong to a site with
relative IP (PAUS et al. 2005):
Fig. 2.19 Distribution of peri- and intrafollicular CD4+, CD8+ T cells and CD1a+ cells. The distribution of immune cells in and around human HF differs from that in and below the interfollicular epidermis. Around the human anagen hair bulb, CD4+, CD8+ and CD1a+ cells are only rarely found. (ITO et al. 2008b)
• In the proximal portion of the HF, there is a sharply reduced number of
apparently non-functional, MHC class II antigen-negative, CD1a+ Langerhans cells,
(PAUS et al. 1994b; CHRISTOPH et al. 2000) as compared with the distal part of hair
follicles (upper portion of HFs) (Fig. 2.19).
• In contrast to the ORS distal of the infundibulum of the sebaceous gland, the
anagen hair bulb displays almost none intraepithelial T cells; and in mice, γ/δ-TCR+
lymphocytes are not detected below the bulge region (CHRISTOPH et al. 2000).
Human anagen-VI scalp HFs extremely rarely show CD4+ T cells, and a CD8+

LITERATURE
61
lymphocyte is almost never caught trafficking through the proximal follicle epithelium
(PAUS et al. 1994b) (Fig. 2.19).
• Like the other well defined immune privileged tissues (HEAD and
BILLINGHAM 1985; STREILEIN 1993), the hair bulb is characterized by the absence
of lymphatic drainage pathway and the presence of a special extracellular matrix
barrier around the hair follicle, both of these conditions may contribute to hinder
immune cell trafficking (BROECKER et al. 1987; STENN and PAUS 2001).
• Importantly, anagen hair bulbs in mice and humans express potent
immunosuppressants, such as TGF-β1 (TOKURA et al. 1997; STENN and PAUS
2001), ACTH (SLOMINSKI et al. 1993), and α-MSH (SLOMINSKI et al. 1993, 2000;
BOTCHKAREV et al. 1999; PAUS et al. 1999a; ITO et al. 2005a).
2.5.2 Downregulation of MHC class Ia and NK cells
NK cells are large granular, non-T, non-B lymphocytes, which destroy certain tumor
cells. NK cells are important in innate immunity against viruses and other intracellular
pathogens, as well as in antibody-dependent cell-mediated cytotoxicity (ADCC).
Normally, NK cells attack autologous (e.g. virally infected or malignantly transformed)
cells, with both the allogeneic MHC class I type or absent or low MHC class I
expression. In sites of IP, where weak or absent MHC class Ia expression is the main
characteristic, this becomes a dilemma, because NK cells are attracted, which are
programmed to destroy MHC class Ia-negative cells. In human anagen HFs and
adjacent tissue only very few perifollicular NK cells have been found (CHRISTOPH et
al. 2000). The same is true for the nail apparatus, where only very rarely CD56+ cells
were detected in the mesenchyme adjacent to the PNM (ITO et al. 2005c).
Nevertheless, a small number of T cells and NK cells also exists in sites of IP (i.e.
HF, nail apparatus) (CHRISTOPH et al. 2000; ITO et al. 2007), so there have to be at
least some compensatory mechanisms available against NK cell effects.

LITERATURE
62
NK cells express two types of surface receptors of opposing kinds that control their
cytotoxic activity and help to distinguish infected/foreign from uninfected/self cells.
One type is an ´activating receptor`, the second type are inhibitory receptors, Killer
cell immunoglobulin (Ig)-like receptors (KIR) and heterodimer CD94/NKG2A (LONG
1999; BOEHM 2006), which are specific for MHC class Ia alleles. If target cells
express MHC class I molecules, NK cell activation is prevented through interaction
with KIR by phosphorylation of immunoreceptor tyrosine inhibitory motif (ITIM)
followed by binding to phosphatases (LONG 1999; VIVIER et al. 2002; JANEWAY et
al. 2005; LANIER 2005). NKG2D is expressed not only in NK cells but also in CD8+
T cells and recognizes the MHC class I chain-related A (MICA) molecule on target
cells, which stimulates these immune cells to attack the target cells (BAUER et al.
1999; WU et al. 1999; MIDDLETON et al. 2002).
Fig. 2.20 Activation of NK cell activity In some circumstances, inhibitory receptors recognizing ligands other than MHC class I proteins may suppress NK cell responses. When interacting with target cells expressing ligands for both inhibitory and activating receptors, the outcome is determined by the summation of the strength of signals. The amount of activating and inhibitory receptors on the NK cells and the amount of ligands on the target cell, as well as the qualitative differences in the signals transduced, determine the extent of the NK cell response. (LANIER 2005)

LITERATURE
63
MICA is induced in response to heat or cellular stress. NK cells are therefore active
only against target cells which have lost MHC expression (e.g. after viral infection or
malignant transformation) because then the interaction with KIR is not possible, and
because in these cells NKG2D is expressed on NK cells recognizing the MHC class I
chain-related A gene (MICA) on target cells (BAUER et al. 1999; WU et al. 1999)
(Fig. 2.20).
In order to investigate NK cell activity in the context of HF IP, ITO et al. (2008a)
studied an organ-specific, cell-mediated autoimmune disease on skin from patients
with AA, thought to result from a collapse of HF IP (GILHAR et al. 2005; PAUS et al.
2005; GILHAR and KALISH 2006). Indeed, in skin of AA patients both, a marked up-
regulation of NK cell numbers with a defect in inhibiting perifollicular NK cell
activation was observed (ITO et al. 2008a) and an upregulation of the activating
NKG2D molecule and MICA. The normal anagen HFs may escape from NK cell
attack by combining active suppression of NK cell (e.g. by MIF and KIR up-
regulation) with reducing the chance of NK cells to receive stimulatory signals (e.g.
by down-regulation of NKG2D expression on NK cells and of its ligand, MICA, on the
potential target - HF epithelium). These recently revealed defects must now be taken
into account in AA pathogenesis, and in the development of more effective treatment
scenarios for AA and related autoimmune diseases that are also characterized by IP
collapse (ITO et al. 2005d).
MHC class Ib molecules such as HLA-G and E inhibit cell killing by NK cells (see
chapter 3.10.4). In IP sites, i.e. the fetotrophoblast, HLA-G is recognized by an
inhibitory receptor on the NK cell, which prevents the NK cell from killing placental
cells (JANEWAY et al. 2005). HLA-E binds a very restricted subset of peptides,
derived from the leader peptide HLA-G and by forming a complex with this HLA-G,
HLA-E can bind to the receptor NKG2a on NK cells. This interaction will lead to an
inhibition of the NK cell dependent cytotoxicity (JANEWAY et al. 2005).
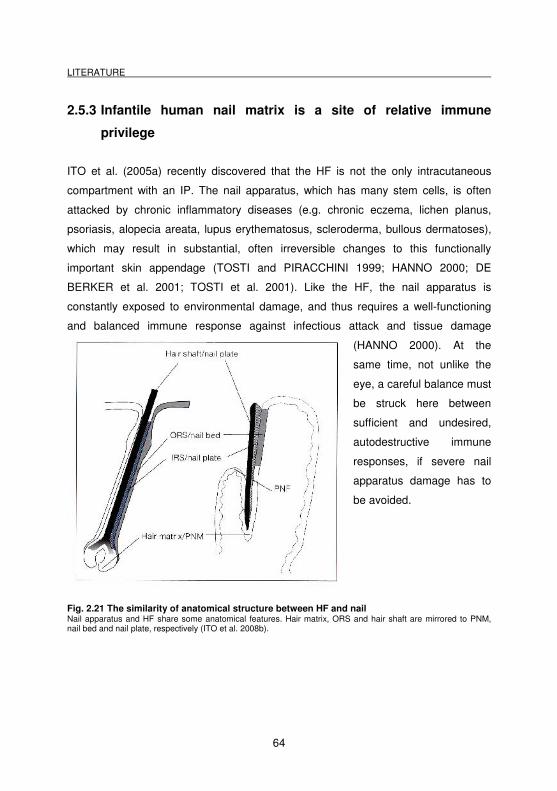
LITERATURE
64
2.5.3 Infantile human nail matrix is a site of relative immune
privilege
ITO et al. (2005a) recently discovered that the HF is not the only intracutaneous
compartment with an IP. The nail apparatus, which has many stem cells, is often
attacked by chronic inflammatory diseases (e.g. chronic eczema, lichen planus,
psoriasis, alopecia areata, lupus erythematosus, scleroderma, bullous dermatoses),
which may result in substantial, often irreversible changes to this functionally
important skin appendage (TOSTI and PIRACCHINI 1999; HANNO 2000; DE
BERKER et al. 2001; TOSTI et al. 2001). Like the HF, the nail apparatus is
constantly exposed to environmental damage, and thus requires a well-functioning
and balanced immune response against infectious attack and tissue damage
(HANNO 2000). At the
same time, not unlike the
eye, a careful balance must
be struck here between
sufficient and undesired,
autodestructive immune
responses, if severe nail
apparatus damage has to
be avoided.
Fig. 2.21 The similarity of anatomical structure between HF and nail Nail apparatus and HF share some anatomical features. Hair matrix, ORS and hair shaft are mirrored to PNM, nail bed and nail plate, respectively (ITO et al. 2008b).

LITERATURE
65
On the basis of the substantial architectural similarities between hair follicle and nail
apparatus (e.g. nail and hair matrix) (Fig. 2.21), it seemed expectable that the –
previously only ill-defined – nail apparatus immune system shows striking similarities
with the HF immune system (ITO et al. 2005c). Immunohistological analysis, in
particular, suggested that nail tissue also harbors a distinct area of IP. ITO et al.
(2005c) showed that HLA-A/B/C expression is prominently down-regulated on both
keratinocytes and melanocytes of the proximal nail matrix (PNM), compared to other
regions of the nail epithelium (Fig. 2.22).
The PNM also shows moderate HLA-G immunoreactivity and strong IR for locally
generated immunosuppressants such as TGF-β1, α-MSH and ACTH, as well as for
macrophage migration inhibitory factor (MIF) as prominent inhibitor of NK cell activity.
Around the PNM, there are only very few CD1a+, CD4+ or CD8+ cells, compared to
nail fold and hyponychium (Fig. 3.5). CD1a+ cells in and around the PNM show
reduced MHC class II and CD209
expression, indicating diminished antigen-
presenting capacity (ITO et al. 2005c).
Fig. 2.22 Distribution of HLA-ABC, CD4+ and CD8+ T cells in the murine nail apparatus (ITO et al. 2008b)

LITERATURE
66
Taken together, this suggests that the nail immune system strikingly differs from the
skin immune system, but shows intriguing similarities to the HF immune system,
including the establishment of an area of relative immune privilege in the PNM. The
functional advantages of establishing IP in the nail matrix are still uncertain (e.g.
stringent containment of inflammation in a skin appendage that is notoriously under
the attack of infectious agents and is prone to trauma-induced inflammatory tissue
damage?). Also, it is unknown yet which key autoantigens are shared between nail
apparatus and HFs, and whether the collapse of IP in the nail matrix may also
underlie the frequent involvement of the nail in AA. However, it is already clear that
our – as yet, poor - understanding of the immunopathogenesis of nail growth
disorders necessitates systemic dissection of the nail immune system and its IP.

LITERATURE
67
2.6 The immune privilege collapse model of AA pathogenesis
Fig. 2.23 ´Immune privilege collapse model` of alopecia areata pathogenesis Some endo-/exogenous factors induce IFN-γ production that upregulate MHC class I in hair bulb. Then, AA-associated follicular autoantigens are presented via MHC class I to autoreactive T cells that result in secondary autoimmune phenomena (ITO et al. 2008b).
More than ten years ago, PAUS and co-workers claimed the hypothesis, which was
the first to apply Billingham’s concept of HF IP to the pathogenesis of AA (PAUS et
al. 1993; PAUS et al. 1994a; PAUS et al. 1999b) (Fig. 2.23). Although this concept
was over years ignored, it is widely accepted today in AA research. While the
pathogenesis of AA still has not been fully elucidated, recent consensus appears that
AA reflects an organ-restricted, T cell-mediated autoimmune disease (ITO et al.
2008b). The most important results in this respect were reported by GILHAR and
KALISH (2006, 2007), and colleagues who demonstrated that AA lesions can be

LITERATURE
68
induced by the transfer of MHC class I-restricted CD8+ T cells alone, that anagen HF
antigens are needed to stimulate T cells for effective triggering of AA lesions after cell
transfer, and that these antigens can be substituted for by HF melanocyte antigens
(WHITING 2003; GILHAR et al. 2005). The key histological feature of AA is a
lymphocytic infiltrate around the lower HF, which may show a characteristic ´swarm
of bees` pattern (SPERLING and LUPTON 1995), but - in chronic AA - can also be
much more discreet than widely assumed (WHITING 2003). CD4+ T-cells
predominate in the infiltrate surrounding the HF, while T-cells within the follicular
epithelium are predominantly of the CD8+ type (TODES-TAYLOR et al. 1984).
In AA lesions, antigen presenting cells including Langerhans cells and macrophages,
infiltrate the dystrophic HFs, and often melanin deposition is observed around these
HFs (HORDINGSKY 2003). Trigger for the pathogenesis could be infectious foci,
bacterial superantigens, psychoemotional stressors, skin microtrauma, or other
damage to the HF, which are possibly aided by predisposing immunogenetic factors.
The result is a peri- and/or intrafollicular rise in AA of IFN-γ secretion, ectopically up-
regulating MHC class Ia expression in the proximal HF epithelium (PAUS et al. 2003;
ITO et al. 2008b). This MHC class I up-regulation occurs in the normally MHC class I-
negative hair matrix of anagen hair bulbs, where immunogenic melanogenesis-
associated antigens are massively generated. Once the HF enters anagen and its
pigmentary unit engages in active melanogenesis [i.e., during anagen III/VI
(SLOMINSKI and PAUS 1993b)], these anagen- and/or melanogenesis-associated
autoantigens are no longer sequestered but can be ectopically presented in the
normally MHC class I-negative epithelial hair bulb and, thus, seriously endangering
maintenance of the HF IP. If an individual has pre-existing autoreactive CD8+ cells,
which in addition receive appropriate co-stimulatory signals and help from CD4+ T
cells (and, possibly, additional signals via CD4 as well), a cytotoxic T cell attack on
the hair matrix is the result. This attack activates a vicious circle of secondary,
follicle-damaging autoimmune phenomena, whose quality and magnitude largely
determine the resulting degree of HF damage (dystrophy) and thus the actual clinical
manifestation, progression, and course of AA (ITO et al. 2008b).

LITERATURE
69
This basic hypothesis was later extended to account for a key role of locally
generated immunosuppressants [e.g. α-MSH, TGF-β1 and IGF-1 (ITO et al. 2004)]
and NK cell-suppressing activities [e.g. MIF (ITO et al. 2005d)] as ´guardians of HF
immune privilege`, the insufficient activity/function of which was thought to
predispose individuals towards AA development, while immune privilege repair via
these agents was suggested to underlie spontaneous AA remission and hair
regrowth (PAUS et al. 2005).
Collapse and restoration of IP in the anagen hair bulb in vitro
As described before, IFN-γ plays an important role in the pathogenesis of AA.
Compared to other cytokines such as IL-1 and TNF-α, IFN-γ offers the most potent
cytokine stimulus for ectopic MHC class I expression in murine pelage HFs in vivo
(RUECKERT et al. 1998). In addition, recent studies have demonstrated that IFN-γ
deficient mice are resistant to the development of alopecia areata (FREYSCHMIDT-
PAUL et al. 2006). ITO et al. have recently developed a standardized and highly
reproducible in vitro assay that recreates the key feature of IP collapse postulated
above in the human system: the ectopic upregulation of HLA-A/B/C expression in the
matrix of normal human anagen scalp HFs. Using this new in vitro assay and very
sensitive immunostaining techniques, confirmed by in situ hybridization and RT-PCR,
they could show that IFN-γ is indeed a very potent stimulator of ectopic MHC class I
expression in microdissected, organ-cultured human scalp HFs from healthy donors
in anagen VI (ITO et al. 2004). Low-dose IFN-γ (75 IU/ml) can be exploited
experimentally to induce IP collapse of normal human anagen HFs in vitro, likely via
an IRF-1-mediated mechanism (ITO et al. 2004). Higher doses of IFN- γ (500 IU/ml)
also act as potent catagen inducer in human scalp HFs (ITO et al. 2005b). The ability
of IFN-γ to induce follicular MHC class I and II was used to test the hypothesis that
AA results from loss of immune privilege. C3H/HeJ female mice were injected
intravenously with IFN-γ to induce follicular MHC. These injected mice demonstrated
an increased rate of development of alopecia areata (GILHAR et al. 2005).

LITERATURE
70
Thus, this in vitro model allows also screening for candidate agents that restore IP by
down-regulating the IFN-γ induced, ectopic MHC class I expression in human
anagen HFs. So far, three immunomodulators, which are known to be locally
produced in the anagen hair bulb, α-MSH, TGF-β1, and IGF-1 (SLOMINSKI et al.
1993, 2000; BOTCHKAREV et al. 1999; PAUS et al. 1999a), demonstrated the
capacity of down-regulating ectopic MHC class Ia expression, on both the protein
and the mRNA level, when added to the culture medium after IFN-γ administration
(ITO et al. 2004). The fact, that natural and by the HF itself generated
immunomodulators might be used for IP restoration in AA is encouraging. These and
other related agents also promise to carry with them minimal risks of toxicity, which
may be further reduced by their topical application (e.g., via HF-targeted liposome
preparations). Today, AA treatment focuses on targeting the secondary autoimmune
phenomena associated with AA, but it is still not convincing or satisfying. Instead, it
would be much more promising to concentrate on the restoration of the HFs lost or
compromised IP - both for preventing the progression of AA lesions and for inducing
hair regrowth. This IP restoration therapy does not require any prior knowledge of the
relevant key autoantigens or the specific autoreactive T cells, and it can resort to the
administration of well-known nonspecific immunomodulators that chiefly down-
regulate ectopic MHC class I expression in the anagen hair bulb.
These reasons explain the need of further evaluating and testing different substances
as possible IP restorer, in order to find a suitable candidate with minimal side effects.

LITERATURE
71
2.7 The human bulge region as a site of relative immune privilege?
Recent gene and protein expression data obtained from mice have raised the
possibility that follicular IP is not purely limited to the anagen hair bulb (COTSARELIS
2006a). By microarray analysis of keratin 15 promoter-driven-GFP labelled ORS cells
of murine pelage HFs, which included bulge stem cells. MORRIS et al. (2004)
identified downregulated transcript levels for several histocompatibility genes (e.g. β2-
microglobulin, H2-Q8, H-2K2, H-2D). Independently, transcriptional profiling of
murine and human bulge label-retaining cells revealed an increase of TGF-β2 mRNA
(TUMBAR et al. 2004; OHYAMA et al. 2006), a molecule that is known to help
maintain the immune-privileged milieu in the eye and brain (NIEDERKORN 2006;
SIGLIENTI et al. 2007).
The concept of bulge IP is further supported by the observation that CD200 IR
preferentially occurs in the bulge region of mice and humans. The cell surface
glycoprotein attenuates inflammatory responses, while CD200-deficient mice develop
an inflammatory, cicatricial alopecia (ROSENBLUM et al. 2004). This has invited the
proposal that CD200 may function as an immunosuppressive, ´no danger` signal for
the HF (ROSENBLUM et al. 2006). OHYAMA et al. (2006) also demonstrated that
the bulge region of normal human scalp HFs prominently expresses CD200 at the
gene and protein level, along with other stem cell markers. Since protection of these
epithelial stem cells from immune destruction is essential for preserving the
regenerative and cycling capacity of HFs (PAUS and COTSARELIS 1999;
COTSARELIS 2006a; TIEDE et al. 2007a), it seems logical that the bulge region
should also have an established relative IP (Fig. 2.24).

LITERATURE
72
Fig. 2.24 The human HF bulge region HE staining shows the human HF bulge with the insertion point of the arrector pili muscle. APM: Arrector pili muscle, BM: Basement membrane, CTS: Connective tissue sheath, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland. Bar 100µm.
However, previous immunohistological analyses have provided contradictory
evidence on this matter. The first study published on MHC class Ia antigen
expression in normal terminal human HFs reported prominent IR in the infundibular
region of the ORS, with diminished expression of HLA-A, B, C and β2-microglobulin
in the isthmus region, although the bulge region had not been specifically analysed
(HARRIST et al. 1983). Subsequent immunohistological studies of MHC class Ia
expression in murine pelage HFs (PAUS et al. 1994b, 1998) did not identify a
prominent downregulation of MHC class Ia antigens in the bulge. However, the

LITERATURE
73
murine MHC class Ia analogue, H-2Db antigen, was slightly downregulated in both
the isthmus and bulge (PAUS et al. 1994b). In a study of normal human skin, the
isthmus region of the ORS in terminal anagen VI HFs clearly expressed HLA-A/B/C
antigens, though possibly at a somewhat reduced level of IR compared to the distal
ORS (CHRISTOPH et al. 2000). In addition, IR for β2-microglobulin (CHRISTOPH et
al. 2000), which stabilizes MHC class Ia molecules allowing proper (self-) peptide
presentation, was also identified (JANEWAY et al. 2005). Furthermore, CD1a+
and/or MHC class II+ Langerhans cells were detected in this region of the ORS,
although their numbers were reduced compared with the distal follicle (CHRISTOPH
et al. 2000). On superficial analysis, these findings supported the concept that HF IP
is restricted to the anagen hair bulb (PAUS et al. 2003, 2005).
Thus, although the murine gene profiling data derived from isolated cultured cells
suggests the existence of an IP within the bulge, convincing protein evidence for this
aspect is still missing. Since IP is a phenomenon that refers to entire tissue
compartments, we aimed to provide in situ protein IR evidence that would support or
refute the hypothesis of bulge IP in human HFs.
2.8 Bulge IP collapse and the pathogenesis of PCA
Cicatricial alopecias (PCA; scarring alopecias, permanent alopecias) are an
uncommon, but clinically important group of inflammatory disorders that result in
permanent loss of human scalp hair. Histological examination reveals the
characteristic replacement of HF with scar-like fibrous tissue (WHITING 2001) and
perifollicular lymphocytic infiltrates that lead to hydrophic degeneration of the basal
layer of the outer root sheath, besides other skin architecture changes.
In contrast to reversible forms of inflammation-induced, autoimmune hair loss (e.g.
alopecia areata) that characteristically attack the actual hair shaft factory (i.e. the
anagen hair bulb), the infiltrate in cicatricial lesions is centered round the eHFSC
located in the bulge (Fig. 2.25 A,B) (PAUS and COTSARELIS 1999; COTSARELIS
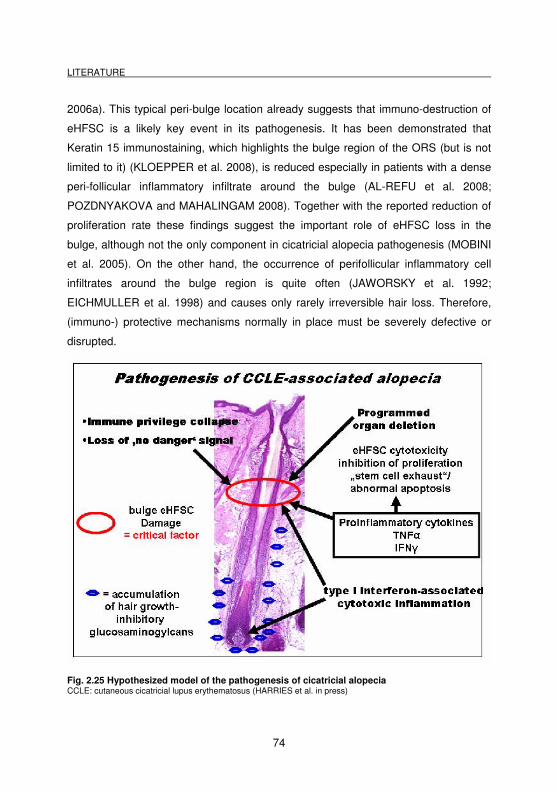
LITERATURE
74
2006a). This typical peri-bulge location already suggests that immuno-destruction of
eHFSC is a likely key event in its pathogenesis. It has been demonstrated that
Keratin 15 immunostaining, which highlights the bulge region of the ORS (but is not
limited to it) (KLOEPPER et al. 2008), is reduced especially in patients with a dense
peri-follicular inflammatory infiltrate around the bulge (AL-REFU et al. 2008;
POZDNYAKOVA and MAHALINGAM 2008). Together with the reported reduction of
proliferation rate these findings suggest the important role of eHFSC loss in the
bulge, although not the only component in cicatricial alopecia pathogenesis (MOBINI
et al. 2005). On the other hand, the occurrence of perifollicular inflammatory cell
infiltrates around the bulge region is quite often (JAWORSKY et al. 1992;
EICHMULLER et al. 1998) and causes only rarely irreversible hair loss. Therefore,
(immuno-) protective mechanisms normally in place must be severely defective or
disrupted.
Fig. 2.25 Hypothesized model of the pathogenesis of cicatricial alopecia CCLE: cutaneous cicatricial lupus erythematosus (HARRIES et al. in press)

LITERATURE
75
On the basis of the recent new microarray data and our current study it was
hypothesized, that loss of relative bulge IP, in combination with the exposure of
eHFSC to a directly cytotoxic immune attack, may therefore play a key role in the
pathogenesis of cicatricial alopecia (HARRIES et al. 2008) (Fig. 2.25). Preliminary
work in patients with cicatricial alopecia indicate that MHC class Ia, β2-microblobulin
and MHC class II IR is significantly up-regulated in the bulge region of lesional skin,
compared with un-involved skin (HARRIES and PAUS, unpublished observations).
While loss of bulge IP offers a plausible explanation for how eHFSC can become
exposed to destructive immune attack, it is still unknown, which intrafollicular
target(s) or stimulators [e.g. IFN-γ (TORO et al. 2000; WENZEL and TUTING 2007;
WENZEL et al. 2007)] of such auto-aggressive immune responses remain to be
identified. It is also unclear whether IP collapse occurs early in the disease process
(i.e. before inflammation develops), or only later on, as a secondary phenomenon.
Since of the irreversible nature of this disease and unsatisfactory therapeutic options
(HARRIES et al. 2008), it is important to further explore the immuno pathogenesis of
cicatrical alopecias and to search for future therapeutic strategies in immuno
protection of eHFSC and restitution of their IP.
2.9 Function of immunoprivileged sites
While it is now clear that IP is indispensable for normal eye function to avoid
inflammatory reactions, and that IP in the fetomaternal placentar unit is vital for the
suppression of fetal rejection (STREILEIN 1993; BRENT 1997; MELLOR and MUNN
2000; EHRLEBACHER 2001; NIEDERKORN 2002, 2003), it still remains to be
satisfactorily demonstrated by convincing functional evidence what exactly HF IP
might be good for. With regard to ocular IP, NIEDERKORN concludes that the
existence of ocular IP is necessary for the normal function of the visual axis, the
preservation of which requires that ocular inflammation is regulated very stringently
and innocent bystander immune damage that may result in irreversible eye damage
is scrupulously avoided (NIEDERKORN 2002; STREILEIN 2003). Along the same

LITERATURE
76
lines, establishment of an IP milieu in the HF may be needed to ensure a safe
environment for HF cycling, the process of which depends on a constitutively active
safeguarding system to protect it against immune injury.
In this context it is noteworthy that, apart from epidermal melanocytes (in vitiligo),
thyroid epithelium (in autoimmune thyroiditis) and the synovium (in rheumatoid or
psoriatic arthritis) (PAUL 1999; JANEWAY et al. 2001), the HF represents one of the
most frequent targets of immune-mediated disease, resulting in the development of
alopecia areata (AA), or even in permanent hair loss (due to immune destruction of
epithelial HF stem cells) as it is seen in the scarring alopecia associated with lichen
planopilaris, lupus erythematodes, scleroderma, and folliculitis decalvans (HERMES
and PAUS 1998; PAUS and COTSARELIS 1999). Therefore, one reasonable
general hypothesis is that HF IP serves to reduce the chances of autoimmune HF
damage (PAUS et al. 2005).
Skin melanocytes are prone to be the target of immune-mediated injury (e.g., in
vitiligo, halo nevi, regressing malignant melanoma, and during immunotherapy of
metastasizing melanoma). Notably, in AA lesions, the characteristic inflammatory cell
attack on lesional HFs almost exclusively targets anagen hair bulbs, which are in the
process of active pigment production (i.e. anagen III-VI HFs). In addition, recovering
HFs in AA patients typically generate white hair shafts (PAUS et al. 1994a; DAWBER
and FENTON 1997). Thus, a more specific hypothetical function of HF-IP is to
sequester melanocyte-/melanogenesis-associated autoantigens from immune
recognition and to protect the hair bulb from potentially deleterious autoaggressive
immune responses. Immunogenetically distinct individuals may differ in their relative
level of protection from, and relative risk of, anti-HF autoimmunity. In case of
constitutively insufficient IP capacity or functional collapse of the HF IP, this would
result in a greatly enhanced risk of autoimmune attack on the follicle (PAUS et al.
1994a, 1999b, 2005).

LITERATURE
77
It is interesting to note that during neonatal HF morphogenesis in mice, in contrast to
the epidermis, distal follicular keratinocytes begin to express MHC class I only at the
late stage of development when almost all skin cells already express MHC class I
molecules, paralleling the timing of maturation of the skin immune system (PAUS et
al. 1998, 1999b; OHYAMA et al. 2006). This observation raises the question if IP is
also needed for the proper development of HF epithelium. Perhaps IP needs to be
established by the time, when the full complement of immunocytes (intraepithelial T
cells and Langerhans cells), immunoregulatory surface molecules, secreted
immunomodulators, and HF melanocytes has been assembled and expressed in
their designated locations.
In any case, the currently available – largely phenomenological – evidence, taken
together, suggests that the HF IP is generated and maintained during each anagen
phase (and then disassembled again during catagen and telogen phases) in order to
sequester potentially deleterious, anagen- and/or melanogenesis-associated
autoantigens from immune recognition by appropriately sensitized CD8+ T cells with
cognate receptors, primarily via down-regulation of MHC class I and by the local
production and secretion of potent immunosuppressants (PAUS et al. 1993, 1999b;
HARRIES et al. 2008; HARRIES et al. in press).
2.10 Immune privilege markers
IP is a relative, not an absolute state, which reflects the net result of multiple
interconnected active and passive mechanisms (FERGUSON and GRIFFITH 2006;
SIMPSON 2006). But it is an ongoing debate, how many and which of these
mechanisms have to be established in a given tissue location to justify the term ´IP`
for this site (FERGUSON and GRIFFITH 2006). Since no universally accepted
criteria have as yet been defined that apply to all IP tissues, ultimate proof for the
human bulge, the murine vibrissal follicle or nail apparatus as sites of relative IP can
only arise from functional studies, such as Billingham’s famous (as yet never
repeated) melanocyte allotransplant experiment in guinea pigs which still represents

LITERATURE
78
the only existing functional evidence of bulb IP (BILLINGHAM 1971; PAUS et al.
2005). Also, in order to demonstrate that a putative ´IP` status of any tissue
compartment is to truly relevant to organ or system homeostasis, it needs to be
shown that loss of IP actually can lead to pathology (FERGUSON and GRIFFITH
2006). Nonetheless, in the following part, we discuss some of the as important
agreed IP markers with their expression patterns and supposed mechanisms.
2.10.1 MHC class I molecules
The major histocompatibility complex (MHC) is a cluster of genes on human
chromosome 6 or mouse chromosome 17. It encodes a set of membrane
glycoproteins called the MHC molecules (JANEWAY et al. 2005) which are grouped
into three classes (I-III). MHC class I is separated to classical (MHC Ia) and non-
classical (MHC Ib). The classical MHC class I gene, is subdivided into HLA-A, HLA-B
and HLA-C in human and Qa, Tla and M in mice. These molecules present peptides
generated in the cytosol in particular to CD8+ T cells. The MHC class I molecule
consists of a single three domain α chain, which is non-covalently associated with β2-
microglobulin. MHC class I is basically expressed on almost all nucleated cells in the
body (LE GAL et al. 1999; JANEWAY et al. 2005), except those in areas of an
established IP (PAUS et al. 2005; NIEDERKORN 2006), which often do have little or
no capacity for regeneration (LAMPSON and FISHER 1984; ABI-HANNA et al. 1988;
LE BOUTEILLER 1994). The lack or down-regulation of classical MHC class I
expression is probably the most characteristic feature of immunoprivileged sites and
tissues.
On the one hand, limited or absent MHC class I expression hinders (auto-)antigen
presentation to autoreactive CD8+ T cells (PAUS et al. 2005) and protects them from
lysis by these cells, contributing to IP. But on the other hand, the reduced or absent
expression of MHC class I molecules creates an immunological dilemma, as it
attracts the attention of NK cells, which are programmed to destroy both cells, with
incompatible MHC class I type and MHC class I-negative cells (LJUNGGREN 1991).

LITERATURE
79
This is especially important for the allogeneic fetus with maternal and paternal
antigens, as NK cells account for 70% of the lymphocytes found in the pregnant
human uterus (NIEDERKORN 2006). Thus, other mechanisms have to compensate
for the paucity or frank absence of MHC class Ia, such as non-classical MHC class Ib
molecules, TGF-β or MIF.
Both skin appendages, the proximal nail matrix and the proximal epithelial anagen
hair bulb of the normal mammalian HF - e.g. in human (BROECKER et al. 1987),
mice (PAUS et al. 1994a) and rat HFs (WESTGATE et al. 1991) - demonstrate virtual
absence of MHC class I-antigen expression.
Interestingly, IR for classical MHC class Ia antigen in the HF is hair cycle dependent,
both in mice and human (PAUS et al. 1994a, 2005). These details have been first
investigated using the murine hair cycle as a model, exhibiting significant differences
between various follicle compartments (Fig. 2.26) (PAUS et al. 1994a). The cycle-
dependency of murine HF MHC class I antigen were illustrated in experiments in
which anagen was induced in the back skin of the mouse (e.g. C57BL/6) by
depilation of the HFs in telogen (PAUS et al. 1990). During the entire hair cycle, the
distal part of the HF continuously showed strong classical MHC class I IR. In the
telogen stage, the entire HF compartments had a strong classical MHC class I IR
except for the dermal papilla. Shortly after anagen induction, this IR pattern changed.
In anagen stages II-III, anagen hair matrix and dermal papilla showed negative
classical MHC class I IR. In anagen stage VI, the IRS, in addition to the hair matrix,
had no classical MHC class I IR, which the MHC class Ia IR in dermal papilla
reappeared. In spontaneously developed catagen HFs, only the slowly receding IRS
keratinocytes and dermal papilla remained MHC class Ia negative (PAUS et al.
1994a; ITO et al. 2008b).
In the human HF cycle corresponding expression changes remain to be elucidated in
greater detail, but MHC class I expression in human anagen VI scalp has also been
investigated (CHRISTOPH et al. 2000; ITO et al. 2004). So far, MHC class I IR was

LITERATURE
80
found on all interfollicular cutaneous cells. In comparison with the epidermis and the
distal ORS, the isthmus region of the ORS showed substantially reduced MHC class
I IR, and a striking restriction of MHC class I IR to basal keratinocytes. The proximal
ORS, IRS, all hair bulb keratinocytes and most of the DP displayed no detectable
MHC class I IR. Sporadically, single MHC class I+ cells were found within the DP. In
contrast, perifollicular fibroblasts and immune cells of the perifollicular CTS were
MHC class I+. In the murine vibrissal follicle or nail apparatus, MHC class I IR has
not been studied yet, although MHC class Ia is one of the most important immune
parameters, and although whisker hairs are often used in experiments on hair
growth.
Fig. 2.26 The cycle-dependency of murine HF MHC class I antigen During the entire hair cycle, the distal part of the HF continuously shows strong classical MHC class I IR, but IR in the proximal or lower HF changes. (PAUS et al. 2005)

LITERATURE
81
2.10.2 β2-microglobulin
β2-microglobulin is a smaller non-covalently associated chain of MHC class Ia
(TOSTI and PIRACCHINI 2008, JANEWAY et al. 2005), is not polymorphic and is not
encoded in the MHC locus. It often serves as an additional verification marker of
MHC class Ia expression. As a result, β2-microglobulin is expressed on almost every
nucleated cell, except for those of IP (CHRISTOPH et al. 2000; PAUS et al. 2005;
NIEDERKORN 2006) demonstrated in the human anagen HF a prominent
expression pattern in the epidermis and distal ORS, whereas IR decreases towards
the lower HF part and was absent in the proximal ORS, IRS, and hair bulb. So far, a
detailed analysis of the bulge region has not been done. In contrast, the
immunoprivileged PNM did not display reduced β2-microglobulin (ITO et al. 2005c).
In addition, β2-microglobulin interacts with the so-called MHC class I pathway-
associated molecules, which are likely to contribute to the compensation work of
MHC class I negativity, and currently available information suggests that they are
intimately involved in maintaining HF IP (ITO et al. 2008b). Briefly, antigen
presentation in the context for the MHC class I molecules to CD8+ T cells requires
the ATP-dependent transporter in antigen presentation (TAP), consisting of the
subunits TAP1 and TAP2 (PAMER and CRESSWELL 1998). TAP1 and TAP2 genes
are in the MHC class II region that must be expressed for MHC class I molecules to
be assembled efficiency (MOMBURG et al. 1994). Peptides generated in the cytosol
by the proteasome are translocated by TAP1 and TAP2 into rough endoplasmatic
reticulum (RER) by a process that requires the hydrolysis of ATP. Within the RER
membrane, newly synthesized class I α chain associates with calnexin until β2-
microglobulin binds to the α chain. The class I α chain-β2 microglobulin heterodimer
then binds to calreticulin and the TAP-associated protein tapasin. When a peptide
delivered by TAP is bound to the class I molecules, folding of MHC class I is
complete and it is released from the RER and transported through the Golgi to the
surface.

LITERATURE
82
The expression of these MHC class I pathway molecules, such as TAP and β2
microglobulin was analysed in human and murine HFs. ITO et al. (2004) showed that
β2-microglobulin and TAP2 IR mirrored the low or absent expression level of HLA-
A/B/C molecules in the proximal ORS and hair matrix of anagen hair bulbs compared
to distal ORS and epidermis in the normal human scalp skin sections (Fig. 2.27)
Therefore, this immunoprivileged region of the skin epithelium is characterized by a
down-regulation not only of HLA-A/B/C and β2 microglobulin but also of major MHC
class I pathway molecules. This reflects the situation in murine HFs in vivo and
underscores the relative defect of these follicular tissue compartments in the
presentation of self-peptides to CD8+ T cells (RUECKERT et al. 1998).
Fig. 2.27 The expression of MHC class I pathway molecules on the murine anagen HF. The expression of the MHC class I pathway molecules TAP and β2-microglobulin is mirroring the IR of HLA-A/B/C molecules. (ITO et al. 2008b)
2.10.3 MHC class II molecules
MHC class II antigens are encoded by the class II region of the MHC complex,
expressed mainly on B cells, macrophages (CD11b on mice) and dendritic cells, and
are involved in the interaction of these with CD4+ T cells. MHC class II molecules are
composed of two non-covalently associated transmembrane glycoprotein chains, α
and β, which also have two domains each. In IP sites it is considered that the
capacity of APCs (i.e. LC and macrophages) may be impaired by a downregulation of

LITERATURE
83
MHC class II (NIEDERKORN 2003; PAUS et al. 2003, 2005). As a consequence, no
or less activation of naïve T cells occurs, because activation and antigen
presentation requires the simultaneous delivery of a co-stimulatory signal, together
with the expression of both MHC molecules (MHC class I and II) (FITZPATRICK
2008; JANEWAY et al. 2005).
In eye IP it was found, that the single corneal endothelial cell layer that lines the
anterior chamber expresses little or no detectable class II MHC (and MHC class I)
determinants (NIEDERKORN 2002). In addition, other factors like TGF-β2 alter the
behavior of APCs, downregulate their MHC class II molecules and their production of
the TH1-inducing cytokine IL-12, while stimulating their synthesis of IL-10 (which
activates TH2 response) (D`ORAZIO and NIEDERKORN 1998; NIEDERKORN
2002). Since IP is composed of several different mechanisms it is unclear, which
effect was first and which are induced by others.
MHC class II+ cells or NLDC-145+ Langerhans cells are essentially absent (PAUS et
al. 1998, 1999b) in the murine anagen hair bulb. Very few CD1a+ or ultrastructurally
identified LCs must be also functionally impaired because they do not express MHC
class II (MOSELEY 1997; CHRISTOPH et al. 2000). In the human HF, CHRISTOPH
et al (2000) reported that HLA-DR+ cells representing LC were prominently
expressed in the epidermis, but were particularly densely distributed in the ORS
surrounding the follicular canal and around the sebaceous gland. The isthmus region
and the proximal ORS showed sharply reduced numbers of MHC II+ cells compared
with the distal ORS and the bulb displayed only very rarely positive cell, the DP none.
In the CTS MHC class II+ cells were, in particular, rare in the isthmus region,
whereas in the distal CTS and around the hair bulb the number was higher. Although
the work of CHRISTOPH et al. (2000) was detailed, they had not focused on the
bulge region or distinguished between isthmus, bulge and proximal ORS.
In the nail apparatus, LC and macrophages in or around the PNF showed the
expected strong HLA-DP/DQ/DR IR, suggesting that these CD1a+ or CD68+ cells

LITERATURE
84
normally perform their well-recognized ´professional` antigen-presentation functions
via MHC class II. In contrast, intraepithelial LC in dorsal nail matrix and dermal
macrophages surrounding the entire nail matrix revealed no or very low IR for HLA-
DP/DQ/DR (ITO et al. 2005c).
2.10.4 MHC class Ib molecules
The non-classical MHC class Ib molecules in humans are HLA-E, -F, -G and –H, in
mice they are called H-2Qa1, to Qa10, H-2T1 to T24, and H-2M1 to M7. These
molecules are encoded within the MHC gene but not highly polymorphic like the
MHC class Ia and MHC class II molecules, and present only a restricted set of
antigens. Non-classical MHC class Ib molecules such as HLA-G and HLA-E in
humans or Qa-2 in mice are expressed in the immunoprivileged fetotrophoblast
(KOWATS 1990; JANEWAY et al. 2005; MOSCOSO et al. 2006; YIP et al. 2006), the
eye (NIEDERKORN et al. 1999; LE DISCORDE et al. 2003), and the proximal nail
matrix (ITO et al. 2005c), and are therefore thought to help maintaining IP
(JANEWAY et al. 2001; FUZZI et al. 2002).
HLA-G and HLA-E impair specific cytolytic T cell functions and have the capacity to
engage the NK cell inhibitory receptor CD94/NKG2 preventing lysis by NK cells
(KOWATS 1990; BRAUD et al. 1998; LEE 1998; LE GAL et al. 1999; ROUAS-
FREISS et al. 2000; TRIPATHI et al. 2006). Originally it was proposed that HLA-G
functions as a ligand with CD94/NKG2A receptor inducing the inhibitory effect on NK
cells. Today, it seems as if HLA-G only acts as a leader peptide, which brings the
mesenchymal-facing intracellular located HLA-E to the cell surface by forming a
complex, and that the following interaction of HLA-E with the CD94/NKG2A receptor
may result in inhibition of NK cell- and cytotoxic T cell-dependent lysis. HLA-E also
might interact with CD8+ T cells directly (GAO et al. 1997), and it is reported that
trophoblast cells also elaborate a soluble isoform of HLA-G that induces apoptosis of
activated CD8+ T cells (CAROSELLA et al. 2000; NIEDERKORN 2006).

