
Aus dem Institut für Pathologie
der Medizinischen Hochschule Hannover
dem
Institut für Pathologie
der Tierärztlichen Hochschule Hannover
und
dem Forschungsbereich Biotechnologie
des Instituts für Tierzucht und Tierverhalten (FAL) Mariensee
Pathomorphologische Phänotypisierung und Expressionsanalyse von hCD59-transgenen Schweinen für Xenotransplantationen
PhD-THESE
Zur Erlangung des Grades eines
DOCTOR OF PHILOSOPHY
(PhD)
durch die Tierärztliche Hochschule Hannover
Vorgelegt von
Stefanie Deppenmeier aus Lügde
Hannover 2005

Wissenschaftliche Betreuung:
1. Gutachter: Prof. Dr. H. H. Kreipe
Prof. Dr. A. D. Gruber, Ph.D
2. Gutachter: Prof. Dr. H. Niemann
3. Gutachter: Prof. Dr. E. Töpfer-Petersen
Externer Gutachter: Prof. Dr. H.-J. Hedrich
Tag der mündlichen Prüfung: 01.06.2005

In Liebe und Dankbarkeit
meiner Familie gewidmet

Vorab öffentlich präsentierte Teilergebnisse dieser Arbeit:
S. DEPPENMEIER, O. BOCK, M. MENGEL, A. D. GRUBER, H. H. KREIPE (2004):
Pathomorphologic and expression pattern analysis of human CD59-transgenic pigs
designed for xenotransplantation.
Abstract Proceedings of the 22nd Meeting of the European Society of Veterinary
Pathology, 15.-18. September, Olsztyn, Polen, S. 72
S. DEPPENMEIER, O. BOCK, M. MENGEL, H. NIEMANN, W. KUES, E. LEMME, D.
WIRTH, K. WONIGEIT, H.H. KREIPE (2005):
Health Status of transgenic pigs expressing the human complement regulatory
protein CD59
Transgenic Research, submitted

Inhaltsverzeichnis 1 Einleitung 1 2 Literaturübersicht 2 2.1 Einleitung 2 2.1.1 Allotransplantation 2
2.1.2 Xenotransplantation – erste Versuche 2
2.1.3 Schweine als Organspender 3
2.2 Probleme und Risiken 4 2.2.1 Zoonosen 4
2.2.2 Transplantatabstoßung 6
2.2.3 Hyperakute Abstoßung 6
2.2.4 Akute Abstoßung 8
2.2.5 Chronische Abstoßung 9
2.3 Herstellung transgener Tiere 10 2.4 Lösungsansätze für die Transplantatabstoßung 11 2.4.1 Hemmung oder Ausschaltung des Gal-Epitops 11
2.4.2 Hemmung der xenoreaktiven Antikörper 13
2.4.3 Akkommodation und Toleranz 13
2.4.4 Hemmung der Komplementkaskade 14
2.4.5 Komplementregulatoren 15
2.5 CD59 und hCD59-transgene Tiere 18 2.5.1 CD59 im Menschen (hCD59) 18
2.5.2 CD59 im Schwein (pCD59) 19
2.5.3 CD59 bei anderen Tierarten 20
2.5.4 hCD59-transgene Tiere 20
2.6 Charakterisierung der Expression in transgenen Schweinen 21 2.6.1 Expression des hCD59-Transgens 22
2.6.2 Expression von mehreren Transgenen 24
2.6.3 Weitere Transplantationsversuche 26

2.7 Morphologische Phänotypen transgener Tiere 28 2.7.1 Ektopische Expression 29
2.7.2 Large Offspring Syndrome (LOS) 29
2.7.3 Kardiopulmonale und plazentale Phänotypen 30
2.7.4 Muskel- und Skelett-Phänotypen 31
2.7.5 Sonstige Phänotypen 31
2.8 Genkonstrukt 32 3 Ziel der Studie 34 4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 35 4.1 Hintergrund und Bedeutung 35 4.2 Experimentelles Design 35 4.2.1 Material 35
4.2.2 Pathologisch-anatomische und histopathologische Beurteilung 38
4.2.3 Virologische Untersuchung 38
4.3 Ergebnisse und Interpretation 39 4.3.1 Virologische Untersuchung 39
4.3.2 Pathomorphologische Untersuchung 39
5 Eigene Ergebnisse: hCD59 - Proteinexpression 46 5.1 Hintergrund und Bedeutung 46 5.2 Experimentelles Design 46 5.2.1 Material 46
5.2.2 Herstellung der Multiblöcke 47
5.2.3 Durchführung der immunhistochemischen Färbung 47
5.2.4 Auswertung 49
5.3 Ergebnisse und Interpretation 49 5.3.1 hCD59-Proteinexpression 49

6 Eigene Ergebnisse: hCD59 - mRNA-Expression 57 6.1 Hintergrund und Bedeutung 57 6.2 Experimentelles Design 57 6.2.1 Material 57
6.2.2 Durchführung der RNA-Isolierung 58
6.2.3 Durchführung der cDNA-Synthese 59
6.2.4 Auswahl der Primer 60
6.2.5 Durchführung der Reversen Transkriptase – PCR 60
6.2.6 Auswertung 62
6.3 Ergebnisse und Interpretation 62 6.3.1 EF1α – mRNA – Expression 62
6.3.2 hCD59 – mRNA – Expression 63
6.3.3 Vergleich hCD59 – Protein und mRNA – Expression 67
7 Eigene Ergebnisse: hCD59 – DNA – Detektion 69
7.1 Hintergrund und Bedeutung 69 7.2 Experimentelles Design 69 7.2.1 Material 69
7.2.2 Durchführung der DNA – Isolierung 70
7.2.3 Auswahl der Primer 71
7.2.4 Durchführung der PCR 72
7.2.5 Auswertung 73
7.3 Ergebnisse und Interpretation 74 7.3.1 TNF – Expression 74
7.3.2 hCD59 – DNA – Expression 74
7.3.3 Vergleich hCD59 – Protein, mRNA und DNA 75
8 Diskussion 77 9 Zusammenfassung 91 10 Summary 93

11 Literaturverzeichnis 95 12 Anhang 123 13 Eidesstattliche Erklärung 127 14 Danksagung 129
Verzeichnis der in Text und Bild verwendeten Abkürzungen Abb. Abbildung
AVR Acute Vascular Rejection (Akute Abstoßung)
BALT Bronchus-assoziiertes lymphatisches Gewebe
bp Basenpaare
cDNA komplementäre Desoxyribonukleinsäure
CMV Cytomegalovirus
CVF Cobra Venom Factor
DAF Decay-accelerating Factor
DIC Disseminierte Intravasale Koagulopathie
DNA Desoxyribonukleinsäure
DXR Delayed Xenograft Rejection (Akute Abstoßung)
EF1α Elongation Factor 1α
EGFP Enhanced Green Flourescent Protein
Gal Galactose α1-3Galactose
HAR Hyperacute Rejection (Hyperakute Abstoßung)
HE Hämatoxylin-Eosin
HT H-Transferase / α 1-2 Fucosyltransferase
H2Kb murines MHC Klasse 1 Gen
ICAM-2 interzelluläres Adhäsionsmolekül-2
IRES Internal Ribosome Entry Side
KO nicht transgenes Kontrolltier
LCR Locus Control Region
Ln./Lnn. Lymphknoten (Singular/Plural)
LOS Large Offspring Syndrome
MAC Membrane attack complex
männl. männlich
MCP Membrane Cofactor Protein
MHC Major Histocompatibility Complex
MIEP Major Immediate-Early Promoter

mRNA messenger Ribonukleinsäure
NTA Transaktivator
PAA Polyacrylamid
PCR Polymerase Kettenreaktion
PCV II Porzines Circovirus Typ II
PDNS Porzines Dermatitis und Nephropathie Syndrom
PERV Porzine Endogene Retroviren
PNP Proliferative und Nekrotisierende Pneumonie
PNH Paroxysmale Nächtliche Hämoglobinurie
PMWS Postweaning Multisystemic Wasting Syndrome
PRRSV Porzines respiratorisches und reproduktives Syndrom-Virus
RNA Ribonukleinsäure
RT-PCR Reverse Transkriptase - Polymerase Kettenreaktion
sCR1 löslicher Komplementrezeptor Typ 1
SPF spezifiziert pathogenfrei
Tab. Tabelle
TG transgenes Tier
TNF Tumornekrosefaktor
weibl. weiblich
XNA Xenoreactive Natural Antibodies

1
1 Einleitung
Der Fortschritt auf dem Gebiet der Transplantationschirurgie in den letzten
Jahrzehnten und der erhöhte Bedarf nach Transplantaten, haben in den letzten
Jahren zu einem Mangel an Spenderorganen geführt. Der wachsende Bedarf an
geeigneten Transplantaten hat in jüngster Zeit verstärkt zu einer Diskussion über die
Nutzung von tierischen Xenotransplantaten zum Zweck der Transplantation in
menschliche Patienten geführt. Auf der Suche nach einer geeigneten Spezies rückte
aufgrund eines niedrigen Zoonose-Risikos, relativ hoher Reproduktionsraten, zu
humanen Organen vergleichbarer Organgrößen und relativ geringer ethischer
Bedenken das Schwein in den Mittelpunkt der Diskussion.
Das größte Problem der Xenotransplantation stellt derzeit die hyperakute Abstoßung
der Transplantate dar, die innerhalb von Minuten zu einer Zerstörung des
Spenderorgans führt und im Wesentlichen auf der Komplementreaktion beruht. Um
das Problem der hyperakuten Abstoßung zu lösen, wurden transgene Schweine
erzeugt, die in ihrem Genom humane Komplementregulatoren tragen, um die
Komplementreaktion zu unterdrücken und damit die hyperakute Abstoßungsreaktion
zu unterbinden. Da die transgenen Schweine mangels besserer verfügbarer
Methoden über eine Mikroinjektion des Genkonstruktes, welches den humanen
Komplementregulator hCD59 beinhaltet, in den Vorkern einer befruchteten Zygote
erzeugt wurden, ist nicht sicher, ob das Genkonstrukt überhaupt in das porzine
Genom integriert wird und ob es zu einer Expression führt.
Ziel dieser Arbeit war es, zu klären, ob diese transgenen Schweine durch die
Insertion des humanen Genkonstruktes einen morphologisch manifesten Phänotyp
entwickeln und ob die Expression des Komplementregulators hCD59 auf DNA-
Ebene, mRNA-Ebene und Protein-Ebene in diversen Organen und Geweben erfolgt.
Diese Ergebnisse sollten einen wesentlichen Beitrag leisten zur Einschätzung der
Eignung von Organen dieser Tiere als Xenotransplantate für den Menschen.

2
2 Literaturübersicht
2.1 Einleitung 2.1.1 Allotransplantation Transplantationen zwischen Individuen der gleichen Spezies, also z.B. von Mensch
zu Mensch werden als Allotransplantationen bezeichnet. Auf diesem Gebiet sind in
den letzten Jahrzehnten beträchtliche Fortschritte gemacht worden. Von den
Patienten, die Allotransplantate erhalten, überleben, abhängig vom transplantierten
Organ, im Schnitt 85% für ein Jahr, 65% für 5 Jahre und 50% für 10 Jahre (COOPER
et al., 2002). Diese Erfolge führten zu einem erheblich gesteigerten Bedarf an
Spenderorganen. In den USA hat sich die Zahl der Menschen, die auf ein
Spenderorgan warten, im Zeitraum von 1990 bis 1999 von 21.914 auf 72.110
verdreifacht (UNITED NETWORK FOR ORGAN SHARING, 2000). In diesem
Zeitraum stieg die Zahl der Spenderorgane jedoch nur von 15.009 auf 21.175 an.
Während in den USA täglich 60 Patienten ein Spenderorgan erhalten, sterben täglich
17 Patienten, die auf der Warteliste für ein Spenderorgan stehen.
Dieses unausgeglichene Verhältnis zwischen dem Bedarf an Spenderorganen und
den zur Verfügung stehenden Organen hat in den letzten Jahren zu einer verstärkten
Suche nach einer alternativen Spenderorganquelle geführt.
2.1.2 Xenotransplantation – erste Versuche Die Xenotransplantation, also die Transplantation von Organen zwischen Individuen
verschiedener Spezies, stellt eine mögliche Alternative zur Allotransplantation dar.
Bereits im frühen 17. Jahrhundert wurden Bluttransfusionen von Tieren auf
Menschen durchgeführt (COOPER et al., 2002) und im 19. Jahrhundert wurden
diverse Organe, insbesondere Haut von Tieren, in Menschen transplantiert. Auch im
20. Jahrhundert wurden wiederholt Xenotransplantationen durchgeführt, doch bis auf
ein oder zwei Ausnahmen scheiterten all diese Versuche (TANIGUCHI und
COOPER, 1997). Die bemerkenswertesten Erfolge wiesen Reemtsma und seine

2 Literaturübersicht 3
Kollegen 1964 auf, die Schimpansennieren in Dialysepatienten transplantierten. Die
Überlebensraten der Empfänger lagen zwischen 11 Tagen und 2 Monaten. Ein
Patient überlebte sogar 9 Monate mit der Schimpansenniere und starb an einer
Elektrolytstörung, ohne dass die transplantierte Niere Anzeichen einer Abstoßung
aufwies (REEMTSMA et al., 1964). CZAPLIKI und BLONSKA transplantierten 1992
ein nicht-transgenes Schweineherz in einen Mann mit einer Herzinsuffizienz in Folge
eines Marfan-Syndroms, da sich der Zustand des Patienten akut verschlechterte.
Das Schweineherz arbeitete zunächst normal, doch nach knapp 24 Stunden verstarb
der Mann. Dabei zeigte das Herz keinerlei Abstoßungsreaktion, sondern erbrachte
lediglich nicht den nötigen Blutauswurf.
Insgesamt waren die Erfolge auf dem Feld der Xenotransplantation jedoch eher
bescheiden und fast alle Organe wurden von den Transplantatempfängern
abgestoßen.
2.1.3 Schweine als Organspender Wenn es um die Frage geht, welche Spezies sich für die Xenotransplantation eignen,
wird die Auswahl dadurch eingeschränkt, dass lediglich eine geringe Anzahl an
Tierarten die erforderlichen Organgrößen und die geeigneten anatomischen
Merkmale aufweisen. Weiterhin ist die Überlebenszeit der transplantierten Organe
umso kürzer, je größer die phylogenetische Distanz zwischen den beiden Spezies ist
(CHALINE et al., 1994). Aufgrund der Tatsache, dass nicht-menschliche Primaten,
insbesondere Altweltaffen, zoologisch den Menschen am nächsten stehen, wurden
sie zunächst als Organspender in Betracht gezogen. Es gibt jedoch eine Reihe von
Gründen, die gegen diese Primaten sprechen:
Zunächst einmal sind viele Primatenarten vom Aussterben bedroht und daher
geschützt. Eine gezielte Zucht von Primaten zum Zweck der Xenotransplantation
wäre auf Grund der langen Reproduktionsintervalle und der benötigten intensiven
Pflege teuer und aufwendig und wird zudem in der Bevölkerung häufig als unethisch
betrachtet. Ein weiterer, sehr wichtiger Grund ist, dass das Risiko der Übertragung
von Zoonosen, wie z.B. Hepatitis oder Herpes auf Grund der nahen
phylogenetischen Verwandschaft zu den Menschen sehr hoch ist. Zudem liegt die

