
Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Methodenstandardisierung Makrozoobenthos 1
Bestimmungsschlüssel für die aquatischen Zweiflügler (Diptera) in Anlehnung an die
Operationelle Taxaliste für Fließgewässer in Deutschland
ANDREA SUNDERMANN & SUSANNE LOHSE
Der vorliegende Schlüssel ist für die Bestimmung der Zweiflügler (Diptera) konzipiert, die im
Rahmen von Makrozoobenthos-Untersuchungen in Fließgewässern zur Umsetzung der EU-
WRRL in Deutschland zu bearbeiten sind. Das taxonomische Niveau, auf dem die Diptera zu
bearbeiten sind, wird in der Operationellen Taxaliste vorgegeben. Aus praktischen Gründen
wurden in diese Liste und damit auch in den Schlüssel lediglich Taxa mit Bezug zu aquati-
schen Lebensräumen aufgenommen. Darüber hinaus blieben solche Taxa, die nur äußerst sel-
ten in Makrozoobenthosproben vorhanden sind, unberücksichtigt.
In seltenen Fällen kann es bei der Bestimmung solcher (oder terrestrischer) Taxa mit diesem
Schlüssel unter Umständen zu einer Fehlbestimmung der Taxa kommen. Diese Fehlerquelle
wird jedoch im Rahmen der routinemäßigen Bearbeitung von Makrozoobenthosproben in
Kauf genommen, da der Bearbeitungsaufwand mit einem umfassenderen Bestimmungs-
schlüssel ungleich höher wäre.
Zum Aufbau des Schlüssels:
Der erste Teil des Schlüssels beinhaltet den Bestimmungsteil zur Differenzierung der einzel-
nen Familien. Eine Ausnahme bilden hier die Familien der Ceratopogonidae und Muscidae.
Taxa dieser Familien wurden aus praktischen Gründen bereits im ersten Teil des Bestim-
mungsschlüssels über das Familienniveau hinaus ausgewiesen.
In der Operationellen Taxaliste ist darüber hinaus eine weiterführende Bestimmung einzelner
Familien vorgesehen. Betroffene Familien werden daher im zweiten Teil des Bestimmungs-
schlüssels behandelt und so weit aufgeschlüsselt, wie es die Operationelle Taxaliste vorsieht.
Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Familienschlüssel für die letzten Larvenstadien
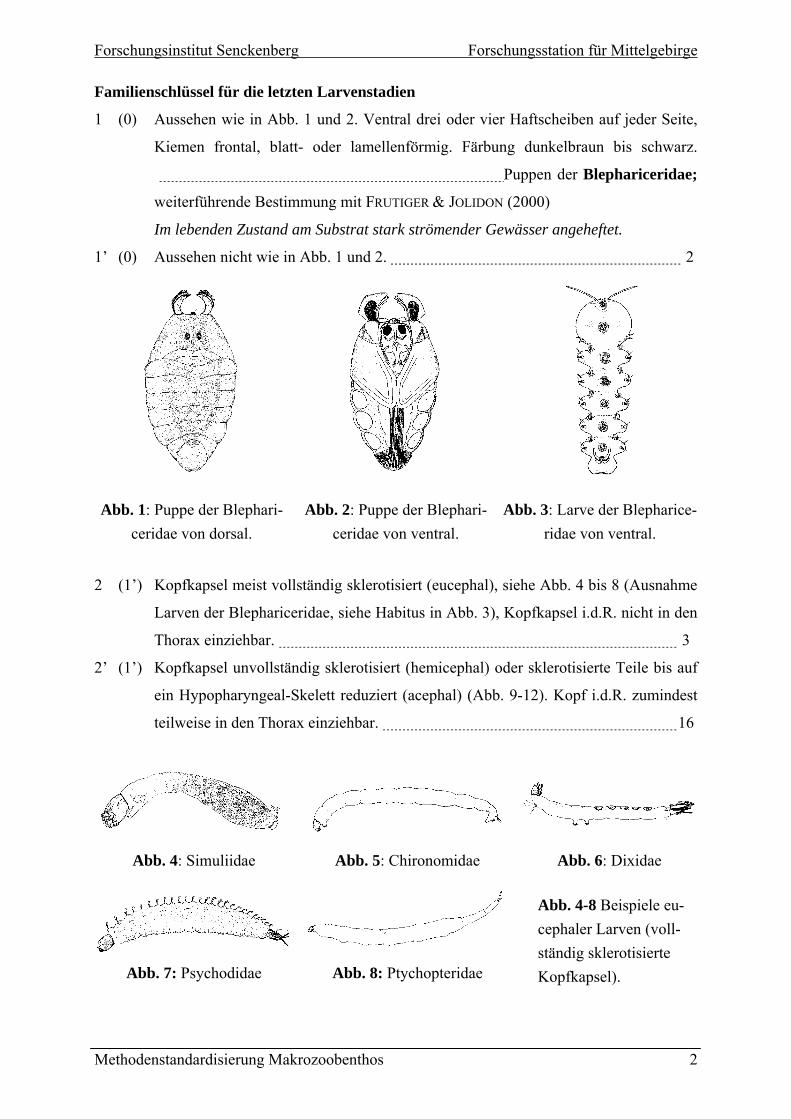
1 (0) Aussehen wie in Abb. 1 und 2. Ventral drei oder vier Haftscheiben auf jeder Seite,
Kiemen frontal, blatt- oder lamellenförmig. Färbung dunkelbraun bis schwarz.
Puppen der Blephariceridae;
weiterführende Bestimmung mit FRUTIGER & JOLIDON (2000)
Im lebenden Zustand am Substrat stark strömender Gewässer angeheftet.
1’ (0) Aussehen nicht wie in Abb. 1 und 2. 2
Abb. 1: Puppe der Blephari-
ceridae von dorsal. Abb. 2: Puppe der Blephari-
ceridae von ventral. Abb. 3: Larve der Blepharice-
ridae von ventral.
2 (1’) Kopfkapsel meist vollständig sklerotisiert (eucephal), siehe Abb. 4 bis 8 (Ausnahme
Larven der Blephariceridae, siehe Habitus in Abb. 3), Kopfkapsel i.d.R. nicht in den
Thorax einziehbar. 3
2’ (1’) Kopfkapsel unvollständig sklerotisiert (hemicephal) oder sklerotisierte Teile bis auf
ein Hypopharyngeal-Skelett reduziert (acephal) (Abb. 9-12). Kopf i.d.R. zumindest
teilweise in den Thorax einziehbar. 16
Abb. 4: Simuliidae Abb. 5: Chironomidae Abb. 6: Dixidae
Abb. 7: Psychodidae Abb. 8: Ptychopteridae
Abb. 4-8 Beispiele eu-cephaler Larven (voll-ständig sklerotisierte Kopfkapsel).
Methodenstandardisierung Makrozoobenthos 2