LITERATURE
85
In fact, expression of a mouse non-classical MHC class I equivalent, Qa-2, could be
detected in the periinfundibular region of the murine outer root sheath throughout the
entire hair cycle (PAUS et al. 1994a). The hair cycle-independent expression of Qa-2
raises the possibility that these MHC class Ib molecules are involved in the regulation
of follicular IP, rather than in the intriguing, non-specific anti-infection defense of the
HF immune system (HIS) (PAUS et al. 1994a; PAUS et al. 1998). It also encourages
the hypothesis that MHC class Ib expression may play a similar T and NK cell-
inhibitory role in the context of the HF-IP as it does in the fetomaternal placentar unit
(LE GAL et al. 1999). In addition, ITO et al. (2005c) demonstrated a strong up-
regulation of HLA-G expression in the proximal nail matrix compared to the proximal
nail fold. They also concluded that HLA-G expression may serve to inhibit an attack
of NK cells on this MHC class I negative tissue.
2.10.5 CD4+ and CD8+ T cells
CD4 (cluster of differentiation 4) and CD8 are transmembrane glycoproteins. CD4 is
expressed on the surface of T helper cells, regulatory T cells, monocytes,
macrophages, and dendritic cells (JANEWAY et al. 2005). On T cells, CD4 is a co-
receptor that assists the T cell receptor (TCR) to activate its T cell following an
interaction with an antigen presenting cell. CD4 amplifies the signal generated by the
TCR by recruiting a tyrosinase kinase, which is essential for activating many
molecules involved in the signaling cascade of an activated T cell. In addition, CD4
also interacts directly with MHC class II molecules on the surface of the APC. CD8
also serves as a co-receptor for the TCR. Like the TCR, CD8 binds to MHC class I
protein and attacks the cell, if it has no or foreign MHC class I protein on its surface.
T is predominantly expressed on the surface of cytotoxic T cells, but can also be
foundon NK cells. To function, CD8 forms a dimer, consisting of a pair of CD8 chains.
It was demonstrated, that contrary to the epidermis, the sebaceous gland and the
rest of the hair follicle epithelium, the anagen hair bulb is almost devoid of
intraepithelial T cells; in humans, CD4+ or CD8 T cells here are extremely rare in the

LITERATURE
86
proximal HF epithelium (CHRISTOPH et al. 2000), whereas in mice no γδ-TCR
lymphocytes are found at all here (PAUS et al. 1994 ; PAUS et al. 1998).
In the human nail apparatus ITO et al. (2004) demonstrated a high density of CD4+ T
cells in PNF, accumulated CD4+ T cells in epithelium and mesenchyme, that were
not observed in normal human skin, and very rarely detected CD4+ T cells in
adjacent proximal nail matrix (PNM) resembling the hair bulb. Essentially, the same
observation was made for CD8+ T cells, even though their total number was
2.10.6 α-MSH and ACTH
α-melanocyte-stimulating hormone is a tricapeptide derived from a precursor
hormone called propiomelanocortin (EIPPER and MAINS 1980). This POMC
molecule is source for several bioactive peptides including adrenocorticotropin
(ACTH), melanocyte-stimulating hormone (MSH) and the endogenous opioid β-
endorphin (LUGER et al. 2000). Although POMC peptides were originally considered
as neuropeptides, it is now well established that many peripheral tissues including
the skin (BOEHM et al. 2000; SLOMINSKI et al. 2000) autonomously express POMC
and process it via expression of distinct enzymes to POMC-derived peptides. For
example, human HFs are an important extrapituitary site of melanocortin expression
and show prominent α-MSH expression (ITO et al. 2005a; KAUSER et al. 2005). It is
important to note that the tridecapeptide sequence of α-MSH is contained within
ACTH, so that sometimes both molecules are detected by immunohistochemistry and
are expected to display similar functions.
Many studies over the last years revealed a lot of effects on different levels. Besides
its pigmentary effects, α-MSH also has potent protective and anti-inflammatory
effects on central cells and on peripheral non-immune cells, that express
melanocortin receptors (MC-R). Five MC-Rs exist, which belong to the superfamily of
G-coupled receptors with 7 transmembrane domains (CONE et al. 1996; LUGER et
al. 1999; ROOSTERMAN et al. 2006). In many animal models for different diseases,
these anti-inflammatory effects have been validated and it was shown that α-MSH

LITERATURE
87
affects various pathways, i.e. NF-κB activation, expression of adhesion molecules
and chemokine receptors, production of pro-inflammatory cytokines and mediators,
IL-10 synthesis, cell proliferation and activity, inflammatory cell migration and
expression of anti-oxidative enzymes and apoptosis (LUGER and BRZOSKA 2007;
TAYLOR 2007) (Fig. 2.28).
Fig. 2.28 The different roles of α-MSH as immunomodulator (LUGER et al. 2000)
In addition or as a result, it is thought that these molecules provide the special
environment in immunoprivileged sites. TAYLOR and co-workers detected α-MSH in
pM levels in the aqueous humor of the eye and found that such naturally occurring
doses of the peptide had suppressive effects on the production of IFN-γ by antigen-
stimulated primed murine lymph node cells (TAYLOR et al. 1992, 1994; TAYLOR
2007). Further studies investigated the effect of α-MSH on T cells and showed that
aqueous humor treated T cells were capable of suppressing inflammation induced by
delayed-type hypersensitivity T cells and that CD25+CD4+ Tregs were induced,

LITERATURE
88
which suppressed the in vitro production of IFN-γ by other inflammatory T cells
(TAYLOR et al. 2000; NAMBA et al. 2002). In the meantime, at least four different
aqueous humor factors have been identified, suppressing DTH responses in the eye,
namely α-MSH, TGF-β, vasoactive intestinal peptide (VIP), and calcitonin gene-
related peptide (CGRP) (TAYLOR 1999).
In astrocytes and microglia of the CNS α-MSH suppressed TNF-α production
presumably via the MC1R (WRONG et al. 1997), as also shown for other cell types
from other tissues (TAHERZADEH et al. 1999). Moreover, recent studies
demonstrated that α-MSH suppresses experimental autoimmune encephalomyelitis
(EAE), and can promote re-establishment of immune tolerance to autoantigens in the
brain (TAYLOR and KITAICHI 2007) and in the human anagen hair bulb (ITO et al.
2004; LUGER and BRZOSKA 2007). Since α-MSH also blocks accessory signals
such as CD86 and CD40 and additionally induces suppressor factors such as IL-10,
it may be one of the signals required for the downregulation of an immune response
and possibly the induction of tolerance (LUGER et al. 1999).
Earlier studies have also demonstrated that the POMC-derived peptides α-MSH and
ACTH are principal inducers of human epidermal pigmentation via their action at the
melanocortin-1 receptor (MC-1R) (ROUSSEAU et al. 2007). α-MSH also promotes
human HF pigmentation by up-regulation of intrafollicular melanogenesis,
melanocyte dendricity, and melanocyte proliferation (KAUSER et al. 2005).
POMC gene transcription and translation is again hair cycle-dependent in the skin of
C57BL/6 mice and increases significantly in the active growth stage of the hair cycle
(anagen) (SLOMINSKI et al. 1992). Since these molecules have immunosuppressive
effects they may provide the special environment for HF IP. On normal skin, ACTH
immunoreactivity is localized on the ORS cells of anagen HFs that is not detected in
the epidermis and dermis of corporal skin (SLOMINSKI et al. 1993). During anagen,
keratinocytes of the ORS and hair matrix express α-MSH (PAUS et al. 1999a). The
concentration of ACTH significantly increases during depilation-induced anagen
measured by radioimmunoassay (SLOMINSKI et al. 1998). Together with the finding

LITERATURE
89
that the epithelium of human anagen VI HFs in scalp skin also prominently expresses
α-MSH and ACTH, it became obvious that ACTH stimulates intrafollicular cortisol
generation (ITO et al. 2005a), and α-MSH is a potent general immunosuppressant
(COOPER et al. 2005). These results indicate that both melanocortins may contribute
the maintenance of IP milieu in anagen stage.
2.10.7 TGF-β
The transforming growth factor β belongs to a family of multifunctional cytokines that
have wide-ranging and opposing effects on many cellular processes, including
development, cell growth, cell differentiation, carcinogenesis, wound healing, fibrosis,
inflammation and host defense (LETTERIO and ROBERTS 1998). Three isoforms of
TGF-β are described in mammals (TGF-β1, TGF-β2, TGF-β3), which are all encoded
by a different gene and have a different expression, regulation and function
(DERYNCK et al. 1985; DERYNCK et al. 1988; SIGLIENTI et al. 2007). The
signalling occurs through transmembrane serin/threonine kinase receptors (KHALIL
1999; OKUNIEFF et al. 2005). Knockout mice for TGF-β have nonoverlapping
phenotypes, indicating diverse noncompensated functions of TGF family members
(SHULL et al. 1992; KULKARNI et al. 1993).
In addition, pathways mediated by TGF-β may obscure immune surveillance
mechanisms, resulting in failure to recognize or respond adequately to self, foreign,
or tumor-associated antigens. This is why TGF-β is also considered to play an
important role for improvising niches of IP, by recruitment of a cohort of powerful
immunosuppressive cells (COBBOLD et al. 2006; WAHL et al. 2006). The role of
TGF-β2 in IP has been demonstrated in the brain (FABRI et al. 1995; SIGLIENTI et
al. 2007) and the anterior chamber of the eye (STREILEIN et al. 2002;
NIEDERKORN 2003, 2006; CAPSI 2006; TAYLOR and KITAICHI 2007). In the brain
for example, it was shown that downregulation of TGF-β2 facilitates inflammation in
the CNS by astrocyte/microglia interactions (SIGLIENTI et al. 2007). In the eye
chamber, TGF-β is present in the aqueous humor at concentrations that produce

LITERATURE
90
profound inhibition of NK cell-mediated cytoloysis (APTE and NIEDERKORN 1996;
APTE 1997; APTE et al. 1998; NIEDERKORN 2003, 2006).
TGF-β influences the behaviour of APCs in promoting differentiation of Ag-specific T
cells into CD4þCD25þFoxp3þ regulatory T cells (Tregs), which are found in infiltrating
tumors and other sites of IP, where they influence CD8+ T cells and CD4+ T-helper
cells (Th1, Th2, Th17) leading to immune tolerance (D`ORAZIO and NIEDERKORN
1998; TAKEUCHI et al. 1998; KEZUKA and STREILEIN 2000; STREILEIN et al.
2002). Through a similar mechanism Tregs also impair NK cell activity to block
immune surveillance. Moreover, TGF-β inhibits IL-2 dependent proliferation of Ag-
specific T cells (WAHL et al. 2006).
TGF-β1 and TGF-β2 are also considered to contribute to the HF IP (PAUS et al.
2003; PAUS et al. 2005; NIEDERKORN 2006). TGF-β1 is prominently expressed in
the HF epithelium of mice and man, and is strongest during late anagen and the
onset of catagen in cells of the ORS and epithelial strand (FOITZIK et al. 1999,
2000). TGF-βRII positive cells were also found in the proximal and central region of
the ORS during the late anagen and catagen induced hair growth in mice (WELKER
et al. 1997; FOITZIK et al. 2000; NIEDERKORN 2003; ITO et al. 2008b). In addition,
TGF-β1 was shown to have the capacity to restore IP after an IFN-γ induced collapse
(ITO et al. 2004). Besides that, in the nail apparatus and, in particular, in the PNM
prominent expression of TGF-β1 besides other immunosuppressants (ACTH, α-MSH,
IGF-1, MIF) was found (ITO et al. 2005c). In addition, several other effects of TGF-β1
have been reported, such as an elusive endogenous regulator of catagen induction in
vivo in mice (FOITZIK et al. 2000).
In HF biology it is not known yet, whether the anagen HF recruits all these factors to
maintain and restore its IP (PAUS et al. 2005; ITO et al. 2008b). In addition, the
production/secretion of α-MSH, TGF-β, IGF-1, IL-10, and neutrophins (with the latter
putatively being capable of deleting autoreactive CD8+ T cells via stimulation of the
p75 neurotrophin receptor) may be up-regulated whenever the HF becomes the

LITERATURE
91
target of immune injury and/or is threatened with a collapse of its IP. On the other
hand, HF-derived α-MSH, TGF-β, IGF-1, IL-10, well-recognized natural
immunosuppressants (LUGER et al. 1998; PAUL 1999; JANEWAY et al. 2001), may
efficiently dampen the secondary autoimmune phenomena that we envisage as
driving force behind the specific clinical characteristics and the progression of AA in
any given patient (SLOMINSKI et al. 1992). Therefore, together with ACTH, α-MSH,
and cortisol, TGF-β1 likely constitutes an integral part of an entire system of
intrafollicularly generated, secreted ´IP maintenance/protective factors` (PAUS et al.
2005), which remains to be dissected comprehensively.
2.10.8 MIF
MIF is a highly conserved protein, with homologues found in evolutionary divergent
organisms including vertebrates, insects, worms and plants (VIGANO et al. 2007).
Originally, it was described as a factor able to inhibit the random migration of
macrophages (RICH and LEWIS 1932) produced by activated T cells (GEORGE and
VAUGHAN 1962), and has been shown to regulate the immune response by
affecting a number of macrophage and lymphocyte features including cytokine
synthesis, cell activation, and phagocytosis. In addition, it has been implicated to play
a role in several immune and inflammatory conditions, including sepsis (BOZZA et al.
1999), delayed type hypersensitivity (BLOOM and BENNET 1966; BERNHAGEN et
al. 1996), and rheumatoid arthritis (LEECH et al. 1999).
MIF protein is ubiquitously expressed by a variety of cells (although primarily by T
cells and macrophages) indicating a more far-reaching non-immunological
involvement in a variety of pathologic states (SHIMIZU 2005). In normal human skin
(SHIMIZU et al. 1996) localized cytoplasmic MIF IR in keratinocytes, especially in the
epidermal basal layer, in endothelial cells, eccrine sweat ductal cells and
myoepithelial cells of eccrine sweat glands, whereas in lesional psoriatic human skin
IR was up-regulated (STEINHOFF et al. 1999).

LITERATURE
92
MIF is also expressed in the classical IP sites as the fetotrophoblast (VIGANO et al.
2007), eye (APTE et al. 1998), brain (GALAT et al. 1993), human proximal nail matrix
(ITO et al. 2005c), and human anagen scalp HFs (ITO et al. 2007), and is considered
to play a pivotal role in IP maintenance by suppression of NK cell activity (APTE et
al., 1998; TAYLOR 2003; CASPI 2006; PAUS et al. 2005). MIF may inhibit NK-cell
mediated cytotoxicity against MHC class I-negative cells by preventing the release of
cytolytic perforin granules from NK cells (APTE and NIEDERKORN 1996; APTE et al.
1998; ARCURI et al. 2006; NIEDERKORN 2006). This would explain why immune-
privileged tissues that are highly vulnerable to lysis by NK cells, due to their lack of
MHC class I expression, are not under constant NK cell attack. So far, MIF IR was
shown to be present throughout most of the epithelium of normal anagen scalp HFs,
particularly in the proximal IRS and ORS, whereas the epithelium of lesional AA HFs
displayed greatly reduced or absent MIF IR. This suggests that HFs in established
AA exhibit a decreased capacity for suppressing undesired NK cell functions. In the
human nail apparatus, MIF IR is upregulated in the PNM compared with the PNF and
the epidermis (ITO et al. 2005c).
2.10.9 IDO
IDO is a nonomeric heme-containing and rate-limiting enzyme for tryptophan
degradation in extrahepatic tissues (HAINZ et al. 2007; MUNN and MELLOR 2007).
It catabolizes tryptophan into N-formylkynurenine which is converted into 3-
hydroxyanthranilic acid (HIGUCHI and HAYAISHI 1967; KWIDZINSKI and
BECHMANN 2007), ultimately leading to the biosynthesis of the cellular co-factor
nicotinamide adenine dinucleotide (NAD+). Tryptophan is an essential amino acid,
and is required for normal T-cell proliferation, but also for protein synthesis and for
the generation of the neurotransmitter serotonin (HAINZ et al. 2007).
IDO expression is inducible in response to pro-inflammatory cytokines such as IFN-γ,
LPS and IFN-β (CARLIN et al. 1989; CURRIER et al. 2000) in many tissues and cell
types including macrophages, dendritic cells, tumor cells, bone marrow stromal cells,
astrocytes, endothelial cells (HAINZ et al. 2007; KWIDZINSKI and BECHMANN

LITERATURE
93
2007), but most prominently in APCs (HAINZ et al. 2007). Previous observations
made from other immunoprivileged sites, e.g. lung or allografts (SWANSON et al.
2004; LI et al. 2006) skin (VON BUBNOFF et al. 2004), anterior chamber of the eye
(CHEN et al. 2007), fetus (MELLOR and MUNN 2008) and pancreatic islet cells
(JALILI et al. 2007) indicated, that IDO has a yet un-known important role in the
maintenance of IP.
The catabolism of tryptophan by IDO expression in tissues or APCs limits T cell
proliferation by creating a tryptophan-depleted environment and also affects NK cell
function, reduces inflammation and enhances Fas-mediated T cell apoptosis via their
tryptophan degradation products (MUNN et al. 1998; FALLARINO et al. 2002;
MELLOR and MUNN 2008). For example, the number of allogeneic concepti in
female mice treated with IDO inhibitor (1-methyl tryptophan) was reduced
significantly compared to control mice (COBBOLD et al. 2006; ITO et al. 2008b).
Thus, IP may be maintained as a result of the enzymatic activity IDO (MELLOR and
MUNN 2000), which downregulates T cell function by tryptophan depletion (MUNN et
al. 1998). In addition, it was found, that murine regulatory T cells can condition
dendritic cells to express IDO functional activity and to, thus, exercise
immunosuppressive functions in the context of IP (XU et al. 2000; FALLARINO et al.
2003). Downregulation of MHC class I levels on the surface of IDO-expressing
keratinocytes (LI et al. 2004), inhibition of complement activation (XU et al. 2000) and
a placenta-specific, β2-microglobulin-dependent process linked to MHC class I
antigen presentation (HOBBS et al. 2002) are other examples for additional, newly
recognized mechanisms that favour maternal immunotolerance of pregnancy; uterine
natural killer cells may also be involved (EHRLEBACHER 2001). But so far, no IDO
protein expression in normal skin has been shown.

LITERATURE
94
Fig. 2.29 Molecular mechanisms of IDO-induced immunosuppression (A) IDO initiates the rate-limiting step in the degradation of tryptophan along the kynurenine pathway. (B) IDO enzymatic activity results in the local depletion of tryptophan and a local increase in the concentration of downstream metabolites. The decrease in tryptophan can cause a rise in the level of uncharged transfer RNA (tRNA) in neighboring T cells, resulting in activation of the amino acid-sensitive GCN2 stress-kinase pathway. In turn, GCN2 signaling can cause cell cycle arrest and energy induction in responding T cells. The local increase in tryptophan metabolites can cause cell cycle arrest, apoptosis, and differentiation of new Tregs from uncommitted CD4+ T cells. (MUNN and MELLOR 2007)

LITERATURE
95
2.10.10 CD200
The type-1 transmembrane cell surface glycoprotein CD200 (formerly Ox-2) is
expressed in a diversity of tissues on cells of both hemopoietic and non-hemopoietic
origin, including thymocytes, neurons of the central nervous system and the retina,
cells of the glomeruli, cells of degenerating ovarian follicles, vascular endothelium,
some T cells, B cells and dendritic cells in both peripheral blood and lymphoid tissue
(BUKOVSKY et al. 1983; CLARK et al. 1985, 2003). This pattern of CD200
expression is largely conserved in mice, rats and humans (WRIGHT et al. 2001).
Recently, it has been demonstrated that the normal HF epithelium in mice and man
also prominently expresses CD200, a prototypic ´no-danger` signal (OHYAMA et al.
2006; ITO et al. 2007; MEYER et al. 2008a). The expression of CD200 has
previously been shown in the mouse HF, in which it was predominantly detected in
the outermost layer of ORS throughout the length of the mouse HF and could not be
localized in the bulge area. But gene expression studies have identified CD200
transcripts as being up-regulated in the hair bulge region of humans and mice, and
CD200 has become a useful bulge marker for the selection of epithelial stem cells
(TUMBAR et al. 2004; KLOEPPER et al. 2008). In the human anagen HF
homogenously CD200 IR was found in the outermost layer of the ORS, in the bulge,
in the proximal part of the isthmus, in the DP (including blood vessels), and in the
companion layer. The arrector pili muscle and the sweat gland mesenchyme
displayed most prominently IR (KLOEPPER et al. 2008).
When CD200 interacts with the CD200 receptor (CD200R), it induces intracellular
signalling within receptor bearing cells (WRIGHT et al. 2001, 2003) and delivers a
immune-inhibitory signal, which reduces inflammation and promotes tolerance
(PAUS et al. 2003; MORRIS et al. 2004). In vitro, CD200+ dendritic cells inhibit the
secretion of pro-inflammatory cytokines by activated T-cells (MORRIS et al. 2004),
and CD200R binding on APC down-regulates APC activity (ROSENBLUM et al.
2004). Therefore, CD200-CD200R interactions are thought to act as a “no danger”

LITERATURE
96
signal that links innate and adaptive immunity in a given tissue location so as to
dampen the chance of undesired immune responses (MORRIS et al. 2004). In
addition, CD200 is thought to influence the IDO production (FALLARINO et al. 2002),
that could play a yet undefined role in the maintenance of relative IP (see 3.10.9).
Loss of this ´no danger` signal could be important in the pathogenesis of autoimmune
primary cicatricial alopecia (HARRIES et al 2008). The concept that CD200-CD200R
interactions are important for protecting the HF epithelium from immune-mediated
attack and eHFSC damage comes from the CD200 knock-out mouse model
(MORRIS et al. 2004). When skin was grafted from CD200-/- mice onto gender-
matched wild-type hosts (CD200+/+) inflammation, hair loss and follicular scarring in
the grafted skin was caused. Of note was that the grafted skin survived long-term
and the epidermis remained normal throughout. Histologically, a peri- and intra-
follicular mononuclear inflammatory cell infiltrate was seen with intra-follicular
oedema and increased levels of apoptosis. Inflammation eventually resolved after all
hairs were lost and replaced with tracks of scar tissue. This supports that CD200 is a
key component in protecting eHFSC from immune attack and also plays a central
role in maintaining bulge IP.
2.10.11 Mast cells
Mast cells arise from pluripotential stem cells, reside and mature in tissues including
the skin and mucosal membranes, particularly at the interface of the host and its
environment, but also next to nerves, blood vessels and glandular structures
(BOTCHKAREV et al. 1997b; WEBER et al. 2003; BROWN et al. 2007). Mast cells
are characterized by their content of cytoplasmic granules, which contain pro-
inflammatory mediators, that are released upon mast cell activation (i.e. histamine,
leukotriens, prostaglandins, proteases, cytokines) (YOUNG 1997; MEKORI 2004;
THEOHARIDES and KALOGEROMITROS 2006; CHRISTY and BROWN 2007;
METZ et al. 2007).

LITERATURE
97
Mast cells have long been only recognized for their role in the genesis of allergic
inflammation [antigen-specific, IgE-mediated degranulation and secretory activities in
type 1 hypersensitivity reactions (BOS 1997)]. This changed since recently an
alternative activation pathway in addition to those elicited by FcεRI cross-linking was
found. Mast cells are equipped with many different receptors or their surface, through
these can be stimulated by complement components, neuropeptides, stress
hormones and TLR (bacteria and viruses), indicating a participation in innate and
acquired immune responses (GALLI et al. 2005; GILFILLAN and TKACZYK 2006).
Mast cell activation and expression on the cell surface can, besides other effects,
modulate T cell and DC function (METZ et al. 2008b; SAYED et al. 2008). For
example, MHC class II and T cell co-stimulatory molecule expression by mast cells
provides potential antigen-presenting cell capability. Mast cells also express most
cytokines (i.e. TNF-α) that control T cell differentiation pathways and directly regulate
T cell fate decision through processing and presenting antigen to T cells through
class I and class II MHC routes (MCLACHLAN et al. 2003; SUTO et al. 2006;
JAHANYAR et al. 2008; SAYED et al. 2008). For example, it has been demonstrated,
that mast cell-deficient mice do not get disease in models of rheumatoid arthritis or
bullous pemphigoid, indicating an essential role in these Ab-mediated, organ-specific
autoimmune diseases. In contrast, in Ab-mediated systemic lupus erythematosus
mast cell-deficient mice have normal or enhanced disease progression, since they
appear to be protected against multi-organ autoimmunity (CHRISTY and BROWN
2007) and it has been demonstrated that mast cells are required for normal
cutaneous wound healing (WELLER et al. 2006).

LITERATURE
98
Fig. 2.30 Stimulatory and immunosuppressive functions of mast cells. (CHRISTY and BROWN 2007)
Interestingly, recent reports indicate, that MCs may also be critical in suppressing
skin immune responses and in maintaining IP (Fig. 2.30). DEPINAY et al. (2006)
explained the reduced delayed-type hypersensitivity in the skin of MC knock-in mice,
that have been bitten by Anopheles mosquitoes, by MC degranulation and induction
of the immunosuppressive IL-10 (DEMEURE et al. 2005; DEPINAY et al. 2006). And
recently LU et al. (2006) suggested that MCs are crucial for optimal expression of

LITERATURE
99
peripheral tolerance to skin allografts. In addition, MCs can also dampen an immune
response directly through expression of certain cytokines such as IL-10 or TGF-β or
indirectly by facilitating T regulatory cell (Treg) activity (SAYED et al. 2008). In
summary, mast cells are now regarded to be ´good and bad guys`, just like every
other cell (MAURER et al. 2003b; MAURER and METZ 2005).
Mast cells show hair-cycle dependent functional and numerical changes in rats
(MORETTI et al. 1963) and mice (MAURER et al. 1994, 1997; PAUS et al. 1994c).
Furthermore, the ratio of degranulated mast cells increases substantially in the
transition periods (between anagen-catagen, catagen-telogen and telogen-anagen)
(PAUS et al. 1994c; BOS 1997). In human HFs, mast cells were shown to be
localized homogenously in the whole CTS (CHRISTOPH et al. 2000), although, a
detailed analysis remains to be done. There are reports from HF organ cultures that
the number and activation status of mast cells change in response to treatment (VAN
BEEK et al. 2008; MEYER et al. unpublished data). However, one has to keep in
mind, that mast cells in mice and man are very similar regarding their distribution,
responses to stimulating signals, and mediators, but that they also differ in several
respects. For example, human MCs do not contain serotonin, some proteases and
cytokines such as IL-3 and IL-4 can differ in their effects (MAURER and METZ 2005).
So there is need to clarify the role of MCs.
In the human nail matrix, dermal mast cells were demonstrated to be reduced in
vicinity of the nail matrix, compared to the PNF (ITO et al. 2005c). This low number of
periunguinal mast cells is in contrast to high density of mast cells found in
mesenchyme surrounding the human hair matrix (CHRISTOPH et al. 2000). Given
that mast cells are important cellular elements of innate immunity (MAURER et al.
2003b), this low mast cell number may compromise the defense capacity of this
region of nail mesenchyme. In contrast, in the more distally located, terminally
differentiated nail epithelium, production of antimicrobial peptides was found
(DORSCHNER et al. 2004), which may compensate and supply the innate immune
defense.

LITERATURE
100
2.10.12 ICAM-1
Intercellular adhesion molecules belong to the immunoglobulin superfamily. They are
cell-surface ligands for the leukocyte integrins and are crucial in the binding of
lymphocytes and other leukocytes to certain cells, including APCs and endothelial
cells. ICAM-1 (CD54) is the most prominent ligand for the integrin CD11a:CD18 or
LFA-1. It is rapidly inducible on endothelial cells by infection, and plays a major role
in local inflammatory responses (CAMELI et al. 1994; BOS 1997; GOLDSBY et al.
2003; JANEWAY et al. 2005).
Many studies have demonstrated ICAM-1 effects in normal inflammation (facilitation
of lymphocytes to migrate to the location of inflammation). ICAM-1 is considered to
be one of the components in uninflamed tissue, which endangers the status of IP. In
mice, the peri- and infrainfundibular ORS constitutively expresses ICAM-1 and thus
attract LFA-1 expressing immunocytes (e.g. macrophages) (MUELLER-ROEVER et
al. 2000). This is not surprising because the ORS and the bulge region are the
preferred sites for the occurrence of dense perifollicular inflammatory cell infiltrates
under stress (ARCK et al. 2001, 2003). In addition, it has been shown that in normal,
uninflamed mouse skin, a very small, but notable percentage of HFs are permanently
deleted by an inflammatory cell attack on the ICAM-1+ bulge region of anagen and
catagen pelage HFs, without any macroscopic evidence for this striking phenomenon
of ´programmed organ deletion (POD)` (EICHMUELLER et al. 1998). ICAM-1 IR in
normal human skin was found on most CTS and dermal fibroblasts and immune
cells, but the strongest ICAM-1 IR was seen on perifollicular inflammatory cell
clusters (PICC) (JAWORSKY et al. 1992). In addition, ICAM-1 expression was found
on HF keratinocytes in the close vicinity of perifollicular inflammatory cell clusters
(CHRISTOPH et al. 2000).
Whether such physiological POD of isolated (or possibly malfunctioning) HFs also
occurs in human skin, and whether at least some forms of cicatricial alopecias only
represent an excessive form of POD (PAUS et al. 1999) remains to be investigated.

LITERATURE
101
Furthermore, it was reported that human nail epithelium does not express ICAM-1
and that nail matrix cells express a low level of ICAM-1 in vitro (PICARDO et al.
1994). But in situ studies in the infantile human nail apparatus did not demonstrate
ICAM-1 IR in any epithelial compartment; only the expected IR in blood vessels was
found (ITO et al. 2005c).
2.10.13 β-defensin 2
β-defensins (also designated BD, and HBD in human) belong to the group of the
cationic antimicrobial peptides (CAPs or AMPs), that represent a highly conserved
element of innate immune defense from the evolutionary point of view, because they
are also present in plants, insects and lower vertebrates. The substances are
involved in the protection of epithelial surfaces, such as the skin or airway surfaces,
against microbial colonization (BOMAN 1995; NISSEN-MEYER and NES 1997;
HARDER et al. 2001; GLAESER et al. 2005; MOOKHERJEE and HANCOCK 2007;
ABTIN et al. 2008). Since the 1980s many peptides with antimicrobial activity were
also found and investigated in humans (HARDER et al. 1997, 2001; BRAFF et al.
2005a, 2005b; GLAESER et al. 2005; ABTIN et al. 2008), like ß defensins,
cathelicidin (LL-37) (LARRICK et al. 1995; FROHM et al. 1997) and the S100 protein
psoriasin.
In general, CAPs or AMPs not only act as an antibiotic, but also in the context of
wound healing (CARRETERO et al. 2008; GALLO 2008), stimulation of the migration
and proliferation of epidermal keratinocytes (NIYONSABA et al. 2007) angiogenesis
(KOCZULLA et al. 2003) and, furthermore, of modulating the immune response [e.g.
influencing the migration of macrophages by modulating their chemokine and
chemokine receptor expression and increasing the production of various cytokines
such as IL-6, IL-8, and GMCSF (NIYONSABA et al. 2005; GALLO 2008)]. For
cathelicidin (LL-37) it was even shown to affect the antimicrobial activity of mast cells
(DI NARDO et al. 2003).

LITERATURE
102
β-defensins are small cationic peptides with broad-spectrum antimicrobial activity.
Human β-defensin 2 is locally regulated by inflammation and is the first member of
the β-defensin family that is locally inducible by inflammation. The murine homolog of
human β-defensin 2, which is called β-defensin 3, is present in the respiratory system
and in low levels in the epithelial cells of the intestine and lung (BALS et al. 1999).
The unique murine β-defensin 2 (Defβ2) is not expressed in airways of untreated
mice (MORRISON et al. 1999), but is upregulated in the airways by
lipopolysaccharide and may contribute to host defense at the mucosal surface of the
airways. Despite the fact that β-defensins are looked upon as classic antimicrobial
peptides, it has recently been demonstrated that they have also immunomodulatory
activity and link between innate and adaptive immunity, being chemotactic for CD4+
T cells and immature dendritic cells (YANG et al. 1999).
Although the HF ostium of skin and the nail hyponychium in the nail apparatus
represent a major potential port of microbial entry into the skin, clinical signs of
bacterial infection are exceptionally rare. This suggests the presence of a highly
effective AMP defense system. The ORS and sebaceous gland of human
pilosebaceous units express prominent IR for human β defensin-1 and 2 (HBD1 and
2), which appear to be up-regulated in acne vulgaris lesions (CHRONNELL et al.
2001). Toll-like receptors (TLRs) -2, -4 and -5 are expressed in murine pelage HFs
and their activation by microbial stimuli induced the production of β defensin-2
(SELLERI et al. 2007). In addition, it was reported, that human, porcine, and murine
nails contain AMPs, and that the human cathelicidin LL-37 can destroy Candida
albicans (DORSCHNER et al. 2004).

LITERATURE
103
2.11 K(D)PT – a candidate as hair growth modulator and IP restorer
in anagen hair bulbs?
α-Melanocyte-stimulating hormone (α-MSH) is a cleavage product of the
neuropeptide pro-hormone pro-opiomelanocortin (POMC), which plays an important
role, e.g., in immune and pigmentary homeostasis (LIPTON and CATANIA 1997;
LUGER et al. 2000; BOEHM et al. 2006; WOOD et al. 2006). Earlier studies have
demonstrated that the POMC-derived peptides α-MSH and ACTH are principal
inducers of human epidermal pigmentation via their action at the MC-1R (LIN and
FISHER 2007; ROUSSEAU et al. 2007). α-MSH also promotes human HF
pigmentation by up-regulation of intrafollicular melanogenesis, melanocyte dendricity,
and melanocyte proliferation (KAUSER et al. 2005). In addition, numerous studies
over the last few years have provided plenty of evidence that α-MSH is a well
tolerated immuno modulator with cytoprotective and antiinflammatory effects
(LUGER et al. 1999, 2000, 2003; LUGER and BRZOSKA 2007). α-MSH is supposed
to be a mediator of tolerance induction in the brain (TAYLOR and KITAICHI 2007)
and eye (NIEDERKORN 2006), affecting several pathways with the capacity to re-
establish IP in anagen human hair bulbs (ITO et al. 2004). Such a collapse is
hypothesized as the reason for the development of the autoimmune disease alopecia
areata (AA).
Synthetic, α-MSH related tripeptides can imitate these pigmentary and
immunomodulatory effects, and promise minimal side effects upon clinical use, since
they are closely related to a well-tolerated endogenous neuropeptide (HADLEY and
DORR 2006). Most of the anti-inflammatory activities of α-MSH can be exerted by its
C-terminal tripeptide KPV. In this study, K(D)PT, a derivative of KPV, is of particular
interest. In K(D)PT, the hydrophobic amino acid valine of KPV is substituted by the
more polar amino acid threonine. K(D)PT also corresponds to amino acid 193-195 of
IL-1β, one of the major pro-inflammatory cytokines which also inhibits human HF
growth in vitro (PHILPOTT et al. 1996; HOFFMANN et al. 1997).

LITERATURE
104
Recent studies indicated that K(D)PT itself exerts anti-inflammatory effects in human
sebocytes (MASTROFRANCESCO et al. 2007) (suppression of IL-1β- and LPS
induced NFκB activation, IL-6 and IL-8 expression) but so far, it is unknown which
effect K(D)PT has on HF biology. On the other hand and in contrast to α-MSH, C-
and N-terminal fragments of α-MSH were shown to have no melanotropic effect in
frog and lizard skin bioassays (HASKELL-LUEVANO et al. 1996), so that it was
expected that K(D)PT has no melanogenic effect. Therefore we used our organ-
cultured human anagen HF model, comparing normal and pro-inflammatory
conditions.
The physiochemical properties, expected low costs of production of K(D)PT, its
combined anti-inflammatory and anti-microbial effects and the small molecular size,
would make this substance suitable for the future local treatment of immune-
mediated inflammatory skin disease, and as an IP restorer or as a hair growth
modulator (BRZOSKA et al. 2008).
Fig. 2.31 Biosynthesis of POMC peptides an natural melanocortins. Posttranslational processing of POMC by enzymes at specific cleavage sites, yields peptide hormones such as ACTH, α-MSH, γ-MSH and β-endorphin peptides. (B) Peptide sequences of the natural melanocortins, the central melanotropic pharmacophor of α-MSH and the α-MSH-related C-terminal tripeptides. Structural homologies are labeled in red (BRZOSKA 2008).

LITERATURE
105
2.12 Questions addressed in this study
1. Does the human HF bulge region belong to a site of relative immune privilege?
2. Do any differences exist between the IP in the anagen hair bulb and the bulge
region?
3. Which effects does the α-MSH related tripeptide K(D)PT have on organ cultured
HF biology?
4. Can K(D)PT influence the constitutive and IFN-γ induced MHC class-I and MHC
class II expression in isolated organ-cultured HFs and restore IP?
5. Which effects has K(D)PT on mast cells and degranulation?
6. Does K(D)PT effect the human HF cycle or does it have effects on hair matrix
proliferation and –apoptosis?
7. Is the murine vibrissal follicle a site of relative immune privilege?
8. Is the murine proximal nail matrix a site of relative immune privilege?
2.13 Experimental design
For this purpose, human uninflamed scalp skin from a total of 16 different patients
was randomly collected during routine face-lift surgery, and, additionally, murine
snout skin and nail apparatus from a total of 18 control mice was used. Both routine
and increased-sensitivity immunohistochemical staining techniques were employed,
and the corresponding IR patterns were evaluated by quantitative
immunohistochemistry. For functional evidence, we performed full thickness human

LITERATURE
106
scalp skin organ cultures to investigate whether IFN-γ, a key inducer of IP collapse in
hair bulbs, has a similar effect on the putative bulge IP.
In addition, we investigated the effects of the new synthetic α-MSH related tripeptide
K(D)PT onto HF IP and biology in our organ-culture HFs model under pro-
inflammatory conditions, complemented by immunohistochemistry.
For staining controls, internal positive controls of the reproduction of published
follicular IR patterns were chosen, if available. Otherwise appropriate positive
controls (e.g. lymphnodes, spleen, brain) were used. Only specific IR patterns that
were well reproducible between at least (3-)5 different individuals were
photodocumented and are reported here.
MATERIAL AND METHODS
107
3 Materials and methods
3.1 Tissue collection
3.1.1 Human tissue collection
Fig. 3.1 Human scalp skin specimen
Fronto-temporal and occipital human skin scalp biopsies were obtained from 16
females undergoing routine face-lift surgery after informed consent. The donors were
selected randomely (aged 45-67 years, mean age 52.5 years) and all experiments
were performed according to the Helsinki guidelines.
Scalp specimens were placed immediately after surgery into William’s E serum-free
medium supplemented with 2.5 fold concentrated penicillin (250 IU/ml), streptomycin

MATERIAL AND METHODS
108
(250mg/ml) and fungazone (12.5mg/ml amphotericin B), and transported overnight to
the labaratory at 4°C (all cell culture reagents were obtained from Gibco BRL,
Paisley, Scotland.
The skin samples were cut into thin strips (approximately 0.5 cm x 1 cm) (Fig. 3.1)
with a scalpel, embedded in Cryomatrix Shandon (Pittburg, PA, USA), snap-frozen in
liquid nitrogen, and stored at –80°C until use.
For immunohistochemistry, 5-7 µm thick cryosections were processed. The common
way of processing cutaneous tissue is by using vertically oriented sections of the
skin, with the epidermis at one pole and the hypodermis at the other. One difficulty is
that vertically orientated sections of the skin very rarely match the slanting of the
HFs, because they do not grow uniformely in a 90° angle to the epidermis. As a
consequence, complete longitudinal sections of the HF from the bulb to the epidermis
can be rarely produced and the HFs almost always appear incomplete. For every
analyzed antigen, scalp skin cryosections of at least 3-5 different individuals were
examined (3-10 cryosections per patient).
3.1.2 Murine tissue collection
Mouse tissue samples were gained from 18 six to 14 weeks old female C57BL/6 and
Balb/c mice of control groups from different experiments, that were housed in
community cages under standardized conditions (12 hours light/dark-cycle, water
and mouse chow ad libitum) in the animal facility of the University Hospital
Schleswig-Holstein. The mice were kindly provided from a cooperation with Prof. Dr.
D. Zillikens from the Department of Dermatology, Venerology and Allergology,
University of Lübeck. Samples that had been fixated in Bouin’s solution were kindly
collected by Prof. Dr. W. Meyer (Institute of Anatomy, University of Veterinary
Medicine Hannover) in cooperation with the Institute of Pharmacology, of the same

MATERIAL AND METHODS
109
university. The mice were anaesthesized and euthanized by cervical dislocation.
Immediately after death, samples were taken as described below.
For dissecting the nail samples, a nail was gripped with a forceps and cut with a
scissors proximal to the joint of the first and second phalanx. Every single nail of the
fore- and hindlimb was taken. One or two specimens each were placed horizontally
into a little plastic tablet form, embedded in Cryomatrix Shandon (Pittburg, PA, USA),
and snap frozen in liquid nitrogen. This embedding technique allows sagittal
cryosections of the whole nail apparatus.
In order to macrodissect the whisker pad, first, the hair in this location was trimed
with clippers or carefully removed with a scalpel. Second, the skin was cut with a
scalpel over the bridge of the snout posterior to the whisker pad. Then the incision
was extended medially along both sides of the mouth, paralleling the upper lips to the
nose. It was necessary to cut the deeper tendinous tissues in order to free the snout
skin and to prevent cutting of the large vibrissal follicles. Using a sterile forceps, the
whisker pad was pulled forward of the face, separating the tissue from the nasal
bridge. These tissue samples were placed in a vertical orientation (epidermis left,
dermis right) in a plastic tablet, embedded with Cryomatrix Shandon (Pittsburg, PA,
USA), snap frozen in liquid nitrogen and stored at –80°C until use. The samples were
cut into ~6-8 µm thick cryosections and then processed for immunohistochemistry (or
stored at –80°C until use). Vertically orientated sections of the whisker pad were
performed, showing the epidermis at one pole and the hypodermis at the other in
order to get complete longitudinal sections of the vibrissal follicle from the bulb to the
epidermis. The nail apparatus was processed for sagittal sections. Per antigen,
whisker pad and nail cryosections deriving from at least 5 different mice were
examined (3-8 cryosections per mouse).
Specimens, which were collected in the Institute of Pharmacology in Hannover, were
immersed immediately after removal in Bouin’s solution (BOECK 1989) [for
preparation 1500 ml picric acid, 500 ml formaldehyde (37-40%) and 100 ml glacial

MATERIAL AND METHODS
110
acetic acid were mixed) and fixed for at least 48 hours. The cut samples were then
placed in labeled plastic tablets and run through different fixing steps with
intermediate changes of the solution: 70 % ethanol (>1 day), 80% ethanol (> 1 day),
96 % ethanol (1 ½ h), isopropanol (1 1/2 h), xyelene I and II (each 1 ½ h), and
paraffine I (over night), paraffine II (2 h), paraffine III (1 ½ h). In the end, the samples
were positioned into a metallic form, effused with paraffin, and were then placed for
hardening in the fridge. After performing sections by using a microtome (5µm thin),
the slides were dried overnight at 37°C.
3.2 Human hair follicle isolation
Human anagen VI HFs were microdissected from frontotemporal and occipital human
scalp skin obtained from female donors (age range: 46-65 years) undergoing face-lift
surgery with informed consent and ethics committee as previously described
(PHILPOTT et al. 1990; MAGERL et al. 2002; MAGERL et al. 2004).
Before and during isolation, the skin samples were maintained in isolation medium
(William’s E medium, Biochrom, Cambridge UK) with 1% antibiotic/antimycotic
mixture (100x, Gibcom Germany, Karlsruhe) containing penicillin G, streptomycin
and amphotericin B to reduce the number of infection.
First, human scalp specimens were divided into 0.5 cm x 1 cm thin strips. Then the
skin strips were cut under sterile conditions at the dermo-hypodermal fat interface
using a scalpel blade (Fig. 3.2). The epidermis was removed and discarded. Under a
binocular dissecting microscope, the dermis was fixed with a blunt forceps to partially
extrude the upper portion of the HFs from the hypodermis. Simultaneously, the
CTS/ORS of the HF was gently gripped with watchmaker’s forceps and the intact HF
pulled out. This technique resulted in the isolation of intact HFs without any visible
damage (the HF was surrounded completely by the intact CTS). Otherwise, HFs had
not survived during the following culture period.

