
Aus dem Institut für Pharmakologie, Toxikologie und Pharmazie der Tierärztlichen Hochschule Hannover
__________________________________________________________
Untersuchung zur Dexamethasonverteilung im Katzenauge nach lokaler Applikation
INAUGURAL-DISSERTATION
Zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
(Dr. med. vet.)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Julia Bessonova aus Uchta, Russland
Hannover 2006

Wissenschaftliche Betreuung: Univ.-Prof. Dr. M. Kietzmann
1. Gutachter: Univ.-Prof. Dr. M. Kietzmann
2. Gutachter: Prof. Dr. M. H. Boevé
Tag der mündlichen Prüfung: 08.11.2006

Meinen Eltern und meinem Mann


1 EINLEITUNG 11
2 LITERATURÜBERSICHT 12
2.1 Aufbau des Katzenauges 12
2.1.1 Augenhöhle, Orbita 12
2.1.2 Augenlider 13
2.1.3 Augapfel, Bulbus oculi 15
2.1.4 Sclera und Cornea 15
2.1.5 Netzhaut, Retina 19
2.1.5.1 Pars optica retinae 19
2.1.5.2 Pars ceca retinae 22
2.1.6 Tränenapparat, Apparatus lacrimalis 23
2.1.7 Augenkammern und Kammerflüssigkeit 25
2.1.7.1 Kammerwasserbildung 25
2.1.7.2 Kammerwasserabfluss 28
2.1.8 Glaskörper, Corpus vitreum 29
2.1.9 Regenbogenhaut (Iris) 29
2.1.10 Linse, Lens 31
2.2 Corticosteroide 34
2.2.1 Glucocorticoide 34
2.2.2. Synthese und Sekretion 35
2.2.3 Wirkung der Glucocorticoide 36
2.2.3.1 Allgemeine Wirkmechanismen 37
2.2.3.2 Wirkungen auf den Stoffwechsel 39
2.2.3.3 Wirkungen auf den Wasser- und Elektrolythaushalt 40
2.2.3.4 Wirkungen über den Mineralocorticoidrezeptor 40
2.2.3.5 Wirkungen über den Glucocorticoidrezeptor 41
2.2.3.6 Wirkungen auf das cardiovaskuläre System 41
2.2.3.7 Wirkungen am ZNS 42
2.2.3.8 Wirkungen auf das Blut, die Skelettmuskulatur und das
Wachstum 43
2.2.3.9 Antiinflammatorische und immunsuppressive Wirkungen 43

2.2.4 Nebenwirkungen der Glucocorticoide 44
2.2.5 Cortisol und seine synthetischen Derivate 46
2.2.6 Pharmakokinetik von Ophtalmica 49
3 MATERIAL UND METHODE 52
3.1 Geräte 52
3.2 Verbrauchsmaterialien 52
3.3 Medikament 53
3.4 Chemikalien und Reagenzien 54
3.5 Patientengut 54
3.5.1 Klinische Untersuchung 57
3.5.2 Spezielle ophthalmologische Untersuchung 57
3.6 Probengewinnung und Lagerung 58
3.7 Versuchsaufbau 58
3.7.1 Enukleation der Bulbi oculi 59
3.7.2 Entnahme des Kammerwassers 60
3.7.3 Entnahme von 3. Augenlid, Cornea, Iris und Linse 61
3.7.4 Entnahme von Glaskörperanteilen und der Retina (Choroidea) 61
3.8 Probenaufarbeitung 61
3.9 Radioimunassay (RIA) 62
3.9.1 Prinzip 62
3.9.2 Spezifität des Antiserums 64
3.9.3 Puffer und Lösungen 64
3.9.3.1 Gelatine-Phosphat-Puffer (GPP) 65
3.9.3.2 Dextran-Aktivkohle-Suspension 65
3.9.3.3 Dexamethason-Standardreihe 65
3.9.3.4 3H-Dexamethason-Lösung 65
3.10 Untersuchung der Proben 66
3.11 Kalibrationskurve und Berechnung 68
3.12 Statistische Auswertung 68
3.13 Wiederholbarkeit und Nachweisgrenzen 69
4 ERGEBNISSE 72

4.1 Dexamethasonkonzentrationen in den einzelnen Kompartimenten
des Katzenauges 71
5 DISKUSSION 82 5.1 Fragestellung und Patientengut 82
5.2 Lokale Behandlung des Auges 83
5.3 Glucocorticoide am Auge 84
5.3.1 Dexamethason in der Cornea und im 3. Augenlid 86
5.3.2 Dexamethason im Kammerwasser 86
5.3.3 Dexamethason in der Iris 87
5.3.4 Dexamethason in der Linse, dem Glaskörper und der Retina 89
5.4 Therapeutisch wirksame Konzentration 91
6 ZUSAMMENFASSUNG 93 7 SUMMARY 95 8 LITERATURVERZEICHNIS 97 9 ANHANG 116 Verzeichnis der Abbildungen 116
Verzeichnis der Tabellen 119
DANKSAGUNG 122

Abkürzungen A, Aa. Arteria, Arteriae
Abb. Abbildung
ACTH Adrenocorticotropes Hormon
Ag Antigen
Ag* markiertes Antigen (3H-Dexamethason)
Ak Antikörper
cAMP zyklisches Adenosinmonophosphat
cpm counts per minute
CRH Corticotropin-Releasing-Hormon
DNA Desoxyribonukleinsäure
d.h. das heisst
DHEA Dehydroepiandrosteron
DXM Dexamethason
EEG Elektroenzaphalogramm
EKH Europäsche Kurzhaar (Katzenrasse)
ENaC epitheliale Natriumkanal
et al. et alii
Fa. Firma
FeLV Felines Leukämie Virus (Katzenleukose)
FIP Feline Infektiöse Peritonitis" (ansteckende
Bauchfellentzündung)
FIV Felines Immundefizienz-Virus
g Gramm
GC Glucocorticoide
GCR Glucocorticoid-Rezeptor
GPP Gelatine–Phosphat –Puffer
GRE Glucocorticoid-responsiven Elemente
HPA Hypothalamisch-hypophysär-adrenale Achse
HSP Hitzeschockprotein
kg Kilogramm

Konz. Konzentration
Lsg. Lösung
mg Milligramm
ml Milliliter
MR Mineralocorticoidrezeptor
mRNA messenger Ribonukleinsäure
ng Nanogramm
N. Nervus
NPE nicht pigmentierte Epithelium
NG Nachweisgrenze
NNR Nebennierenrinde
NSB Non-specific binding (unspez. Bindung)
PE pigmentierte Epithelium
pg Picogramm
Procc. Processus (Plural)
RIA Radioimunassay
SGK-Kinase Serum-and glucocorticoid-induced kinase
TC Total count (Gesamtradioaktivität)
u. und
U Umdrehungen
v.a. vor allem
Vk Verteilungskoeffizient
z.B. zum Beispiel
ZNS Zentralnervensystem
z.T. zum Teil
µg Mikrogramm
µl Mikroliter

Einleitung
11
1 Einleitung Entzündete und tränende Augen sind häufige Beschwerden, mit welchen vor allem
jüngere Katzen zu kämpfen haben. Die Konjunktivitis (Bindehautentzündung) ist bei
der Katze niemals eine Bagatellerkrankung, und häufig sind gleichzeitig andere
Symptome wie Schnupfen vorhanden. Es lohnt sich in solchen Fällen die Ursache
der Augenentzündung abzuklären und eine spezifische Therapie einzuleiten, damit
keine Langzeitschäden entstehen können. Ursächlich kommen für eine Entzündung
am Auge z.B. Fremdkörper, einwachsende Haare oder Allergien in Frage. In dieser
Übersicht wird aber auf die weitaus häufigeren infektiösen Ursachen und die
diagnostischen Möglichkeiten eingegangen.
Glucocorticoide stellen eine in der Behandlung des Auges häufig zum Einsatz
gelangende Stoffgruppe dar. Bei entzündlichen Erkrankungen kann ihr Einsatz
aufgrund ihrer starken antiinflammatorischen und imunsuppressiven Wirkung von
großem Nutzen sein, er sollte jedoch nicht ohne gründliche Abwägung der Risiken
geschehen.
Das Ziel der Arbeit ist die Untersuchung der Verteilung von Dexamethason in den
Kompartimenten des Auges nach Anwendung einer handelsüblichen Augensalbe
(Isopto®-Dex) und einer Augentropfsuspension (Isopto-Dex ®). Die diesbezüglichen
Kenntnisse sind noch unzureichend. Durch die Untersuchung soll die Anwendung bei
verschiedenen Erkrankungen des Auges abgesichert werden.

Literaturübersicht
12
2 Literaturübersicht 2.1 Aufbau des Katzenauges 2.1.1 Augenhöhle, Orbita Die knöcherne Wand der Orbita, die den Augapfel medial, dorsal und medioventral
umschließt, wird von den Ossa lacrimale, zygomaticum, frontale, praesphenoidale,
palatinum und maxillare gebildet. Temporal ist die Orbita nur bindegewebig
umschlossen. Caudal steht sie mit der Schläfengrube und der Fossa pterygopalatina
in direkter Verbindung. Der Augenhöhleneingang ist nicht völlig knöchern, sondern
wird zwischen Stirnbein und Jochbogen durch eine straffe Bandverbindung, das
Ligamentum orbitale, begrenzt. Dieses Band kann aber auch verknöchern (ULMER
et al. 1971). Abgesehen von den verschiedenen Durchtritten für Nerven und Gefäße
stellt die Orbita einen umschlossenen Raum dar, in dem sich der Bulbus oculi, N.
opticus und andere Nerven, die extraokulären Muskeln, Blutgefäße, Gl. lacrimalis, Gl.
zygomatica, Fasciae orbitales und das Corpus adiposum orbitae befinden. Bei einer
Größenzunahme einer dieser Strukturen kann es zu einer Verlagerung des Bulbus
oculi kommen, wobei sich der Augapfel aufgrund der anatomischen Gegebenheiten
entweder nach dorsolateral, ventral oder rostral verlagert.
Bei Hunden und Katzen weichen die Augenachsen lediglich um 10º – 20º von einer
senkrecht geradeaus gehenden Nullachse nach rostrolateral ab (PRINCE et al. 1960,
BÖHME 1992).

Literaturübersicht
13
2.1.2 Augenlider Neben dem oberen und dem unteren Augenlid, Palpebra superior und Palpebra
inferior, kommt bei den Säugetieren noch 3.Augenlid, Palpebra tetria, vor. Die
Augenlider sind bewegliche Hautfalten, die zum Schutz des Auges dienen.
Das obere und das untere Augenlid schmiegen sich der freien Bulbusvorderfläche
dicht an und begrenzen mit ihrem Rand die Lidspalte, Rima palpebrarum, die sich
dank der Beweglichkeit, namentlich des oberen Augenlides, mehr oder weniger weit
öffnen oder reflektorisch schließen lässt. Die Wischbewegungen des oberen
Augenlides reinigen die Bulbusvorderfläche von Fremdkörpern. Der Lidschlußreflex
stellt einen Schutzmechanismus dar. Die Bewegungen der Augenlider tragen auch
wesentlich zum mimischen Ausdrucksvermögen der Tiere bei (BÖHME 1992).
Vom 3. Augenlid (Nickhaut), erkennt man bei geöffneter Lidspalte nur einen
schmalen, halbmondförmigen Saum, der am medialen Augenwinkel in die Rima
palpebrarum vorragt. Es wird von einer nahezu senkrecht stehenden Bindehautfalte,
Plica semilunaris conjunctivae, gebildet, die von einem Knorpel, dem Blinzknorpel,
Cartilago palpebrae tertia, gestützt wird und der Bulbusvorderfläche dicht aufliegt.
Bei Druck auf den Augapfel oder, wenn dieser durch Kontraktion des M. retractor
bulbi in die Orbita zurückgezogen wird, fällt das dritte Augenlid über die Cornea
hinweg passiv vor (BERG 1995).
Bei der Katze geht ein glatter Muskel aus der Fascienscheide des M. rectus bulbi
ventralis hervor. Eine Lamelle dieses Muskels zieht ins untere Augenlid, eine zweite
in den unteren Teil der Membrana nictitans. In ähnlicher Weise verhält sich ein
glatter Muskel, der mit dem M. rectus bulbi medialis in Verbindung steht und zum
oberen Augenlid bzw. in den dorsomedialen Teil der Membrana nictitans zieht.
Keiner der Muskeln dringt weit in das dritte Augenlid ein. Die Muskeln sind
sympathisch innerviert und bewirken eine Retraktion der Nickhaut (KÖNIG 1992).
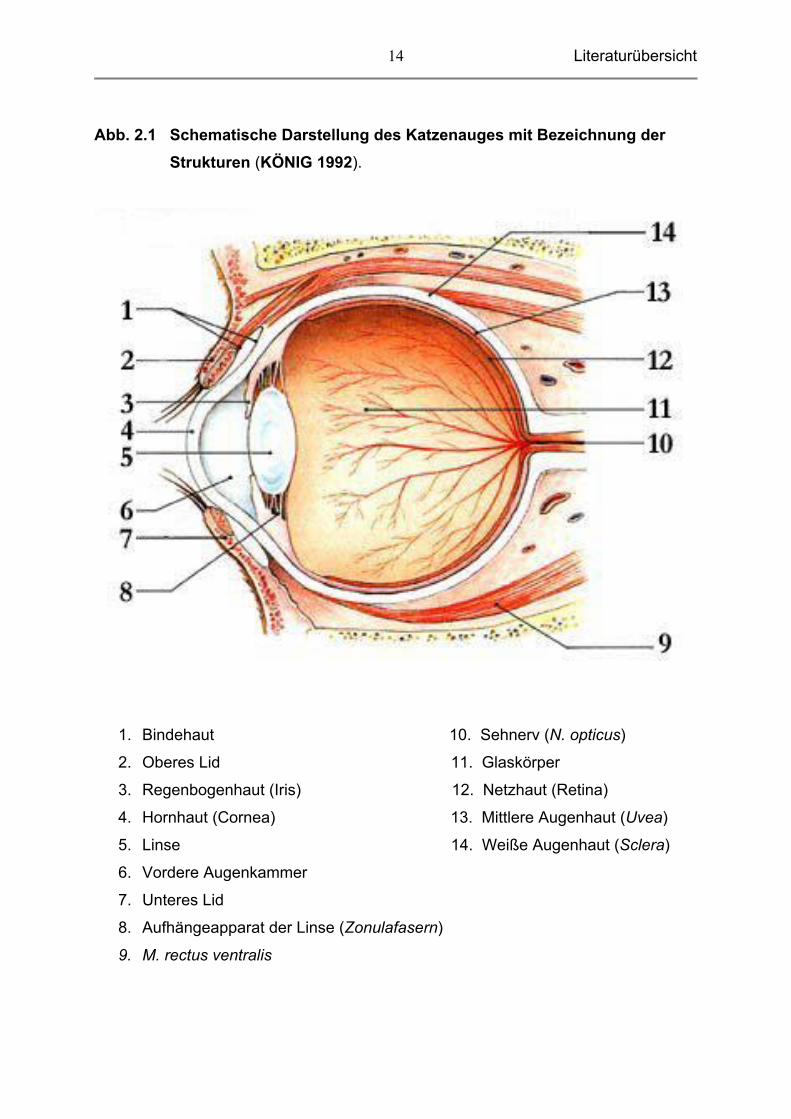
Literaturübersicht
14
Abb. 2.1 Schematische Darstellung des Katzenauges mit Bezeichnung der Strukturen (KÖNIG 1992).
1. Bindehaut 10. Sehnerv (N. opticus)
2. Oberes Lid 11. Glaskörper
3. Regenbogenhaut (Iris) 12. Netzhaut (Retina)
4. Hornhaut (Cornea) 13. Mittlere Augenhaut (Uvea)
5. Linse 14. Weiße Augenhaut (Sclera)
6. Vordere Augenkammer
7. Unteres Lid
8. Aufhängeapparat der Linse (Zonulafasern)
9. M. rectus ventralis

Literaturübersicht
15
2.1.3 Augapfel, Bulbus oculi Der Augapfel stellt eine Hohlkugel dar, deren Wand aus drei konzentrischen Häuten
aufgebaut ist:
1. Die äußere Augenhaut, Tunica fibrosa bulbi (Sklera und Cornea)
2. Die mittlere Augenhaut, Tunica vasculosa bulbi (Choroidea, Ziliarkörper und
Iris)
3. Die äußerst zarte innere Augenhaut, Tunica interna bulbi (Retina)
Den Inhalt des Bulbus oculi bilden der Glaskörper, die Linse mit ihrem
Aufhängeapparat und das Kammerwasser in der vorderen und hinteren
Augenkammer (BÖHME 1992).
Im Verhältnis zur Körpergröße besitzt die Katze einen sehr großen Bulbus, dann
folgen der Hund, das Schaf, das Pferd, das Rind und das Schwein. Er wiegt ca. 4,7 g
und hat ein Volumen von 4,5 ml (FATH EL BAB et al. 1981).
Das Verhältnis von mittlerem Längsdurchmesser (= Länge der äußeren Augenachse,
Verbindungslinie zwischen vorderem und hinterem Augenpol) zum mittleren
Querdurchmesser (in Höhe des Bulbusäquators) zum mittleren Vertikaldurchmesser
(Verbindungslinie zwischen proximalem und distalem Augenpol) beträgt bei der
Katze 21,3: 20,1: 20,2 mm (KÖNIG 1992).
2.1.4 Sclera und Cornea
Die äußere Augenhaut (Tunica fibrosa bulbi) stellt eine formgebende, fibröse Hülle
dar. Sie teilt sich in dem Verhältnis 4:1 in die proximale, undurchsichtige Sclera,
sowie die distale durchsichtige Cornea auf.
Die weiße Sclera ist eine dicke, vorwiegend aus kollagenen Fasern aufgebaute
dehnungsfeste Bindegewebskapsel, die unterstützt durch den Augeninnendruck die

Literaturübersicht
16
Form des Bulbus aufrechterhält (LIEBICH 2004). Ihr liegt außen die gefäßreiche
Lamina episcleralis auf.
Die Sclera besteht hauptsächlich aus drei Schichten: der Episclera, dem Stroma und
der Lamina fusca (KOMAI und USHIKI 1991).
Im Übergangsbereich zur Cornea wird die Sclera außen von der Tunica conjunctiva
bulbi, einem Abschnitt der Bindehaut (Konjunktiva), überzogen. Im Grenzbereich
zwischen Cornea und Sclera, dem sogenannten Limbus, überdeckt die Sclera die
Hornhaut dachziegelartig (Corneoskleralfalz). Die Sclera ist an dieser Stelle durch
einen innen anliegenden Bindegewebsring (Anulus sclerae) zum Scleralwulst
verdickt, wo auch der Musculus ciliaris verankert ist (HUBEL 1989)
Eine Besonderheit bei Hund und Katze stellt das Vorhandensein eines intraskleralen
Plexus dar. Dieser repräsentiert ein venöses Netzwerk im äußeren Stromabereich,
welches die Drainage von Kammerwasser im iridokornealen Winkel unterstützt.
Die lichtdurchlässige Cornea umgibt den distalen Augenpol. Ihre Dicke ist am
zentralen Vertex corneae etwas geringer (0,4 – 0,5 mm) als am peripheren Limbus
corneae (0,5 – 0,65 mm) (GILGER et al. 1993). Die Cornea hat die Form eines
Uhrschälchens und ist mit ihrem scharfen Rand in den Scleralfalz eingelassen. Die
gesunde Cornea ist klar, hat keine Gefäße und bricht das einfallende Licht mit einer
Brechkraft von 40 – 42 Dioptrien.
Die Vorderfläche der Cornea ist von einem mehrschichtigen nichtverhornenden
Plattenepithel überzogen. Bei Hund und Katze ist diese Epithelschicht 25 bis 40 µm
dick.

Literaturübersicht
17
Abb. 2.2 Aufbau der Cornea (OHRLOFF 1987)
Die äußere lipophile Schicht der Hornhaut bildet das Epithel (Abb. 2), welchen aus
einem unverhornten, geschichteten Plattenepithel mit fünf bis sechs Zellschichten
besteht. Zwischenräume sind sehr eng, so dass für Substanzen mit einer relativen
Molekülmasse > 1000 kaum eine Chance besteht, in das Epithel einzudringen.
Bevorzugt lipophile Substanzen mit einem hohen VK können durch das Epithel
diffundieren (LEIBOWITZ et al. 1972, LEIBOWITZ und KUPFERMANN 1974).
Die subepitheliale, sehr starke Basalmembran, Bowman-Membran, ist homogen und
azellulär. Sie trennt das Epithel vom Stroma. Die Substantia propria (Stroma) macht
neun Zehntel der Dicke der Hornhaut aus und besteht überwiegend aus Fibroblasten
und kollagenen Fasern, die in Form sich überkreuzender Lamellen angeordnet sind.
Die regelmäßige Anordnung der kollagenen Fasern, die in eine
mucopolysaccharidreiche Kittsubstanz eingelagert sind, sowie das Fehlen von
Blutgefäßen bedingen die Durchsichtigkeit des Gewebes (ALVERDES 1974). Ein
Wassergehalt von 70% bewirkt den hydrophilen Charakter des Stromas als
elastische und mikroporöse Schicht. Eine rasche Diffusion durch diese Schicht setzt
eine nicht zu geringe Wasserlöslichkeit des Wirkstoffs voraus. Daher penetrieren
Wirkstoffe dann besonders gut durch die Cornea, wenn sie neben lipophilen
(Epitheltransport) in gewissem Umfang auch hydrophile Eigenschaften besitzen
(KISHIDA 1973).

Literaturübersicht
18
Die Descemet-Membran stellt ebenfalls eine kräftige Basalmembran dar, die sich
endothelseitig zum Stroma befindet. Sie weist eine außerordentlich hohe Elastizität
auf und bleibt selbst dann noch intakt, wenn die darüberliegenden Schichten zerstört
sind. Das Endothel schließt die Cornea nach innen ab und besteht lediglich aus einer
Schicht relativ weit auseinanderstehender, abgeflachter, epithelähnlicher Zellen,
deren Hauptaufgabe darin besteht, das Stroma zu dehydrieren und damit die Dicke
und Transparenz der Cornea sicherzustellen. Außerdem spielt das Endothel eine
wichtige Rolle bei aktiven Transportprozessen (HUBEL 1989) und ist als gut
durchlässige Membran wesentlich für den Stoffaustausch mit anderen Geweben
verantwortlich. Das Endothel weist, genau wie das Epithel, lipophile Eigenschaften
auf, d.h. es ist leichter für lipophile Stoffe passierbar, jedoch ist die Barrierefunktion
des Epithels als wesentlich bedeutsamer einzuschätzen. Die Hornhaut wird durch
Diffusion von Substanzen aus dem Kammerwasser, der Tränenflüssigkeit und
arteriellen Gefäßen am Hornhautrand (Sclera) ernährt (HUBEL 1989).
Die Tunica vasculosa bulbi setzt sich aus der Aderhaut (Chorioidea), dem
Ziliarkörper (Corpus ciliare) und der Regenbogenhaut (Iris) zusammen. Den
hintersten Abschnitt bildet die Chorioidea. Sie ist intensiv vaskularisiert und
pigmentiert. Etwa die obere Hälfte des Augenhintergrundes (Fundus) nimmt ein
halbmondförmiges lichtreflektierendes Feld, das Tapetum lucidum, ein. Es ist eine
besondere Vorrichtung der Haussäugetiere zur optimalen Ausnutzung des
einfallenden Lichtes durch die Photorezeptoren der inneren Augenhaut. Der
Ziliarkörper mit enthaltenem M. ciliaris ist die vordere Fortsetzung der Chorioidea. Er
reicht bis zum Ansatz der Iris und umgibt die Linse ringförmig. Die Oberfläche des
Ziliarkörpers ist durch Falten (Plicae ciliares) vergrößert, die mit den abgehenden
Zonulafasern die Verbindung zur Augenlinse herstellen. Der Ziliarmuskel dient zur
Akkomodation der Linse. Bei seiner Kontraktion entspannt sich der Aufhängeapparat
der Linse und diese wölbt sich aufgrund ihrer Elastizität (Nahsehen). Wenn der
Muskel entspannt ist, sind die Zonulafasern angespannt und die Linse wird flacher
(BÖHME 1992).

Literaturübersicht
19
2.1.5 Netzhaut, Retina
Die innere Augenhaut (Tunica interna bulbi), die auch Netzhaut oder Retina genannt
wird, kleidet die innere Oberfläche des Augapfels vom Pupillarrand der Iris bis zum
Sehnervenaustritt aus und ist entwicklungsgeschichtlich als eine spezielle
Modifikation der embryonalen Vorderhirnwand aufzufassen (BÖHME 1992). Retinale
sowie choroidalen Kapillargefäßen versorgte die Retina, die eine der höchsten
Metabolismusgeschwindigkeit aller Gewebe des Gesamtorganismus hat. Bei der
Katze strahlen die retinalen Gefäße, kräftig entwickelt, radiär vom blinden Fleck aus
(KÖNIG und LIEBICH 2005).
In der Netzhaut wird das auftreffende Licht, nachdem es die Hornhaut, die Linse und
den Glaskörper durchquert hat, in Nervenimpulse umgewandelt. Diese Impulse
werden von den Retinazellen an die Nervenzellen des Sehnervs, der im Thalamus
des Gehirns endet. Es lassen sich eine lichtempfindlicher Teil, die Pars optica retinae
von einen lichtunempfindliche Pars caeca retinae unterscheiden (WISSDORF et al.
1998). Infolge der beschriebenen Embryonalentwicklung bestehen die beiden Teile
aus einem Außen- und einem Innenblatt (LIEBICH 1993).
2.1.5.1 Pars optica retinae
SAMUELSON (1991) unterscheidet histologisch zehn Sichten beim Aufbau des Pars
optica retinae.
Das Stratum pigmentosum, ist das einschichtige, stark pigmentierte Außenblatt der
Pars optica retinae. Dieses retinale Pigmentepithel hat sich aus dem Außenblatt des
embryonalen Augenbechers entwickelt (LIEBICH 1993). Die Zellen der
Pigmentzellschicht sind mit der Choroidea über die Bruchmembrann (MÄTZ-
RENSING 1993) stärker in Verbindung als andere retinale Zellschichten, was ihre
besondere Bedeutung für die Ernährung der inneren Retinazellagen verdeutlicht.
Zahlreiche Einfältelungen der Zellularmembranen erleichtern den Nährstofftransport
(SAMUELSON 1991).