2 Literaturübersicht 4
Organgröße der meisten Affenarten unter der Organgröße von adulten,
menschlichen Organen, so dass fraglich ist, inwiefern eine vollständige Funktion
gewährleistet wäre (HAMMER et al., 1998).
Aus diesen Gründen wurde von den nicht-humanen Primaten als potentielle
Organspender wieder Abstand genommen und das Schwein rückte in den
Mittelpunkt. Die Organe von Schweinen und Menschen haben in etwa die gleiche
Größe und vergleichbare anatomische und physiologische Merkmale (HAMMER et
al., 1998). Die Zucht von Schweinen zum Zweck der Organspende wäre auf Grund
der recht kurzen Reproduktionsintervalle und der komplikationslosen Aufzucht
einfacher zu realisieren. Zudem werden Schweine als Organspender in der
Bevölkerung ethisch erheblich besser akzeptiert, da Schweine auch als
lebensmittelliefernde Tiere gezüchtet werden. Nicht zuletzt ist die Gefahr der
Übertragung von Zoonosen auf den Menschen bei Schweinen erheblich geringer,
wenn auch nicht gänzlich gebannt (HAMMER et al., 1998).
All diese Gründe sprechen für das Schwein als potentiellen Organspender.
Im Folgenden sollen die Probleme und Risiken der Xenotransplantation im
Allgemeinen und des Schweins als Organspender im Besonderen dargestellt
werden.
2.2 Probleme und Risiken 2.2.1 Zoonosen Auch wenn die Gefahr der Übertragung von Zoonosen auf den Menschen beim
Schwein geringer ist als bei nicht-humanen Primaten, gibt es dennoch eine Reihe
von Krankheitserregern, die eine Gefahr für den potentiellen Organempfänger
darstellen. Dies gilt insbesondere, wenn man in Betracht zieht, dass die
Organempfänger eine immunsuppressive Therapie erhalten und dadurch noch
anfälliger für Krankheitserreger jeglicher Art sind.
Bei den Zoonosen kommt den tierartspezifischen bakteriellen, mykotischen und
parasitären Infektionen eher eine untergeordnete Bedeutung zu. Hier läge das Risiko

2 Literaturübersicht 5
am ehesten bei allgemeinen Infektionen im Rahmen einer Parasitose, einer
systemischen Mykose oder einer bakteriellen Sepsis mit relativ unspezifischen
Erregern wie z.B. Streptokokken oder Staphylokokken.
Das größte Risiko stellt eine Übertragung von viralen Erregern auf den Menschen
dar. Das Virus, das in den letzten Jahren am meisten für Aufsehen gesorgt hat, ist
das Porzine Endogene Retrovirus (PERV). Die Porzinen Endogenen Retroviren sind
für Schweine apathogene Viren, die in der porzinen DNA integriert sind und daher in
allen Schweinen nachweisbar sind. Die virale RNA wird in diversen
transplantationsrelevanten Organen, wie z.B. Herz, Lunge, Leber, Niere, Haut und
Pankreasinselzellen exprimiert (BLUSH et al., 2002). LANGFORD et al. zeigten 2001
eine Expression von PERV-RNA in allen untersuchten Organen von transgenen
Schweinen, wobei die Expressionslevel zwischen den einzelnen Organen sehr
differierten. Allerdings konnten sie keine Freisetzung von Virionen nachweisen.
Mittlerweile haben einige Arbeitsgruppen eine Infektion mit PERV von humanen
Zelllinien in vitro nachweisen können (LE TISSIER et al., 1997; WILSON et al., 1998;
MARTIN et al., 2000; SPECKE et al., 2001). Zudem entdeckten ERICSSON et al.
2003 humane Proteine, die als Rezeptoren für PERV funktionieren. NOD/SCID-
Mäuse (non-obese diabetic/ severe combined immunodeficiency), denen porzine
Pankreasinselzellen transplantiert wurden, zeigten eine Infektion mit PERV in
diversen Organen. OGLE et al. zeigten 2004 eine Fusion von Menschen- und
Schweinezellen nach einer Infusion von humanen hämatopoietischen Stammzellen
in Schweineuteri. Die Hybridzellen enthielten DNA von beiden Spezies und
exprimierten eine Mischung aus porzinen und humanen Proteinen. Die Hybridzellen
enthielten zudem PERV und waren auch in der Lage, PERV in einer Co-Kultur auf
rein humane Zellen zu übertragen. Diese Ergebnisse zeigen, dass PERV eine
ernstzunehmende Gefahr für Empfänger von Schweineorganen darstellen, wenn
auch eine Infektion von Menschen die porzine Gewebetransplantate, wie z.B.
Herzklappen erhalten haben, bislang nicht nachgewiesen werden konnte (HENEINE
et al., 1998; HERRING et al., 2001).
Aber auch andere virale Erreger, wie z.B. das porzine Hepatitis Virus E (YOO und
GIULIVI, 2000), das Porzine Cytomegalovirus (CLARK et al., 2003) oder die

2 Literaturübersicht 6
Porzinen Circoviren Typ I und II (HATTERMANN et al., 2004) stellen mögliche
Zoonoserisiken dar.
Um die Infektionsgefahr so gering wie möglich zu halten, müssten die Schweine
strengen Kontrollen unterliegen und vor einer Organspende auf alle möglichen
Zoonoseerreger untersucht werden. In der gängigen Literatur existieren bereits
Vorschläge für Hygiene-Protokolle, die einer Xenotransplantation vorausgehen
sollten, um das Zoonoserisiko so gering wie möglich zu halten (ONIONS et al.,
2000).
2.2.2 Transplantatabstoßung Das größte Problem der Xenotransplantation ist die Transplantatabstoßung. Je nach
Dauer bis zur Abstoßung werden die Abstoßungsreaktionen als „hyperakute
Abstoßung“ (Hyperacute Rejection, HAR), „akute Abstoßung“ (Acute Vascular
Rejection, AVR / Delayed Xenograft Rejection, DXR) und „chronische Abstoßung“
bezeichnet.
Aufgrund der phylogenetischen Distanz zwischen Schweinen und Menschen wird
das porzine Transplantat vom Menschen hyperakut abgestoßen.
2.2.3 Hyperakute Abstoßung Die hyperakute Abstoßung (HAR) beschreibt die Abstoßung eines Spenderorgans
innerhalb von Minuten bis Stunden (COOPER et al., 1996).
Die hyperakute Abstoßung wird dadurch eingeleitet, dass präexistente, xenoreaktive
Antikörper an Epitope auf der Oberfläche des Spenderorgans binden. Das Epitop,
das in der Xenotransplantation die größte Rolle spielt, ist ein α-galactosyl-Epitop,
gegen das spezifische Antikörper gerichtet sind (GALILI et al., 1984). Genauer
handelt es sich bei diesem Epitop um den von der α 1-3-Galactosyltransferase
synthetisierten Zucker Galactose α 1-3Galactose, welcher spezifische anti-Gal-
Antikörper bindet. Das porzine α 1-3-Galactosyltransferase-Gen besteht aus
mindestens 10 Exons. 4 verschiedene Promotoren kontrollieren die Transkription des
Gens in porzinen Zellen (MERCIER et al., 2002). Die spezifischen anti-Gal-
Antikörper sind in Menschen und Altweltaffen in hohen Titern präexistent vorhanden,

2 Literaturübersicht 7
da diese Spezies das Galactose α 1-3Galactose-Epitop nicht exprimieren (GALILI et
al., 1987). Neuweltaffen und andere Säuger exprimieren dagegen das Galactose α 1-
3Galactose-Epitop (GALILI et al., 1988). Somit bietet dieses Epitop auf dem Endothel
der Schweineorgane eine geeignete Angriffsstelle für die im Menschen präexistenten
anti-Gal-Antikörper. Der Grund für die kontinuierliche anti-Gal-Antikörperproduktion in
Menschen und Altweltaffen sind Gal α 1-3Gal-Epitope in Außenmembranen von
Bakterien, die im Intestinaltrakt von Menschen und Altweltaffen vorkommen und
damit einen ständigen Stimulus zur Antikörperproduktion darstellen (GALILI et al.,
1988). Die Theorie der Bindung von Anti-Gal-Antikörpern an das Gal α 1-3Gal-Epitop
konnte auch bereits in Transplantationsversuchen bewiesen werden (COLLINS et al.,
1995). Im Schwein wird das Gal α 1-3Gal-Epitop auf dem Gefäßendothel in allen
Organen exprimiert (DOR et al., 2004).
Bei den xenoreaktiven Antikörpern handelt es sich vorwiegend um IgM-Antikörper
(SANDRIN et al., 1993), die zu 80% spezifisch für die terminalen α-Galactose-
Determinanten sind (PARKER et al., 1994). Zudem gibt es eine kleine Subgruppe
von Antikörpern, die nicht gegen das Gal α 1-3Gal-Epitop gerichtet sind und daher
auch, wenn auch nur zu einem geringen Teil, zur hyperakuten Abstoßung beitragen
(ZHU, 2000).
Nachdem die xenoreaktiven IgM-Antikörper an die Gal α 1-3Gal-Epitope auf dem
Endothel gebunden haben, wird dadurch die Aktivierung der Komplementkaskade,
und zwar des klassischen Weges, eingeleitet. Nach Xenotransplantationsversuchen
konnten auf dem Endothel des Spenderorgans immunhistochemisch sowohl
Komponenten der Komplementkaskade als auch IgM nachgewiesen werden (PLATT
et al., 1991; DALMASSO et al., 1992). Weiterhin korrelierte die Lokalisation der Gal α
1-3Gal-Epitope mit der Lokalisation von IgM- und Membrane attack complex (MAC)-
Deposition (CHEN et al., 1999). Der MAC ist das Endprodukt der
Komplementkaskade und führt zu einer Endothelzellaktivierung und -schädigung und
einer anschließenden Thrombozytenaggregation mit einer Ausbildung von
Fibrinthromben und Infiltration von neutrophilen Granulozyten (PLATT et al., 1991).
Die Abstoßung des Spenderorgans erfolgt über eine Endothelzellschädigung und

2 Literaturübersicht 8
Thrombosierung der Gefäße mit einer anschließenden ischämischen Infarzierung
des Organs (ROSE und COOPER, 2000).
Histopathologisch sind Spenderorgane, die hyperakut abgestoßen wurden,
gekennzeichnet durch massive interstitielle Blutungen und Ödeme sowie
Thrombosen mit einer konsekutiven Zerstörung des Parenchyms (ROSE et al., 1991;
SCHUURMAN et al., 2003).
2.2.4 Akute Abstoßung Nach Überwindung der hyperakuten Abstoßungsreaktion stellt die akute Abstoßung
die zweite und mittlerweile bedeutendste Hürde der Xenotransplantation dar. Die
akute Abstoßung wird in der Literatur mit den zwei Termini „Delayed Xenograft
Rejection“ (DXR) oder „Acute Vascular Rejection“ (AVR) bezeichnet (COOPER et al.,
1996). Diese Art der Abstoßung geschieht zumeist nach einigen Tagen bis Wochen.
Die akute Abstoßung stellt eine humorale Immunantwort dar, bei der vor allem IgM-
und IgG-Antikörper gegen das Gal-Epitop eine große Rolle spielen (LIN et al., 1998).
Im Fall der akuten Abstoßung handelt es sich dabei jedoch nicht um präexistente,
xenoreaktive Antikörper, sondern um induzierte Antikörper, die wiederum zur
Aktivierung der klassischen Komplementkaskade führen (LOSS et al., 2000). Es
kommt zu einer Aktivierung der Endothelzellen mit einer verstärkten Expression von
Adhäsionsmolekülen und Produktion von Cytokinen. Monozyten und natürliche
Killerzellen infiltrieren das Spenderorgan und es kommt zu einer
Entzündungsreaktion und Thrombose (BACH et al., 1996). ASHTON-CHESS et al.
konnten 2003 zusätzlich zu der humoralen Immunantwort eine zelluläre Reaktion
nachweisen, indem sie in abgestoßenen Xenotransplantaten Monozyten und
Makrophagen sowie CD8+ und CD4+ T-Lymphozyten nachwiesen. Zudem wurden
Interleukin 1α, Tumor-Nekrosefaktor α, Interferon γ und Granzym B nachgewiesen.
IERINO et al. zeigten 1998, dass systemische Koagulationsstörungen zu einer
disseminierten intravaskulären Gerinnung (DIC) in dem Spenderorgan führen. Dabei
kommt es zunächst zu einer Zerstörung von Kapillaren und Ausbildung einer so
genannten thrombotischen Mikroangiopathie (SHIMIZU et al., 2000). Insgesamt stellt
die akute Transplantatabstoßung ein multifaktorielles Geschehen dar, bei dem

2 Literaturübersicht 9
natürliche Killerzellen, Monozyten, Thrombozyten und die Aktivierung des
Gerinnungssystems eine Rolle spielen.
Histologisch ist die akute Abtoßung gekennzeichnet durch großflächige
Parenchymnekrosen und Infarkte mit einzelnen Blutungen sowie durch massive
intravaskuläre Fibrinthromben. Sowohl die Infarkte als auch die Fibrinthromben
zeigen dabei verschiedene Organisationsstadien, was für eine langsame Entwicklung
der Abstoßung spricht (GODDARD et al., 2002). Etwa 24 Stunden nach der
Reperfusion des Organs sind erste Veränderungen in Form von Fibrindepositionen,
fibrinösen Mikrothromben und eine Verdickung des Endothels zu beobachten. Als
nächstes kommt es zu fokalen Ischämien, die häufig mit einer Infiltration von
Entzündungszellen in Gefäßwände und in das Interstitium assoziiert sind. In den
darauf folgenden Tagen entwickeln sich aus den Ischämien großflächige Nekrosen
mit Entzündungszellinfiltraten (PLATT et al., 1998).
2.2.5 Chronische Abstoßung Die chronische Abstoßung bezeichnet die Abstoßung des Spenderorgans nach
Wochen, Monaten oder Jahren. In der Xenotransplantation spielt sie bislang keine
große Rolle, da das Problem der akuten Abstoßung noch nicht überwunden ist, und
wird daher lediglich bei Knorpeltransplantaten beobachtet. Auch bei der chronischen
Abstoßung spielt das Gal-Epitop eine entscheidende Rolle. Das Epitop stimuliert B-
Lymphozyten zur Produktion von anti-Gal-IgG, die an die Xenotransplantat-Zellen
binden (GALILI et al., 1997) und zu einer zellulären Immunantwort von T-
Lymphozyten und Makrophagen führen (STONE et al., 1998). Nach der Entfernung
der Gal-Epitope aus den Knorpeltransplantaten wurden zwar keine anti-Gal-
Antikörper mehr gebildet, dafür kam es jedoch zur Produktion von Antikörpern, die
spezifisch gegen den porzinen Knorpel gerichtet waren. Diese Antikörper führten
jedoch im Vergleich zu den anti-Gal-Antikörpern zu einer geringeren
Entzündungsreaktion (STONE et al., 1998).