MATERIAL AND METHODS
111
Fig. 3.2 Isolation of human HFs Isolation of human HFs. A,B) Cut through the skin at the dermo-hypodermal fat interface. C,D) Parts of the dermis are removed. E) The skin is compressed, so that the HF steps out. F) The ORS of the follicle is gripped in the forceps and pulled out. G) Isolated pigmented HFs.
3.3 Human HF organ culture
Normally pigmented human anagen VI HFs were microdissected from temporal and
occipital human scalp skin obtained from female donors (age range: 46-65 years);
grey and white HFs were excluded from study. The isolated HFs were maintained
and cultured in serum-free William’s E medium (Biochrom AG, Berlin, Germany),
which was supplemented with 2 mmol/L L- glutamine (Invitrogen, Paisley, UK), 10
µg/ml insulin (Sigma-Aldrich, Taufkirchen, Germany), 10 ng/ml hydrocortisone
(Sigma-Aldrich), and 1% antibiotic/antimycotic mixture (100x, Gibco, Germany,
A
C
E
D
B
F G

MATERIAL AND METHODS
112
Karlsruhe). HFs were placed in a threesome in a 24-well plate with 500 µl complete
medium (Figs. 3.3, 3.4) and incubated free-floating in the wells in an atmosphere of
5% CO2 and 95% air at 37°C. On day 1, medium was changed and treatment began.
Fig. 3.3 Hair follicles in a 24-well plate
Fig. 3.4 Isolated hair follicles

MATERIAL AND METHODS
113
Six independent HF organ cultures with two different set-ups were used (three
HFs/well, three test groups and three controls) and compared.
In the first assay (Restoration assay) (Fig. 3.5), HFs were pre-treated with 75IU/ml
IFN-γ (Peprotech, Rocky Hill, NJ) for 4 days, followed by addition of 10-8, 10-9 or 10-
10M K(D)PT on day 3 (Assay 1). Control HFs were cultured for 4 days with K(D)PT
(10-8 M), IFN-γ (75 IU/ml), or with the vehicle (distilled water).
Alternatively (Assay 2, protection assay) (Fig. 3.5), HFs were first treated with
K(D)PT (10-8, 10-9 or 10-10M) for 48 hours (control groups were cultured in complete
medium only), followed by addition of 75 IU/ml IFN-γ (treatment for 4 days). Control
HFs were exclusively treated with K(D)PT, IFN-γ, or in the vehicle from day 3 on. We
chose low-dose 75IU/ml IFN-γ on the basis of our previous studies (ITO et al. 2004;
ITO et al. 2005b) in which we had demonstrated that this low dose of IFN-γ sufficed
to induce immune privilege collapse in the human anagen hair bulb (i.e. induction of
ectopic MHC class I immunoreactivity) without inducing premature catagen
development (the latter had to be avoided since premature catagen-induction shuts
off HF pigmentation) (SLOMINSKI et al. 1994). Every second day, hair shaft
elongation was measured using an inverted microscope and medium and substances
were renewed. 5 (assay 1) or 7 days (assay 2) after microdissection, HFs were
frozen in liquid nitrogen, stored at -80°C and processed for cryosectioning.
Fig. 3.5 Experimental design of the HF organ culture

MATERIAL AND METHODS
114
3.4 Full thickness human scalp skin organ culture
Two independent full-thickness organ cultures with adult human scalp skin were
performed. Human scalp skin was transferred and washed in William’s E medium
(Biochrom, Cambridge, UK) supplemented with 100 IU/ml penicillin G (Sigma, St
Louis, MO, USA), 10 lg/ml streptomycin (Sigma), 0.25 lg/ml amphotericin B (Gibco,
Karlsruhe, Germany) for the preparation. The outgrowing hair shafts were shaved
down to the level of the epidermis. Then 3-4 mm biopsies of intact scalp skin were
punched out parallel to the direction of hair growth, using an Acu-puncher (STIEFEL,
Offenbach am Main, Germany).
Human scalp tissue pieces were carefully placed randomised into William’s E
medium, which was supplemented with 100 IU/ml penicillin/10 lg/ml streptomycin, 10
lg/ml of insulin (Sigma), 10 ng/ml of hydrocortisone (Sigma) and 2 mmol/l of l-
glutamine (Invitrogen, Paisley, UK). Each treatment group consisted of three to four
punch biopsies and was placed in a 6-well-multi well plate with 3 ml complete
medium (Fig. 3.6).
Fig. 3.6 Full thickness human scalp skin punch biopsies in 6-well multi well plate

MATERIAL AND METHODS
115
The skin fragments were left to float freely in the medium, with the epidermis up at
the air/liquid interface and the dermis/hypodermis down. The cultures were
maintained at 37°C in a gassed incubator with 95% air and 5% CO2. After the first
over-night incubation, medium was changed and treatment began.
Control punch biopsies were treated only with PBS; the other five groups were
treated with five different IFN-γ (Peprotech, Rocky Hill, NJ) concentrations (100
IU/ml, 500 IU/ml, 1000 IU/ml). The treatment was always combined with a medium
change on day 1 and 3. After 5 days of cultivation, the specimens were embedded,
snap-frozen in liquid nitrogen and stored at-80°C until use.
3.5 Histological stainings
3.5.1 Hematoxylin-Eosin staining
The hematoxylin-eosin staining is a dichromatic staining, based on the basic dye
hematoxylin and the ethanol-based acidic dye eosin (BOECK 1989).
Cryosections (stored at -80°C) were fixed in acetone for 10 min at -20°C followed by
air drying for 30 min at room temperature and two times washing in Tris-buffered
saline (TBS) and 10 min in distilled water.
For the preparation of TBS buffer, 6.1 g Tris-base and 8.8 g sodium chloride (NaCl)
were diluted in 800 ml distilled water. Then the pH was adjusted with 1N hydrochloric
acid (HCL) to 7.6, and the solution was dropped up to 1000 ml with distilled water.
The sections were stained in Mayers hemalum (Merck, #1.04302) for 10 min and
rinsed under tap water approximately 15 min. The counterstaining was performed
with 0.1% eosin (Sigma, #E4382) for approximately 1 min. The final differentiation
was as follows via an ascending ethanol series: one time in 70%, two times in 96%,
two times in 100% ethanol and two times in xylene, all steps with carefully dipping 10
to 15 times. The slides were mounted with synthetic resin (Eukitt, O.Kindler &Co,
Freiburg, Germany).

MATERIAL AND METHODS
116
3.5.2 Trichromatic staining
This method was used for slides fixed in Bouin’s solution; with cryosections the
staining results were not successful.
The method was performed according to Masson-Goldner (BOECK 1989) and used
mainly as general map for the detection of connective tissue (collagen fibres, green),
that can well be distinguished from, i.e., muscles (red). For deparaffinization, the
slides were treated as follows: two times washing in xylene for 10 min each, one time
washing for 2 min each in isopropanol, 96 % ethanol, 80% ethanol, 70 % ethanol,
and clearing in distilled water.
Then the sections were stained in hemalum according to Delafield for 8 min, dipped
carefully in 0.1 % HCl in distilled water and rinsed under tap water approximately 15
min. For the following staining solution, 0.2 g Ponceau de xyline and 0.1 g acid
fuchsin were solved in 300 ml distilled water and 0.6 ml glacial acetic acid was
added. The slides were stained for 5 min in acid fuchsin-Ponceau, followed by 5 min
washing in 0.1 % acetic acid, 10 min washing in a solution of 10 g phosphortungstic
acid and 5 g orange G in 250 ml distilled water and a second time of 5 min. washing
in 0.1 % acetic acid. The counterstaining was performed using light green (0.5 g light
green + 250 ml distilled water + 0.5 ml acetic acid) for 5 min followed again by a 5
min washing step in 0.1 % acetic acid, and dehydration via the ascending ethanol
series as follows: three min washing each in 80 % ethanol, in 96 % ethanol and 96%
isopropanol, and two times clearing in xylene substitute (Merck, Darmstadt,
Germany) for 5 min each. The slides were mounted with synthetic resin (Eukitt, O.
Kindler GmbH Co., Freiburg, Germany).
3.5.3 Toluidine blue staining
This staining method is simple and widely used as a technique for the detection of
mast cells. Mast cells are often present in connective tissue, and their cytoplasm

MATERIAL AND METHODS
117
contains granules composed of heparin and histamine. Toluidine blue should colour
mast cells violet or red purple (metachromatic staining) and the background blue
(orthochromatic staining).
Cryosections (6-8 µm thick) which had been stored at -80°C were air dried for 10
min. Then the slides were washed for 2 min in distilled water at room temperature
followed by staining for 1 min in toluidine blue according to RICHARDSON et al.
(1960) (working solution: 1g toluidine blue 0,1g di-sodiumtetraborate-10-hydrate,
0,2g paraformaldehyde in 100ml distilled water). The staining step was stopped by
three times washing in distilled water, and the slides were then differentiated and
dehydrated via an ascending ethanol series as follows: three times dipping in 70%
ethanol, three times dipping in 96% ethanol, 3 min washing in 96% ethanol, two
times washing 3 min in 100% ethanol and two times clearing in xylene substitute
(Merck, Darmstadt, Germany) for 10 min each. The slides were coversliped with a
synthetic resin mounting medium (Eukitt, O. Kindler GmbH Co., Freiburg, Germany).
3.5.4 Leder`s esterase staining
Leder`s esterase staining (LEDER 1964) uses the enzyme chloroacetate esterase as
a marker for cells of the neutrophil series, monocytes, and mast cells in tissues, and
produces a strong red to deep pink cytoplasmic staining of mast cells, whereas the
nuclei appear in green (light green) or blue (hematoxylin). We performed this method
mainly to detect mast cells in the CTS of isolated HFs.
Cryosections of cultured HFs which have been stored at -80°C were air dried for 10
min at room temperature. The slides were fixed in 1% paraformaldehyde (in PBS) for
10 min and then washed three times in distilled water for 5 min each.
For the preparation of the incubation medium several different working buffers were
prepared: The Sörensen working buffer consisted of 32.8 ml of the Sörensen stock
solution A (2.73 g Na2HPO4 in 250 ml distilled water) and 7.2 ml Sörensen stock
solution B (2.27 g KH2PO4 in 250 ml distilled water). A pararosanilin solution was

MATERIAL AND METHODS
118
made of 0.5 g pararosanilin powder (Merck, Germany), 20 ml of distilled water and
2,5 ml concentrated hydrochloric acid, which was heated carefully, filtered and stored
in the refridgerator. Immediately before staining, the nitrosylated pararosanilin was
prepared by mixing 150 µl of the pararosanilin solution with 150 µl 4% sodium nitirite
in distilled water. In the end the incubating medium was prepared by mixing and
filtering of 10 mg naphtol-ASD chloroacetate (Merck, Germany), 1 ml N,N-
dimethylformamide, 35 ml of Sörensen working buffer and 200 µl of the nitrosylated
pararosanilin The slides were stained in this incubating medium for 40 min, followed
by washing in running tap water for 5 min. The counterstaining was performed either
using light green or hematoxylin for approximately 1 to 2 min, followed by another 5
min wash in running tap water. Afterwards the slides were air dried and dehydrated
via an ascending ethanol series as follows: three times dipping in 70% ethanol, three
times dipping in 96% ethanol, 3 min washing in 96% ethanol, two times washing 3
min in 100% ethanol and two times clearing in xylene substitute (Merck, Darmstadt,
Germany) for 10 min each. The slides were coversliped with synthetic resin mounting
medium (Eukitt, O. Kindler GmbH Co., Freiburg, Germany).
3.5.5 Ki-67 / TUNEL
To evaluate apoptotic cells in co-localization with the proliferation marker Ki-67, a Ki-
67 / TUNEL (terminal dUTP nickendlabeling) double-staining method was used.
The Ki-67 antigen is a nuclear protein, which is identified by its reactivity with
monoclonal antibody from Ki-67 clone (GERDES et al. 1984); two isoforms of 345
and 395 kDa have been identified (GERDES et al. 1991). The Ki-67 antigen is
preferentially expressed during all active phases of the cell cycle (G1, S, G2 and M-
phase), but it is absent in resting cells (G0-phase) (GERDES et al. 1984). During
interphase, the antigen can be exclusively detected within the nucleus, whereas in
mitosis most of the protein is relocated to the surface of the chromosomes. The
antigen is rapidly degraded as the cell enters the non-proliferative state (SCHOLZEN

MATERIAL AND METHODS
119
and GERDES 2000), and there appears to be no expression of Ki-67 during DNA
repair process (KEY et al. 1994).
Apoptosis is a form of cell death that eliminates compromised or superfluous cells.
During this process, the DNA is cut into fragments via endonuclease activity. The
DNA strand breaks are detected by enzymatically labeling the free 3’-OH termini with
modified nucleotides. These nucleotides are enzymatically added to the DNA by the
terminal deoxynucleotidyltransferase (TdT), catalysing a template-independent
addition of nucleotide triphosphates to the 3’-OH ends of double-stranded or single-
stranded DNA. DNA fragments which have been labelled with the
digoxigenin-nucleotide are then allowed to bind an anti-digoxigenin antibody that is
conjugated to FITC.
Cryostat sections of treated (with 12G10 (+), TS2/16 (+) and mAb13 (-) β1 integrin
antibodies) and untreated cultured HFs were fixed in paraformaldehyde (10 min,
room temperature) and ethanol-acetic acid (2:1; 5 min, -20°C), using washing steps
with PBS in-between (three times for 5 min). Then the slides were labelled with a
digoxigenin-deoxyUTP (ApopTag Fluorescein In Situ Apoptosis detection kit;
Intergen, Purchase, USA) in the presence of TdT (60 min, 37°C), followed by
incubation with a mouse anti-Ki-67 antiserum (DAKO, Glostrup, Denmark) overnight
at 4°C.
Using washing steps with PBS, TUNEL-positive cells were visualized by an
anti-digoxigenin FITC-conjugated antibody (ApopTag kit) (30 min, room
temperature), whereas Ki-67 was detected by a rhodamine-labelled goat anti-mouse
antibody (Jackson ImmunoResearch, West Grove, USA) (45 min, room temperature).
Finally the cryosections were counterstained with DAPI (Boehringer Mannheim,
Germany) for 1 min and mounted with Fluoromount-G (Southern Biotechnologies,
Birmingham, USA).
Negative controls were performed by omitting TdT and the Ki-67antibody.

MATERIAL AND METHODS
120
3.6 Immunohistochemistry
3.6.1 Primary antibodies
Tab. 3.1 Primary antibodies and secondary detection systems Anitbodies used for immunohistological stainings are listed and described in detail. ABC-AP: Avidin-biotin complex, alkaline phosphatase, ABC-HRP: Avidin-biotin complex, horseradish peroxidase, FITC: Fluorescein isothiocyanate, IF: Immunofluorescence, Ig: Immuno globulin, NA: Not applicable, Rh: Rhodamine, TSA: Tyramide signal amplification conjugated.
Anti-human antibodies Primary
antibody
Origin Clone Ig
class
Vendor Dilu-
tion
Secon-
dary
system
References
HLA-ABC mouse W6/32 IgG2akappa
DAKO, Glostrup, Denmark
1:50 1:50 1:1500
EnVision® IF-FITC/Rh TSA
ITO et al. (2004); ITO et al. (2005c)
HLA-DP/ DQ/ DR
mouse CR3/43 IgG1, kappa
DAKO, Glostrup, Denmark
1:50 1:50 1:1500
EnVision® IF-FITC/Rh TSA
ITO et al. (2004)
CD200 mouse MRC OX 104
IgG1 Serotec, Oxford, UK
1:400 TSA OHYAMA et al. (2006); TIEDE et al. (2007a)
β2-micro-globulin
mouse Tü99 IgM, kappa
BD Pharmingen, Heidelberg, Germany
1:100 1:100
EnVision® IF-FITC/Rh
HARRIST et al. (1983); CHRISTOPH et al. (2000); BLASCHITZ et al. (2001)
α-MSH rabbit - IgG Sigma-Aldrich, München, Germany
1:2000 TSA KONO et al. (2001)
TGF-β2 rabbit - - Santa Cruz, Heidelberg, Germany
1:4000 TSA DE KOZAK et al. (2007)
IDO mouse 10.1 - AbD Serotec, Oxford, UK
1:50 ABC-HRP SCHELER et al. (2007)
MIF mouse 12302 IgG1 R&D Systems, Minneapolis, USA
1:50 ABC-HRP ITO et al. (2007)
HLA-E mouse - - kindly produced and provided by Daniel Geraghty, Fred Hutchinson, Re-search Centre, Seattle, USA
1:100 IF-FITC -

MATERIAL AND METHODS
121
Tab. 3.2 Primary anti-mouse antibodies and secondary detection systems Anitbodies used for immunohistological stainings on mouse tissue are listed and described in detail. ABC-AP: Avidin-biotin complex, alkaline phosphatase, ABC-HRP: Avidin-biotin complex, horseradish peroxidase, FITC: Fluorescein isothiocyanate, IF: Immunofluorescence, Ig: Immuno globulin, NA: Not applicable, Rh: Rhodamine, TSA: Tyramide signal amplification conjugated.
Anti-mouse antibodies Primary
antibody
Origin Clone Ig
class
Vendor Dilution Secon-
dary
system
References
MHC I (H-2d, H-2b,p,q)
rat ER-HR52
IgG2a BMA Biomedical,
Augst, Switzerland
1:50 1:150
ABC-HRP RUECKERT et al. (1998)
MHC II rat ER-TR3 IgG2b BMA Biomedical,
Augst, Switzerland
1:200 1.200
ABC-HRP IF-
FITC/Rh
PAUS et al. (1998); EICHMUELLER et
al (1998)
CD4 (L3T4)
rat RM4-5 IgG2a BD Pharmingen, Heidelberg, Germany
1:100 ABC-HRP PAUS et al. (1998); EICHMUELLER et
al (1998)
CD8a (Ly-2)
rat 53-6.7 IgG2a kappa
BD Pharmingen, Heidelberg, Germany
1:10 ABC-HRP PAUS et al. (1998); EICHMUELLER et
al (1998) CD11b (Ly-40)
rat M1/70 IgG2b BMA Biomedicals,
Augst, Switzerland
1:8000 ABC-HRP PAUS et al. (1998)
ICAM-1 (CD54)
hamster 3E2 IgG BD Pharmingen, Heidelberg, Germany
1:50 ABC-HRP EICHMUELLER et al (1998)
TGF-β1 rabbit - IgG Santa Cruz, Heidelberg, Germany
1:750 ABC-HRP CHUNN et al (2006)
β-defen-sin 2
goat - IgG Santa Cruz, Heidelberg, Germany
1:10 ABC-HRP -
3.6.2 Avidin Biotin Complex-Peroxidase
Cryosections (6-7 µm thick) which had been stored at -80°C were air dried for 10 min
and fixed in acetone at -20°C for 10 min. Then the slides were washed tree times for
5 min each in TBS buffer (for preparation see chapter 3.5.1) followed by a blocking
step against the endogenous peroxidase in 3 % H2O2 in TBS for 15 min at room
temperature. After three times of 5 min washing the samples, sections were circled

MATERIAL AND METHODS
122
with a fat marker and then preincubated with normal serum 10% (DAKO, Glostrup,
Denmark) in TBS for 20 min in a moist chamber. The normal serum was of the same
origin as the secondary antibody (i.e. goat, rabbit, hamster).
After preincubation the slides were not washed but the normal serum was only
dropped, and the primary antibody (see Tabs. 3.1, 3.2) was applicated with its
appropriate dilution in TBS together with 2 % of normal serum overnight at 4 °C.
After three times washing for 5 min each in TBS, the slides were stained either with
goat anti-mouse or goat anti-rabbit (1:200 in TBS, Jackson ImmunoResearch,
Cambridgeshire, UK), goat anti-rat (1:200 in TBS, Beckman Coulter, Marseille,
France) or rabbit anti-goat (1:200 in TBS, Santa Cruz, Heidelberg, Germany)
biotinylated secondary antibodies. After 45 min incubation at room temperature,
slides were washed three times for 5 min in TBS. A 30 min application of ABC-
peroxidase solution (avidin-biotin-complex, peroxidase conjugated, Vector
Laboratories, Burlingame, CA, USA) followed at room temperature. It is important to
note is, that the solution had to be prepared 30 min before usage.
The sections were labelled with the peroxidase-chromogen 3,3’-diaminobenzidine
(DAB) (Vector Laboratories, Burlingame, CA, USA) or 3-amino-9-ethylcarbazole
(AEC) (DAKO, Burlingame, CA, USA) for 2-10 min (see Tabs. 3.1, 3.2). Both
substrate solutions DAB and AEC were prepared according to their protocols
immediately before use as follows: For preparation, 2 drops of Buffer stock solution,
5 drops of DAB stock solution, i.e. 3 drops of the AEC stock solution and 2 drops of
the hydrogen peroxide solution, were added to 5 ml of distilled water, and mixed well
in between. If necessary or desired, the sections were counterstained with Mayer’s
hemalum (Merck, Darmstadt, Germany) using washing steps in between. Finally the
slides were mounted with DAKO Faramount (DAKO, Glostrup, Denmark).
As negative controls, the primary antibodies were omitted, performing the same
staining steps as described before.

MATERIAL AND METHODS
123
3.6.3 EnVision®-alkaline phosphatase
Cryosections (6-8 µm thick) which had been stored at -80°C were air dried for 10 min
and then fixed in acetone at -20°C for 10 min. After a three times washing step for 5
min each in TBS (as described before, see 3.5.1), the section samples were
preincubated with normal serum 10%, that was of the same origin as the secondary
antibody (i.e. goat, rabbit, hamster; DAKO, Glostrup, Denmark), in TBS for 20 min.
The slides were washed three times for 5 min in TBS and then preincubated with
10% normal goat serum (DAKO, Glostrup, Denmark) in TBS for 20 min. The
application of the primary antibodies (see Tabs. 3.1, 3.2) followed in their appropriate
dilution at 4 ˚C in TBS overnight.
A three times washing step in TBS for 5 min each was followed by the incubation
with EnVision® solution (alkaline phosphatase, anti rabbit / mouse, IgG-labelled
polymer, DAKO, Glostrup, Denmark) for 30 min and another three times washing
step. Afterwards Fast Red AP-chromogen (Sigma, Saint Louis, USA) was employed
for 3 to 10 min to visualize the immunosignals, and finally, sections were
counterstained with Mayer’s hemalum (Merck, Darmstadt, Germany) and mounted
with DAKO Faramount (DAKO, Glostrup, Denmark).
3.6.4 Immunofluorescence
To identify the IR in the bulge region of human HFs via immunofluorescence staining,
we used primary antibodies (see Tabs. 3.1, 3.2) and the secondary antibodies goat
anti-mouse, anti-rat or anti-rabbit IgG conjugated with fluorescein isothiocyanate
(FITC) or rhodamine (1:200 in TBS, Jackson ImmunoResearch, Cambridgeshire,
UK).
For Immunofluorescence staining, cryosections (6-8 µm thick) which had been stored
at -80°C were first air dried for 10 min and then fixed in acetone at -20°C for 10 min.
A three times washing step in TBS (produced as described before, see 4.5) for 5 min
each followed, and then the section samples were preincubated for 20 min with

MATERIAL AND METHODS
124
normal serum 10% (DAKO, Glostrup, Denmark) in TBS at room temperature. Then
primary antibodies (see Tabs. 3.1, 3.2) in their appropriate dilution in TBS were
applicated and the sections were incubated overnight at 4°C. After washing three
times for 5 min in TBS, a 45 min incubation at room temperature with the appropriate
secondary antibody in TBS was arranged. Then the slides were washed three times
for 5 min in TBS again, counterstained with DAPI (4’,6-diamidin-2’-phenylindol-
dihydrochlorid, Boehringer Mannheim, Germany) for 1 min and mounted with
Fluoromount-G (Southern Biotechnologies, Birmingham, USA).
The fluorescent dye DAPI binds selectively to DNA (deoxyribonucleic acid) and forms
strongly fluorescent DNA-DAPI complexes with high specificity. On adding DAPI to
the tissue it is rapidly taken up into cellular DNA yielding highly fluorescent nuclei and
no detectable cytoplasmatic fluorescence.
3.6.5 Tyramide signal amplification (TSA)
After standard fixation for 10 min in acetone at -20°C, the cryosections (6-7µm thick)
were washed three times for 5 min using TNT buffer (0.1 mol/l Tris-HCl, pH 7.5;
containing 0.15 mol/l NaCl and 0.05 % Tween 20). Then endogenous horseradish
peroxidase was blocked by washing with 3% H2O2 in PBS (phosphate buffered
saline) for 15 min.
For the preparation of PBS, 1.8 g NaH2PO4 x H2O sodium dihydrophosphate
monohydrate and 8.0 g sodium chloride (NaCl) were diluted in 800 ml distilled water.
The pH was adjusted to 7.2 with 1N sodium hydroxide (NaOH). At the end the
solution had to be dropped up to 1000 ml with distilled water.
Preincubation was performed with the incubation of avidin and biotin for 15 min each,
with three times 5 min washing steps in TNT in between followed by another
preincubation step with 5% normal goat serum (DAKO, Glostrup, Denmark) in TNT
for 30 min. Afterwards the primary antibodies (see Tabs. 3.1. 3.2) diluted in TNT
were added and the sections were incubated overnight at 4˚C. Next day, the

MATERIAL AND METHODS
125
application of the appropriate biotinylated secondary antibody (goat anti-mouse
(1:200 in TNT, Jackson ImmunoResearch, Cambridgeshire, UK) followed with an
incubation for 45 min at room temperature. After a three times washing step,
streptavidin horseradish peroxidase (TSA kit; Perkin-Elmer, Boston, USA) was
administrated (1:100 in TNT) for 30 min at room temperature, and the reaction was
amplified by tetramethylrhodamine- or FITC-tyramide amplification reagent at room
temperature for 5 min (1:50 in amplification diluent provided with the TSA kit). The
cryosections were counterstained with DAPI (Boehringer Mannheim, Germany) for 1
min and mounted with Fluoromount-G (Southern Biotechnologies, Birmingham,
USA).
For all immunostaining assays, primary antibodies were omitted as negative control.
3.7 Histomorphometry
3.7.1 Assessment of hair cycle stages
In order to observe spontaneous or induced catagen development, immunostained
HF slides were used for HF cycle staging according to previously well-defined
morphological criteria (MUELLER-ROEVER et al. 2001; STENN and PAUS 2001),
and the percentage of HFs in anagen, early, mid and late catagen were determined.
3.7.2 Assessment of proliferating matrix keratinocytes
Cryosections of organ-cultured HFs treated with different concentrations of the test α-
MSH related tripeptide K(D)PT, with or without a combination with IFN-γ were then
stained for Ki-67 / TUNEL and investigated at 400x magnification. All Ki-67-positive
(red) and all TUNEL-positive cells (green) below Auber’s line (AUBER 1952) were
counted as well as the total number of keratinocytes, stained with DAPI (blue) (Fig.
3.7). The amount of Ki-67-positive cells, and, TUNEL-positive cells respectively
relating to DAPI-positive cells was shown in per cent.

MATERIAL AND METHODS
126
Fig. 3.7 Auber’s line marked in the human HF The Auber’s line runs through the widest part of the hair bulb at the level of the maximal diameter of the dermal papilla; Below this line every Ki67+ cell, TUNEL+ cell and DAPI+ cell is counted and the percentage for proliferation and apoptosis calculated; dotted white line: Auber’s line (AUBER 1952).
3.7.3 Assessment of immunostaining intensity
The immunostaining intensity for selected antigens examined was compared by
quantitative immunohistochemistry as previously described (ITO et al. 2004b, 2005b,
2005c), using NIH image software (National Institute of Health, Bethesda, Maryland).
In regard to the special tissue, different reference areas were defined and analyzed.
In general, the human scalp HF was divided anatomically into five reference
compartments: (1) epidermis, (2) distal ORS: middle of the isthmus, (3) bulge:
insertion point of the APM, (4) proximal ORS, (5) hair matrix, and analyzed deriving
from 3-5 different individuals.
The vibrissal follicle was divided according to the human HFs into five compartments
1) isthmus, (2) distal ORS: superior swelling of the ORS, (3) bulge: inferior swelling of

MATERIAL AND METHODS
127
the ORS, (4) proximal ORS, (5) hair matrix, and analyzed, deriving from 5 different
mice.
The nail apparatus was analyzed within two reference areas (1) the proximal nail
matrix and (2) the proximal nail fold as shown.
3.7.4 Assessment of MHC class I IR in isolated HFs
Immunostaining intensity was compared by quantitative immunohistochemistry as
previously described (ITO et al. 2004), using NIH image software (NIH, Bethesda,
Maryland). In the run-up to analyse, reference areas were defined according to the
objective of the study. The mean intensity was measured at five previously defined
reference areas in the hair matrix, proximal ORS and CTS/DP (Fig. 3.8) and the
average was calculated.
Fig. 3.8 Reference areas for the quantitative analysis of MHC class I IR CTS: Connective tissue sheath, DP: Dermal papillam IRS: Inner root sheath, MK: Matrix keratinocytes, ORS: Outer root sheath. Original magnification: 200x.

MATERIAL AND METHODS
128
3.7.5 Assessment of mast cells
By evaluating the toluidine blue and with Leder`s esterase method stained HFs, the
total number of mast cells was counted in the defined reference areas. In isolated
HFs, all mast cells located in the CTS were counted, whereas in the human and
murine tissue in situ mast cells were counted at a 400x magnification of the defined
compartments (see 3.7.3) and the mean value was calculated. In addition, non-
degranulated and degranulated mast cells were distinguished. Mast cells with more
then six granules outside of the cell membrane were regarded as ´degranulated`.
3.7.6 Assessment of MHC class II, CD4, CD54, CD11b, mast cells
and c-kit positive cells
Positively stained cells were counted by quantitative immmuno histomorphometry
following our previously described morphometrical techniques (PAUS et al. 1998;
CHRISTOPH et al. 2000; ITO et al. 2004).
Again, the human scalp HF was divided anatomically into five reference
compartments: (1) epidermis, (2) distal ORS: middle of the isthmus, (3) bulge:
insertion point of the APM, (4) proximal ORS, (5) hair matrix. In these compartments
HLA-DP/DQ/DR, CD54 positive cells and mast cells were counted at a 400x
magnification and analyzed on deriving from 3-5 different individuals.
In the same way, the vibrissal follicle was divided into (1) isthmus, (2) distal ORS:
superior swelling of the ORS, (3) bulge: inferior swelling of the ORS, (4) proximal
ORS, (5) hair matrix, and analyzed at 400x magnification for MHC class II, CD4,
CD11, CD54 positive cells and mast cells (N= 5 mice).
The murine nail was divided into the nail matrix and the PNF as shown anatomically,
and MHC class II, CD4, CD11b, CD54, and mast cells (toluidine blue staining) in
these compartments cells were counted at 400x magnification per macroscopic field
from one nail (N=5-7 individuals).

MATERIAL AND METHODS
129
3.7.7 Microscopical equipment
For fluorescence and light microscopy, a Keyence Biozero-8000 microscope
(Keyence Corporation, Biozero-8000, Higashi-Nakajima, Osaka, Japan) was used, in
combination with Nikon lenses (Japan).
3.7.8 Statistical analysis
For the evaluation of statistical significance, all data obtained from the experiments
were pooled and analyzed with the Mann-Whitney-U-Test for unpaired samples
(GraphPad Prism version 4.00 for Windows, GraphPad Software, San Diego,
California, USA). The results were expressed as standard error of the mean (mean
±SEM) and values of p<0.05 were regarded as significant.
The IR for MHC class I in human scalp skin in situ and in the full thickness punch
biopsies was analysed by quantitative immuno histomorphometry, using NIH image
software. The reference area for ‘distal ORS’ was on the level of the insertion of the
sebaceous duct, the ´bulge` at the insertion point of the arrector pili muscle and the
proximal ORS point was situated in the middle between the bulge and the hair bulb.
(N=5, *p<0.001, mean+/-SEM), p value was calculated with the Mann-Whitney-Test
for unpaired samples.
The mean immunostaining intensity for MHC class I in isolated HFs was measured
for four previously defined reference areas in the hair matrix of anagen hair bulbs
(Fig. 3.8) by NIH image, and the average staining intensity was calculated (N= 6-36
HFs from 6 patients/group).
The percentage of HFs in the different hair cycle stages treated with the K(D)PT was
calculated as compared to the control (N=8-12 follicles / group, six representative
experiments), p value was calculated with the Mann-Whitney-U-Test for unpaired
samples.

MATERIAL AND METHODS
130
Ki-67+ / TUNEL+ cells were counted below Auber’s line were counted as well as the
total number of keratinocytes, stained with DAPI. The amount of Ki-67+ cells, and,
TUNEL+ cells respectively relating to DAPI-positive cells was shown in per cent.
N=6-18 follicles / group, *p<0.05, mean ± SEM, p value was calculated with the
Mann-Whitney-U-Test for unpaired samples.

RESULTS
131
4 Results
4.1 Immune privilege and the human hair follicle bulge
4.1.1 Demonstration of MHC class Ia and β2-microglobulin
expression on CD200+ cells
In order to accurately define MHC class Ia protein expression in the bulge region, we
used the broad-specificity antibody HLA-A/B/C against all MHC class Ia antigens,
which are expressed on all nucleated cells, except in areas of IP (NIEDERKORN
2006). Compared to the distal ORS or the epidermis, immunohistochemistry clearly
showed that the ORS bulge region indeed expressed substantially lower or absent
MHC class Ia IR (Fig. 4.1 A,B). This was evident both by light microscopy (Fig. 4.1)
and Immunofluorescence (Fig. 4.4), complemented by quantitative analysis of MHC
class I IR (***p<0.001) (Fig. 4.3). In the distal ORS, especially in the infundibulum
and upper parts of the isthmus both basal and suprabasal staining was observed,
whereas IR in the bulge ORS was mainly limited to the basal layer of the ORS.
The proximal ORS, IRS and all matrix keratinocytes of the immunoprivileged anagen
hair bulb displayed extremely weak or absent MHC class Ia IR (Fig. 4.1 C).
Interestingly, compared to the bulge, the proximal ORS showed lower, although not
significant MHC class I IR (Fig. 4.3) whereas the level of HLA-A/B/C associated IR in
the anagen hair matrix compared to the bulge was significantly reduced (*p<0.05)
(Figs. 4.2, 4.3).

RESULTS
132
Fig. 4.1 IR is downregulated in the human bulge region for MHC class I (A-C) Immunohistological identification (EnVision®) of MHC class I expression. (A,B) HLA-A/B/C IR is prominently downregulated in the bulge region compared to the distal ORS and the epidermis. (C) The anagen hair matrix shows no MHC class I expression. APM: Arrector pili muscle, B: Bulge, CTS: Connective tissue sheath, E: Epidermis; HS: Hair shaft, MK: Matrix keratinocytes, ORS: Outer root sheath, SG: Sebaceous gland, Bars: (A): 200µm; (B,C): 100µm.
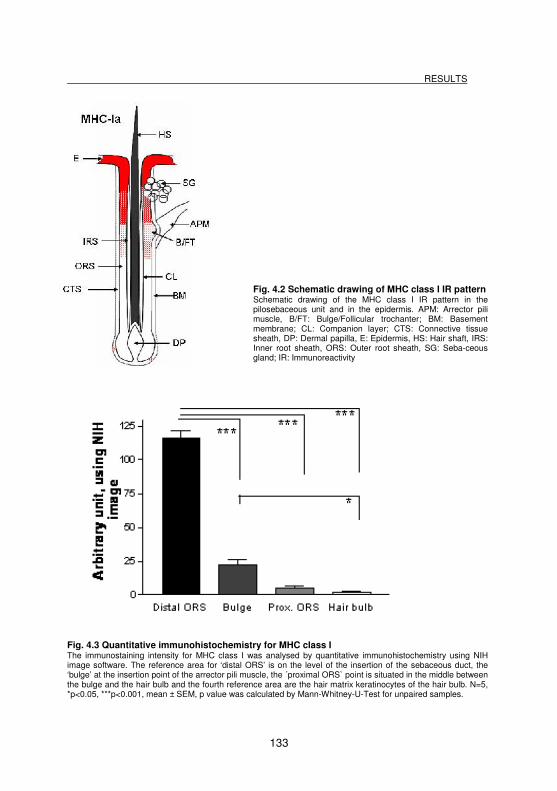
RESULTS
133
Fig. 4.2 Schematic drawing of MHC class I IR pattern Schematic drawing of the MHC class I IR pattern in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Seba-ceous gland; IR: Immunoreactivity
Fig. 4.3 Quantitative immunohistochemistry for MHC class I The immunostaining intensity for MHC class I was analysed by quantitative immunohistochemistry using NIH image software. The reference area for ‘distal ORS’ is on the level of the insertion of the sebaceous duct, the ‘bulge’ at the insertion point of the arrector pili muscle, the ´proximal ORS` point is situated in the middle between the bulge and the hair bulb and the fourth reference area are the hair matrix keratinocytes of the hair bulb. N=5, *p<0.05, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.

RESULTS
134
Since the immunoregulatory molecule CD200 belongs to the so-called bulge marker
(although it is not restricted to it), we performed a double TSA staining with MHC
class I molecule. We found prominent CD200 IR in the outermost layer of the ORS
between the insertion of the APM and the insertion of the SG duct (parts of the
isthmus and the bulge region) besides some staining in the DP including blood
vessels, the APM, the homogeneously stained companion layer and SW
mesenchyme and epithelium (data not shown). Most importantly, the intensity of
MHC class Ia-like IR was inversely correlated with that of CD200 (Fig. 4.4 A,B):
Again, MHC class I protein expression was only prominent in the epidermis, and in
the ORS of infundibulum and isthmus, but absent or extremely weak in the basal
bulge ORS (Figs. 4.4 B, 4.5).
Fig. 4.4 MHC class I IR is downregulated in CD200+ bulge ORS cells Double Immunofluorescence staining (TSA) shows the expression pattern of CD200 molecule (FITC, green signal) and MHC class I (Rhodamine, red signal) (counterstained with DAPI, blue). CD200 is upregulated in the bulge whereas MHC class I protein expression is downregulated compared to the distal ORS and epidermis. (A) High magnification of the bulge ORS displays very strong CD200 expression and almost none MHC class I expression. APM: Arrector pili muscle, HS: Hair shaft, ORS: Outer root sheath, SG: Sebaceous gland, Bars: (A): 100µm; (B): 20µm.

RESULTS
135
Fig. 4.5 Schematic drawing of CD200 IR pattern Schematic drawing of the CD200 IR pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.
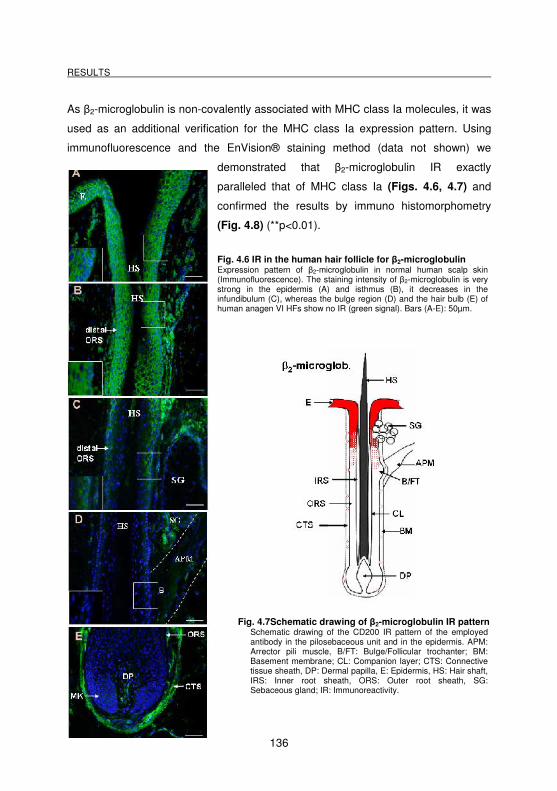
RESULTS
136
As β2-microglobulin is non-covalently associated with MHC class Ia molecules, it was
used as an additional verification for the MHC class Ia expression pattern. Using
immunofluorescence and the EnVision® staining method (data not shown) we
demonstrated that β2-microglobulin IR exactly
paralleled that of MHC class Ia (Figs. 4.6, 4.7) and
confirmed the results by immuno histomorphometry
(Fig. 4.8) (**p<0.01).
Fig. 4.6 IR in the human hair follicle for β2-microglobulin Expression pattern of β2-microglobulin in normal human scalp skin (Immunofluorescence). The staining intensity of β2-microglobulin is very strong in the epidermis (A) and isthmus (B), it decreases in the infundibulum (C), whereas the bulge region (D) and the hair bulb (E) of human anagen VI HFs show no IR (green signal). Bars (A-E): 50µm.
Fig. 4.7Schematic drawing of β2-microglobulin IR pattern
Schematic drawing of the CD200 IR pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.

RESULTS
137
Fig. 4.8 Quantitative immunohistochemistry for β2-microglobulin The immunostaining intensitiy for β2-microglobulin was analysed by quantitative immunohistochemistry using NIH image software. The reference area for ‘distal ORS’ is on the level of the insertion of the sebaceous duct and for the ‘bulge’ at the insertion point of the arrector pili muscle. N=5, **p<0.01, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
4.1.2 Demonstration of MHC class II+ cells in the bulge
A monoclonal antibody specific against HLA-DP/DQ/DR was used for the localization
of intraepithelial MHC class II+ antigen-presenting cells such as Langerhans cells
and macrophages (SMITH et al. 1987). The most prominent expression of MHC
class II was epidermal (data not shown) and expression was particularly dense in the
infundibulum and upper parts of the isthmus (Fig. 4.9 A,B). Here we show, that the
bulge region, as the proximal ORS, exhibited a significantly reduced expression of
MHC class II compared to the upper parts of the HF (Figs. 4.9 C,D). As expected,
the bulb region only rarely showed single MHC class+ cells (Fig. 4.9 D) that were not
found in the DP. This was confirmed by quantitative analysis using NIH image (Figs.
4.10, 4.11).

RESULTS
138
While the distal perifollicular CTS displayed a high density of MHC class II+ cells,
most of which may represent macrophages, the lowest density was observed in the
isthmus region of the CTS. The CTS around the proximal HF and the bulb showed
more MHC class II+ cells than the CTS at the level of the isthmus region.
Fig. 4.9 IR in the human bulge region for MHC class II Immunohistochemical identification of MHC class II protein expression (HLA-DP/-DQ/DR) in normal human scalp skin (EnVision®) (A-D) /red). (A) MHC class II+ cells can be found in high numbers in the epidermis (A) and distal ORS of isthmus and infundibulum (B) whereas the bulge region and the hair bulb show only rare MHC class+ cells. APM: Arrector pili muscle, B: Bulge, CTS: Connective tissue sheath, E: Epidermis; HS: Hair shaft, MK: Matrix keratinocytes, ORS: Outer root sheath, SG: Sebaceous gland, Bars (A-D): 100µm.
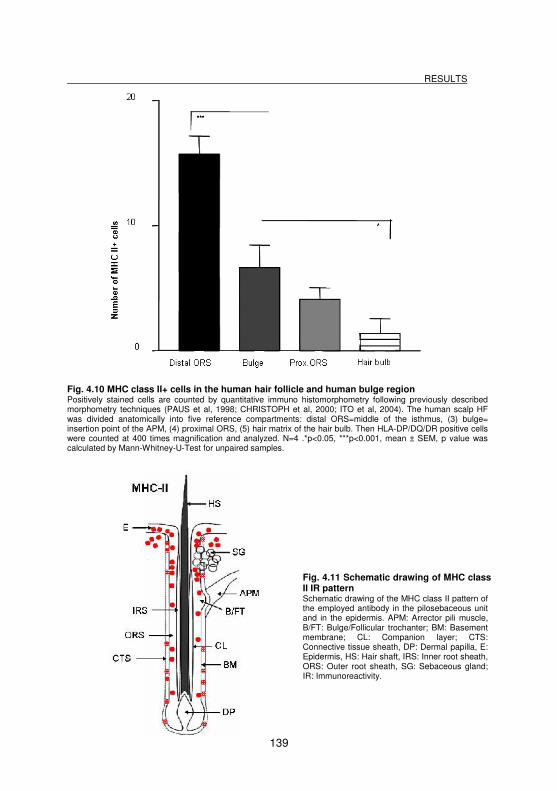
RESULTS
139
Fig. 4.10 MHC class II+ cells in the human hair follicle and human bulge region Positively stained cells are counted by quantitative immuno histomorphometry following previously described morphometry techniques (PAUS et al, 1998; CHRISTOPH et al, 2000; ITO et al, 2004). The human scalp HF was divided anatomically into five reference compartments: distal ORS=middle of the isthmus, (3) bulge= insertion point of the APM, (4) proximal ORS, (5) hair matrix of the hair bulb. Then HLA-DP/DQ/DR positive cells were counted at 400 times magnification and analyzed. N=4 .*p<0.05, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.11 Schematic drawing of MHC class II IR pattern Schematic drawing of the MHC class II pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.
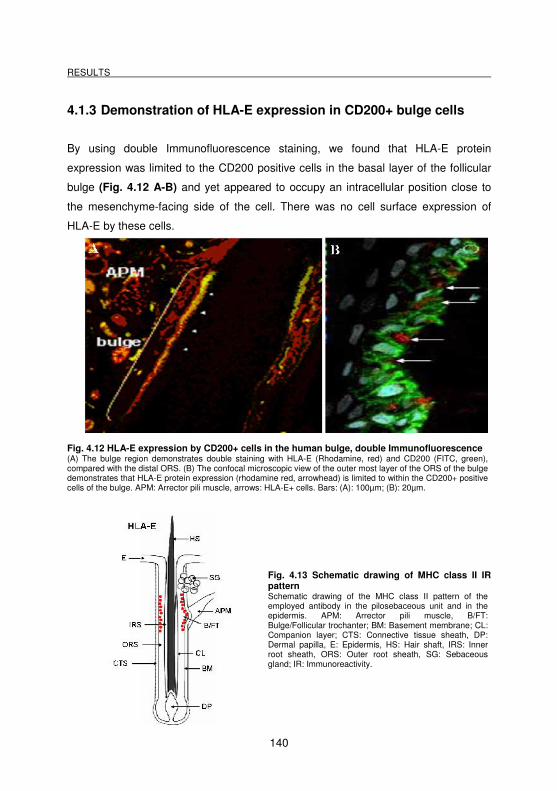
RESULTS
140
4.1.3 Demonstration of HLA-E expression in CD200+ bulge cells
By using double Immunofluorescence staining, we found that HLA-E protein
expression was limited to the CD200 positive cells in the basal layer of the follicular
bulge (Fig. 4.12 A-B) and yet appeared to occupy an intracellular position close to
the mesenchyme-facing side of the cell. There was no cell surface expression of
HLA-E by these cells.
Fig. 4.12 HLA-E expression by CD200+ cells in the human bulge, double Immunofluorescence (A) The bulge region demonstrates double staining with HLA-E (Rhodamine, red) and CD200 (FITC, green), compared with the distal ORS. (B) The confocal microscopic view of the outer most layer of the ORS of the bulge demonstrates that HLA-E protein expression (rhodamine red, arrowhead) is limited to within the CD200+ positive cells of the bulge. APM: Arrector pili muscle, arrows: HLA-E+ cells. Bars: (A): 100µm; (B): 20µm.
Fig. 4.13 Schematic drawing of MHC class II IR pattern Schematic drawing of the MHC class II pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.