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 9: Empididae
Abb. 10: Empididae
Abb. 11: Athericidae
Abb. 12: Limoniidae
Abb. 9-12 Beispiele hemi- oder acephaler Larven (unvollständig sklerotisierte Kopf-kapsel).
3 (2) Eine Reihe von Saugnäpfen auf der Ventralseite ausgebildet (Abb. 3), nur sechs
Körpersegmente. Larven der Blephariceridae;
weiterführende Bestimmung mit FRUTIGER & JOLIDON (2000).
Larven in Bereichen mit starker Strömung.
3’ (2) Saugnäpfe fehlen oder anders angeordnet, Anzahl der Körpersegmente ≥ 9. 4
4 (3’) Pseudopodien (Scheinfüßchen mit apikalem Hakenkranz) an Thorax- und/oder Ab-
dominalsegmenten vorhanden (Abb. 4-6, 9-11 und 13-14). 5
4’ (3’) Pseudopodien fehlen (Abb. 7, 8 und 12). 11
5 (4) Pseudopodien nur an den Abdominalsegmenten vorhanden. 6
5’ (4) Pseudopodien an Thorax- und/oder Abdominalsegmenten vorhanden. 7
6 (5) Pseudopodien an Abdominalsegment 1 und 2, charakteristischer Habitus (Abb. 6).
Ventralseite einiger Abdominalsegmente mit Hakenreihen ausgestattet.
Dixidae (weiterführende Bestimmung auf Seite 12)
Lebensraum in der Uferzone der Gewässer im Bereich der Grenzschicht. Die Tiere
nehmen im lebenden Zustand eine charakteristische, verkehrt U-förmige Haltung
ein.
6’ (5) Pseudopodie (unpaar) am letzten Abdominalsegment einziehbar und flexibel, mit
langen Häkchen ausgestattet (Abb. 13-14). Dasyheleinae (Ceratopogonidae)
7 (5’) Pseudopodie (unpaar) am 1. Thoraxsegment. Körper am Hinterende am breitesten,
mit kreisförmig angeordneten Reihen von Häkchen am letzten Abdominalsegment
(Haftscheibe). Habitus wie in Abb. 4. Simuliidae
(weiterführende Bestimmung auf Seite 14)
7’ (5’) Merkmale anders, Aussehen nicht wie in Abb. 4. 8
Methodenstandardisierung Makrozoobenthos 3

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
8 (7’) Körper lateral mit zahlreichen Anhängen oder langen Borsten, Aussehen wie in Abb.
15 - 17. Körper z.T. dorsoventral abgeflacht, Pseudopodien vorne und hinten unpaar.
9
8’ (7’) Körper ohne derartig lange Auswüchse. 10
Abb. 13: Dasyheleinae (Ceratopogonidae)
Abb. 14: Dasyheleinae (Ceratopogonidae),
letzte Abdominalsegmente mit Pseudopodium. Verändert nach HENNIG (1950).
Abb. 15: Atrichopogon
(Ceratopogonidae)
Abb. 16: Atrichopogon
(Ceratopogonidae)
Abb. 17: Forcipomyia
(Ceratopogonidae)
9 (8) Körper seitlich mit langen Auswüchsen, i.d.R. dorsoventral abgeflacht. Habitus wie
in Abb. 15-16. Atrichopogon (Ceratopogonidae)
9’ (8) Körper rundlich mit langen Borsten, Habitus wie in Abb. 17. Forcipomyia
(Ceratopogonidae)
10 (8’) Thorax und Abdominalsegmente dorsal stark sklerotisiert, Dorsalseite daher deutlich
dunkler als Ventralseite. Letztes Abdominalsegment mit Hakenkranz ausgestattet.
Stigmen (Atemorgane) lateral am 1. Thorax- und am 8. Abdominalsegment (Abb.
18). Thaumaleidae
Lebensraum der Larven in Bereichen dünner Wasserfilme (hygropetrische
Lebensweise).
Methodenstandardisierung Makrozoobenthos 4