2 Literaturübersicht 10
2.3 Herstellung transgener Tiere
Die Biotechnologie bietet vermehrt Möglichkeiten, um Organe und Gewebe von
Tieren gezielt und effektiv für den Menschen zu nutzen. Dabei kommen
verschiedene biotechnologische Techniken, wie z.B. das Klonen oder die Insertion
fremder Gene, zum Einsatz.
Während die gezielte Manipulation des Erbgutes durch homologe Rekombination
und das Klonen bei Mäusen seit Jahren etabliert ist, ist diese Technik bei den großen
Haussäugetieren wie Rindern oder Schweinen schwierig durchzuführen, da die
Nutzung embryonaler Stammzellen bei diesen Spezies nicht vollständig etabliert ist
(CAPECCHI, 2000).
Um die Abstoßung der Schweineorgane in der Xenotransplantation zu verhindern,
werden Schweine generiert, die menschliche Gensequenzen enthalten, die eine
Abstoßung hemmen sollen. Wird speziesfremde DNA in ein Genom eingefügt, so
spricht man von der Herstellung transgener Individuen. Die Herstellung von
transgenen Tieren erfolgte bislang über eine Mikroinjektion der fremden DNA in den
Vorkern einer befruchteten Zygote. Die Ausbeute bei diesem Verfahren ist sehr
gering, die Integration der fremden DNA-Sequenz in das Genom geschieht rein
zufällig, und das Expressionsmuster für das fremde Protein ist sehr variabel
(NIEMANN und KUES 2000).
Die Nutzung von Klonen oder der homologen Rekombination könnte die Effektivität
der Genmanipulation in Haussäugetieren erheblich verbessern. Doch auch wenn
Schweine (ONISHI et al., 2000) und andere Haussäuger (CAMPBELL et al., 1996;
CIBELLI et al., 1998) bereits erfolgreich geklont wurden, wird diese Technik noch
nicht routinemäßig eingesetzt. Insgesamt ist die reproduktive Biotechnologie beim
Schwein auch noch nicht so weit fortgeschritten wie bei Rindern oder Schafen
(NIEMANN und RATH, 2001).
Zusätzlich zu der Herstellung transgener Schweine durch Mikroinjektion ist es in den
letzten Jahren durch eine Kombination von homologer Rekombination und Klonen
gelungen, Tiere zu produzieren, bei denen das Hauptepitop der hyperakuten
Abstoßung ausgeknockt ist (LAI et al., 2002; RAMSOONDAR et al., 2003).

2 Literaturübersicht 11
Die Herstellung von Knockout-Tieren könnte in Kombination mit dem Einfügen
transgener Gene in den nächsten Jahren zu einem großen Fortschritt auf dem
Gebiet der Xenotransplantation führen. Dabei wird vor allem die Fähigkeit, porzine
Gene spezifisch zu modifizieren, im Vordergrund stehen (PIEDRAHITA und MIR,
2004).
2.4 Lösungsansätze für die Transplantatabstoßung
Um die Abstoßung der Transplantate zu verhindern, wurden bislang verschiedenste
Methoden angewandt. Hier soll ein kurzer Überblick über dese Methoden gegeben
werden. Da die hyperakute Abstoßung in der Xenotransplantation zunächst das
größte Problem darstellt, zielen die Lösungsansätze zunächst auf die Überwindung
dieser Art der Abstoßung.
2.4.1 Hemmung oder Ausschaltung des Gal-Epitops Da das Gal-Epitop den initialen Angriffspunkt der xenoreaktiven Antikörper darstellt,
zielen viele Methoden darauf ab, dieses Epitop zu hemmen oder auszuschalten.
Eine Methode zu diesem Zweck ist die Infusion von synthetischen Oligosacchariden,
wie z.B. Galα1-3Galβ1-4GlcNAc oder Melibiose (GALILI et al., 1996). Die
synthetischen Oligosaccharide sollen die gegen das Gal-Epitop gerichteten
Antikörper binden und damit eine Bindung an das Endothel des Spenderorgans
verhindern. In Transplantationsversuchen konnte gezeigt werden, dass die
intravenöse Infusion von Galα1-3Gal Oligosacchariden eine hyperakute Abstoßung
verhindert (SIMON et al., 1998).
Ein weiter Ansatz ist der Verdau des terminalen α-Galactosylrests mit Hilfe eines
Enzyms (Glycosidase). Dadurch können die Antikörper das Epitop nicht mehr
erkennen und es kommt zu einer massiv reduzierten Bindung von XNAs
(LAVECCHIO et al., 1995; WATIER et al., 1996).
Ein weiterer Ansatz ist der Ersatz des Gal-Epitops durch das sogenannte H-Antigen,
welches durch α1-2Fucosyltransferase (H-Transferase/HT) synthetisiert wird. Die

2 Literaturübersicht 12
Galactosyltransferase, die das Gal-Antigen synthetisiert, und die Fucosyltransferase
konkurrieren um die gleichen Substrate. Tiere, die transgen für die
Fucosyltransferase sind, produzieren daher weniger Gal-Epitope (SHARMA et al.,
1996).
Auch eine Herstellung von Mäusen und Schweinezellen, die transgen für α1-
2Fucosyltransferase und für α-Galactosidase waren, zeigte eine massiv reduzierte
Antikörperbindung (OSMAN et al., 1997). Dabei konkurriert die Fucosyltranferase mit
der Galactosyltransferase um Substrate, während die α-Galactosidase den
terminalen Galactosylrest des Gal-Epiops spaltet.
In den letzten Jahren wurde durch Fortschritte auf dem Gebiet der Biotechnologie
auch die Herstellung von Schweinen, bei denen das Gal-Epitop ausgeknockt ist,
möglich. Da bei Mäusen schon länger die Möglichkeit der homologen Rekombination
besteht, gelang es TEARLE et al. bereits 1996, eine α-1-3-Galactosyltransferase
knockout Maus zu erzeugen, die durch das fehlende Enzym das Gal-Epitop nicht
mehr exprimieren konnte. 2002 berichteten LAI et al. zum ersten Mal über die
Herstellung eines Schweins, bei dem ein Allel der α-1-3-Galactosyltransferase
ausgeknockt war. Auch die Arbeitsgruppe von DAI et al. generierte 2002 fünf Ferkel,
die nur ein Allel der α-1-3-Galactosyltransferase besaßen. 2003 wurden von
PHELPS et al. vier Schweine mit einem α-1-3-Galactosyltransferase Doppelknockout
produziert.
2003 gelang RAMSOONDAR et al. zudem das Klonen eines Ferkels, bei dem ein
Allel der α-1-3-Galactosyltransferase ausgeknockt ist und zusätzlich noch α-1-
2Fucosyltransferase synthetisiert wird.
Durch weitere Zucht mit den existierenden Gal-knockout-Tieren sollen in den
nächsten Jahren Linien mit Schweinen entstehen, die das Gal-Epitop nicht mehr
exprimieren. Expressionsanalysen und Transplantationsversuche müssen dann
klären, wie effektiv dieser Genotyp die Abstoßung von Xenotransplantaten hemmt.

2 Literaturübersicht 13
2.4.2 Hemmung der xenoreaktiven Antikörper Ein anderer Ansatz zur Überwindung der hyperakuten Abstoßung ist die Hemmung
der präexistenten, xenoreaktiven Antikörper (XNAs).
Besonders in den ersten Jahren der Xenotransplantation wurde eine Hemmung der
XNAs durch eine Perfusion von diversen Schweineorganen mit dem Blut der
Empfänger erreicht. Dabei verblieben die Antikörper auf dem Endothel der
perfundierten Organe und im Blut der Empfänger wurde so eine Reduzierung der
XNAs erreicht (FISCHEL et al., 1990).
Eine mittlerweile häufiger verwendete Methode zu diesem Zweck ist die
Immunadsorption von Antikörpern mittels einer spezifischen Galα1-3Gal-
Immunaffinitätssäule (TANIGUCHI et al., 1996; XU et al., 1998). Diese Methode
führte auch ohne Immunsuppression der Empfänger zur Überwindung der
hyperakuten Abstoßung (MANEZ et al., 2000). Jedoch erreichten die Antikörper nach
einigen Tagen wieder ihre Ausgangswerte und führten so zu einer akuten
Abstoßung.
Weiterhin besteht die Möglichkeit, die Produktion der anti-Gal-Antikörper
unspezifisch zu verhindern. Dies geschieht zumeist durch eine
Ganzkörperbestrahlung, Knochenmarkbestrahlung, Splenektomie oder durch eine
medikamentelle Immunsuppression (ALWAYN et al., 1999).
Mit all diesen Methoden konnte bisher zwar eine hyperakute, nicht jedoch eine akute
Abstoßungsreaktion verhindert werden. In den meisten Fällen werden diese
Methoden lediglich begleitend in Kombination mit anderen Verfahren eingesetzt.
2.4.3 Akkommodation und Toleranz In der Humanmedizin kommt es im Fall einer Blutgruppenunverträglichkeit zwischen
Organspender und Empfänger ebenfalls zu einer hyperakuten Abstoßung. Nach
einer Depletion der anti-AB0-Antikörper vor der Transplantation kam es zwar nach
der Organtransplantation wieder zu einem Anstieg dieser Antikörper im Blut, das
Organ wurde jedoch auf Grund einer sehr schwachen Immunantwort trotzdem nicht
abgestoßen. Dieses Phänomen wird als „Akkommodation“ bezeichnet (BACH et al.,
1991).

2 Literaturübersicht 14
Dieses Phänomen wird in der Xenotransplantation genutzt, indem nicht nur eine
Depletion der XNAs (z.B. durch Betrahlung und Organperfusion) vor der
Transplantation durchgeführt wird, sondern zusätzlich noch Knochenmarkzellen des
Spenders in den Empfänger infundiert werden. Durch diese Methode entsteht ein
lymphohämatopoetischer, so genannter gemischter, xenogener Chimärismus in dem
Empfänger (AKSENTIJEVICH et al., 1992). Dieser Chimärismus führt zur Toleranz
der Xenotransplantate durch Entwicklung einer T-Zell-Toleranz. SABLINSKI et al.
zeigten 1997, dass durch eine solche Knochemarkinfusion und Elimination von XNAs
die hyperakute Abstoßung verhindern kann. Obwohl die hyperakute Abstoßung
verhindert werden kann, erreichen die XNA-Level bereits nach wenigen Tagen
wieder ihre Ausgangswerte (KOZLOWSKI et al., 1999) und auch der
lymphohämatopoetische Chimärismus ist nicht stabil und nach einigen Tagen nicht
mehr nachweisbar (TANAKA et al., 1994), so dass die Xenotransplantate schließlich
akut abgestoßen werden.
2.4.4 Hemmung der Komplementkaskade Eine weitere Möglichkeit der Hemmung der hyperakuten Abstoßung stellt die
Hemmung der Komplementkaskade dar, da sie hauptsächlich für die
Abstoßungsreaktion verantwortlich ist.
Eine Depletion der Komplementaktivität kann durch den Einsatz von Cobra Venom
Factor (CVF) erreicht werden. Das Cobra venom enthält ein C3b-Analogon, das mit
anderen Komponenten der alternativen Komplementkaskade einen Enzymkomplex
bildet, der zu einem massiven Verbrauch von C3, Faktor B und Komponenten des
Membrane attack complex (MAC) führt und dadurch die Komplementkaskade hemmt
(LEVENTHAL et al., 1993). Mehrere Transplantationsversuche haben gezeigt, dass
die hyperakute Abstoßungsreaktion durch CVF-Gabe verhindert werden kann
(LEVENTHAL et al., 1993; KOBAYASHI et al., 1997). Jedoch wird auch durch CVF
nur die hyperakute Abstoßung des Xenotransplantates gehemmt, während eine
akute Abstoßung weiterhin stattfindet. Zum einen könnten komplement-unabhängige
Mechanismen für die Abstoßung verantwortlich sein (KOBAYASHI et al., 1997), zum
anderen konnte auch durch die CVF-Therapie eine Deposition von Komplement in