RESULTS
141
4.1.4 Locally generated immunosuppressants complement the
bulge IP
Since immunosuppressants like TGF-β and α-MSH play a major role in IP, it is
important that we could show a substantial increase of α-MSH IR in the bulge and
proximal ORS - in particular in the suprabasal layers - (Fig. 4.14 A-C) compared to
the epidermis and the distal ORS (Figs. 4.14 A, 4.16).
Fig. 4.14 IR in the human bulge region for α-MSH α-MSH (green) shows upregulated IR in the ORS of the bulge region and proximal ORS compared to the upper part of the HF and the epidermis. APM: Arrector pili muscle, E: Epidermis; FT: Follicular trochanter, ORS: Outer root sheath, SG: Sebaceous gland, Bars (A): 200µm; (B,C): 20µm.
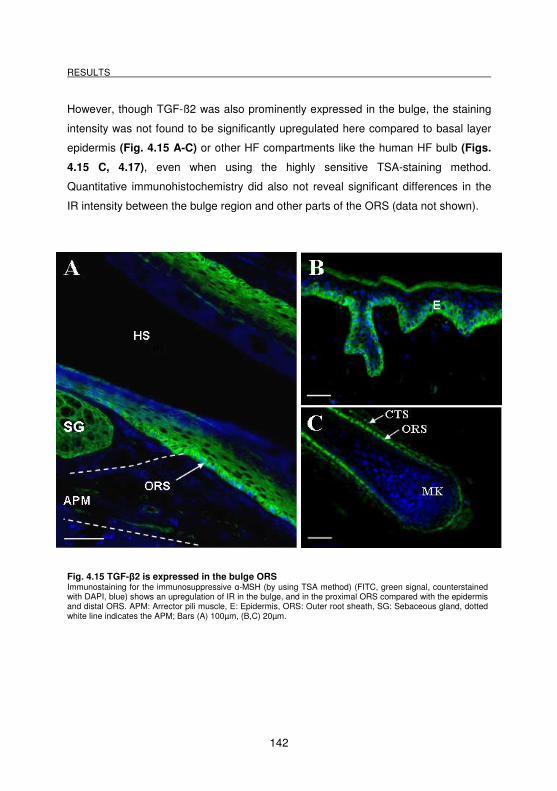
RESULTS
142
However, though TGF-ß2 was also prominently expressed in the bulge, the staining
intensity was not found to be significantly upregulated here compared to basal layer
epidermis (Fig. 4.15 A-C) or other HF compartments like the human HF bulb (Figs.
4.15 C, 4.17), even when using the highly sensitive TSA-staining method.
Quantitative immunohistochemistry did also not reveal significant differences in the
IR intensity between the bulge region and other parts of the ORS (data not shown).
Fig. 4.15 TGF-β2 is expressed in the bulge ORS Immunostaining for the immunosuppressive α-MSH (by using TSA method) (FITC, green signal, counterstained with DAPI, blue) shows an upregulation of IR in the bulge, and in the proximal ORS compared with the epidermis and distal ORS. APM: Arrector pili muscle, E: Epidermis, ORS: Outer root sheath, SG: Sebaceous gland, dotted white line indicates the APM; Bars (A) 100µm, (B,C) 20µm.

RESULTS
143
Fig. 4.16 Schematic drawing of α-MSH IR pattern Schematic drawing of the α-MSH IR pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.
Fig. 4.17 Schematic drawing of TGF-β2 IR pattern Schematic drawing of the TGF-β2 IR pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.

RESULTS
144
4.1.5 Demonstration of macrophage migration inhibitory factor and
indoleamine-2,3-dioxygenase
Next, we investigated the local expression of the immunosuppressant factor MIF.
Compared to the epidermis and the distal ORS (Fig. 4.18 A,B), MIF-IR was
upregulated in the bulge and proximal ORS (Fig. 4.18 A), where it was limited to the
suprabasal layer. Quantitative analysis of MIF-IR in the bulge confirmed this
significant upregulation compared to the distal ORS and to the epidermis (Figs. 4.19,
4.23) (*p<0.01).
Fig. 4.18 IR is upregulated in the human bulge region for MIF MIF (red) IR can be found throughout the whole ORS, with an upregulation in the bulge region and proximal parts of the ORS. The epidermis and APM are weakly stained for MIF. APM: Arrector pili muscle, BM: Basement membrane, HS: Hair shaft, ORS: Outer root sheath, SG: Sebaceous gland, dotted white line indicates the APM; Bars: (A) 200µm; (B) 50µm.
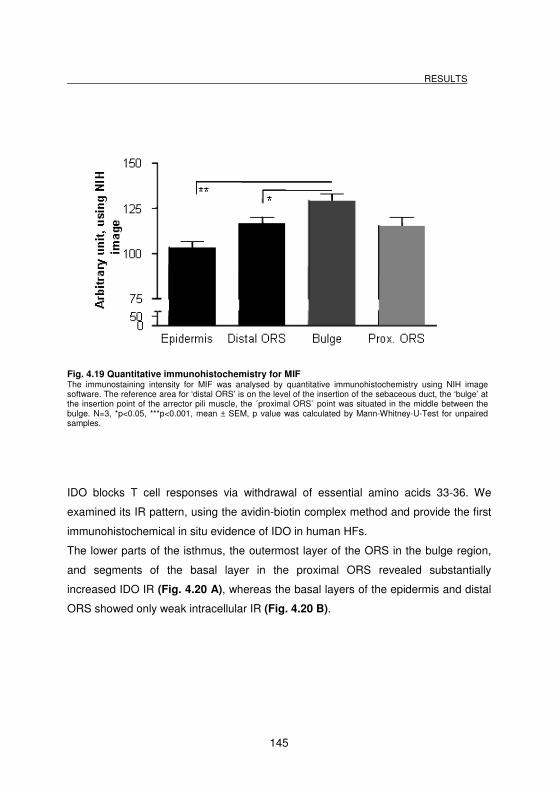
RESULTS
145
Fig. 4.19 Quantitative immunohistochemistry for MIF The immunostaining intensity for MIF was analysed by quantitative immunohistochemistry using NIH image software. The reference area for ‘distal ORS’ is on the level of the insertion of the sebaceous duct, the ‘bulge’ at the insertion point of the arrector pili muscle, the ´proximal ORS` point was situated in the middle between the bulge. N=3, *p<0.05, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
IDO blocks T cell responses via withdrawal of essential amino acids 33-36. We
examined its IR pattern, using the avidin-biotin complex method and provide the first
immunohistochemical in situ evidence of IDO in human HFs.
The lower parts of the isthmus, the outermost layer of the ORS in the bulge region,
and segments of the basal layer in the proximal ORS revealed substantially
increased IDO IR (Fig. 4.20 A), whereas the basal layers of the epidermis and distal
ORS showed only weak intracellular IR (Fig. 4.20 B).

RESULTS
146
Fig. 4.20 IR is upregulated in the human bulge region for IDO IDO IR (brown, DAB substrate) is upregulated in the basal layers of the bulge ORS (A) compared to the distal ORS and epidermis (B). The APM is weakly stained positive. APM: Arrector pili muscle, BM: Basement membrane, E: epidermis, HS: Hair shaft, SG: Sebaceous gland; Bars: (A) 50µm; (B) 200µm.
Interestingly, the staining intensity decreased in the proximal ORS with a sparse
distribution of IDO+ keratinocytes (Fig. 4.21 B) and no IR was found in the hair bulb
(data not shown) (Fig. 4.23). Quantitative analysis corroborated a significantly
increased IR in the bulge, compared to distal ORS and epidermis (Fig. 4.22)
(***p<0.001).

RESULTS
147
Fig. 4.21 IR in different parts of the proximal ORS for IDO A heterogenous staining intensity of IDO is seen in the proximal ORS (red, AEC substrate). Compared to the suprabulge ORS (A), IR decreased in lower parts of the proximal ORS with a sparse distribution of IDO+ keratinocytes (B). No IR was found in the IRS or the hair bulb (data not shown). APM: Arrector pili muscle, BM: Basement membrane, HS: Hair shaft; IRS: Inner root sheath; ORS: Outer root sheath; Bars: (A, B) 50µm.
Fig. 4.22 Quantitative immunohistochemistry for IDO The immunostaining intensity for IDO was analysed by quantitative immunohistochemistry using NIH image software. The reference area for ‘distal ORS’ is on the level of the insertion of the sebaceous duct, the ‘bulge’ at the insertion point of the arrector pili muscle. N=7, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.

RESULTS
148
Fig. 4.23 Schematic drawings of MIF and IDO IR pattern Schematic drawing of the MIF and IDO IR pattern of the employed antibody in the pilosebaceous unit and in the epidermis. APM: Arrector pili muscle, B/FT: Bulge/Follicular trochanter; BM: Basement membrane; CL: Companion layer; CTS: Connective tissue sheath, DP: Dermal papilla, E: Epidermis, HS: Hair shaft, IRS: Inner root sheath, ORS: Outer root sheath, SG: Sebaceous gland; IR: Immunoreactivity.

RESULTS
149
4.1.6 Influence of IFN-γ on ectopic MHC class I protein expression
in the bulge ORS
In order to check whether IFN-γ also causes ectopic MHC class I expression in the
bulge epithelium such as in the hair matrix, we performed organ culture of full-
thickness human scalp skin fragments under serum-free conditions and treated them
with IFN-γ or vehicle, and evaluated MHC class I IR by quantitative immuno
histomorphometry. The cultured human anagen VI HFs in situ showed a consistent
tendency towards ectopic MHC class I expression in the bulge region (Fig. 4.24).
However, only high-dose IFN-γ treatment (1000 IU/ml) reached the level of
significance (Fig. 4.25) (*p<0.05). Thus, just as the anagen hair bulb IP, the MHC
class I-based IP of the human bulge collapses after exposure to sufficient amounts of
IFN-γ.
Fig. 4.24 Influence of IFN-γ on MHC class I expression of human HFs in full thickness organ culture Full-thickness human scalp skin punch biopsies were organ cultured and groups were treated with IFN-γ (100, 500, 1000 IU/ml) or PBS. Then, Immunofluorescence staining for MHC class I (green) was performed and IR was compared. (A) PBS control group, (B) 1000 IU/ml IFN-γ treated sample. APM: Arrector pili muscle, HS: Hair shaft, ORS: Outer root sheath, SG: Sebaceous gland, Bars: (A,B) 50µm.

RESULTS
150
Fig. 4.25 Quantitative analysis of MHC class I staining intensity in the bulge Full-thickness human scalp skin punch biopsies were organ cultured and groups were treated with IFN-γ (100, 500, 1000 IU/ml) or PBS. Then, IF staining for MHC class I (green) was performed and IR in the bulge region of anagen VI HFs was compared. The group treated with 1000 IU/ml IFN-γ revealed a significant increase in MHC class I IR. N=2, *p<0.05, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.

RESULTS
151
4.2 Influence of the α-MSH related tripeptide K(D)PT on human hair
follicle biology in situ under pro-inflammatory conditions
4.2.1 Influence of K(D)PT on IFN-γ induced MHC class I and II
protein expression
IFN-γ significantly upregulated MHC class Ia IR in the hair bulb of human anagen VI
HFs, whereas the control HFs treated with K(D)PT 10-8M alone displayed no
differences compared to vehicle treated HFs (Fig. 4.2 C). In the majority of
experiments none of the K(D)PT doses could significantly and reliably reduce the
IFN-γ-induced MHC class I IR in the hair matrix and proximal ORS (Fig. 4.27 A,B),
although K(D)PT 10-8 often showed the tendency to and evoked in one patient a
significant downregulation of MHC class Ia IR (Figs. 4.26, 4.27 C).
Fig. 4.26 Isolated HFs stained for MHC class I EnVision® staining of isolated and cultured HFs for MHC class I (red).
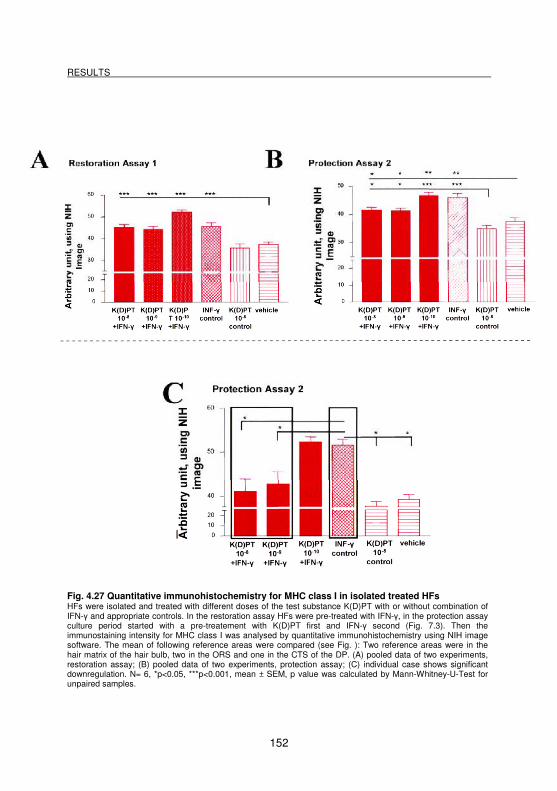
RESULTS
152
Fig. 4.27 Quantitative immunohistochemistry for MHC class I in isolated treated HFs HFs were isolated and treated with different doses of the test substance K(D)PT with or without combination of IFN-γ and appropriate controls. In the restoration assay HFs were pre-treated with IFN-γ, in the protection assay culture period started with a pre-treatement with K(D)PT first and IFN-γ second (Fig. 7.3). Then the immunostaining intensity for MHC class I was analysed by quantitative immunohistochemistry using NIH image software. The mean of following reference areas were compared (see Fig. ): Two reference areas were in the hair matrix of the hair bulb, two in the ORS and one in the CTS of the DP. (A) pooled data of two experiments, restoration assay; (B) pooled data of two experiments, protection assay; (C) individual case shows significant downregulation. N= 6, *p<0.05, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.

RESULTS
153
Similarly, the expected significant upregulation of MHC class II+ cells in the hair bulb
of IFN-γ treated control HFs (Fig. 4.28) could not be significantly and reliably
downregulated by any K(D)PT dose, neither in the number of MHC class II+ cells
(Fig. 4.28), nor in the staining intensity (Fig. 4.29). However, K(D)PT showed the
tendency for downregulation and we also observed interindividual differences and
susceptibility (data not shown).
Fig. 4.28 Quantitative analysis of the mean number of MHC class II+ cells Quantitative immuno histomorphometry using NIH Image of MHC class II+ cells (A) Assay 1: pre-treatment with IFN-γ, pooled data of two experiments, N=10-30 HFs/group, (B) Assay 2: pre-treatment with K(D)PT, pooled data of two experiments, N=10-30 HFs/group, N= 6-18 HFs/group, *p<0.05, **p<0.01, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples. Original magnification 630x.
RESULTS
154
Fig. 4.29 Staining intensity of MHC class II+ cells Immunostaining for MHC class II+ cells in K(D)PT treated HFs (A) versus control HFs (B) [using the tyramide signal amplification (TSA) method, green signal] did not reveal any marked differences in the IR level (no formal quantification done).

RESULTS
155
4.2.2 Reaction of K(D)PT on the IFN-γ induced upregulation of total
mast cell numbers and mast cell degranulation
Following treatment, cryosections of organ-cultured HFs were stained with the Leder
esterase staining (Fig. 4.30 A) and for c-kit (Fig. 4.30 B), in order to investigate how
K(D)PT influenced the number of mast cells and their activation status.
Fig. 4.30 Staining for mast cells (A) Representative photodocument of a Leder esterase stained HF, using the standard leder esterase protocol, cytoplasmic mast cell granules stained red, green counterstain, original magnification 400x. (B) Representative photodocument of c-Kit staining (green signal/FITC), original magnification 200x.
Stimulation with IFN-γ induced both a significant upregulation of mast cell numbers
and an increased degranulation compared to K(D)PT– or vehicle-treated control
groups (Fig. 4.31). High dose K(D)PT 10-8M always showed a strong reduction of this
increase in mast cell numbers (Fig. 4.31), although significance was only reached in
the restoration assay (assay 2) with IFN-γ pre-treatment. This was confirmed by c-Kit
immunofluorescence staining in the CTS of the HF (Fig. 4.30).

RESULTS
156
Fig. 4.31 Quantitative analysis of the number of mast cells and their degranulation ratio Every Leder estease stained HF was analysed quantitatively regarding its number of mast cells (non-degranulated/degranulated) and the mean was calculated. Representative experiment of assay 2, n=10-18 hair follicles/group. Mean ± SEM, *p<0.05. p value was calculated by Mann-Whitney-U-Test for unpaired samples.

RESULTS
157
4.2.3 Influence of K(D)PT on the hair cycle
Microdissected and cultured HFs were assessed with respect to their hair cycle
status. As shown in Fig. 4.32, this procedure revealed significant differences in the
ratio of HFs in distinct hair cycle stages: In the vehicle and the K(D)PT control groups
most of the HFs were still in anagen (~ 75%) and only a small part in early catagen
stage. The treatment with the pro-inflammatory cytokine IFN-γ reduced significantly
the percentage of HFs, which were in anagen (~ 50%) and displayed more HFs in
mid catagen. We could demonstrate, that supplementation with high dose K(D)PT
(10–8M, 10-9M) significantly counteracted against the effects of IFN-γ, both in assay 1
and assay 2. In addition, the effects seemed to inhibit even more catagen
transformation than in the vehicle treated control groups (~ 90% of the HFs in anagen
stage) (Fig. 4.32).
Fig. 4.32 Influence of K(D)PT on the hair cycle stage The total number (%) of HFs in the different hair cycle stages is shown. High-dose K(D)PT treated HFs under pro-inflammatory stimulus counteract the IFN-γ induced catagen transformation, and display a similar or even higher percentage of HFs being in anagen. N=8-12 follicles/group, pooled data of four experiments, Mean ± SEM, *p<0.05, **p<0.01. p value was calculated by Mann-Whitney-U-Test for unpaired samples.
* *
* *
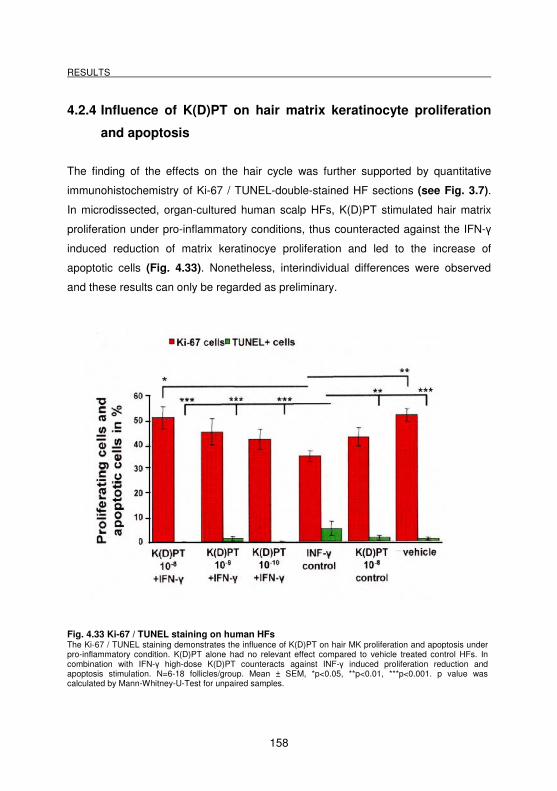
RESULTS
158
4.2.4 Influence of K(D)PT on hair matrix keratinocyte proliferation
and apoptosis
The finding of the effects on the hair cycle was further supported by quantitative
immunohistochemistry of Ki-67 / TUNEL-double-stained HF sections (see Fig. 3.7).
In microdissected, organ-cultured human scalp HFs, K(D)PT stimulated hair matrix
proliferation under pro-inflammatory conditions, thus counteracted against the IFN-γ
induced reduction of matrix keratinocye proliferation and led to the increase of
apoptotic cells (Fig. 4.33). Nonetheless, interindividual differences were observed
and these results can only be regarded as preliminary.
Fig. 4.33 Ki-67 / TUNEL staining on human HFs The Ki-67 / TUNEL staining demonstrates the influence of K(D)PT on hair MK proliferation and apoptosis under pro-inflammatory condition. K(D)PT alone had no relevant effect compared to vehicle treated control HFs. In combination with IFN-γ high-dose K(D)PT counteracts against INF-γ induced proliferation reduction and apoptosis stimulation. N=6-18 follicles/group. Mean ± SEM, *p<0.05, **p<0.01, ***p<0.001. p value was calculated by Mann-Whitney-U-Test for unpaired samples.
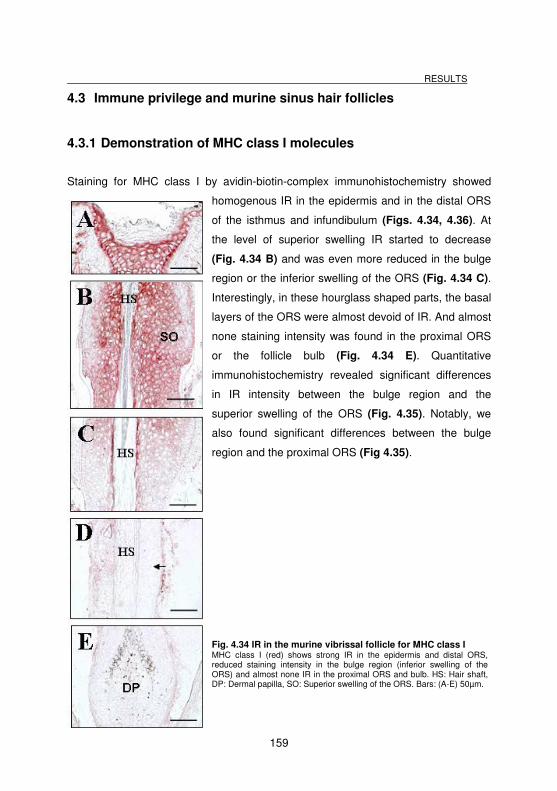
RESULTS
159
4.3 Immune privilege and murine sinus hair follicles
4.3.1 Demonstration of MHC class I molecules
Staining for MHC class I by avidin-biotin-complex immunohistochemistry showed
homogenous IR in the epidermis and in the distal ORS
of the isthmus and infundibulum (Figs. 4.34, 4.36). At
the level of superior swelling IR started to decrease
(Fig. 4.34 B) and was even more reduced in the bulge
region or the inferior swelling of the ORS (Fig. 4.34 C).
Interestingly, in these hourglass shaped parts, the basal
layers of the ORS were almost devoid of IR. And almost
none staining intensity was found in the proximal ORS
or the follicle bulb (Fig. 4.34 E). Quantitative
immunohistochemistry revealed significant differences
in IR intensity between the bulge region and the
superior swelling of the ORS (Fig. 4.35). Notably, we
also found significant differences between the bulge
region and the proximal ORS (Fig 4.35).
Fig. 4.34 IR in the murine vibrissal follicle for MHC class I MHC class I (red) shows strong IR in the epidermis and distal ORS, reduced staining intensity in the bulge region (inferior swelling of the ORS) and almost none IR in the proximal ORS and bulb. HS: Hair shaft, DP: Dermal papilla, SO: Superior swelling of the ORS. Bars: (A-E) 50µm.

RESULTS
160
Fig. 4.35 Quantitative immunohistochemistry for MHC class I The immunostaining intensity for MHC class I was analysed by quantitative immunohistochemistry using NIH image software. The vibrissal follicle was divided according to the human HFs into five compartments 1) isthmus, (2) distal ORS=superior swelling of the ORS, (3) bulge= inferior swelling of the ORS, (4) proximal ORS, (5) hair matrix, and analyzed. deriving from 5-10 different mice. N=5, *p<0.05, **p<0.01, ***p<0.001, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.36 Schematic drawing of MHC class I IR pattern Schematic drawing of the MHC class I IR pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity.
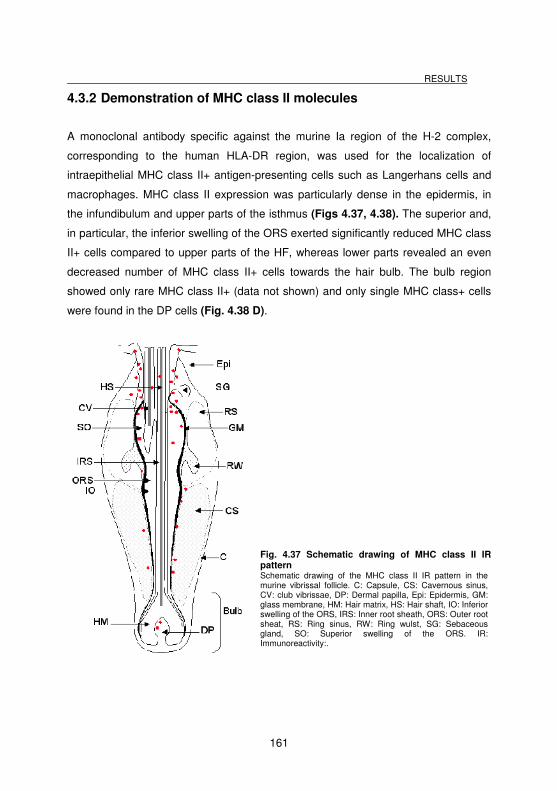
RESULTS
161
4.3.2 Demonstration of MHC class II molecules
A monoclonal antibody specific against the murine Ia region of the H-2 complex,
corresponding to the human HLA-DR region, was used for the localization of
intraepithelial MHC class II+ antigen-presenting cells such as Langerhans cells and
macrophages. MHC class II expression was particularly dense in the epidermis, in
the infundibulum and upper parts of the isthmus (Figs 4.37, 4.38). The superior and,
in particular, the inferior swelling of the ORS exerted significantly reduced MHC class
II+ cells compared to upper parts of the HF, whereas lower parts revealed an even
decreased number of MHC class II+ cells towards the hair bulb. The bulb region
showed only rare MHC class II+ (data not shown) and only single MHC class+ cells
were found in the DP cells (Fig. 4.38 D).
Fig. 4.37 Schematic drawing of MHC class II IR pattern Schematic drawing of the MHC class II IR pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity:.

RESULTS
162
Fig. 4.38 MHC class II positive cells in the murine vibrissal follicle Immunohistochemical identification of MHC class II+ cells (brown) in normal murine snout skin (ABC method). MHC class II+ cells can be found in a relative high number in the ORS of upper parts of the vibrissal follicle and isthmus, whereas the inferior swelling of the ORS and lower parts of the follicle show only single or no positive cells in the ORS. (A) epidermis, isthmus, superior swelling and inferior swelling of the ORS; (B) lower parts of the vibrissal follilce: hair shaft and hair bulb; (b) superior swelling of the ORS. Bars: (A-B) 100µm, (b) original magnification 400x.

RESULTS
163
HS
4.3.3 Demonstration of CD4+ T cells
CD4+ T cells were predominantly expressed in upper parts of the vibrissal follicle, in
the isthmus, whereas in the superior swelling of the ORS and even more in parts of
the inferior swelling of the ORS, a significantly reduced number of this cell type was
found (Fig. 4.39). CD4+ T cells were also found in lower parts of the vibrissal follicle,
although most of them were localized in the sinus. The bulb region showed only
single CD4+ T cells (Figs 4.39, 4.41) and no CD4+ T cell could be detected in the
dermal papilla. Quantitative analysis corroborated the signigicant differences
between the compartments (Fig. 4.40).
Fig. 4.39 IR in the murine vibrissal follicle for CD4+ T cells Immunohistochemical identification of CD4+ T cells (brown) in normal murine snout skin (ABC method). CD4+ T cells can be found in high numbers in upper parts of the vibrissal follicle and isthmus, whereas the inferior swelling of the ORS and lower parts of the follicle show only rare CD4+ T cells. HS: Hair shaft, DP: Dermal papilla, SO: Superior swelling of the ORS. Bars: (A-E) 50µm.

RESULTS
164
Fig. 4.40 Quantitative immunohistochemistry for CD4 The immunostaining intensity for CD4 was analysed by quantitative immunohistochemistry using NIH image software. The vibrissal follicle was divided according to the human HFs into five compartments 1) isthmus, (2) distal ORS=superior swelling of the ORS, (3) bulge= inferior swelling of the ORS, (4) proximal ORS, (5) hair matrix, and analysed. deriving from 5-10 different mice. N=5, *p<0.05, **p<0.01, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.41 Schematic drawing of CD4 pattern Schematic drawing of the CD4 IR pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity.
Isthmus SO IO prox. ORS Hair bulb0
1020
25
30
35*
***
**
nu
mb
er
of
CD
4+
ce
lls

RESULTS
165
4.3.4 Demonstration of CD11b molecules
In order to detect CD11b on myeloid cells, we used again the avidin-biotin-complex
staining method. CD11b+ cells were often visible particularly in the epidermis, the
infundibulum and upper parts of the isthmus (Fig. 4.42). The superior and inferior
swelling of the ORS, as well as lower parts
of the vibrissal follicle, demonstrated a
reduced number of CD11+ cells.
The bulb region and DP showed only
single CD11b+ cells (Figs. 4.42, 4.44).
Interestingly, many CD11b+ cells occurred
in the surroundings of the vibrissal capsule.
This was confirmed by quantitative analysis
(Fig. 4.43).
Fig. 4.42 IR in the murine vibrissal follicle for CD11b Immunohistochemical identification of CD11b cells (brown) in normal murine snout skin (ABC method). CD11b+ cells can be found in high numbers in upper parts of the vibrissal follicle and isthmus, whereas the inferior swelling of the ORS and lower parts of the follicle show only rare CD11b+ cells. DP: Dermal pailla, IO: Inferior swelling of the ORS, SO: Suprior swelling of the ORS, 100x maginification
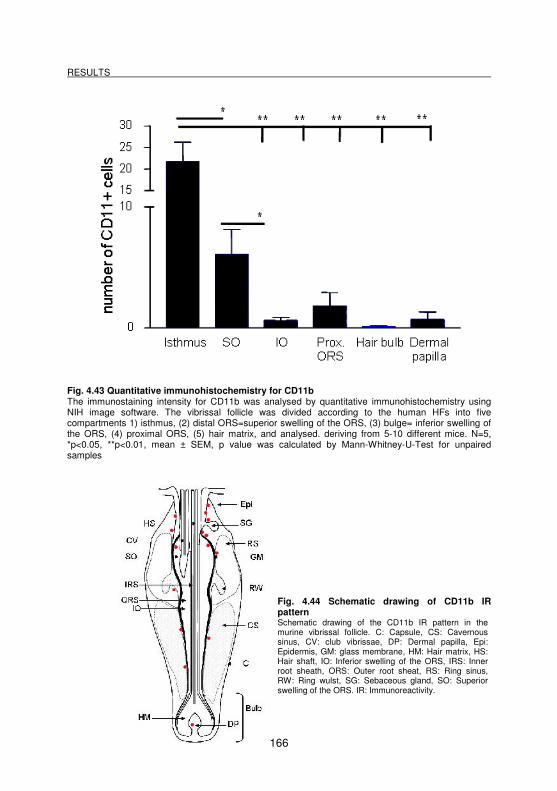
RESULTS
166
Fig. 4.43 Quantitative immunohistochemistry for CD11b The immunostaining intensity for CD11b was analysed by quantitative immunohistochemistry using NIH image software. The vibrissal follicle was divided according to the human HFs into five compartments 1) isthmus, (2) distal ORS=superior swelling of the ORS, (3) bulge= inferior swelling of the ORS, (4) proximal ORS, (5) hair matrix, and analysed. deriving from 5-10 different mice. N=5, *p<0.05, **p<0.01, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples
Fig. 4.44 Schematic drawing of CD11b IR pattern Schematic drawing of the CD11b IR pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity.

RESULTS
167
4.3.5 Demonstration of TGF-β1 molecules
A polyclonal antibody against TGF-β1 was used for the localization of precursor and
mature TGF-β1+ and TGF-β2+ cells. The most prominent expression of TGF-β was
epidermal (Fig. 4.45 A) and in the ORS of the infundibulum - showing both a basal
and suprabasal cytoplasmic staining. In the
superior swelling of the ORS
immunohistochemistry decreased and was
almost exclusively evident in the basal
layers of the ORS. No staining was detected
in the IRS (Fig. 4.45 B). In contrast, the
bulge region (namely the inferior swelling of
the ORS) exerts a highly increased
immunostaining intensity compared with the
SO parts (Fig. 4.45 C). Interestingly, the
ORS of the bulge region did not
demonstrate a uniform expression pattern,
but an increased IR in the basal layers with
a sparse distribution of more intense stained
TGF-β1+ keratinocytes. Staining intensity
was reduced in the lower vibrissal follicle
and in the hair bulb (Fig. 4.45 D).
Fig. 4.45 IR in the murine vibrissal follicle for TGF-β1 on cryosections Immunohistochemical identification of TGF-β1 (brown) in normal murine sinus follicle cryosections (ABC method). TGF-β1 IR can be observed in high numbers in upper parts of the vibrissal follicle and isthmus, whereas the inferior swelling of the ORS and lower parts of the follicle show only rare CD4+ T cells. E: HS: Hair shaft, DP: Dermal papilla, SO: Superior swelling of the ORS. Bars: (A-E) 50µm.
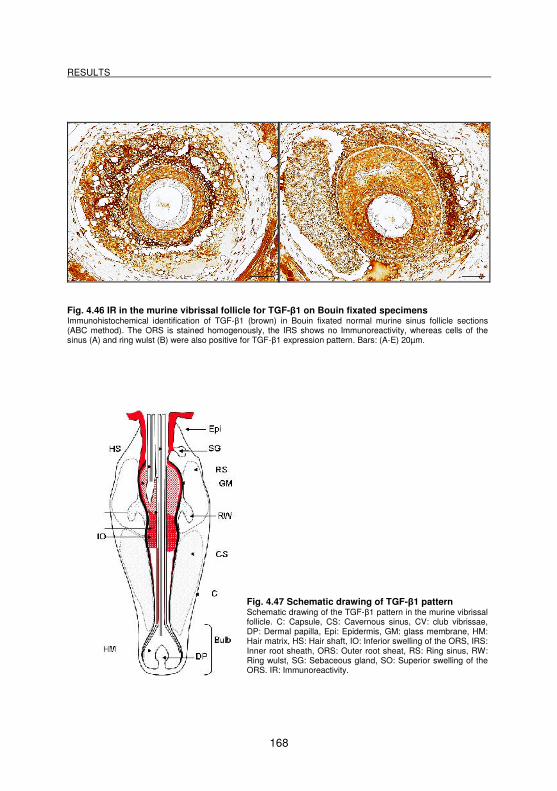
RESULTS
168
Fig. 4.46 IR in the murine vibrissal follicle for TGF-β1 on Bouin fixated specimens Immunohistochemical identification of TGF-β1 (brown) in Bouin fixated normal murine sinus follicle sections (ABC method). The ORS is stained homogenously, the IRS shows no Immunoreactivity, whereas cells of the sinus (A) and ring wulst (B) were also positive for TGF-β1 expression pattern. Bars: (A-E) 20µm.
Fig. 4.47 Schematic drawing of TGF-β1 pattern Schematic drawing of the TGF-β1 pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity.
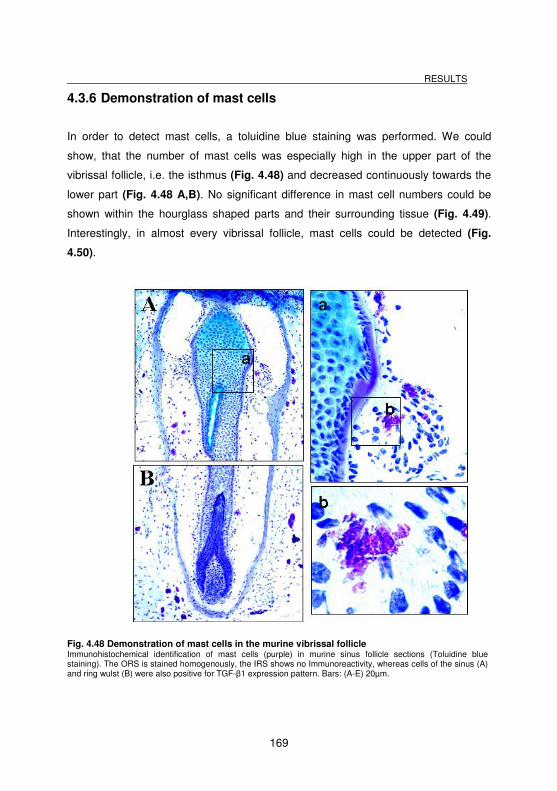
RESULTS
169
4.3.6 Demonstration of mast cells
In order to detect mast cells, a toluidine blue staining was performed. We could
show, that the number of mast cells was especially high in the upper part of the
vibrissal follicle, i.e. the isthmus (Fig. 4.48) and decreased continuously towards the
lower part (Fig. 4.48 A,B). No significant difference in mast cell numbers could be
shown within the hourglass shaped parts and their surrounding tissue (Fig. 4.49).
Interestingly, in almost every vibrissal follicle, mast cells could be detected (Fig.
4.50).
Fig. 4.48 Demonstration of mast cells in the murine vibrissal follicle Immunohistochemical identification of mast cells (purple) in murine sinus follicle sections (Toluidine blue staining). The ORS is stained homogenously, the IRS shows no Immunoreactivity, whereas cells of the sinus (A) and ring wulst (B) were also positive for TGF-β1 expression pattern. Bars: (A-E) 20µm.
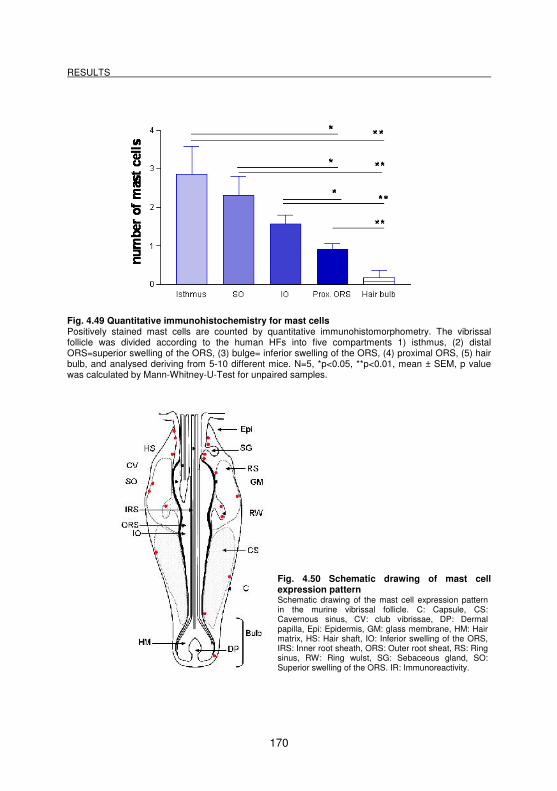
RESULTS
170
Fig. 4.49 Quantitative immunohistochemistry for mast cells Positively stained mast cells are counted by quantitative immunohistomorphometry. The vibrissal follicle was divided according to the human HFs into five compartments 1) isthmus, (2) distal ORS=superior swelling of the ORS, (3) bulge= inferior swelling of the ORS, (4) proximal ORS, (5) hair bulb, and analysed deriving from 5-10 different mice. N=5, *p<0.05, **p<0.01, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.50 Schematic drawing of mast cell expression pattern Schematic drawing of the mast cell expression pattern in the murine vibrissal follicle. C: Capsule, CS: Cavernous sinus, CV: club vibrissae, DP: Dermal papilla, Epi: Epidermis, GM: glass membrane, HM: Hair matrix, HS: Hair shaft, IO: Inferior swelling of the ORS, IRS: Inner root sheath, ORS: Outer root sheat, RS: Ring sinus, RW: Ring wulst, SG: Sebaceous gland, SO: Superior swelling of the ORS. IR: Immunoreactivity.