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
10’ (8’) Keine starke Sklerotisierung der Thorax- und Abdominalsegmente ausgeprägt. Dor-
salseite nicht dunkler als Ventralseite. Am letzten Abdominalsegment paarige Pseu-
dopodien (Nachschieber). Habitus wie in Abb. 5. Chironomidae
(weiterführende Bestimmung auf Seite 11)
11 (4’) Thorax zu einem einheitlichen Segment verwachsen, breiter als die übrigen
Körpersegmente. 12
11’ (4’) Thorax als drei getrennte Segmente erkennbar. 13
12 (11) Antenne als Greiforgan ausgebildet, 7. Abdominalsegment i.d.R. mit paarigen
(pigmentierten) Tracheenblasen ausgestattet (Abb. 19). Chaoboridae
Siedlungsschwerpunkt in Stillgewässern.
12’ (11) Antenne nicht als Greiforgan ausgebildet, nicht mit paarigen Tracheenblasen
Habitus wie in Abb. 20. Culicidae
Siedlungsschwerpunkt in Stillgewässern.
13 (11’) Ausbildung einer Pseudosegmentierung (scheinbar 26 Körpersegmente) (Abb. 21).
Psychodidae (weiterführende Bestimmung auf Seite 13)
13’ (11’) Keine Pseudosegmentierung ausgebildet, weniger als scheinbar 26 Segmente. 14
Abb. 18: Thaumaleidae
Abb. 19: Chaoboridae
Abb. 20: Culicidae
Abb. 21: Psychodidae
Stigmen Stigmen
Tracheenblase
Antenne als Greiforgan ausgebildet
Methodenstandardisierung Makrozoobenthos 5

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 22: Stratiomyidae
Abb. 23: Ceratopogoninae
(Ceratopogonidae)
Abb. 24: Cylindrotomidae; verändert nach HENNIG (1950)
14 (13’) Körperoberfläche vollständig sklerotisiert, erscheint granuliert und fest. Abdomen
i.d.R. dorsoventral abgeflacht (Abb. 22). Stratiomyidae
(weiterführende Bestimmung auf Seite 15)
14’ (13’) Körperoberfläche nicht vollständig sklerotisiert. Abdomen rundlich, lang und
schlank. 15
15 (14’) Mit ausziehbarem Atemrohr am Ende des Abdomens (Abb. 8). Ptychoptera
(Ptychopteridae)
Lebensraum der Larven in Bereichen mit hohem Detritusanteil im Uferbereich
stehender oder fließender Gewässer.
15’ (14’) Ohne ausziehbares Atemrohr am Ende des Abdomens. Habitus nadelförmig und
rund (Abb. 23). Ceratopogoninae (Ceratopogonidae)
16 (2’) Mandibeln arbeiten gegeneinander in horizontaler Ebene (Abb. 26). 17
16’ (2’) Mandibeln arbeiten nebeneinander in vertikaler Stellung (Abb. 25). 19
Abb. 25: Athericidae; Teile des
Hypopharyngealskelettes von lateral.
Ausrichtung der Mandibeln in vertikaler
Stellung.
Abb. 26: Limoniidae; Kopfkapsel von
ventral. Ausrichtung der Mandibeln in
horizontaler Stellung.
Methodenstandardisierung Makrozoobenthos 6

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
17 (16) Abdominalsegmente (dorsal) mit zweigeteilten oder gezähnten Anhängen, die min-
destens die Länge der halben Körperbreite erreichen (Abb. 24 ).
Cylindrotomidae (weiterführende Bestimmung auf Seite 12)
Als Lebensraum dienen submerse Makrophyten oder Moose.
17’ (16) Abdominalsegmente ohne oder mit deutlich kürzeren Anhängen. 18
18 (17’) Stigmenfeld (letztes Abdominalsegment) mit 6 Anhängen (Abb.27-28), Randlappen
rundlich oder abgeflacht, z.T. auch reduziert, Habitus wie in Abb. 29. Tipulidae
(weiterführende Bestimmung auf Seite 17)
18’ (17’) Stigmenfeld mit ≤ 5 Randlappen umgeben. Limoniidae/Pediciidae
(weiterführende Bestimmung auf Seite 17)
Abb. 27: Tipulidae, Stigmenfeld am letzten
Abdominalsegment. Ventral zusätzlich 3 Paar Analkiemen.
Abb. 28: Tipulidae, Stigmenfeld am letzten
Abdominalsegment. Anhänge z.T. reduziert.
Abb. 29: Tipulidae Abb. 30: Tabanidae
1 62 5
3 4 Analkiemen
19 (16’) 3 oder 4 Paar Pseudopodien an jedem Abdominalsegment ausgebildet (Abb. 30).
Tabanidae (weiterführende Bestimmung auf Seite 16)
19’ (16’) Maximal ein Paar Pseudopodien oder Kriechwülste (an der Grenze zweier Segmente
ausgebildet und meist mit einer Querreihe kleiner Dörnchen besetzt) an jedem
Abdominalsegment ausgebildet. 20
20 (16’) Abdominalende mit unpaarem, ausziehbaren Atemrohr. 21
20’ (16’) Abdominalende ohne ausziehbares Atemrohr. 22
21 (20) Atemrohr so lang oder deutlich länger als der Körper (Abb. 31), oder Habitus wie in
Abb. 32. Syrphidae
Als Lebensraum werden häufig Gewässer mit einem hohen Anteil an organischem
Material besiedelt.
21’ (20) Letztes Segment des Atemrohrs in zwei Äste aufgeteilt (Abb.33). Ephydridae
Methodenstandardisierung Makrozoobenthos 7