2 Literaturübersicht 15
dem Spenderorgan nicht vollständig verhindert werden (CHEN et al., 2004). Zudem
ist das CVF toxisch und führt zu systemischen Entzündungsreaktionen.
Ein anderes Verfahren nutzt die Komplementhemmung durch den rekombinanten,
löslichen Komplementrezeptor Typ 1 (sCR1). sCR1 hemmt den klassischen und den
alternativen Weg der Komplementkaskade (PRUITT et al., 1994). In
Transplantationsversuchen konnte sCR1 zwar die hyperakute Abstoßung hemmen,
doch die Organe wurden nach einigen Tagen akut abgestoßen (PRUITT et al.,
1997).
Eine weitere Methode der Hemmung der Komplementkaskade stellt C1 Inhibitor dar.
Dies ist ein Regulatorprotein, das die klassische Komplementkaskade hemmt, sobald
C1 durch schwache Aktivatoren wie z.B. Dextransulfat oder Nukleinsäuren aktiviert
wird. Allerdings ist sein Effekt begrenzt, sobald C1 durch starke Aktivatoren wie z.B.
Antigen-Antikörper-Komplexe aktiviert wird (DALMASSO und PLATT, 1993). Damit
ist C1 Inhibitor für das Gebiet der Xenotransplantation nur begrenzt geeignet.
2.4.5 Komplementregulatoren Die so genannten Komplementregulatoren stellen ein weiteres Prinzip zur
Verhinderung der hyperakuten Abstoßungsreaktion dar. Komplementregulatoren sind
Proteine, die auf der Oberfläche von Blutzellen, Endothelzellen und anderen Zellen
exprimiert werden und eine überflüssige oder autologe Komplementaktivierung
verhindern sollen. Dabei hemmen die Komplementregulatoren die
Komplementreaktion sehr speziesspezifisch, d.h. die porzinen
Komplementregulatoren können die humane Komplementkaskade nach einer
Transplantation nicht hemmen. Es gibt verschiedene Komplementregulatoren, die an
verschiedenen Lokalisationen der Komplementkaskade eingreifen (siehe Abb. 1).
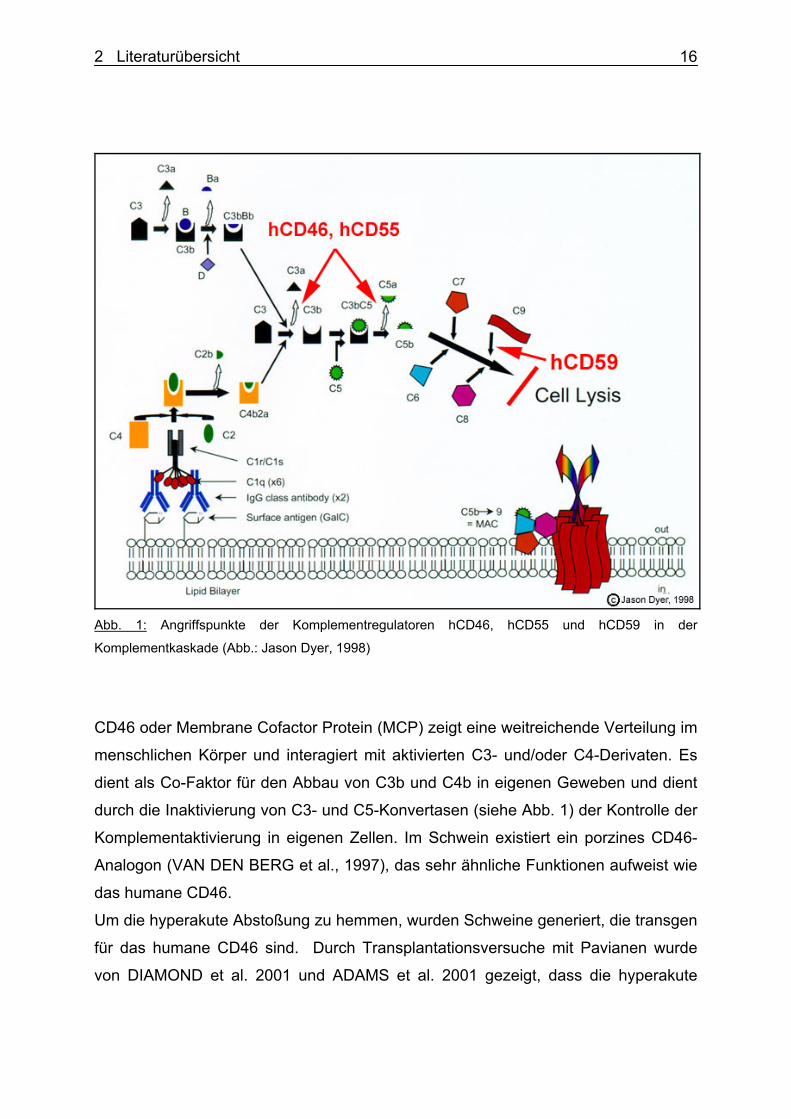
2 Literaturübersicht 16
Abb. 1: Angriffspunkte der Komplementregulatoren hCD46, hCD55 und hCD59 in der
Komplementkaskade (Abb.: Jason Dyer, 1998)
CD46 oder Membrane Cofactor Protein (MCP) zeigt eine weitreichende Verteilung im
menschlichen Körper und interagiert mit aktivierten C3- und/oder C4-Derivaten. Es
dient als Co-Faktor für den Abbau von C3b und C4b in eigenen Geweben und dient
durch die Inaktivierung von C3- und C5-Konvertasen (siehe Abb. 1) der Kontrolle der
Komplementaktivierung in eigenen Zellen. Im Schwein existiert ein porzines CD46-
Analogon (VAN DEN BERG et al., 1997), das sehr ähnliche Funktionen aufweist wie
das humane CD46.
Um die hyperakute Abstoßung zu hemmen, wurden Schweine generiert, die transgen
für das humane CD46 sind. Durch Transplantationsversuche mit Pavianen wurde
von DIAMOND et al. 2001 und ADAMS et al. 2001 gezeigt, dass die hyperakute

2 Literaturübersicht 17
Abstoßung durch das CD46-Transgen verhindert wird. Die Komplementkaskade
wurde durch das humane Transgen an dem C3-Konvertase-Schritt gehemmt. In den
transplantierten Organen fanden sich weiter keine Ablagerungen von C5b oder
MAC. Obwohl in Transplantationsversuchen mit CD46 immer wieder eine weite
Verteilung und Expression des humanen Proteins in den Schweineorganen
nachgewiesen werden konnte und die Organe nach der Transplantation nicht
hyperakut abgestoßen wurden, gelang es bisher nicht, die akute Abstoßung zu
überwinden (LOVELAND et al., 2004).
CD55 oder Decay-Accelerating Factor (DAF) ist ein weiterer Komplementregulator,
der speziesspezifisch sowohl den klassischen als auch den alternativen Weg der
Komplementkaskade hemmt. Dabei hemmt CD55 ebenfalls, wie CD46, die
Aktivierung der C3- und C5-Konvertasen (SMITH UND SMITH, 2001).
Im Rahmen der Forschung auf dem Gebiet der Xenotransplantation wurden
Schweine generiert, die transgen für das humane CD55 sind. Diese Tiere
produzierten das CD55-Protein in einer Vielzahl von Organen und die hCD55-Level
waren dabei höher als im Menschen (COZZI et al., 1997). Dabei zeigten
CARRINGTON at al. 2001, dass ab einer gewissen Konzentration an hCD55-
Molekülen pro Zelle der protektive Effekt nicht mehr gesteigert werden kann. In
zahlreichen Schwein-zu-Primaten Transplantations- und Perfusionsversuchen konnte
demonstriert werden, dass die Expression von hCD55 in porzinen Organen eine
hyperakute Abstoßung verhindert (SCHMOECKEL et al., 1998; WATERWORTH et
al., 1998; COZZI et al., 2000; LUO et al., 2003). Obwohl in fast allen
Transplantationsversuchen die HAR durch hCD55 gehemmt werden konnte, wurden
bei einzelnen Tieren transgene Herztransplantate hyperakut abgestoßen
(SCHUURMAN et al., 2002). Wiederum gelang es jedoch nicht, die akute Abstoßung
der Organe zu überwinden.
Häufig werden die aufgeführten Methoden zur Verhinderung der
Transplantatabstoßung in Kombination genutzt. So werden z.B. transgene Organe in
Primaten transplantiert, die medikamentell immunsupprimiert sind und zusätzlich

2 Literaturübersicht 18
noch eine Depletion der XNAs durch Immunaffinitätssäulen und/oder Bestrahlungen
erhalten haben.
Ein weiterer Komplementregulator ist das humane CD59. Da es sich bei den Tieren
der vorliegenden Studie um hCD59-transgene Schweine handelt, soll im Folgenden
genauer auf dieses Protein eingegangen werden.
2.5 CD59 und hCD59-transgene Tiere
2.5.1 CD59 im Menschen (hCD59) Das hCD59, auch Protectin genannt, wird durch ein einzelnes Gen kodiert, dass auf
Chromosom 11p13 lokalisiert ist (Qian et al., 2000). Das hCD59-Protein ist ein 20 kD
großes Membranprotein, das in großen Mengen im gesamten Organismus, vor allem
auf Lymphozyten und Granulozyten sowie auf Thrombozyten, Erythrozyten und
aktivierten T-Zellen nachgewiesen wurde und mittels Glycosyl Phosphatidylinositol
(GPI) in der Membran verankert ist (DAVIES et al., 1989). MERI et al. zeigten 1991
mittels Immunfloureszenzmikroskopie, dass hCD59 im Gefäßendothel des gesamten
Körpers, aber auch in extravaskulärem Gewebe vorkommt. Dabei zeigten duktale
Epithelien in Pankreas, Gallenblase und Speicheldrüse, Bronchien, Sammelrohre der
Nieren, die Epidermis und Synzytiotrophoblasten der Plazenta eine hCD59-
Expression.
hCD59 hemmt die Komplementreaktion relativ spät während der Kaskade, nämlich
vor der Ausbildung des Membrane Attack Complex (MAC) (DAVIES et al., 1989).
Dabei hemmt hCD59 den C9 Input in den C5b-8 und C5b-9 Komplex und blockiert
zusätzlich die funktionelle Aktivität des im C5b-9 Komplex inkorporierten C9 (MERI et
al., 1990; ROLLINS et al., 1990). So wird die Ausbildung des C5b-9 MAC verhindert.
Zusätzlich hat hCD59 noch eine co-stimulierende Aktivität für T-Zellen (MAHER et
al., 1998). Obwohl zunächst angenommen wurde, dass die komplementhemmende
Wirkung von CD59 streng speziesspezifisch war, zeigten VAN DEN BERG et al.
1994 an einer Untersuchung von CD59 in Menschen, Ratten, Schafen und

2 Literaturübersicht 19
Schweinen, dass CD59 nicht so stark speziesspezifisch reagiert, wie bisher
angenommen wurde. Besonders das CD59 von Ratten und Schafen zeigte auch in
anderen Spezies (z.B. Meerschweinchen) eine protektive Wirkung.
Eine Fehlfunktion von hCD59 kann bei Menschen schwere Folgen haben. Ein
Beispiel dafür ist die Paroxysmale Nächtliche Hämoglobinurie (PNH), eine erworbene
hämatologische Erkrankung. Bei der PNH ist die Glycosyl Phosphatidylinositol-
Verankerung defekt und dadurch enthalten betroffene Blutzellen von PNH-Patienten
kein hCD59 oder hCD55. Aufgrund des Fehlens dieser Komplementregulatoren
kommt es zu einer autologen, komplementvermittelten Lyse von Erythrozyten und
Thrombozyten, die zu einer hämolytischen Anämie und Thrombose führt (QIAN et
al., 2000).
2.5.2 CD59 im Schwein (pCD59) Das porzine Ortholog zu hCD59, pCD59 wird durch ein einziges Gen kodiert, das in
enger Verbindung mit dem follikel-stimulierenden Hormon Beta Polypeptid (FSHB),
auf dem gleichen Chromosom wie das humane CD59 im Menschen lokalisiert ist
(MAHER et al., 1998). pCD59-Protein zeigt eine starke Expression ubiquitär im
Organismus. Am stärksten ist die pCD59-Expression auf den Endothelzellen von
Blutgefäßen. Aber auch Erythrozyten, ein Teil der mononukleären Zellen und
Granulozyten im Blut, Epithelien aller Organe, die graue Substanz im Gehirn und die
T-Zellen der Lymphfollikel in Lymphknoten und Milz zeigen eine starke pCD59-
Expression (HANNA et al., 1998). Im Unterschied zu dem humanen CD59 zeigen ein
Großteil der mononukleären Zellen im Blut, Thrombozyten sowie B-Zellen keine
pCD59-Expression. Das pCD59-Protein hat eine β-Faltblattstruktur und daher ist es
schwierig, die Domäne zu identifizieren, die für die komplementhemmende Wirkung
verantwortlich ist. Man nimmt jedoch an, dass die Aminosäure-Seitenketten zwischen
Position 42 und 58 für die Interaktion mit C8 oder C9 verantwortlich sind (MAHER et
al., 1998).
Der Wirkungsmechanismus von pCD59 entspricht dem Wirkungsmechanismus des
humanen CD59. Auch pCD59 hemmt die Ausbildung des MAC durch die Blockierung
der Interaktion von C5b-8 mit C9.

2 Literaturübersicht 20
HANNA et al. zeigten 1998, dass das pCD59 in der Lage ist, auch humanes
Komplement zu hemmen, somit also auch nicht streng speziesspezifisch reagiert.
Die hyperakute Abstoßung durch humanes Komplement ist daher möglicherweise
auf eine nicht ausreichende Kapazität des pCD59 zurückzuführen.
2.5.3 CD59 bei anderen Tierarten Orthologe von hCD59 sind auch in vielen anderen Tierarten untersucht worden, wie
z.B. in Ratten, Mäusen, Schafen und Primaten. Im Allgemeinen weisen alle
Orthologe die gleiche Struktur im Hinblick auf das Molekulargewicht (18-20 kD) und
die GPI-Verankerung in der Membran auf. Auch die Verteilung in den bisher
untersuchten Spezies ist ähnlich, da CD59, mit einzelnen Variationen, ubiquitär auf
Blutzellen, Endothelien und Epithelien vorhanden ist.
Die Maus weist in ihrem CD59-Ortholog im Vergleich zum Mensch, zur Ratte und
zum Schwein eine Besonderheit auf: Während diese Spezies nur ein Gen für das
CD59 besitzen, sind in der Maus zwei Gene, CD59a und CD59b, nachgewiesen
worden. Während CD59a weit im gesamten Organismus exprimiert wird, wird CD59b
selektiv im Hoden der Mäuse exprimiert. Man nimmt an, dass die zwei CD59-Gene
der Maus verschieden reguliert sind und sich ihre physiologischen Aufgaben nicht
überschneiden (QIAN et al., 2000).
2.5.4 hCD59-transgene Tiere Da die Komplementreaktion den Hauptmechanismus der hyperakuten Abstoßung
darstellt, ist die Hemmung der Komplementkaskade ein Ansatz, um die Abstoßung
des Xenotransplantates zu verhindern. Da das porzine CD59 die
Komplementreaktion an den transplantierten Organen nur sehr begrenzt hemmen
kann, wurden Tiere generiert, die in ihren Organen das humane CD59 exprimieren.
Das hCD59 soll dann direkt auf den Xenotransplantaten die Aktivierung der
Komplementkaskade verhindern.
Da die Herstellung solcher transgener Schweine sehr teuer, ineffektiv und
zeitaufwendig ist, wurden auch hCD59-transgene Mäuse hergestellt, da sie schneller

2 Literaturübersicht 21
und leichter zu generieren sind. An den hCD59-transgenen Mäusen wurde der
protektive Effekt des humanen Transgens getestet. In extrakorporalen Perfusionen
von transgenen Mäuseorganen mit humanem Serum zeigte das humane CD59 einen
protektiven Effekt. Weiterhin wurden porzine Nierenzellen (PK1-Zellen) mit humaner
CD59-cDNA transfiziert und auch hier zeigte sich ein protektiver Effekt bei Kontakt
mit humanem Serum (SOMERVILLE et al., 1994).
FODOR et al. gelang 1994 neben der Produktion von hCD59-transgenen Mäusen
auch die Herstellung eines hCD59-transgenen Schweins. Das Tier wurde mittels
Mikroinjektion eines hCD59-Konstruktes in die Pronuklei einer Zygote generiert. Als
genomische Expressionskassette diente dabei das murine MHC Klasse I Gen
(H2Kb).
Die Methode der Mikroinjektion stellte lange Zeit die einzige Methode dar, um
transgene Schweine zu produzieren. Die Nachteile der Mikroinjektion liegen in einer
geringen Effektivität und in einer zufälligen Integration des humanen Konstrukts, so
dass unklar ist, ob, wo und in welcher Quantität das humane CD59 integriert wird.
Zusätzlich können Positionseffekte des zufällig eingebauten Konstrukts
Auswirkungen auf den Phänotyp des Schweins haben (NIEMANN et KUES, 2000).
Nach der Etablierung von Klonen und Knockouts bei Schweinen kann jetzt eine
gezieltere Manipulation der Tiere erfolgen und in den nächsten Jahren wird die
Technik wahrscheinlich durch eine Kombination von Knock-outs und Transgenen
noch erheblich verbessert werden.
2.6 Charakterisierung der Expression in transgenen Schweinen
Nach der gelungenen Herstellung von transgenen Schweinen muss geklärt werden,
ob und in welcher Verteilung das humane Transgen in den Geweben und Organen
des Schweins exprimiert wird. Weiterhin gilt es zu untersuchen, ob das humane
Transgen durch Positions- oder andere Effekte zu einem morphologischen Phänotyp
bei den Tieren führt.