RESULTS
171
4.4 Immune privilege and the murine mouse nail apparatus
4.4.1 Demonstration of MHC class I molecules
Fig. 4.51 IR in the murine nail apparatus for MHC class I Expression pattern of MHC class I in normal murine nail apparatus (brown signal). The staining is very strong in the PNF and decreases towards the PNM. Bars: (A) 100µm, (B,C) 20µm.
By using avidin biotin-complex staining method with a monoclonal antibody against
H-2Dd, H-2b,p,q, we found that MHC class I IR in the murine nail apparatus changed
interindiviudally and also in regard on the fixation method applied (cryosections
versus Bouin-fixated tissue).
In most of the mice cryosections samples we found that the nail matrix epithelium,
and very prominently the PNM, displayed significantly downregulated MHC class I
Immunoreactivity (Fig. 4.51 A,C), compared with PNF (Fig. 4.51 B), nail bed, and
hyponychium (Fig. 4.51 A). This low or absent MHC class I expression in nail matrix
epithelium was mirrored in the immediately adjacent nail mesenchyme (Figs. 4.51-
4.53). The pad epithelium was only weak or not stained.
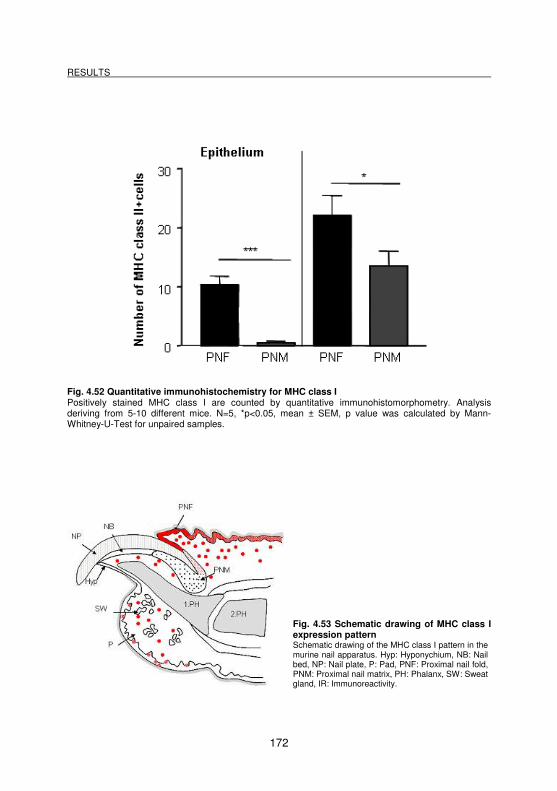
RESULTS
172
Fig. 4.52 Quantitative immunohistochemistry for MHC class I Positively stained MHC class I are counted by quantitative immunohistomorphometry. Analysis deriving from 5-10 different mice. N=5, *p<0.05, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.53 Schematic drawing of MHC class I expression pattern Schematic drawing of the MHC class I pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
173
4.4.2 Demonstration of MHC class II molecules
In order to detect MHC class II+ cells we used again avidin-biotin-complex method.
MHC class II+ cell were found in high numbers in the epidermis and in the vicinity of
the PNF (Fig. 4.54 A), whereas no MHC class II+ cells were detectable in the PNM
(*p<0.05) and in its mesenchymal vicinity (***p<0.001) (Fig. 4.54 B). The nail bed
only rarely showed MHC class II+ cells. In the foot pad epithelium, the greatest
number of intraepithelial MHC class II+ cells could be demonstrated (Fig. 4.54 D),
whereas the dermis showed a reduced number compared to the PNF and PNM
dermis (Fig. 4.56). This was confirmed by quantitative analysis using NIH imaging
(Fig. 4.55).
Fig. 4.54 IR in the murine nail apparatus for MHC class II Expression pattern of MHC class II in normal murine nail apparatus (brown signal). (A) MHC class II+ cells can be found in high numbers in the epidermis and in the vicinity of the PNF, whereas the NB shows only single (B) and the PNM show no MHC class II+ cells (C). In pad epithelium most of the intraepithelial MHC class II+ cells can be demonstrated. Bars: (A-D) 200µm.
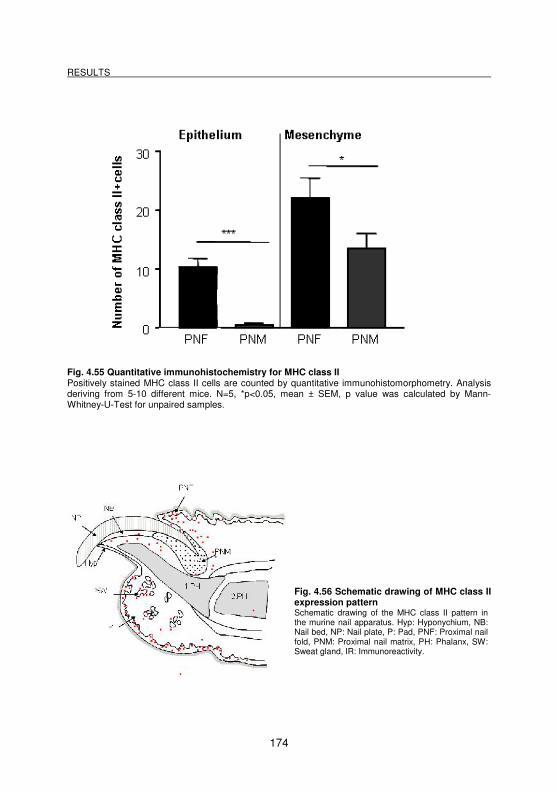
RESULTS
174
Fig. 4.55 Quantitative immunohistochemistry for MHC class II Positively stained MHC class II cells are counted by quantitative immunohistomorphometry. Analysis deriving from 5-10 different mice. N=5, *p<0.05, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.56 Schematic drawing of MHC class II expression pattern Schematic drawing of the MHC class II pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
175
4.4.3 Demonstration of CD4+ T cells
CD4+ T cell localization was identified by avidin-biotin-complex staining. We could
show, that the number of CD4+ T cells was especially high in the vicinity of the PNF
(Fig. 4.57 A) and decreased continuously towards the PNM (Fig. 4.57 B).
Intraepithelial CD4+ T cells were only found in the PNF and foot pad, but not in the
PNM; quantification was not done (Fig. 4.58).
Fig. 4.57 IR in the murine nail apparatus for CD4+ T cells Expression pattern of CD4 in normal human scalp skin (brown). NP: Nail plate, PNF: Proximal nail fold, PNM: Proximal nail matrix, Bars: (A,B) 100µm.
Fig. 4.58 Schematic drawing of CD4 expression pattern Schematic drawing of the CD4 expression pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
176
4.4.4 Demonstration of TGF-β1 molecules
A polyclonal antibody against TGF-β1 was used for the localization of precursor and
mature TGF-β1+ and TGF-β2+ cells. The most prominent expression of TGF-β was
visible in the epidermal basal layer (Fig. 4.59 A) and in the ORS of HFs (Fig. 4.59
B). In addition, other epithelial parts also showed homogenously TGF-β1 IR like the
suprabasal layers of the epidermis, the epidermis in the foot pad and the PNM.
Moreover, we found IR, although with less intensity, in sweat glands and single other
dermal cells.
Fig. 4.59 IR in the murine nail apparatus for TGF-β1 Expression pattern of TGF-β1 in normal murine nail apparatus (brown). (A) The epidermal basal layers (A) and
Outer root sheath of the HFs are intensely stained for TGF-β1. HF: Hair follicle, NP: Nail plate, PNF: Proximal nail fold, PNM: Proximal nail matrix, Bars: (A-D) 200µm.
Fig. 4.60 Schematic drawing of TGF-β1 expression pattern Schematic drawing of the TGF-β1 expression pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
177
4.4.5 Demonstration of CD54 molecules
By using avidin-biotin-complex staining, we observed that cells positive for
intracellular adhesion molecule 1 (ICAM-1) were limited to the mesenchyme (Fig.
4.61 A), except for single positive ICAM-1+ cells in the hair follicle ORS (data not
shown). Most of the ICAM-1 positive cells could easily be detected in endothelial
cells (Fig. 4.61 B) of small blood vessels near the epidermis of the PNF, whereas the
number of positive cells was slightly reduced in the vicinity of the PNM and the foot
pad. The nail bed showed only rarely contained ICAM-1+ cells (Figs. 4.61, 4.62).
Fig. 4.61 IR in the murine nail apparatus for CD54 Immunohistological identification of CD54 expression (brown). NP: Nail plate, PNF: Proximal nail fold, PNM: Proximal nail matrix, Bars: (A) 100µm, (B) 20µm.
Fig. 4.62 Schematic drawing of CD54 expression pattern Schematic drawing of the CD54 expression pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
178
4.4.6 Demonstration of mast cells
In order to detect mast cells, again, toluidine blue staining was performed. We found
no significant differences of mast cell numbers in the vicinity of the PNM, compared
to the PNF (Fig. 4.63 A) or the foot pad. This was confirmed by quantitative
immunohistochemistry. Interestingly, the nail bed displayed a significantly reduced
number of mast cells than the PNF (*p<0.05) (Fig.4.64).
Fig. 4.63 IR in the murine nail apparatus for mast cells Immunohistochemical identification of mast cells in normal murine nail apparatus (purple). (A) Mast cells are uniformly distributed in the vicinity of PNF and PNM and foot pad. NP: Nail plate, PNF: Proximal nail fold, PNM: Proximal nail matrix, Bars: (A) 100µm, (B) 20µm.
NP
PNM
PNF
A
B
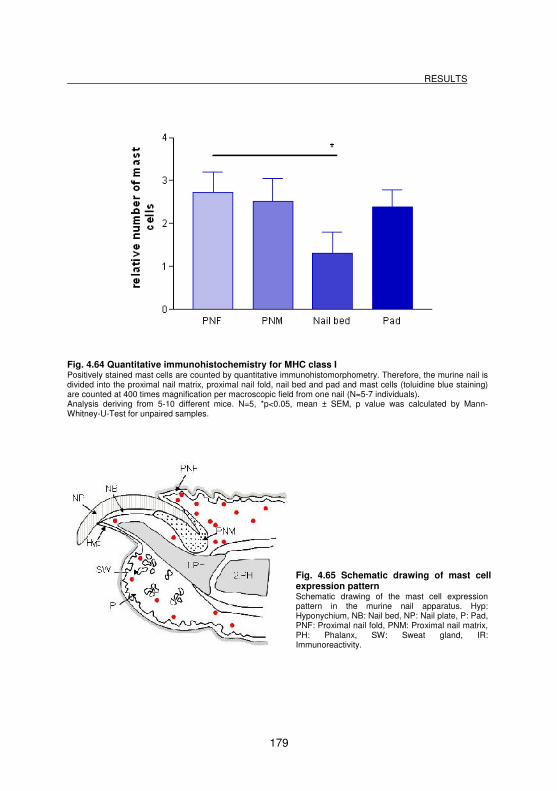
RESULTS
179
Fig. 4.64 Quantitative immunohistochemistry for MHC class I Positively stained mast cells are counted by quantitative immunohistomorphometry. Therefore, the murine nail is divided into the proximal nail matrix, proximal nail fold, nail bed and pad and mast cells (toluidine blue staining) are counted at 400 times magnification per macroscopic field from one nail (N=5-7 individuals). Analysis deriving from 5-10 different mice. N=5, *p<0.05, mean ± SEM, p value was calculated by Mann-Whitney-U-Test for unpaired samples.
Fig. 4.65 Schematic drawing of mast cell expression pattern Schematic drawing of the mast cell expression pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

RESULTS
180
4.4.7 Demonstration of β-defensin 2
We investigated the local expression of precursor and mature β-defensin 2 producing
cells in the murine nail apparatus. A strong up-regulation of staining intensity
appeared in parts of the
PNM close to the nail plate
compared to the PNF and
parts of the foot pad (Fig.
4.66). The staining was
cytoplasmic and a descreet
IR could also be shown in
the epidermal basal layers of
the PNF and pad (Figs. 4.66,
4.67).
Fig. 4.66 IR in the murine nail apparatus for β-defensin 2 Immunohistochemical identification of β-defensin 2 in normal murine nail apparatus (brown). (A) Staining intensitiy is in the PNF reduced compared to dorsal parts of the PNM (B). NP: Nail plate, PNF: Proximal nail fold, PNM: Proximal nail matrix, Bars: (A,B) 100µm.
Fig. 4.67 Schematic drawing of β-defensin 2 expression pattern Schematic drawing of β-defensin 2 expression pattern in the murine nail apparatus. Hyp: Hyponychium, NB: Nail bed, NP: Nail plate, P: Pad, PNF: Proximal nail fold, PNM: Proximal nail matrix, PH: Phalanx, SW: Sweat gland, IR: Immunoreactivity.

DISCUSSION
181
5 Discussion
5.1 Introductory remarks
Recent gene profiling data in mice have raised the possibility, that follicular IP is not
purely limited to the anagen hair bulb, but that the epithelial stem cell region in the
ORS of HFs, termed the bulge, may also represent an area of relative IP. Since
protection of these epithelial stem cells from immune destruction is essential for
preserving the regenerative and cycling capacity of the follicle, it seems logical that
the bulge region should also have an established relative IP. However, convincing
protein evidence for this idea is still missing. In the current study, we aimed to
support or refute the hypothesis of bulge IP in human HFs.
By using highly sensitive immunohistochemistry, including Immunofluorescence, we
provide the first comprehensive in situ-protein expression evidence, which follows-up
leads from gene profiling data that suggests, contrary to previous assumptions, that
the HF bulge fully meets classical criteria for immune privileged tissues: MHC class
Ia, β2-microglobulin and MHC class II immunoreactivity (IR) were found to be down-
regulated in the human bulge. The immuno supressants α-melanocyte stimulating
hormone (α-MSH), transforming growth factor-β2 (TGF-β2), macrophage migration
inhibitory factor (MIF) and indoleamine-2,3-dioxygenase (IDO) are upregulated in the
CD200+, stem cell-rich bulge region. In addition, these CD200+ cells also co-express
HLA-E.
Our systematic bulge immuno phenotyping experiments were complemented with a
functional study, which should determine, whether IFN-γ, a recognized key inducer of
IP collapse in human anagen hair bulbs, had a similar effect on the putative bulge IP.
We showed a significant ectopic upregulation of MHC class Ia in bulge cells of organ-
cultured human scalp skin. By this skin organ culture assay other IP markers can be
evaluated in future experiments, particularly substances to restore IP after IFN-γ
induced collapse.

DISCUSSION
182
In the current study, the effects of the α-MSH related tripeptide K(D)PT were
explored with our organ-culture HF model, and it was demonstrated that it failed to
act as an agent to protect IP or to re-establish IP. Nevertheless, it revealed many
other interesting effects on HF biology (e.g. melanogenesis, mast cells, hair cycle),
suggesting further investigation.
Moreover, we contributed substantially to the immunological characterization of the
murine vibrissal follicle and murine nail apparatus and showed that the vibrissal bulb,
and in a less extent the bulge and proximal nail matrix, can be considered to be also
immunoprivileged to a certain. However, many gaps in the immunobiology of the
murine vibrissal follicle and nail apparatus remain to be closed in future studies.
5.2 Methods employed
Tissue specimens:
In the current study, normal human fronto-temporal and occipital uninflamed scalp
skin was obtained from females only, undergoing routine face-lift surgery after
informed consent. By choosing samples from the ´female` sex only, the influence of
hormonal differences was prevented. The patients were supposed to have no pre-
treatment and did not take any drugs to exclude any non-biological influences,
although a guarantee did not exist. The donors were selected randomly (aged 45-67
years, mean age 52.5 years) and all experiments were performed according to the
Helsinki guidelines.
For the in situ protein immunohistochemistry of the bulge, scalp skin of 16 patients
was investigated, which was obtained from both regions of the skin (occipital and
fronto-temporal). However, no region-related or age-dependent differences in the IR
pattern of the markers were obvious. Per antigen, 3-10 cryosections per patient of at
least 5 different individuals were examined. This representative amount of patients
substantially limited the possibility of inter-individual differences, which always also
occur within a population. For the isolated HF organ culture, only occipital human

DISCUSSION
183
scalp skin from six donors (age range: 46-65 years) was used. Per experiment the
isolated HFs from one donor were randomly and uniformly distributed into the six
different test- and control groups, so that interindividual differences between the test
groups could be excluded. Despite this aspect, we found interindividual differences
when comparing the experiments. Per test group normally 18 HFs were cultured,
taking into account losses associated with the treatment or with the cutting, so that a
statistical evaluation was possible. Two experiments started with a number of 12 HFs
per group, due to a limited availability of HFs in the sample. For the full-thickness
skin organ culture, scalp skin of different regions from two patients was cultivated
(age 53 and 63 years). Again, no age- or patient-related difference, as well as no
region-related difference in the IR pattern of the staining could be observed.
Important for this culture was the appropriate size of the skin sample, i.e. in case that
the size was too large, apoptosis in the skin specimens occurred, because the
medium could not penetrate into the centre of a sample.
In the cooperating hospitals, scalp specimens were placed immediately after surgery
into supplemented William’s E serum-free medium, and transported overnight to the
laboratory at a temperature of 4°C. One critical point of specimen collection was the
time period between tissue removal and its immersion into tempered William’s E
medium. The optimal procedure is to place the sample into the medium immediately
after removal, but since the hospitals were widely distributed over Germany, it was
not possible to take the samples oneself. A second critical point was to keep the
specimen refrigerated at a temperature of 4°C during the transport overnight (only
one hospital in Lübeck delivered scalp skin on the same day of surgery). High
outdoor temperatures might have influenced tissue quality, although the samples
were transported on ice packs, it could not be checked whether the material was sent
well tempered during transport. After arriving in our laboratory, nevertheless, the skin
was immediately processed for the in situ immunohistochemistry experiments, for HF
organ culture or for full-thickness skin culture. Despite all critical aspects that could
have affected tissue quality, we ensured that no negative influence on tissue and
reaction quality was observable light microscopically during the course of our study.

DISCUSSION
184
Mouse tissue samples were taken from 18 control C57/BL6 and Balb/c mice from
other cooperating working groups on the campus. Specimens were collected
immediately after cervical dislocation to minimize autolysis. For obtaining a
representative result, 3-10 cryosections per patient of at least 5 different individuals
were examined per antigen. Although we collected samples of different mouse
strains (C57BL6 or Balb/c), which had a different immunological background and
susceptibility for autoimmune diseases, we could not detect any difference in the
pattern of the IR staining. The same observation was made for possible age-related
differences or anatomical differences between the fore- and the hind limb.
Fixation:
The routine fixation method was to immerse the sample into Shandon Cryomatrix,
followed by snap-freezing in liquid nitrogen and storing at -80°C until use. In contrast
to fixation in Bouin’s solution, this method is supposed to be less efficient. Bouin’s
fixation has the advantage that the fluid penetrates faster into the tissue and
therefore produces good protein conservation and tissue morphology excluding also
shrinkage artefacts. Most experiments were based on cryosections, because the
experimental laboratory was specialized in such methodical approach, including the
fact that there was much experience available for the use of anti-mouse antibodies
on cryosections. Furthermore it is important to note, that the freezing technique had
the advantage in the Cryomatrix embedding step compared to Bouin fixation with
paraffin embedding: All specimens could be accurately placed in an optimal angle
into the cups, so that longitudinal or sagittal cuts became possible. Furthermore, the
possibility of adjusting the cutting level (angle) facilitates the production of
longitudinal sections. For future research it should be considered to use the more
rapid freezing technique at lower temperatures.
Performing cryosections:

DISCUSSION
185
We took the rather common way of processing cutaneous tissue and carried out
vertical cuts, showing the epidermis at one pole and the hypodermis at the other. The
aim was to obtain as many longitudinal scalp HF and vibrissal sections, respectively
sagittal sections of the murine nail apparatus, as possible. Since the follicles do not
grow in the same angle or parallel, it was difficult to receive longitudinal sections from
the bulb to the epidermis. Cutting the organ cultured punch biopsies was particularly
difficult, since they were of such a small size (4 mm diameter) and often contained
two or three HFs only. If a section on a the isthmus level was complete in one HF,
and another HF of the follicular unit next to it showed a complete lower HF, the skin
sample was taken into evaluation. An additional aspect which had to be taken into
account, was the fact that IR of several antigens were shown to be hair cycle
dependent. Since IP in the bulge or hair bulb is only present in anagen VI, it was
important to ensure, that only HFs and vibrissal follicles in anagen were evaluated.
Therefore, the best way to differentiate was to investigate complete longitudinal
follicles, or, at least, to obtain the lower HF to examine the shape and structure of the
hair bulb and DP (STENN and PAUS 2001; WHITING 2004) to exclude HFs of a
different cycling stage from evaluation.
Despite the fact, that it was not sure at the beginning of the experiments, whether the
bone tissue of the phalanges could be cut with a normal cryostat blade, we learned
that it was possible to obtain sections of good quality without decalcification and with
an optimal cutting technique. Sometimes the bone structure was slightly disrupted,
but this fact did not affect the staining quality or evaluation. In contrast, in Bouin-
fixated samples a step of decalcification with 25% EDTA was included.
Immunohistochemistry and evaluation:
In this work not only standard immunohistochemistry, including Immunofluorescence
was employed, but also highly sensitive visualization methods (EnVision®, TSA
immunofluorescence) (see e.g. ROTH et al. 1999; ITO et al. 2004b, 2005c; BODO et
al. 2005; GAISER and BERNHARDS 2007). Since previous studies had shown
contrary results compared to the recent gene profiling data, these special sensitive

DISCUSSION
186
methods should provide more reliable results. Regarding the nail apparatus, we
observed that ABC methods based on alkaline phosphatases cross-reacted
unspecifically with the bone tissue. For this reason, all stainings based on avidin-
biotin-complex method were performed with horseradish peroxidase. In general, it
was also found, that the substrate AEC could not be reliably used on murine tissue.
Sometimes no staining was seen, even though DAB provided good results. Whether
this observation is due to the staining kit (although different ones were used) or to the
special tissue, could not be clarified. In addition, Masson Goldner staining could not
be performed on cryosections, due to the fixation method, which leads to a reduced
staining intensity of epithelial tissue. As a consequence most overview stainings were
performed with HE.
Furthermore, MHC class Ia staining on murine nail apparatus specimens revealed
contradictory results, although positive controls worked. On cryosections, sometimes
parts of the epidermis in the PNF were stained, sometimes not. The PNM has not
produced any positive staining and the epithelium of the foot pad was often negative
for MHC class I IR, although dermal cells were stained. In contrast, Bouin-fixed tissue
showed a homogenous staining all over the epithelium (including the PNM, PNF and
foot pad). However, region-related, age-or sex- or breed-related differences in the IR
pattern of the staining could not be detected.
5.3 Immune privilege in the human hair follicle bulge, murine nail
and sinus hair follicle
5.3.1 The human HF bulge
The aim of the current study was to show by immunohistochemistry and organ
culture, whether the human anagen hair bulb fully meets the classical criteria for
immune privileged tissues, as recently shown in gene profiling data (MORRIS et al.

DISCUSSION
187
2004; TUMBAR et al. 2004; OHYAMA et al. 2006), or whether previous contradictory
assumptions on the protein level are correct.
Since downregulation of MHC class Ia IR is one hallmark of IP, it was very important
to verify its exact and distinctive expression pattern in the human HF. HARRIST et al.
(1983) published the first MHC class Ia antigen expression data in normal terminal
human HFs, reporting prominent IR in the infundibular region of the ORS with
diminished expression of HLA-A, B, C and β2-microglobulin in the isthmus region,
whereas the bulge was not referred to. Later CHRISTOPH et al. (2000)
demonstrated strong HLA-A/B/C IR and β2-microglobulin IR in the ORS isthmus
region of the ORS, though possibly at a somewhat reduced level of IR compared to
the distal ORS (CHRISTOPH et al. 2000). They concluded, that the anagen hair bulb
was the only site of IP within the HF (PAUS et al. 2003), which became generally
accepted. However, with the recent gene and profiling data, this theory had to be
abandoned. We demonstrated a convincing and significant downregulation of MHC
class Ia and β2-microglobulin in the bulge region, compared to the isthmus and
epidermis (Figs. 4.1, 4.6), by using high sensitive immunohistochemistry. Intriguingly,
we could also present a difference in MHC class Ia IR between the bulge and the
bulb.
There are several aspects which might explain why a bulge IP had not been
suggested before: 1) At that time, object of interest was to investigate the
pathogenesis of AA, with the IP collapse model in the pathogenesis of AA (PAUS et
al. 2003, 2005), other HF disorders were not discussed. 2) The bulge was ignored as
a prominent and important structure or as a reservoir of eSCs in the HF. Originally, it
was thought that stem cells mainly reside in the hair bulb and have to be protected
(PAUS et al. 1999b; PAUS and COTSARELIS 1999). It was not considered that an
attack of the bulge eSCs could cause irreversible HF damage. 3) The bulge is not
easily detected as a part of the HF, so that detailed analyses remained inconclusive.
Since the characterization of the bulge by KLOEPPER et al. (2008), with the finding

DISCUSSION
188
of the leading structure ´follicular trochanter` and the insertion point of the APM, this
structure became more distinctive.
In addition, we confirmed a significant downregulation of antigen-presenting functions
of intraepithelial dendritic cells (seen as downregulation of MHC class II expression)
(Figs. 4.9-4.11). These finding was expected, since CHRISTOPH et al. (2000) had
already shown significantly reduced numbers of MHC class II+ cells in the isthmus
and proximal ORS compared with the distal ORS, although again, no detailed
analysis of the bulge had been carried out. It is important to note, that this
downregulation much more severely hampered antigen presentation in the bulb
compared to the bulge, as also true for the MHC class II-dependent antigen. Here,
very few CD1a positive or ultrastructurally detectable Langerhans cells were clearly
MHC class II-negative, while the number of dendritic cells in the bulge region still
displayed some MHC class II expression, albeit the fact that their number was
substantially reduced.
We could detect an upregulation of non-classical MHC class Ib molecules, which has
previously been shown to be important in the IP maintenance of other systems, like
the fetotrophoblast (MOSCOSO et al. 2006; YIP et al. 2006), the eye (NIEDERKORN
et al. 1999; LE DISCORDE et al. 2003), and the proximal nail matrix (ITO et al.
2005c). We focused on HLA-E, since its expression in human skin has not been
extensively studied until now. In addition, its effect is more directly than, for example,
the one of HLA-G. HLA-E is a ligand of the NK cell inhibitory receptor CD94/NKG2A
and may result in inhibition of NK cell- and cytotoxic T cell-dependent lysis (GAO et
al. 1997; BRAUD et al. 1998; PACASOVA et al. 1999; TRIPATHI et al. 2006),
although HLA-E is also regarded to interact with CD8+ T cells directly (GAO et al.
1997). In our study, we observed a mesenchymal-facing intracellular location of HLA-
E protein in CD200+ keratinocytes within the bulge (Figs. 4.12., 4.13), which
corresponds to a previous report of HLA-E sequestration to the endoplasmic
reticulum in other tissues (ULBRECHT et al. 1992), and suggests that HLA-E is
stored in the bulge ORS, ready to be transported to the cell surface upon recruitment.

DISCUSSION
189
The role of the immunosuppressants has long been underestimated, although recent
studies showed that α-MSH suppresses experimental autoimmune
encephalomyelitis, and that α-MSH and TGF-β can promote re-establishment of
immune tolerance to autoantigens in the brain (TAYLOR and KITAICHI 2007) and
the human anagen hair bulb (PAUS et al. 2003; PAUS et al. 2005; NIEDERKORN
2006). TGF-β1 for example, is prominently expressed in the HF epithelium of mice
and humans, and shows its highest IR during late anagen and the onset of catagen
in cells of the ORS and epithelial strand (FOITZIK et al. 2000).
Our finding of the upregulation of the potent, locally generated “IP guardian” α-MSH
in the bulge (Fig. 4.14), along with strong IR of the immunosuppressant TGF-β2 (Fig.
4.15), probably contributes to the maintenance of bulge IP in a manner similar to that
recognized in the eye, brain and fetal IP (WAHL et al. 2006; SIGLIENTI et al. 2007).
TGF-β is thought to inhibit NK cell-mediated cytoloysis, contributing to the
mechanism of IDO (APTE and NIEDERKORN 1996; APTE 1997; APTE et al. 1998;
NIEDERKORN 2003, 2006), and influences the behavior of APCs in promoting
differentiation of Ag-specific T cells into CD4þCD25þFoxp3þ regulatory T cells
(Tregs) (D`ORAZIO and NIEDERKORN 1998; TAKEUCHI et al. 1998; KEZUKA and
STREILEIN 2000; STREILEIN et al. 2002).
We also present the first in situ immunohistochemical evidence of IDO expression in
human HFs and demonstrated an upregulation of IDO in the bulge region (Figs.
4.20-4.22), that underscores previous observations made from other
immunoprivileged sites [(e.g. lung or allografts (SWANSON et al. 2004; LI et al.
2006); skin (VON BUBNOFF et al. 2004), anterior chamber of the eye (CHEN et al.
2007) and fetus (MELLOR and MUNN 2008)]. IDO is the rate limiting tryptophan-
catabolizing enzyme, which limits T cell proliferation by creating a tryptophan-
depleted environment, and also affects NK cell function, reduces inflammation and
enhances Fas-mediated T cell apoptosis through their tryptophan degradation
products (MUNN et al. 1998; FALLARINO et al. 2002; MELLOR and MUNN 2008).
Even though T cells (CD4+ or CD8+ cells) are predominantly distributed in the distal
HF epithelium, a small number of T cells and NK cells exists in and around the

DISCUSSION
190
human HF bulge (CHRISTOPH et al. 2000; ITO et al. 2007). In addition, since every
CD8+ and NK cell could endanger or attack the ORS bulge cells upon their MHC
class Ia negativity, IDO might be vital in preserving HF immune integrity and keeping
these cells inactivated.
In addition, in the light of the presented upregulation of MIF in the human HF bulge,
which is expressed in several IP sites such as the fetotrophoblast (VIGANO et al.
2007), eye (APTE et al. 1998), brain, human proximal nail matrix (ITO et al. 2005c),
and human anagen scalp HFs (ITO et al. 2007), the bulge IP concepts also gains
importance. We confirmed the previously reported MIF IR pattern in human
epidermis (SHIMIZU et al. 1996; ITO et al. 2007), and were able to show an
increased MIF IR in the bulge region, proximal of the ORS and IRS of human HFs
(Figs. 4.18, 4.19). MIF may inhibit NK-cell mediated cytotoxicity against MHC class I-
negative cells by preventing the release of cytolytic perforin granules from NK cells
(APTE et al. 1998; ARCURI et al. 2006). This would explain why immune-privileged
tissues, that are highly vulnerable to lysis by NK cells due to their lack of MHC class I
expression, are not under constant NK cell attack. By comparative studies of human
scalp skin from healthy donors and patients with AA, it has been already shown that
the epithelium of lesional AA HFs displayed a greatly reduced or absent MIF IR. This
observation suggests that HFs in established AA have a decreased capacity for
suppressing undesired NK cell functions.
We could confirm the previously demonstrated strong upregulation of the ´no
danger`-signal CD200 (ROSENBLUM et al. 2004; OHYAMA et al. 2006; KLOEPPER
et al. 2008), that inhibits pro-inflammatory cytokine release of CD200 receptor+ mast
cells, downregulates APC activity (ROSENBLUM et al. 2006), and is thought to
influence IDO production (FALLARINO et al. 2002). As recently reported (OHYAMA
et al. 2006; KLOEPPER et al. 2008), CD200 is present in the outermost ORS layer,
between the insertion of the APM and the insertion of the SG duct [note that this
region is classically defined as isthmus (WHITING 2004), while only its proximal end
includes the bulge region]. This cell surface protein is upregulated in the human

DISCUSSION
191
bulge, although not restricted to it, and therefore does not qualify as a specific human
epithelial HF stem cell marker (KLOEPPER et al. 2008). But for our purpose the
expression pattern was distinct enough, and because of its immunomodulatory
functions contributing to IP, we used CD200 for double Immunofluorescence
stainings with MHC class I and HLA-E to confirm the IR of both antigens within the
bulge.
In order to obtain functional evidence, some experiments were run with our
established full-thickness human skin culture. It was shown that the putative bulge IP
is subject to the same IP collapse-inducing pro-inflammatory stimulus (IFN-γ) (Fig.
4.25) as the bulb IP in mice and humans (RUECKERT et al. 1998; ITO et al. 2004),
providing the strongest evidence available so far, that the human bulge also
represents an area of relative IP. Interestingly, more than ten times higher doses of
IFN-γ were required to induce a statistically significant collapse in the bulge than in
the bulb (1000 IU/ml vs. 75 IU/ml). This could be a result of a different experimental
design: ITO et al. (2005) used the isolated HF organ culture and 75 IU/ml IFN-γ to
induce an ectopic upregulation of MHC class Ia IR in hair matrix keratinocytes,
whereas higher doses like 500 or 1000 IU/ml IFN-γ induced catagen transformation
in the HFs. In contrast, to explore the bulge region regarding its effects on IFN-γ, it
was necessary to use the full thickness organ culture model (LU et al. 2007) (for the
isolation of HFs, the epidermis and bulge region is cut off). Since the specimens were
much larger than isolated HFs, including a longer penetration distance, it could be
expected that much higher doses of agent had to be added.
However, our immunohistochemical study also suggests that the bulge and bulb IP in
human scalp HFs differ in several important aspects: The level of constitutive MHC
class Ia, as associated with β2-microglobulin and MHC class II protein expression in
the bulge, is much higher than in the bulb (Figs. 4.3, 4.8, 4.10). Since β2-
microglobulin protein expression is virtually absent in the human anagen bulb
(CHRISTOPH et al. 2000; ITO et al. 2004), but not in the bulge, in theory, leaving
unclear how the rest of HLA-A/B/C molecules can be stabilized here (and thus

DISCUSSION
192
rendered fully functional by association with β2-microglobulin). This suggests that the
presentation of MHC class I-dependent autoantigens, virally encoded or tumor
antigens, is still possible in the bulge region of the ORS (albeit its significant
reduction compared to other ORS regions), while this presentation is almost
impossible under physiological conditions in the anagen bulb. Whether such
observation indicates a more labile relative IP in the bulge than in the bulb or not,
cannot be assessed easily. The state of IP is a complex of different mechanisms
(e.g. IDO, TGF-β or MIF), which are upregulated in the bulge and might reduce the
risk of IP collapse. Furthermore, for such an important site as the bulge with its eSCs,
it might be necessary to have a capacity of defense via the immune system in case
of inflammation.
Taken together, even though this concept of bulge IP remains to be supported by
additional functional evidence, the in situ protein expression data presented here
emphasize that the bulge in fact has a strong - and perhaps stronger - immune
protection than that of the anagen bulb. IDO and MIF showed higher IR levels around
the bulge compared to the proximal follicle, and CD200 and HLA-E were solely
expressed in the bulge, having both marked immunomodulatory functions. These are
likely to be the key aspect in protecting the bulge and the stem cells from immune
attacks, not only in anagen but throughout the complete hair cycle. Supportive
evidence for this finding is provided by ROSENBLUM et al. (2006), who showed that
CD200-/-skin grafts quickly develop permanent alopecia when grafted onto wild type
mice.
On the other hand, MHC I, MHC II and β2-microglobulin IR levels were not as low as
those in the hair bulb, which may indicate that the bulge IP is constitutively more
prone to inciting an immune response than the bulb, resulting in an inflammatory cell
attack on bulge epithelial stem cells (COTSARELIS and MILLAR 2001). The latter
aspect is the most characteristic histological feature of cicatricial alopecia. However,
from a clinical point of view, alopecia areata (AA) (the prototypical bulb IP collapse
model) is far more common than any of the primary cicatricial alopecias (PCA). It
seems logical that, if PCA were the ´clinically visible consequence of bulge IP failure`

DISCUSSION
193
and the bulge more susceptible to IP collapse, we should see far more cases of PCA
than actually done. Furthermore, the required IFN-γ dose to induce IP collapse within
the organ culture was much higher than in the bulb (ITO et al. 2004), suggesting that
the bulge IP is less susceptible to IP collapse. This theory is corroborated by
evidence from molecular studies that show a down-regulation of INF-γ receptors in
the bulge (MORRIS et al. 2004).
5.3.2 The murine sinus hair follicle and nail apparatus
In order to immunologically characterize murine vibrissal follicles and nail apparatus,
and to get a basic understanding if parts of these tissues also might have the benefit
of relative IP, we focused on a small amount of IP marker and some general
immunological parameters. Although MHC class Ib, MIF, IDO and CD200 are
important actors in IP, we waived investigating these antigen IR on murine tissue due
to temporary and economical reasons. In addition, in the case of MIF and IDO for
example, cross reaction of the anti-human Abs with the mouse tissue impeded its
usage. Moreover, concerning the murine specimens, we focused on MHC class Ia
expression only and did not analyze the β2-microglobulin expression pattern,
because it was likely to parallel the findings for MHC class Ia, with which it is
associated. Inspection of human HFs was more important, since a lot of data on this
matter have been published before (HARRIST et al. 1983; CHRISTOPH et al. 2000)
and recent microarray data have claimed a β2-downregulation (MORRIS et al. 2004;
TUMBAR et al. 2004).
Since MHC class Ia IR is one of the striking features of IP (JANEWAY et al. 2005;
NIEDERKORN and WANG 2005; PAUS et al. 2005), we analyzed the MHC class Ia
expression pattern. Staining for MHC class I showed homogenous IR in the
epidermis and in the distal ORS of the isthmus and infundibulum (Figs. 4.35, 4.37).
At the level of superior swelling, IR started to decrease (Fig. 4.35 B) and was even
more and significantly reduced in the bulge region or the inferior swelling of the ORS

DISCUSSION
194
(Fig. 4.34 C). Interestingly, in these hourglass shaped parts, the basal layers of the
ORS were almost devoid of IR. And almost no staining intensity was found in the
proximal ORS or the follicle bulb (Fig. 4.34 E). Thus, in the vibrissal follicle the
expression pattern was similar to the human HF, apart from structural differences.
The negativity in the basal layers of the bulge is different from the human HF and
could be important to reduce immune regulation, because the basal layers are facing
to the blood sinus and environment. But since a residual level of MHC class Ia+ cells
is still presented in the other layers of the ORS, they could be activated in case of
need. Otherwise, MHC class I negativity may sequester (auto) antigens from
presentation to CD8+ T cells and NK cells.
We also demonstrated that the nail matrix epithelium, and most prominently the
PNM, displayed substantially downregulated MHC class I IR, compared with PNF,
nail bed, and hyponychium (Fig. 4.51), as it was also shown for the human nail
apparatus (ITO et al. 2005c). This low or absent MHC class I expression in nail
matrix epithelium was mirrored in the immediately adjacent nail mesenchyme (Fig.
5.52). The pad epithelium was only weak or not stained. The absence of MHC class I
in the PNM compared to the PNF, suggested the PNM to be also immunoprivileged.
On the one hand, it is not fully comprehensible, why the epithelium of the foot pad
also displayed a reduced IR. One possible explanation could be that the foot pad
epithelium is the part, on which the mice weigh themselves down, thus, the stratum
corneum is more distinctive and acts as the main barrier against pathogens, water,
and pressure. On the other hand, another explanation could be that our staining
problems influenced the result. As mentioned before, we obtained varying results,
although the positive controls (or the vibrissal follicle) worked. On cryosections, parts
of the epidermis in the PNF sometimes were stained and sometimes not. The PNM
has never demonstrated positive staining, and the epithelium of the foot pad was
often negative for MHC class I IR, although fibroblasts / fibrocytes in the dermis were
stained. In contrast, Bouin-fixed tissue showed a homogenously staining of the entire
epithelium (including the PNM, PNF and foot pad). It would be easy to suggest that
staining of the cryosections did not work at all. But the fact that we obtained
convincing staining results in normal murine back skin and snout / vibrissal follicles

DISCUSSION
195
contradicts this hypothesis. Because of the lack of data in the literature and despite
the exchange of information with international experts in this field, it was difficult to
decide which IR pattern was reliable. Given that the PNM is indeed negative for MHC
class Ia IR, it could be possible that the PNM is another site of IP.
In accordance with the findings in the human bulge (MEYER et al. 2008a), we could
demonstrate a significant downregulation of antigen-presenting functions of
intraepithelial dendritic cells (seen as downregulation of MHC class II expression) in
the vibrissal bulb, bulge and PNM. These findings are in line with the theory of IP
analogue to the HF (MEYER et al. 2008a) or human nail apparatus (ITO et al.
2005c). The MHC class II expression was particularly dense in the epidermis, in the
infundibulum and upper parts of the isthmus (Fig. 4.38), whereas the superior and
inferior swelling of the ORS exerted significantly reduced MHC class II+ cells. Similar
to the human HF, lower parts revealed an even decreased number of MHC class II+
cells towards the hair bulb, and only single MHC class+ cells were present in the DP.
In the murine nail matrix, MHC class II+ cells were found in high numbers in the
epidermis and in adjacent PNF (Fig. 4.54 A), whereas no MHC class II+ cells were
detectable in the PNM and in its mesenchymal vicinity (Fig. 4.54 B). The nail bed
only rarely showed MHC class II+ cells (Fig. 4.54 B). In the foot pad epithelium, the
greatest number of intraepithelial MHC class II+ cells could be identified (Fig. 4.54
D), whereas the dermis showed a reduced number compared to the PNF and PNM
dermis (Figs. 4.55, 4.56). We focused on the detection of surface MHC class II
molecules, since their downregulation correlates with the suppressing of the APC
capacity, independent of the cell type (LC, macrophages, DC). The results of the
CD11b IR in the vibrissal follicle and the nail apparatus underlines the MHC class II
findings and reveals that most of the MHC class II positive cells were macrophages.
Nonetheless, in future experiments it would be interesting to perform double
stainings, as ITO et al. (2005) have done to distinguish APCs in detail, and also to
evaluate how many APC are localized in the different parts and which percentage is
impaired of their antigen-presenting capacity (seen as downregulation of MHC class
II+ cells on the surface).

DISCUSSION
196
As demonstrated for the human HF (CHRISTOPH et al. 2000), we also detected that
CD4+ T cells and CD8+ T cells (data not shown) were predominantly expressed in
the epidermis and upper parts of the vibrissal follicle (in particular in the isthmus and
around the sebaceous gland), whereas in the superior swelling of the ORS and even
more in the bulge, a significantly reduced number of this cell type was obvious (Fig.
4.39). The bulb region showed only single CD4+ T cells (Fig. 4.39) and no CD4+ T
cells in the dermal papilla. The same observation was made for CD8+ T cells, even
though their total number was lower (data not shown). This similarity is also true for
the murine nail apparatus compared to the human system. As known from the human
nail apparatus (ITO et al. 2005c), we could identify a high density of CD4+ T cells in
the vicinity of the PNF, but very rarely in adjacent PNM. Intraepithelial CD4+ T cells
were only present in the PNF and foot pad. This absence or downregulation of CD4+
T cells may contribute to an immunoprivileged environment since the mechanism of
antigen presenting is also dependent on CD4+ T cells and co-stimulatory signals. In
addition, a low number of CD8+ T cells could reduce the risk of an attack on cells
without MHC class I surface molecules to protect the stem cells.
We demonstrated a similar expression pattern of the immunosuppressant TGF-β1, in
both the vibrissal follicle (Fig. 4.45) and the nail apparatus (Fig. 4.59) compared to
the human HF. In the vibrissal follicle, the bulge exerts, when compared to the SO
and lower parts of the follicle, an upregulated TGF-β1 IR, indicating a special function
of TGF-β1. In the nail apparatus, we found homogenously TGF-β1 IR (i.e. the
suprabasal layers of the epidermis, the epidermis in the foot pad and the PNM),
besides prominent expression of TGF-β in epidermal basal layers and in the ORS of
HFs (Fig. 4.59 B). As already described recently (WAHL et al. 2006; SIGLIENTI et al.
2007), TGF-β1 is considered to play an important role in the maintenance of IP by
inhibiting NK cell-mediated cytolysis and CD8+ T cell attacks, in particular, in areas of
low or absent MHC class Ia expression. The aforementioned findings suggest that
besides these and other IP-associated effects, TGF-β1 somehow influences and
supplies cells and tissues in another way, because it is so widely expressed.