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 31: Syrphidae (verändert nach HENNIG 1952)
Abb. 32: Syrphidae Abb. 33: Ephydridae
22 (20’) Längere Abdominalanhänge dorsal und/oder lateral an den Abdominalsegmenten
vorhanden, Habitus wie in Abb. 11 oder Abb. 43-44. Athericidae
(weiterführende Bestimmung auf Seite 10)
22’ (20’) Längere Abdominalanhänge fehlen. 23
23 (22’) Pseudopodien vorhanden (Abb. 9-10). Empididae
(weiterführende Bestimmung auf Seite 13)
23’ (22’) Maximal Kriechwülste, jedoch keine Ausbildung von Pseudopodien. 24
24 (23’) Vier konische oder kegelförmige Anhänge am letzten Abdominalsegment (Abb. 35,
37). Habitus wie in Abb. 34 oder 36. 25
24’ (23’) Anhänge am letzten Abdominalsegment anders gestaltet, nicht wie in Abb. 34 oder
36. 26
25 (24) Anhänge am letzten Abdominalsegment dorsal und ventral von gleicher Länge
(Abb. 34-35). Chrysopilus (Rhagionidae)
25’ (24) Ventrale Anhänge am letzten Abdominalsegment länger als dorsale (Abb. 36-37).
Dolichopodidae
Abb. 34: Chrysopilus (Rhagionidae)
Abb. 35: Chrysopilus
(Rhagionidae), letztes
Abdominalsegment
von dorsal.
Abb. 36: Dolichopodidae
Abb. 37:
von dorsal.
Dolichopodidae,
letztes
Abdominalsegment
Methodenstandardisierung Makrozoobenthos 8

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
26 (24’) Vier nach außen bzw. nach vorne divergierende Anhänge am letzten Abdominal-
segment, Anhänge starr, wenig beweglich. Habitus wie in Abb. 38.
Limnophora (Muscidae)
26’ (24’) Habitus nicht wie in Abb. 38, Abdominalanhänge nicht divergierend. 27
27 (26’) Abdominalsegment ohne Anhänge, lediglich zwei hervorstehende Stigmenfelder
(Abb. 40). Habitus wie in Abb. 39. Lispe (Muscidae)
27’ (26’) Stigmenfeld am letzten Abdominalsegment von mehreren Anhängen umgeben (Abb.
41-42). Sciomycidae
Abb. 38: Limnophora (Muscidae)
Abb. 39: Lispe (Muscidae)
Abb. 40: Lispe (Muscidae), Stigmenfeld
am letzten Abdominalsegment.
Abb. 41: Sciomycidae
Abb. 42: Sciomycidae, letztes
minalsegmAbdo ent
Methodenstandardisierung Makrozoobenthos 9

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Weiterführende Bestimmung einzelner Familien:
Athericidae:
1 (22) Abdominalsegmente 6-8 mit langen fädigen Fortsätzen, die die Hälfte der
Körperlänge erreichen können (Abb. 43). Atrichops crassipes
1’ (22) Abdominalsegmente 6-8 nicht mit derartig langen Fortsätzen. 2
2 (1’) Krallen an den Pseudopodien in der äußeren und mittleren Reihe etwa gleich lang
(Abb. 45), Merkmal jedoch erst im letzten Larvenstadium ausgeprägt!
Abdominalanhänge dorsal und lateral etwa gleich lang (Abb. 44). Ibisia marginata
2’ (1’) Reihen der Krallen an den Pseudopodien verschieden lang, äußere Reihe kürzer als
die mittlere Reihe (Abb. 47), Merkmal erst im letzten Larvenstadium ausgeprägt!
Abdominalanhänge dorsal kürzer als lateral (Abb. 46). Atherix ibis
Abb. 43: Atrichops crassipes
Abb: 44: Ibisia marginata
Abb. 45: Ibisia marginata,
Krallenreihen an den Pseudopodien.
Abb. 46: Atherix ibis
Abb. 47: Atherix ibis, Krallenreihe an
den Pseudopodien.
äußere Reihemittlere Reihe
innere Reihe
innere Reihe
äußere Reihe
mittlere Reihe
Methodenstandardisierung Makrozoobenthos 10