Literaturübersicht
20
Das Stratum nervosum, ein neunschichtiges Innennervenblatt der Pars optica
retinae, ist spezifisch umgebauter Teil der embryonalen Hirnwand (Innenwand des
Augenbechers). Von außen (choroidaseitig) nach innen (glaskörperseitig) wird das
Innenblatt durch die folgenden Schichten dargestellt (Abb. 3) (BÖHME 1992,
WISSDORF et al. 1998):
1. Stäbchen- und Zapfenschicht, Stratum neuroepitheliale,
2. äußere Grenzschicht, Stratum limitans externum, aus Gliafortsätzen bestehend,
3. äußere Körnerschicht (kernhaltige Zellkörper der Stäbchen- u. Zapfenzellen),
Stratum nucleare externum,
4. äußere retikuläre Schicht (Neuriten der Stäbchen- und Zapfenschicht),
Stratum plexiforme externum,
5. innere Körnerschicht (Kerne der Ganglienzellen, Müller-Stützzellen und amakrinen
Zellen), Stratum nucleare internum,
6. innere retikuläre Schicht, Stratum plexiforme internum,
7. Ganglienzellschicht, Stratum ganglionare n. optici,
8. Nervenfaserschicht, Stratum neurofibrarum,
9. innere Grenzschicht (Gliazellen, Müller-Stützzellen), Stratum limitans internum.
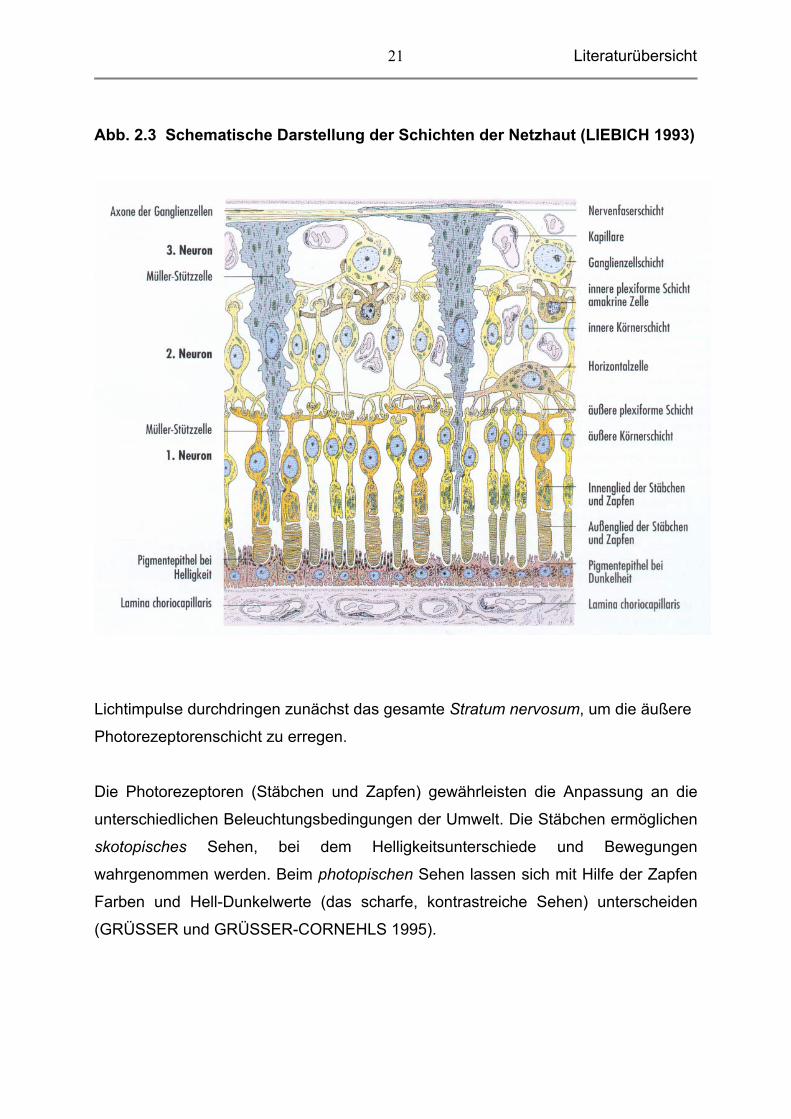
Literaturübersicht
21
Abb. 2.3 Schematische Darstellung der Schichten der Netzhaut (LIEBICH 1993)
Lichtimpulse durchdringen zunächst das gesamte Stratum nervosum, um die äußere
Photorezeptorenschicht zu erregen.
Die Photorezeptoren (Stäbchen und Zapfen) gewährleisten die Anpassung an die
unterschiedlichen Beleuchtungsbedingungen der Umwelt. Die Stäbchen ermöglichen
skotopisches Sehen, bei dem Helligkeitsunterschiede und Bewegungen
wahrgenommen werden. Beim photopischen Sehen lassen sich mit Hilfe der Zapfen
Farben und Hell-Dunkelwerte (das scharfe, kontrastreiche Sehen) unterscheiden
(GRÜSSER und GRÜSSER-CORNEHLS 1995).

Literaturübersicht
22
Katzen verfügen über eine reflektierende Schicht, Tapetum lucidum, denen Zellen
Guaninkristallen haben, hinter der Netzhaut im Auge, die jene Lichtanteile, die die
Netzhaut durchdrungen haben, zurückspiegelt, so dass diese noch ein zweites Mal
auf die Netzhaut treffen (OLIPHANT et al. 1992). Bei blauäugigen Katzen mit
weißem Fell fehlt das Tapetum lucidum (DYCE et al. 1991). Außerdem verschafft ein
weiterer Unterschied in der Anatomie der Augen den Katzen gegenüber Menschen
einen Vorteil in der Dämmerung: Ihre Pupille kann den Lichteintritt wesentlich stärker
regulieren (FREWEIN und VOLLMEHRHAUS 1994). Das Stäbchen/Zapfenverhältnis
der Netzhautrezeptoren liegt bei etwa 63:1 (beim Mensch: 20:1), variiert jedoch sehr
stark zwischen Zentrum der Netzhaut (10:1) und der Peripherie (200:1) (KOCH und
RUBIN 1972, STEINBERG et al. 1973). Im Vergleich zum Menschen reicht
Katzenaugen eine etwa siebenmal geringere Lichtintensität, um überhaupt etwas
wahrzunehmen.
Im Wesentlichen hält der Glaskörper, der durch den Augeninnendruck gegen die
Retina gepresst wird, das Innenblatt der Retina in dessen Lage, denn nur im Bereich
der Sehnervenpapille (Discus n. optici) und des Orbiculus ciliaris sind Außen- und
Innenblatt fest miteinander verbunden (BRATTON et al. 1988, WISSDORF et al.
1998). Der Discus n. optici (blinder Fleck) ist der im ventrotemporalen Bereich des
Auges gelegene Nervenfaseraustritt aus dem Auge (BRATTON et al. 1988). Dieser
Bereich besitzt keine Rezeptoren. Über die Aa. und Vv. ciliares posteriores breves
erfolgt eine direkte Versorgung des Discus n. optici. Diese radiär um die Sehnerven
austretenden Gefäße zeigen ein tierartspezifisches Muster und reichen bei Katzen
nur wenige Millimeter in die Retina (ULMER et al. 1971, BÖHME 1992).
2.1.5.2 Pars ceca retinae
Die Pars ceca retinae schließt sich der Pars optica retinae an und überzieht als
zweischichtige Epithellage, an der Ora serrata beginnend, die innere Fläche der
Procc. ciliares (Pars ciliares retinae) und der Iris (Pars iridica retinae).

Literaturübersicht
23
2.1.6 Tränenapparat, Apparatus lacrimalis Der Tränenapparat besteht aus den die Tränenflüssigkeit liefernden Drüsen (Gl.
lacrimalis, Gl. superficialis palpebrae tertiae) und einem Gangsystem, das die
Tränenflüssigkeit, nachdem sie das Auge umspült hat, in die Nasenhöhle befördert,
wo sie verdunstet (Abb. 4). Die Tränendrüse, Gl. lacrimalis, liegt bei der Katze unter
dem Ligamentum orbitale, dorsolateral auf dem Bulbus in der Orbita (BORBE 1989,
DYCE et al. 1991).
Durch die Bewegungen der Augenlider wird Tränenflüssigkeit über den exponierten
Anteilen des Auges verteilt. So wird die Cornea feucht gehalten und zusätzlich
werden Fremdkörper weggespült und die Cornea mit Nährstoffen versorgt. Die
Ausführungsgänge der Tränendrüse münden lateral in den Bindehautsack des
oberen Augenlides. Die Gl. lacrimalis wird von Nervenvasern der Äste des N.
trigeminus und des N. sympathicus versorgt (BRATTON et al. 1988). Ihre
Blutversorgung erfolgt durch die A. lacrimalis, einen Ast der A. ophtalmica externa
(BORBE 1989). Die Tränendrüse ist in der Lage, den Ausfall der Nickhautdrüse
annähernd zu kompensieren, dagegen kann der Ausfall der Tränendrüse nicht durch
die Nickhautdrüse ausgeglichen werden (BORBE 1989).
Die Tränenflüssigkeit sammelt sich im medialen Augenwinkel und wird über die zwei
schlitzförmigen Tränenpünktchen, die sich jeweils am Rande des oberen und unteren
Augenlides befinden, abgeleitet. Über die Tränenkanälchen, Canaliculi lacrimales
(Abb. 4), gelangt sie in den Tränensack, Saccus lacrimalis, der den Anfang des
Tränennasenkanals, Ductus nasolacrimalis, darstellt.
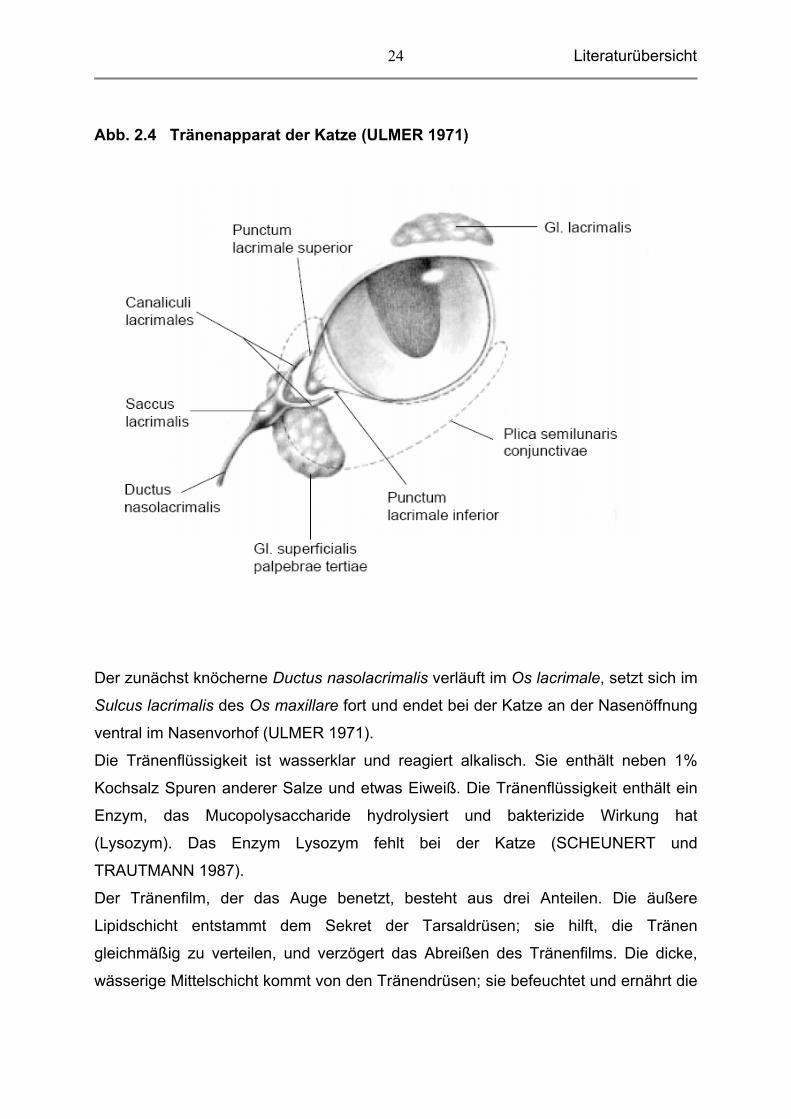
Literaturübersicht
24
Abb. 2.4 Tränenapparat der Katze (ULMER 1971)
Der zunächst knöcherne Ductus nasolacrimalis verläuft im Os lacrimale, setzt sich im
Sulcus lacrimalis des Os maxillare fort und endet bei der Katze an der Nasenöffnung
ventral im Nasenvorhof (ULMER 1971).
Die Tränenflüssigkeit ist wasserklar und reagiert alkalisch. Sie enthält neben 1%
Kochsalz Spuren anderer Salze und etwas Eiweiß. Die Tränenflüssigkeit enthält ein
Enzym, das Mucopolysaccharide hydrolysiert und bakterizide Wirkung hat
(Lysozym). Das Enzym Lysozym fehlt bei der Katze (SCHEUNERT und
TRAUTMANN 1987).
Der Tränenfilm, der das Auge benetzt, besteht aus drei Anteilen. Die äußere
Lipidschicht entstammt dem Sekret der Tarsaldrüsen; sie hilft, die Tränen
gleichmäßig zu verteilen, und verzögert das Abreißen des Tränenfilms. Die dicke,
wässerige Mittelschicht kommt von den Tränendrüsen; sie befeuchtet und ernährt die

Literaturübersicht
25
Cornea. Die innere, muköse Schicht wird von den Becherzellen der Conjunctiva
geliefert und sorgt für ein enges Anhaften des Tränenfilms an der Cornea (DYCE et
al. 1991).
2.1.7 Augenkammern und Kammerflüssigkeit
Zu den Binnenräumen des Auges gehören die beiden mit einer klaren, wäßrigen
Flüssigkeit, dem Kammerwasser, Humor aquosus, gefüllten Augenkammern, von
denen die vordere, Camera anteriror bulbi, zwischen Cornea, Iriswinkel und
Irisvorderfläche liegt, während die hintere, Camera posterior bulbi, zwischen
Irishinterfläche, Ziliarkörper, Zonula ciliaris und Linse eingeschoben ist. Die vordere
Augenkammer ist relativ geräumig und ophthalmoskopisch übersehbar, die hintere
stellt dagegen nur einen ringförmigen Spaltraum dar. Beide Kammern
kommunizieren durch die Pupille miteinander. Vom Füllungsgrad der Augenkammern
wird der Innendruck des Bulbus maßgeblich bestimmt.
2.1.7.1 Kammerwasserbildung
Bei der Katze enthält die vordere Augenkammer ca. 0,6-0,8 ml Kammerwasser und
die ist nach einer operativen Eröffnung innerhalb von ca. 1 Stunde wieder
vollständich gefüllt.
Die Bildung des Kammerwassers ist ein komplexer Vorgang, in dem die Blut-
Kammerwasser-Schranke eine besondere Rolle übernimmt. Diese Schranke ist im
Bereich der Ziliarfortsätze zwischen den Kapillarwänden und dem Ziliarepithel
entwickelt. Der aktive Transport von Natriumionen über das Ziliarepithel und der
osmotische Flüssigkeitstransport in die hintere Augenkammer sind die
Hauptmechanismen der Neubildung des Kammerwassers. Von dort fließt das
Kammerwasser durch das Sehloch in die vordere Kammer und tritt im
Kammerwinkel, Angulus iridocornealis, durch die tierartlich unterschiedlichen
spaltenförmigen Öffnungen des Ligamentum pectinatum und durch das trabekuläre

Literaturübersicht
26
Maschenwerk in den Plexus venosus sclerae über. Bestimmte Plasmaproteine
werden auch durch Diffusion durch die Kapillarwände reabsorbiert. Das
Kammerwasser dient der Ernährung der gefäßfreien Strukturen des Auges, der
Kornea und der Linse (LIEBICH 2003).
Beim Ziliarkörper handelt es sich um den verdickten, radiärstrahlig gegliederten Teil
der mittleren Aderhaut, der, bedeckt von Ziliarkörperepithel (Pars ciliaris retinae), von
der Ora serrata (Pars plana) bis zur Basis der Iris (Pars plicata) reicht. Die Pars
plicata bildet etwa 70 villiforme Ziliarkörperfortsätze aus, deren Stroma sich aus einer
Grundsubstanz (Mukopolysaccharide, Proteine und gelöste Plasmaanteile),
kollagenen Bindegewebsfasern sowie wandernden Zellen aus Bindegewebe und Blut
zusammensetzt. Das Stroma enthält ein netzartiges Kapillarsystem sowie freie
Nervenendigungen (postsynaptische sympathische und parasympathische Fasern)
für Ziliarmuskel, Gefäßsystem und/oder Ziliarkörperepithel. Das bedeckende Epithel
weist 2 Zelllagen auf, die dem inneren und äußeren Blatt des Augenbechers
entsprechen: eine äußere pigmentierte Epithelschicht und eine innere nicht
pigmentierte Epithelschicht, die an der Ora serrata in sensorische Retina und
retinales Pigmentepithel übergehen. Aufgund der embryologischen Invagination des
Augenbechers stehen die beiden Zelllagen mit ihren apikalen Seiten zueinander. Die
basolaterale Seite des pigmentierten Ziliarkörperepithels (PE) weist zum Stroma hin,
die des nicht pigmentierten Epithels (NPE) zur hinteren Augenkammer. Beide
Schichten sind untereinander und miteinander eng über ``gap junctions`` verbunden,
die es erlauben, dass elektrisches Potenzial und Ionenzusammensetzung annähernd
identisch sind und PE und NPE als Synzytium fungieren. NPE-Zellen verfügen
weiterhin über ``tight junctions``, die das Epithel zur hinteren Augenkammer hin
abdichten.
Die Kammerwasserbildung, deren Ort der Ziliarkörper ist, erfolgt in 3 Schritten:
1. über den Blutfluss zu den Ziliarkörperfortsätzen,
2. durch den Übertritt von Plasma in das Ziliarkörperstroma und
3. über aktive und passive Transportprozesse in die hintere Augenkammer.

Literaturübersicht
27
Die Blutversorgung des Ziliarkörpers erfolgt über die Aa. ciliares anteriores und
Aa. ciliares posteriores longae, Äste der A. ophthalmica, die nahe der Iriswurzel
anastomisieren und den Circulus arteriosus major der Iris aufbauen, der Iris,
Ziliarkörper und Choroidea versorgt. Kollateralkreisläufe ergänzen das Gefäßsystem.
Die Ziliarkörperfortsätze werden durch 2 verschiedene Äste aus dem Circulus
arteriosus major versorgt: die anterioren und posterioren Arteriolen, die über
Anastomosen miteinander verbunden sind. Der venöse Blutabstrom erfolgt über die
Aderhautvenen. Die Kapillaren, die als dichtes Netzwerk jeden Ziliarkörperfortsatz
durchziehen, besitzen ein dünnes, gefenstertes Endothel, durch das Plasma in den
umgebenden interstitiellen Raum gelangen kann (FREWEIN und VOLLMERHAUS
1994). Der durchschnittliche Blutfluss im Processus ciliaris liegt bei etwa 115–
154 µl/min. Etwa 4% des Plasmas treten durch die falschen Poren des
Kapillarendothels in das Stroma über. Dies entspricht einer Filtrationsrate von etwa
2,7 µl/min. Ein Teil des Filtrates gelangt nicht in das Kammerwasser, sondern
verlässt das Stroma direkt über einen uveoskleralen Abflussweg. Die
Kammerwasserproduktion erfolgt überwiegend in den vorderen Anteilen der Pars
plicata. Der Prozess der aktiven Sekretion erfolgt über die ca. 4 Mio. nicht
pigmentierte Epithelzellen, die zusammen ein Zellvolumen von etwa 8 µl aufweisen.
Die Sekretionsmenge des Kammerwassers beträgt ungefähr 2 µl/min.
Der Intraokulardruck ist das Ergebnis von Kammerwasserbildung und
Kammerwasserabfluss. Der Prozess der Kammerwasserbildung ist letztlich das
Resultat aus schneller unidirektionaler Sekretion und langsamer kontradirektionaler
Reabsorption. Die Regulation der Kammerwasserbildung erfolgt sowohl über die
Beeinflussung der Sekretion als auch der Reabsorption. Dass die
Kammerwasserbildung einem endogenen regulativen Prozess untergeordnet ist,
macht sich anhand der zirkadianen Rhythmik deutlich. Die durchschnittliche,
unbeeinflusste Sekretionsmenge des Kammerwassers liegt bei etwa 2,0–2,4 µl/min.
Während des Schlafes kommt es zu einer deutlichen Absenkung der
Kammerwassersekretionsrate von bis zu 45±20%. Änderungen des endogenen
Adrenalinspiegels werden für die tageszeitlichen Schwankungen verantwortlich
gemacht. Aufgrund von Veränderungen in der Ultrastruktur der Epithelzellen nimmt

Literaturübersicht
28
die Kammerwasserproduktion mit zunehmendem Alter ab. Pro Lebensdekade sinkt
die Sekretionsrate um etwa 2,4–3,2%. Entgegen früherer Meinung muss heute
davon ausgegangen werden, dass die Kammerwassersekretionsmenge relativ
augendruckunabhängig ist, da das als Pseudofazilität beschriebene Phänomen nur
von kurzer Dauer und meist nur gering ausgeprägt ist (FREWEIN und
VOLLMERHAUS 1994).
2.1.7.2 Kammerwasserabfluss
Der Kammerwasserabfluss erfolgt über 2 Wege: die konventionellen und die
unkonventionellen Abflusswege.
Konventionelle Abflusswege
Unter dem konventionellen Abflussweg versteht man den Kammerwasserabfluss
über das Trabekelmaschenwerk und Schlemm-Kanal. Es wird angenommen, dass
83–96% des Kammerwassers über die konventionellen Wege abfließen und sich mit
dem venösen Blut der episkleralen oder konjunktivalen Venen vermischen.
Unkonventionelle Abflusswege
Unter den unkonventionellen Abflusswegen versteht man den uveoskleralen Abfluss
und den Uveovortexabfluss, bis zu 25–50% des Kammerwassers abfließen können.
Beim uveoskleralen Abfluss fließt das Kammerwasser über die Iriswurzel und die
interstitiellen Räume des Ziliarmuskels in den Suprachoroidalraum. Den
Hauptwiderstand bildet der Ziliarmuskel. Vom Suprachoroidalraum gelangt es
entweder über Gewebsräume neben den Emissarien der Sklera zu den episkleralen
Geweben, oder es fließt entlang spezieller Kollagenstrukturen der Sklera bzw.
entlang der Blutgefäße der Optikusscheiden ab. Als treibende Kraft wird die
Druckdifferenz zwischen dem Kammerwasser und dem Suprachoroidalraum
angenommen. Beim Uveovortexabfluss gelangt das Kammerwasser über einen nicht
energieabhängigen vesikulären Transport in die venösen Gefäße der Iris, des

Literaturübersicht
29
Ziliarmuskels und der anterioren Choroidea und fließt über die Vortexvenen ab
(GRÜB und MIELKE 2004).
2.1.8 Glaskörper, Corpus vitreum
Der Glaskörper, Corpus vitreum, füllt den zwischen Linse, Zonula ciliaris, Ziliarkörper
und dem Sehteil der Netzhaut gelegenen Glaskörperraum, Camera vitrea bulbi
vollständig aus. Er besteht aus einer gallertartigen, wasserreichen und klar
durchsichtigen Masse (mit einem Wassergehalt von 99%), deren Stroma aus einem
Gerüstwerk sich durchflechtender, zartester Fibrillen aufgebaut ist, die sich an der
Glaskörperoberfläche zur Membrana vitrea verdichten. Das Maschenwerk dieses
durchsichtigen Fasergeflechtes ist mit einer wässerigen Flüssigkeit, dem Humor
vitreus, gefüllt. Die Vorderfläche des Glaskörpers wird durch die Linse zur
Linsengrube oder Fossa Hyaloidea eingedellt (BÖHME 1992).
Der Glaskörper zählt zu den dioptrischen Medien des Auges und dient dem
Metabolismus und der Homöostase der Retina. Durch Steuerung des intraokulären
Drucks hält der Glaskörper die Netzhaut in ihrer Position zum Pigmentepithel. Sinkt
der Binnendruck, so kann die Retina sich in den hinteren Bereichen ablösen,
während sie am Ziliarepithel stets mit der Pigmentschicht verbunden bleibt (LIEBICH
1993).
2.1.9 Regenbogenhaut (Iris) Die Regenbogenhaut bildet den vordersten, in das Augeninnere ziehenden Abschnitt
der mittleren Augenhaut. Sie entspringt dem Corpus ciliare, bedeckt teilweise die
Linse und begrenzt mit ihrem freien Rand Margo pupillaris das Sehloch, die Pupille.
Die Iris unterteilt als ein undurchsichtiges Diaphragma den vorderen Hohlraum des
Auges in eine vordere Augenkammer Camera anterior bulbi und eine hintere
Augenkammer Camera posterior bulbi, die über das Sehloch in Verbindung stehen
(NICKEL et al. 1992).

Literaturübersicht
30
Das Irisstroma besteht aus einem lockeren, äußerst zarten Geflecht kollagener
Faserbündel in einer amorphen Matrix mit Gefäßen, glatten Muskelzellen,
Pigmentzellen und Nervenfasern (MAURICE 1980). Die Kollagenbündel sind
bogengitterartig angeordnet. Sie können sämtlichen Größenveränderungen der Iris
während der Verengung (Miosis) oder Erweiterung (Mydriasis) folgen. Eng mit dem
Bindegewebsgerüst verbunden ist im Stroma der Iris ein ausgeprägtes Gefäßnetz
entwickelt, das neben nutritiven Aufgaben auch mechanisch-stabilisierende Funktion
übernimmt. Das Irisstroma schließt zwei glatte Muskelbündel ein, die die Größe der
Pupille regulieren, den M. sphincter pupillae und den M. dilatator pupillae (NICKEL et
al. 1992, LIEBICH 1993)
Die Iris schließt Pigmentzellen ein, die sich als Stromamelanozyten aus der
Neuralleiste und als Pigmentepithelien aus dem Neuroektoderm ableiten.
Melaninpigmente schützen die Retina vor übermäßiger Lichtstrahlung und vor
Streulicht, in dem sie in der Iris einen neutralen Dichtefilter aufbauen. Der Grad der
Pigmentierung (Größe und Anzahl der Melanosomen) bestimmt die Farbe der Iris
und damit die „Farbe der Augen“.
Die Farbe der Iris, die dem Ausdruck des Auges seinen besonderen Charakter
verleiht, schwankt nach Tierart und Rasse, aber auch individuell, beträchtlich. Je
dichter die Pigmentzellen liegen, umso dunkler braun ist sie gefärbt. Die hell-braune
und gelbliche Irisfarbe, wie sie vor allem bei Hund vorkommt, beruht im Wesentlichen
auf einem geringeren Pigmentgehalt. Die schillernd gelbe oder gelbgrüne Farbe der
Regenbogenhaut der Katze wird auf gelöstes Pigment in den Stromazellen
zurückgeführt. Einer Blau- oder Graublaufärbung der Iris begegnet man oft bei rein
weißen Katzen. In solchen Fällen fehlt das Stromapigment vollständig, während das
Pigment der Pars iridica retinae durch die farblosen Schichten der Stromas
hindurchschimmern (KÖNIG 1992). Im albinotischen Auge fehlt das Pigment auch in
der Pars iridica retinae so wie in den Schichten des Augenhintergrundes mehr oder
weniger vollständig, womit dann infolge der durchschimmernden Blutgefäße die

Literaturübersicht
31
bekannte Rotfärbung zustande kommt. In der Regel besitzen beide Augen die
gleiche Irisfarbe (LIEBICH 1993, WILCOCK 1993).
2.1.10 Linse, Lens Das sich vom Ektoderm abschnürende Linsenbläschen wird zur kompakten,
bikonvexen Linse, Lens, von nahezu kreisrundem Umriss (Abb. 5). Die glasklare,
durchsichtige Linsensubstanz, Substantia lentis, besteht aus einer weicheren Rinde,
Cortex lentis, und einem konsistenteren Kern, Nucleus lentis, der dank seiner
Elastizität die Tendenz hat, die Linse zur Kugelform abzurunden. Der vordere, der
Pupille zugekehrte Pol der Linse, Polus anterior lentis, ist flacher, der hintere, dem
Glaskörper zugewandte Linsenpol, Polus posterior lentis, stärker gewölbt. Der
Linsendurchmesser beträgt bei der Hauskatze bis zu 14 mm, die Längsachse bis zu
9 mm (FATH EL BAB et al. 1981, FREEWEIN und VOLLMERHAUS 1994)
Abb. 2.5 Schematische Darstellung des Augenvordergrundes (LIEBICH 1993)

Literaturübersicht
32
Die vom Kammerwasser bespülte vordere Linsenfläche, Facies anterior lentis, liegt
direkt hinter der Pupille und wird von den Pupillarränder der Iris berührt. Die hintere
Linsenfläche, Facies posterior lentis, dagegen liegt in die Linsengrube des
Glaskörpers eingebettet. In der Linse kommen weder Blutgefäße noch Nerven vor
(FATH El BAB et al. 1981). Sie ist das einzige Organ des gesamten Organismus, das
nur aus Epitelgewebe besteht. Die Ernährung des Linsengewebes erfolgt über das
Kammerwasser (BÖHME 1992).
Die Akkomodation auf kurze Distanz der auf Fernsicht eingestellten Linse wird durch
Kontraktion des in die Grundplatte des Ziliarkörpers eingebauten M. ciliaris
bewerkstelligt, dessen Muskelfasern teils zirkulär, teils meridional verlaufen. Die
Kontraktion des M. ciliaris bewirkt eine Verlagerung des Ziliarkörpers gegen die
vordere Augenkammer, wodurch der Durchmesser des Aufhängeapparates
verkleinert, dieser der Augenachse genähert wird und die Zonulafasern entspannt
werden. Die Linse rundet sich daraufhin dank ihrer Elastizität ab, ein Vorgang, der
funktionell die Akkomodation bedeutet.
Gut entwickelt ist der M. ciliaris bei den Fleischfressern, bei denen ein scharfes
Sehen auf kurze Distanz vor allem beim Beutefang wichtig ist.
Besondere Strukturen der Linse sind Linsenkapsel (Capsula lentis), Linsenepithel
(Epithelium lentis) und Linsenfasern (Fibrae lentis) (KÖNIG 1992).
Die Linse wird von einer lichtbrechenden, in der Regel sehr dehnungsfähigen Kapsel
umgeben, die aus Sekreten des Linsenepithels entsteht. Am äußeren Rand
inserieren die Zonulafasern des Aufhängeapparates der Linse und verbinden sich mit
den Filamentbündeln der Linsenkapsel. Die Linsenkapsel ist eine semipermeable
Grenzschicht, durch die metabolisch aktive Stoffe penetrieren (LIEBICH 1993)
Das Linsenepithel liegt an der Vorderfläche der Linse unter der Linsenkapsel, es ist
einschichtig isoprismatisch. Die Epithelzellen der Linse sind die einzigen
teilungsfähigen Zellen dieses Organs, die sich verlängern und zu Linsenfasern
differenzieren. Vorzugsweise am Linsenäquator teilen sich die Linsenepithelzellen

Literaturübersicht
33
und lagern sich als neue Linsenfasern appositionell dem vorhandenen Linsenkörper
auf. Die Linse wächst zeitlebens. Das Wachstum ist eng mit dem Alter des Tieres
verbunden (WISSDORF et al. 1998; LIEBICH 1993).
Die Linsenfasern sind lang gestreckte, prismatische Zellen, die die Hauptmasse der
Linse bilden. Die einzelnen Linsenfasern stehen durch reißverschlußähnliche
Verzahnungen untereinander in Verbindung. Dieser Bau ermöglicht bei der
Akkomodation der Linse die Plastizität des gesamten Organs.
Die Linsenfasern schließen Wasser (70%) und im Wesentlichen 35%
Membranproteine, Zytoskelettproteine, Enzyme, Kristalline und Elektrolyte ein (GUM
1991). Die Form der Linse wird entscheidend von der Struktur ihres Zytoskeletts
geprägt, dazu tragen Mikrofilamente und Mikrotubuli bei (FATH El BAB et al. 1981).
Die geordnete Struktur und der Metabolismus der Linsenfasern sind für die
Aufrechterhaltung der Transparenz von entscheidender Bedeutung. Jede Änderung
führt zur Trübung der Linse.