2 Literaturübersicht 22
2.6.1 Expression des hCD59-Transgens
Ziel der Herstellung transgener Tiere ist häufig, eine möglichst ubiquitäre und starke
Expression des Transgens in den Organen und Geweben zu erreichen. Häufig hat
die Wahl des Promotors einen großen Einfluss auf das Verteilungsmuster und die
Expressionsstärke des Transgens, da einige Promotoren gewebsspezifisch sind.
FODOR et al. setzten 1994 für die Generierung von hCD59-transgenen Mäusen und
einem hCD59-transgenen Schwein ein murines MHC Klasse 1 Gen, H2Kb als
Promotor ein. H2Kb wird ubiquitär exprimiert und ist vor allem ein wichtiges
Endothelzell-Oberflächenantigen. Bei der Expressionsanalyse wiesen FODOR et al.
bei drei transgenen hCD59-Foundermäusen eine Expression von hCD59-Protein auf
dem Endothel von Blutgefäßen im Herz und auf dem Endokard nach. Um das
transgene Schwein nicht töten zu müssen, wurde von diesem Tier ein Stück
Schwanz untersucht und auch hier wurde eine Expression von hCD59-Protein in
Fibroblasten, Epithelzellen, Endothelzellen und glatten Muskelzellen nachgewiesen.
Somit führte der eingesetzte H2Kb-Promotor zu einer Expression des Transgens in
einer Vielzahl von Zelltypen und Geweben. Zudem waren Zellen des transgenen
Schweins bei Kontakt mit humanem Serum vor einer Komplementreaktion geschützt.
Auch DIAMOND et al. charakterisierten 1996 mehrere hCD59-transgene Schweine.
In der RNA-Analyse mittels Northern Blot zeigten die Tiere eine Expression von
hCD59-RNA in Leber, Herz, Niere, Lunge, Milz, Muskel, Haut und Pankreas, wobei
die Level des hCD59 etwa 10fach geringer waren als im Menschen. Das hCD59-
Protein wurde auch auf dem Endothel exprimiert, allerdings nur in größeren Gefäßen
und mit einem sehr viel schwächeren Signal als im Menschen. Trotzdem zeigten die
porzinen Endothelzellen nach Inkubation mit humanem Serum keine Zytotoxizität.
Bei einer heterotopen Transplantation von Herzen der hCD59-transgenen Schweine
in Paviane überlebten die transgenen Organe zwar nur etwas länger als die Herzen
der nicht-transgenen Kontrolltiere, aber dafür waren die histopathologischen
Merkmale der hyperakuten Abstoßung in den transgenen Herzen nicht so
ausgeprägt, und zudem zeigten die transgenen Herzen einen deutlich verringerten

2 Literaturübersicht 23
Nachweis von MAC im Vergleich zu den Kontrollherzen. Diese Ergebnisse sprachen
dafür, dass trotz der geringen Expressionslevel von hCD59 ein gewisser protektiver
Effekt vorhanden war.
COWAN et al. generierten 1998 hCD59-transgene Mäuse, die das Transgen
entweder unter dem H2Kb-Promotor oder unter dem endothel-spezifischen,
humanen, interzellulären Adhäsionsmolekül-2-Promotor (ICAM-2) exprimierten. Die
Expressionsmuster der beiden Promotoren waren deutlich unterschiedlich: Alle
Leukozyten der H2Kb-Linie exprimierten hCD59 unterschiedlich stark, während die
ICAM-2-Linie hCD59 vorwiegend auf neutrophilen Granulozyten, aber nicht auf
Lymphozyten exprimierten. Hingegen zeigten kardiale Endothelzellen der ICAM-2-
Linie eine 25-30fach höhere hCD59-Expression als die H2Kb-Linie. Auch in
Perfusionsversuchen mit humanem Serum zeigten die ICAM-2-Herzen eine längere
Vitalität als die H2Kb-Herzen. Diese Ergebnisse zeigten, dass eine hohe endotheliale
Expression von hCD59 eine bessere Protektivität bietet.
NIEMANN et al. untersuchten 2001 das Expressionsmuster von hCD59-transgenen
Schweinen, die das Transgen unter dem Cytomegalovirus (CMV)-Promotor
exprimierten. Der CMV-Promotor induzierte eine starke und ubiquitäre Expression in
einer Reihe von Zelllinien und transgenen Mäusen. In den transgenen Schweinen
führte der Promotor zu einer Expression von hCD59 in einer Reihe von Organen und
Geweben, wobei die Expressionslevel zwischen Linien verschiedener Founder
erheblich schwankte. Zwei Linien zeigten eine Expression von hCD59 in Herz,
Muskel, Lunge, Niere, Pankreas und Haut. Andere Linien exprimierten hCD59
lediglich im Pankreas. In der immunhistochemischen Untersuchung konnte eine
zelltyp-spezifische Expression dargestellt werden. Im Herz zeigten die
Kardiomyozyten eine starke Anfärbung, während die Endothelzellen weniger stark
reagierten. In der Haut färbte sich lediglich die Epidermis und in der Niere
vorwiegend die Tubulusepithelzellen. Zudem lag in der Niere ein sogenanntes
„patchy pattern“ vor, bei dem sich nur einzelne Areale positiv färbten. Bei einer
Inkubation von transgenen Fibroblasten und Endothelzellen mit humanem Serum

2 Literaturübersicht 24
sowie bei einer extrakorporalen Nierenperfusion mit humanem Blut zeigte sich bei
den transgenen Zellen und Nieren eine deutlich ausgeprägte Protektion im Vergleich
zu den nicht-transgenen Zellen und Organen. Diese Ergebnisse zeigten, dass der
CMV-Promotor zu einer weit verteilten Expression von hCD59 führt, wenn auch nicht
in allen Organen und Zellen eine Expression nachgewiesen werden konnte.
Insgesamt führten alle eingesetzten Promotor zu der Expression des Transgens und
zu einem protektiven Effekt, wenn auch die Expression nicht immer ubiquitär
vorhanden war und nur die hyperakute Abstoßung, nicht jedoch die akute Abstoßung
gehemmt werden konnte.
2.6.2 Expression von mehreren Transgenen Um den protektiven Effekt noch zu steigern, werden transgene Tiere konstruiert, die
nicht nur ein Transgen, sondern mehrere Transgene exprimieren. Diese doppelt-
oder dreifach-transgenen Tiere sollen durch eine Kombination und/oder Addition der
Transgeneigenschaften zu einer höheren Xenotransplantatakzeptanz führen.
COSTA et al. untersuchten 1999 drei Mauslinien: Eine Mauslinie war doppelt
transgen für hCD59 und α1-2Fucosyltransferase (HT), eine Mauslinie war transgen
für hCD59 und besaß zusätzlich einen α1-3Gal-knockout und die dritte Linie war
doppelt transgen für hCD59 und HT und besaß zusätzlich noch einen knockout für
α1-3Gal. Als Promotor wurde ein β-actin-Promotor gewählt. Alle drei Linien zeigten
eine mRNA- und Protein-Expression der Transgene in allen untersuchten Organen.
Dabei gab es zwischen den drei Linien keine signifikanten qualitativen oder
quantitativen Unterschiede. In einer Untersuchung der Hemmung der Zytolyse durch
humanes Serum zeigte sich, dass die hCD59xHT-Kombination genauso effektiv war
wie die hCD59xα1-3Gal-ko-Kombination. Überraschenderweise war auch in der
dritten Linie, die doppelt transgen war und einen α1-3Gal-ko aufwies, die Protektion
nicht merklich verbessert.

2 Literaturübersicht 25
COWAN et al. generierten 2000 dreifach-transgene Schweine, die unter dem H2Kb-
Promotor hCD59, hCD55 und HT exprimierten. In der Expressionsanalyse konnte
eine starke Expression von hCD59 und hCD55 in allen untersuchten Geweben und
Organen nachgewiesen werden, während HT nur sehr schwach exprimiert wurde
und zu keiner signifikanten Reduktion des Gal-Epitops führte. Bei einer
Transplantation von dreifach transgenen Nieren in nicht-immunsupprimierte Paviane
konnte die hyperakute Abstoßung dennoch gehemmt werden.
COSTA et al. erstellten 2002 hCD59/HT doppelt-transgene Schweine. Die Founder
der doppelt-transgenen Tiere exprimierten das jeweilige Transgen unter dem K2Kb-
oder dem CMV-Promotor. Die doppelt-transgenen Tiere zeigten ein
unterschiedliches Expressionsmuster. Während eine Linie eine starke Expression
beider Transgene zeigte, wies die andere Linie ein Mosaikmuster und eine schwache
Expression der Transgene auf. hCD59-mRNA konnte in allen untersuchten Geweben
mittels RT-PCR nachgewiesen werden. Die HT-mRNA-Expression wurde unter dem
CMV-Promotor in nahezu allen Geweben nachgewiesen, während unter dem H2Kb-
Promotor nur in der Milz eine Expression festgestellt wurde. Monozyten und
Endothelzellen der doppelt-transgenen Tiere zeigten bei Kontakt mit humanem
Serum eine deutlich bessere Protektion gegenüber einer Zytolyse als Zellen von
Tieren, die nur eines der beiden Transgene exprimieren. Diese Ergebnisse stehen im
Widerspruch zu den Ergebnissen von COWAN et al., 2000.
BYRNE et al. konstruierten 1997 transgene Schweine, die doppelt-transgen für die
Komplementregulatoren hCD59 und hCD55 waren. In der Expressionsanalyse dieser
Tiere wurde eine variierend starke Expression von hCD59 in allen untersuchten
Organen gefunden. Dabei waren die Expressionslevel niedriger als in humanen
Geweben. Die Expression von hCD55 war vom Verteilungsmuster ähnlich wie
hCD59, jedoch waren die Expressionslevel niedriger als bei hCD59 und auch
niedriger als in humanen Geweben. Trotz der geringen hCD55-Expression konnte in
Transplantationsversuchen mit Pavianen eine Hemmung der Komplementreaktion
nachgewiesen werden. Dabei waren sowohl C3 und C5b, als auch MAC in

2 Literaturübersicht 26
reduzierten Mengen vorhanden. Diese Ergebnisse zeigen, dass beide
Komplementregulatoren an der Hemmung der Komplementkaskade beteiligt waren,
trotz der niedrigen Expression von hCD55.
CHEN et al. generierten 1999 ebenfalls hCD59/hCD55 doppelt-transgene Schweine,
die mit Hilfe von CD59 und CD55 humanen genomischen Klonen konstruiert wurden.
Immunhistochemisch wurde in den doppelt-transgenen Herzen Expressionlevel für
hCD59 und hCD55 nachgewiesen, die den in humanen Herzen in etwa entsprechen.
Nach heterotoper Herztransplantation zeigten beide Transgene einen protektiven
Effekt mit Hemmung der hyperakuten Abstoßung.
Insgesamt wurde bei der Konstruktion von doppelt-transgenen Tieren in den meisten
Fällen gezeigt, dass ein Transgen stärker exprimiert wurde als das andere (BYRNE
et al., 1997; ZHOU et al., 2002). Trotzdem zeigten beide Transgene einen
protektiven Effekt, wenn auch nur die hyperakute Abstoßung, nicht jedoch die akute
Abstoßung gehemmt werden konnte.
Auch für die beiden anderen Komplementregulatoren hCD55 und hCD46 wurden
Expressionsanalysen an transgenen Tieren durchgeführt. Dabei waren die
Expressionslevel teilweise höher als beim Menschen (COZZI et al., 1997). Doch
auch für diese Transgene gilt, dass sich der protektive Effekt auf die hyperakute
Abstoßung beschränkt.
2.6.3 Weitere Transplantationsversuche Mit Schweinen, die transgen für Komplementregulatoren sind, wurden mittlerweile
viele Transplantationsversuche durchgeführt, über die im Folgenden ein kurzer
Überblick gegeben werden soll.
1996 führten NORIN et al. orthotope Lungentransplantationen von hCD59-
transgenen Schweinen in Paviane durch und konnte zeigen, dass eine hyperakute

2 Literaturübersicht 27
Abstoßung im Vergleich zu nicht-transgenen Organen (Abstoßung nach 3 Stunden
post operationem) verhindert wurde. Allerdings wurden die Xenotransplantate 12
Stunden post operationem akut abgestoßen.
1998 führten SCHMOECKEL et al. orthotope Herztransplantationen von hCD55
transgenen Schweinen in Paviane durch und zeigte, dass eine hyperakute
Abstoßung verhindert wurde und die Schweineherzen zunächst ihre physiologische
Funktion anfnahmen, bevor sie akut abgestoßen wurden.
ZAIDI et al. machten 1998 eine überraschende Beobachtung, als sie nach
Nierentransplantationen zwischen hCD55-transgenen Schweinen und
Cynomolgusaffen zwar keine hyperakute Abstoßung der Organe nachwiesen, aber
auch in den Kontrollgruppen mit nicht-transgenen Nieren die Nieren teilweise nicht
hyperakut abgestoßen wurden. Bereits 1995 hatten KAPLON et al. das gleiche
Phänomen nach Transplantationen von nicht-transgenen Schweinelungen in Paviane
beobachtet.
2000 erreichten COZZI et al. die bislang längste Überlebenszeit eines
Xenotransplantates. Nieren von hCD55-transgenen Schweinen wurden orthotop in
immunsupprimierte Cynomolgusaffen transplantiert. Eine Niere wurde erst nach 78
Tagen abgestoßen und 4 von 9 Tieren überlebten für 50 oder mehr Tage. Dabei
lagen die Nierenwerte wochenlang im physiologischen Bereich.
2000 schlossen LEVY et al. zwei humane Patienten mit einem akuten, totalen
Leberversagen an extrakorporale, hCD55- und hCD59-transgene Schweinelebern an
und konnten so einen Zeitraum von 6,5, bzw. 10 Stunden überbrücken, bis ein
Allotransplantat zur Verfügung stand. Damit zeigten sie, dass transgene
Schweineorgane zur Überbrückung der Wartezeit auf ein geeignetes Allotransplantat
genutzt werden könnten.