DISCUSSION
197
Interestingly, according to the findings in the human HF, the matrix keratinocytes in
the bulb revealed a reduced staining intensity. This indicates that the assumed IP
within the bulb either is not as strong as in the bulge, or that the role of TGF-β1 in the
bulb is not really essential, because MHC class I negativity persists.
In addition to these antigens directly associated with IP, we also investigated some
other key immunological parameters on the mouse specimens: Mast cells, CD54 and
β-defensin 2: Mast cells are multifunctional and involved in many different
interactions of innate and adaptive immunity (YOUNG 1997; MCLACHLAN et al.
2003; MEKORI 2004; SUTO et al. 2006; THEOHARIDES and KALOGEROMITROS
2006; CHRISTY and BROWN 2007; METZ et al. 2007; JAHANYAR et al. 2008;
SAYED et al. 2008). They are also considered to contribute to IP (DEPINAY et al.
2006; SAYED et al. 2008), and we aimed to define the expression of mast cells
pattern in murine specimens.
Contrary to previous findings in the human HF (CHRISTOPH et al. 2000), where
mast cells were distributed homogenously all over the CTS of the human HF, we
could show a compartment-dependent distribution of mast cells in the vibrissal
follicle. Here we found that the number of mast cells was especially high in the upper
part and the isthmus, and decreased continuously towards the lower part, whereas
almost no mast cell was evident around the vibrissal bulb (Figs. 4.48 A,B, Fig. 4.49).
However, no significant difference in mast cell numbers could be demonstrated within
the hourglass shaped parts and their surrounding tissue, whereas many mast cells
were localized outside the capsule around the vibrissal follicle.
On the one hand, such decreased mast cell numbers towards the bulb suggest a
contribution of mast cell interaction to the maintenance of IP in the vibrissal follicle,
although the mechanisms are still unclear. Since it is supposed that mast cells play a
role in the pathogenesis of AA, which is based on hair bulb IP collapse, this idea
would underline our findings in mice. On the other hand, it cannot be fully explained,
why the mast cell results strongly differ between the murine vibrissal follicle and the
human HF. An explanation would be that the vibrissal capsule is the cause of the
difference and acts as a strong barrier against the invasion of mast cells from the

DISCUSSION
198
surrounding. Furthermore, it is unknown so far under which circumstances mast cells
reveal pro-inflammatory or immunosuppressive functions, or when they act as ´good`
or ´bad guys` (MAURER et al. 2003b; MAURER and METZ 2005). For this reason, it
is also important to keep in mind that not only the mast cell numbers but also the
activation status or ratio of degranulated mast cells compared to non-degranulated
have to be considered. Either the previous findings of CHRISTOPH et al (2000) were
not distinctive enough, or the ratio of degranulation is the main limiting or important
factor.
In the murine nail apparatus, we found a homogenous distribution of mast cells
without any significant differences in mast cell numbers in the compartments of the
murine nail apparatus (Fig. 4.64). The only exception were the surroundings of the
nail bed, where significantly less mast cells were detectable than in the PNF. Since
this part has a sterile environment and is lying between two important hard tissue
structures, namely the nail plate and the bone where much pressure occurs, it might
be that a low number of mast cells should reduce the risk of unnecessary
inflammation induced by trauma. However, according to the human nail system, the
expected reduced numbers of dermal mast cells in the vicinity of the proximal nail
matrix was not confirmed. The explanation of ITO et al. (2005) for these results
emphasized that low mast cell number may compromise the defense capacity of this
region of nail mesenchyme, whereas in the terminally differentiated nail epithelium
locally produced AMPs (DORSCHNER et al. 2004) may compensate and supply the
innate immune defense. However, the authors did not investigate the mast cell
activation status. Since we did not find such differences, the explanation suggested is
not convincing and it remains to be controlled by future experiments whether the
activation status of mast cells differs in the defined tissue compartments, and plays a
more important role in IP than mast cell numbers.
ICAM-1 is rapidly inducible on endothelial cells by infection, playing a major role in
local inflammatory responses (CAMELI et al. 1994; BOS 1997; GOLDSBY et al.
2003; JANEWAY et al. 2005). Since it is may be involved in IP maintenance or,

DISCUSSION
199
collapse respectively, we also examined this molecule. In this connection, it is
noteworthy to recognize that ICAM-1 expression pattern obviously differs
substantially between the follicle and the nail apparatus immune system. ITO et al
(2005) reported in their in situ studies that the infantile human nail apparatus did not
demonstrate ICAM-1 IR in any epithelial compartment, except for the expected IR in
blood vessels. In the current study, we could confirm this expression pattern also for
the murine nail apparatus. We identified ICAM-1+ positive cells as limited to the
mesenchyme (Fig. 4.61 A), except for endothelial cells of small blood vessels near
the epidermis of the PNF. The number of positive cells was slightly reduced in the
vicinity of the PNM and the foot pad. In contrast, studies of the HFs in mice and
humans have demonstrated constitutively expression of ICAM-1 in the peri- and
infrainfundibular ORS, thus attracting LFA-1 expressing immunocytes (MUELLER-
ROEVER et al. 2000). This is not surprising, because the ORS and the bulge region
has been shown to be the preferred site for the occurrence of dense perifollicular
inflammatory cell infiltrates under stress (ARCK et al. 2001, 2003). In addition, it was
reported that in normal, uninflamed mouse skin, a very small, but notable percentage
of HFs are permanently deleted by an inflammatory cell attack on the ICAM-1+ bulge
region of anagen and catagen pelage HFs, without any macroscopic evidence for this
striking phenomenon of ´programmed organ deletion (POD)` (EICHMULLER et al.
1998). ICAM-1 IR in normal human skin was found on most CTS and dermal
fibroblasts and immune cells, but the strongest ICAM-1 IR was seen on perifollicular
inflammatory cell clusters (PICC) (JAWORSKY et al. 1992). In addition, ICAM-1
expression was found on HF keratinocytes in the close vicinity of perifollicular
inflammatory cell clusters (CHRISTOPH et al. 2000), in particular in the perifollicular
bulge region (MEYER et al. 2008c). Whether such physiological POD of isolated (or
possibly malfunctioning) HFs also occurs in human skin, and if at least some forms of
cicatricial alopecias only represent an excessive form of POD (PAUS et al. 1999),
remains to be investigated.
Furthermore, in the current study a strong up-regulation of the staining intensity of β-
defensin 2 was found in parts of the PNM close to the nail plate, as compared to the

DISCUSSION
200
PNF and parts of the foot pad (Figs. 4.66, 4.67). The staining was cytoplasmic and a
discreet IR could also be shown in other epithelial parts. These results are slightly
contradictive and unexpected, because APMs are expected to be secreted in
epithelial parts facing to the environment (DOERFL 1982; DORSCHNER et al. 2004).
However, results from wild mammals on the production of antimicropial peptides
have demonstrated that not only the epidermis, but also the complete hair follicle
complex, i.e. including the entire ORS, the apocrine tubular glands and the
sebaceous glands can express CAPs or ß-glucan receptors (MEYER et al. 2003,
2008b, MEYER and SEEGERS 2004).
Taken together, our results suggest that the nail immune system shows similarities to
the HF immune system, including the possible establishment of an area of relative IP
in the PNM. The functional advantages of establishing IP in the nail matrix are still
uncertain. Also, it is unknown yet, which key autoantigens are shared between nail
apparatus and HFs, and whether the collapse of IP in the nail matrix may also
underlie the frequent involvement of the nail in AA.
5.3.3 Comparison of the human HF bulge, murine sinus hair follicle
and nail apparatus
The human HF, the murine vibrissal follicle and the nail apparatus show one common
aspect: They all belong to skin appendages and are relatives from the evolutionary
point of view. They consist of epithelial and mesenchymal structures and are in direct
contact with the environment. In addition, many structures and their functions of the
human HF and the vibrissal follicle are exactly the same ones.
Despite the above mentioned similarities, there are several aspects in which these
tissues differ, and which make a direct comparison difficult, if they belong to sites of
IP or whether the supposed function of IP could be similar. In the following, only the
most relevant aspects will be emphasized: Human HFs (as well as murine pelage

DISCUSSION
201
HFs) undergo life-long cycles of growth (anagen), regression (catagen) and relative
quiescence (telogen), the so-called HF cycle. During anagen development, i.e. when
a new hair shaft (HS) factory - the anagen hair bulb - is reconstructed, the HF
recapitulates in part key morphological events of its own morphogenesis. This
astounding phenomenon of cyclic organ regeneration during adult life is only possible
because this ´miniorgan` is richly endowed with eSCs. Terminal HFs for example,
function as a protection against the environment, act as a heat balance and
mechanoreceptor, or contribute to the individual and characteristic appearance.
In contrast, vibrissal follicles show also life-long cycling, but termination of hair growth
in the previous hair cycle and initiation of hair regeneration in the following cycle
partially overlaps. Thus, telogen is not obvious; otherwise the specific sensory
function of the tactile organ would be interrupted. The nail apparatus, however, is a
permanent structure without cycling. The nail plate is continuously produced by the
nail matrix and nail bed. In conclusion, the signals for eSCs are considered to differ
considerably from each other, which might affect also their respective defense
mechanisms.
Nonetheless, apart from such differences, the suggested function of bulge IP and IP
in the PNM is the protection of epithelial stem cells. Since IP is a phenomenon, which
is described in many tissues (SIMPSON 2006), independent of the fact that these
often strongly differ from each other (brain, fetus, anterior chamber of the eye, testes,
hamster cheek pouch), so comparisons are possible, regarding to the widely
accepted markers. In this study, we presented a down-regulation of MHC class Ia -
one of the key IP mechanism - and MHC class II molecules in all three tissues
investigated. In addition, the expression of important immunosuppressive molecules
like TGF-β could be demonstrated. The distribution pattern of CD4+ and CD8+ T
cells in the vibrissal follicle and nail apparatus also resemble the one of human HF.
Taken together, although we observed only a small amount of IP marker in the
murine specimens, available data suggest that the vibrissal bulge and bulb, as well
as the PNM also belong to a site of relative IP.

DISCUSSION
202
5.4 Effects of K(D)PT on the hair follicle immune system
α-MSH is known to have melanotropic, immunomodulatory and restoration capacity
in anagen hair bulbs (ITO et al. 2004), and acts as a mediator of tolerance induction
in the brain (TAYLOR and KITAICHI 2007) and eye (NIEDERKORN 2006).
One of the key questions investigated in this study was whether the synthetic α-MSH
related tripeptide K(D)PT can restore HF IP after IFN-γ induced collapse, or whether
it has protective functions, respectively, as it was shown for other immuno
suppressants (ITO et al. 2004). Since low or absent expression of MHC class I
protein in hair matrix keratinocytes is the hallmark of HF IP (ITO et al. 2004), we
imitated an IP collapse by application of IFN-γ (ITO et al. 2004). We chose a dose of
75 IU/ml IFN-γ to prevent a transition of the anagen VI HFs into catagen, because
hair bulb IP is hair cycle dependent and occurs in anagen VI only.
Our MHC class I results (Fig. 4.27) show that K(D)PT in the concentrations tested
failed to act as an agent for either re-establishing IP or for protecting against its
collapse. As shown previously (ITO et al. 2004), 75 IU/ml IFN-γ significantly
upregulated MHC class Ia IR in the hair bulb of human anagen VI HFs, but none of
the applicated K(D)PT doses could significantly and reliably reduce the IFN-γ I-
induced MHC class I IR in the hair matrix and proximal ORS. This is corroborated by
the observed, comparably limited effects on the downregulation of MHC class II+
cells (Fig. 4.28). However, high-dose K(D)PT (10-8M) may well down-regulate the
number of MHC class I and II+ cells (and/or their expression level!) in susceptible
individuals. So far, it is unclear, which factors make the individual susceptible or not.
In accordance to previous studies, the tested dose range of K(D)PT was chosen in
relation to the concentration of α-MSH, which was detected as an IP restorer (ITO et
al. 2007).
Intriguingly, K(D)PT in dose-dependent manner reduced both, the number of mature
mast cells and their degranulation / activation status and counteracted IFN-γ pro-
inflammatory mast cell-directed stimuli (Fig. 4.31). Since α-MSH mast cell interaction
probably takes place via the MC-1R (LUGER et al. 2000), this might be also relevant

DISCUSSION
203
for K(D)PT. Mast cells not only elicit the first and fastest innate immune response
after recognition through Toll-like receptors and cytokine release (JAHANYAR et al.
2008), but are also material to the modulation of adaptive immune response
(secretion of pro-inflammatory and anti-inflammatory mediators, acting as antigen-
presenting cells and expression of co-stimulatory molecules) (JAHANYAR et al.
2008). This functions could lead into excessive inflammation and are required for
cutaneous wound healing (WELLER et al. 2006). In addition, recent studies showed
that inhibition of mast cell activation confers to the maintenance of immune privilege
(FIJAK and MEINHARDT 2006), i.e. the optimal expression of peripheral tolerance to
skin allografts (MAURER et al. 2003a; METZ et al. 2008a). Thus, K(D)PT obviously
exerts immunomodulatory capacity, and deserves full exploration as a mast cell
inhibitor in all inflammatory skin conditions where mast cells play a major role (e.g.
allergy, atopic eczema, urticara).
It should also be noted, that K(D)PT inhibits inflammation-induced premature catagen
(Fig. 4.32) development, which also matches with the Ki67/TUNEL results (Fig.
4.33). Such unexpected effect of an α-MSH-related peptide on HF cycling suggests
that K(D)PT might become exploitable for counteracting telogen effluvium induced by
various pro-inflammatory scalp skin processes that induce premature catagen
development (e.g. seborrhoic dermatitis). In addition, we found that K(D)PT does not
appear to greatly influence hair shaft formation (data not shown) or to stimulate hair
matrix keratinocyte proliferation indicating that K(D)PT is unlikely to have major or
general hair growth-stimulatory properties. On the contrary, the observation that
K(D)PT tended to reduce apoptosis in IFN-γ treated HFs raises the possibility, that
K(D)PT under inflammatory conditions may operate as a general ´epithelial
proliferation protector` in the HF and elsewhere in human skin.
In conclusion, despite its failure to impress as a reliable HF IP protectant / restorer,
K(D)PT may yet prove to be of importance in the management of inflammatory skin
diseases in which mast cells play a major role. Specifically, topical application of
K(D)PT, allowing the administration of high peptide doses with a favourable benefit /

DISCUSSION
204
risk ratio, may become exploitable for the treatment of post-inflammatory poliosis, or
for treating the recalcitrant poliosis that is characteristically seen during the recovery
phase of alopecia areata, an organ-specific autoimmune hair growth disorder, in the
pathogenesis of which IFN-γ is recognized to play a key role (GILHAR et al. 2007).
Beyond these effects there may be plenty other so far unknown functions useful for
the treatment of skin diseases.
5.5 Conclusions
On the basis of current and generally accepted knowledge, we present suggestive
phenomenological and functional evidence that the bulge of human anagen VI HFs
represents another site of relative IP within human skin, as predicted by previous
gene expression profiling data. However, our in situ-protein expression results also
suggest that this IP site, the most vital part of the follicle with its stem cell reservoir,
displays a number of features that distinguish it from the bulb IP. It remains to be
clarified, whether these differences are signs of different IP strength, development or
importance. In any case, the full thickness scalp skin organ culture offers good
opportunities to further explore bulge IP collapse and restoration in regard to other
parameters.
The immunophenotyping results obtained in the course of this study shed further light
on the in situ-expression patterns of some immunological and IP associated markers
in the murine vibrissal follicle and nail apparatus. The results suggest that the
vibrissal bulb, in particular, but also the bulge and to a lesser extent the proximal nail
matrix, probably meet the criteria to name these sites as immunoprivileged.
Nonetheless, some parameters remain to be studied, and functional evidence should
be provided before a definite statement regarding IP establishment can be made.
Irrespective of whatever function the IP of the mammalian vibrissal follicle and nail
apparatus has, these findings contribute to immuno biology in general and fill
scientific gaps, facilitating further research in the field of IP.

DISCUSSION
205
When interpreting the current data, it has to be pointed out that IP is a relative, not an
absolute state, reflecting the net result of multiple interconnected active and passive
mechanisms (FERGUSON and GRIFFITH 2006; SIMPSON 2006). In addition, it is
an ongoing debate, how many and which of these mechanisms have to be
established in a given tissue location to justify the term ´IP` (FERGUSON and
GRIFFITH 2006). Since no universally accepted criteria have as yet been defined
that apply to all IP tissues, ultimate proof for the human bulge, the vibrissal follicle or
the proximal nail matrix as sites of relative IP can only arise from functional studies,
such as the famous (as yet never repeated) melanocyte allotransplant experiment
(BILLINGHAM 1971; PAUS et al. 2005).
Furthermore, the physiologically relevant HF organ culture assay employed here,
simulating IP collapse via IFN-γ treatment, offers a highly instructive experimental
tool for the research of substances as IP restorer, like K(D)PT, and ideally
complements human cell culture models available (BELL et al. 1979). Despite its
failure to act as a reliable HF IP protectant / restorer, K(D)PT may yet prove to be
very useful in the management of inflammatory skin diseases in which mast cells
play a major role.
5.6 Perspectives
In order to demonstrate that a putative ´IP` status of any tissue compartment is truly
relevant to organ or system homeostasis, it needs to be shown that loss of IP actually
can lead to pathology (FERGUSON and GRIFFITH 2006). While it may be argued
that primary cicatricial alopecias (PCA), such as lichen planopilaris and CCLE-
associated scarring alopecia, represent just such examples for pathology as a result
of bulge IP loss, where an autoaggressive inflammatory cell attack on the HF‘s stem
cell region damages and destroys its stem cell-based regenerative capacity.
Definitive evidence that this is indeed the case has not yet been published.
In cooperation with Dr. M. Harries from the University of Manchester a current study
follows-up this lead and investigates, whether loss of bulge IP causes primary

DISCUSSION
206
cicatricial alopecias (HARRIES et al. 2008; HARRIES et al. submitted). Therefore,
punch scalp biopsies from healthy donors, and patients with PCA (unaffected vs.
affected scalp skin) are collected and compared immunohistochemically in respect of
IP parameters.
In a second step, full thickness organ culture experiments should be performed, to
evaluate substances, which are capable of restoring bulge IP. Another optimal test
system, in which this missing proof of pathology may be generated, would be to use
xenotransplants of hair-bearing human skin onto SCID mice (GILHAR et al. 1998), as
ideally after reconstitution with a human immune system upon transfer of appropriate
human stem cells.
In regard to the murine vibrissal follicles and nail apparatus, some parameters remain
to be investigated with respect to the in situ expression pattern, e.g. MHC class Ib
molecules, the ´no danger signal` CD200, IDO, MIF, and other immunosuppressants.
In addition, it would be helpful to perform also full-thickness skin organ culture, to
prove whether or not the suspected IP site could be induced to collapse.
With regard to the common autoimmune disease AA, which is considered to be
caused by the loss of IP within the human anagen hair bulb, it would be promising to
concentrate on the restoration of the HFs lost or compromised IP, both for preventing
the progression of AA lesions and for inducing hair regrowth. This IP restoration
therapy does not require any prior knowledge of the relevant key autoantigens or the
specific autoreactive T cells, and it can resort to the administration of well-known
nonspecific immunomodulators that drastically down-regulate ectopic MHC class I
expression in the anagen hair bulb. Although K(D)PT failed to restore IP, it exerts
several other interesting functions, which should be explored systematically.

SUMMARY
207
6 Summary
Katja C. Meyer
Detailed analysis of immunoprivileged tissues in skin appendages
Recent gene profiling data in mice have raised the possibility, that follicular IP is not only
limited to the anagen hair bulb, but that the epithelial stem cell region in the ORS of HFs,
termed the bulge, may also represent an area of relative IP. Since protection of these
epithelial stem cells from immune destruction is essential for preserving the regenerative and
cycling capacity of the follicle, it seems logical that the bulge region should also have an
established relative IP.
The current study on the immune status of human HFs and murine sinus hair follicles and
nail apparatus, is based on both routine and increased-sensitivity immunohistochemical
(including Immunofluorescence) staining techniques, and evaluation of these IR patterns by
quantitative immunohistochemistry, histochemical and histomorphometric assessments.
Additionally, full thickness human scalp skin organ culture and the HF organ culture model
were used for functional evidence.
The in situ protein expression data presented here provide evidence, that the bulge region of
anagen VI HFs meets all criteria of an IP site, and suggests that the bulge has a perhaps
stronger immune protection than that of the anagen bulb. Contrary to previous reports, we
demonstrated that MHC class Ia, β2-microglobulin and MHC class II IR were down-regulated
in the human bulge relative to the distal ORS. The locally secreted immunosuppressants α-
MSH and TGF-β were prominently expressed in the bulge. In addition, IDO and MIF showed
higher IR levels around the bulge compared to the proximal follicle and CD200 and HLA-E
were solely expressed in the stem cell-rich bulge region. Besides these systematic bulge
immuno phenotyping experiments, we obtained functional evidence for an established IP in
the bulge, by showing that IFN-γ can induces significant ectopic MHC class Ia expression in
bulge cells of organ-cultured human scalp skin, as it had been shown for the anagen hair
bulb. However, our results also revealed that the bulge and bulb IP in human scalp HFs differ
in several important aspects: The level of constitutive MHC I, MHC II and β2-microglobulin IR
was not as low as in the hair bulb, which may indicate that the bulge IP is constitutively more
prone to inciting an immune response than the bulb, resulting in an inflammatory cell attack
on bulge epithelial stem cells.

SUMMARY
208
This thesis also contributes considerably towards the immunological characterization of the
murine vibrissal follicle and murine nail apparatus, and showed that the vibrissal bulb, and to
a lesser extent, the bulge as well as the proximal nail matrix can also be considered to be
sites of relative IP. Both tissues showed a substantial downregulation of MHC class I and II in
the examined IP sites. Prominent IR of the immunosuppressant TGF-β1 was demonstrated
in the PNM, bulge and weaker in the bulb, exerting also an important role in IP. Furthermore,
the presented distribution pattern of CD4+ and CD8+ T cells was similar to the human HF.
Besides these IP-associated findings, we could observe that mast cell numbers decreased in
vibrissal follicles towards the vibrissal bulb, whereas no relevant differences in the
distribution could be shown in the murine nail apparatus. In addition, we identified ICAM-1+
positive cells to be limited to the mesenchyme of the nail apparatus. The number of these
cells was slightly reduced in the vicinity of the PNM and the foot pad, but no intraepithelial
clusters were observed, in accordance with previous studies on the human nail apparatus.
Unexpected results were obtained for β-defensin 2, showing substantially higher levels of IR
in the PNM and in the entire ORS of the vibrissal follicle, and only weak IR in epidermal
layers. Nonetheless, whether mast cells, ICAM-1, and β-defensin 2 contribute to the
mechanisms for or against IP, remains to be further explored.
The analyses of this study were concluded by exploring the effects of the α-MSH related
tripeptide K(D)PT by using an organ-culture HF model. K(D)PT failed to act as a protection or
restoration agent of IP. However, K(D)PT counteracted IFN-γ-induced upregulation of mast
cell numbers and degranulation. In addition, we demonstrated that K(D)PT inhibited
inflammation-induced premature catagen development, whereas it did not considerably
influence hair shaft formation in vitro or stimulate the proliferation of hair matrix keratinocytes.
In summary, the current thesis has provided evidence for a second epithelial IP within the
human HF, which could even be stronger than that of the bulge. Moreover, the presented
results indicate that the murine PNM, and the bulge and bulb of vibrissal follicles also enjoy
IP, so that some gaps in basic immunological research could be filled. This was
complemented with the exploration of the new candidate K(D)PT for IP-restoration or
protection and the demonstration of several effects, which may involve that K(D)PT may be
used pharmacologically as a hair growth modulator.

ZUSAMMENFASSUNG
209
7 Zusammenfassung
Katja C. Meyer
Detaillierte Analyse von immunpriviligierten Geweben in Hautanhangsorganen
Neueste Daten auf Genebene von Mäusen haben die Vermutung nahegelegt, dass nicht nur
die anagene Haarwurzel zum follikulären Immunprivileg (IP) zählt, sondern auch die
sogenannte Wulstregion in der äußeren Wurzelscheide des Haarfollikels, die dessen
epitheliale Stammzellregion darstellt.
Methodische Grundlagen für die vorliegende Studie an Haarfollikeln des Menschen und an
Sinushaarfollikeln und dem Nagelapparat der Maus waren primär hochspezifische
immunhistochemische Färbungen (inklusive Immunfluoreszenz), die qualitativ und quantitativ
ausgewertet wurden. Weiterhin wurden für die Erarbeitung funktioneller Daten
Vollhautorgankulturen von humanen Kopfhautproben sowie Kulturen mit mikrodissezierten
humanen Haarfollikeln verwendet.
Die hier präsentierten in situ Protein Reaktivitätsverteilungsmuster erbringen den Beweis,
dass die Wulstregion des Anagen VI Haarfollikels alle Kriterien eines immunprivilegierten
Gewebes erfüllt. Sie lassen sogar vermuten, dass die Wulstregion einen stärkeren
Immunschutz aufweist als die anagene Haarwurzel. Im Gegensatz zu früheren
Veröffentlichungen zeigen wir in unserer Arbeit, dass die Immunreaktivität von MHC Klasse
Ia, β2-Mikroglobulin und MHC Klasse II Molekülen in der Wulstregion, verglichen mit der
distalen äußeren Wurzelscheide, stark herabgesetzt ist. Die lokal sezernierten
Immunosuppressoren α-MSH und TGF-β waren ebenfalls in der Wulstregion prominent.
Zusätzlich zeigten IDO und MIF höhere Immunintensitäten im Bereich der Wulstregion als im
proximalen Haarfollikel und die Moleküle CD200 und HLA-E waren alleinig in der
stammzellreichen Wulstregion vertreten. Neben dieser systematischen,
immunphänotypischen Analyse, konnten wir zusätzlich einen funktionellen Beweis für das
Bestehen eines IP in der Wulstregion erbringen: Ähnlich den Beobachtungen vorheriger
Experimente an der immunprivilegierten anagenen Haarwurzel, konnten wir demonstrieren,
dass eine Behandlung mit IFN-γ in organ-kultivierten humanen Skalp-Vollhautstanzen eine
Hochregulation von MHC Klasse Ia IR in der Wulstregion bewirkte. Gleichzeitig zeigen die
Ergebnisse jedoch auch, dass sich die Wulstregion und die anagene Haarwurzel hinsichtlich

ZUSAMMENFASSUNG
210
verschiedener Aspekte unterscheiden: Die Intensitätsstärke von MHC Klasse I, Klasse II und
β2-Mikroglobulin ist in der Wulstregion höher als in der Haarwurzel, was darauf hin deutet,
dass das IP der Wulstregion unmittelbarer eine Immunreaktion zulässt als die Haarwurzel
und die Folge eine entzündliche Zellattacke auf die epithelialen Stammzellen in der
Wulstregion und Zerstörung sein könnte.
Mit dieser Arbeit wurde auch ein Beitrag zur immunologischen Charakterisierung von
Sinushaarfollikel und dem Nagelapparat der Maus geleistet. Es konnte gezeigt werden, dass
die Haarwurzel des Sinushaarfollikels, und zu einem geringeren Maße dessen Wulstregion
sowie die proximale Nagelmatrix, ebenfalls zu immunologisch privilegierten Regionen
zählen. Beide Gewebetypen in den genannten Regionen wiesen eine stark reduzierte
Färbungsintensität für MHC Klasse I und II Moleküle auf. Ferner war TGF-β1 stark präsent in
der proximalen Nagelmatrix, in der Wulstregion und, weniger intensiv, in der Haarwurzel, so
dass eine wichtige Funktion für diese Substanz zu unterstellen ist. Das Verteilungsmuster
von CD4+ und CD8+ T Zellen war ebenfalls ähnlich den Verhältnissen im humanen
Haarfollikel. Neben diesen IP-assoziierten Ergebnissen konnten wir zeigen, dass die Zahl an
Mastzellen innerhalb des Sinushaarfollikels von der Epidermis zur Haarwurzel kontinuierlich
abnimmt, wohingegen kein relevanter Unterschied im Verteilungsmuster innerhalb des
Maus-Nagelapparates zu verzeichnen war. Zusätzlich wurden ICAM-1 positive Zellen
identifiziert, die auf das Mesenchym des Nagelapparates begrenzt waren. In direkter Nähe
zur proximalen Nagelmatrix und zum Fußballen waren etwas weniger ICAM-1+ Zellen zu
beobachten. Intraepitheliale Anhäufungen dieser Zellen wurden, entgegen vorheriger
Berichte, im humanen Nagelsystem nicht gefunden. Unerwartete Resultate wurden ebenfalls
für β-Defensin 2 erhoben, da die proximale Nagelmatrix und die gesamte ORS des
Sinushaarfollikels substantiell höhere Reaktionsintensitäten aufwiesen, als in den
epidermalen Zellschichten. Ob und welche Rolle Mastzellen, ICAM-1+ Zellen und das
antimikrobielle Peptid β-Defensin 2 zur Erhaltung des IP spielen, sollte in zukünftigen
Versuchen weiter erforscht werden.
Die Analysen dieser Studie wurden ergänzt mit der Erforschung von K(D)PT hinsichtlich
seiner Effekte auf den Haarfollikel durch den Einsatz unseres HF-Organ-Kulturmodels. Diese
Substanz, ein neues synthetisches, mit α-MSH verwandtes Tripeptid, war jedoch als
Wirkstoff zur Protektion oder Wiederherstellung des IP nicht überzeugend. Wir konnten
allerdings demonstrieren, dass K(D)PT der IFN-γ induzierten Zunahme von Mastzellen und

ZUSAMMENFASSUNG
211
der Rate an Degranulation entgegenwirkte; zusätzlich inhibierte K(D)PT die
entzündungsinduzierte vorzeitige Catagenentwicklung.
Zusammenfassend gesehen, erbringt die vorliegende Dissertation den Beweis, dass die
Wulstregion ein zweites epitheliales IP innerhalb des HFs darstellt. Dieses IP ist
möglicherweise stärker oder bedeutender, als dasjenige der Haarwurzel. Zusätzlich weisen
die hier vorgestellten Ergebnisse darauf hin, dass die murine PNM des Nagelapparates,
sowie die Wulstregion und Haarwurzel des Sinushaarfollikels ebenfalls immunologisch
privilegiert sein könnten; zudem war es möglich, einige Lücken in der immunologischen
Grundlagenforschung zu schließen. Ergänzt wurde diese Studie durch die Erforschung eines
neuen Haarmodulator-Kandidatens, dem K(D)PT, das zwar nicht als IP-Modulator
einzusetzen ist, jedoch zur Behandlung von Haarausfall und gegen Vergrauung genutzt
werden könnte.

ANNEX
212
8 References
ABI-HANNA, D., WAKEFIELD, D. and WATKINS, S. (1988): HLA antigens in ocular tissues. I. In vivo expression in human eyes. Transplantation 45, 610-613 ABTIN, A., ECKHART, L., MILDNER, M., GRUBER, F., SCHROEDER, J. M. and TSCHACHLER, E. (2008): Flagellin is the principal inducer of the antimicrobial peptide S100A7c (psoriasin) in human epidermal keratinocytes exposed to Escherichia coli. FASEB J. 22, 2168-2176 AHMAD, W., PANTELEYEV, A. A., HENSON-APOLLONIO, V., SUNDBERG, J. P. and CHRISTIANO, A. M. (1998): Molecular basis of a novel rhino (hr(rhChr)) phenotype: a nonsense mutation in the mouse hairless gene. Exp. Dermatol. 7, 298-301 ALONSO, L. and FUCHS, E. (2006): The hair cycle. J. Cell Sci. 119, 391-393 ANDRES, K. H. (1966): Zur Feinstruktur des Merkelschen Tastsinnesapparates am Sinushaar. Naturwissenschaften 53, 706 APTE, R. S. (1997): Local inhibition of natural killer cell activity promotes the progressive growth of intraocular tumores. Invest. Ophtalmol. Vis. Sci. 38, 1277-1282 APTE, R. S. and NIEDERKORN, J. Y. (1996): Isolation and characterization of a unique natural killer cell inhibitory factor present in the anterior chamber of the eye. J. Immunol. 160, 5693-5696 APTE, R. S., SINHA, D., MAYHEW, E., WISTOW, G. J. and NIEDERKORN, J. Y. (1998): Cutting edge: role of macrophage migration inhibitory factor in inhibiting NK cell activity and preserving immune privilege. J. Immunol. 160, 5693-5696

REFERENCES
213
ARCK, P. C., HANDJISKI, B., HAGEN, E., JOACHIM, R., KLAPP, B. F. and PAUS, R. (2001): Indications for a ´brain-hair follicle axis (BHA)`: inhibition of keratinocyte proliferation and up-regulation of keratinocyte apoptosis in telogen hair follicles by stress and substance P. FASEB J. 15, 2536-2538 ARCK, P. C., HANDJISKI, B., PETERS, E. M., PETER, A. S., HAGEN, E., FISCHER, A., KLAPP, B. F. and PAUS, R. (2003): Stress inhibits hair growth in mice by induction of premature catagen development and deleterious perifollicular inflammatory events via neuropeptide substance P-dependent pathways. Am. J. Pathol. 162, 803-814 ARCURI, F., CINTORINO, M., CARDUCCI, A., PAPA, S., RIPARBELLI, M. G., MANGIONI, S., DI BLASIO, A. M., TOSI, P. and VIGANO, P. (2006): Human decidual natural killer cells as a source and target of macrophage migration inhibitory factor. Reproduction 131, 175-182 AUBER, L. (1952): The anatomy of follicles producing wool fibres with special reference to keratinization. Trans. Royal Soc. Edinburgh. 62, 191-254 BADEN, H. P. and KVEDAR, J. C. (1993): Epithelial cornified envelope precursors are in the hair follicle and nail. J. Invest. Dermatol. 101, 72S-74S BALAKRISHNAN, S. and PURI, R. K. (1973): Fetal age and morphological features. Indian Pediatr. 10, 427-433 BALS, R., WANG, X., MEEGALLA, R. L., WATTLER, S., WEINER, D. J., NEHLS, M. C. and WILSON, J. M. (1999): Mouse ß-defensin 3 is an inducible antimicrobial peptide expressed in the epithelia of multiple organs. Infect. Immun. 67, 3542-3547 BAUER, S., GROH, V., WU, J., STEINLE, A., PHILIPS, J. H., LANIER, L. L. and SPIES, T. (1999): Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285, 727-729 BEDDARD, F. E. (1902): Observation upon the carpal vibrissae in mammals. Proc. Zool. Soc. 3, 127-136

ANNEX
214
BERNHAGEN, J., BACHER, M., CALANDRA, T., METZ, C. N., DOTY, S. B., DONNELY, T. and BUCALA, R. (1996): An essential role for macrophage migration inhibitory factor in the tuberculin delayed-type hypersensitivity reaction. J. Exp. Med. 183, 277-282 BILLINGHAM, R. E. (1971): Transplantation immunity evoked by skin homografts and expressed in intact skin. Adv Biol Skin. 11, 183-198 BILLINGHAM, R. and BOSWELL, T. (1953): Studies on the problem of corneal homografts. Proc. R. Soc. Lond. B. 141, 392-406 BILLINGHAM, R. E. and SILVERS, W. K. (1971): A biologist's reflections on dermatology. J. Invest. Dermatol. 57, 227-240 BILLINGHAM, R., BRENT, L. and MEDAWA, P. (1954): Quantitative studies on tissue transplantation immunity. II. The origin, strength, and duration of actively and adoptively acquired immunity. Proc. R. Soc. Lond B. 143, 58-80 BLAND-SUTTON, J. (1887): On the arm glands of the Lemurs. Proc. zool. Soc. 369-372 BLANPAIN, C., LOWRY, W. E., GEOGHEGAN, A., POLAK, L. and FUCHS, E. (2004): Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell 118, 635-648 BLASCHITZ, A., HUTTER, H. and DOHR, G. (2001): HLA Class I protein expression in the human placenta. Early Pregnancy 5, 67-69 BLOOM, B. R. and BENNET, B. (1966): Mechanism of a reaction in vitro associated with delayed-tape hypersensitivity. Science 153, 80-82

REFERENCES
215
BODO, E., TOBIN, D. J., KAMENISCH, Y., BIRO, T., BERNEBURG, M., FUNK, W. and PAUS, R. (2007): Dissecting the impact of chemotherapy on the human hair follicle: a pragmatic in vitro assay for studying the pathogenesis and potential management of hair follicle dystrophy. Am. J. Pathol. 171, 1153-1167 BODO, E., VAN BEEK, N., NAUMANN, V., OHNEMUS, U., BRZOSKA, T., ABELS, C. and PAUS, R. (2008): Modulation of chemotherapy-induced human hair follicle damage by 17-ß estradiol and prednisolone: Potential stimulators of normal hair regrowth by ´dystrophic catagen` promotion? J. Invest. Dermatol. (in press) BOECK, P. (ed.)(1989) Romeis – Mikroskopische Technik. 17th Edition. Urban and Schwarzenberg, München, Wien BOEHM, T. (2006): Quality control in self/nonself discrimination. Cell 125, 845-858 BOEHM, M., LUGER, T., TOBIN, D. J. and GARCIA-BORRÓN, J. G. (2000): Melanocortin receptor ligands: new horizons for skin biology and clinical dermatology. J. Invest. Dermatol. 126, 1966-1975 BRZOSKA, T., LUGER, T.A., MAASER, C., ABELS, C AND BÖHM, M. (2008) α-melanocyte-stimulating hormone and related tripeptides: biochemistry, antiinflammatory and protective effects in vitro and in vivo, and future perspectives for the treatment of immune-mediated inflammatory diseases. Endocr. Rev. 29, 581-602. BOMAN, H. G. (1995): Peptide antibiotics and their role in innate immunity. Annu. Rev. Immunol. 13, 61-92 BORGENS, R. B. (1982): Mice regrow the tips of their foretoes. Science 217, 747-750 BOTCHKAREV, V. A., BOTCHKAREVA, N. V., SLOMINSKI, A., ROLOFF, B., LUGER, T. and PAUS, R. (1999): Developmentally regulated expression of α-MSH and MC-1 receptor in C57BL/6 mouse skin suggests functions beyond pigmentation. Ann. N. Y. Acad. Sci. 885, 433-439

ANNEX
216
BOTCHKAREV, V. A., EICHMUELLER, S., JOHANSSON, O. and PAUS, R. (1997a): Hair cycle-dependent plasticity of skin and hair follicle innervation in normal murine skin. J. Comp. Neurol. 386, 379-395 BOTCHKAREV, V. A., EICHMUELLER, S., PETERS, E. M. J., PIETSCH, P., JOHANSSON, O., MAURER, M. and PAUS, R. (1997b): A simple immunofluorescence technique for simultaneous visualization of mast cells and nerve fibres reveals selectivity and hair cycle-dependent changes in mast cell - nerve fibre contacts in murine skin. Arch. Dermatol. Res. 289, 292-302 BOTCHKAREV, V. A., WELKER, P., ALBERS, K. M., BOTCHKAREVA, N. V., METZ, M., LEWIN, G. R., BULFONE-PAUS, S., PETERS, E. M., LINDNER, G. and PAUS, R. (1998b): A new role for neurotrophin-3: involvement in the regulation of hair follicle regression (catagen). Am J Pathol. 153, 785-799 BOTEZAT, E. (1897): Die Nervenendigungen an den Tasthaaren von Säugetieren. Arch. mikr. Anat. 50, 142-169 BOZZA, M., SATOSKAR, A. R., LIN, G., LU, B., HUMBLES, A. A., GERARD, C. and DAVID, J. R. (1999): Targeted disruption of migration inhibitory factor gene reveals its critical role in sepsis. J. Exp. Med. 189, 341-346 BRAFF, M. H., BARDAN, A., NIZET, V. and GALLO, R. L. (2005a): Cutaneous defense mechanisms by antimicrobial peptides. J. Invest. Dermatol. 125, 9-13 BRAFF, M. H., ZAIOU, M., FIERER, J., NIZET, V. and GALLO, R. L. (2005b): Keratinocyte production of cathelicidin provides direct activity against bacterial skin pathogens. Infect. Immun. 73, 6771-6781 BRAUD, V. M., ALLAN, D. S., O'CALLAGHAN, C. A., SODERSTROM, K., D'ANDREA, A., OGG, G. S., LAZETIC, S., YOUNG, N. T., BELL, J. I., PHILLIPS, J. H., LANIER, L. L. and MCMICHAEL, A. J. (1998): HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 391, 795-799

REFERENCES
217
BRECHT, M., PREILOWSKI, B. and MERZENICH, M. M. (1997): Functional architecture of the mystacial vibrissae. Behav. Brain Res. 84, 81-97 BROECKER E.B., ACHTERNACHT-HAPPLE K. and HAMM, R.H. (1987): Abnormal expression of class I and class II major histocompatibility antigen in alopecia areata. J. Invest. Dermatol. 88, 564-568 BROWN, J. M., WILSON, T. M. and METCALFE, D. D. (2007): The mast cell and allergic diseases: role in pathogenesis and imlications for therapy. Clin. Exp. Allergy 38, 4-18 BRUNO, R., SABATER, L., SOSPEDRA, M., FERRER-FRANCESCH, X., ESCUDERO, D., MARTINEZ-CACERE, E. and PUJOL-BORRELL, R. (2002): Multiple sclerosis candidate autoantigens except myelin oligodendrocyt glycoprotein are transcribed in human thymus. Eur. J. Immunol. 32, 2737-2747 BUKOVSKY, A., PRESL, J., ZIDOVSKY, J. and MANCAL, P. (1983): The localization of Thy-1.1, MRC OX 2 and Ia antigens in the rat ovary and fallopian tube. Immunology 48, 587-596 CAMELI, N., PICARDO, M., TOSTI, A., PERRIN, C., PISANI, A. and ORTONNE, J. (1994): Expression of integrins in human nail matrix. Br. J. Dermatol. 130, 583-588 CAPSI, R. R. (2006): Ocular autoimmunity: the price of privilege. Immunol. Rev. 213, 23-35 CARLIN, J. M., BORDEN, E. C., SONDEL, P. M. and BYRNE, G. I. (1989): Interferon-induced indoleamine 2,3- dioxygenase activity in human monoculear phagocytes. J. Leukoc. Biol. 45, 29-34 CAROSELLA, E. D., PAUL, P., MOREAU, P. and ROUAS-FREISS, N. (2000): HLA-G and HLA-E: fundamental and pathophysiological aspects. Immunol. Today 21, 532-534 CARRETERO, M., ESCAMEZ, M. J., GARCIA, M., DUARTE, B., HOLGUIN, A., RETAMOSA, L., JORCANO, J. L., RIO, M. D. and LARCHER, F. (2008): In vitro and in vivo wound healing-promoting activities of human cathelicidin LL-37. J. Invest. Dermatol. 128, 223-236

ANNEX
218
CHASE, H. B. (1954): Growth of the hair. Physiol. Rev. 34, 113-126 CHEN, P. W., MELLON, J. K., MAYHEW, E., WANG, S., HE, Y. G., HOGAN, N. and NIEDERKORN, J. Y. (2007): Uveal melanoma expression of indoleamine 2,3-deoxygenase: Establishment of an immune privileged environment by tryptophan depletion. Exp. Eye Res. 85, 617-625 CHRISTIANO, A. M. (2004): Epithelial stem cells: stepping out of their niche. Cell 118, 530-532 CHRISTOPH, T., MUELLER-ROEVER, S., AUDRING, H., TOBIN, D. J., HERMES, B., COTSARELIS, G., RUECKERT, R. and PAUS, R. (2000): The human hair follicle immune system: cellular composition and immune privilege. Br. J. Dermatol. 142, 862-873 CHRISTY, A. L. and BROWN, M. A. (2007): The multitasking mast cell: positive and negative roles in the progression of autoimmunity. J. Immunol. 179, 2673-2679 CHRONNELL, C. M., GHALI, L. R., ALI, R. S., QUINN, A. G., HOLLAND, D. B., BULL, J. J., CUNLIFFE, W. J., MCKAY, I. A., PHILPOTT, M. P. and MUELLER-ROEVER, S. (2001): Human ß defensin-1 and -2 expression in human pilosebaceous units: upregulation in acne vulgaris lesions. J. Invest. Dermatol. 117, 1120-1125 CLARK, D. A., KEIL, A., CHEN, Z., MARKERT, U., MANUEL, J. and GORCZYNSKI, R. M. (2003): Placental trophoblast from successful human pregnancies expresses the tolerance signaling molecule, CD200 (OX-2). Am. J. Reprod. Immunol. 50, 187-195 CLARK, L. W. E. (1936): The problem of the claw in primates. Proc. Zool. Soc. Part II, 1-24 CLARK, M. J., GAGNON, J., WILLIAMS, A. F. and BARCLAY, A. N. (1985): MRC OX-2 antigen: a lymphoid/neuronal membrane glycoprotein with a structure like a single immunoglobulin light chain. EMBO J. 4, 113-118

REFERENCES
219
COBBOLD, S. P., ADAMS, E., GRACA, L., DALEY, S., YATES, S., PATERSON, A., ROBERTSON, N. J., NOLAN, K. F., FAIRCHILD, P. J. and WALDMANN, H. (2006): Immune privilege induced by regulatory T cells in transplantation tolerance. Immunol. Rev. 213, 239-255 CONE, R. D., LU, D., KOPPULA, S., VAGE, D. I., KLUNGLAND, H., BOSTON, B., CHEN, W., ORTH, D. N., POUTON, C. and KESTERSON, R. A. (1996): The melanocortin receptors: agonists, antagonists, and the hormonal control of pigmentation. Recent. Prog. Horm. Res. 51, 287-318 COOPER, A., ROBINSON, S. J., PICKARD, C., JACKSON, C. L., FRIEDMANN, P. S. and HEALY, E. (2005): α-melanocyte-stimulating hormone suppresses antigen-induced lymphocyte proliferation in humans independently of melanocortin 1 receptor gene status. J. Immunol. 175, 4806-4813 COTSARELIS, G. (2006a): Epithelial stem cells: a folliculocentric view. J. Invest. Dermatol. 126, 1459-1468 COTSARELIS, G. (2006b): Gene expression profiling gets to the root of human hair follicle stem cells. J. Clin. Invest. 116, 19-22 COTSARELIS, G. and MILLAR, S. E. (2001): Towards a molecular understanding of hair loss and its treatment. Trends Mol. Med. 7, 293-301 COTSARELIS, G., SUN, T. T. and LAVKER, R. M. (1990): Label-retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell 61, 1329-1337 COTSARELIS, G., KAUR, P., DHOUAILLY, D., HENGGE, U. and BICKENBACH, J. (1999): Epithelial stem cells in the skin: definition, markers, localization and functions. Exp. Dermatol. 8, 80-88 CRISSMAN, R. S., WARDEN, R. J., SICILIANO, D. A., KLEIN, B. G., RENEHAN, W. E., JACQUIN, M. F. and RHOADES, R. W. (1991): Numbers of axons innervating mystacial vibrissa follicles in newborn and adult rats. Somatosens. Mot. Res. 8, 103-109