Literaturübersicht
34
2.2 Corticosteroide Corticosteroide sind Hormone, die im ganzen Organismus wichtige physiologische,
biochemische und pathologische Prozesse beeinflussen (ESTLER 1995).
Unter Corticosteroiden versteht man Steroidhormone der Nebennierenrinde
einschließlich ihrer synthetischen Derivate. Nach ihrer biologischen Wirkung lassen
sich die Nebennierenrindenhormone in drei Gruppen einteilen:
1. Glucocorticoide, mit bevorzugter Wirkung auf den Kohlenhydratstoffwechsel,
2. Mineralocorticoide, mit bevorzugter Wirkung auf den Elektrolythaushalt, und
3. Androgene wie Androstendion, Dehydroepiandrosteron (DHEA) und sein
Sulfat (KNEPEL 2005).
2.2.1 Glucocorticoide Glucocorticoide (GC) sind seit ihrer erstmaligen klinischen Anwendung 1948 durch
HENCH et al. (1949) zu einem unverzichtbaren Bestandteil der medikamentösen
Therapie in der modernen Medizin geworden.
Sie werden wie alle Steroidhormone aus Cholesterin synthetisiert. Die wichtigsten
physiologischen Vertreter sind Cortisol, Corticosteron und Cortison, eine Vorstufe
des Cortisols (KAISER und KLEY 1992). Synthetisch hergestellte GC leiten sich
überwiegend vom Prednisolon ab. Ein wichtiger Vertreter ist Dexamethason. Es ist
lang wirksam und hat eine sehr starke entzündungshemmende Wirkung aller GC
(KAISER und KLEY 1992).
Synthetische GC zeigen prinzipiell das gleiche Wirkspektrum wie endogene GC,
jedoch sind die meist unerwünschten Effekte auf den Wasser- und Elektrolythaushalt
reduziert oder wie bei Dexamethason fast vollständig eliminiert (ROHDEWALD et al.
1986).

Literaturübersicht
35
Glucocorticoide sind Steroidhormone, welche in der Zona fasciculata der
Nebennierenrinde aus Cholesterol synthetisiert und grundsätzlich als katabole
Stoffwechselhormone betrachtet werden (SHARPE et al. 1986). Daneben wirken sie
in pharmakologischen Dosierungen immunsuppressiv und entzündungshemmend, so
daß sie therapeutisch bei allergischen und besonders chronisch entzündlichen
Prozessen eingesetzt werden (THUN und SCHWARTZ-PORSCHE 1994).
2.2.2. Synthese und Sekretion
Die Glucocorticoide werden überwiegend in der Zona fasciculata der
Nebennierenrinde gebildet.
Die Sekretion der Glucocorticoide erfolgt auf hypothalamisch-hypophysärer Achse
über Stimulation durch das Corticotropin-releasing Hormon (CRH) und das
Adrenocorticotrope Hormon (ACTH). Die Glucocorticoide selbst hemmen die
Freisetzung von CRH und ACTH. Für die Sekretion der Glucocorticoide wurde beim
Menschen sowie bei verschiedenen Tierarten eine circadiane Rhythmik gefunden,
mit höchsten Konzentrationen in den frühen Morgenstunden und niedrigsten in den
Abendstunden (JANNING 1993, THUN und SCHWARTZ-PORSCHE 1994).
ACTH bindet an Rezeptoren von Nebennierenzellen der Zona fasciculata und
aktiviert (über Erhöhung der Konzentration von cAMP) die Synthese der
Glucocorticoide Cortisol, Cortison und Corticosteron aus Cholesterin und die
Proliferation der aktivierten Zellen. Glucocorticoide wirken über spezifische
cytosolische Rezeptoren, die in nahezu allen Geweben nachgewiesen werden
konnten. Der bedeutendste Teil des Glucocorticoid-Rezeptors (GCR) wurde erst
kürzlich sequenziert (GUTSCHER et al. 2001). Nach Rezeptorbindung erfolgt die
Translokation in den Zellkern, wo Interaktionen auf Gen-Ebene die Transkription
verschiedenster Faktoren modulieren und so auch in die Regulation der meisten
Hormone eingreifen. Glucocorticoide können zusätzlich die Stabilität der mRNA über
Enzyme und Bindungsproteine verändern (GELEY et al. 1997).

Literaturübersicht
36
Die NNR ist nicht imstande, ihre Sekretionsprodukte zu speichern und erst bei Bedarf
auszuschütten. Glucocorticoide diffundieren nach der Synthese aus der Zelle. Im Blut
werden sie an das α-Globulin Transcortin gebunden transportiert. Die Konzentration
liegt bei 200-500ng/ml. Die Halbwertszeiten betragen 1,5-2 Stunden für Cortisol und
etwa 1 Stunde für Corticosteron. Diesem Sekretionstyp ist ein circadianer Rhythmus
überlagert (LABHART und MÜLLER 1978, HERRTAGE 1996).
Bei der Abnahme der Konzentrationen an freiem Cortisol im Blut wird im
Hypothalamus CRH abgegeben, das in der Hypophyse die Freisetzung von ACTH
bewirkt. Auch beim Hund werden Maximalkonzentrationen morgens und
Minimalkonzentrationen abends beachtet, bei der Katze ist es dagegen umgekehrt
(FREY 1996). Bei der Katze treten Maximalwerte abends auf (UNGEMACH 1999).
Über eine negative Rückkopplung (negatives Feedback) wirken die Glucocorticoide
sowohl auf den Hypothalamus als auch auf die Hypophyse. Die negative
Rückkopplung besitzt zwei Phasen. Die schnelle Phase erfolgt beim Anfluten der
Glucocorticoide, wahrscheinlich durch Sekretionshemmung von CRH und ACTH. Die
Synthesehemmung von ACTH erfolgt in der langsameren Phase (OETTEL 1996).
Neben der Steuerung durch negative Rückkopplung beeinflussen auch Stress und
ein circadianer Rhythmus die Konzentration der Glucocorticoide im Blut (OETTEL
1996). HEUSER (1995) beobachtet eine Schwächung der Glucocorticoid–
Rückkopplung bei jungen Tieren unter Stressbedingungen. Der circadiane Rhythmus
überlagert die episodische Sekretion von CRH, ACTH und von Glucocorticoiden
(THUN et al.1990).
2.2.3 Wirkung der Glucocorticoide
Die glucocorticoiden Wirkungen treten erst nach einer Latenzzeit auf und bleiben
über die Elimination aus der Blutbahn hinaus erhalten. Auch wenn sich keine

Literaturübersicht
37
Glucocorticoid-Rezeptor-Komplexe mehr im Zellkern befinden, kann noch eine
Wirkung bestehen (JUSKO et al. 1980, GUSTAFSSON et al. 1989).
2.2.3.1 Allgemeine Wirkmechanismen
Alle Glucocorticoide binden an den spezifischen Glucocorticoidrezeptor, der im
Cytosol in inaktiver Form an das Hitzeschockprotein HSP 90 gebunden ist. Nach
Hormonbindung kommt es nach Dissoziation von diesem Protein zu einer
Dimerisierung, und der Rezeptor-Glucocorticoid-Komplex transloziert in den Zellkern,
wo er an die Glucocorticoid-responsiven Elemente (GRE) der DNA bindet. Durch
diese Elemente wird die Transkriptionsrate von zahlreichen Proteinen verändert.
Neben der erhöhten Synthese vor allem von Enzymproteinen kann die Synthese
anderer Proteine abgeschaltet werden (KNEPEL 2005).
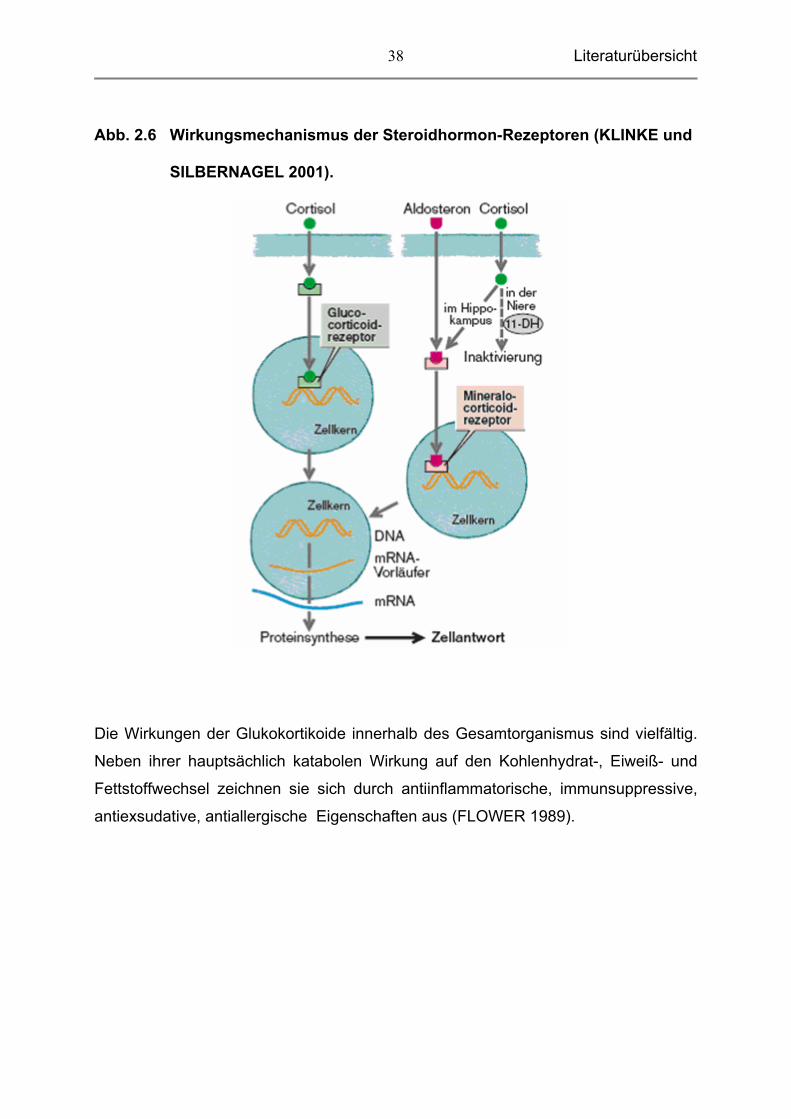
Literaturübersicht
38
Abb. 2.6 Wirkungsmechanismus der Steroidhormon-Rezeptoren (KLINKE und
SILBERNAGEL 2001).
Die Wirkungen der Glukokortikoide innerhalb des Gesamtorganismus sind vielfältig.
Neben ihrer hauptsächlich katabolen Wirkung auf den Kohlenhydrat-, Eiweiß- und
Fettstoffwechsel zeichnen sie sich durch antiinflammatorische, immunsuppressive,
antiexsudative, antiallergische Eigenschaften aus (FLOWER 1989).

Literaturübersicht
39
2.2.3.2 Wirkungen auf den Stoffwechsel
Glucocorticoide fördern die Gluconeogenese aus Aminosäuren, die durch Abbau von
Proteinen frei werden. Die neu gebildete Glucose wird z.T. als Glukogen in der
Leber gespeichert. Der Glucoseumsatz wird gesteigert, die Glucosetoleranz und die
Insulinempfindlichkeit nehmen ab (KNEPEL 2005, GELEY et al. 1996). Durch
Glucocorticoide kann ein Prädiabetes in einen latenten oder klinisch manifesten
Diabetes überführt werden – Steroiddiabetes (BENKER 1995). Im Gegensatz zum
Diabetes mellitus ist der Steroiddiabetes nach Absetzen der Glucocorticoide
reversibel (OETTEL 1996).
Glucocorticoide fördern den Proteinabbau und führen zu einer negativen
Stickstoffbilanz, z.T. auch bedingt durch vermehrte Ausscheidung von Aminosäuren
und Harnsäure (KNEPEL 2005). Die katabole Wirkung zeigt sich vor allem in der
Muskulatur, der Haut (OETTEL 1996) und dem Skelett. Sie führt beim Jungtier zu
einer Wachstumshemmung (THUN und SCHWARTZ-PORSCHE 1994).
Glucocorticoide beeinflussen den Fettstoffwechsel, indem sie die lipolytische
Wirkung von Katecholaminen und Wachstumshormon fördern. Andererseits führen
hohe Konzentrationen von Glucocorticoiden zu einer Umverteilung von Fettgewebe.
Es kommt zur Steigerung der zirkulierenden Triglyceride (KÖBBERLING und
ROTENBERGER 1993). Durch Glucocorticoide wird Fettgewebe mobilisiert
(Lipolyse), gleichzeitig wird die Fettsäuresynthese in der Leber gehemmt (MÖSTL
2000). Es tritt Fettverlust an den Extremitäten und Fettzunahme am Stamm sowie im
Nacken und Gesicht auf. Dies führt zum typischen Bild der Stammfettsucht beim
Cushing-Syndrom (OETTEL 1996, KNEPEL 2005).

Literaturübersicht
40
2.2.3.3 Wirkungen auf den Wasser- und Elektrolythaushalt
Die Effekte der Corticoide auf Wasser- und Elektrolythaushalt werden durch zwei
homologe Steroidrezeptoren vermittelt: den Mineralocorticoidrezeptor (MR) und den
Glucocorticoidrezeptor (GR) (DE KLOET et al. 1986). Beide Rezeptoren gehören zu
der Familie der ligandenabhängigen Transkriptionsfaktoren (MANGELSDORF et al.
1995). Der MR wird vor allem in limbischen Strukturen, wie dem Hippocampus, dem
cerebralen Septum, und den Amygdala exprimiert und trägt wesentlich zur
Regulation der basalen Aktivität der Hypothalamus-Hypophysen-Nebennierenrinden-
Achse (HPA-Achse) bei (DE KLOET et al. 1993, DEUSCHLE et al. 1998). Der GR
hingegen wird in weiten Bereichen des Gehirns exprimiert, vor allem im limbischen
System, in parvozellulären Neuronen des paraventriculären Kerns und im
Hirnstamm. Aber auch in anderen Organen, wie der Lunge, der Leber, dem
Knochenmark und dem Immunsystem, ist der GR an wichtigen, physiologischen
Prozessen beteiligt (EVANS-STORMS und CIDLOWSKI 1995, SCHMID et al. 1995).
2.2.3.4 Wirkungen über den Mineralocorticoidrezeptor
Die natürlichen Glucocorticoide sowie einige synthetische Analoga besitzen
mineralocorticoide Wirkung. Sie steigern im distalen Tubulus und an den
Sammelrohren der Niere die Reabsorption von Natrium; gleichzeitig steigern sie die
renale Ausscheidung von K+ und H+. Dieser Wirkung liegt wahrscheinlich die über
den Mineralocorticoidrezeptor vermittelte schnelle Steigerung der Transkription des
Gens der SGK-Kinase (serum- and glucocorticoid-induced kinase) zu Grunde. Diese
phosphoryliert und aktiviert einen amiloridsensitiven epithelialen Natriumkanal
(ENaC) (DE KLOET et al. 1986, MC EWEN et al. 1986).
Langsamer (über Stunden) erfolgt auch eine Synthese von Untereinheiten des
Natriumkanals und der Na+-K+-ATPase. Durch Förderung der Natrium- und damit
Wasserretention nimmt das Volumen des Extrazellularraums zu, durch vermehrte

Literaturübersicht
41
Kaliumausscheidung kommt es zu Hypokaliämie und metabolischer Alkalose. Auch
die Magnesiumausscheidung wird durch Mineralocorticoide gesteigert (KNEPEL
2005).
2.2.3.5 Wirkungen über den Glucocorticoidrezeptor
Der bedeutendere Rezeptor ist jedoch der Glucocorticoid-Rezeptor (DE KLOET et al.
1986, MC EWEN et al. 1986). Über ihn werden die meisten Glucocorticoid-Effekte
vermittelt. Dies sind die Wirkungen auf den Stoffwechsel wie die Förderung der
Gluconeogenese, die Steigerung des Glucoseumsatzes, die Senkung der
Glucoseutilisation und der Glucosetoleranz, sowie letztendlich die Steigerung der
Glucosekonzentration im Plasma. Auf den Proteinstoffwechsel wirken
Glucocorticoide katabol. Sie fördern die lipolytische Wirkung von Katecholaminen
und bewirken eine Fettumverteilung von den Extremitäten zum Körperstamm.
Von einigen Wirkungen der Corticosteroide auf Wasser- und Elektrolythaushalt muss
angenommen werden, dass sie über den Glucocorticoidrezeptor vermittelt werden.
Unter Cortisol ist die glomeruläre Filtration gesteigert und wird die renale
Ausscheidung von Wasser über Vasopressinabhängige und –unabhängige
Mechanismen erhöht. Glucocorticoide hemmen auch die Resorption von Calcium
über die Nieren. Diese Mechanismen sowie die erhöhte Phosphatclearance bewirken
eine vermehrte Calcium- und Phosphatausscheidung und fördern die Entwicklung
einer Osteoporose. Die hypocalcämie Wirkung wird beim Gesunden kompensiert,
kann aber therapeutisch bei Hypercalcämien genutzt werden (OETTEL 1996,
KNEPEL 2005).
2.2.3.6 Wirkungen auf das cardiovaskuläre System
Die Effekte von Glucocorticoiden auf das cardiovaskuläre System beruhen zum Teil
auf mineralocorticoiden und zum Teil auf glucocorticoiden Wirkungen.

Literaturübersicht
42
Glucocorticoide wirken positiv inotrop, erhöhen die Ansprechbarkeit der
Mikrozirkulation auf Noradrenalin und verbessern dadurch die lokale Durchblutung
(FREY 2002).
Auf das Herz üben die Glucocorticoide einen positiven inotropen Effekt aus. Unter
Einwirkung der Glucocorticoide nehmen Herz-Zeit-Volumen und arterieller Blutdruck
zu, der periphere und pulmonale Gefäßwiderstand sinken. Glucocorticoide
reduzieren die Kapillarpermeabilität. Die Ansprechbarkeit der kleinen Gefäße auf
Catecholamine (permissive Wirkung) wird gesteigert, so dass es zu einer
verbesserten Mikrozirkulation beim anaphylaktischen Schock kommt (OETTEL
1996).
2.2.3.7 Wirkungen am ZNS
Am ZNS wirken Glucocorticoide euphorisierend und lösen ein subjektives Gefühl des
Wohlbefindens aus. Als Folge kommt es bei Tieren, insbesondere bei Rindern, im
Zusammenwirken mit der Steigerung des Blutzuckerspiegels zu einer
Appetitanregung, und es beginnen auch schwerkranke Tiere wieder zu fressen, ohne
dass sich die Grundkrankheit gebessert hat. Diese Wirkung wird häufig als
scheinbare Besserung fehlinterpretiert. Verschiedentlich kommt es zu
Erregbarkeitssteigerungen und bei Hunden zu Bösartigkeit. Hunde und Katzen
können vereinzelt auch mit Depression reagieren (UNGEMACH et al 2002).
Unabhängig von der indirekten Beeinflussung des ZNS durch Effekte auf
Stoffwechsel, Elektrolythaushalt und Zirkulation scheinen Glucocorticoide auch
direkte Effekte zu haben. Sie steigern die Erregbarkeit des Gehirns, die Reizschwelle
für eine Reihe von Stimuli wird gesenkt; EEG-Veränderungen werden beobachtet
(OETTEL 1996).
Zu den Wirkungen im Gehirn zählt auch die Hemmung der Synthese und Sekretion
von CRH im Hypothalamus im Rahmen der negativen Rückkopplung durch

Literaturübersicht
43
Glucocorticoide. Dabei scheinen sowohl Mineralocorticoid- als auch
Glucocorticoidrezeptoren im Hippocampus beteiligt zu sein.
2.2.3.8 Wirkungen auf das Blut, die Skelettmuskulatur und das Wachstum
Glucocorticoide können die Zunahme der Erythrocytenzahl und der
Hämoglobinkonzentration bewirken, vermutlich durch Verzögerung des
Erythrocytenabbaus (THUN und SCHWARTZ-PORSCHE 1994).
Die Zahl der Thrombocyten und neutrophilen Granulocyten im Blut nimmt unter
Glucocorticoiden zu. Dagegen nimmt die Zahl der Lymphocyten, Monocyten,
eosinophilen und basophilen Granulocyten als Folge einer Umverteilung ab. Die
Skelettmuskulatur wird durch Glucocorticoide indirekt durch Beeinflussung der
Zirkulation und des Stoffwechsels, aber auch direkt beeeinflusst. Wachstum und
Zellteilung werden durch Glucocorticoide beeinflusst.
Die DNA-Synthese und die Zellteilung in einer Reihe von Geweben, z.B.
Fibroblasten, Magenschleimhaut, Epidermis, Thymocyten u.a., werden durch
Glucocorticoide gehemmt. Dieser Effekt scheint selektiv zu sein, da andere Gewebe
wie Knochenmark und Darmschleimhaut nicht betroffen sind (KNEPEL 2005).
2.2.3.9 Antiinflammatorische und immunsuppressive Wirkungen
Glucocorticoide wirken antiinflammatorisch und immunsuppressiv. Sie beeinflussen
zahlreiche andere endokrine Systeme wie die Schilddrüse, das Reproduktionssystem
sowie das Wachstum (CHARMANDARI et al. 2003). Speziell auf diesen Wirkungen
beruht die breite therapeutische Anwendung der Glucocorticoide. Obwohl sie nicht
die zu Grunde liegende Ursache einer Erkrankung beiseitigen, besitzt die potente
entzündungshemmende und immunsupressive Wirkung der Glucocorticoide einen
enormen Klinischen Wert.

Literaturübersicht
44
2.2.4 Nebenwirkungen der Glucocorticoide
Unter hochdosierter Langzeittherapie werden Cushing-ähnliche Effekte beobachtet
(FELDMANN und NELSON 1996, FERGUSON und HOENIG 2001). Katzen können
schneller eine Glukoseintoleranz entwickeln als Hunde. Mehr als 95% der Katzen mit
einem spontanen Cushing Syndrom leiden gleichzeitig an Diabetes mellitus (IMMNIK
et al. 1992, NELSON et al. 1988); unter der Glucocorticoid -Therapie wurde eine
Hyperglykämie bereits mit einer Dosierung von 2 mg/kg Prednisolon während 8
Tagen festgestellt (MIDDLETON und WATSON 1985, MIDDLETON et al. 1987,
SCOTT et al. 1979).
Typische unerwünschte Wirkungen bei der Katze: Polyurie, Polydipsie, Polyphagie
mit Gewichtszunahme, Durchfall, Depression.Glucocorticoide wirken am Auge
infektionsbegünstigend. Zubereitungen ohne antibiotischen Zusatz bergen die Gefahr
der Infektion, teilweise mit Penetration der Keime in die Cornea. Glucocorticoide
verhindern die Heilung von Ulzera und vor allem beim Vorliegen von Epithelläsionen
kann es zur Entwicklung von Ulzera kommen. Des Weiteren erhöhen sie den
Augeninnendruck, was insbesondere bei Langzeitanwendung zum Glaukom führen
kann (UNGEMACH 1999); die durch Glucocorticoide induziererten morphologische
Veränderungen im trabekulären Netzwerk werden hauptsächlich für die Entstehung
eines Glaukoms verantwortlich gemacht (TRIPATHI et al. 1999). Bei lokaler Therapie
tritt diese Komplikation häufiger auf als bei langer systemischer Behandlung. Vor
allem die Dauer der Therapie ist entscheidend (STRAUB 1992). Die Druckerhöhung
ist in etwa 30 % der Fälle reversibel, solange noch keine morphologischen
Veränderungen des Kammerwinkels eingetreten sind (ROHEN 1973). Auch die
Katarakt ist eine mögliche Folge der Glucocorticoide Anwendung (HUNTER et al.
1973, UNGEMACH 1999, SMEETH et al. 2003). Linsentrübungen wurden auch nach
systemischer Anwendung von Steroiden beschrieben (BLACK et al. 1960). In Tabelle
1 sind die häufigsten Nebenwirkungen zusammengefasst.
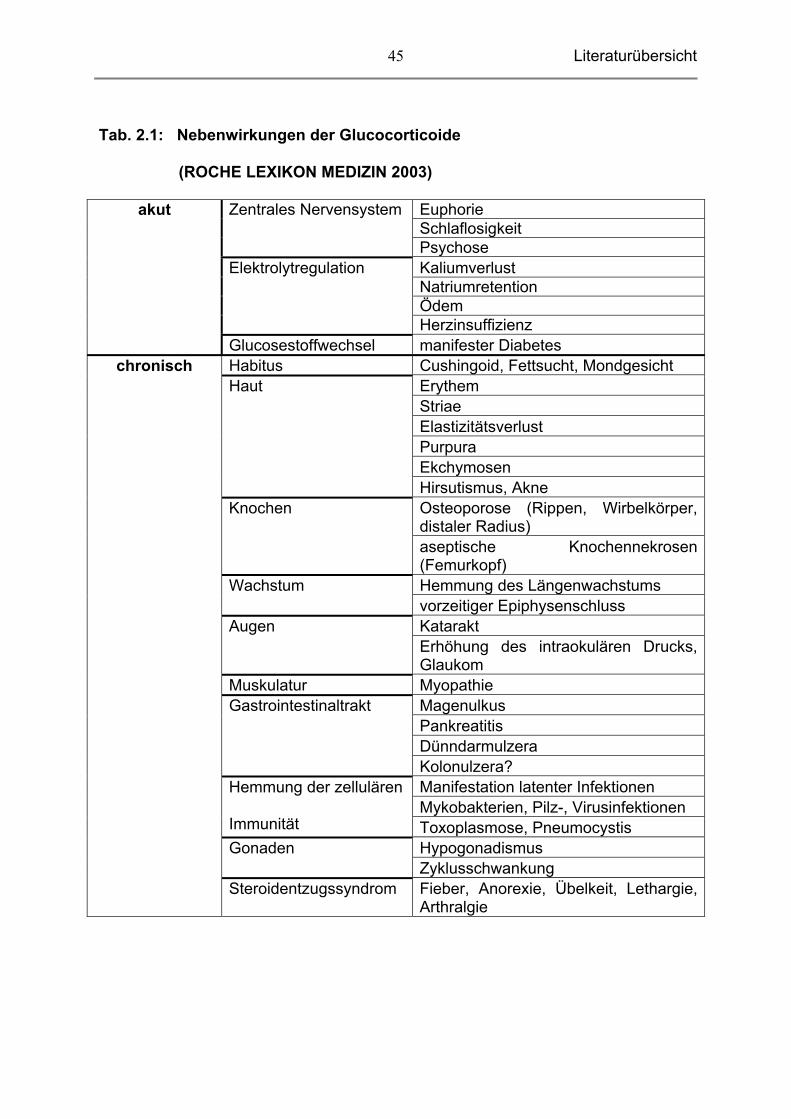
Literaturübersicht
45
Tab. 2.1: Nebenwirkungen der Glucocorticoide
(ROCHE LEXIKON MEDIZIN 2003)
Euphorie Schlaflosigkeit
Zentrales Nervensystem
Psychose Kaliumverlust Natriumretention Ödem
Elektrolytregulation
Herzinsuffizienz
akut
Glucosestoffwechsel manifester Diabetes Habitus Cushingoid, Fettsucht, Mondgesicht
Erythem Striae Elastizitätsverlust Purpura Ekchymosen
Haut
Hirsutismus, Akne Osteoporose (Rippen, Wirbelkörper, distaler Radius)
Knochen
aseptische Knochennekrosen (Femurkopf) Hemmung des Längenwachstums Wachstum vorzeitiger Epiphysenschluss Katarakt Augen Erhöhung des intraokulären Drucks, Glaukom
Muskulatur Myopathie Magenulkus Pankreatitis Dünndarmulzera
Gastrointestinaltrakt
Kolonulzera? Manifestation latenter Infektionen Mykobakterien, Pilz-, Virusinfektionen
Hemmung der zellulären
Immunität Toxoplasmose, Pneumocystis Hypogonadismus Gonaden Zyklusschwankung
chronisch
Steroidentzugssyndrom Fieber, Anorexie, Übelkeit, Lethargie, Arthralgie