2 Literaturübersicht 28
Insgesamt zeigen die bisher weltweit durchgeführten Versuche, dass trotz der
Fortschritte auf dem Gebiet der Erzeugung transgener Tiere und der knockout-
Technik die Erfolge auf dem Gebiet der Xenotransplantation bisher eher bescheiden
sind. Zwar gelingt es mittlerweile, die hyperakute Abstoßung der Xenotransplantate
zu verhindern, die Organe werden jedoch trotz der Expression von humanen
Transgenen oder Gal-knockouts immer noch akut abgestoßen. Man hofft, durch die
Kombination von Knockout-Technik und der Expression von humanen Transgenen in
den nächsten Jahren auch die akute Abstoßung zu überwinden.
2.7 Morphologische Phänotypen transgener Tiere
Die Manipulation am Genom von Tieren kann zur Ausprägung von
pathomorphologischen Phänotypen führen. Am besten ist dieses Phänomen bei
Mäusen erforscht, da bei dieser Spezies bereits seit Jahren gezielte Veränderungen
des Genoms vorgenommen werden können. In vielen Fällen ist ein morphologisch
manifester Phänotyp der Tiere geplant und erwünscht, z. B. wenn es um die
Erzeugung von Mausmodellen für bestimmte Krankheiten geht. Manchmal
entwickeln die Tiere jedoch einen im Vorfeld ungeplanten Phänotyp durch die
genetischen Veränderungen.
Bei den landwirtschaftlichen Nutztieren sind bislang nicht so viele Phänotypen
beschrieben worden, da bei ihnen die biotechnologischen Methoden noch nicht so
lange und so häufig eingesetzt werden. Trotzdem sind auch bei Schafen, Rindern
und Schweinen pathomorphologische Phänotypen in Folge genetischer
Manipulationen beschrieben worden. Besonders die Mikroinjektion birgt die Gefahr
ungeplanter Phänotypen, da der Einbau der mikroinjizierten DNA rein zufällig erfolgt
und dazu führen kann, dass das Transgen gar nicht exprimiert wird, verstärkt
exprimiert wird oder in einem anderen Organ oder Gewebe exprimiert wird als
ursprünglich geplant (ektopische Expression) (VAN REENEN et al., 1999). Zudem

2 Literaturübersicht 29
kann das Transgen benachbarte Gene des Tieres beeinflussen und auch dadurch zu
einem spezifischen Phänotyp führen.
Auf die Ausprägung einiger Phänotypen soll im Folgenden genauer eingegangen
werden.
2.7.1 Ektopische Expression NIEMANN et al. generierten 1999 transgene Schafe, die den humanen
Gerinnungsfaktor Faktor VIII in ihrer Milch exprimieren sollten. Dies sollte mit einem
Milchdrüsen-spezifischen Promotor des ovinen beta-Lactoglobulin-Gens erreicht
werden. Zwar zeigten einige Tiere eine Expression des gewünschten Proteins in der
Milch, ein Tier exprimierte die mRNA jedoch zusätzlich noch ektopisch in Gehirn,
Herz, Milz, Niere und Speicheldrüse. Dies zeigte, dass der gewählte Promotor nicht
zu einer ausschließlichen Produktion des Transgens in der Milch führte.
1996 generierten MASSOUD et al. transgene Kaninchen, die humanes Erythropoetin
über einen Kaninchen Whey Acid Promotor in der Milchdrüse exprimieren sollten.
Die transgenen Kaninchen hatten einen abnormal erhöhten Gehalt an Erythrozyten,
konnten sich nicht fortpflanzen, produzierten keine Milch und starben sehr jung. In
den betroffenen Tieren wurde das Erythropoetin-Gen in niedrigen Mengen auch in
anderen Organen als der Milchdrüse produziert.
2.7.2 Large Offspring Syndrome (LOS) Bei Schafen und Rindern haben in vitro Technologien bereits häufig zum so
genannten Large Offspring Syndrome (LOS) geführt. Das deutlichste Symptom des
LOS ist das hohe Geburtsgewicht der Tiere. Dabei variieren die Geburtsgewichte
stark, ein doppelt erhöhtes Geburtsgewicht ist keine Seltenheit und es sind sogar
Geburtsgewichte bis zum fünffachen des Normalen beschrieben worden. Da häufig
eine Dystokie mit dem hohen Geburtsgewicht vergesellschaftet ist, müssen die
Lämmer bzw. Kälber oft per Kaiserschnitt entwickelt werden. Die Trächtigkeitszeit ist
häufig verlängert und unter der Geburt kommt es oftmals zu Atemschwierigkeiten,
Schwierigkeiten beim Saugen und zu plötzlichem perinatalen Tod. Während der
Trächtigkeit sind auch pränatale Verluste, besonders in der ersten Trächtigkeitshälfte

2 Literaturübersicht 30
zu beobachten. Für einige Organe sind makroskopische Läsionen beschrieben
worden, wie z.B. vermehrte Muskelmasse, Muskelfaserveränderungen, zerebelläre
Dysplasie und Missbildungen des Skelettsystems und des Kopfes. Die
allometrischen Wachstumskoeffizienten für fetale Leber, Herz, Niere und Muskel sind
in von LOS betroffenen Schaffeten erhöht. Dies weist darauf hin, dass auch die
Entwicklung dieser Organe gestört sein könnte (YOUNG et al., 1998).
Das LOS ist bislang nur bei Rindern und Schafen beschrieben, während es bei
Schweinen und Mäusen nach der Anwendung von in vitro Technologien bisher nicht
aufgetreten ist.
2.7.3 Kardiopulmonale und plazentale Phänotypen 1998 klonten CIBELLI et al. Kälber aus fetalen Fibroblasten. Im Verlauf des
Versuches kam es bei einigen Kälbern zu Plazentationsstörungen in Form einer
Hydroallantois, vergrößerten Plazentomen und einer ödematiserten Chorioallantois
und Amnion. Ein abortiertes Kalb zeigte einen vergrößerten rechten Herzventrikel,
ein Lungenödem und stark vergrößerte Nabelgefäße. Ein lebend geborenes Kalb
verstarb im Alter von fünf Tagen mit einem vergrößerten rechten Herzventrikel, einer
pulmonalen Hypertension, einem Ductus arteriosus persistens, einer Lungenarterie,
die größer war als die Aorta und dreifach vergrößerten Nabelgefäßen.
HILL et al. untersuchten 1999 morphologische Phänotypen von dreizehn aus
Fibroblasten geklonten Kälbern und fanden ähnliche Befunde wie CIBELLI et al.
1998. Auch in ihren Untersuchungen stellten sie dilatative Kardiomyopathien,
pulmonale Hypertension, plazentale Ödeme und Hydroallantois fest. Außerdem
zeigten 8 Kälber ein neonatales respiratorisches distress Syndrom und ein Kalb wies
eine pulmonale Surfactant-Insuffizienz auf. Insgesamt standen bei den untersuchten
Kälbern kardiopulmonale Phänotypen, häufig im Zusammenhang mit
Plazentaveränderungen im Vordergrund. Es wurde vermutet, dass die Technik des
Klonens und/oder die Bedingungen der in vitro Embryokulturen für die Phänotypen
verantwortlich waren, der Mechanismus konnte jedoch nicht geklärt werden.

2 Literaturübersicht 31
Ähnliche Phänotypen wie die oben beschriebenen beobachteten auch LAI et al. 2002
bei geklonten Ferkeln, die ein Allel des Gal-Epitops ausgeknockt hatten. Auch bei
diesen Tieren lagen ein dilatierter rechter Ventrikel und eine verdicktes Myokard vor.
Auch CARTER et al. beobachteten 2002 bei zwei von zehn EGFP (Enhanced Green
Flourescent Protein)-transgenen Ferkeln eine Herzinsuffizienz in Folge einer
dilatativen Kardiomyopathie des rechten Ventrikels.
2.7.4 Muskel- und Skelett-Phänotypen PARK et al. generierten 2001 mittels nukleärem Transfer transgene Schweine, die
das Enhanced Green Flourescent Protein (EGFP) unter der Kontrolle eines
Cytomegalovirus (CMV)-Promotors exprimierten. Die Ferkel zeigten
Beugesehnenkontrakturen an den Vorderbeinen und ein Tier wies eine bilaterale
tarsale Hyperextension auf. Diese Läsionen verschwanden jedoch innerhalb einiger
Tage wieder.
Auch LAI et al. beschrieben 2002 zusätzlich zu den kardialen Veränderungen in den
Ferkeln mit einem ausgeknockten Allel der α-1-3-Galactosyltransferase
Beugesehnenfehlbildungen. Zudem zeigten die Gal-knockout Ferkel im Vergleich zu
Kontroll-Ferkeln der gleichen Rasse geringere Geburtsgewichte (450g – 650 g,
Normalwert: ca. 860g). Im Gegensatz dazu konnten DAI et al., die ebenfalls 2002
Ferkel mit einem ausgenockten Allel der α-1-3-Galactosyltransferase produzierten,
bei ihren Tieren keinerlei pathomorphologische Veränderungen feststellen.
2.7.5 Sonstige Phänotypen SHAMAY et al. beschrieben 1992 die Produktion von sechs verschiedenen Linien
transgener Schweine, die in ihrer Milch murines Whey Acidic Protein in hohen Dosen
produzierten. In drei der sechs transgenen Linien zeigten die Säue eine stark
eingeschränkte Milchproduktion und unreifes Milchdrüsenparenchym. Dieses
Phänomen war bereits bei transgenen Mäusen, die ebenfalls das Whey Acidic
Protein in der Milchdrüse produzierten, als „Milchlos Phänotyp“ beschrieben worden.
SHAMAY et al. postulierten 2001, dass das Whey Acidic Protein zu einer

2 Literaturübersicht 32
eingeschränkten Entwicklung und Funktion der Milchdrüse und damit zu dem
Milchlos Phänotyp führt.
2000 produzierten McCREATH et al. Schafe mittels nukleärem Transfer, die über
gene-targeting ein therapeutisches Transgen in dem ovinen α1(I)Prokollagen
(COL1A1)-Lokus exprimierten. Von den 14 lebend geborenen Lämmern starben 7
innerhalb der ersten 30 Lebensstunden und 4 weitere in den nächsten 12 Wochen.
Bei den verstorbenen Tieren ergaben die durchgeführten Sektionen eine Reihe von
Abnormalitäten. Obwohl sich kein konstantes Muster erkennen ließ, zeigte sich ein
hoher Anteil an Nierendefekten, wie z.B. Nierenbeckendilatation, und an Leber- und
Gehirnläsionen (nicht näher spezifiziert).
Insgesamt zeigen die Phänotypen, die nach genetischen Manipulationen auftreten
eine hohe Variabilität, häufig kann man aber auch spezifische Phänotypen
beobachten, die mit einem bestimmten Genotyp in Zusammenhang stehen. In den
meisten Fällen ist es jedoch schwierig zu bestimmen, welcher genaue Mechanismus
für die Entstehung eines Phänotyps verantwortlich ist.
2.8 Genkonstrukt
Die für die vorliegende Studie untersuchten transgenen Tiere wurden im Institut für
Tierzucht und Tierverhalten der FAL Mariensee durch Mikroinjektion der
Genkonstrukte in den Vorkern einer Zygote hergestellt (NIEMANN et al., 2001;
Konstruktsequenz siehe Anhang).
Die verwendeten Genkonstrukte bestanden aus einer bicistronischen Kassette, in der
der Komplementregulator hCD59 entweder mit EGFP (Enhanced Green Flourescent
Protein), oder mit NTA (Transaktivator, Lokalisation im Konstrukt: bp 445-951) über
ein IRES-Element (Internal Ribosome Entry Side) (Lokalisation im Konstrukt: bp 952-
1579) verbunden war und durch einen Cytomegalovirus (CMV)–Promotor getrieben

2 Literaturübersicht 33
wurde. Der IRES-Sequenz folgte das Transkativator-Gen (Lokalisation im Konstrukt:
bp 1580-2380) und schließlich eine Poly-A Erkennungssequenz.
Das EGFP sollte durch die Floureszenz einer leichteren, floureszenzmikroskopischen
Detektion des Transgens dienen. Die IRES-Sequenz stellt einen internen
Zugangspunkt für die Initiation der Translation durch eukaryotische Ribosomen dar.
Das Transaktivator-Gen ist eine Fusion aus einem bakteriellen Tet-Repressor und
einer viralen VP 16 Domäne. Der Tet-Repressor bindet spezifisch an Tet-Operator-
Sequenzen im Promotor während die virale VP 16 Domäne interagiert und die
eukaryontische Polymerase II aktiviert. Auf diesem Weg erhöht der Transaktivator
die Expression. Zudem kann der Tet-Repressor des Transaktivators Tetrazyklin
binden, wodurch er seine Konformation ändert und dann nicht mehr an Tet-Operator-
Sequenzen bindet. Dadurch ist die VP 16 Domäne nicht mehr in der Nähe der
Polymerase II, deren Aktivierung ausbleibt, und so wird schließlich die Expression
verringert (FURTH et al., 1994).

34
3 Ziel der Studie
Diese Studie sollte mittels pathologisch-anatomischer und histopathologischer
Untersuchungen klären, ob die per Mikroinjektion erzeugten hCD59-transgenen
Schweine einen spezifischen pathomorphologischen Phänotyp aufwiesen, und ihn
gegebenenfalls beschreiben.
Weiterhin sollte mittels immunhistochemischer Untersuchungen mit einem
spezifischen hCD59-Antikörper geklärt werden, ob das humane Protein in den
porzinen Organen und Geweben exprimiert wurde und in welchen Zellen die
Expression nachweisbar war. In diesem Zusammenhang wurde auch das
Verteilungsmuster und die Intensität der hCD59-Proteinexpression beurteilt.
Des weiteren wurde mittels einer qualitativen Reversen-Transkriptase-PCR mit
einem spezifischen hCD59-Primer untersucht, ob die porzinen Organe und Gewebe
die hCD59-mRNA enthielten, welches Expressionsmuster für die mRNA vorlag und
ob dieses Muster mit dem Expressionsmuster für das hCD59-Protein übereinstimmt.
Eine qualitative PCR mit einem spezifischen hCD59-Primer diente dazu, die
Detektion und das Verteilungsmuster der hCD59-DNA in den verschiedenen
porzinen Organen und Geweben zu untersuchen. Auch hier wurde ein Vergleich des
DNA-Nachweises mit dem Expressionsmuster für das hCD59-Protein und die
hCD59-mRNA durchgeführt.
Diese Untersuchungen sollten wesentliche Hinweise zur Beantwortung der Frage
liefern, ob die untersuchten hCD59-transgenen Tiere als Spendertiere für
Organtransplantate für den Menschen in Frage kommen.