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Chironomidae:
1 (10) Kopfkapsel mit einziehbaren Antennen. Antennen häufig unterschiedlich weit
eingezogen, d.h. Antennen scheinbar ungleich lang. Kopfkapsel oval, innerhalb
dieser ist i.d.R. ein Schaft erkennbar, in den die Antenne eingezogen werden kann
(Abb. 48). Tanypodinae
1’ (10) Antennen nicht einziehbar, Kopf rundlich. 2
2 (1’) Kopfkapsel auf der Ventralseite mit charakteristischer Zeichnung (Abb. 51),
Ventromentalplatten mit langen Haaren ausgestattet. Prodiamesa olivacea
2’ (1’) Merkmale anders, charakteristische Zeichnung (Abb. 51) nicht in Kombination mit
langen Haaren an den Ventromentalplatten. 3
3 (2’) Augenstellung in vertikaler Ebene, meistens zwei, nicht miteinander verschmolzene
Augen (Abb. 49), Ausbildung von Ventromentalplatten (siehe Pfeil in Abb. 52-53).
4 Chironominae
3’ (2’) Augenstellung horizontal. Augenstellung in vertikaler Ebene nicht in Kombination
mit Ausbildung von Ventromentalplatten. Augen z.T. miteinander verschmolzen.
Chironomidae Gen. sp. (p.p.)
4 (3) Antenne häufig kürzer als die halbe Kopflänge, Ventromentalplatten durch großen
Zwischenraum voneinander getrennt (Abb. 52). Chironomini
4’ (3) Antenne häufig halb so lang wie der Kopf, Antenne auf einem Antennensockel
(Pfeil in Abb. 49), Ventromentalplatten meistens nicht weiter als die mittlere
Zahnbreite des Mentums voneinander getrennt (Abb. 53). Tanytarsini
Abb. 48: Tanypodinae; Kopfkapsel von dorsal. Kopfkapsel von lateral. Augen
in vertikaler Ebene. hor e.
Abb. 49: Chironominae; Abb. 50: Chironomidae Gen. sp., Kopfkapsel von
lateral. Augen in izontaler Eben
Methodenstandardisierung Makrozoobenthos 11

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 51: Prodiamesa olivacea; Kopfkapsel von
ventral. Kopfkapsel von ventral. Kopfkapsel von ventral. Abb. 52: Chironomini; Abb. 53: Tanytarsini;
Cylindrotomidae:
1 (17) Körperanhänge zweigeteilt, Habitus wie in Abb. 54. Phalacrocera replicata
Körperanh och ähnlich wie in 1’ (17) änge nicht zweigeteilt, sondern gezähnt. Habitus jed
Abb. 54. Triogma trisulcata
Ab ta
(verändert nach HENNIG 1950) b. 54: Phalacrocera replica
Abb. 55: Triogma trisulcata, gesägte Abdominalanhänge.
1 (6) mente dorsal
Dixidae:
Fünf oder mehr Abdominalseg mit einem Haarkranz ausgestattet (Abb.
57). Habitus wie in Abb. 56. Dixa
A1’ (6) bdominalsegmente dorsal ohne Haarkranz. Habitus sonst ähnlich wie in
Abb. 56.
Dixella
Abb. 57: Dixa; Detailzeichnung, Abdo-Abb. 56: Dixa; A mente dorsal mit
Haarkranz. minalsegment mit Haarkranz von dorsal. bdominalseg
Methodenstandardisierung Makrozoobenthos 12

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Empididae:
1 (23) Sieben Paar Pseudopodien (Abb. 58). 2
Acht Paar Pseudopodien (A am letzten 1’ (23) bb. 60). Dorsolaterale Anhänge
Abdominalsegment i.d.R. lang (Abb. 61). Clinocerinae
Letztes Abdominalsegment gerundet, mit se2 (1) hr kleinen Warzen, jede mit zwei oder
drei langen Haaren ausgestattet. (Abb. 58). Chelifera
Dorsolaterale Fo2’ (1) rtsätze am letzten Abdominalsegment sehr kurz, apikale Forsätze
lang (Abb. 59). Hemerodromia
Abb. 58: Chelifera
59: Hemerodromia, leAbb. tztes
Abdominalsegment von dorsal.
Abb. 60: Clinocerinae
. 61: Clinocerinae, letAbb ztes
Abdominalsegment von dorsal.
1 (13)
lerotisierten Platten auf den Tergiten z.T. stark reduziert (Abb. 62).
Psychodidae:
Larven blass, zwischen den Tergiten weniger stark sklerotisiert, Größe der
sk
Psychoda, Tinearia & Jungiella (p.p.)
L1’ (13) arven dunkel gefärbt, zwischen den Tergiten stärker sklerotisiert (Abb. 63).
Psychodidae Gen. sp.
Abb. 63: Psychodidae Gen. sp.
2: Jungiella (Psychodidae). Abb. 6
Methodenstandardisierung Makrozoobenthos 13