Literaturübersicht
46
2.2.5 Cortisol und seine synthetischen Derivate
Bald nach der Entdeckung der klinischen Effekte des Cortisons bemühte man die
Substanz so abzuwandeln, dass die Wirkung erhalten blieb, die unerwünschten
Nebenwirkungen aber gemindert wurden. Dabei sollten die mineralocorticoiden
"Nebenwirkungen" von den erwünschten glucocorticoiden Effekten getrennt werden
(BETTENDORF 1995). Ein erster Schritt bei der Synthese neuer Glucocorticoide war
die Einführung einer Doppelbindung zwischen Position 1 und 2. Prednison und
Prednisolon waren als Produkt dieser Molekularveränderung um den Faktor 4-5
wirksamere Glucocorticoide. Ihre mineralocorticoide Wirkung war auf etwa die Hälfte
reduziert. Eine wichtige Epoche begann mit der Halogenierung des Steroidmoleküls
in Position 6,7 und/oder 9. Mit den halogenierten Glucocorticoiden wurde eine
erhebliche Wirkungssteigerung bei nahezu völligem Fehlen der Mineralcorticoid-
Wirkung erreicht (z.B. Dexamethason und Triamcinolon) (FREY 1996).
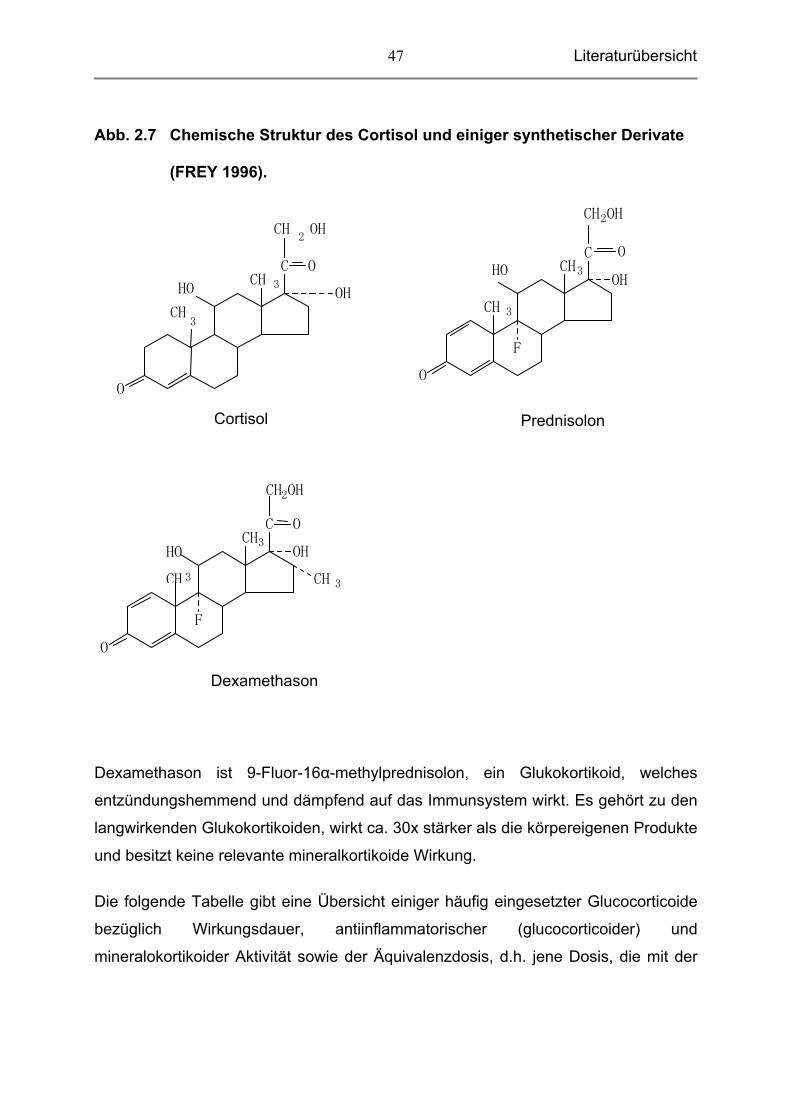
Literaturübersicht
47
Abb. 2.7 Chemische Struktur des Cortisol und einiger synthetischer Derivate
(FREY 1996).
Dexamethason ist 9-Fluor-16α-methylprednisolon, ein Glukokortikoid, welches
entzündungshemmend und dämpfend auf das Immunsystem wirkt. Es gehört zu den
langwirkenden Glukokortikoiden, wirkt ca. 30x stärker als die körpereigenen Produkte
und besitzt keine relevante mineralkortikoide Wirkung.
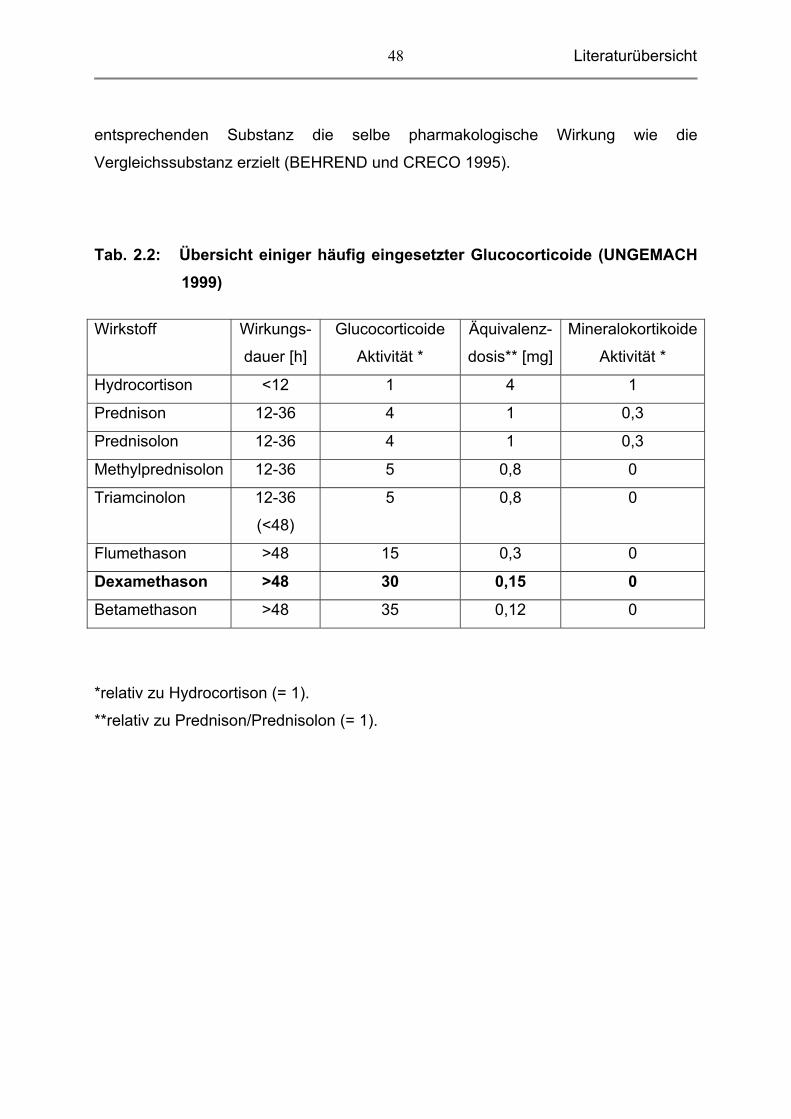
Die folgende Tabelle gibt eine Übersicht einiger häufig eingesetzter Glucocorticoide
bezüglich Wirkungsdauer, antiinflammatorischer (glucocorticoider) und
mineralokortikoider Aktivität sowie der Äquivalenzdosis, d.h. jene Dosis, die mit der
O
CH3
HO
CH 3
OH
F
CH 3 C O
CH 2 OH
Dexamethason
HO
O
CH 3 C
CH 3
O
OH
2
Cortisol
OH
F
O
C CH 3 HO
CH 3
O
CH 2 OH
Prednisolon
CH OH
Literaturübersicht
48
entsprechenden Substanz die selbe pharmakologische Wirkung wie die
Vergleichssubstanz erzielt (BEHREND und CRECO 1995).
Tab. 2.2: Übersicht einiger häufig eingesetzter Glucocorticoide (UNGEMACH 1999)
Wirkstoff Wirkungs-
dauer [h]
Glucocorticoide
Aktivität *
Äquivalenz-
dosis** [mg]
Mineralokortikoide
Aktivität *
Hydrocortison <12 1 4 1
Prednison 12-36 4 1 0,3
Prednisolon 12-36 4 1 0,3
Methylprednisolon 12-36 5 0,8 0
Triamcinolon 12-36
(<48)
5 0,8 0
Flumethason >48 15 0,3 0
Dexamethason >48 30 0,15 0
Betamethason >48 35 0,12 0
*relativ zu Hydrocortison (= 1).
**relativ zu Prednison/Prednisolon (= 1).

Literaturübersicht
49
2.2.6 Pharmakokinetik von Ophthalmica
Bei Behandlung von Augenerkrankungen bestehen lokale so wie auch systemische
Applikationsmöglichkeiten. Die systemische Applikation, wie z.B. intravenös oder
oral, hat den Nachteil, dass die Medikamente nicht nur lokale Wirkungen, sondern
auch systemische Nebenwirkungen zeigen (SCHOENWALD 1993), die noch
verstärkt werden, weil zur Überwindung der Diffusionsbarrieren im Auge
Blut/Netzhaut und Blut/Kammerwasser- sowie die Blut/Glaskörper-Schranke
(RAVIOLA 1977) sehr hohe Blutwirkstoffspiegel nötig sind. Ziel der medikamentösen
Therapie von Augenerkrankungen ist eine hohe Bioverfügbarkeit im Auge bei
möglichst geringen systemischen Nebenwirkungen. Neben dem geringen
Vorhandensein oder dem vollständigen Fehlen von Blutgefäßen in verschiedenen
Abschnitten des Auges bedingt die aktive Sekretion von intraokulären Flüssigkeiten
allgemein einen nur geringen Übertritt von Komponenten aus dem Blutplasma in
intraokuläre Bereiche (BINKHORST 1987).
Aus diesen Gründen ist die lokale Therapie der systemischen oft vorzuziehen
(MAUGER 1994). Die lokalen Applikationsarten umfassen die topische Anwendung
sowie die subkonjunktivale oder retrobulbäre Injektion (CLERC und KRÄHENMANN
1990). Die am häufigsten angewendete lokale Anwendung in der Augenheilkunde ist
die topische Applikation von Augentropfen und Augensalben (COX et al 1972).
Die physiologischen und anatomischen Besonderheiten des Auges müssen bei der
medikamentellen Therapie bedacht werden, um einen ausreichend hohen
Arzneimittelspiegel in den Kompartimenten des Auges zu bekommen. Die inneren
Strukturen des Auges zählen zu den am besten geschützten Regionen des Körpers.
Es ist bekannt, dass die auf Untersuchungen systemisch applizierter Wirkstoffe
beruhende klassische Pharmakokinetik sich nicht direkt auf ophthalmologisch
angewandte Wirkstoffe übertragen lässt (SCHOENWALD 1993). Die Wirkstoffe
topischer Arzneiformen zur Anwendung am Auge sollen nach ihrer Liberation
entweder direkt an der Konjunktiva angreifen oder sollten durch die Cornea oder

Literaturübersicht
50
Konjunktiva penetrieren, um dann in der vorderen oder hinteren Augenkammer und
an den Rezeptoren des mittleren und hinteren Auges ihre Wirkung zu bringen.
Zur topischen Anwendungin stehen wässrige Lösungen, Suspensionen, Emulsionen
sowie Salben zur Verfügung. Die Auswahl der Formulierung hat dabei besonders
durch die unterschiedliche Viskosität Einfluss auf die Verweildauer des
Medikamentes am Auge (SCHOENWALD 1985).
Glucocorticoide penetrieren nach lokaler Applikation gut in die vorderen
Augenabschnitte und erreichen die Kammerflüssigkeit. Sie zeichnen sich somit durch
eine gute Hornhautpenetration aus und sind in der Lage Gefäßeinsprossung,
Zellinfiltration und Ödem zu reduzieren (MATTHEWS et al. 1983). (COX et al. 1972)
konnte allerdings zeigen, dass Dexamethasonacetat aus einer Salbe trotz der
verlängerten Kontaktzeit weniger gut in die Kornea übertritt als aus einer Lösung.
Den Grund dafür liegt in der großen Affinität von Dexamethasonacetat für den
Salbenträger, der nur wenig Wirkstoff in den präkornealen Tränenfilm übertreten ließ
(COX et al. 1972).
Ein Nachteil wässriger Augentropfen ist deren rascher Abtransport über den Tränen-
Nasenkanal. Weiterhin kommt es durch den Reiz bei der Applikation wässriger
Augentropfen häufig zur vermehrten Tränenproduktion und damit zur Steigerung der
Drainagerate. Auf Grund dessen sind weniger als 10% des ursprünglich applizierten
Arzneistoffs in der Lage, das Hornhautepithel zu durchdringen und einen
therapeutischen Effekt im Auge zu entfalten. Bis zu 90% der Arzneistoffmenge
können durch die conjunktivalen und nasalen Blutgefäße zur systemischen
Zirkulation gelangen (KEIPERT 1997) und zu unerwünschten Effekten führen.
LEOPOLD und MAYLATH (1952), die topische, subkonjunktivale und systemische
Applikation von Cortisonacetat beim Menschen verglichen, haben festgestellt, dass
Erkrankungen der vorderen Segmente des Auges mit gleichem Erfolg lokal,
subkonjunktival oder systemisch behandelt werden können. Eine Behandlung der

Literaturübersicht
51
tieferen Strukturen ist nach ihrer Meinung lediglich durch eine systemische Therapie
sowie durch subkonjunktivale Injektion möglich.
Arbeiten von REICHENBECKER (2002) und KAISER (2003) beschäftigen sich mit
Untersuchungen zur Verteilung von Dexamethason im Auge des Pferdes und im
Hundeauge. KAISER (2003) konnte feststellen, dass sich Dexamethason bereits
nach einer einmaligen lokalen Behandlung des Hundeauges mit einer
Dexamethason-Augensalbe in der Cornea, Iris und im dritten Augenlid, sowie in
geringerem Ausmaß auch in der Retina / Choroidea nachweisen lässt. Für die Katze
lassen sich aus der zurzeit vorhandenen Literatur keine Daten für die Verteilung
pharmakologischer Substanzen in den Kompartimenten des Auges finden. Die
vorliegende Arbeit beschäftigt sich daher mit dieser bisher nur wenig behandelten
Fragestellung.

Material und Methode
52
3 MATERIAL UND METHODE In der vorliegenden Arbeit sollte das Verteilungsverhalten von Dexamethason (DXM)
nach einmaliger lokaler Anwendung am Katzenauge untersucht werden. 20 Katzen
wurden dazu jeweils 3 und 6 Stunden vor Euthanasie und Probengewinnung mit
einer Dexamethason-Augensalbe oder Dexamethason-Augentropfsuspension
behandelt. Die benötigten Materialien und Geräte werden im folgenden Abschnitt,
ebenso wie Versuchsaufbau und Messmethoden aufgelistet.
3.1 Geräte Dispensette II® (Brand, Deutschland)
Kühlzentrifuge, Typ 5403 Eppendorf
Magnetrührer mit Heizplatte (Heidolf MR 3001K)
pH-Meter, Typ pH 320 (WTW, Weilheim)
Pipetten: einstellbar von 10-100 µl, 100-1000 µl, 500-2500 µl (Eppendorf, Hamburg)
Präparierbesteck (VWR International GmbH, Deutschland)
Rüttler, Typ Reax Top (Heidolph, Deutschland)
Überkopfschwenker,Typ PFAX 2 (Heidolph, Deutschland)
Ultra Turrax TP 18/10 (IKA-Werk, Deutschland)
Wärmeschrank (Memmert, Schwarbach)
β- Counter: LS-5000-TA (Beckmann, Deutschland)
Vibrax VXR (IKA Werk, Deutschland)
TCS- Metallblock- Thermostat (Labor Technik, Barkey)
3.2 Verbrauchsmaterialien Einmalkanülen 23Gx1“ (Terumo Europe N.V., 3001 Leuven, Belgium)
Pasteurpipetten, 150 mm Glas, Nr. 747715 (Brand GmbH&Co., Wertheim)
Pipettenspitzen: blau, Nr. 686290 (Greiner BIO-ONE GmbH, Frickenhausen)
Pipettenspitzen: gelb, Nr. 685290 (Greiner BIO-ONE GmbH, Frickenhausen)

Material und Methode
53
Pipettenspitzen: weis, Nr. 0030000.978 (Eppendorf, Hamburg)
PP-Reaktionsgefäße 1,5 ml (Greiner, Frickenhausen)
Spritzen 2,0 ml (BRAUN Melsungen AG)
3.3 Medikament Dexamethasonhaltige Augentropfsuspension Isopto-Dex® und Augensalbe Isopto-
Dex® (Alkon-Pharma GmbH, Freiburg) enthalten jeweils 1 mg Dexamethason / ml
Suspension oder / g Salbe.
Folgende arzneilich nicht wirksame Bestandteile sind in der Augentropfsuspension
enthalten:
• Benzalkoniumchlorid
• Citronensäure und/oder Natriumhydroxid zur pH-Einstellung
• Dinatriumedetat 2H2O
• gereinigtes Wasser
• Hypromellose
• Natriumchlorid
• Natriummonohydrogenphosphat wasserfrei
• Polysorbat 80
Folgende arzneilich nicht wirksame Bestandteile sind in der Augensalbe enthalten:
• flüssiges Wollwachs Wasser-frei
• Methyl-4-Hydroxybenzoat (E218)
• Propyl-4-Hydroxybenzoat (E216)
• weiße Vaseline
Dosierungsanleitung:
Isopto-Dex Augentropfsuspension: 1 Tropfen zwei- bis sechsmal täglich einträufeln.
Isopto-Dex Augensalbe sollte drei- bis viermal pro Tag in den unteren Bindehautsack
des Auges gestrichen werden.

Material und Methode
54
3.4 Chemikalien und Reagenzien Aquasafe 300 Plus Szintillator (Zinsser Analytic, U.K.)
[1, 2, 3, 4-3H] Dexamethason Lösung in Ethanol, (Amersham Pharmacia Biotech,
U.K.)
Spezifische Aktivität: 1,44 TBq/mmol; 39,0 Ci/mmol
Aktivkohle zur Analyse (E. Merck, Darmstadt)
Dexamethason (Sigma, München)
Dextran T 70 (Carl Roth GmbH & Co., Karlsruhe)
Di-Natriumhydrogencarbonat-Dihydrat (E. Merck, Darmstadt)
Ethanol (absolut) (Riedel-de Haen, Seetze)
Ethylacetat (LAB-SCAN, Dublin)
Gelatine (SIGMA, Steinheim)
Natriumchlorid (NaCl) (E. Merck, Darmstadt)
Salzsäure (HCl) (E. Merck, Darmstadt)
Stickstoff (Westfalen AG, Göttingen)
3.5 Patientengut Die im Rahmen dieser Arbeit untersuchten Proben stammen von 20 Katzen. Alle
Katzen wurden in 4 Gruppen aufgeteilt und einmal lokal mit der Augensalbe oder
Augentropfsuspension behandelt. Es handelt sich bei diesen Katzen um Patienten
der Kleintierklinik der Tierärztlichen Hochschule Hannover, die in der Zeit von
Oktober 2004 bis Juni 2006 behandelt und wegen anderer Erkrankungen
euthanasiert wurden. Die Patienten der 1 Gruppe und 2 Gruppe wurden 6 Stunden,
die Patienten der 3 Gruppe und 4 Gruppe 3 Stunden, nach der Salben- und
Tropfenapplikation euthanasiert (Tab. 4). Klinische Befunde sind den Tabellen 3.1 bis
3.4 zu entnehmen.
Material und Methode
55
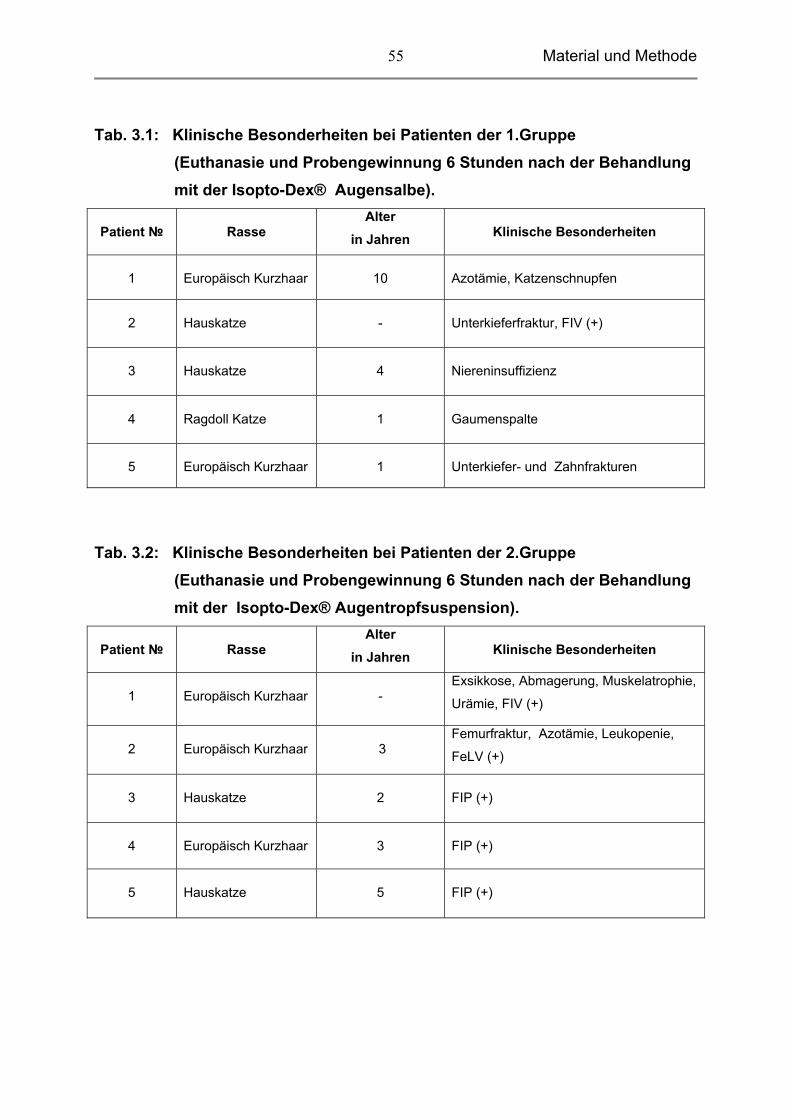
Tab. 3.1: Klinische Besonderheiten bei Patienten der 1.Gruppe (Euthanasie und Probengewinnung 6 Stunden nach der Behandlung mit der Isopto-Dex® Augensalbe).
Patient №
Rasse
Alter in Jahren
Klinische Besonderheiten
1
Europäisch Kurzhaar
10
Azotämie, Katzenschnupfen
2
Hauskatze
-
Unterkieferfraktur, FIV (+)
3
Hauskatze
4
Niereninsuffizienz
4
Ragdoll Katze
1
Gaumenspalte
5
Europäisch Kurzhaar
1
Unterkiefer- und Zahnfrakturen
Tab. 3.2: Klinische Besonderheiten bei Patienten der 2.Gruppe (Euthanasie und Probengewinnung 6 Stunden nach der Behandlung mit der Isopto-Dex® Augentropfsuspension).
Patient №
Rasse
Alter in Jahren
Klinische Besonderheiten
1
Europäisch Kurzhaar
-
Exsikkose, Abmagerung, Muskelatrophie,
Urämie, FIV (+)
2
Europäisch Kurzhaar
3
Femurfraktur, Azotämie, Leukopenie,
FeLV (+)
3
Hauskatze
2
FIP (+)
4
Europäisch Kurzhaar
3
FIP (+)
5
Hauskatze
5
FIP (+)
Material und Methode
56
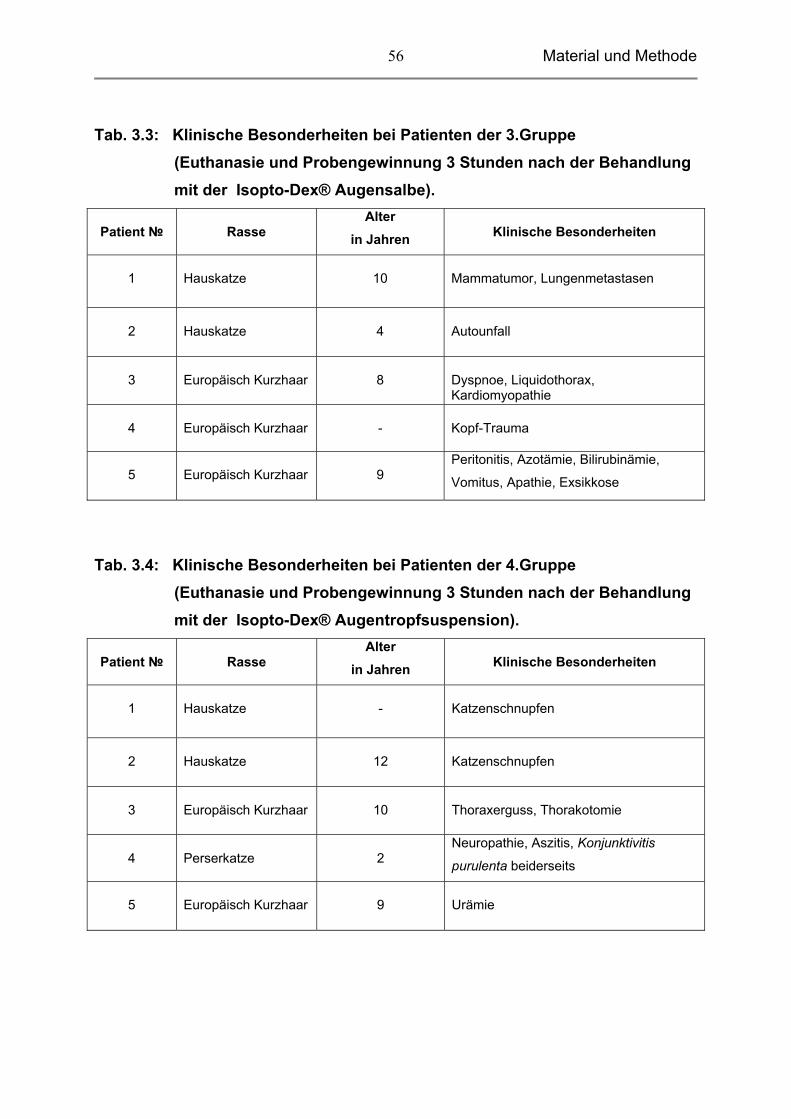
Tab. 3.3: Klinische Besonderheiten bei Patienten der 3.Gruppe (Euthanasie und Probengewinnung 3 Stunden nach der Behandlung mit der Isopto-Dex® Augensalbe).
Patient №
Rasse
Alter in Jahren
Klinische Besonderheiten
1
Hauskatze
10
Mammatumor, Lungenmetastasen
2
Hauskatze
4
Autounfall
3
Europäisch Kurzhaar
8
Dyspnoe, Liquidothorax, Kardiomyopathie
4
Europäisch Kurzhaar
-
Kopf-Trauma
5
Europäisch Kurzhaar
9
Peritonitis, Azotämie, Bilirubinämie,
Vomitus, Apathie, Exsikkose
Tab. 3.4: Klinische Besonderheiten bei Patienten der 4.Gruppe (Euthanasie und Probengewinnung 3 Stunden nach der Behandlung mit der Isopto-Dex® Augentropfsuspension).
Patient №
Rasse
Alter in Jahren
Klinische Besonderheiten
1
Hauskatze
-
Katzenschnupfen
2
Hauskatze
12
Katzenschnupfen
3
Europäisch Kurzhaar
10
Thoraxerguss, Thorakotomie
4
Perserkatze
2
Neuropathie, Aszitis, Konjunktivitis
purulenta beiderseits
5
Europäisch Kurzhaar
9
Urämie