ANNEX
220
CURRIER, A. R., ZIEGLER, M. H., RILEY, M. M., BABCOCK, T. A., TELBIS, V. P. and CARLIN, J. M. (2000): Tumor necrosis factor-α and lipopolysaccharide enhances interferon-induced antichlamydial indoleamine dioxygenase activity independently. J. Interferon Cytokine Res. 20, 369-376 CYGAN, J. A., JOHNSON, R. L. and MCMAHON, A. P. (1997): Novel regulatory interactions revealed by studies of murine limb pattern in Wnt-7a and En-1 mutants. Development 124, 5021-5032 D`ORAZIO, T. J. and NIEDERKORN, J. Y. (1998): A novel role for TGF-β and IL-10 in the induction of immune privilege. J. Immunol. 160, 2089-2098 DANFORTH, C. H. (1925a): Hair in its relation to questions of homology and phylogeny. Am. J. Anat. 36, 47-68 DANFORTH, D. H. (1925b): Studies on hair, with specil reference to hypertrichosis. Arch. Derm. Syph. 11, 494-508 DAVIDSON, P. and HARDY, M. H. (1952): The development of mouse vibrissae in vivo and in vitro. J. Anat. 86, 342-356 DAWBER, R. P. (1980): The ultrastructure and growth of human nails. Arch. Dermatol. Res. 269, 197-204 DE BERKER, D. and ANGUS, B. (1996): Proliferative compartments in the normal nail unit. Br. J. Dermatol. 135, 555-559 DEMEURE, C. E., BRAHIMI, K., HACINI, F., MARCHAND, F., PERONET, R., HUERRE, M., ST-MEZARD, P., NICOLAS, J. F., BREY, P., DELESPESSE, G. and MERCHERI, S. (2005): Anopheles mosquito bites activate cutaneous mast cells leading to a local inflammatory response and lymph node hyperplasia. J. Immunol. 174, 3932-3940 DEPINAY, N., HACINI, F., BEGHDADI, W., PERONET, R. and MECHERI, S. (2006): Mast cell-dependent down-regulation of antigen-specific immune responses by mosquito bites. J. Immunol. 176, 4141-4146

REFERENCES
221
DERYNCK, R., JARETT, J. A., CHEN, E. Y., EATON, D. H., BELL, J. R., ASSOIAN, R. K., ROBERTS, A. B., SPORN, M. B. and GOEDDEL, D. V. (1985): Human transforming growth factor-ß complementary DNA sequence and expression in normal and transformed cells. Nature 316, 701-705 DERYNCK, R., LINDQUIST, P. B., LEE, A., WEN, D., TAMM, J., GRAYEAR, J. L., RHEE, L., MASON, A. J., MILLER, D. A., COFFEY, R. J., MOSES, H. L. and CHEN, E. Y. (1988): A new type of transforming growth factor-ß, TGF-ß3. EMBO J. 7, 3737-3743 DI NARDO, A., VITIELLO, A. and GALLO, R. L. (2003): Cutting edge: mast cell antimicrobial activity is mediated by expression of cathelicidin antimicrobial peptide. J. Immunol. 170, 2274-2278 DOERFL, J. (1982): The musculature of the mystacial vibrissae of the white mouse. J. Anat. 135, 147-154 DORSCHNER, R. A., LOPEZ-GARCIA, B., MASSIE, J., KIM, C. and GALLO, R. L. (2004): Innate immune defense of the nail unit by antimicrobial peptides. J. Am. Acad. Dermatol. 50, 343-348 DRY, F. W. (1926): The coat of the mouse (Mus musculus). J. Genetics. 16, 287-340 DUERDEN, J. E. R., M.I.F. (1924): Development of the merino wool fibre. S. Afric. J. Sci. 21, 480-497 DUN, R. B. and FRASER, A. S. (1958): Selection for an invariant character; vibrissa number in the house mouse. Nature 181, 1018-1019 EHRLEBACHER, A. (2001): Why isn't the fetus rejected? Curr. Opin. Immunol. 13, 590-593 EICHMUELLER, S., VAN DER VEEN, C., MOLL, I., HERMES, B., HOFFMANN, U., MUELLER-ROEVER, S. and PAUS, R. (1998): Clusters of perifollicular macrophages in normal murine skin: physiological degeneration of selected hair follicles by programmed organ deletion. J. Histochem. Cytochem. 46, 361-370

ANNEX
222
EIPPER, B. A. and MAINS, R. E. (1980): Strcture and biosynthesis of pro-adrenocorticotropin/endorphin and related peptides. Endocr. Rev. 1, 1-27 FABRI, Z., TOPHAM, D. J. and FEE, D. (1995): TGF-β 2 decreases migration of lymphocytes in-vitro and homing of cells into the central nervous system in-vivo. J. Immunol. 155, 325-332 FALLARINO, T., GROHMANN, U., HWANG, K., ORABONA, C., VACCA, C., BIANCHI, R., BELLADONNA, M., FIORETTI, M., ALEGRE, M. and PUCCETTI, P. (2003): Modulation of tryptophan catabolism by regulatory T cells. Nat. Immunol. 4, 1206-1212 FALLARINO, F., VACCA, C., ORABONA, C., BELLADONNA, M. L., BIANCHI, R., MARSHALL, B., KESKIN, D. B., MELLOR, A. L., FIORETTI, M. C., GROHMANN, U. and PUCCETTI, P. (2002): Functional expression of indoleamine 2,3-dioxygenase by murine CD8 alpha(+) dendritic cells. Int. Immunol. 14, 65-68 FERGUSON, T. A. and GRIFFITH, T. S. (2006): A vision of cell death: Fas ligand and immune privilege 10 years later. Immunol. Rev. 213, 228-238 FILIPPINI, A., RICCIOLI, A., PADULA, F., LAURETTI, P., D'ALESSIO, A., CESARIS, P., GANDINI, L., LENZI, A. and ZIPARO, E. (2001): Control and impairment of immune privilege in the testis and in semen. Hum. Reprod. Update. 7, 444-449 FLECKMAN, P., LEE, J. and ASTION, M. L. (1997): Nail-Tutor: an image-based personal computer program that teaches the anatomy, patterns of pathology, and disorders of the nails. Dermatol. Online J. 3, 2 FOITZIK, K., LINDNER, G., MUELLER-ROEVER, S., MAURER, M., BOTCHKAREVA, N., BOTCHKAREV, V., HANDJISKI, B., METZ, M., HIBINO, T., SOMA, T., DOTTO, G. P. and PAUS, R. (2000): Control of murine hair follicle regression (catagen) by TGF-β1 in vivo. FASEB J. 14, 752-760 FOITZIK, K., PAUS, R., DOETSCHMAN, T. and DOTTO, G. P. (1999): The TGF-β2 isoform is both a required and sufficient inducer of murine hair follicle morphogenesis. Dev. Biol. 212, 278-289

REFERENCES
223
FOURNEL, S., AGUERRE-GIRR, M., HUC, X., LENFANT, F., ALAM, A., TOUBERT, A., BENSUSSAN, A. and LE BOUTEILLER, P. (2000): Cutting edge: soluble HLA-G1 triggers CD95/CD95 ligand-mediated apoptosis in activated CD8+ cells by interacting with CD8. J. Immunol. 164, 6100-6104 FREYSCHMIDT-PAUL, P., MCELWEE, K., HOFFMANN, R., SUNDBERG, J., VITACOLONNA, M., KISSLING, S. and ZOLLER, M. (2006): Interferon-γ-deficient mice are resistant to the development of alopecia areata. Br. J. Dermatol. 155, 515-521 FROHM, M., AGERBERTH, B., AHANGARI, G., STAHLE-BACKDAHL, M., LIDEN, S., WIGZELL, H. and GUDMUNDSSON, G. H. (1997): The expression of the gene coding for the antibacterial peptide LL-37 is induced in human keratinocytes during inflammatory disorders. J. Biol. Chem. 272, 15258-15263 FUCHS, E. (2007): Scratching the surface of skin development. Nature 445, 834-842 FUZZI, B., RIZZO, R., CRISCUOLI, L., NOCI, I., MELCHIORRI, L., SCARSELLI, B., BENCINI, E., MENICUCCI, A. and BARICORDI, O. (2002): HLA-G expression in early embryos is a fundamental prerequisite for the obtainment of pregnancy. Euro. J. Immunol. 32, 311-315 GALAT, A., RIVIERE, S. and BOUET, F. (1993): Purification of macrophage migration inhibitory factor (MIF) from bovine brain cytosol. FEBS Lett. 319, 233 GALLI, S. J., NAKAE, S. and TSAI, M. (2005): Mast cells in the development of adaptive immune responses. Nat. Immunol. 6, 135-142 GALLO, R. L. (2008): Sounding the alarm: multiple functions of host defense peptides. J. Invest. Dermatol. 128, 5-6 GAO, G. F., TORMO, J., GERTH, U. C., WYER, J. R., MCMICHAEL, A. J., STUART, D. I., BELL, J. I., JONES, E. Y. and JAKOBSEN, B. K. (1997): Crystal structure of the complex between human CD8α (alpha) and HLA-A2. Nature 387, 630-634 GEORGE, M. and VAUGHAN, J. H. (1962): In vitro cell migration as a model for delayed hypersensitivity. Proc. Soc. Exp. Biol. Med. 111, 514-521

ANNEX
224
GIBBS (1938): A study of the development of the skin and hair of the Australian opossum, Trichosurus vulpecula. Proc. zool. Soc. Lond. 108B, 611-648 GILFILLAN, A. M. and TKACZYK, C. (2006): Integrated signalling pathways for mast-cell activation. Nat. Rev. Immunol. 6, 218-230 GILHAR, A. and KALISH, R. S. (2006): Alopecia areata: a tissue specific autoimmune disease of the hair follicle. Autoimmun. Rev. 5, 64-69 GILHAR, A., KARN, Y., ASSY, B. and KALISH, R. S. (2005): Alopecia areata induced in C3H/heJ mice by interferon-γ: evidence for loss of immune privilege. J. Invest. Dermatol. 124, 288-289 GILHAR, A., PAUS, R. and KALISH, R. S. (2007): Lymphocytes, neuropeptides, and genes involved in alopecia areata. J. Clin. Invest. 117, 2019-2027 GILHAR, A., ULLMANN, Y., BERKUTZKI, T., ASSY, B. and KALISH, R. S. (1998): Autoimmune hair loss (alopecia areata) transferred by T lymphocytes to human scalp explants on SCID mice. J. Clin. Invest. 101, 62-67 GLAESER, R., HARDER, J., LANGE, H., BARTELS, J., CHRISTOPHERS, E. and SCHROEDER, J. M. (2005): Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat. Immunol. 6, 57-64 GODWIN, A. R. and CAPECCHI, M. R. (1998): Hoxc13 mutant mice lack external hair. Genes Dev. 12, 11-20 GOSLINGS, W. R., PRODEUS, A. P., STREILEIN, J. W., CARROLL, M. C., JAEGER, M. J. and TAYLOR, A. W. (1998): A small molecular weight factor in aqueous humor acts on C1q to prevent antibody-dependent complement activation. Invest. Ophthalmol. Vis. Sci. 39, 989-995 GREENE, H. (1947): The use of the mouse eye in transplantation experiments. Cancer Res. 7, 491

REFERENCES
225
GRUNEBERG, H. (1943): The development of some external features in mouse embryos. J. Hered. 34, 89-92 HADLEY, M. E. and DORR, R. T. (2006): Melanocortin peptide therapeutics: historical milestones, clinical studies and commercialization. Peptides 27, 921-930 HAINZ, U., JUERGENS, B. and HEITGER, A. (2007): The role of indoleamine 2,3-dioxygenase in transplantation. Transpl. Int. 20, 118-127 HALATA, Z. (1993): Sensory innervation of the hairy skin (light- and electronmicroscopic study). J. Invest. Dermatol. 101, 75S-81S HARDER, J., BARTELS, J., CHRISTOPHERS, E. and SCHROEDER, J. M. (1997): A peptide antibiotic from human skin. Nature 387, 861 HARDER, J., BARTELS, J., CHRISTOPHERS, E. and SCHROEDER, J. M. (2001): Isolation and characterization of human β-defensin-3, a novel human inducible peptide antibiotic. J. Biol. Chem. 276, 5707-5713 HARDY, M. H. (1949): The development of mouse hair in vitro with some observations on pigmentation. J. Anat. 83, 364-384, 363 pl HARDY, M. H. (1951): The development of pelage hairs and vibrissae from skin in tissue culture. Ann. N. Y. Acad. Sci. 53, 546-561 HARRIES, M. J., MEYER, K. C. and PAUS, R. (2009): Hair loss as a result of cutaneous autoimmunity: Frontiers in the immunopathogenesis of primary cicatricial alopecia. Autoimmun. Rev. (in press) HARRIES, M. J., SINCLAIR, R. D., MACDONALD-HULL, S., WHITING, D. A., GRIFFITHS, C. E. and PAUS, R. (2008): Management of primary cicatricial alopecias: options for treatment. Br. J. Dermatol. 159, 1-22 HARRIST, T. J., RUITER, D. J., MIHM, M. C., JR. and BHAN, A. K. (1983): Distribution of major histocompatibility antigens in normal skin. Br. J. Dermatol. 109, 623-633

ANNEX
226
HASKELL-LUEVANO, C., SAWYER, T. K., HENDRATA, S., NORTH, C., PANAHINIA, L., STUM, M., STAPLES, D. J., CASTRUCCI, A. M., HADLEY, M. F. and HRUBY, V. J. (1996): Truncation studies of α-melanotropin peptides identify tripeptide analogues exhibition prolonged agonist bioactivity. Peptides 17, 995-1002 HAUSMANN, L. A. (1924): Further studies of the relationships of the structural characteristics of mammalian hair. Amer. Nat. 58, 544-557 HEAD, J. and BILLINGHAM, R. (1985): Immunologically privileged sites in transplantation immunology and oncology. Persp. Biol. Med. 29, 115-131 HEADINGTON, J. T. (1984): Transverse microscopic anatomy of the human scalp. A basis for a morphometric approach to disorders of the hair follicle. Arch. Dermatol. 120, 449-456 HEID, H. W., MOLL, I. and FRANKE, W. W. (1988): Patterns of expression of trichocytic and epithelial cytokeratins in mammalian tissues. II. Concomitant and mutually exclusive synthesis of trichocytic and epithelial cytokeratins in diverse human and bovine tissues (hair follicle, nail bed and matrix, lingual papilla, thymic reticulum). Differentiation 37, 215-230 HERMES, B. and PAUS, R. (1998): ´Vernarbende` Alopezien Anmerkungen zur Klassifikation, Differentialdiagnose und Pathobiologie. Hautarzt 49, 462-472 HIGASHI, N. (1968): Melanocytes of nail matrix and nail pigmentation. Arch. Dermatol. 97, 570-574 HIGASHI, N. and SAITO, T. (1969): Horizontal distribution of the dopa-positive melanocytes in the nail matrix. J. Invest. Dermatol. 53, 163-165 HIGUCHI, K. and HAYAISHI, O. (1967): Enzymatic formation of d-kynurenine from d-tryptophan. Arch. Biochem. Biophys. 120, 397-403

REFERENCES
227
HOBBS NK, BONDAREVA AA, S, B., MR, C. and EE, S. (2002): Removing the vertebrate specific TBP N terminus disrupts placental ß2-dependent interactions with the maternal immune system. Cell 12, 43-54 HOFFMANN, R., EICHELER, W., WENZEL, E. and HAPPLE, R. (1997): Interleukin-1β-induced inhibition of hair growth in vitro is mediated by cyclic AMP. J. Invest. Dermatol. 108, 40-42 HORNE, K. A., JAHODA, C. A. and OLIVER, R. F. (1986): Whisker growth induced by implantation of cultured vibrissa dermal papilla cells in the adult rat. J. Embryol. Exp. Morphol. 97, 111-124 IBRAHIM, L. and WRIGHT, E. A. (1975): The growth of rats and mice vibrissae under normal and some abnormal conditions. J. Embryol. Exp. Morphol. 33, 831-844 IBRAHIM, L. and WRIGHT, E. A. (1978a): The effect of a single plunking at different times in the hair cycle on the growth of individual mouse vibrissae. Br. J. Dermatol. 99, 365-370 IBRAHIM, L. and WRIGHT, E. A. (1978b): The long term effect of repeated pluckings on the function of the mouse vibrissal follicles. Br. J. Dermatol. 99, 371-376 IIDA, M., IHARA, S. and MATSUZAKI, T. (2007a): Follicular epithelia and dermal papillae of mouse vibrissal follicles qualitatively change their hair-forming ability during anagen. Differentiation 75, 371-381 IIDA, M., IHARA, S. and MATSUZAKI, T. (2007b): Hair cycle-dependent changes of alkaline phosphatase activity in the mesenchyme and epithelium in mouse vibrissal follicles. Dev. Growth Differ. 49, 185-195 ITO, N., ITO, T., KROMMINGA, A., BETTERMANN, A., TAKIGAWA, M., KEES, F., STRAUB, R. H. and PAUS, R. (2005a): Human hair follicles display a functional equivalent of the hypothalamic-pituitary-adrenal axis and synthesize cortisol. FASEB J. 19, 1332-1334

ANNEX
228
ITO, T., ITO, N., BETTERMANN, A., TOKURA, Y., TAKIGAWA, M. and PAUS, R. (2004): Collapse and restoration of MHC class-I-dependent immune privilege: exploiting the human hair follicle as a model. Am. J. Pathol. 164, 623-634 ITO, T., ITO, N., SAATHOFF, M., BETTERMANN, A., TAKIGAWA, M. and PAUS, R. (2005b): Interferon-gamma is a potent inducer of catagen-like changes in cultured human anagen hair follicles. Br. J. Dermatol. 152, 623-631 ITO, T., ITO, N., SAATHOFF, M., STAMPACHIACCHIERE, B., BETTERMANN, A., BULFONE-PAUS, S., TAKIGAWA, M., NICKOLOFF, B. J. and PAUS, R. (2005c): Immunology of the human nail apparatus: the nail matrix is a site of relative immune privilege. J. Invest. Dermatol. 125, 1139-1148 ITO, T., ITO, N., SAATOFF, M., HASHIZUME, H., FUKAMIZU, H., NICKOLOFF, B. J., TAKIGAWA, M. and PAUS, R. (2008a): Maintenance of hair follicle immune privilege is linked to prevention of NK cell attack. J. Invest. Dermatol. 128, 1196-1206 ITO, T., MEYER, K. C., ITO, N. and PAUS, R. (2008b): Immune privilege and the skin. Curr. Dir. Autoimmun. 10, 27-52 ITO, T., SAATHOFF, M., NICKOLOFF, B. J., TAKIGAWA, M. and PAUS, R. (2005d): Novel aspects of hair follicle immune privilege and their relevance to alopecia areata. J. Invest. Dermatol. 124, A:103 JAHANYAR, J., KOERNER, M. M., LOEBE, M., YOUKER, K. A., TORRE-AMIONE, G. and NOON, G. P. (2008): The role of mast cells after solid organ transplantation. Transplantation 85, 1365-1371 JAHODA, C. A. (1992): Induction of follicle formation and hair growth by vibrissa dermal papillae implanted into rat ear wounds: vibrissa-type fibres are specified. Development 115, 1103-1109 JAHODA, C. A. (2003): Cell movement in the hair follicle dermis - more than a two-way street? J. Invest. Dermatol. 121, DOI 10.1111/j.1523-1747.2003.12585.x

REFERENCES
229
JAHODA, C. A. and REYNOLDS, A. J. (1996): Dermal-epidermal interactions. Adult follicle-derived cell populations and hair growth. Dermatol. Clin. 14, 573-583 JAHODA, C. A., HORNE, K. A. and OLIVER, R. F. (1984): Induction of hair growth by implantation of cultured dermal papilla cells. Nature 311, 560-562 JAHODA, C. A., MAUGER, A., BARD, S. and SENGEL, P. (1992): Changes in fibronectin, laminin and type IV collagen distribution relate to basement membrane restructuring during the rat vibrissa follicle hair growth cycle. J. Anat. 181, 47-60 JALILI, R. B., RAYAT, G. R., RAJOTTE, R. V. and GHAHARY, A. (2007): Suppression of islet allogeneic immune response by indoleamine 2,3 dioxygenase-expressing fibroblasts. J. Cell Physiol. 213, 137-143 JAWORSKY, C., KLIGMAN, A. M. and MURPHY, G. F. (1992): Characterization of inflammatory infiltrates in male pattern alopecia: implications for pathogenesis. Br. J. Dermatol. 127, 239-246 JIARAVUTHISAN, M. M., SASSEVILLE, D., VENDER, R. B., MURPHY, F. and MUHN, C. Y. (2007): Psoriasis of the nail: anatomy, pathology, clinical presentation, and a review of the literature on therapy. J. Am. Acad. Dermatol. 57, 1-27 JOHNSON, E. and EBLING, F. J. (1964): The effect of plucking hairs during different phases of the follicular cyle. J. Embryol. Exp. Morphol. 12, 465 JOHNSON, M., COMAISH, J. S. and SHUSTER, S. (1991): Nail is produced by the normal nail bed: a controversy resolved. Br. J. Dermatol. 125, 27-29 KAUSER, S., THODY, A. J., SCHALLREUTER, K. U., GUMMER, C. L. and TOBIN, D. J. (2005): A fully functional proopiomelanocortin/melanocortin-1 receptor system regulates the differentiation of human scalp hair follicle melanocytes. Endocrinology 146, 532-543 KAWAKAMI, Y., WADA, N., NISHIMATSU, S. and NOHNO, T. (2000): Involvement of frizzled-10 in Wnt-7a signaling during chick limb development. Dev. Growth Differ. 42, 561-569

ANNEX
230
KEZUKA, T. and STREILEIN, J. W. (2000): Analysis of in vivo regulatory properties of T cells activated in vitro by TGF ß2-treated antigen presenting cells. Invest. Ophtalmol. Vis. Sci. 41, 1410-1421 KHALIL, N. (1999): TGF-ß: from latent to active. Microbes Infect. 1, 1255-1263 KITAHARA, T. and OGAWA, H. (1993): Coexpression of keratins characteristic of skin and hair differentiation in nail cells. J. Invest. Dermatol. 100, 171-175 KITAHARA, T. and OGAWA, H. (1994): Variation of differentiation in nail and bovine hoof cells. J. Invest. Dermatol. 102, 725-729 KITAHARA, T. and OGAWA, H. (1997): Cellular features of differentiation in the nail. Microsc. Res. Tech. 38, 436-442 KLOEPPER, J. E., TIEDE, S., BRINCKMANN, J., REINHARDT, D. P., MEYER, W., FAESSLER, R. and PAUS, R. (2008): Immunophenotyping of the human bulge region: the quest to define useful in situ markers for human epithelial hair follicle stem cells and their niche. Exp. Dermatol. 17, 592-609 KOCZULLA, R., VON DEGENFELD, G., KUPATT, C., KROTZ, F., ZAHLER, S., GLOE, T., ISSBRUCKER, K., UNTERBERGER, P., ZAIOU, M., LEBHERZ, C., KARL, A., RAAKE, P., PFOSSER, A., BOEKSTEGERS, P., WELSCH, U., HIEMSTRA, P. S., VOGELMEIER, C., GALLO, R. L., CLAUSS, M. and BALS, R. (2003): An angiogenic role for the human peptide antibiotic LL-37/hCAP-18. J. Clin. Invest. 111, 1665-1672 KOLLAR, E. J. (1970): The induction of hair follicles by embryonic dermal papillae. J. Invest. Dermatol. 55, 374-378 KOWATS, S. (1990): HLA-G, expressed in human trophoblasts. Science 248, 220-223

REFERENCES
231
KULKARNI, A. B., HUH, C. G., BECKER, D., GEISER, A., LYGHT, M., FLANDERS, K. C., ROBERTS, A. B., SPORN, M.B., WARD, J. M., KARLSSON, S. (1993): Transforming growth factor ß 1 null mutation in mice causes excessive inflammatory response and early death. Proc. Natl. Acad. Sci USA. 90, 770-774 KWIDZINSKI, E. and BECHMANN, I. (2007): IDO expression in the brain: a double-edged sword. J. Mol. Med. 85, 1351-1359 LAKO, M., ARMSTRONG, L., CAIRNS, P. M., HARRIS, S., HOLE, N. and JAHODA, C. A. (2002): Hair follicle dermal cells repopulate the mouse haematopoietic system. J. Cell Sci. 115, 3967-3974 LAMPSON, L. A. and FISHER, C. A. (1984): Weak HLA and ß 2-microglobulin expression of neuronal cell lines can be modulated by interferon. Proc. Natl. Acad. Sci. U S A. 81, 6476-6480 LANGBEIN, L., ROGERS, M. A., PRAETZEL, S., AOKI, N., WINTER, H. and SCHWEIZER, J. (2002): A novel epithelial keratin, hK6irs1, is expressed differentially in all layers of the inner root sheath, including specialized huxley cells (Fluegelzellen) of the human hair follicle. J. Invest. Dermatol. 118, 789-799 LANGBEIN, L., ROGERS, M. A., WINTER, H., PRAETZEL, S., BECKHAUS, U., RACKWITZ, H. R. and SCHWEIZER, J. (1999): The catalog of human hair keratins. I. Expression of the nine type I members in the hair follicle. J. Biol. Chem. 274, 19874-19884 LANGBEIN, L., ROGERS, M. A., WINTER, H., PRAETZEL, S. and SCHWEIZER, J. (2001): The catalog of human hair keratins. II. Expression of the six type II members in the hair follicle and the combined catalog of human type I and II keratins. J. Biol. Chem. 276, 35123-35132 LANIER, L. L. (2005): NK cell recognition. Annu. Rev. Immunol. 23, 225-273

ANNEX
232
LARRICK, J. W., HIRATA, M., BALINT, R. F., LEE, J., ZHONG, J. and WRIGHT, S. C. (1995): Human CAP18: a novel antimicrobial lipopolysaccharide-binding protein. Infect. Immun. 63, 1291-1297 LASS, J. H., WALTER, E. I., BURRIS, T. E., GROSSNIKLAUS, H. E., ROAT, M. I., SKELNIK, D. L., NEEDHAM, L., SINGER, M. and MEDOF, M. E. (1990): Expression of two molecular forms of the complement decay-accelerating factor in the eye and lacrimal gland. Invest. Ophthalmol. Vis. Sci. 31, 1136-1148 LE BOUTEILLER, P. (1994): HLA class I chromosomal region, genes, and products: facts and questions. Crit. Rev. Immunol. 14, 89-129 LEDER, L.D. (1964): Uber die selektive fermentcytochemische Darstellung von neutrophilen myeloischen Zellen und Gewebsmastzellen im Paraffinschnitt. Klin. Wschr. 42, 553 LE DISCORDE, M., MOREAU, P., SABATIER, P., LEGEAIS, J. M. and CAROSELLA, E. D. (2003): Expression of HLA-G in human cornea, an immune-privileged tissue. Hum. Immunol. 64, 1039-1044 LE GAL, F. A., RITEAU, B., SEDLIK, C., KHALI-DAHER, I., MENIER, C., DAUSSET, J., GUILLET, J., CAROSELLA, E. D. and ROUAS-FREISS, N. (1999): HLA-G-mediated inhibition on antigen-specific cytotoxic T lymphocytes. Int. Immunol. 11, 7-18 LEE, N. (1998): HLA-E is a major ligand for the natural killer inhibitory receptor CD94/NKG2A. Proc. Natl. Acad. Sci. 31, 724-725 LEECH, M., METZ, C. N., HALL, P., HUTCHINSON, P., GIANIS, K., SMITH, M., WEEDON, H., HOLDSWORTH, S., BUCALA, R. and MORAND, E. F. (1999): Macrophage migration inhibitory factor in rheumatoid arthritis: evidence of pro-inflammatory function and regulation by glucocorticoids. Arthritis Rheum. 42, 1601-1608 LETTERIO, J. J. and ROBERTS, A. B. (1998): Regulation of immune responses by TGF-ß. Annu. Rev. Immunol. 16, 137-161

REFERENCES
233
LEWIS, B. L. (1954): Microscopic studies of fetal and mature nail and surrounding soft tissue. Am. Arch. Derm. Syphil. 70, 733-747 LI, Y., TREDGET, E. E. and GHAHARY, A. (2004): Cell surface expression of MHC class I antigen is suppressed in IDO genetically modified keratinocytes: implications in allogeneic skin substitute engraftment. Hum. Immunol. 65, 114-123 LI, Y., TREDGET, E. E., GHAFFARI, A., LIN, X., KILANI, R. T. and GHAHARY, A. (2006): Local expression of indoleamine 2,3-dioxygenase protects engraftment of xenogeneic skin substitute. J. Invest. Dermatol. 126, 128-136 LIN, M. H. and KOPAN, R. (2003): Long-range, nonautonomous effects of activated Notch1 on tissue homeostasis in the nail. Dev. Biol. 263, 343-359 LINDNER, G., BOTCHKAREV, V. A., BOTCHKAREVA, N. V., LING, G., VAN DER VEEN, C. and PAUS, R. (1997): Analysis of apoptosis during hair follicle regression (catagen). Am. J. Pathol. 151, 1601-1617 LINK, R. E., PAUS, R., STENN, K. S., KUKLINSKA, E. and MOELLMANN, G. (1990): Epithelial growth by rat vibrissae follicles in vitro requires mesenchymal contact via native extracellular matrix. J. Invest. Dermatol. 95, 202-207 LIPTON, J. M. and CATANIA, A. (1997): Anti-inflammatory actions of the neuroimmunomodulator α-MSH. Immunol. Today. 18, 140-145 LITTLE, J. C., WESTGATE, G. E., EVANS, A. and GRANGER, S. P. (1994): Cytokine gene expression in intact anagen rat hair follicles. J. Invest. Dermatol. 103, 715-720 LJUNGGREN, H. G. (1991): The RMA-S lymphoma mutant: consequences of a peptide loading defect on immunological recognition and graft rejection. Int. J. Cancer Suppl. 6, 38-44

ANNEX
234
LOBO-YEO, A., SENALDI, G., PORTMANN, B., MOWAR, A., MIELI-VERGANI, G. and VERGANI, D. (1990): Class I and class II major histocompatibility complex antigen expression on hepatocytes: a study in children with liver disease. Hepatology 12, 224-232 LONG, E. O. (1999): Regulation of immune responses through inhibitory receptors. Annu. Rev. Immunol. 17, 875-904 LU, Z., HASSE, S., BODO, E., ROSE, C., FUNK, W. and PAUS, R. (2007): Towards the development of a simplified long-term organ culture method for human scalp skin and its appendages under serum-free conditions. Exp. Dermatol. 16, 37-44 LUGER, T. A. and BRZOSKA, T. (2007): α-MSH related peptides: a new class of anti-inflammatory and immunomodulating drugs. Ann. Rheum. Dis. 66, Suppl 3, iii52-55 LUGER, T. A., BRZOSKA, T., SCHOLZEN, T. E., KALDEN, D. H., SUNDERKOTTER, C., ARMSTRONG, C. and ANSEL, J. (2000): The role of α-MSH as a modulator of cutaneous inflammation. Ann. N. Y. Acad. Sci. 917, 232-238 LUGER, T. A., KALDEN, D., SCHOLZEN, T. E. and BRZOSKA, T. (1999): α-melanocyte-stimulating hormone as a mediator of tolerance induction. Pathobiology 67, 318-321 LUGER, T. A., SCHOLZEN, T., BRZOSKA, T., BECHER, E., SLOMINSKI, A. and PAUS, R. (1998): Cutaneous immunomodulation and coordination of skin stress responses by α-melanocyte-stimulating hormone. Ann. N. Y. Acad. Sci. 840, 381-394 LUGER, T. A., SCHOLZEN, T. E., BRZOSKA, T. and BOHM, M. (2003): New insights into the functions of α-MSH and related peptides in the immune system. Ann. N.Y. Acad. Sci. 994, 133-140 LYLE, S., CHRISTOFIDOU-SOLOMIDOU, M., LIU, Y., ELDER, D. E., ALBELDA, S. and COTSARELIS, G. (1999): Human hair follicle bulge cells are biochemically distinct and possess an epithelial stem cell phenotype. J. Invest. Dermatol. Symp. Proc. 4, 296-301

REFERENCES
235
LYNCH, M. H., O'GUIN, W. M., HARDY, C., MAK, L. and SUN, T. T. (1986): Acidic and basic hair/nail (´hard`) keratins: their colocalization in upper cortical and cuticle cells of the human hair follicle and their relationship to ´soft` keratins. J. Cell Biol. 103, 2593-2606 MA, D. and NIEDERKORN, J. Y. (1995): Transforming growth factor-β down-regulates major histocompatibility complex class I antigen expression and increases the susceptibility of uveal melanoma cells to natural killer cell-mediated cytolysis. Immunology 86, 263-269 MAGERL, M., KAUSER, S., PAUS, R. and TOBIN, D. J. (2002): Simple and rapid method to isolate and culture follicular papillae from human scalp hair follicles. Exp. Dermatol. 11, 381-385 MAGERL, M., PAUS, R., FARJO, N., MULLER-ROVER, S., PETERS, E. M., FOITZIK, K. and TOBIN, D. J. (2004): Limitations of human occipital scalp hair follicle organ culture for studying the effects of minoxidil as a hair growth enhancer. Exp. Dermatol. 13, 635-642 MAROTTE, L. R., RICE, F. L. and WAITE, P. M. (1992): The morphology and innervation of facial vibrissae in the tammar wallaby, Macropus eugenii. J. Anat. 180, 401-417 MASTROFRANCESCO, A., KOKO, A., STRPZYK, E., SCHILLER, M., ZOUBOULIS, C. C., PICARDO, M., LUGER, T. A. and BOEHM, M. (2007): Suppression of inflammatory responses in human sebocyte by K(D)PT, a peptide fragment related to the C-terminal domain of alpha-melanocyte-stimulating hormone. J. Invest. Dermatol. 127, S70 MAURER, M., FISCHER, E., HANDJISKI, B., V.STEBUT, E., ALGERMISSEN, B., BAVANDI, A. and PAUS, R. (1997): Activated skin mast cells are involved in murine hair follicle regression (catagen). Lab. Invest. 77, 319-332 MAURER, M., FISCHER, E. L., HANDJISKI, B., CZARNETZKI, B. M. and PAUS, R. (1994): Do mast cells play a role in hair follicle regressen (catagen)? J. Invest. Dermatol. 103, 446a MAURER, M. and METZ, M. (2005): The status quo and quo vadis of mast cells. Exp. Dermatol. 14, 923-929

ANNEX
236
MAURER, M., SEIDEL-GUYENOT, W., METZ, M., KNOP, J. and STEINBRINK, K. (2003a): Critical role of IL-10 in the induction of low zone tolerance to contact allergens. J. Clin. Invest. 112, 432-439 MAURER, M., THEOHARIDES, T., GRANSTEIN, R.D., BISCHOFF, S.C., BIENENSTOCK, J., HENZ, B., KOVANEN, P., PILIPONSKY, A. M., KAMBE, N., VLIAGOFTIS, H., LEVI-SCHAFFER, F., METZ, M., MIYACHI, Y-, BEFUS, D., FORSYTHE, P., KITAMURA, Y., GALLI, S. (2003b): What is the physiological function of mast cells? Exp. Dermatol. 12, 886-910 MCELWEE, K. J., KISSLING, S., WENZEL, E., HUTH, A. and HOFFMANN, R. (2003): Cultured peribulbar dermal sheath cells can induce hair follicle development and contribute to the dermal sheath and dermal papilla. J. Invest. Dermatol. 121, 1267-1275 MCLACHLAN, J. B., HART, J. P., PIZZO, S. V., SHELBURNE, C. P., STAATS, H. F., GUNN, M. D., ABRAHAM, S. N. (2003): Mast cell-derived tumor necrosis factor induces hypertrophy of draining lymph nodes during infection. Nat. Immunol. 4, 1199-1205 MECKLENBURG, L., PAUS, R., HALATA, Z., BECHTOLD, L. S., FLECKMAN, P. and SUNDBERG, J. P. (2004): FOXN1 is critical for onycholemmal terminal differentiation in nude (Foxn1) mice. J. Invest. Dermatol. 123, 1001-1011 MECKLENBURG, L., TOBIN, D. J., MUELLER-ROEVER, S., HANDJISKI, B., WENDT, G., PETERS, E. M., POHL, S., MOLL, I. and PAUS, R. (2000): Active hair growth (anagen) is associated with angiogenesis. J. Invest. Dermatol. 114, 909-916 MECKLENBURG, L., TYCHSEN, B. and PAUS, R. (2005): Learning from nudity: lessons from the nude phenotype. Exp. Dermatol. 14, 797-810 MEDAWAR, P. (1945): A second study of the behavior and fate of skin homografts in rabbits. J. Anat. Lond. 79, 157-176 MEDAWAR, P. B. (1948): Immunity to homologous grafted skin. III. The fate of skin homografts transplanted to the brain and to the anterior chamber of the eye. Br. J. Exp. Pathol. 29, 58-69

REFERENCES
237
MEKORI, Y. A. (2004): The mastocyte: the ´other` inflammatory cell in immunopathogenesis. J. Allergy Clin. Immunol. 114, 52-57 MELLOR, A. L. and MUNN, D. H. (2000): Immunology at the maternal-fetal interface: lessons for T cell tolerance and suppression. Annu. Rev. Immunol. 18, 367-391 MELLOR, A. L. and MUNN, D. H. (2008): Creating immune privilege: active local suppression that benefits friends, but protects foes. Nat. Rev. Immunol. 8, 74-80 METZ, M., SIEBENHAAR, F. and MAURER, M. (2008a): Mast cell functions in the innate skin immune system. Immunobiology 213, 251-260 METZ, M., GRIMBALDESTON, M. A., NAKAE, S., PHILIPONSKY, A. M., TSAI, M. and GALLI, S. J. (2007): Mast cells in the promotion and limitation of chronic inflammation. Immunol. Rev. 217, 304-328 MEYER, K. C., KLATTE, J. E., DINH, H. V., HARRIES, M. J., REITHMAYER, K., MEYER, W., SINCLAIR, R. and PAUS, R. (2008c): Evidence that the bulge region is a site of relative immune privilege in human hair follicles. Br. J. Dermatol. (in press) MEYER, W. (1999): Zur Bedeutung von Sinushaaren für unsere Haussäugetiere. Amtstierärztl. Dienst Lebensmittelkontr. 6, 294-296 MEYER, W. (2009a): Hair follicles in domesticated mammals, with comparison to laboratory animals and humans. In: MECKLENBURG, L., LINEK, M. and TOBIN, D.J. (eds.): Hair Loss Disorders in Domestic Animals. Blackwell Publ., Ames, Iowa, USA (in press) MEYER, W. (2009b): Haut und Hautorgane. In: WISSDORF, H., GERHARDS, H., HUSKAMP, B. and DEEGEN, E. (eds.): Praxisorientierte Anatomie und Propädeutik des Pferdes. 3rd Edition. Schlüter, Hannover, Germany (in press)

ANNEX
238
MEYER, W. and GOERGEN, S. (1986): Development of hair coat and skin glands in fetal porcine integument. J. Anat. (Lond.) 144, 201-220 MEYER, W. and ROEHRS, M. (1986): Von der Reptilienschuppe zu Feder und Haar - zur Evolution der Hautanhangsorgane. Dtsch. tierärztl. Wschr. 93, 245-251 MEYER, W., GODYNICKI, S. and TSUKISE, A. (2008a): Lectin histochemistry of the endothelium of blood vessels in the mammalian integument, with remarks on the endothelial glycocalyx and blood vessel nomenclature. Ann. Anat. 190, 264-276 MEYER, W., KLOEPPER, J. E. and FLEISCHER, L. (2008b): Demonstration of ß-glucan receptors in the skin of aquatic mammals - a preliminary report. Eur. J. Wildl. Res. 54, 479-486 MEYER, W., NEURAND, K. and SCHWARZ, R. (1980): Der Haarwechsel der Haussäugetiere. II. Topographischer Ablauf, Vergleich Haustier - Wildtier und Steuerungsmechanismen. Dtsch. tierärztl. Wschr. 87, 96-102 MEYER, W., SEEGERS, U., HERRMANN, J. and SCHNAPPER, A. (2003): Further aspects of the general antimicrobial properties of pinniped skin secretions. Dis. Aquat. Organ. 53, 177-179 MIDDLETON, D., CURRAN, M. and MAXWELL, L. (2002): Natural killer cells and their receptors. Transpl. Immunol. 10, 147-164 MOBINI, N., TAM, S. and KAMINO, H. (2005): Possible role of the bulge region in the pathogenesis of inflammatory scarring alopecia: lichen planopilaris as the prototype. J. Cutan. Pathol. 32, 675-679 MOLL, I., HEID, H. W., FRANKE, W. W. and MOLL, R. (1988): Patterns of expression of trichocytic and epithelial cytokeratins in mammalian tissues. III. Hair and nail formation during human fetal development. Differentiation 39, 167-184 MOLL, I. and MOLL, R. (1993): Merkel cells in ontogenesis of human nails. Arch. Dermatol. Res. 285, 366-371

REFERENCES
239
MOMBURG, F., ROELSE, J., HAMMERLING, G. J. and NEEFJES, J. J. (1994): Peptide size selection by the major histocompatibility complex-encoded peptide transporter. J. Exp. Med. 179, 1613-1623 MOOKHERJEE, N. and HANCOCK, R. E. (2007): Cationic host defense peptides: innate immune regulatory peptides as a novel approach for treating infections. Cell Mol. Life Sci. 64, 922-933 MOOKHERJEE, N., WILSON, H. L., DORIA, S., POPOWYCH, Y., FALSAFI, R., YU, J. J., LI, Y., VEATCH, S., ROCHE, F. M., BROWN, K. L., BRINKMAN, F. S., HOKAMP, K., POTTER, A., BABIUK, L. A., GRIEBEL, P. J. and HANCOCK, R. E. (2006): Bovine and human cathelicidin cationic host defense peptides similarly suppress transcriptional responses to bacterial lipopolysaccharide. J. Leukoc. Biol. 80, 1563-1574 MORETTI, G. M., GIACOMETTI, C., BOIDO, V. and REBORA, A. (1963): Histamine, serotonin and mast cells in the skin of the rat during the hair cycle. J. Invest. Dermatol. 40, 205-212 MORRIS, R. J., LIU, Y., MARLES, L., YANG, Z., TREMPUS, C., LI, S., LIN, J. S., SAWICKI, J. A. and COTSARELIS, G. (2004): Capturing and profiling adult hair follicle stem cells. Nat. Biotechnol. 22, 411-417 MORRISON, G. M., DAVIDSON, D. J. and DORIN, J. R. (1999): A novel mouse ß defensin, Defb2, which is upregulated in the airways by lipopolysaccharide. FEBS Lett. 442, 112-116 MOSCOSO, J., SERRANO-VELA, J. I., PACHECO, R. and ARNAIZ-VILLENA, A. (2006): HLA-G, -E and -F: allelism, function and evolution. Transpl. Immunol. 17, 61-64 MOSELEY, R. P. (1997): An immunocytochemical study of MHC class I expression on human Langerhans cells and melanocytes. J. Pathol. 181, 419-425 MUELLER-ROEVER, S., BULFONE-PAUS, S., HANDJISKI, B., WELKER, P., SUNDBERG, J. P., MCKAY, I. A., BOTCHKAREV, V. A. and PAUS, R. (2000): Intercellular adhesion molecule-1 and hair follicle regression. J. Histochem. Cytochem. 48, 557-568