35
4 Eigene Ergebnisse:
Pathomorphologische Charakterisierung
4.1 Hintergrund und Bedeutung
Die pathomorphologische Charakterisierung der transgenen Tiere diente dazu, einen
möglicherweise vorhandenen Phänotyp zu identifizieren und zu beschreiben. Es ist
bekannt, dass genetische Manipulationen am Erbgut zu der Ausbildung eines
Phänotyps führen können.
Die Erstellung der transgenen Schweine mittels Mikroinjektion führt dazu, dass das
humane Konstrukt zufällig im Genom des Schweins integriert wird. Daher lässt sich
nicht sagen, wo die Insertionsstelle des Konstrukts liegt, und ob durch die Position
des Konstruktes möglicherweise porzine Gensequenzen durch so genannte
Positionseffekte beeinflusst werden. Diese Positionseffekte können dazu führen,
dass es zu einer Hemmung oder andersartigen Beeinflussung (z.B. einer verstärkten
Expression) von porzinen Gensequenzen kommt und dadurch möglicherweise ein
pathomorphologischer Phänotyp entsteht.
Um zu klären, ob die Integration des humanen Konstruktes zu der Ausbildung eines
Phänotyps geführt hat, wurde eine pathomorphologische Charakterisierung der
transgenen Tiere im Vergleich zu den nicht-transgenen Kontrolltieren durchgeführt.
4. 2 Experimentelles Design
4.2.1 Material Für die pathomorphologischen Untersuchungen wurden 19 transgene Schweine
(gekennzeichnet mit Kürzel TG und fortlaufender Nummer) und 9 nicht-transgene
Kontrollschweine (gekennzeichnet mit KO und fortlaufender Nummer) der Rasse
Deutsche Landrasse im Alter von 1 bis 32 Monaten untersucht. Beide Gruppen

4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 36
wurden unter gleichen Bedingungen gehalten und es handelte sich bei den Tieren
um spezifiziert-pathogen-freie (SPF) Schweine (Tab. 1).
18 der 19 untersuchten Schweine waren transgen für hCD59. Ein Tier (TG23) war
transgen für den Komplementregulator hCD55 und diente als zusätzliche
Negativkontrolle. Bei dem Tier TG26 handelte es sich um ein doppelt transgenes
Tier, das sowohl den Komplementregulator hCD55, als auch hCD59 trug.
Die genauen Genotypen der einzelnen Schweine sowie Alter und Zuchtlinie der
untersuchten Tiere sind in Tabelle 1 dargestellt.
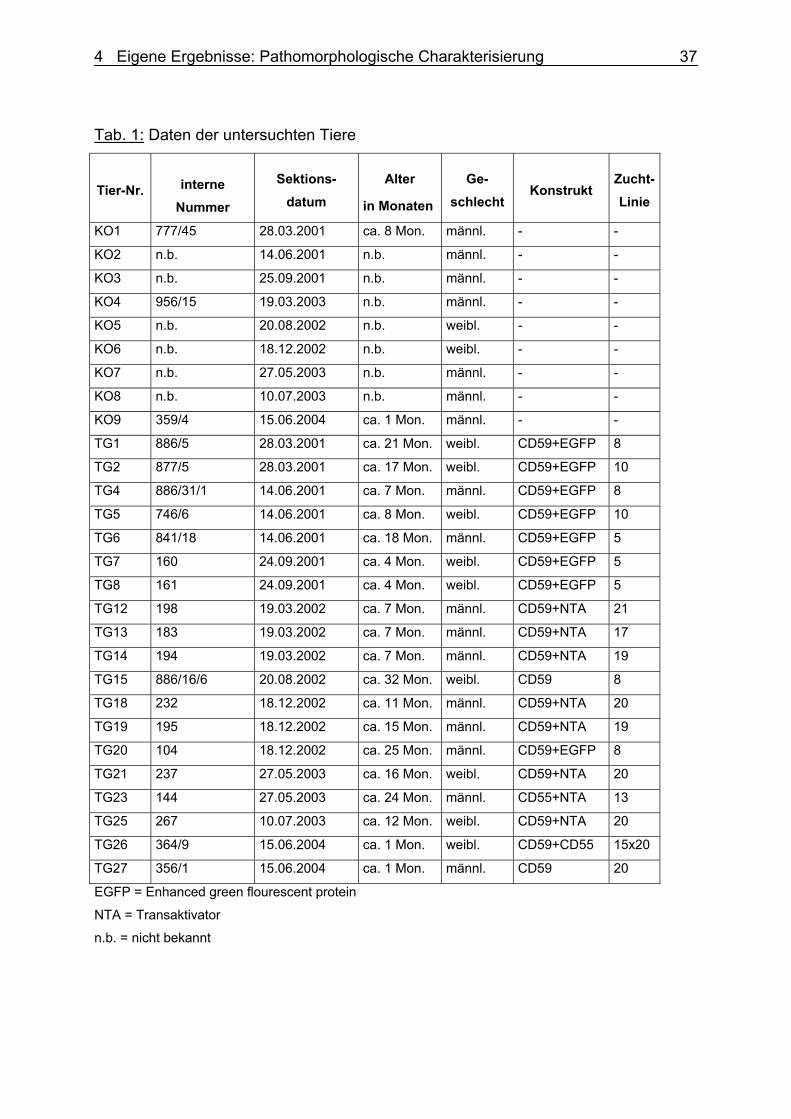
4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 37
Tab. 1: Daten der untersuchten Tiere
Tier-Nr.
interne
Nummer
Sektions-datum
Alter
in Monaten
Ge-schlecht
Konstrukt Zucht-Linie
KO1 777/45 28.03.2001 ca. 8 Mon. männl. - -
KO2 n.b. 14.06.2001 n.b. männl. - -
KO3 n.b. 25.09.2001 n.b. männl. - -
KO4 956/15 19.03.2003 n.b. männl. - -
KO5 n.b. 20.08.2002 n.b. weibl. - -
KO6 n.b. 18.12.2002 n.b. weibl. - -
KO7 n.b. 27.05.2003 n.b. männl. - -
KO8 n.b. 10.07.2003 n.b. männl. - -
KO9 359/4 15.06.2004 ca. 1 Mon. männl. - -
TG1 886/5 28.03.2001 ca. 21 Mon. weibl. CD59+EGFP 8
TG2 877/5 28.03.2001 ca. 17 Mon. weibl. CD59+EGFP 10
TG4 886/31/1 14.06.2001 ca. 7 Mon. männl. CD59+EGFP 8
TG5 746/6 14.06.2001 ca. 8 Mon. weibl. CD59+EGFP 10
TG6 841/18 14.06.2001 ca. 18 Mon. männl. CD59+EGFP 5
TG7 160 24.09.2001 ca. 4 Mon. weibl. CD59+EGFP 5
TG8 161 24.09.2001 ca. 4 Mon. weibl. CD59+EGFP 5
TG12 198 19.03.2002 ca. 7 Mon. männl. CD59+NTA 21
TG13 183 19.03.2002 ca. 7 Mon. männl. CD59+NTA 17
TG14 194 19.03.2002 ca. 7 Mon. männl. CD59+NTA 19
TG15 886/16/6 20.08.2002 ca. 32 Mon. weibl. CD59 8
TG18 232 18.12.2002 ca. 11 Mon. männl. CD59+NTA 20
TG19 195 18.12.2002 ca. 15 Mon. männl. CD59+NTA 19
TG20 104 18.12.2002 ca. 25 Mon. männl. CD59+EGFP 8
TG21 237 27.05.2003 ca. 16 Mon. weibl. CD59+NTA 20
TG23 144 27.05.2003 ca. 24 Mon. männl. CD55+NTA 13
TG25 267 10.07.2003 ca. 12 Mon. weibl. CD59+NTA 20
TG26 364/9 15.06.2004 ca. 1 Mon. weibl. CD59+CD55 15x20
TG27 356/1 15.06.2004 ca. 1 Mon. männl. CD59 20
EGFP = Enhanced green flourescent protein
NTA = Transaktivator
n.b. = nicht bekannt

4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 38
4.2.2 Pathologisch-anatomische und histopathologische Beurteilung Zur Untersuchung gelangten sowohl Foundertiere als auch Tiere der F1- und F2-
Generation.
Zum Zweck der pathologisch-anatomischen Untersuchung wurden die Schweine
mittels Elektroschock betäubt und durch Entblutung über die Vena jugularis getötet.
Anschließend wurden die Tiere seziert und alle Organe und Gewebe makroskopisch
beurteilt.
Für die histopathologische Untersuchung wurden Gewebeproben von folgenden
Organen und Geweben in 10%igem, gepufferten Formalin fixiert: Haut
(Innenschenkel), Skelettmuskulatur (M. gracilis), Herz (linke Ventrikelwand, rechte
Ventrikelwand, Septum), Lunge (Lobus cranialis, Lobus caudalis), Milz, Leber, Niere,
Harnblase, Pankreas, Gehirn (Kleinhirn, Großhirn), Schilddrüse, Lymphknoten (Ln.
pulmonalis, Ln. cervicalis superficialis, Lnn. ileofemorales und Lnn. mesenteriales),
Oesophagus, Magen, Duodenum, Jejunum, Ileum, Colon und Aorta.
Von den beiden Tieren KO8 und TG25 wurden direkt nach der Tötung der Tiere
Herz, Lunge, Milz, Leber und Niere über zuführende Arterien mit 4°C kalter
Natriumchloridlösung perfundiert, um die Blutgefäße von Plasma zu befreien.
Nach 24 Stunden wurden die Organ- und Gewebeproben in Paraffin eingebettet und
von jedem Paraffinblock wurde ein 4 µm dicker Paraffinschnitt auf Objektträgern
angefertigt, entparaffiniert und mit Hämatoxylin-Eosin (HE) gefärbt.
Die histopathologische Auswertung erfolgte an einem Lichtmikroskop (Axioskop, Fa.
Zeiss, Göttingen).
4.2.3 Virologische Untersuchung
Von allen untersuchten Tieren wurde natives Material vom Lobus caudalis der Lunge
und von den Lungenlymphknoten zu Laboklin (Bad Kissingen) geschickt und dort
mittels PCR auf Porzines Circovirus Typ II (PCV II) und Porzines Respiratorisches
und Reproduktives Syndrom Virus (PRRSV) untersucht.

4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 39
4.3 Ergebnisse und Interpretation
4.3.1 Virologische Untersuchung Es wurden 8 nicht-transgene Kontrolltiere und 17 transgene Tiere der Studie mittels
PCR auf PCV II und PRRSV untersucht.
Davon waren 6 Kontrolltiere (entspricht 75%) und 11 transgene Tiere (entspricht
65%) positiv für PCV II.
PRRSV konnte bei keinem untersuchten Tier nachgewiesen werden.
Die Tabelle 2 gibt einen Überblick über die PCV II – Untersuchungsergebnisse.
Tab. 2: PCV II – Untersuchungsergebnisse
Nicht-transgene Kontrolltiere Transgene Tiere PCV II- positiv KO1, KO2, KO4, KO6, KO7,
KO8 TG1, TG2, TG4, TG5, TG6, TG8, TG18, TG19, TG20, TG21, TG23
PCV II- negativ KO3, KO5 TG7, TG12, TG13, TG14, TG15, TG25
4.3.2 Pathomorphologische Untersuchung
In der pathomorphologischen Untersuchung konnte kein spezifischer Phänotyp für
die hCD59-transgenen Tiere nachgewiesen werden.
Allerdings wurden sowohl bei den nicht-transgenen Kontrolltieren als auch bei den
transgenen Tieren in verschiedenen Organen Läsionen (siehe Tab. 3 und Tab. 4)
festgestellt, die jedoch auf Grund der Tatsache, dass transgene Tiere und nicht-
transgene Kontrolltiere in gleicher Zahl und im gleichen Ausmaß betroffen waren, als
unspezifische Hintergrundpathologie interpretiert wurden.
Dabei ließen sich zwei Arten von Befunden differenzieren:
Einerseits lagen Befunde vor, die lediglich bei Einzeltieren diagnostiziert wurden, wie
z.B. Pneumonie (siehe Abb. 2), Epi- und Perikarditis, mesenteriale Verknöcherungen
(siehe Abb. 3) oder Nierenzysten. Von diesen Einzeltierbefunden waren die nicht-
transgenen Kontrolltiere und die transgenen Tieren in gleichem Ausmaß betroffen.
Eine genaue Auflistung dieser Einzeltierbefunde ist in Tab. 3 dargestellt.
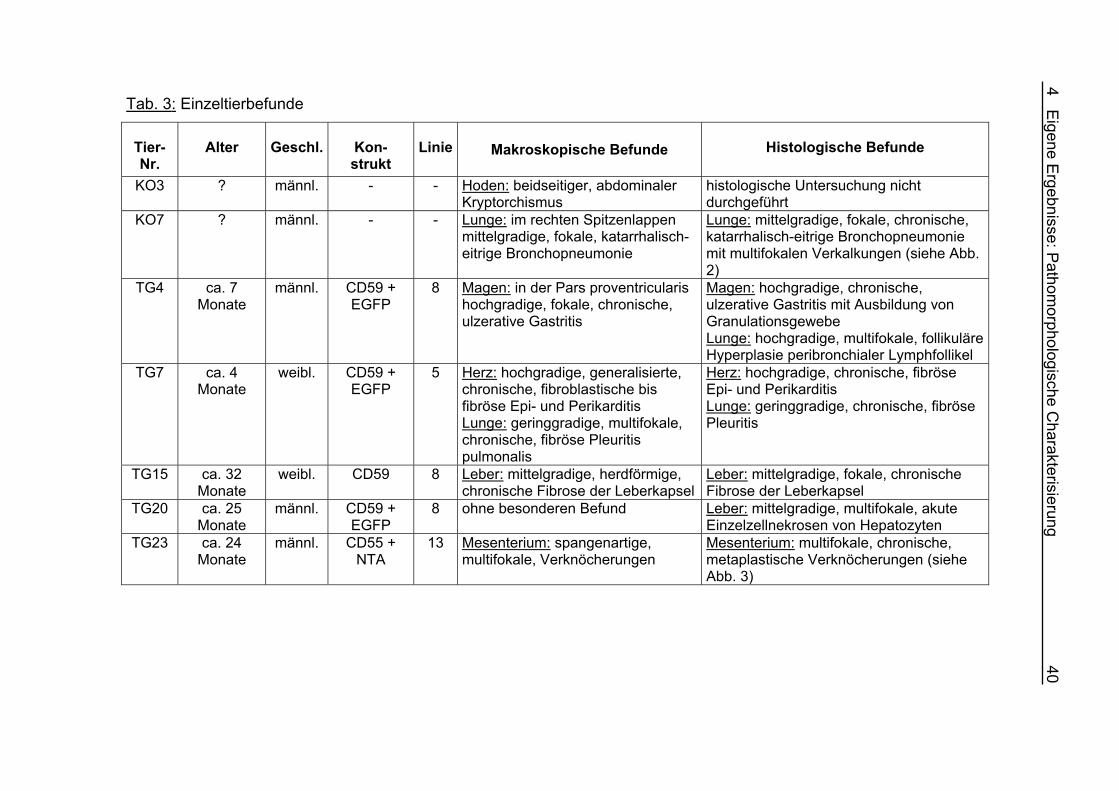
4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 40
Tab. 3: Einzeltierbefunde
Tier-Nr.
Alter Geschl. Kon-strukt
Linie Makroskopische Befunde Histologische Befunde
KO3 ? männl. - - Hoden: beidseitiger, abdominaler Kryptorchismus
histologische Untersuchung nicht durchgeführt
KO7 ? männl. - - Lunge: im rechten Spitzenlappen mittelgradige, fokale, katarrhalisch-eitrige Bronchopneumonie
Lunge: mittelgradige, fokale, chronische, katarrhalisch-eitrige Bronchopneumonie mit multifokalen Verkalkungen (siehe Abb. 2)
TG4 ca. 7Monate
männl. CD59 +EGFP
8 Magen: in der Pars proventricularis hochgradige, fokale, chronische, ulzerative Gastritis
Magen: hochgradige, chronische, ulzerative Gastritis mit Ausbildung von Granulationsgewebe Lunge: hochgradige, multifokale, follikuläre Hyperplasie peribronchialer Lymphfollikel
TG7 ca. 4Monate
weibl. CD59 +EGFP
5 Herz: hochgradige, generalisierte, chronische, fibroblastische bis fibröse Epi- und Perikarditis Lunge: geringgradige, multifokale, chronische, fibröse Pleuritis pulmonalis
Herz: hochgradige, chronische, fibröse Epi- und Perikarditis Lunge: geringgradige, chronische, fibröse Pleuritis
TG15 ca. 32Monate
weibl. CD59 8 Leber: mittelgradige, herdförmige, chronische Fibrose der Leberkapsel
Leber: mittelgradige, fokale, chronische Fibrose der Leberkapsel
TG20 ca. 25Monate
männl. CD59 +EGFP
8 ohne besonderen Befund Leber: mittelgradige, multifokale, akute Einzelzellnekrosen von Hepatozyten
TG23 ca. 24Monate
männl. CD55 +NTA
13 Mesenterium: spangenartige, multifokale, Verknöcherungen
Mesenterium: multifokale, chronische, metaplastische Verknöcherungen (siehe Abb. 3)
4 E
igene Ergebnisse: P
athomorphologische C
harakterisierung 40