Material und Methode
57
Tab. 3.5: Bechandlungsschema
Gruppe
Katze
Behandlung mit
Probengewinnung nach Bechandlung
1
1 - 5
Isopto-Dex® Augensalbe
6 Stunden
2
6 - 10
Isopto-Dex®
Augentropfsuspension
6 Stunden
3
11 - 15
Isopto-Dex® Augensalbe
3 Stunden
4
16 - 20
Isopto-Dex®
Augentropfsuspension
3 Stunden
3.5.1 Klinische Untersuchung Die Katzen dieser Studie wurden aufgrund verschiedener Erkrankungen
euthanasiert. Unmittelbar vor dem Eintritt eines Patienten in die Studie erfolgte eine
Untersuchung. Hierbei wurden neben der Körpertemperatur, die Atmung, der
Zustand der Schleimhäute und die Mandibularlymphknoten untersucht. Die
Auskultation des Herzens und der Lunge schloss die klinische Untersuchung ab. Ein
Großteil der Patienten wurde vor dem Eintritt in die Studie über einen unterschiedlich
langen Zeitraum bis zur Euthanasie mit verschiedenen Arzneimitteln therapiert. Es
gelangten keine Patienten in die Studie, die mit Dexamethason vorbehandelt waren.
3.5.2 Spezielle ophthalmologische Untersuchung
Vor Auswahl eines Patienten erfolgte eine spezielle ophthalmologische
Untersuchung zur Feststellung des Status praesens der Augengesundheit des

Material und Methode
58
Tieres. Die Untersuchung erfolgte in einem abgedunkelten Raum am nicht
narkotisierten Tier. Es wurde die Umgebung des Auges kontrolliert, dabei wurde auf
Entzündungserscheinungen, die Lidstellung, Spuren eines übermäßigen oder
verminderten Tränenflusses sowie Veränderungen des 3. Augenlides (soweit ohne
Anästhesie möglich) geachtet.
Die Untersuchung wurde mit einem Ophthalmoskop bei jedem zur Untersuchung
anstehenden Tier vorgenommen. Sie erfolgte im darauffallenden, seitlichen und im
durchfallenden Licht. Auf diese Weise wurde Cornea, vordere Augenkammer, Linse,
hintere Augenkammer und der vordere Glaskörper untersucht. Ein Patient der 4.
Gruppe (3 Stunden nach der Tropfenapplikation) wies eine Konjunktivitis purulenta
beiderseits auf. Alle weiteren Patienten waren unauffällig. 3.6 Probengewinnung und Lagerung Die Entnahme und Präparation der Augen erfolgte in unmittelbarem Anschluß an die
Euthanasie der Patienten. Zur Etablierung der Entnahmetechnik der einzelnen
Proben wurde die Entnahmetechnik vorab in der Tierärztlichen Klinik Dr. S. Kaiser /
Dr. H. Lindenstruth (Werl / Westfalen) und im Institut für Pathologie der Tierärztlichen
Hochschule Hannover erprobt. Dabei wurden die Proben von 3. Augenlid, Kornea,
Kammerwasser, Iris, Linse, Glaskörper und Retina entnommen. Die Aufbewahrung
erfolgte bei -20°C.
3.7 Versuchsaufbau Die in den Tabellen 3.1 bis 3.4 aufgeführten Tiere erhielten einmalig eine Gabe von
0,05 ml eines dexamethasonhaltigen Medikament Isopto® Dex je nach der
Patientengruppe in Form einer Augensalbe oder Augentropfsuspension in den
Konjunktivalsack eingegeben. Die Menge entspricht 0,05 mg Dexamethason. Die
Medikamentgabe erfolgte am wachen Tier unter Praxisbedingungen. 6 Stunden

Material und Methode
59
(Gruppe 1 und 2) und 3 Stunden (Gruppe 3 und 4) nach der Applikation wurden die
Tiere euthanasiert.
3.7.1 Enukleation der Bulbi oculi
Die Enukleation beider Bulbi erfolgte unmittelbar im Anschluß an die Euthanasie des
Patienten. Der Augapfel wird möglichst weit freigelegt und gleich eine Kanthotomie
vorgenommen, um die Lidspalte zu erweitern. Die Durchtrennung der Bindehaut
erfolgt zirkulär dem Limbus entlang, wobei die Tenonsche Kapsel ebenfalls losgelöst
wird (1). Nun werden die Augenmuskeln sichtbar. Die 4 geraden und die 2 schrägen
Muskeln werden in der nähe ihres Sehnenansatzes an der Lederhaut abgesetzt (2).
Es verbleibt indessen noch ein Muskelkranz rund um die Hülle des Sehnervs. Die
Muskeln werden durchschnitten, so dass nur noch der Nervenstrang den Bulbus in
der Orbitalhöhle zurückhält (3). Zur Vermeidung einer Blutung wird der gesamte
Nervenstrang mit einer gebogenen und stumpfen Klemme erfasst und oberhalb
dieser durchtrennt (4). Die Nickhaut samt Nickhautdrüse wird entfernt.

Material und Methode
60
Abb. 3.1 Entfernung des Augapfels (CLERC und KRÄHENMANN 1990).
3.7.2 Entnahme des Kammerwassers Mittels einer Kanüle (23G x 1“) und einer 2 ml Spritze wurde im ersten Sektionsschritt
Kammerwasser entnommen. Dazu wurde im medialen Augenwinkel im
Übergangsbereich von Sclera und Kornea in flachem Winkel in die vordere
Augenkammer eingegangen und zwischen 0,7 und 1,5 ml Kammerwasser
gewonnen. Das so gewonnene Kammerwasser wurde dann 5 Minuten bei 3000 g
zentrifugiert und der zellfreie Überstand bis zur Probenaufarbeitung gelagert.

Material und Methode
61
3.7.3 Entnahme von 3. Augenlid, Cornea, Iris und Linse Nach Entnahme des Kammerwassers wurde das 3. Augenlid mit einer Augenpinzette
und einer spitzen Schere in toto inklusive der Nickhautdrüse vorgelagert und
entnommen. Durch die Inzision im Bereich der Einstichstelle für die
Kammerwasserentnahme wurde anschließend die vordere Augenkammer eröffnet.
Die Cornea wurde nun zirkulär am Limbus abpräpariert. In gleicher Weise wurde die
Iris entnommen. Die so zugängliche Linse wurde vorsichtig aus ihrem Halteapparat
gelöst.
3.7.4 Entnahme von Glaskörperanteilen und der Retina (Choroidea) Durch den nach der Entnahme der Linse entstandenen Zugang zur hinteren
Augenkammer wurden mittels einer Spritze Anteile des Glaskörpers aufgenommen.
Die Menge lag bei 1,2 bis 2,2 ml. Nach Zentrifugation (5 Minuten bei 3000 g) wurde
etwa 1 ml zellfreier Überstand abpipettiert und gemäß Angaben unter bei –20 °C
aufbewahrt. Mit einer feinen Pinzette konnte nun die Retina in Verbund mit der
Choroidea vom Ziliarkörper bis zum Sehnervenkopf, welcher mit einer feinen Schere
umschnitten wurde, in toto gelöst werden.
3.8 Probenaufarbeitung Die Einwaage von Kornea, Linse sowie Retina / Choroidea erfolgte in gefrorenem
Zustand. Die durchschnittlichen Mengen der eingewogenen Proben betrugen: 0,21 g
für die Cornea, 0,61 g für das 3. Augenlid, 0,07 g für die Iris, 0,78 g für die Linse
sowie 0,47 g für Retina / Choroidea. Nach Zerkleinerung der Proben mittels eines
feinen Präparierbestecks wurden alle festen Proben (ausschließlich Kammerwasser
und Glaskörper) in einem Reaktionsgefäß mit 1 ml GPP versetzt und homogenisiert
(Ultra-Turrax, 8.000 U/Minute). Kammerwasser und Glaskörper wurden bei

Material und Methode
62
Zimmertemperatur aufgetaut und Aliquote von 500 µl in ein Reaktionsgefäß pipettiert.
Alle Proben wurden mit 2 ml Ethylacetat versetzt und 12 Minuten geschüttelt
(Überkopfschwenker). Danach erfolgte eine Zentrifugation (3.000 g, 10 Minuten). Die
Überstände wurden unter Stickstoff eingedampft. Die Lagerung der eingedampften
Proben erfolgte bei –80 °C.
Vor der radioimmunologischen Messung von DXM wurden die Proben bei
Raumtemperatur aufgetaut und anschließend in 210 µl GPP gelöst. Hierzu wurden
alle Proben fünf Minuten in das Ultraschallbad verbracht und unmittelbar vor der
Messung geschüttelt. Für die nachfolgende Messung wurden 200 µl der so gelösten
Probe verwendet.
3.9 Radioimunassay (RIA) 3.9.1 Prinzip Der RIA beruht auf einer Antigen-Antikörperreaktion (BERSON und YALOW 1964).
Der zu bestimmenden Substanz, dem Antigen Dexamethason (Ag), stehen eine
bekannte Menge Antikörper (Ak) sowie eine ebenfalls bekannte Menge des
markierten Antigens 3H-Dexamethason (Ag*) gegenüber.
Die Antigen-Antikörperreaktion läuft bei der RIA in einem geeigneten
Reaktionsmedium ab, dem unmarkiertes und markiertes Antigen zugesetzt sind.
Dabei sollte die Menge des markierten Antigens in etwa der zu bestimmenden
Menge Antigen entsprechen. Im anschließenden Schritt wird soviel Antiserum
hinzugegeben, dass etwa die Hälfte des markierten Antigens gebunden wird.

Material und Methode
63
In einer definierten Inkubationszeit stellt sich ein Gleichgewicht zwischen den
Komplexen A und B ein:
Komplex A: Komplex B:
Ag + Ak ↔ [ Ag Ak] Ag* + Ak ↔ [ Ag* Ak]
Ag + Ag* + Ak ↔ [ Ag Ak] + [ Ag* Ak]
Ag = unmarkiertes Antigen (zu bestimmendes Dexamethason in der Probe)
Ag* = markiertes Antigen (3H-Dexamethason)
Ak = Antikörper
Im Meßansatz wird nach der Inkubationszeit freies Antigen durch die Zugabe der
Dextran-Aktivkohle-Suspension gebunden und abgetrennt. Das freie Antigen wird
von der Aktivkohle adsorbiert und anschließend abzentrifugiert. Der großmolekulare
Antigen-Antikörper-Komplex verbleibt im Überstand. Die gemessene Radioaktivität
entspricht der Menge der verbliebenen Antigen*-Antikörper-Komplexe und damit
auch der Menge des zu bestimmenden Antigens. Je mehr Antigen also in der
untersuchten Probe vorliegt, desto weniger radioaktives Antigen (3H-Dexamethason)
wird vom Antikörper gebunden.
Für die Konzentrationsbestimmung in unbekannten Proben wird eine
Kalibrationskurve erstellt, anhand welcher die Konzentration der Proben abgelesen
werden kann. Zu diesem Zweck wird dem Inkubationsmedium unmarkiertes Antigen
in bekannten Konzentrationen zugesetzt. Dadurch wird das Gleichgewicht in
Abhängigkeit zur Konzentration zu Gunsten des nichtmarkierten Komplex A
verschoben. Über die Messung der Gesamtradioaktivität in den jeweiligen
Versuchsansätzen kann somit auf die Konzentration von unmarkiertem Antigen
rückgeschlossen werden.
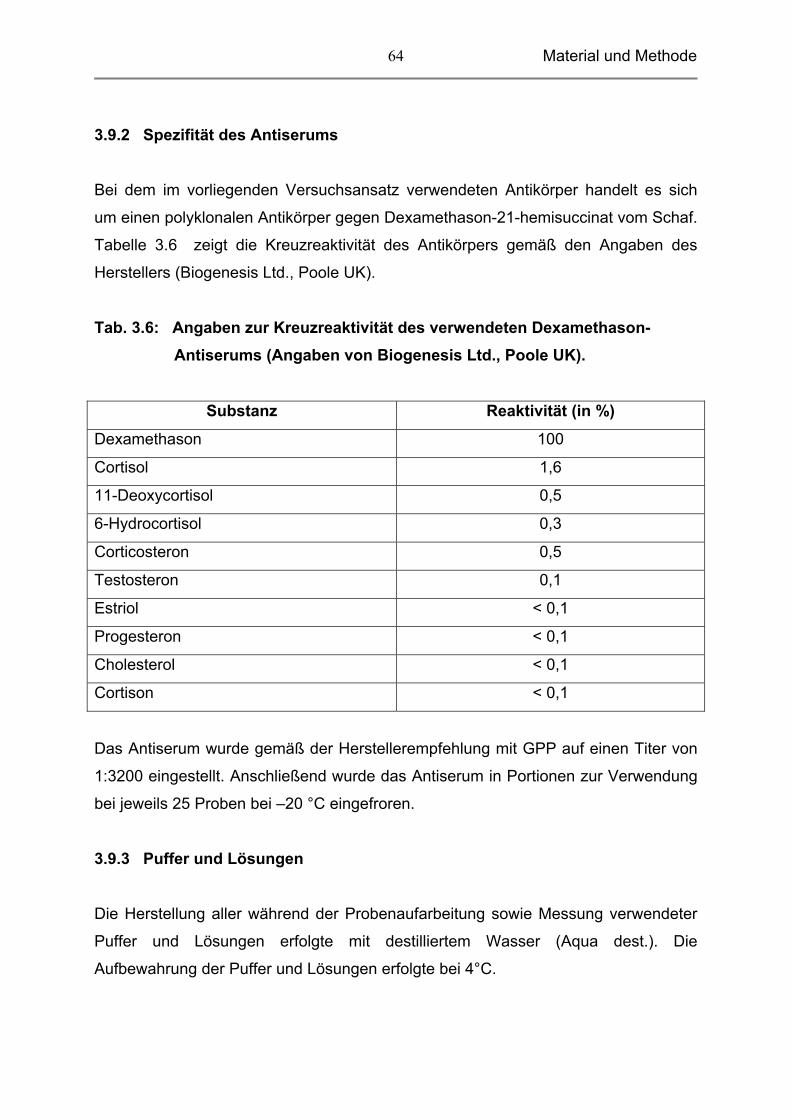
Material und Methode
64
3.9.2 Spezifität des Antiserums Bei dem im vorliegenden Versuchsansatz verwendeten Antikörper handelt es sich
um einen polyklonalen Antikörper gegen Dexamethason-21-hemisuccinat vom Schaf.
Tabelle 3.6 zeigt die Kreuzreaktivität des Antikörpers gemäß den Angaben des
Herstellers (Biogenesis Ltd., Poole UK).
Tab. 3.6: Angaben zur Kreuzreaktivität des verwendeten Dexamethason- Antiserums (Angaben von Biogenesis Ltd., Poole UK).
Substanz Reaktivität (in %)
Dexamethason 100
Cortisol 1,6
11-Deoxycortisol 0,5
6-Hydrocortisol 0,3
Corticosteron 0,5
Testosteron 0,1
Estriol < 0,1
Progesteron < 0,1
Cholesterol < 0,1
Cortison < 0,1
Das Antiserum wurde gemäß der Herstellerempfehlung mit GPP auf einen Titer von
1:3200 eingestellt. Anschließend wurde das Antiserum in Portionen zur Verwendung
bei jeweils 25 Proben bei –20 °C eingefroren.
3.9.3 Puffer und Lösungen Die Herstellung aller während der Probenaufarbeitung sowie Messung verwendeter
Puffer und Lösungen erfolgte mit destilliertem Wasser (Aqua dest.). Die
Aufbewahrung der Puffer und Lösungen erfolgte bei 4°C.

Material und Methode
65
3.9.3.1 Gelatine-Phosphat-Puffer (GPP) Ein Liter einer 0,01 molaren Di-Natriumhydrogenphosphat-Lösung wird unter Zugabe
von 0,01 molarer Natriumdihydrogenphosphat-Lösung auf einen pH-Wert von 7,8
eingestellt. In einem Liter dieser Lösung werden 8,77 g NaCl gelöst. Es erfolgt eine
Korrektur des pH-Wertes der Lösung auf 7,6 unter Zugabe von Natriumhydroxid oder
Salzsäure. 200 mg Gelatine werden abschließend in 200 ml dieser Pufferlösung
gelöst.
3.9.3.2 Dextran-Aktivkohle-Suspension In 100 ml Gelatine-Phoshat-Puffer werden 500 mg Aktivkohle und 50 mg Dextran 70
gelöst.
3.9.3.3 Dexamethason-Standardreihe Zur Herstellung einer Stammlösung werden 10 mg Dexamethason in 10 ml Ethanol
gelöst. Diese Lösung wird soweit mit GPP verdünnt, dass am Ende Lösungen mit
16.000, 8.000, 4.000, 2.000, 1.000, 500, 250, 125 pg DXM/200 µl vorliegen.
3.9.3.4 3H-Dexamethason-Lösung Die 3H-Dexamethason-Lösung wurde mit Ethanol auf 7.500 - 8.000 CPM / 100 µl
eingestellt.

Material und Methode
66
3.10 Untersuchung der Proben In Reaktionsgefäße (1,5 ml) wurden 100 µl 3H-Dexamethason-Ethanol-Lösung
gegeben und unter Stickstoff eingedampft. Nun wurden zu jedem Ansatz 200 µl der
zuvor in 210 µl GPP gelösten Probe sowie 100 µl der Antikörperlösung gegeben. Es
folgte eine Inkubation über eine Stunde bei 30 °C. Unmittelbar anschließend wurden
alle Proben 15 Minuten bei 4°C gekühlt. Nach der Zugabe von 500 µl Dextran-
Aktivkohle-Suspension wurden alle Probenansätze weitere 17 Minuten bei 4 °C
inkubiert. Nach zehnminütiger Zentrifugation (3000 g, 4 °C) wurden die Überstände
abpipettiert und in 5 ml Szintillator (Aquasafe 300 Plus) gegeben. Die Messung der
enthaltenen Radioaktivität erfolgte mittels β-Counter. Tabelle 3.7 fasst die Methode
zusammen.
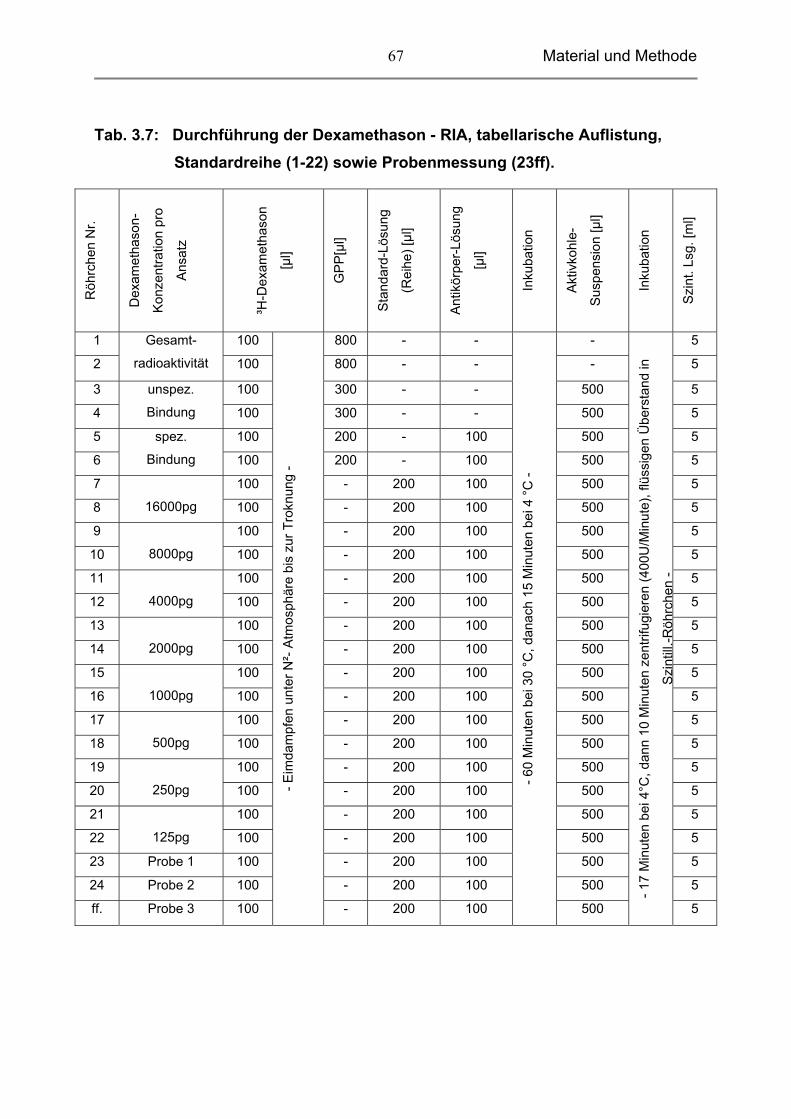
Material und Methode
67
Tab. 3.7: Durchführung der Dexamethason - RIA, tabellarische Auflistung, Standardreihe (1-22) sowie Probenmessung (23ff).
Röh
rche
n N
r.
Dex
amet
haso
n-
Kon
zent
ratio
n pr
o
Ansa
tz
³H-D
exam
etha
son
[µl]
GP
P[µ
l]
Sta
ndar
d-Lö
sung
(Rei
he) [µl
]
Ant
ikör
per-
Lösu
ng
[µl]
Inku
batio
n
Akt
ivko
hle-
Susp
ensi
on [µ
l]
Inku
batio
n
Szi
nt. L
sg. [
ml]
1 100 800 - - - 5
2
Gesamt-
radioaktivität 100 800 - - - 5
3 100 300 - - 500 5
4
unspez.
Bindung 100 300 - - 500 5
5 100 200 - 100 500 5
6
spez.
Bindung 100 200 - 100 500 5
7 100 - 200 100 500 5
8
16000pg 100 - 200 100 500 5
9 100 - 200 100 500 5
10
8000pg 100 - 200 100 500 5
11 100 - 200 100 500 5
12
4000pg 100 - 200 100 500 5
13 100 - 200 100 500 5
14
2000pg 100 - 200 100 500 5
15 100 - 200 100 500 5
16
1000pg 100 - 200 100 500 5
17 100 - 200 100 500 5
18
500pg 100 - 200 100 500 5
19 100 - 200 100 500 5
20
250pg 100 - 200 100 500 5
21 100 - 200 100 500 5
22
125pg 100 - 200 100 500 5
23 Probe 1 100 - 200 100 500 5
24 Probe 2 100 - 200 100 500 5
ff. Probe 3 100
- Eim
dam
pfen
unt
er N
²- A
tmos
phär
e bi
s zu
r Tro
knun
g -
- 200 100
- 60
Min
uten
bei
30
°C, d
anac
h 15
Min
uten
bei
4 °
C -
500
- 17
Min
uten
bei
4°C
, dan
n 10
Min
uten
zen
trifu
gier
en (4
00U
/Min
ute)
, flü
ssig
en Ü
bers
tand
in
Szi
ntill.
-Röh
rche
n -
5

Material und Methode
68
3.11 Kalibrationskurve und Berechnung
Bei der Messung liefert der β-Counter ein Ergebnis in CPM / Probe. Es werden bei
jeder
Messung erfaßt:
• Gesamtradioaktivität (Total count TC)
• Unspezifische Bindung (non-specific binding NSB)
• Spezifische Bindung (maximum specific binding MSB)
• Blanc (Leerwert)
• Dexamethason DXM-Standardreihe
Zur Berechnung der Dexamethasonkonzentration der einzelnen Proben wird die in
Prozent errechnete Bindungsrate (in Bezug auf die Gesamtradioaktivität TC) nach
Probit-Transformation mit Hilfe einer mit linearer Regression errechneten Funktion in
die Dexamethasonkonzentration je Ansatz umgerechnet. Anschließend erfolgt die
Korrektur der Werte mit Hilfe der für die jeweilige Probenmatrix ermittelten
Widerfindungsrate. Die endgültige Umrechnung in ng DXM / g Probe bzw. ng DXM /
ml Probe erfolgt unter Berücksichtigung der Probeneinwaage und aller
Verdünnungsstufen.
3.12 Statistische Auswertung Die statistische Auswertung der Ergebnisse erfolgte mit Hilfe des Programms
GraphPad Prism 4.02®. Für die statistische Auswertung wurden die mit der
Wiederfindungsrate korrigierten und die in DXM/g Probe bzw. DXM/ml Probe
umgerechneten Werte verwendet. Der statistische Vergleich der Gruppen erfolgte mit
dem Mann-Whitney-Test für unabhängige, nicht normalverteilte Proben. Für alle

Material und Methode
69
statistischen Verfahren wurden die in Tabelle 3.8 dargestellten Signifikanzstufen in
Abhängigkeit von der Irrtumswahrscheinlichkeit p ≤ 0,05 festgelegt.
Tab. 3.8: Signifikanzstufen in Abhängigkeit von der Irrtumswahrscheinlichkeit p ≤ 0,05
p- Wert
Signifikanzstufe
Symbol
≤ 0,001
hoch signifikant
***
≤ 0,01
signifikant
**
≤ 0,05
schwach signifikant
*
≥ 0,05
nicht signifikant
3.13 Wiederholbarkeit und Nachweisgrenzen Tabelle 3.9 verdeutlicht die gute Wiederholbarkeit mit dem durchgeführten
radioimmunologischen Verfahren.