ANNEX
240
MUELLER-ROEVER, S., HANDJISKI, B., VAN DER VEEN, C., EICHMUELLER, S., FOITZIK, K., MCKAY, I. A., STENN, K. S. and PAUS, R. (2001): A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Invest. Dermatol. 117, 3-15 MULLER, T. L., NGO-MULLER, V., REGINELLI, A., TAYLOR, G., ANDERSON, R. and MUNEOKA, K. (1999): Regeneration in higher vertebrates: limb buds and digit tips. Semin. Cell Dev. Biol. 10, 405-413 MUNN, A. L. and MUNN, D. H. (2006): Immune privilege: A recurrent theme in immunoregulation. Immunol. Rev. 213, 5-11 MUNN, D. H. and MELLOR, A. L. (2007): Indoleamine 2,3-dioxygenase and tumor-induced tolerance. J. Clin. Invest. 117, 1147-1154 MUNN, D. H., ZHOU, M., ATTWOOD, J. T., BONDAREV, I., CONWAY, S. J., MARSHALL, B., BROWN, C. and MELLOR, A. L. (1998): Prevention of allogeneic fetal rejection by tryptophan catabolism. Science 281, 1191-1193 MURDAN, S. (2002): Drug delivery to the nail following topical application. Int. J. Pharm. 236, 1-26 NAGAE, H., NAKANISHI, H., URANO, Y. and ARASE, S. (1995): Serial cultivation of human nail matrix cells under serum-free conditions. J. Dermatol. 22, 560-566 NAKAMURA, M. and ISHIKAWA, O. (2008): The localization of label-retaining cells in mouse nails. J. Invest. Dermatol. 128, 728-730 NAMBA, K., KITAICHI, N., NISHIDA, T. and TAYLOR, A. W. (2002): Induction of regulatory T cells by the immunomodulating cytokines alpha-melanocyte-stimulating hormone and transforming-growth factor-ß2. J. Leukoc. Biol. 72, 946-952 NIEDERKORN, J. Y. (2002): Immune privilege in the anterior chamber of the eye. Crit. Rev. Immunol. 22, 13-46

REFERENCES
241
NIEDERKORN, J. Y. (2003): Mechanisms of immune privilege in the eye and hair follicle. J. Investig. Dermatol. Symp. Proc. 8, 168-172 NIEDERKORN, J. Y. (2006): See no evil, hear no evil, do no evil: the lessons of immune privilege. Nat. Immunol. 7, 354-359 NIEDERKORN, J. Y. and WANG, S. (2005): Immune privilege of the eye and fetus: parallel universes? Transplantation 80, 1139-1144 NIEDERKORN, J. Y., CHIANG, E. Y., UNGCHUSRI, T. and STROYNOWSKI, I. (1999): Expression of a nonclassical MHC class Ib molecule in the eye. Transplantation 68, 1790-1799 NISSEN-MEYER, J. and NES, I. F. (1997): Ribosomally synthesized antimicrobial peptides: their function, structure, biogenesis, and mechanism of action. Arch. Microbiol. 167, 67-77 NIYONSABA, F., USHIO, H., NAGAOKA, I., OKUMURA, K. and OGAWA, H. (2005): The human ß-defensins (-1, -2, -3, -4) and cathelicidin LL-37 induce IL-18 secretion through p38 and ERK MAPK activation in primary human keratinocytes. J. Immunol. 175, 1776-1784 NIYONSABA, F., USHIO, H., NAKANO, N., NG, W., SAYAMA, K., HASHIMOTO, K., NAGAOKA, I., OKUMURA, K. and OGAWA, H. (2007): Antimicrobial peptides human ß-defensins stimulate epidermal keratinocyte migration, proliferation and production of pro-inflammatory cytokines and chemokines. J. Invest. Dermatol. 127, 594-604 OHYAMA, M., TERUNUMA, A., TOCK, C. L., RADONOVICH, M. F., PISE-MASISON, C. A., HOPPING, S. B., BRADY, J. N., UDEY, M. C. and VOGEL, J. C. (2006): Characterization and isolation of stem cell-enriched human hair follicle bulge cells. J. Clin. Invest. 116, 249-260 OKUNIEFF, P., CORNELISON, T. and MESTER, M. (2005): Mechanism and modification of gastrointestinal soft tissue response to radiation: role of growth factors. Int. J. Radiat. Oncol. Biol. Phys. 62, 273-278

ANNEX
242
OLIVER, R. F. (1966a): Histological studies of whisker regeneration in the hooded rat. J. Embryol. Exp. Morphol. 16, 231-244 OLIVER, R. F. (1966b): Whisker growth after removal of the dermal papilla and lengths of follicle in the hooded rat. J. Embryol. Exp. Morphol. 15, 331-347 PACASOVA, R., MARTINOZZI, S., BOULOUIS, H. J., ULBRECHT, M., VIEVILLE, J. C., SIGAUX, F., WEISS, E. H. and PLA, M. (1999): Cell-surface expression and alloantigenic function of a human nonclassical class I molecule (HLA-E) in transgenic mice. J. Immunol. 162, 5190-5196 PAMER, E. and CRESSWELL, P. (1998): Mechanisms of MHC class I--restricted antigen processing. Annu. Rev. Immunol. 16, 323-358 PANTELEYEV, A. A., JAHODA, C. A. and CHRISTIANO, A. M. (2001): Hair follicle predetermination. J. Cell Sci. 114, 3419-3431 PARKIN, J. M., HEY, E. N. and CLOWES, J. S. (1976): Rapid assessment of gestational age at birth. Arch. Dis. Child. 51, 259-263 PAUS, R. and COTSARELIS, G. (1999): The biology of hair follicles. N. Engl. J. Med. 341, 491-497 PAUS, R. and FOITZIK, K. (2004): In search of the ´hair cycle clock`: a guided tour. Differentiation 72, 489-511 PAUS, R., CHRISTOPH, T. and MUELLER-ROEVER, S. (1999b): Immunology of the hair follicle: a short journey into terra incognita. J. Investig. Dermatol. Symp. Proc. 4, 226-234 PAUS, R., NICKOLOFF, B. J. and ITO, T. (2005): A 'hairy' privilege. Trends Immunol. 26, 32-40

REFERENCES
243
PAUS, R, PEKER, S. and SUNDBERG,J.P. (2007): Biology of hair and nails. In: BOLOGNIA, J.L., JORIZZO, J.L.. and RAPINI, R.P. (eds): Dermatology, 2nd edition. Mosby, London, pp.1007-1032 PAUS, R., STENN, K. S. and LINK, R. E. (1990): Telogen skin contains an inhibitor of hair growth. Br. J. Dermatol. 122, 777-778 PAUS, R., BOTCHKAREV, V. A., BOTCHKAREVA, N. V., MECKLENBURG, L., LUGER, T. and SLOMINSKI, A. (1999a): The skin POMC system (SPS). Leads and lessons from the hair follicle. Ann. N. Y. Acad. Sci. 885, 350-363 PAUS, R., EICHMUELLER, S., HOFMANN, U., CZARNETZKI, B. M. and ROBINSON, P. (1994b): Expression of classical and non-classical MHC class I antigens in murine hair follicles. Br. J. Dermatol. 131, 177-183 PAUS, R., HOFMANN, U., EICHMUELLER, S. and CZARNETZKI, B. (1994a): Distribution and changing density of γ/δ T cells in murine skin during the induced hair cycle. Br. J. Dermatol. 130, 281-289 PAUS, R., ITO, N., TAKIGAWA, M. and ITO, T. (2003): The hair follicle and immune privilege. J. Investig. Dermatol. Symp. Proc. 8, 188-194 PAUS, R., MAURER, M., SLOMINSKI, A. and CZARNETZKI, B. M. (1994c): Mast cell involvement in murine hair growth. Dev. Biol. 163, 230-240 PAUS, R., PETERS, E. M., EICHMUELLER, S. and BOTCHKAREV, V. A. (1997): Neural mechanisms of hair growth control. J. Investig. Dermatol. Symp. Proc. 2, 61-68 PAUS, R., SLOMINSKI, A. and CZARNETZKI, B. M. (1993): Is alopecia areata an autoimmune-response against melanogenesis-related proteins, exposed by abnormal MHC class I expression in the anagen hair bulb? Yale J. Biol. Med. 66, 541-554 PAUS, R., VAN DER VEEN, C., EICHMUELLER, S., KOPP, T., HAGEN, E., MUELLER-ROEVER, S. and HOFFMANN, U. (1998): Generation and cyclic remodeling of the hair follicle immune system in mice. J. Invest. Dermatol. 111, 7-18

ANNEX
244
PETERS, E. M., BOTCHKAREV, V. A., MUELLER-ROEVER, S., MOLL, I., RICE, F. L. and PAUS, R. (2002): Developmental timing of hair follicle and dorsal skin innervation in mice. J. Comp. Neurol. 448, 28-52 PETERS, E. M., BOTCHKAREV, V. A., BOTCHKAREVA, N. V., TOBIN, D. J. and PAUS, R. (2001): Hair-cycle-associated remodeling of the peptidergic innervation of murine skin, and hair growth modulation by neuropeptides. J Invest Dermatol. 116, 236-245 PETERS, E. M., MAURER, M., BOTCHKAREV, V. A., GORDON, D. S. and PAUS, R. (1999): Hair growth-modulation by adrenergic drugs. Exp Dermatol. 8, 274-281 PHILPOTT, M. P., GREEN, M. R. and KEALEY, T. (1990): Human hair growth in vitro. J. Cell Sci. 97 ( Pt 3), 463-471 PHILPOTT, M. P., SANDERS, D. A., BOWEN, J. and KEALEY, T. (1996): Effects of interleukins, colony-stimulating factor and tumour necrosis factor on human hair follicle growth in vitro: a possible role for interleukin-1 and tumour necrosis factor-α in alopecia areata. Br. J. Dermatol. 135, 942-948 PICARDO, M., TOSTI, A., MARCHESE, C., ZOMPETTA, C., TORRISI, M. R., FAGGIONIE, A. and CAMELI, N. (1994): Characterization of cultured nail matrix cells. J. Am. Acad. Dermatol. 30, 434-440 PISANSARAKIT, P. and MOORE, G. P. (1986): Induction of hair follicles in mouse skin by rat vibrissa dermal papillae. J. Embryol. Exp. Morphol. 94, 113-119 POWELL, B. C. and ROGERS, G. E. (1997): The role of keratin proteins and their genes in the growth, structure and properties of hair. Exs. 78, 59-148 POZDNYAKOVA, O. and MAHALINGAM, M. (2008): Involvement of the bulge region in primary scarring alopecia. J. Cutan. Pathol. (in press)

REFERENCES
245
RENEHAN, W. E. and MUNGER, B. L. (1986): Degeneration and regeneration of peripheral nerve in the rat trigeminal system. II. Response to nerve lesions. J. Comp. Neurol. 249, 429-459 REYNOLDS, A. J., LAWRENCE, C., CSERHALMI-FRIEDMAN, P. B., CHRISTIANO, A. M. and JAHODA, C. A. (1999): Trans-gender induction of hair follicles. Nature 402, 33-34 RICE, F. L., MANCE, A. and MUNGER, B. L. (1986): A comparative light microscopic analysis of the sensory innervation of the mystacial pad. I. Innervation of vibrissal follicle-sinus complexes. J. Comp. Neurol. 252, 154-174 RICH, A. R. and LEWIS, M. R. (1932): The nature of allergy in tuberculosis as revealed by tissue culture studies. Bull Johns Hopkins Hosp. 50, 115-131 RICHARDSON K.C., JARETT L. and FINKE, E.H. (1960): Embedding in epoxy resins for ultrathin sectioning in electron microscopy. Stain Technol. 35, 313-323 ROBINSON, M., REYNOLDS, A. J. and JAHODA, C. A. (1997): Hair cycle stage of the mouse vibrissa follicle determines subsequent fibre growth and follicle behavior in vitro. J. Invest. Dermatol. 108, 495-500 ROOK, A. H., KEHRL, J. H., WAKEFIELD, L. M., ROBERTS, A. B., SPORN, M. B., BURLINGTON, D. B., LANE, H. C. and FAUCI, A. S. (1986): Effects of transforming growth factor beta on the functions of natural killer cells: depressed cytolytic activity and blunting of interferon responsiveness. J. Immunol. 136, 3916-3920 ROOSTERMAN, R., GOERGE, T., SCHNEIDER, S. W., BUNNET, N. W. and STEINHOFF, M. (2006): Neuronal control of skin function: the skin as a neuroimmunoendocrine organ. Physiol. Rev. 86, 1309-1379 ROSENBLUM, M. D., OLASZ, E. B., YANCEY, K. B., WOODLIFF, J. E., LAZAROVA, Z., GERBER, K. A. and TRUITT, R. L. (2004): Expression of CD200 on epithelial cells of the murine hair follicle: a role in tissue-specific immune tolerance? J. Invest. Dermatol. 123, 880-887 ROSENBLUM, M. D., YANCEY, K. B., OLASZ, E. B. and TRUITT, R. L. (2006): CD200, a ´no danger` signal for hair follicles. J. Dermatol. Sci. 41, 165-174

ANNEX
246
ROUAS-FREISS, N., PAUL, P., DAUSSET, J. and CAROSELLA, E. D. (2000): HLA-G promotes immune tolerance. J. Biol. Regul. Homeost. Agents. 14, 93-98 ROUSSEAU, K., KAUSER, S., PRITCHARD, L. E., WARHURST, A., OLIVER, R. L., SLOMINSKI, A., WEI, E. T., THODY, A. J., TOBIN, D. J. and WHITE, A. (2007): Proopiomelanocortin (POMC), the ACTH/melanocortin precursor, is secreted by human epidermal keratinocytes and melanocytes and stimulates melanogenesis. FASEB J. 21, 1844-1856 RUECKERT, R., HOFMANN, U., VAN DER VEEN, C., BULFONE-PAUS, S. and PAUS, R. (1998): MHC class I expression in murine skin: developmentally controlled and strikingly restricted intraepithelial expression during hair follicle morphogenesis and cycling, and response to cytokine treatment in vivo. J. Invest. Dermatol. 111, 25-30 RUNNE, U. (1980): Nagelveränderungen durch Gewohnheitsmanipulationen, Artefakte und Traumen. Hautarzt 31, 345 RUNNE, U. and ORFANOS, C. E. (1981): The human nail: structure, growth and pathological changes. Curr. Probl. Dermatol. 9, 102-149 SAYED, B. A., CHRISTY, A., QUIRION, M. R. and BROWN, M. A. (2008): The master switch: The role of mast cells in autoimmunity and tolerance. Annu. Rev. Immunol. 26, 705-739 SELLERI, S., ARNABOLDI, F., PALAZZO, M., GARIBOLDI, S., ZANOBBIO, L., OPIZZI, E., SHIRAI, Y. F., BALSARI, A. and RUMIO, C. (2007): Toll-like receptor agonists regulate ß-defensin 2 release in hair follicle. Br. J. Dermatol. 156, 1172-1177 SHIMIZU, T. (2005): Role of macrophage migration inhibitory factor (MIF) in the skin. J. Dermatol. Sci. 37, 65-73 SHIMIZU, T., OHKAWARA, A., NISHIHIRA, J. and SAKAMOTO, W. (1996): Identification of macrophage migration inhibitory factor (MIF) in human skin and its immmunohistochemical localization. FEBS Lett. 381, 199-202

REFERENCES
247
SHULL, M. M., Ormsby, I., KIER, A. B., PAWLOWSKI, S., DIEBOLD, R. J., YIN, M., ALLEN, R., SIDMAN, C., PROTZEL, G., CALCIN, D. (1992): Targeted disruption of the mouse transforming growth factor-ß-1 gene results in multifocal inflammatory disease. Nature 359, 693-699 SIEBER-BLUM, M., GRIM, M., HU, Y. F. and SZEDER, V. (2004): Pluripotent neural crest stem cells in the adult hair follicle. Dev. Dyn. 231, 258-269 SIGLIENTI, I., CHAN, A., KLEINSCHNITZ, C., JANDER, S., TOYKA, K. V., GOLD, R. and STOLL, G. (2007): Downregulation of transforming growth factor-ß2 facilitates inflammation in the central nervous system by reciprocal astrocyte/microglia interactions. J. Neuropathol. Exp. Neurol. 66, 47-56 SIMPSON, E. (2006): A historical perspective on immunological privilege. Immunol. Rev. 213, 12-22 SLOMINSKI, A., BOTCHKAREVA, N. V., BOTCHKAREVA, V. A., CHAKRABORTY, A., LUGER, T., UENALA, M. and PAUS, R. (1998): Hair cycle-dependent production of ACTH in mouse skin. Biochim. Biophys. Acta. 1448, 147-152 SLOMINSKI, A. and PAUS, R. (1993): Melanogenesis is coupled to murine anagen: toward new concepts for the role of melanocytes and the regulation of melanogenesis in hair growth. J. Invest. Dermatol. 101, 90S-97S SLOMINSKI, A., PAUS, R. and MAZURKIEWICZ, J. (1992): Proopiomelanocortin expression in the skin during induced hair gwoth in mice. Experientia 48, 50-54 SLOMINSKI, A., PAUS, R., PLONKA, P., CHAKRABORTY, A., MAURER, M., PRUSKI, D. and LUKIEWICZ, S. (1994): Melanogenesis during the anagen-catagen-telogen transformation of the murine hair cycle. J. Invest. Dermatol. 102, 862-869 SLOMINSKI, A., WORTSMAN, J., LUGER, T., PAUS, R. and SOLOMON, S. (2000): Corticotropin releasing hormone and proopiomelanocortin involvement in the cutaneous response to stress. Physiol. Rev. 80, 979-1020

ANNEX
248
SLOMINSKI, A., WORTSMAN, J., MAZURKIEWICZ, J. E., MATSUOKA, L., DIETRICH, J., LAWRENCE, K., GORBANI, A. and PAUS, R. (1993): Detection of proopiomelanocortin-derived antigens in normal and pathologic human skin. J. Lab. Clin. Med. 122, 658-666 SMITH, M. E., HOLGATE, C. S., WILLIAMSON, J. M., GRIGOR, I., QUIRKE, P. and BIRD, C. C. (1987): Major histocompatibility complex class II antigen expression in B and T cell non-Hodgkin's lymphoma. J. Clin. Pathol. 40, 34-41 SOHN, J. H., KAPLAN, H. J., SUK, H. J., BORA, P. S. and BORA, N. S. (2000a): Chronic low level complement activation within the eye is controlled by intraocular complement regulatory proteins. Invest. Ophthalmol. Vis. Sci. 41, 3492-3502 SOHN, J. H., KAPLAN, H. J., SUK, H. J., BORA, P. S. and BORA, N. S. (2000b): Complement regulatory activity of normal human intraocular fluid is mediated by MCP, DAF, and CD59. Invest. Ophthalmol. Vis. Sci. 41, 4195-4202 SPERLING, L. and LUPTON, G. (1995): Histopathology of non-scarring alopecia. J. Cutan. Pathol. 22, 97-114 STEINHOFF, M., MEINHARDT, A., STEINHOFF, A., GENSA, D., BUCALA, R. and BACHER, M. (1999): Evidence for a role of macrophage migration inhibitory factor in psoriatic skin disease. Br. J. Dermatol. 141, 1061-1066 STENN, K.S. and PAUS, R. (2001): Control of hair follicle cycling. Physiol. Rev. 81, 449-494 STENN, K. S., COMBATES, N. J., EILERTSEN, K. J., GORDON, J. S., PARDINAS, J. R., PARIMOO, S. and PROUTY, S. M. (1996): Hair follicle growth controls. Dermatol. Clin. 14, 543-558 STREILEIN, J. W. (1993): Immune privilege as the result of local tissue barriers and immunosuppressive microenvironments. Curr. Opin. Immunol. 5, 428-432

REFERENCES
249
STREILEIN, J. W. (2003): Ocular immune privilege: therapeutic opportunities from an experiment of nature. Nat. Rev. Immunol. 3, 879-889 STREILEIN, J. W., MASLI, S. and TAKEUCHI, M. (2002): The eye's view of antigen presentation. Hum. Immunol. 63, 435-443 SUTO, H., NAKAE, S. and KAKURAI (2006): Mast cell-associated TNF promotes dendritic cell migration. J. Immunol. 176, 4102 SWANSON, K. A., ZHENG, Y., HEIDLER, K. M., MIZOBUCHI, T. and WILKES, D. S. (2004): CDllc+ cells modulate pulmonary immune responses by production of indoleamine 2,3-dioxygenase. Am. J. Respir. Cell Mol. Biol. 30, 311-318 TACHIBANA, T. and NAWA, T. (2005): Immunohistochemical reactions of receptors to met-enkephalin, VIP, substance P, and CGRP located on Merkel cells in the rat sinus hair follicle. Arch. Histol. Cytol. 68, 383-391 TACHIBANA, T., ENDOH, M., FUJIWARA, N. and NAWA, T. (2005): Receptors and transporter for serotonin in Merkel cell-nerve endings in the rat sinus hair follicle. An immunohistochemical study. Arch. Histol. Cytol. 68, 19-28 TAFURI, A., ALFERINK, J., MOLLER, P., HAMMERLING, G. J. and ARNOLD, B. (1995): T cell awareness of paternal alloantigens during pregnancy. Science 270, 630-633 TAHERZADEH, S., SHARMA, S., CHHAJLANI, V., GANTZ, I., RAJORA, N., DEMITRI, M. T., KELLY, L., ZHAO, H., ICHIYAMA, T., CATANIA, A. and LIPTON, J. M. (1999): α-MSH and its receptors in regulation of tumor necrosis factor-α production by human monocyte/macrophages. Am. J. Physiol. 276, R1289-1294 TAKEUCHI, M., ALARD, P. and STREILEIN, J. W. (1998): TGF-β promotes immune deviation by altering accessory signals of antigen-presenting cells. J. Immunol. 160, 1589-1597

ANNEX
250
TAYLOR, A. W. (1999): Ocular immunosuppressive microenvironment. Chem. Immunol. 73, 72-89 TAYLOR, A. W. and KITAICHI, N. (2007): The diminishment of experimental autoimmune encephalomyelitis (EAE) by neuropeptide α-melanocyte stimulating hormone (α-MSH) therapy. Brain Behav. Immun. 22, 639-646 TAYLOR, A. W., STREILEIN, J. W. and COUSINS, S. W. (1992): Identification of α-melanocyte-stimulating hormone as a potential immunosuppressive factor in aqueaous humor. Curr. Eye Res. 11, 1199-1206 TAYLOR, A. W., STREILEIN, J. W. and COUSINS, S. W. (1994): α-melanocyte-stimulating hormone supresses antigen-stimulated T cell production of gamma interferon. Neuroimmunomodulation 1, 188-194 TAYLOR, A. W., YEE, D. G., NISHIDA, T. and NAMBA, K. (2000): Neuropeptide regulation of immunity. The immunesuppressive activity of α-melanocyte-stimulating hormone. Ann. N. Y. Acad. Sci. 917, 239-247 THEOHARIDES, T. C. and KALOGEROMITROS, D. (2006): The critical role of mast cells in allergy and infalmmation. Ann. N. Y. Acad. Sci. 1088, 78-99 THORNDYKE, E. (1966): Microscopic studies of the marmoset claw an nail. Am. J. Phys. Anthropol. NS. 28, TIEDE, S., KLOEPPER, J. E., BODO, E., TIWARI, S., KRUSE, C. and PAUS, R. (2007a): Hair follicle stem cells: Walking the maze. Eur. J. Cell Biol. 86, 355-376 TIEDE, S., KLOEPPER, J. E., WHITING, D. A. and PAUS, R. (2007b): The ´follicular trochanter`: an epithelial compartment of the human hair follicle bulge region in need of further characterization. Br. J. Dermatol. 157, 1013-1016 TOBIN, D. J., GUNIN, A., MAGERL, M., HANDIJSKI, B. and PAUS, R. (2003): Plasticity and cytokinetic dynamics of the hair follicle mesenchyme: implications for hair growth control. J. Invest. Dermatol. 120, 895-904

REFERENCES
251
TOBIN, D. J. and PAUS, R. (2001): Graying: gerontobiology of the hair follicle pigmentary unit. Exp. Gerontol. 36, 29-54 TODES-TAYLOR, N., TURNER, R., WOOD, G., STRATTE, P. and MORHENN, V. (1984): T cell subpoulations in alopecia areata. J. Am. Acad Dermatol. 11, 216-223 TOKURA Y, HOFMANN U, MUELLER-ROEVER S, PAUS, R., WAKITA H, YAGI H, SEO N, FURUKAWA F and M, T. (1997): Spontaneous hair follicle cycling may influence the development of murine contact photosensitivity by modulating kerantinocate cytokine production. Cell. Immunol. 178, 172-179 TORO, J. R., FINLAY, D., DOU, X., ZHENG, S. C., LEBOIT, P. E. and CONNOLLY, M. K. (2000): Detection of type 1 cytokines in discoid lupus erythematosus. Arch. Dermatol. 136, 1497-1501 TOSTI, A. and PIRACCHINI, B. (2008): Biology of nails. In: WOLFF, K., KATZ, S.I. and GOLDSMITH, L.A. (eds.): Fitzpatrick's Dermatology in General Medicine. McGraw-Hill, New York. TOSTI, A., BARAN, R. and DAWBER, R.P.R. (2001): The nail in systemic diseases and drug-induced changes. In: Baran, R., Dawber, R.P.R., de Berker, D.A.R., Haneke, R., Tosti, A. (eds.): Diseases of the nails and their management. Blackwell Science, Oxford, U.K. TOSTI, A., CAMELI, N., PIRACCINI, B. M., FANTI, P. A. and ORTONNE, J. P. (1994): Characterization of nail matrix melanocytes with anti-PEP1, anti-PEP8, TMH-1, and HMB-45 antibodies. J. Am. Acad. Dermatol. 31, 193-196 TRIPATHI, P., NAIK, S. and AGRAWAL, S. (2006): HLA-E and immunobiology of pregnancy. Tissue Antigens 67, 207-213 TUMBAR, T., GUASCH, G., GRECO, V., BLANPAIN, C., LOWRY, W. E., RENDL, M. and FUCHS, E. (2004): Defining the epithelial stem cell niche in skin. Science 303, 359-363

ANNEX
252
UHR, G. (1984): Vergleichende Untersuchungen an Haut und Haarkleid von Mus musculus domesticus Rutty, 1772 und Rattus norvegicus Berkenhout, 1769 Thesis (doctoral), University Hannover ULBRECHT, M., HONKA, T., PERSON, S., JOHNSON, J. P. and WEISS, E. H. (1992): The HLA-E gene encodes two differentially regulated transcripts and a cell surface protein. J. Immunol. 149, 2945-2953 VAN BEEK, N., BODO, E., KROMMINGA, A., GASPAR, E., MEYER, K., ZMIJEWSKI, M. A., SLOMINSKI, A., WENZEL, B. E. and PAUS, R. (2008): Thyroid hormones directly alter human hair follicle functions: Anagen prolongation, and stimulation of both hair matrix keratinocyte proliferation and hair pigmentation. J. Clin. Endocrinol. Metab. (in press) VAN EXAN, R. J. and HARDY, M. H. (1980): A spatial relationship between innervation and the early differentiation of vibrissa follicles in the embryonic mouse. J. Anat. 131, 643-656 VIGANO, P., CINTORINO, M., SCHATZ, F., LOCKWOOD, C. J. and ARCURI, F. (2007): The role of macrophage migration inhibitory factor in maintaining the immune privilege at the fetal-maternal interface. Semin. Immunopathol. 29, 135-150 VINCENT, S. B. (1913): The tactile hair of the white rat. J. Comp. Neurol. 23, 1-38 VIVIER, E., TOMASELLP, E. and PAUL, P. (2002): Lymphocyte activation via NKG2D: towards a new paradigm in immune recognition? Curr. Opin. Immunol. 14, 306-311 VOLLRATH, D., JARAMILLO-BABB, V. L., CLOUGH, M. V., MCINTOSH, I., SCOTT, K. M., LICHTER, P. R. and RICHARDS, J. E. (1998): Loss-of-function mutations in the LIM-homeodomain gene, LMX1B, in nail-patella syndrome. Hum. Mol. Genet. 7, 1091-1098

REFERENCES
253
VON BUBNOFF, D., BAUSINGER, H., MATZ, H., KOCH, S., HACKER, G., TAKIKAWA, O., BIEBER, T., HANAU, D. and DE LA SALLE, H. (2004): Human epidermal langerhans cells express the immunoregulatory enzyme indoleamine 2,3-dioxygenase. J. Invest. Dermatol. 123, 298-304 WAHL, S. M., WEN, J. and MOUTSOPOULOS, N. (2006): TGF-β: a mobile purveyor of immune privilege. Immunol. Rev. 213, 213-227 WAITE, P. M. and CRAGG, B. G. (1982): The peripheral and central changes resulting from cutting or crushing the afferent nerve supply to the whiskers. Proc. R. Soc. Lond. B. Biol. Sci. 214, 191-211 WEBER, A., KNORP, J. and MAURER, M. (2003): Pattern analysis of human cutaneous mast cell populations by total body surface mapping. Br. J. Dermatol. 148, WELKER, E. and VAN DER LOOS, H. (1986): Quantitative correlation between barrel-field size and the sensory innervation of the whiskerpad: a comparative study in six strains of mice bred for different patterns of mystacial vibrissae. J. Neurosci. 6, 3355-3373 WELKER, P., FOITZIK, K., BULFONE-PAUS, S., HENZ, B. M. and PAUS, R. (1997): Hair cycle-dependent changes in the gene expression and protein content of transforming factor ß 1 and ß2 in murine skin. Arch. Dermatol. Res. 289, 554-557 WELLER, K., FOITZIK, K., PAUS, R., SYSKA, W. and MAURER, M. (2006): Mast cells are required for normal healing of skin wounds in mice. Faseb J. 20, 2366-2368 WENZEL, J. and TUTING, T. (2007): Identification of type I interferon-associated inflammation in the pathogenesis of cutaneous lupus erythematosus opens up options for novel therapeutic approaches. Exp. Dermatol. 16, 454-463 WENZEL, J., ZAHN, S., MIKUS, S., WIECHERT, A., BIEBER, T. and TUTING, T. (2007): The expression pattern of interferon-inducible proteins reflects the characteristic histological distribution of infiltrating immune cells in different cutaneous lupus erythematosus subsets. Br. J. Dermatol. 157, 752-757

ANNEX
254
WESTGATE GE, CRAGGS RI and WT, G. (1991): Immune privilege and hair growth. J. Invest. Dermatol 97, 417-420 WHITING, D. A. (2001): Cicatricial alopecia: clinico-pathological findings and treatment. Clin. Dermatol. 19, 211-225 WHITING, D. A. (2003): Histopathologic features of alopecia areata: a new look. Arch. Dermatol. 139, 1555-1559 WHITING, D. A. (2004): The Structure of the Human Hair Follicle. Canfield Publishing, Fairfield New Jersey WIENDL, H., MITSDOERFFER, M., HOFMEISTER, V., WISCHHUSEN, J., WEISS, E. H., DICHGANS, J., LOCHMULLER, H., HOHLFELD, R., MELMS, A. and WELLER, M. (2003): The non-classical MHC molecule HLA-G protects human muscle cells from immune-mediated lysis: implications for myoblast transplantation and gene therapy. Brain 126, 176-185 WINKELMANN, R. K. (1959): The innervation of a hair follicle. Ann. N. Y. Acad. Sci. 83, 400-407 WOOD, J. M., GIBBONS, N. C. and SCHALLREUTER, K. U. (2006): Melanocortins in human melanocytes. Cell Mol. Biol. (Noisy-le-grand). 52, 75-78 WOOLSEY, T. A. and VAN DER LOOS, H. (1970): The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 17, 205-242 WRENN, J. T. and WESSELLS, N. K. (1984): The early development of mystacial vibrissae in the mouse. J. Embryol. Exp. Morphol. 83, 137-156

REFERENCES
255
WRIGHT, G. J., CHERWINSKI, H., FOSTER-CUEVAS, M., BROOKE, G., PUKLAVEC, M. J., BIGLER, M., SONG, Y., JENMALM, M., GORMAN, D., MCCLANAHAN, T., LIU, M. R., BROWN, M. H., SEDGWICK, J. D., PHILLIPS, J. H. and BARCLAY, A. N. (2003): Characterization of the CD200 receptor family in mice and humans and their interactions with CD200. J. Immunol. 171, 3034-3046 WRIGHT, G. J., JONES, M., PUKLAVEC, M. J., BROWN, M. H. and BARCLAY, A. N. (2001): The unusual distribution of the neuronal/lymphoid cell surface CD200 (OX2) glycoprotein is conserved in humans. Immunology 102, 173-179 WRONG, K. Y., RAJORA, N., BOCCOLI, G., CATANIA, A. and LIPTON, J. M. (1997): A potential mechanism of local anti-inflammatory action of α-melanocyte-stimulating hormone within the brain: modulation of tumor necrosis factor-α production by human astrocytic cells. Neuroimmunomodulation 4, 37-41 WU, H., SONG, Y., BAKKER, A. B., BAUER, S., SPIES, T., LANIER, L. L. and PHILIPS, J. H. (1999): An activating immunoreceptor comlex formed by NKG2D and DAP10. Science 285, 730-732 XU, C., MAO, D., HOLERS, V., PALANCA, B., CHEN, A. and MOLINA, H. (2000): A critical role for murine complement regular Crry in fetomaternal tolerance. Science 287, 498-501 YAMAKADO, M. and YOHRO, T. (1979): Subdivision of mouse vibrissae on an embryological basis, with descriptions of variations in the number and arrangement of sinus hairs and cortical barrels in BALB/c (nu/+; nude, nu/nu) and hairless (hr/hr) strains. Am. J. Anat. 155, 153-173 YANG, D., CHERTOV, O., BYKOVSKAIA, S. N., CHEN, Q., BUFFO, M. J., SHOGAN, J., ANDERSON, M., SCHROEDER, J. M., WANG, J. M., HOWARD, O. M. and OPPENHEIM, J. J. (1999): ß-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science 286, 525-528 YANO, K., BROWN, L. F. and DETMAR, M. (2001): Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Invest. 107, 409-417

ANNEX
256
YIP, L., MCCLUSKEY, J. and SINCLAIR, R. (2006): Immunological aspects of pregnancy. Clin. Dermatol. 24, 84-87 YOUNG, L. C. J. (1997): The mast cell: origin, morphology, distribution, and function. Exp. Toxic Pathol. 49, 409-424 YOUNG, R. D. and OLIVER, R. F. (1976): Morphological changes associated with the growth cycle of vibrissal follicles in the rat. J. Embryol. Exp. Morphol. 36, 597-607 ZAIAS, N. (1963): Embryology of the human nail. Arch. Dermatol. 87, 37-53 ZAIAS, N. and ALVAREZ, J. (1968): The formation of the primate nail plate. An autoradiographic study in squirrel monkey. J. Invest. Dermatol. 51, 120-136 ZIEGLER, H. (1954): [The formation of human nail and hors’s hoof.] Z. Mikrosk. Anat. Forsch. 60: 556-572 ZIETZSCHMANN (1920): Beiträge zum Bau und zur Entwicklung von Hautorganen bei Säugetieren. 7 Die früheste Entwicklung der Sinushaare des Schweines. Anat. Anz.. 52, 332-349

ANNEX
9 Annex
Parts of this thesis were published as follows:
Articles: ITO, T., MEYER, K.C., ITO, N., PAUS, R. (2008): Immune Privilege and the Skin. Curr. Dir. Autoimmun. 10, pp 27–52 MEYER, K.C., KLATTE, J.E., REITHMAYER, K., DINH, H.D., HARRIES, M.J., MEYER, W., SINCLAIR, R., PAUS, R. (2008) Evidence that the bulge region is a site of relative immune privilege in human hair follicles Br. J. Dermatol. Br J Dermatol. 2008 Sep 15. [Epub ahead of print], PMID: 18795933 HARRIES, M.J., MEYER, K.C., PAUS, R. (2009)
Hair loss as a result of cutaneous autoimmunity: Frontiers in the immunopathogenesis of primary cicatricial alopecia Autoimmun. Rev. 2008 Oct 13. [Epub ahead of print] PMID: 18926937 Meyer, K.C., BRZOSKA, T., ABELS, C., PAUS, R. The role of α-MSH related tripeptide K(D)PT in human hair follicle immunology and biology in situ under pro-inflammatory conditions Br. J. Dermatol. 2008 Oct 25. [Epub ahead of print] PMID: 19016700 Abstracts: MEYER, K.C., KLOEPPER, J.E., TIEDE, S., PAUS, R. (2007): The bulge – a second site of epithelial immune privilege within the human anagen hair follicle? Exp. Dermatol., 16, 3, P087, 221-222 KLOEPPER, J.E., MEYER, K. C., TIEDE, S., PAUS, R. (2007): The human hair follicle bulge: Immunohistological examination of potential stem cell-niche elements, and further indications that it displays relative immune privilege J. Invest. Dermatol. 127, S198 MEYER, K. C., KLOEPPER, J.E., DINH, H.V., SINCLAIR, R., PAUS, R. (2008): The immunoprivileged bulge: Regeneration pool and immunological “Achilles’ heel” of the hair follicle? Exp. Dermatol., 17, Number 3, March 2008 P134, Page 264

ANNEX
258
REITHMAYER, K., MEYER, K.C., GLAESER, R., HARDER, H., SCHROEDER, J.M., PAUS, R. (2008): Antimicrobial defense of human hair follicle epithelium: inducible expression of RNase 7 and psoriasin and constitutive expression of hornerin Dermatol., 17, 3, P51, Page 264 VAN BEEK, N., BODÓ, E., KROMMINGA, A., GÁSPÁR, E., MEYER, K.C., ZMIJEWSKI, M.A., SLOMINSKI, A., WENZEL, B.E., PAUS, R. (2008): Human hair follicles are direct targets for thyroid hormones: involvement in anagen prolongation, hair matrix proliferation, hair pigmentation and metabolism J. Invest. Dermatol. 128, S156-S156
Additional publications generated during this thesis project: MEYER, K.C., BRZOSKA, T., ABELS, C., PAUS, R. (2008) The α-MSH related tripeptide K(D)PT stimulates hair follicle pigmentation in situ under pro-inflammatory conditions Br. J. Dermatol. (in press) VAN BEEK, N., BODÓ, E., KROMMINGA, A., GÁSPÁR, E., MEYER, K.C., ZMIJEWSKI, M.A., SLOMINSKI, A., WENZEL, B.E., PAUS, R. (2008) Thyroid hormones directly alter human hair follicle functions: Anagen prolongation, and stimulation of both hair matrix keratinocyte proliferation and hair pigmentation. J. Clin. Endocrinol. Metab. 2008 Nov;93(11):4381-8. Epub 2008 Aug 26. REITHMAYER, K., MEYER, K.C., GLAESER, R., HARDER, H., SCHROEDER, J.M., PAUS, R. Antimicrobial defense of human hair follicle epithelium: inducible expression of RNase 7 and psoriasin and constitutive expression of hornerin Br. J. Dermatol. (submitted) HARRIES, M.J., MEYER, K.C., CHAUDHRY, I., GRIFFITHS, C., PAUS, R.
Does collapse of the hair follicle bulge immune privilege play an important role in the pathogen? Br. J. Dermatol. (submitted)


260
Mein besonderer Dank gilt…
… meinem lieben Doktorvater, Herrn Prof. Dr. Wilfried Meyer (Anatomisches
Institut, Stiftung Tierärztliche Hochschule Hannover), für die Vermittlung meines
Dissertationsthemas an der Universität zu Lübeck, für die hervorragende und
verantwortungsbewusste Betreuung aus der Ferne und bei meinen Aufenthalt in
Hannover, für die zahlreichen hilfreichen Hinweise und Anregungen und vor allem für
die zu jedem Zeitpunkt gewährte fachliche und menschliche Unterstützung und sein
klassisch-motivierenden Zuspruch „Ach, das schaffen wir schon…!“.
… meinem lieben Betreuer und hervorragenden Chef vor Ort, Herrn Prof. Dr. Ralf
Paus (Klinik für Dermatologie, Allergologie und Venerologie, Universität zu Lübeck),
für die Vergabe des Themas, für die ständigen fachlichen Anregungen und
Hilfestellungen, für das Vertrauen in mich und meine Arbeit, das jederzeit offene Ohr
und seinen ständigen Optimismus und Enthusiasmus bei all meinen Vorhaben.
… dem Direktor der Hautklinik der Universität zu Lübeck, Herrn Prof. Dr. D.
Zillikens, für die Möglichkeit, in seiner Klinik als Doktorandin zu arbeiten, für den
fachlichen Austausch im Rahmen des Autoimmunitätsschwerpunktes der
Medizinischen Fakultät der Universität zu Lübeck und für das gute Arbeitsklima
innerhalb seines Hauses. Ebenso gilt mein Dank Herrn PD. Dr. Ralf Ludwig für den
inhaltlichen Austausch und die Bereitstellung meiner Maushautproben.
… unseren kollaborierenden plastischen Chirurgen, Herrn Dr. Dr. W. Funk (Klinik Dr.
Koslowski, München), Herrn W. Moser (Moser-Klinik, Augsburg) und Herrn J. Levy
(Atriumklinik in Holzkirchen, Holzkirchen), ohne deren wertvolle Lieferungen von
menschlicher Skalphaut aus elektiven Eingriffen diese Arbeit niemals hätte
durchgeführt werden können. Dieser Dank gilt natürlich auch den anonymen
Spendern, deren großzügige Bereitschaft, ihre Hautexzidate zu Forschungszwecken
zur Verfügung zu stellen, die Grundvorausetzung zur Durchführung dieser Arbeit
war.

... Herrn Dr. Christoph Abels und Dr. Thomas Brzoska von der Firma Wolff,
Bielefeld Arzneimittel für die Möglichkeit, die erzielten Ergebnisse über K(D)PT in
meine Dissertation zu integrieren .
… meinen Kooperationspartnern Dr. Matthew J. Harries, Prof. Dr. Taisuke Ito,
Prof. Dr. Rodney Sinclair und Hope V. Dinh für die Bereitstellung diverser
Antikörper und sonstige professionelle Hilfestellungen.
… der AG Wollenberg (Leiter: PD Dr. Ralph Pries) und der AG Wenzel (Leiter: Dr.
Björn Wenzel) an der Universität zu Lübeck für die Erlaubnis, diverse Instrumente
mitnutzen zu dürfen und für die immerwährend gute Zusammenarbeit. Besonderer
Dank gilt hier unmittelbar meinem Tischnachbarn Dr. Björn Wenzel und Frau Silvia
Grammersdorf, für jegliche Unterstützung und interessanten Austausch.
… meiner treuesten Mitdoktorandin, Dr. Jennifer Klatte, die mir den Einstieg
besonders erleichterte, ihr Wissen mit mir teilte, mir in jeglicher Hinsicht und zu jeder
Zeit beiseite stand und steht, und am allerbesten wusste, in welcher emotionalen
Lage der Promotionsphase ich mich gerade befand.
... meinem Laborchef Herrn Dr. Stephan Tiede, der mit seiner besonderen Art und
seinem trockenem Humor den Alltag mitbestimmt hat und auf den ich in
unwegsamen Situationen zählen konnte.
… Frau Dr. Enikö Bodó und Frau Katrin Reithmayer für deren Anteilnahme, Rat
und Tat im Labor und auf allen Ebenen der Dissertation.
… der Chef-MTA Frau Astrid Becker, Frau Nadine Dörwald, Frau Gaby Scheel
und Frau Antje Winter-Keil für ihre menschliche und fachliche Unterstützung, und
die sehr gute Organisation des Labors.

262
… allen bisher nicht genannten Mitgliedern der AG Paus für die tolle
Arbeitsatmosphäre, für die geduldige und engagierte Unterstützung und alle
gemeinsamen Aktivitäten: Nina van Beek, Nina Biedermann, Nancy Ernst, Tobias
Fischer, Lütfiye Hastedt, Elisabeth Gáspár, Céline Hardenbicker, Sybille Hasse,
Benedikt Kany, Patrick Kleditzsch, Dorothee Langan, Viktoria Naumann, Burkhard
Poggeler, Christian Plate, Yuval Ramot, Katrin Reithmayer, Anne Spiegelberger, Koji
Sugawara, Sai Kailash Uppalapati, Ronny Wachtel, Friederike Wiersma, Annika
Hanning.
… meinen lieben Freunden und meiner gesamten Familie für das offene Ohr, den
Rückhalt, das Vertrauen und die Zuversicht
... meinem Freund Robert, für die Geduld, die Hilfestellung und den Stolz, den er für
mich und mein Vorhaben empfand.
… meiner lieben Mutter, Ernst und meinem lieben Bruder für ihre guten Gedanken
und die liebevolle Unterstützung.