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Simuliidae:
K1 (7) opffächer fehlend, Form des Analsklerites y-förmig (Abb. 64-65).
Twinnia hydroides
1’ (7) Kopffächer vorhanden, Form des Analsklerites x-förmig (Abb. 66 und 67). 2
Stirnsklerit vor der Basis am breitesten, Ventralausschnitt der Kopfkapsel breiter als
tief, Cervalsklerite m um-Mittelzahn
2 (1’)
it der Kopfkapsel verbunden, Hypostomi
dreigeteilt (Abb. 68 bis 70). Prosimulium
Stirnsklerit an der Basis am breitesten, Ventralausschnitt tiefer als breit,
C ach (Abb. 71
2’ (1’)
ervalsklerite von Kopfkapsel separiert, Hypostomium-Mittelzahn einf
bis 73).
3 Simulium
A z am Abdomen dorsal 3 (2) temfäden dick und darmähnlich gewunden, Hakenkran
entralseite
wesentlich mehr Haken pro Reihe als an der V (Abb. 75).
Simulium (Wilhelmia)
3’ (2) Atemfäden dünn und schneckenartig gewunden (Abb. 74). Simulium
Ab ia Ab ia hydroides
Analsklerit. Prosimulium, Kopf mit
opffächer (Pfeil).
Abb m / Prosimulium, Analsklerit.
b. 64: Twinnhydroides, Kopf ohne
Kopffächer.
b: 65: Twinn Abb. 66: Simulium /
K
. 67: Simuliu
Ventralausschnitt
Abb. 68: Prosimulium, Kopfkapsel von dorsal.
Abb. 69: Prosimulium, Kopfkapsel von ventral.
Abb. 70: Prosimulium; Hypostomium.
Cervalsklerit
Methodenstandardisierung Makrozoobenthos 14

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Methodenstandardisierung Makrozoobenthos 15
bb. 71: Simulium, A
Kopfkapsel von dorsal.
Kopfkapsel von ventral mit
Ventralausschnitt.
Abb. 72: Simulium,
Abb. 73: Simulium, Hypostomium.
Abb. 74: Simulium / Prosimulium,
Atemanlage.
Abb. 75: Simulium (W helmia), Atemanlage.
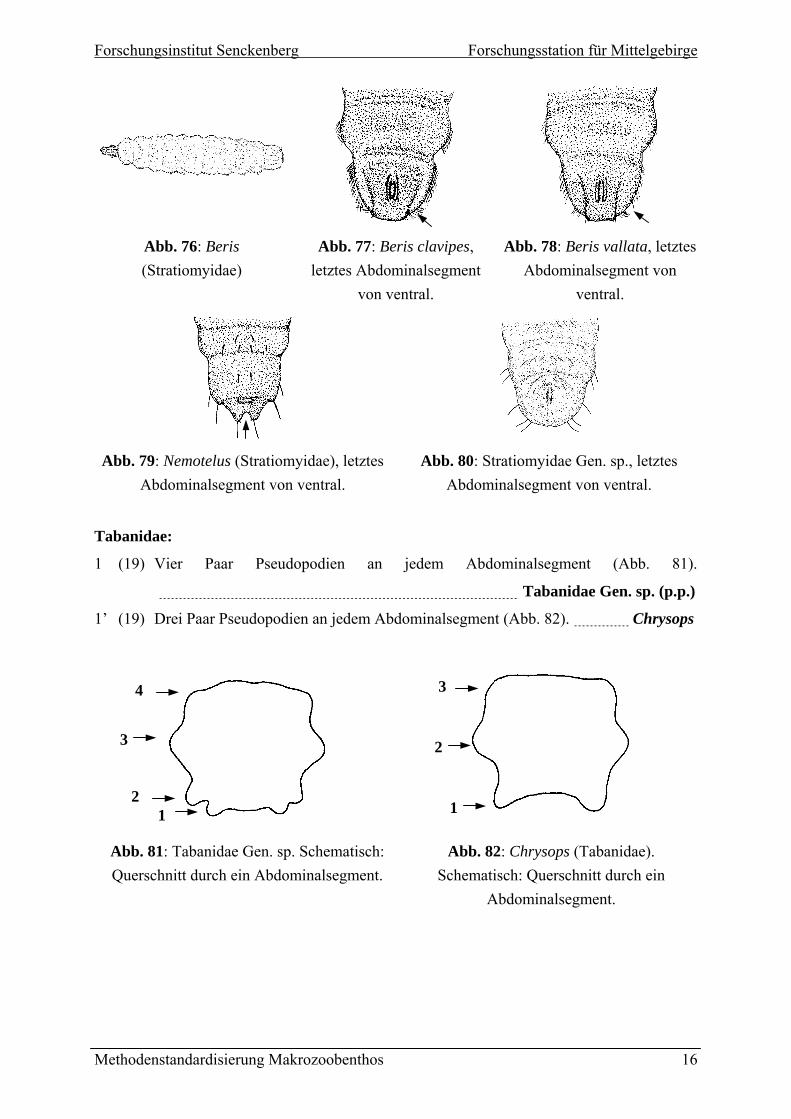
1 (14) rundet und mit charakteristischen Haarsaum (Abb. 78,
il
Stratiomyidae:
Letztes Abdominalsegment ge
79), Habitus wie in Abb. 76. 2
1’ (14) Abdominalsegment anders. 3
R2 (1) andborsten auf dem Analsegment nicht in zwei Gruppen geteilt (Abb. 77).
Beris clavipes
R2’ (1) andborsten auf dem Analsegment in zwei Gruppen geteilt (Abb. 78).
Beris vallata
3 (2’) Randlappen an der Basis eingeschnürt (Abb. 79). Nemotelus
Randlappen langgezogen, rechtwinklig oder gerundet aber ohne Randborsten und
ni cht wie in Abb. 76-79.
3’ (2’)
cht eingeschnürt (Abb. 80), Aussehen ni
Stratiomyidae Gen. sp. (p.p.)
Ventralausschnitt Cervalsklerit
Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 76: Beris (Stratiomyidae)
Abb. 77: Beris clavipes, letztes Abdominalsegment
von ventral.
Abb. 78: Beris vallata, letztes Abdominalsegment von
ventral.
Abb. 79: Nemotelus (Stratiomyidae), letztes Abdominalsegment von ventral.
Abb. 80: Stratiomyidae Gen. sp., letztes Abdominalsegment von ventral.
Tabanidae:
1 (19) Vier Paar Pseudopodien an jedem Abdominalsegment (Abb. 81).
Tabanidae Gen. sp. (p.p.)
1’ (19) Drei Paar Pseudopodien an jedem Abdominalsegment (Abb. 82). Chrysops
Abb. 81: Tabanidae Gen. sp. Schematisch: Querschnitt durch ein Abdominalsegment.
Abb. 82: Chrysops (Tabanidae). Schematisch: Querschnitt durch ein
inalseg
3
2
1
4
21
3
Abdom ment.
Methodenstandardisierung Makrozoobenthos 16