Material und Methode
70
Tab. 3.9: Angaben zur Wiederholbarkeit (Total count = 100 %).
Bindungsrate in Relation zur Gesamtradioaktivität (TC) in Prozent
125 ng
DXM/200 ml
250 ng
DXM/200 ml
2000 ng
DXM/200 ml
Mittelwert
33,5
24,4
9,25
Standardabweichung
1,4
0,8
0,6
Für Kornea, Iris, 3. Augenlid, Linse und Retina (Tab. 3.9), sowie Kammerwasser und
Glaskörperflüssigkeit ergeben sich unter Berücksichtigung der entsprechenden
Wiederfindungsraten die in der Tabelle 3.10 aufgeführten Nachweisgrenzen. Die
Wiederfindung liegt für die Kornea bei im Mittel 79 %, für das Kammerwasser bei 90
%, für die Iris bei 83 %, für die Linse bei 76 %, für den Glaskörper bei 90 % sowie für
Retina bei 81 %.
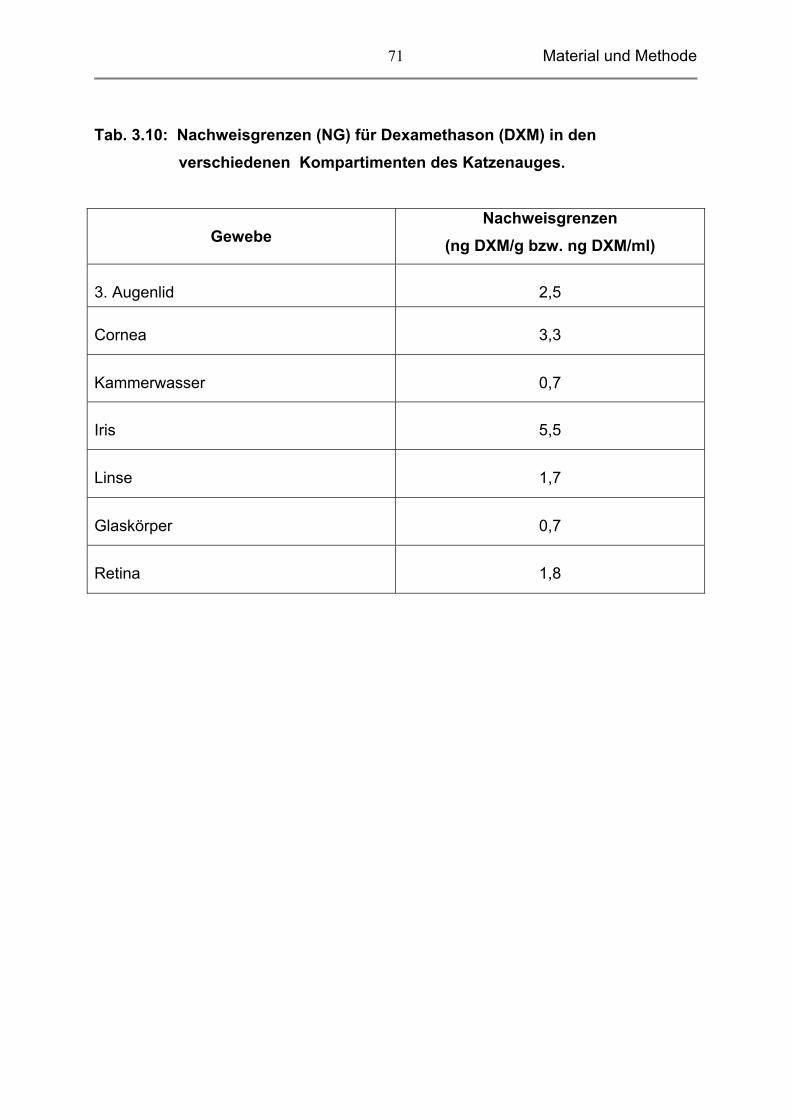
Material und Methode
71
Tab. 3.10: Nachweisgrenzen (NG) für Dexamethason (DXM) in den verschiedenen Kompartimenten des Katzenauges.
Gewebe
Nachweisgrenzen (ng DXM/g bzw. ng DXM/ml)
3. Augenlid
2,5
Cornea
3,3
Kammerwasser
0,7
Iris
5,5
Linse
1,7
Glaskörper
0,7
Retina
1,8

Ergebnisse
72
4 ERGEBNISSE 4.1 Dexamethasonkonzentrationen in den einzelnen Kompartimenten des
Katzenauges Die Tabellen 4.1 bis 4.4 enthalten Angaben der Einzelwerte sowie von Mittelwert,
Standardabweichung und Median der ermittelten Dexamethasonkonzentrationen in
den einzelnen Augenkompartimenten. Die Abbildungen 4.1 bis 4.7 zeigen die
Ergebnisse in Form von Median-Boxen.
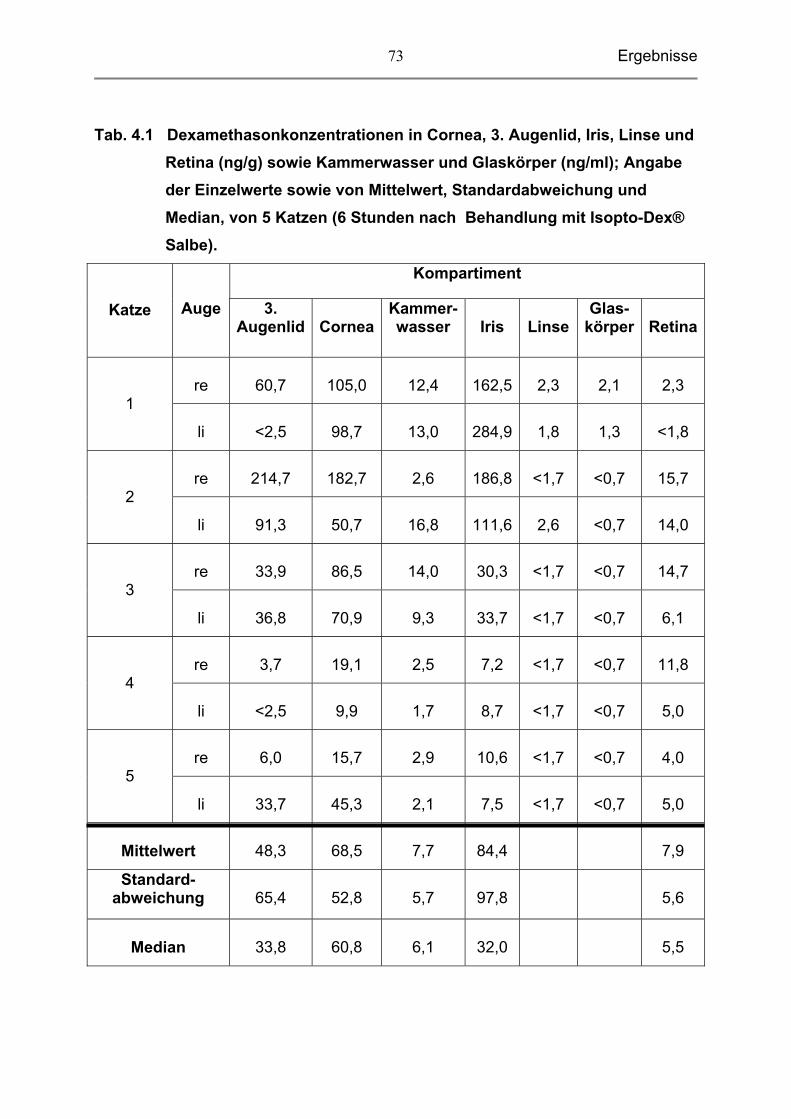
Ergebnisse
73
Tab. 4.1 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml); Angabe der Einzelwerte sowie von Mittelwert, Standardabweichung und Median, von 5 Katzen (6 Stunden nach Behandlung mit Isopto-Dex® Salbe).
Kompartiment
Katze Auge 3. Augenlid
Cornea
Kammer-wasser
Iris
Linse
Glas-körper
Retina
re
60,7
105,0
12,4
162,5
2,3
2,1
2,3
1 li
<2,5
98,7
13,0
284,9
1,8
1,3
<1,8
re
214,7
182,7
2,6
186,8
<1,7
<0,7
15,7
2 li
91,3
50,7
16,8
111,6
2,6
<0,7
14,0
re
33,9
86,5
14,0
30,3
<1,7
<0,7
14,7
3 li
36,8
70,9
9,3
33,7
<1,7
<0,7
6,1
re
3,7
19,1
2,5
7,2
<1,7
<0,7
11,8
4 li
<2,5
9,9
1,7
8,7
<1,7
<0,7
5,0
re
6,0
15,7
2,9
10,6
<1,7
<0,7
4,0
5 li
33,7
45,3
2,1
7,5
<1,7
<0,7
5,0
Mittelwert
48,3
68,5
7,7
84,4
7,9
Standard- abweichung
65,4
52,8
5,7
97,8
5,6
Median
33,8
60,8
6,1
32,0
5,5
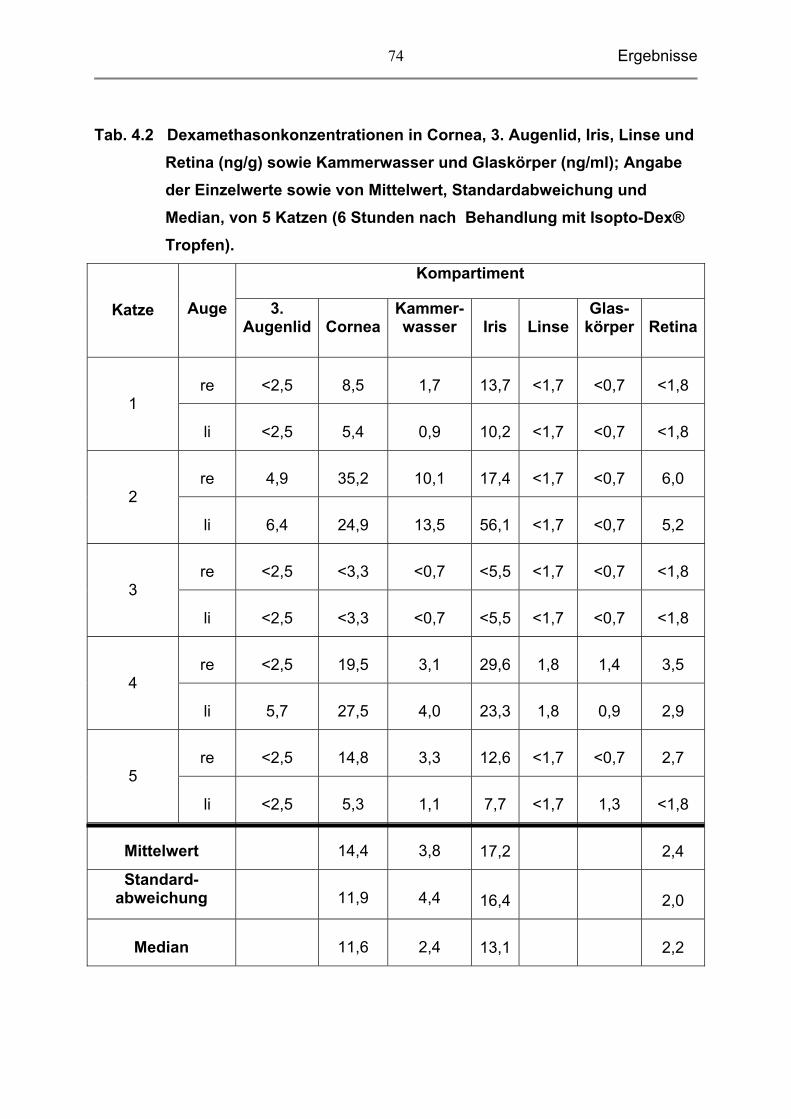
Ergebnisse
74
Tab. 4.2 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml); Angabe der Einzelwerte sowie von Mittelwert, Standardabweichung und Median, von 5 Katzen (6 Stunden nach Behandlung mit Isopto-Dex® Tropfen).
Kompartiment
Katze Auge 3. Augenlid
Cornea
Kammer-wasser
Iris
Linse
Glas-körper
Retina
re
<2,5
8,5
1,7
13,7 <1,7 <0,7 <1,8
1
li
<2,5
5,4
0,9
10,2 <1,7 <0,7 <1,8
re
4,9
35,2
10,1
17,4 <1,7 <0,7 6,0
2
li
6,4
24,9
13,5
56,1 <1,7 <0,7 5,2
re
<2,5
<3,3
<0,7
<5,5 <1,7 <0,7 <1,8
3
li
<2,5
<3,3
<0,7
<5,5 <1,7 <0,7 <1,8
re
<2,5
19,5
3,1
29,6 1,8 1,4 3,5
4
li
5,7
27,5
4,0
23,3 1,8 0,9 2,9
re
<2,5
14,8
3,3
12,6 <1,7 <0,7 2,7
5
li
<2,5
5,3
1,1 7,7 <1,7 1,3 <1,8
Mittelwert
14,4
3,8 17,2 2,4
Standard- abweichung
11,9
4,4 16,4 2,0
Median
11,6
2,4 13,1 2,2

Ergebnisse
75
Tab. 4.3 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml); Angabe der Einzelwerte sowie von Mittelwert, Standardabweichung und Median, von 5 Katzen (3 Stunden nach Behandlung mit Isopto-Dex® Salbe).
Kompartiment
Katze Auge 3. Augenlid
Cornea
Kammer-wasser
Iris
Linse
Glas-körper
Retina
re
6,1
11,6
8,3
18,1
<1,7
<0,7
2,8
1 li
7,0
20,1
4,8
26,7
<1,7
<0,7
4,1
re
5,7
17,8
4,4
19,1
<1,7
<0,7
2,5
2 li
5,2
9,3
2,8
18,8
<1,7
<0,7
2,0
re
11,9
62,9
3,7
10,9
<1,7
<0,7
6,1
3 li
54,5
27,4
0,8
<5,5
<1,7
<0,7
6,4
re
14,4
124,7
11,5
19,3
<1,7
0,9
16,3
4 li
59,9
24,3
38,1
30,9
<1,7
<0,7
2,5
re
<2,5
<3,3
<0,7
<5,5
<1,7
<0,7
2,7
5 li
<2,5
4,6
0,7
<5,5
<1,7
<0,7
<1,8
Mittelwert
16,6
30,6
7,6
15,2
4,6
Standard- abweichung
21,8
37,2
11,3
10,1
4,4
Median
6,5
18,9
4,1
18,4
2,7

Ergebnisse
76
Tab. 4.4 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml); Angabe der Einzelwerte sowie von Mittelwert, Standardabweichung und Median, von 5 Katzen (3 Stunden nach Behandlung mit Isopto-Dex® Tropfen).
Kompartiment
Katze Auge 3. Augenlid
Cornea
Kammer-wasser
Iris
Linse
Glas-körper
Retina
re
46,5
16,4
5,7
18,0
<1,7
<0,7
2,1
1 li
235,5
12,6
2,5
12,7
<1,7
<0,7
2,2
re
3,3
16,4
3,7
48,0
<1,7
<0,7
1,8
2 li
6,7
61,6
12,7
38,3
<1,7
<0,7
<1,8
re
<2,5
15,8
1,5
<5,5
<1,7
<0,7
4,3
3 li
<2,5
12,5
<0,7
<5,5
<1,7
<0,7
3,8
re
19,7
62,0
37,1
113,9
2,1
1,3
27,5
4 li
22,8
107,9
13,3
27,2
4,2
<0,7
8,2
re
27,1
68,3
3,4
<5,5
<1,7
<0,7
5,6
5
li
<2,5
66,8
2,7
21,3
<1,7
<0,7
<1,8
Mittelwert
14,3
44,0
8,3
28,7
5,7
Standard- abweichung
15,9
33,5
11,0
33,7
8,0
Median
13,2
39,0
3,5
19,6
3,0
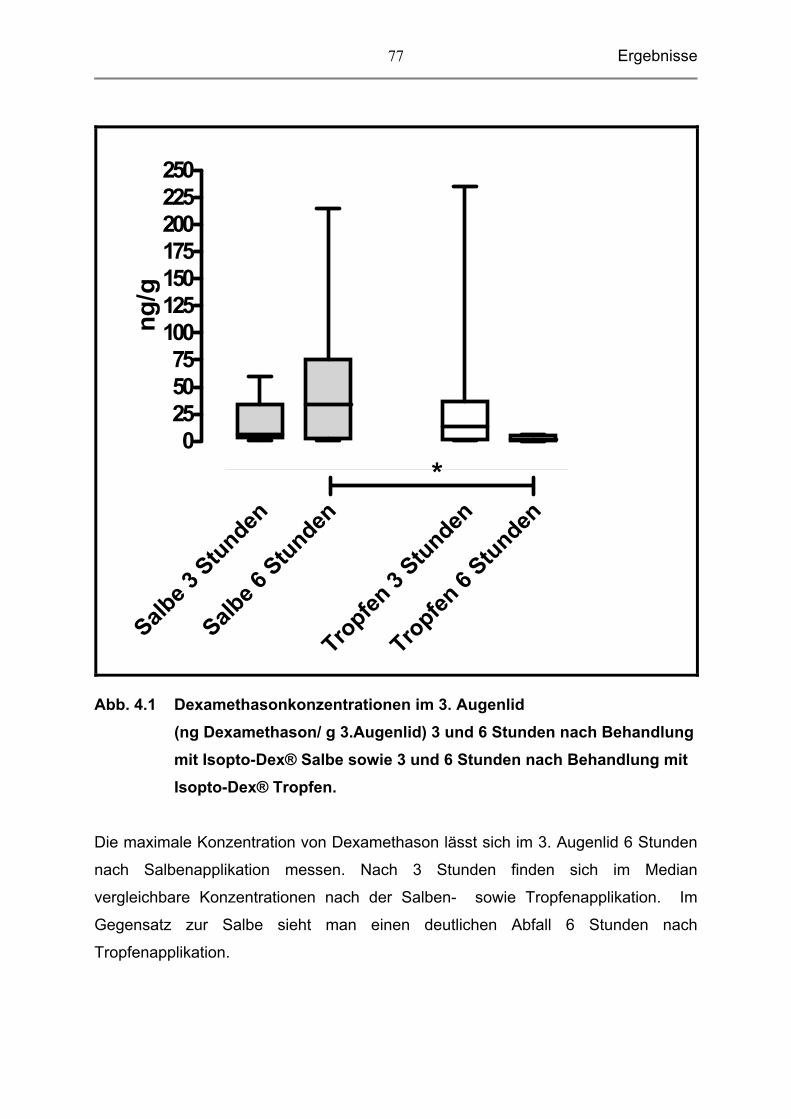
Ergebnisse
77
Salbe 3
Stunden
Salbe 6
Stunden
Tropfen
3 Stunden
Tropfen
6 Stunden
0255075
100125150175200225250
*
ng/g
Abb. 4.1 Dexamethasonkonzentrationen im 3. Augenlid (ng Dexamethason/ g 3.Augenlid) 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Die maximale Konzentration von Dexamethason lässt sich im 3. Augenlid 6 Stunden
nach Salbenapplikation messen. Nach 3 Stunden finden sich im Median
vergleichbare Konzentrationen nach der Salben- sowie Tropfenapplikation. Im
Gegensatz zur Salbe sieht man einen deutlichen Abfall 6 Stunden nach
Tropfenapplikation.

Ergebnisse
78
Salbe 3
Stunden
Salbe 6
Stunden
Tropfen
3 Stunden
Tropfen
6 Stunden
0255075
100125150175200
***
ng/g
Abb. 4.2 Dexamethasonkonzentrationen in der Cornea
(ng Dexamethason/ g Cornea) 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Die maximale Konzentration von Dexamethason lässt sich in der Cornea 6 Stunden
nach Salbenapplikation messen. Im Gegensatz zur Salbe sieht man einen
signifikanten Abfall 6 Stunden nach Tropfenapplikation.

Ergebnisse
79
Salbe 3
Stunden
Salbe 6
Stunden
Tropfen
3 Stunden
Tropfen
6 Stunden
0
10
20
30
40
ng/m
l
Abb. 4.3 Dexamethasonkonzentrationen im Kammerwasser (ng Dexamethason/ ml Kammerwasser) 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Im Kammerwasser bewegen sich die Werte auf deutlich niedrigerem Niveau als in
der Cornea und im 3. Augenlid. Die maximale Konzentration von Dexamethason
lässt sich im Kammerwasser 3 Stunden nach Tropfenapplikation und 6 Stunden
nach Salbenapplikation messen. Im Gegensatz zur Salbe sieht man einen
tendenziellen Abfall 6 Stunden nach Tropfenapplikation.

Ergebnisse
80
Salbe 3
Stunden
Salbe 6
Stunden
Tropfen
3 Stunden
Tropfen
6 Stunden
0
50
100
150
200
250
300
ng/g
Abb. 4.4 Dexamethasonkonzentrationen in der Iris
(ng Dexamethason/ g Iris) 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Die maximale Konzentration von Dexamethason lässt sich in der Iris 6 Stunden nach
Salbenapplikation messen. Während die Konzentration 6 Stunden nach
Salbenapplikation deutlich ansteigt, fällt die Konzentration nach Tropfenapplikation
tendenziell ab.
In der Linse und dem Glaskörper wurden die Konzentrationen von Dexamethason
unter der Nachweisgrenze gemessen.

Ergebnisse
81
Salbe 3
Stunden
Salbe 6
Stunden
Tropfen
3 Stunden
Tropfen
6 Stunden
0
5
10
15
20
25
30
*
ng/g
Abb. 4.5 Dexamethasonkonzentrationen in der Retina
(ng Dexamethason/ g Retina) 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Die gemessenen Dexamethasonkonzentrationen in der Retina liegen niedriger als in
den Kompartimenten der vorderen Augenkammer. Die maximale Konzentration von
Dexamethason lässt sich in der Retina 6 Stunden nach Salbenapplikation messen. 6
Stunden nach Salbenapplikation finden sich im Vergleich zur Tropfenapplikation
signifikant höhere Dexamethasonkonzentrationen in der Retina.

Diskussion
82
5 DISKUSSION 5.1 Fragestellung und Patientengut In der Veterinärmedizin werden oft Glucocorticoide in Form von Augentropfen,
Augensalben oder einer subkonjunktivahlen Injektion bei der Behandlung von
entzündlichen Erkrankungen des Auges sowie verschiedener Nebenorgane des
Auges eingesetzt.
Eine genaue Feststellung der Wirkstoffkonzentrationen in einzelnen Augenstrukturen
des Katzenauges wurde bislang im Gegensatz zu Untersuchungen am Hund
(KAISER 2003), am Pferd (REICHENBECKER 2002) oder Kaninchen (JANES und
STILES 1963, HAMASHIGE und POTTS 1955) nicht durchgeführt. Es war aus
diesem Grund das Ziel dieser Arbeit festzustellen, welche Konzentrationen in
einzelnen Kompartimenten des Auges nach Verabreichung einer
dexamethasonhaltigen Salbe im Vergleich zu dexamethasonhaltigen Tropfen erreicht
werden.
Im Rahmen dieser Arbeit wurden 20 Katzen vor der Euthanasie mit einer
Dexamethason-Augensalbe oder Dexamethason-Augentropfen lokal behandelt. Die
Katzen wurden aufgrund verschiedener Erkrankungen eingeschläfert. Alle Katzen
wurden in 4 Gruppen aufgeteilt. Die versuchsbedingte Einteilung erfolgte unabhängig
von Alter und Geschlecht sowie Rasse der Tiere (siehe Tab. 5.1). Das Alter der Tiere
lag zwischen 1 und 12 Jahren. Es handelt sich bei diesen Katzen um Patienten der
Kleintierklinik der Tierärztlichen Hochschule Hannover, die in der Zeit von Oktober
2004 bis Juni 2006 behandelt wurden.
Der in der vorliegenden Studie verwendete Radioimmunoassay (RIA) hat sich
hinsichtlich der Detektion verschiedener Arzneimittel in biologischen Strukturen
bereits in der Vergangenheit bewährt (KAEMMERER et al. 1984, REICHENBECKER
2002, KAISER 2003).

Diskussion
83
5.2 Lokale Behandlung des Auges
Die lokale Behandlung des Auges ist im Vergleich zur systemischen Behandlung
aufgrund der einfachen Anwendung, niedriger Kosten sowie des selteneren
Vorkommens systemischer Nebenwirkungen generell vorteilhaft (PAPPA 1994).
Weiterer wichtiger Vorteil der lokalen Applikation von Ophthalmica ist die Umgehung
der physiologischen Barrieren zwischen der Blutbahn und Augenstrukturen
(MAUGER 1994, SCHMIDT 1988).
Für diesen Versuch wurde gezielt die lokale Applikation in den Konjunktivalsack
gewählt, da diese Art der Applikation eine starke Praxisrelevanz besitzt, da sie sicher
zu den am meisten durchgeführten Maßnahmen am Auge zählt. Für den lokalen
Einsatz am Auge werden zumeist wässrige und ölige Lösungen, Salben und seltener
auch Augentabletten verwendet. Dabei ist es wichtig, dass sich die Ophthalmika in
der Tränenflüssigkeit lösen und so ihre gleichmäßige Verteilung auf allen cornealen
und konjunktivalen Oberflächen erreicht wird (MATHIS 1999). Die Bewegung der
Augenlider während des Lidschlags führt zu einer stetigen Vermischung von
Tränenflüssigkeit und Arzneimittel innerhalb des präcornealen Tränenfilms. Es ist zu
berücksichtigen, dass die Tränenflüssigkeit das applizierte Arzneimittel verdünnt und
auch die Kontaktzeit am Wirkort verkürzt.
Augentropfen sollen, um gute lokale Verträglichkeit zu gewährleisten, chemisch-
physikalisch der Tränenflüssigkeit weitestgehend entsprechen (HAMACHER 1976,
CAMPELL 1979, LIST et al. 1982). Normale Tränenflüssigkeit enthält etwa 0,15 bis
0,17 % Protein, 0,8 % Natriumchlorid und 0,2 % Natriumbicarbonat. Der
verdünnende Effekt der Tränenflüssigkeit erlaubt es, Lösungen mit einem
Natriumchloridgehalt von 0,6 bis 1,5 % als augenverträglich einzustufen
(HAMACHER 1976). CO2-gesättigt weist die Tränenflüssigkeit einen pH von 7,4 auf.
Aufgrund der Pufferkapazität der Tränenflüssigkeit werden Augentropfen in einem
pH-Bereich von etwa 7,3 bis 9,7 meistens gut vertragen, wohingegen Lösungen mit
einem pH-Wert von mehr als 11,4 und weniger als 5,8 am Auge immer schmerzhaft
und lokal stark reizend sind (HAMACHER 1976, SCHMIDT 1988).

Diskussion
84
Eine Augensalbe gewährleistet einen längeren Kontakt des enthaltenen Wirkstoffes
mit der cornealen und konjunktivalen Oberfläche (CAMPELL 1979). Die Grundlagen
der Augensalben dürfen das Auge nicht reizen und müssen geschmeidig und
sterilisierbar sein. Außerdem soll eine ausreichende Wirkstofffreisetzung erfolgen
(LIST et al. 1982). In Anbetracht dieser Eigenschaften kommen für Augensalben in
erster Linie Kohlenwasserstoffgele (z.B. Vaselin, eventuell mit Zusatz von flüssigem
Paraffin, Wollwachs oder Wollwachsalkohol) in Frage (LIST et al. 1982). Ein Nachteil
der Augensalben ist eine mögliche Verstopfung der tränenableitenden Wege
(CAMPELL 1979, SCHMIDT 1988), was aber relativ selten vorkommt.
5.3 Glucocorticoide am Auge Studien zu der Verteilung verschiedener Glucocorticoide im Kaninchenauge bei
lokaler Applikation wurden bereits mit radioaktiv markiertem Cortisonacetat
(HAMASHIGE und POTTS 1955) und mit radioaktiv markiertem Cortison (JANES
und STILES 1963) durchgeführt. HAMASHIGE und POTTS (1955) konnten nach
wenigen Minuten verschiedene Konzentrationen von Cortison sowie seinen
Metaboliten Hydrocortison im Auge feststellen. Die höchsten Konzentrationen
wurden in der Cornea und im Kammerwasser nachgewiesen, geringere Werte
fanden sich in der Iris. In Linse, Glaskörper und Netzhaut konnte kein Cortison
gefunden werden. JANES und STILES (1963) berichten, dass nach einer lokalen
Behandlung von Kaninchenaugen mit radioaktiv markiertem Cortison nach 30
Minuten bis 2% applizierter Wirkstoff im Auge gemessen werden konnte. Die Autoren
konnten in allen Augenstrukturen Cortison mit Hilfe einer Radioaktivitätsmessung
nachweisen. So ließen sich in der Cornea 0,7 %, in der Iris 0,3 %, in Sklera 0,2 %, in
der Retina 0,16 %, in der Linse 0,04 % und im Glaskörper 0,01 % des applizierten
Wirkstoffs nachweisen.
TCHERNITCHIV et al. (1980) und HERNANDEZ et al. (1981) haben Untersuchungen
am Kaninchen zum Nachweis von Dexamethason im Auge durchgeführt, wobei
TCHERNITCHIV et al. (1980) Dexamethason systemisch und HERNANDEZ et al.

Diskussion
85
(1981) lokal am Auge applizierten. In beiden Untersuchungen wurde Dexamethason
nicht quantitativ in Cornea, Retina und Sklera mittels Autoradiographie
nachgewiesen.
REICHENBECKER (2002) hat eine Studie zur Pharmakokinetik von Dexamethason
am Auge des Pferdes bei lokaler Applikationen dexamethasonhaltiger Augensalbe
durchgeführt. Eine Gruppe des Pferdes erhielt diese Salbe 3x täglich, die andere 7x
täglich über 6 Tage. Dabei konnte er auswertbare Konzentrationen von
Dexamethason mittels Radioimmunoassay (RIA) in der Cornea, dem Kammerwasser
und der Iris nachweisen. Im Gegensatz zur Cornea und dem Kammerwasser, wo
beide Gruppen keine Unterschiede in Konzentration gezeigt haben, wiesen die
Pferde, die 7x täglich behandelt waren signifikant höhere Dexametasonkonzentration
in der Iris auf.
KAISER (2003) untersuchte die Verteilung von Dexamethason im Auge des Hundes
nach lokaler Applikationen tropffähiger dexamethasonhaltiger Augensalbe in
unterschiedlichem Zeitabstand (6, 11 und 16 Stunden) nach der Behandlung. Die
höchste Konzentration von Dexamethason im 3. Augenlid (11 Stunden nach
Applikation), Cornea und Iris (6 Stunden nach Applikation) gemessen werden. Die
Konzentrationen von Dexamethason waren im Kammerwasser, Glaskörper und
Retina nachgewiesen unter den Nachweisgrenzen.
Die Tendenz solcher Verteilung innerhalb des Auges wird auch durch die Daten der
vorgelegten Arbeit bestätigt: die höchsten Dexamethasonkonzentrationen wurden in
der Cornea und der Iris gemessen, weniger im 3. Augenlied und der Retina. Die
Mengen von Dexamethason in Linse und Glasskörper lagen unter der
Nachweisgrenze und konnten nicht quantifiziert werden.