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Tipulidae:
1 (18) Dorsale Randlappen des Stigmenfeldes (letztes Abdominalsegment) eng
nebeneinander stehend, zwei Paar Analpapillen (Abb.83). Dolichopeza
1’ (18) Dorsale Randlappen nicht eng nebeneinander stehend, Randlappen rundlich oder
abgeflacht, z.T. auch reduziert (Abb. 84-85). Tipula
.
Abb. 83: Dolichopeza, Stigmenfeld. Verändert nach
Theowald (1967).
Abb. 84: Tipula, Stigmenfeld mit sechs Randlappen. Ventral mit
drei Paar Analpapillen.
Abb. 85: Tipula, Stigmenfeld mit teilweise
reduzierten Anhängen.
Limoniidae/Pediciidae:
1 (18’) Stigmenfeld mit nur zwei Randlappen (Abb. 88-89). 2
1’ (18’) Stigmenfeld mit mehr als zwei Randlappen. 4
2 (1) Kriechwülste (mit Reihen mikroskopischer Dörnchen besetzt, diese z.T.
hakenförmig) dorsal und ventral an Abdominalsegmenten 2-7 vorhanden (Abb. 86
und 90). Stigmen am letzten Abdominalsegment fehlen. Antocha
2’ (1) Pseudopodien oder Kriechwülste nur ventral an den Abdominalsegmenten
vorhanden. Stigmen am letzten Abdominalsegment vorhanden. 3
3 (2’) Randlappen kurz, Stigmenfeld groß (Abb. 88). Abdominalsegmente mit
Kriechwülsten. Tricyphona
3’ (2’) Randlappen länger, Stigmenfeld klein (Abb. 89). 4
4 (2’) Paarige Pseudopodien mit apikalen Hakenkränzen ventral an Abdominalsegmenten
vorhanden (Abb. 91). Dicranota
4’ (2’) Paarige warzenartige Ausstülpungen ohne apikale Hakenkränze ventral an
Abdominalsegmenten vorhanden (Abb. 92). Pedicia
5 (1’) Stigmenfeld mit ≤ 4 Randlappen. 6
5’ (1’) Stigmenfeld mit 5 Randlappen. 7
Methodenstandardisierung Makrozoobenthos 17

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 86: Antocha Abb. 87: Dicranota
Abb. 88: Tricyphona, letztes
inalsegment von dAbdom orsal.
Abb. 89: Dicranota, letztes
inalsegment von dAbdom orsal.
Abb. 90: Antocha,
Kriechwulst. warzenartige Ausstülpungen. Abb. 91: Dicranota,
Pseudopodien. Abb. 92: Pedicia,
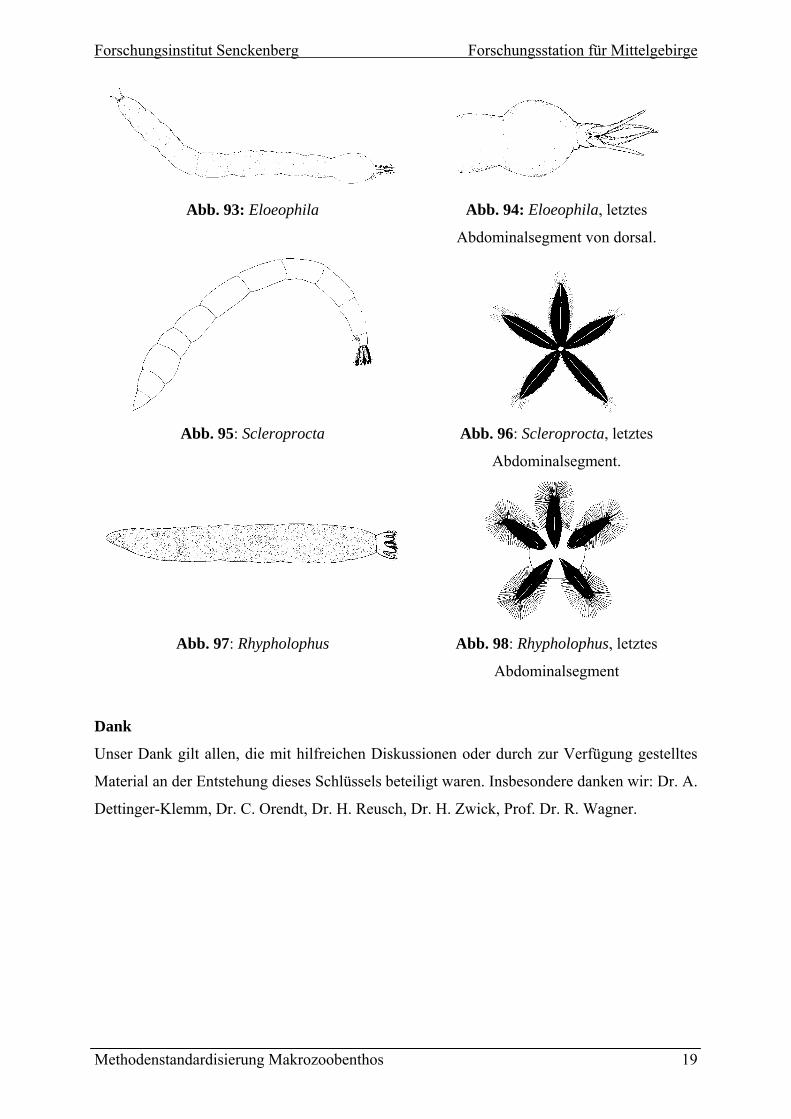
6 (4) Stigmenfeld mit vier Randlappen, alle Randlappen etwa gleich lang, schlank und
fingerförmig (Abb. 94). Abdominalsegmente ohne Kriechwülste, Habitus wie in
Abb. 93. Eloeophila
6’ (4) Randlappen und Stigmenfeld nicht wie in Abb. 94, Merkmalskombination nicht
wie oben beschrieben, teilweise weniger als vier Randlappen ausgebildet.
Kriechwülste können ventral sowie ventral und dorsal vorhanden sein.
Limoniidae Gen. sp. (p.p.)
7 (4’) Fünf Randlappen großflächig schwarz pigmentiert, Randlappen in Form
spatelförmiger Platten mit leicht gezähnten Außenrändern (Abb. 96).
Abdominalsegmente ohne Kriechwülste. Habitus wie in Abb. 95. Scleroprocta
7’ (4’) Randlappen nicht wie in Abb. 96. 8
8 (6’) Fünf Randlappen rundlich und dunkelbraun, Ränder mit charakteristischem
Haarsaum (Abb. 98). Habitus wie in Abb. 97. Rhypholophus
8’ (6’) Ausprägung der Randlappen nicht wie oben beschrieben.
Limoniidae Gen. sp. (p.p.)
Methodenstandardisierung Makrozoobenthos 18
Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Abb. 93: Eloeophila
Abb. 94: Eloeophila, letztes
minalsegment von doAbdo rsal.
Abb. 95: Scleroprocta
Abb. 96: Scleroprocta, letztes
Abdominalsegment.
Abb. 97: Rhypholophus
Abb. 98: Rhypholophus, letztes
Abdominalsegment
Dank
Unser Dank gilt allen, die mit hilfreichen Diskussionen oder durch zur Verfügung gestelltes
Material an der Entstehung dieses Schlüssels beteiligt waren. Insbesondere danken wir: Dr. A.
Dettinger-Klemm, Dr. C. Orendt, Dr. H. Reusch, Dr. H. Zwick, Prof. Dr. R. Wagner.
Methodenstandardisierung Makrozoobenthos 19

Forschungsinstitut Senckenberg Forschungsstation für Mittelgebirge
Methodenstandardisierung Makrozoobenthos 20
Literatur:
FRUTIGER, A.& JOLIDON, C. (2000): Bestimmungsschlüssel für die Larven und Puppen der in
der Schweiz, in Österreich und in Deutschland vorkommenden Netzflügelmücken (Dipte-
ra: Blephariceridae), mit Hinweisen zu ihrer Verbreitung und Phänologie. 4 Abb., 4 Tab. –
Mitteilungen der schweizerischen entomologischen Gesellschaft 73: 93-108, Neuchâtel.
HENNIG, W. (1950): Die Larvenformen der Dipteren. Vol. 2. Berlin.
HENNIG, W. (1952): Die Larvenformen der Dipteren. Vol. 3. Berlin.
THEOWALD, B. (1967): Familie Tipulidae (Diptera, Nematocera) Larven und Puppen. - In:
D'ANGUILAR, J., M. BEIER, H. FRANZ & F. RAW (Hrsg.): Bestimmungsbücher zur Boden-
fauna Europas Lfg. 7, 100 pp., (Akademie-Verlag) Berlin.