Diskussion
86
5.3.1 Dexamethason in der Cornea und im 3. Augenlid Die Menge von Dexamethason, die in die Cornea und das 3. Augenlid gelangt, ist
von der Verweildauer des Arzneistoffes auf diesen Augenstrukturen abhängig, die
sich aus den unterschiedlichen Arzneiformen ergibt (Salbe, Tropfen).
Die Studie von JANES u. STILES (1963) bestätigt, dass Augentropfen sich gut mit
Tränenflüssigkeit vermischen und auch schnell resorbiert werden können.
Augentropfen fließen generell schnell über die beiden Tränenpunkte in den Ductus
nasolacrimalis aus dem Conjunktivalsack ab. Die Autoren zeigten, dass 30 Minuten
nach lokaler Augenbehandlung mit radioaktiv markiertem Cortison bis zu 0,5 % des
verabreichten Cortisons aus Waschproben der Nase gewonnen werden können. So
zeigte sich in eigenen Untersuchungen, dass beispielsweise nach der
Tropfenapplikation im Augenlid die mittlere Dexamethasonkonzentration in den
vorderen Augenkompartimenten von 14,3 ng/g (3 Stunden) auf 2,4 ng/g (6 Stunden)
abfällt und in der Cornea von 44 ng/g (3 Stunden) auf 14,4 ng/g (6 Stunden).
Bei der Salbenapplikation hingegen kommt es zu einem Anstieg über die Zeit: So
werden, vergleichbar zu Tropfengruppe, nach 3 Stunden im 3.Augenlid 16,6 ng/g
gemessen, und nach 6 Stunden eine Konzentration von 48,3 ng/g erreicht. Für die
Cornea kann ein Anstieg von 30,6 ng/g (3 Stunden) auf 68,5 ng/g (6 Stunden)
gemessen werden. Diesen Unterschied kann nur auf der galenischen Formulierung
beruhen, da in beiden Fällen die gleiche Menge Dexamethason verabreicht worden
ist. Ansammlungen von Salbenresten in Spalträumen um das 3. Augenlid entziehen
sich mit zunehmender Konsistenz der Drainage über den Tränen-Nasengang, und
können so eine Rolle für den beobachteten Depoteffekt spielen (CAMPELL 1979).
5.3.2 Dexamethason im Kammerwasser
Es fällt auf, dass die Konzentrationen von Dexamethason im Kammerwasser im
Vergleich zu denen im 3. Augenlid und in der Cornea stark abfallen. Während bei der
Salbenapplikation die mittlere Dexamethasonkonzentration nahezu konstant bleibt

Diskussion
87
(7,6 ng/ml 3 Stunden zu 7,7 ng/ml 6 Stunden), fällt die Konzentration nach
Tropfenapplikation von 8,3 ng/ml (3 Stunden) auf 3,8 ng/ml (6 Stunden) ab. Dieser
Abfall korreliert mit dem Abfall, der auch beim 3.Augenlid und der Cornea beobachtet
werden konnte. Es zeigt sich, dass die Bioverfügbarkeit auch für ein tieferes
Kompartiment höher nach Salbenapplikation ist.
Die im Vergleich zur Cornea niedrigeren Konzentrationen im Kammerwasser erklären
LEOPOLD und KROMAN (1960) damit, dass die höchste Konzentration des
Wirkstoffs hier eventuell deutlich früher (ca. 1 Stunde nach Applikation) vorliegt. Sie
haben gezeigt, dass die lokale Applikation von Dexamethason, Methylprednisolon
und Triamcinolon schon nach 20 Minuten zu messbaren Konzentrationen von
Methylprednisolon und Dexamethason im Kammerwasser des Kaninchenauges führt
(LEOPOLD und KROMAN 1960 und KROMAN und LEOPOLD 1961).
CIVIALE et al. (2004) konnten an Kaninchen feststellen, dass nach einer einmaligen
lokalen Applikation von dexamethasonhaltigen Augentropfen die Konzentration von
Dexamethason im Kammerwasser schon nach 60 Minuten ihr Maximum erreichte
und danach stetig abfiel, so dass sie nach 3 Stunden die Nachweisgrenze erreichte.
Das Kammerwasser wird ständig vom nicht pigmentierten Epithel der Procc. ciliares
aktiv sekretiert (NAUMANN 1997) und in die Augenkammern abgegeben.
Anschließend gelangt es über den Angulus iridocornealis oder die Iriswurzel bzw.
den Ziliarkörper (sekundärer Kammerwasserabfluss) in die Blutbahn. So wird das
Kammerwasser innerhalb von ca. 1 Stunde wieder vollständig erneut (LIEBICH,
2003). Diesem Fluss unterliegt auch das im Kammerwasser befindliche
Dexamethason, so dass es zu einer ständigen Auswaschung von Dexamethason
kommt und so generell nur niedrige Werte zu messen sind.
5.3.3 Dexamethason in der Iris Dexamethason wird in die Iris durch Diffusion über Cornea und Kammerwasser
transportiert (SCHOENWALD 1993). Die Konzentrationsveränderungen von
Dexamethason in der Iris ähneln grundsätzlich denen im Kammerwasser und der
Cornea bei den entsprechenden Gruppen von Tieren.
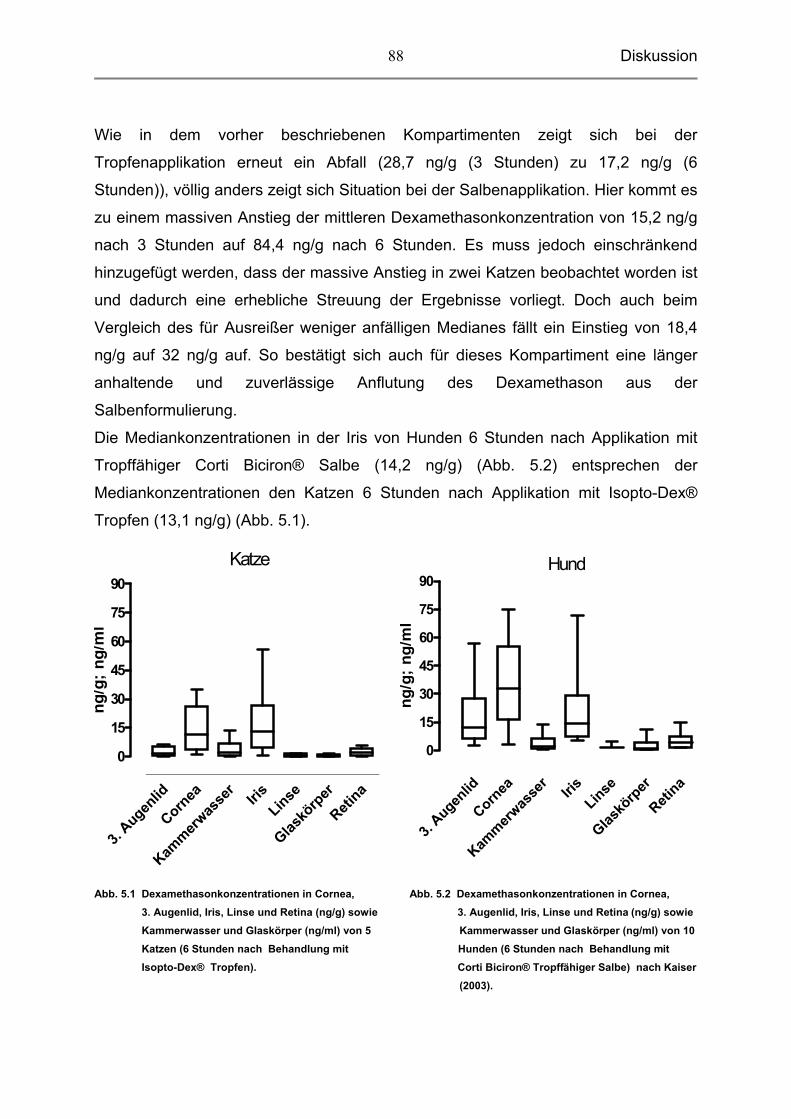
Diskussion
88
Wie in dem vorher beschriebenen Kompartimenten zeigt sich bei der
Tropfenapplikation erneut ein Abfall (28,7 ng/g (3 Stunden) zu 17,2 ng/g (6
Stunden)), völlig anders zeigt sich Situation bei der Salbenapplikation. Hier kommt es
zu einem massiven Anstieg der mittleren Dexamethasonkonzentration von 15,2 ng/g
nach 3 Stunden auf 84,4 ng/g nach 6 Stunden. Es muss jedoch einschränkend
hinzugefügt werden, dass der massive Anstieg in zwei Katzen beobachtet worden ist
und dadurch eine erhebliche Streuung der Ergebnisse vorliegt. Doch auch beim
Vergleich des für Ausreißer weniger anfälligen Medianes fällt ein Einstieg von 18,4
ng/g auf 32 ng/g auf. So bestätigt sich auch für dieses Kompartiment eine länger
anhaltende und zuverlässige Anflutung des Dexamethason aus der
Salbenformulierung.
Die Mediankonzentrationen in der Iris von Hunden 6 Stunden nach Applikation mit
Tropffähiger Corti Biciron® Salbe (14,2 ng/g) (Abb. 5.2) entsprechen der
Mediankonzentrationen den Katzen 6 Stunden nach Applikation mit Isopto-Dex®
Tropfen (13,1 ng/g) (Abb. 5.1).
3. Augen
lid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
15
30
45
60
75
90 Katze
ng/g
; ng/
ml
3. Augen
lid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
15
30
45
60
75
90 Hund
ng/g
; ng/
ml
Abb. 5.1 Dexamethasonkonzentrationen in Cornea, Abb. 5.2 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml) von 5 Kammerwasser und Glaskörper (ng/ml) von 10 Katzen (6 Stunden nach Behandlung mit Hunden (6 Stunden nach Behandlung mit Isopto-Dex® Tropfen). Corti Biciron® Tropffähiger Salbe) nach Kaiser (2003).

Diskussion
89
5.3.4 Dexamethason in der Linse, dem Glaskörper und der Retina In drei hinteren Strukturen des Auges (Linse, Glaskörper und Retina) nehmen die
Konzentrationen von Dexamethason im Vergleich zu den Konzentrationen in den
vorderen Strukturen deutlich ab.
Bei allen 4 Gruppen lagen die Werte in der Linse und im Glaskörper unter den
entsprechenden Nachweisgrenzen (siehe Tab. … Nachweisgrenzen)
Dexamethason könnte durch Diffusion aus dem Kammerwasser auch in die Linse
gelangen. Die Linsenkapsel aber umhüllt als strukturlose, elastische Membran die
Linse und besitzt nach GUM (1991) die Funktion einer semipermeablen Membran.
Das kann ein Grund dafür sein, dass alle Konzentrationen von Dexamethason in der
Linse deutlich unter der Nachweisgrenze liegen.
Eine Reihe von Autoren, wie JANES und STILES (1963), TCHERNITCHIV et al.
(1980) und HERNANDEZ et al. (1981), haben am Kaninchenauge und KAISER
(2003) am Hundeauge festgestellt, dass sich innerhalb der drei hinteren Strukturen
des Auges (Linse, Glaskörper und Retina) die höchsten Konzentrationen von
Dexamethason genau so wie in der vorgelegten Arbeit in der Retina messen lassen.
Auch in der Retina zeigen sich die schon vorgeschriebenen kinetischen Unterschiede
nach Salben- bzw. Tropfenapplikation. Während nach 3 Stunden etwa vergleichbare
mittlere Konzentrationen zu messen sind (4,7 ng/g Salbe zu 5,7 ng/g Tropfen). Zeigt
sich 6 Stunden nach Tropfenapplikation wieder ein Abfall der Konzentration (2,4
ng/g) und ein Anstieg 6 Stunden nach Salbenapplikation (7,9 ng/g). Die mittlere
Dexamethasonkonzentration, die nach 6 Stunden in der Retina des Hundes
gemessen worden ist, liegt mit durchschnittlich 4,9 ng/g im Bereich der in eigenen
Untersuchungen gemessenen Konzentrationen.
Da es einen Stoffaustausch zwischen Retina und dem Glaskörper gibt, kann
Dexamethason auch in den Glaskörper eindringen. Als sicher kann man nur
annehmen, dass aufgrund seiner anatomischen Lage eine mögliche Diffusion von
Dexamethason in den Glaskörper nur von der Retina bzw. Choroidea ausgehen
kann.
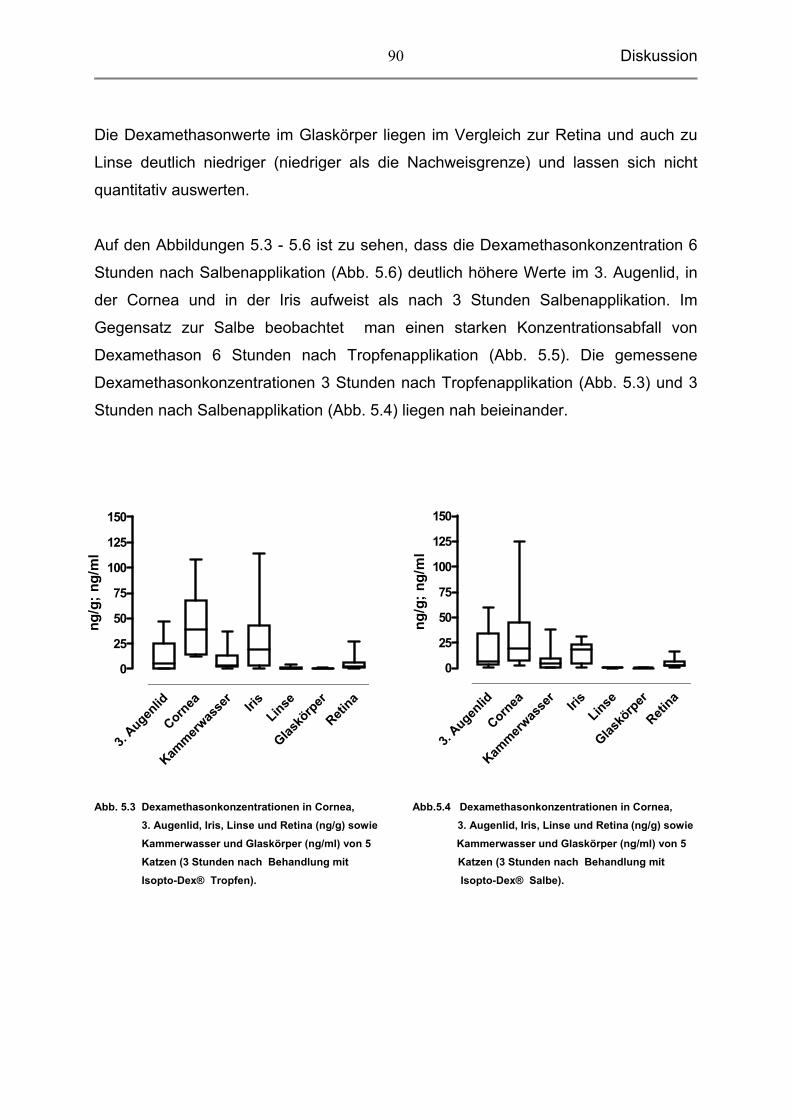
Diskussion
90
Die Dexamethasonwerte im Glaskörper liegen im Vergleich zur Retina und auch zu
Linse deutlich niedriger (niedriger als die Nachweisgrenze) und lassen sich nicht
quantitativ auswerten.
Auf den Abbildungen 5.3 - 5.6 ist zu sehen, dass die Dexamethasonkonzentration 6
Stunden nach Salbenapplikation (Abb. 5.6) deutlich höhere Werte im 3. Augenlid, in
der Cornea und in der Iris aufweist als nach 3 Stunden Salbenapplikation. Im
Gegensatz zur Salbe beobachtet man einen starken Konzentrationsabfall von
Dexamethason 6 Stunden nach Tropfenapplikation (Abb. 5.5). Die gemessene
Dexamethasonkonzentrationen 3 Stunden nach Tropfenapplikation (Abb. 5.3) und 3
Stunden nach Salbenapplikation (Abb. 5.4) liegen nah beieinander.
3.
Augenlid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
25
50
75
100
125
150
ng/g
; ng/
ml
Abb. 5.3 Dexamethasonkonzentrationen in Cornea, Abb.5.4 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml) von 5 Kammerwasser und Glaskörper (ng/ml) von 5 Katzen (3 Stunden nach Behandlung mit Katzen (3 Stunden nach Behandlung mit
Isopto-Dex® Tropfen). Isopto-Dex® Salbe).
3. Augen
lid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
25
50
75
100
125
150
ng/g
; ng/
ml

Diskussion
91
3. Augen
lid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
50
100
150
200
250
300
ng/g
; ng/
ml
Abb. 5.5 Dexamethasonkonzentrationen in Cornea, Abb. 5.6 Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris, Linse und Retina (ng/g) sowie 3. Augenlid, Iris, Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper (ng/ml) von 5 Kammerwasser und Glaskörper (ng/ml) von 5 Katzen (6 Stunden nach Behandlung mit Katzen (6 Stunden nach Behandlung mit Isopto-Dex® Tropfen). Isopto-Dex® Salbe).
5.4 Therapeutisch wirksame Konzentration
Nach der Messung der Dexamethasonkonzentrationen innerhalb verschiedener
Strukturen des Katzenauges stellt sich die Frage, ob die gemessenen
Konzentrationen therapeutisch wirksam sein können.
Die Angaben über wirksame Konzentrationen von Dexamethason variieren erheblich:
SAREEN et al. (1981) zeigte an humanen neutrophilen Granulozyten, dass
Dexamethasonkonzentrationen erst ab ca. 390 bis 3.900 ng /ml einen messbaren
Abfall der Zellaktivität verursacht haben. Niedrigere Konzentrationen (um 40 ng/ml)
waren unwirksam. SHEPARD et al. (2001) konnten Effekte an humanen Zellen aus
3. Augen
lid
Cornea
Kammerw
asse
rIris
Linse
Glaskö
rper
Retina
0
50
100
150
200
250
300
ng/g
; ng/
ml

Diskussion
92
dem trabekulären Netzwerk des iridocornealen Winkels bei
Dexamethasonkonzentrationen um 40 ng/ml nachweisen.
BOURCIER et al. (1999/2000) beobachteten verstärkten Zelluntergang von humanen
cornealen Keratozyten und Epithelzellen bei Dexamethasonkonzentrationen
zwischen 3,9 und 39 ng/ml.
Beim In-vitro-Versuch von BÄUMER (2006) an Mäusekeratinozyten mit
Dexamethason wurde ein signifikanter Abfall der Prostaglandin E2–Synthese im
Konzentrationsbereich um 1 – 10 ng/ml nachgewiesen.
Eine In-Vitro-Studie von FUKOMOTO et. al (1992) an Zellkulturen aus Osteoblasten
und Osteosarkomzellen von Ratten zeigte, dass auch deutlich niedrigere
Dexamethasonkonzentrationen (ca. 0,4 ng/ml) die Zellaktivität unterdrücken.
In der vorgelegten Arbeit wurden Dexamethasonkonzentrationen von bis zu 68,5
ng/g Gewebe in den vorderen Strukturen (Cornea), 84,4 ng/g (Iris), bzw. 7,9 ng/g
Gewebe in den hinteren Strukturen (Retina) gemessen. Die erreichte
Konzentrationen sind nach einer einzigen lokalen Behandlung des Auges gemessen
und nicht nach wiederholten Verabreichungen in kurzen Zeitintervallen (3-4-mal
täglich alle 2 bis 3 Stunden), was der Gebrauchsinformation des Arzneimittels
entspricht. Die mehrfache lokale Applikationen führen zur Akkumulation des
Wirkstoffs und höheren Konzentrationen in Kompartimenten des Auges
(REICHENBECKER 2002). In seinen Untersuchungen zeigt sich zumindest in der Iris
eine signifikante Steigerung der Dexamethasonkonzentrationen nach 7-maliger Gabe
im Vergleich zur 3-maligen Verabreichung.

Zusammenfassung
93
6 ZUSAMMENFASSUNG Julia Bessonova (2006): Untersuchung zur Dexamethasonverteilung im Katzenauge nach lokaler Applikation
Das Ziel der Arbeit war es, festzustellen, welche Konzentrationen von Dexamethason
nach einmaliger lokaler Anwendung einer handelsüblichen Augensalbe (Isopto-
Dex®) und einer Augentropfsuspension (Isopto-Dex®) in den Konjunktivalsack in
verschiedenen Kompartimenten des Katzenauges nachweisbar sind.
In die Studie wurden 20 Katzen einbezogen. Für die Versuchsdurchführung erfolgte
eine Einteilung in vier Gruppen zu je 5 Katzen. Alle Tiere wurden einmal mit 0,05 ml
einer 1 % dexamethasonhaltigen Augentropfsuspension Isopto-Dex® oder 0,05 g 1
% dexamethasonhaltigen Augensalbe Isopto-Dex® behandelt (die Menge entspricht
0,05 mg Dexamethason). Es wurden grundsätzlich beide Augen jedes Tieres
behandelt. Die Tropfen- oder Salbenapplikation erfolgte in den temporalen unteren
Teil des Konjunktivalsackes. Die Tiere wurden 3 oder 6 Stunden nach der
Behandlung euthanasiert. In den aus allen Augen entnommenen Proben von
Cornea, Kammerwasser, Iris, Linse, Glaskörper, Retina / Choroidea sowie drittem
Augenlid wurde die Konzentration von Dexamethason mittels Radioimmunoassay
ermittelt.
Ein Ergebnis der Studie war, dass 3 Stunden nach Salben- oder Tropfenapplikation
die Dexamethasonkonzentrationen nah beieinander lagen. Die höchsten
Konzentrationen wurden in den vorderen Augenkompartimenten bzw. in dem 3.
Augenlid (48,3 ng/g), der Cornea (68,5 ng/g) und der Iris (84,4 ng/g) nachgewiesen.
In den hinteren Kompartimenten des Auges sind deutlich niedrigere
Dexamethasonkonzentrationen beobachtet worden. Im Vergleich zur 3-stündigen
Behandlung waren die Konzentrationen von Dexamethason nach 6-stündiger
Salben- und Tropfenapplikation unterschiedlich. So zeigte sich in allen

Zusammenfassung
94
Kompartimenten ein tendenzieller signifikanter Anstieg der gemessenen
Konzentrationen 6 Stunden nach der Behandlung mit der Isopto-Dex® Salbe. Im
Gegensatz zur Salbe wurde nach 6-stündiger Tropfenapplikation ein starker Abfall
der Dexamethasonkonzentrationen nachgewiesen.
Während die Konzentrationen im Glaskörper und Linse größtenteils unterhalb der
Nachweisgrenze lagen, waren die Dexamethasonkonzentrationen in der Retina noch
quantifizierbar (7,9 ng/g).
Die Ergebnisse dieser Arbeit lassen den Schluss zu, dass eine Salbenapplikation ein
verlängertes Applikationsintervall ermöglicht und insgesamt im Vergleich zur
Tropfenapplikation zu höheren Dexamethasonkonzentrationen führt.

Summary
95
7 SUMMARY Julia Bessonova (2006): Examinations of the distribution of dexamethasone in the feline eye after local administration
The goal of this study was to measure the detectable dexamethasone concentration
in the different compartments of the cat eye after a single topical application of an
eye-ointment or eyedrops into the conjuctival sac.
The study was conducted on 20 cats. The animals were divided into 4 groups of 5
cats each. All animals were treated once with either 0,05 ml of an 1%
dexamethasone eyedropsuspension (Isopto-Dex® Augentropfsuspension) or 0,05 g
of an 1% dexamethasone ointment (Isopto-Dex® Augensalbe) (the amount equals
0,05 mg dexamethasone). Both eyes of each animal were treated. The drug was
applied into temporal canthus of the ventral conjuctival sac. The cats were
euthanized 3 or 6 hours, respectively, after the treatment. The dexamethasone
concentration was evaluated by radioimmunoassay in the cornea, aqueous humor,
iris, lens, vitreous, retina/choroidea and third eyelid.
The result of this study was that 3 hours after the ointment and eyedrop treatment the
dexamethasone concentration was in a similar range regardless of the galenic
formulation. The highest concentration was measured in the anterior compartments
of the eye i.e. the third eyelid (48,3 ng/g), the cornea (68,5 ng/g) and the iris (84,4
ng/g). In the posterior compartments of the eye the dexamethasone concentrations
were markedly lower. Compared to the 3 hour treatment the dexamethasone
concentration 6 hours after application differed within the structures of the eye. The
measured concentrations were significantly higher after treatment with Isopto-Dex®
ointment. Contrary to the ointment, the treatment with eyedrops for 6 hours led to a
strong decrease in dexamethasone concentration. While the concentrations for the

Summary
96
vitreous and lens were generally below the threshold/level of detection, the
dexamethasone concentration for the retina was still quantifiable (7,9 ng/g).
The results of this study lead to the conclusion that the use of ointment results in a
longer application interval and a generally higher concentration of dexamethasone
compared to the use of eyedrops.

Literaturverzeichnis
97
8 LITERATURVERZEICHNIS ALVERDES, K., BERTOLINI, R. (1974)
Grundlagen der Anatomie
VEB Georg Thieme, Leipzig
BINKHORST, G.L. (1987)
Antibiotic levels in bovine lacrimal fluid after single application of ointments
containing procaine benzyl penicillin plus dihydrostreptomycin; and benzathine
cloxacillin
The Veterinary Record, 121, 124-125
BÄUMER W.(2006)
Wirksame Konzentrationen von Dexamethason.
Persönliche Mitteilung durch Dr. W. BÄUMER, Hannover.
BEHREND, E.N., u. GRECO,D.S. (1995)
Clinical applications of glucocorticoid therapy in nonendocrine disease.
In: Kirk's Current Veterinary Therapy XII - Small Animal Practice (JD Bonagura, ed),
WB Saunders Company, Philadelphia (USA)
BENKER, G. (1995)
Glucocorticoide: Wirkung pharmakologischer Dosen auf das Endokrinum.
in: ALLOLIO, B., G. BENKER u. SCHULTE H. M.: Nebennieren und Stress: Von
den Grundlagen zur Klinik.
Verlag Schattauer, Stuttgart, New York

Literaturverzeichnis
98
BERG, R. (1995)
Angewandte und topographische Anatomie der Haustiere
Gustav Fischer, Jena, Stuttgart
BERSON, S. A. u. R. S. YALOW (1964):
Immunoassay of protein hormones.
in: PINCUS, G., K. V. THIMANN u. E. B. ASTWOOD (Hrsg.): The Hormones, vol. IV.
Academic Press, New York
BETTENDORF, G. (1995)
Zur Geschichte der Endokrinologie und Reproduktions-Medizin.
Springer Verlag, Heidelberg.
BLACK, R., R. OBLESBY, L. SALLMANN, J. BUNIM (1960):
Posterior subcapsular cataracts induced by corticosteroids in patients with
rheumatoid arthritis.
J. Am. Med. Ass. 174, 166-169
BORBE, H.-J. (1989)
Untersuchungen über den präokularen Tränenfilm der Katze und dessen Drainage
unter besonderer Berücksichtigung des Schirmer Tränentests
Wiener Tierärztliche Monatsschrift 76, 313-314
BÖHME G. (1991)
Nervensystem, Sinnesorgane, endokrine Drüsen.
In: Lehrbuch der Anatomie der Haustiere Band IV, R. NICKEL, A. SCHUMMER u. E.
SEIFERLE
Parey, Berlin, Hamburg

Literaturverzeichnis
99
BOURCIER, T., BORDERIE, V., FORGEZ, P., LOMBET, A., ROSTENE, W. und
LAROCHE L. (1999)
In vitro effects of dexamethasone on human corneal keratocytes.
Invest Ophthalmol Vis Sci. 40, 1061-1070
BOURCIER, T., FORGEZ, P., BORDERIE, V., SCHEER, S., ROSTENE, W. und
LAROCHE L.
Regulation of human corneal epithelial cell proliferation and apoptosis by
dexamethasone.
Invest Ophthalmol Vis Sci. 41, 4133-4141
BRATTON, G. R., HUDSON, L. C., KLEMM, W. R., u. DZIEZYC, J. (1988)
The Origins of Innervation of the Feline Eyelids
Anat. Histol. Embryol. 17, 362
CAMPELL, C. B. (1979)
Ophthalmic agents. Oinments or drops?
Vet. Med. Small Anim. Clin. 74, 971-980
CHARMANDARI, E., KINO, T., SOUVATZOGLOU, E., CHROUSOS, G.P. (2003)
Pediatric stress : hormonal mediators and human development.
Horm. Res. 59, 161-179.
CIVIALE, C., BUCARIA, F., PIAZZA, S., PERI, O., MIANO, F. und ENEA, V.(2004)
Ocular Permeability Screening of Dexamethasone Esters Through Combined
Cellular and Tissue Systems
J Ocul Pharmacol Ther. 20, 75-84

Literaturverzeichnis
100
CLERC, B. und KRÄHENMANN, A. (1990)
Augenheilkunde Hund und Katze.
Parey-Verlag, Berlin, Hamburg
COX W.V., A. KUPFERMANN u. H.M. LEIBOWITZ (1972)
Topically applied steroids in corneal disease: II. The role of the drug vehicle in
stromal
absorption of dexamethasone.
Arch Ophthalmol 88, 549
DE KLOET, E.R., REUL, J.M., DE RONDE, F.S., BLOEMERS, M., u . RATKA, A. (1986)
Function and plasticity of brain corticosteroid receptor systems: action of
neuropeptides.
J. Steroid Biochem 25, 723-731.
DEUSCHLE, M., WEBER, B., COLLA, M., MULLER, M., KNIEST, A., u. HEUSER, I. (1998)
Mineralocorticoid receptor also modulates basal activity of hypothalamus-
pituitaryadrenocortical system in humans.
Neuroendocrinology. 68, 355-360.
DYCE, K. M., W. O. SACK und G. J. C. WENSING (1991)
Anatomie der Haustiere.
Ferdinand Enke Verlag, Stuttgart
ESTLER, C.-J. (Hrsg.) (1995)
Pharmakologie und Toxikologie: Lehrbuch für Mediziner, Veterinärmediziner,
Pharmazeuten und Naturwissenschaftler. 4. Auflage.
Verlag Schattauer, Stuttgart, New York

Literaturverzeichnis
101
EVANS-STORMS, R.B. u. CIDLOWSKI, J.A. (1995)
Regulation of apoptosis by steroid hormones.
J.Steroid Biochem.Mol.Biol. 53, 1-8.
FATH EL BAB, M. R., MISK, N. A., HINFY, A., KASSEM, A. M. (1981)
Surgical anatomy of the lens in different domestic animals
Zbl., Vet.Med., R.C., Anat. Histol. Embryol. 11, 27-31
FELDMANN, E.C. u. NELSON, R.W. (1996)
Canine and Feline Endocrinology and Reproduction.
WB Saunders Company, Philadelphia, USA
FERGUSON, D.C. u. HOENIG, M. (2001)
Glucocorticoids, Mineralocorticoids, and Steroid Synthesis Inhibitors.
In: Veterinary Pharmacology and Therapeutics (HR Adams, Hrsg.) Iowa State
University Press, Ames (USA) 8, 649-671
FLOWER R.J. (1989)
Glucocorticoids and the inhibition of Phospholipase A2.
In: SCHLEIMER R.P., H.N. CLAMAN u. A.L. ORONSKY : Anti-inflammatory
steroid action : Basic and Clinical Aspects, Academic Press Inc., San Diego.
FREWEIN, J., VOLLMERHAUS, B. (1994)
Anatomie von Hund und Katze
Blackwell, Berlin
FREY, H.-H., LÖSCHER, W. (1996)
Lehrbuch der Pharmakologie für die Veterinärmedizin.
Ferdinand Enke Verlag, Stuttgart

Literaturverzeichnis
102
FUKUMOTO S., ALLAN E.H., ZEHEB R., GELEHTER T.D. u. MARTIN T.J. (1992)
Glucocorticoid regulation of plasminogen activator inhibitor-1 messenger ribonucleic
acid and protein in normal and malignant rat osteoblasts.
Endocrinology, Vol. 130, 797-804
GELEY, S., FIEGL, M., HARTMANN, B.L., KOFLER, R. (1996)
Genes mediating glucocorticoid effects and mechanisms of their regulation
In: Reviews of: Physiology, Biochemistry and Pharmacology (BLAUSTEIN, M.P.,
GRUNICKE, H., HABERMANN, E., PETTE, D., SCHULTZ, G., u. SCHWEIGER, M.,
Hrsg.)
Springer Verlag, Berlin
GILGER, B.C., WRIGHT, J.C., WHITLEY, R.D., McLAUGHLIN, S:A..(1993)
Corneal thickness measured by ultrasonic pachymetry in cats.
Am J Vet Res. 54, 228-30.
GRÜB, M., MIELKE, J. (2004)
Der Ophthalmologie
Springer-Verlag / Heidelberg
GRÜSSER, O. J. u. U. GRÜSSER-CORNEHLS (1995)
Gesichtssinn und Okulomotorik.
in: SCHMIDT, R. F. u. G. THEWS (Hrsg.): Physiologie des Menschen.
Verlag Springer, Berlin, Heidelberg, New York, Barcelona, Budapest, Hong Kong,
London, Mailand, Paris, Tokyo
GUM G.G. (1991)
Physiology in the eye.
in: GELATT K.N.: Textbook of veterinary ophthalmology. Second Edition.
Verlag Lea & Felbinger, Philadelphia

Literaturverzeichnis
103
GUSTAFSSON, J.A., WIKSTRÖM A.C., DENIS M. (1989)
The non-activated glucocorticoid receptor: Structure and activation.
J. Steroid Biochem. 34, 53-62
GUTSCHER, M., EDER, S., MÜLLER, M., CLAUS, R. (2001)
The porcine glucocorticoid receptor: sequencing a 2.1 kb cDNA fragment and raising
antibodies for western bloting, immunoprecipitation and immunohistochemistry
Anim. Sci. 73, 267 – 278
HAMASHIGE S. u. POTTS A. (1955)
The penetration of cortisone and hydrocortisone into the ocular structures.
Am. J. Ophthalmol. 40, 211
HAMACHER, H. (1976)
Augenarzneien, mikrobiologische und biopharmazeutische Aspekte.
Pharmazie in unserer Zeit 5, 77-93
HENCH, P.S., KENDALL, P.D., SLOCUMB, C.H.u. POLLEY HF (1949)
The efect of a hormone of the adrenal cortex (17-hydroxy-11-dehydrocorticosterone:
Compound E) and of pituitary adrenocorticotropic hormone on rheumatoid arthritis.
Pro Staff Meet Mayo Clin. 2, 183
HERRTAGE, M.E. (1996)
Nebennieren.
In: HUTCHISON, M., DÖCKE, F. (deutsche Übersetzung)
Kompendium der Endokrinologie, Hund und Katze
Schlütersche Verlagsanstalt und Druckerei GmbH & Co., Hannover

Literaturverzeichnis
104
HERNANDEZ, M. R., WENK E. J., WEINSTEIN B. I., GORDON G. G., DUNN M. W.,
u. SOUTHREN A. L. (1981)
Corneal conjunctival uptake of topical 3H-dexamethasone in the rabbit eye.
Invest. Ophthalmol. Vis. Sci. 20, 120
HUBEL, D.H. (1989 )
Auge und Gehirn, Neurobiologie des Sehens.
Spektrum der Wissenschaft , Heidelberg
HUNTER, P.J., FOWLER, P.D. u. WILKINSON, P. (1973)
Treatment of anterior uveitis. Comparison of oral oxyphenbutazone and topical
steroids.
Br J Ophthalmol 57, 892-896
IMMNIK, W.F., VAN TOOR, A.J., VOS, J.H., VAN DER LINDE-SIPMAN, J.S. u.
LUBBERINK, A.A. (1992)
Hyperadrenocorticism in four cats.
Vet. Quart. 14, 81-85
JANES, R.G u. STILES J.F. (1963)
The penetration of cortisol into normal and pathologic rabbit eyes.
Am. J. Ophthalmol. 56, 84
JANNING, T. (1993)
Glukokortikoide als katabole Stoffwechselhormone: Methodische Entwicklungen und
Messungen beim Schwein.
Dipl. Ing. agr., Hohenheim
JUSKO, W.J., ROSE J.Q. (1980)
Monitoring prednisone and prednisolone.
Ther. Drug Monit. 2, 169-176

Literaturverzeichnis
105
KAEMMERER, K., KIETZMANN M. u. KREISNER M. (1984)
Untersuchungen mit Magnesium: 2. Mitteilung: Die Wirkung von Magnesiumchlorid
und Magnesiumaspartathydrochlorid auf Stressreaktionen.
Zbl. Vet. Med. Reihe A. 31, 321-333
KAISER, H., KLEY, K. (1992)
Cortisontherapie, 9. Auflage.
Thieme Verlag, Stuttgart.
KAISER, T. (2003)
Untersuchungen zur Verteilung von Dexamethason im Hundeauge nach lokaler
Behandlung
Hannover, Tierärztliche Hochschule, Diss.
KEIPERT, S. (1997)
Ophthalmika: Etablierte Arzneiformen und neue Konzepte
In: MÜLLER, R.H., HILDEBRAND, G.E., Pharmazeutische Technologie: Moderne
Arzneiformen.
Wissenschaftliche Verlagsgesellschaft mbH, Stuttgart
KISHIDA, K., UENO, N., OTORI, T., (1973)
Studies on corneal permeability of drugs. I. Effects of medication of drugs on corneal
permeability.
Acta Soc. Ophthalmol. Jap. 77, 804-808
KLINKE, R., SILBERNAGEL ,S. (2001)
Lehrbuch der Physiologie, 3 Auflage.
Georg Thieme Verlag, Stuttgart

Literaturverzeichnis
106
KNEPEL, W. (2005)
Nebennierenrindenhormone
In: AKTORIES, K., FÖRSTERMANN, U., HOFMANN, F., STARKE, K.: Allgemeine
und spezielle Pharmakologie und Toxikologie, 9. Auflage
Elsevier Urban und Fischer, München, Jena
KÖBBERLING, J. u. ROTENBERGER,J. (1993)
Methodische Konzepte zur Prüfung der Glucocorticoidwirkung.
in: SCHULTE, H. M., G. BENKER u. B. ALLOLIO: Therapie mit Glucocorticoiden:
molekularbiologische, pharmakologische und klinische Aspekte.
Verlag Schattauer, Stuttgart, New York
KOCH, S.A. und RUBIN, L.F. (1972)
Distribution of cones in retina of the normal dog.
Am J Vet Res. 33 , 361-363
KOMAI, Y., USHIKI, T. (1991)
The three-dimensional organization of collagen fibrils in the human cornea and
sclera. Invest. Ophthalmol. Vis. Sci. 32, 2244-2258
KÖNIG, H. E. (1992)
Anatomie der Katze
Gustav Fischer, Jena, Stuttgart
KÖNIG,H.E. und LIEBICH, H.-G. (2005)
Anatomie der Haussäugetiere, 3 Auflage.
Verlag Schattauer, Stuttgart

Literaturverzeichnis
107
KROMANN H.S. u. LEOPOLD I.H. (1961)
Studies upon methyl- and fluoro-substituted prednisolones in the aqueous humor of
the rabbit.
Am J Ophthalmol 52 , 77
LABHART, A. u. MÜLLER, J. (1978)
Physiologie.
In: LABHART, A. (Hrsg.): Klinik der inneren Sekretion, 3 Auflage
Springer Verlag, Berlin, Heidelberg, New York,
LEIBOWITZ, H.M , COX, W.V., KUPFERMANN, A.. (1972)
Topically applied steroids in corneal di-sease. I. The role of inflammation in stromal
absorption of dexamethasone.
Arch. Oph-thalmol. 88, 308-313
LEIBOWITZ, H.M. ,KUPFERMANN, A. (1974)
Topically applied steroids in corneal disease.V. Dexamethasone alcohol:
Arch. Ophthalmol. 92, 329-330
LEOPOLD I.H. u. F. MAYLATH (1952)
Intraocular penetration of cortisone and its effectiveness against experimental cornea
burns.
Am J Ophthalmol 35 , 1125
LEOPOLD I.H. u. KROMANN H.S. (1960)
Methyl- and fluoro-substituted prednisolones in the blood and aqueous humor of the
rabbit.
Arch. Ophthalmol. 63, 943

Literaturverzeichnis
108
LIEBICH, H.-G. (1993)
Funktionelle Histologie, 2. Auflage.
Verlag Schattauer, Stuttgart, New York
LIEBICH, H.-G. (2004)
Funktionelle Histologie der Haussäugetiere, 4 Auflage
Schattauer GmbH Verlag
LIST, P. H., MÜLLER B. W. u. NÜRNBERG E. (1982):
Arzneiformenlehre.
Wiss. Verlagsgesellschaft, Stuttgart
MANGELSDORF, D.J., THUMMEL, C., BEATO, M., HERRLICH, P., SCHÜTZ, G.,
UMESONO, K., BLUMBERG, B., KASTNER, P., MARK, M., u. CHAMBON, P. (1995)
The nuclear receptor superfamily: the second decade.
Cell 83, 835-839.
MATHIS G.A. (1999)
Clinical Opthalmic Pharmacology and Therapeutics. Part 1: Ocular drug delivery.
In: GELATT K.N.: Veterinary ophthalmology 3rded.,
Lippincott Williams & Wilkins,
Baltimore, Maryland
MATTHEWS, A.G., HANDSCOMBE M.C. (1983)
Superficial keratitis in the horse: Treatment with the antiviral drug idoxuridin.
Equine Vet. J. 2 (Suppl.), 29-31
MÄTZ-RENSING, K. (1993)
Gegenüberstellung klinischer, licht- und elektronenmikroskopischer Befunde am
Augenhintergrund des Pferdes.
Hannover, Tierärztliche Hochschule, Diss.

Literaturverzeichnis
109
MAUGER T.F. (1994)
Antibacterials.
In: Havener`s Ocular Pharmacology, 6thed., MAUGER T.F. and CRAIG E.L.
Mosby-Year Book, Inc., St. Louis.
MAURICE , D.V (1980)
Structures and fluids involved in the penetration of topically applied drugs.
Inter. Ophthal. Clinics 20, 7-20
Mc EWEN, B.S., DE KLOET, E.R., ROSTENE, W., (1986)
Adrenal receptors and actions in the nervous system.
Physiol. Rev. 66, 1121
MIDDLETON, D.J. u. WATSON, A.D. (1985)
Glucose intolerance in cats given short-term therapies of prednisolone and megestrol
acetate.
Am J Vet Res 46, 2623-2625
MIDDLETON, D.J., WATSON, A.D., HOWE, C.J. u. CATERSON, I.D. (1987)
Suppression of cortisol responses to exogenous adrenocorticotrophic hormone, and
the occurrence of side effects attributable to glucocorticoid excess, in cats during
therapy with megestrol acetate and prednisolone.
Can J Vet Res 51, 60-65
MÖSTL, E. (2000)
Spezielle Endokrinologie.
In: ENGELHARDT, W. v., BREVES, G. : Physiologie der Haustiere,
Enke, Stuttgart

Literaturverzeichnis
110
NAUMANN. G.O.H. (1997)
Pathologie des Auges.
Verlag Springer, Berlin, Heidelberg
NELSON, R.W., FELDMAN, E.C. u. SMITH, M.C.(1988)
Hyperadrenocorticism in cats: seven cases (1978-1987).
J Am Vet Med Assoc (JAVMA) 193, 245-250
NICKEL, R., SCHUMMER A. u. SEIFERLE E. (1992)
Lehrbuch der Anatomie der Haustiere.
3. Aufl., Bd. 4: Nervensystem, Sinnesorgane, Endokrine Drüsen.
Verlag Parey, Berlin, Hamburg
OETTEL, M. (1996)
Endokrinpharmakologie.
in: FREY, H-H. u. W. LÖSCHER : Lehrbuch der Pharmakologie und Toxikologie für
die Veterinärmedizin.
Verlag Enke, Stuttgart
OHRLOFF, S. (1987)
Permeabilität der zellulären Grenzschichten der Hornhaut in vivo.
Fortschr. Ophthalmol. 84, 307-312
OLIPHANT, L.W., HUDON, J., BAGNARA, J.T. (1992)
Pigment cell refugia in homeotherms--the unique evolutionary position of the iris.
Pigment Cell Res. 5 ,367-71
PRINCE J.H., DIESEN C.D., GLITISI. E. et al. (1960)
Anatomy and histology of the eye and orbit in domestic animals.
Springfield, Il; Charles C. Thomas.

Literaturverzeichnis
111
RAVIOLA G. (1977)
The structural basis of the blood ocular barriers: the ocular and cerebrospinal fluids.
Exp. Eye Res. 25 (Suppl.), 27
REICHENBECKER F. (2002)
Untersuchungen zur Pharmakokinetik von Dexamethason am Auge des Pferdes.
Hannover, Tierärztliche Hochschule, Diss.
ROCHE LEXIKON MEDIZIN (2003)
5. Auflage
Elsevier GmbH, Urban & Fischer Verlag, München-Jena
ROHDEWALD, P., REHDER S. u. WÜRTHWEIN G. (1993):
Rezeptoraffinität synthetischer Glucocorticoide.
in: SCHULTE, H. M., G. BENKER u. B. ALLOLIO: Therapie mit Glucocorticoiden:
molekularbiologische, pharmakologische und klinische Aspekte.
Verlag Schattauer, Stuttgart, New York, S. 39-50
ROHEN, W. (1973)
Feinstrukturelle Veränderungen im Trabekelwerk des menschlichen Auges
bei verschiedenen Glaukomformen.
Klin. Mbl. Augenheilk. 163, 401
SAMUELSON D.A. (1991)
Ophthalmic Anatomy. In: Veterinary Ophthalmology.
Edited by: K.N. GELATT, 3rd Edition.
Lippincott Williams & Wilkins /Pennsylvania.
SAREEN P.M., LODHA S.K., KALRA V.B., UTREJA S.R. u. VIJAY R.K. (1981)
Effect of dexamethasone on bactericidal activity of human neutrophils.
Indian. J. Physiol. Pharmacol. 25, 180-183

Literaturverzeichnis
112
SCHEUNERT, A. u. TRAUTMANN, A.(1987)
Lehrbuch der Veterinär-Physiologie, 7.Auflage
Paul Parey Verlag, Berlin und Hamburg.
SCHMIDT V. (1988)
Augenkrankheiten der Haustiere, 2. Auflage
F. Enke Verlag, Stuttgart.
SCHMID, W., COLE, T.J., BLENDY, J.A., u. SCHÜTZ, G. (1995)
Molecular genetic analysis of glucocorticoid signalling in development.
J.Steroid Biochem.Mol.Biol. 53, 33-35.
SCHOENWALD R. (1985)
The control of drug bioavailability from ophthalmic dosage forms.
In: SMOLEN V.F., L.A. BALL :Controlled drug bioavailability Vol. 3,
New York, John Wiley & Sons
SCHOENWALD R.D. (1993)
Ocular pharmakokinetics / pharmakodynamics.
In: Ophthalmic Drug Delivery Systems, MITRA A.K.
Marcel Dekker Inc., New York,
SCOTT, D.W., KIRK, R.W. u. BENTINCK-SMITH, J. (1979)
Some effects of short-term methylprednisolone therapy in normal cats.
Cornell Vet 69, 104-115
SHARPE P.M., HAYNES, N.B., BUTTERY, P.J. (1986)
Glucocorticoid status and growth.
In: Control and manipulation of animal growth (BUTTERY P.J., HAYNES N.B.,
LINDSAY D.B., Hrsg.).
Butterworth, London Boston Durban Singapore Sidney Toronto Wellington

Literaturverzeichnis
113
SHELL J.W. (1982)
Pharmakokinetics of topically applied ophthalmic drugs.
Surv. Ophthalmol. 26, 207
SHEPARD, A.R., JACOBSON, N., FINGERT, J.H., STONE, E.M., SHEFFIELD, V.C.
und CLARK, A.F( 2001)
Delayed secondary glucocorticoid responsiveness of MYOC in human trabecular
meshwork cells.
Invest Ophthalmol Vis Sci 42, 3173-3181.
SMEETH, L., BOULIS, M., HUBBARD, R. u.FLETCHER, A.E. (2003)
A population based case-control study of cataract and inhaled corticosteroids.
Br J Ophthalmol 87, 1247-1251
STEINBERG R.H., REID M. u. LACY P.L. (1973)
The distribution of rods and cones in the retina of the cat.
J. Comp. Neurol. 148, 229-235
STRAUB, W. (1992):
Augenkrankheiten.
In: KAISER, H., KLEY H.K. (1992):
Cortisontherapie, Corticoide in Klinik und Praxis.
Thieme, Stuttgart, New York
TCHERNITCHIV, A., WENK E. J., HERNANDEZ M. R., WEINSTEIN B. I.,
DUNN M. W., GORDON G. G. u. SOUTHERN A. L. (1980)
Glucocorticoid localization by radioautography in the rabbit eye of 3H-
dexamethasone.
Invest. Ophthalmol. Vis. Sci. 19, 1231

Literaturverzeichnis
114
TRIPATHI, R.C., PARAPURAM, S.K., TRIPATHI, B.J., .ZHONG, Y. u. CHALAM, K.V.
(1999)
Corticosteroids and glaucoma risk.
Drugs Aging 15, 439-450,
THUN, R., EGGENBERGER F. u. ZEROBIN K. (1990)
24-hour profiles of plasma cortisol and testosterone in the male dog: absence of
circadian rhythmicity, seasonal influence and hormonal interrelationships.
Reprod. Dom. Anim. 25, 68-77
THUN, R. u. SCHWARTZ-PORSCHE D. (1994)
Nebennierenrinde.
In: F. DÖCKE, Hrsg.: Veterinärmedizinische Endokrinologie.
Gustav Fischer Verlag, Stuttgart
ULMER, M. J., HAUPT, R. E., HICKS, E. A. (1971):
Anatomy of the cat, an Atlas and Dissection guide
Harper & Row, New York, San Francisco, London
UNGEMACH, F. R. (1999):
Pharmakologische Beeinflussung von Entzündungen.
In: LÖSCHER, W., F. R. UNGEMACH, R. KROKER: Grundlagen der
Pharmakotherapie bei Haus- und Nutztieren, 4. Aufl.,
Parey, Berlin und Hamburg
WILCOCK, B. P. (1993)
The eye and ear.
in: JUBB, K. V. F., P. C. KENNEDY u. N. PALMER: Pathology of domestic animals.
Academic. Press Inc., Orlando

Literaturverzeichnis
115
WISSDORF, H., OTTO B. u. GERHARDS H. (1998)
in: WISSDORF. H., H. GERHARDS u. B. HUSKAMP: Praxisorientierte Anatomie
des Pferdes.
Verlag M. & H. Schaper, Hannover

Anhang
116
9 ANHANG
Verzeichnis der Abbildungen
Abbildung 2.1: Schematische Darstellung des Katzenauges mit
Bezeichnung der Strukturen (WISSDORF und KAPITZKE).
Abbildung 2.2: Aufbau der Cornea (OHRLOFF 1987).
Abbildung 2.3: Schematische Darstellung der Schichten der Netzhaut
(LIEBICH 1993).
Abbildung 2.4: Tränenapparat der Katze (ULMER 1971).
Abbildung 2.5: Schematische Darstellung des Augenvordergrundes
(LIEBICH 1993).
Abbildung 2.6: Wirkungsmechanismus der Steroidhormon-Rezeptoren
(KLINKE und SILBERNAGEL 2001).
Abbildung 2.7: Chemische Struktur des Cortisol und einiger synthetischer
Derivate (FREY 1996).
Abbildung 3.1: Entfernung des Augapfels (CLERC und KRÄHENMANN 1990).
Abbildung 4.1: Dexamethasonkonzentrationen im 3. Augenlid
(ng Dexamethason/ g 3.Augenlid) 3 und 6 Stunden nach
Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden
nach Behandlung mit Isopto-Dex® Tropfen.

Anhang
117
Abbildung 4.2: Dexamethasonkonzentrationen in der Cornea
(ng Dexamethason/ g Cornea) 3 und 6 Stunden nach
Behandlung mit Isopto-Dex® Salbe sowie 3 und 6 Stunden
nach Behandlung mit Isopto-Dex® Tropfen.
Abbildung 4.3: Dexamethasonkonzentrationen im Kammerwasser
(ng Dexamethason/ ml Kammerwasser) 3 und 6 Stunden
nach Behandlung mit Isopto-Dex® Salbe sowie 3 und
6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Abbildung 4.4: Dexamethasonkonzentrationen in der Iris
(ng Dexamethason/ g Iris) 3 und 6 Stunden nach
Behandlung mit Isopto-Dex® Salbe sowie 3 und
6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Abbildung 4.5: Dexamethasonkonzentrationen in der Retina
(ng Dexamethason/ g Retina) 3 und 6 Stunden nach
Behandlung mit Isopto-Dex® Salbe sowie 3 und
6 Stunden nach Behandlung mit Isopto-Dex® Tropfen.
Abbildung 5.1: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 5
Katzen (6 Stunden nach Behandlung mit
Isopto-Dex® Tropfen).
Abbildung 5.2: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 10
Hunden (6 Stunden nach Behandlung mit
Corti Biciron® Tropffähiger Salbe) nach Kaiser (2003).

Anhang
118
Abbildung 5.3: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 5
Katzen (3 Stunden nach Behandlung mit
Is opto-Dex® Tropfen).
Abbildung 5.4: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 5
Katzen (3 Stunden nach Behandlung mit
Isopto-Dex® Salbe).
Abbildung 5.5: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 5
Katzen (6 Stunden nach Behandlung mit
Isopto-Dex® Tropfen). Abbildung 5.6: Dexamethasonkonzentrationen in Cornea,
3. Augenlid, Iris, Linse und Retina (ng/g) sowie
Kammerwasser und Glaskörper (ng/ml) von 5
Katzen (6 Stunden nach Behandlung mit
Isopto-Dex® Salbe).

Anhang
119
Verzeichnis der Tabellen
Tabelle 2.1: Nebenwirkungen der Glucocorticoide (URBAN und
FISCHER 2003).
Tabelle 2.2: Übersicht einiger häufig eingesetzter Glucocorticoide
Tabelle 3.1: Klinische Besonderheiten bei Patienten der 1.Gruppe
(Euthanasie und Probengewinnung 6 Stunden nach
der Behandlung mit der Isopto-Dex® Augensalbe).
Tabelle 3.2: Klinische Besonderheiten bei Patienten der 2.Gruppe
(Euthanasie und Probengewinnung 6 Stunden nach
der Behandlung mit der Isopto-Dex® Augentropfsuspension).
Tabelle 3.3: Klinische Besonderheiten bei Patienten der 3.Gruppe
(Euthanasie und Probengewinnung 3 Stunden nach
der Behandlung mit der Isopto-Dex® Augensalbe).
Tabelle 3.4: Klinische Besonderheiten bei Patienten der 4.Gruppe
(Euthanasie und Probengewinnung 3 Stunden nach
der Behandlung mit der Isopto-Dex® Augentropfsuspension).
Tabelle 3.5: Bechandlungsschema.
Tabelle 3.6: Angaben zur Kreuzreaktivität des verwendeten
Dexamethason- Antiserums (Angaben von Biogenesis
Ltd., Poole UK).
Tabelle 3.7: Durchführung der Dexamethason - RIA, tabellarische
Auflistung, Standardreihe (1-22) sowie Probenmessung (23ff).

Anhang
120
Tabelle 3.8: Signifikanzstufen in Abhängigkeit von der
Irrtumswahrscheinlichkeit p ≤ 0,05.
Tabelle 3.9: Angaben zur Wiederholbarkeit (Total count = 100 %).
Tabelle 3.10: Nachweisgrenzen (NG) für Dexamethason (DXM) in den
Verschiedenen Kompartimenten des Katzenauges.
Tabelle 4.1: Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris,
Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper
(ng/ml); Angabe der Einzelwerte sowie von Mittelwert,
Standardabweichung und Median, von 5 Katzen (6 Stunden
nach Behandlung mit Isopto-Dex® Salbe).
Tabelle 4.2: Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris,
Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper
(ng/ml); Angabe der Einzelwerte sowie von Mittelwert,
Standardabweichung und Median, von 5 Katzen (6 Stunden
nach Behandlung mit Isopto-Dex® Tropfen).
Tabelle 4.3: Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris,
Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper
(ng/ml); Angabe der Einzelwerte sowie von Mittelwert,
Standardabweichung und Median, von 5 Katzen (3 Stunden
nach Behandlung mit Isopto-Dex® Salbe).
Tabelle 4.4: Dexamethasonkonzentrationen in Cornea, 3. Augenlid, Iris,
Linse und Retina (ng/g) sowie Kammerwasser und Glaskörper
(ng/ml); Angabe der Einzelwerte sowie von Mittelwert,
Standardabweichung und Median, von 5 Katzen (3 Stunden
nach Behandlung mit Isopto-Dex® Tropfen).

121
Hiermit erkläre ich, dass ich die Dissertation „Untersuchung zur Dexamethasonverteilung im Katzenauge nach lokaler Applikation“ selbständig
verfasst habe. Bei der Anfertigung wurde keine Hilfe Dritter in Anspruch genommen.
Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten
(Promotionsberater oder andere Personen) in Anspruch genommen. Niemand hat
von mir unmittelbar oder mittelbar entgeltlich Leistungen für Arbeiten erhalten, die im
Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen.
Ich habe die Dissertation am folgenden Institut angefertigt:
Institut für Pharmakologie, Toxikologie und Pharmazie
der Stiftung Tierärztliche Hochschule Hannover
Bünteweg 17, 30559 Hannover.
Die Dissertation wurde bisher nicht für eine Prüfung oder eine Promotion oder für
einen ähnlichen Zweck zur Beurteilung eingereicht.
Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig
und der Wahrheit entsprechend gemacht habe.
Julia Bessonova

Danksagung
122
DANKSAGUNG Ich bedanke mich bei meinem Doktorvater Herrn Univ.-Prof. Dr. M. Kietzmann für die
Überlassung des interessanten Themas meiner Dissertation, die fachliche Betreuung
und die freundliche Unterstützung.
Ein großes Dankeschön gilt allen Mitarbeitern der Abteilung Pharmakologie, Toxikologie und Pharmazie der Tierärztlichen Hochschule Hannover. W. Bäumer, M.
Braun, F. Niedorf, J. Stahl, A. Werner, K. Hünermann und S. Krekeler möchte ich
hierbei besonders hervorheben, ebenso wie G. Ludwig und H.-H. Bohr, die mich in
die Geheimnisse der Labortechniken mit großem Engagement und Engelsgeduld
eingewiesen haben.
Weiterhin möchte ich mich bei Frau Univ.-Prof. Dr. A. Meyer-Lindenberg und Frau M.
Fork aus der Klinik für kleine Haustiere der Tierärztlichen Hochschule Hannover für
die Hilfe mit den Katzen bedanken.
Ebenfalls herzlich bedanken möchte ich mich bei Familie Klug für die persönliche
Unterstützung und Begleitung seit meinem Aufenthalt in Deutschland.
Allen meinen Freunden aus Deutschland und Russland gilt ein herzliches
Dankeschön.
Einen besonders lieben Dank möchte ich an meinen Mann Dmitry und meine Eltern
für die liebe Unterstützung und den Rückhalt aussprechen.