4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 41
Zum anderen lagen Befunde vor, die bei mehreren Tieren auftraten und überwiegend
erst in der histologischen Untersuchung auffielen, wie z.B. follikuläre Hyperplasie in
Lymphknoten, Balantidium coli-Befall oder Eisen-Depositionen in den
Ileofemorallymphknoten.
Diese Befunde waren stets lediglich in geringem Grad ausgeprägt und betrafen
wiederum die Kontrolltiere und die transgenen Tiere in gleicher Zahl und in gleichem
Ausmaß. Eine genaue Auflistung der Befunde, die bei mehreren Tieren auftraten, ist
in Tabelle 4 dargestellt.
Eine Auswahl der Läsionen ist in Abb. 4-7 illustriert.
Insgesamt fanden sich keine Hinweise auf einen spezifischen,
pathomorphologischen Phänotyp in den transgenen Tieren. Die Läsionen, die in der
makroskopischen und histologischen Untersuchung diagnostiziert wurden, stellten
lediglich Einzeltierbefunde dar, eine Häufung von spezifischen Befunden konnte
dabei nicht beobachtet werden.
Auch die Läsionen, die in mehreren Tieren beobachtet wurden, waren nicht nur bei
den transgenen Schweinen vorhanden, sondern wurden bei transgenen Tieren und
Kontrolltieren in gleicher Zahl und in gleichem Ausmaß vorhanden.

4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 42
Tab. 4: Befunde bei mehreren Tieren Organ Histologischer Befund Betroffene Tiere Betroffene
Tiere (%) Haut geringgradige, multifokale, perivaskuläre
Infiltration mit Lymphozyten, Plasmazellen und einzelnen eosionophilen Granulozyten (siehe Abb. 4)
KO1, KO2, KO6, KO7 TG1, TG2, TG5, TG15, TG18, TG19, TG20, TG21
44% KO 47% TG
Leber geringgradige, multifokale, herdförmige Infiltration mit Lymphozyten, Makrophagen und einzelnen neutrophilen Granulozyten (siehe Abb. 5)
KO3, KO4, KO6, KO7, KO8 TG7, TG8, TG12, TG13, TG14, TG15, TG18, TG19, TG20, TG21, TG25
56% KO 58% TG
Niere geringgradige, multifokale, chronische, nichteitrige, interstitielle Nephritis
KO7 TG13, TG20, TG21
11% KO 16% TG
Colon im Lumen geringgradiger Nachweis von Balantidium coli (reaktionslos) (siehe Abb. 6)
KO2 TG5, TG6, TG7, TG18
11% KO 21% TG
Milz gering- bis mittelgradige, multifokale, follikuläre Hyperplasie
KO3
11% KO
multifokale, akute, frische Blutungen KO6, KO7 TG18, TG19, TG22, TG23, TG26
22% KO 26% TG
follikuläre Hyperplasie mit Follikelzentrumsreaktion
KO1, KO2, KO8 TG1, TG2, TG4, TG5, TG13, TG14, TG23, TG26
33% KO 42% TG
Ln. pulmo-nalis
Ödem und Sinushistiozytose KO4, KO6, KO7 TG5, TG7, TG8, TG13, TG15, TG18, TG19, TG21
33% KO 42% TG
follikuläre Hyperplasie mit Follikelzentrumsreaktion
KO2, KO3, KO4, KO5, KO6, KO8 TG4, TG5, TG7, TG12, TG13, TG15, TG18, TG19, TG20, TG21, TG23, TG25
67% KO 63% TG
Lnn. mesente-
riales
Ödem und Sinushistiozytose KO4, KO7, KO8, KO9 TG4, TG8, TG13, TG14, TG18, TG19, TG21, TG25, TG26
44% KO 47% TG
multifokale, akute, frische Blutungen TG18, TG19, TG23 16% TG follikuläre Hyperplasie mit Follikelzentrumsreaktion
KO2, KO4 TG4, TG12, TG13, TG14, TG20
22% KO 26% TG
Ödem und Sinushistiozytose KO2, KO3, KO5, KO6, KO7, KO9 TG4, TG5, TG6, TG7, TG8, TG13, TG14, TG26, TG27
66% KO 47% TG
Lnn. ileofe-morales
Eisen-Depositionen frei oder in Makrophagen in den Sinus (Berliner-Blau positiv) (siehe Abb. 7)
KO3, KO5, KO6, KO7, KO9 TG7, TG8, TG15, TG19, TG20, TG23, TG26, TG27
55% KO 42% TG
follikuläre Hyperplasie mit Follikelzentrumsreaktion
KO2 TG4
11% KO 5% TG
Ln. cervicalis
cran. superf.
Ödem und Sinushistiozytose KO2, KO4 TG5, TG7, TG8
22% KO 16% TG
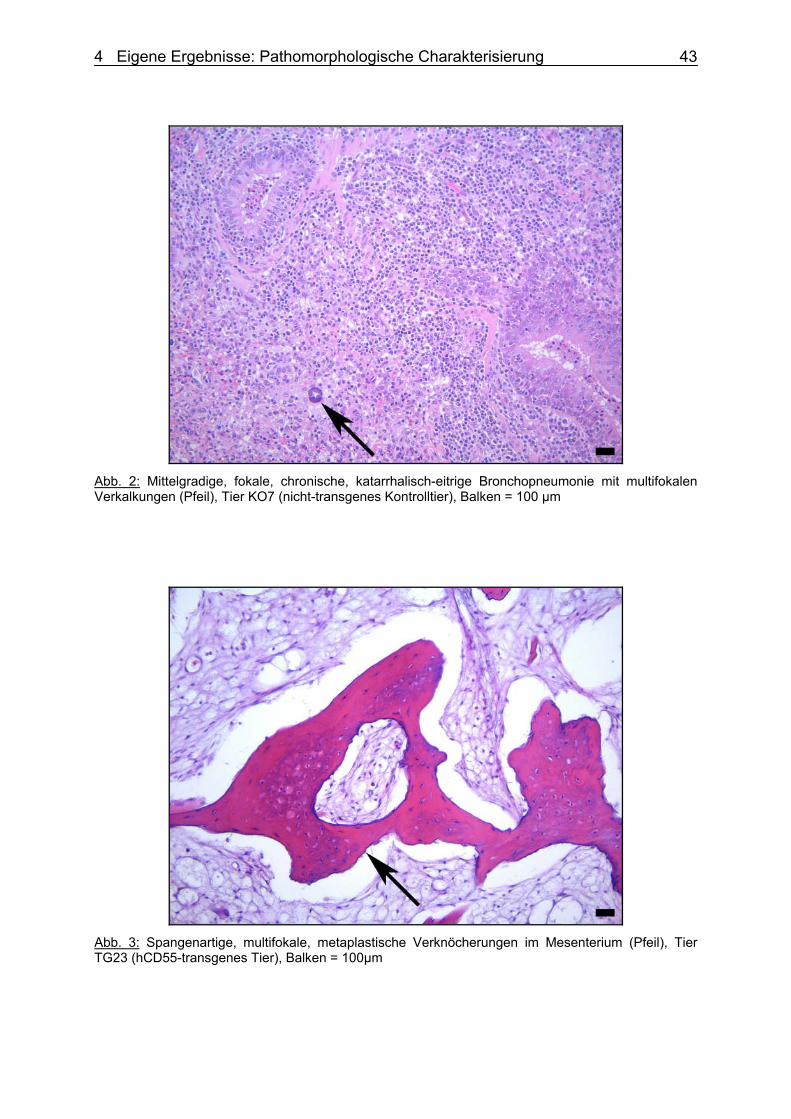
4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 43
Abb. 2: Mittelgradige, fokale, chronische, katarrhalisch-eitrige Bronchopneumonie mit multifokalen Verkalkungen (Pfeil), Tier KO7 (nicht-transgenes Kontrolltier), Balken = 100 µm
Abb. 3: Spangenartige, multifokale, metaplastische Verknöcherungen im Mesenterium (Pfeil), Tier TG23 (hCD55-transgenes Tier), Balken = 100µm
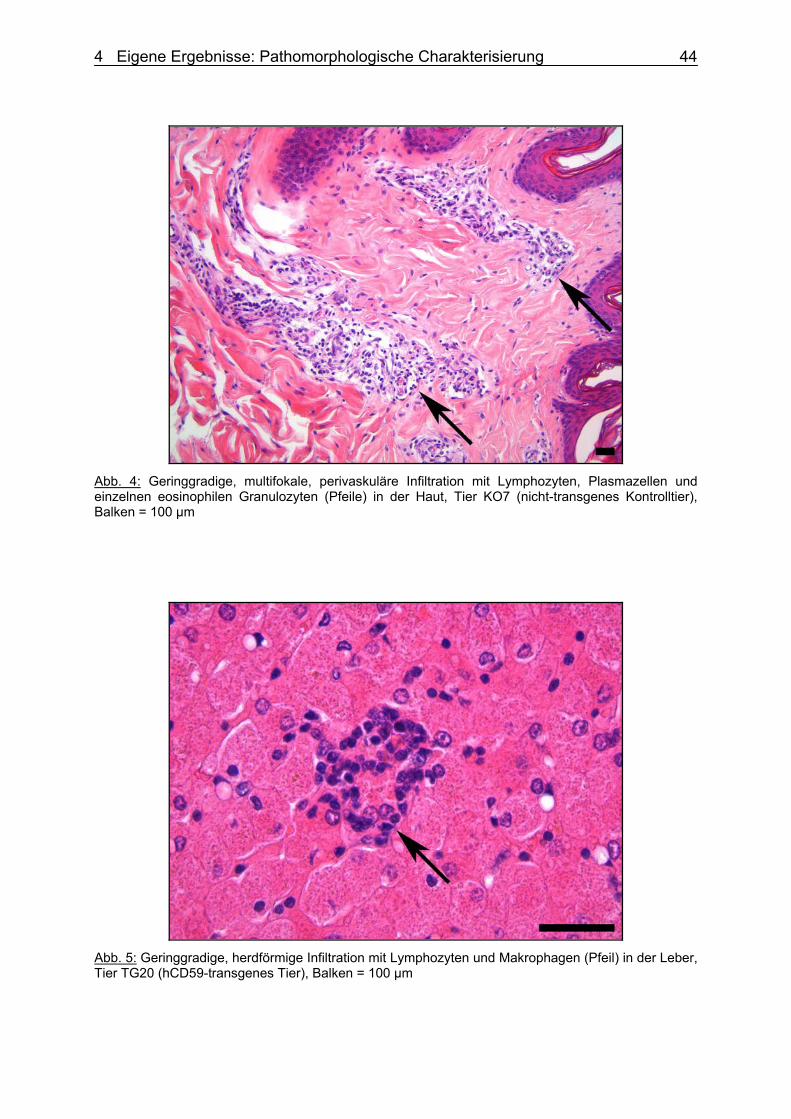
4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 44
Abb. 4: Geringgradige, multifokale, perivaskuläre Infiltration mit Lymphozyten, Plasmazellen und einzelnen eosinophilen Granulozyten (Pfeile) in der Haut, Tier KO7 (nicht-transgenes Kontrolltier), Balken = 100 µm
Abb. 5: Geringgradige, herdförmige Infiltration mit Lymphozyten und Makrophagen (Pfeil) in der Leber, Tier TG20 (hCD59-transgenes Tier), Balken = 100 µm
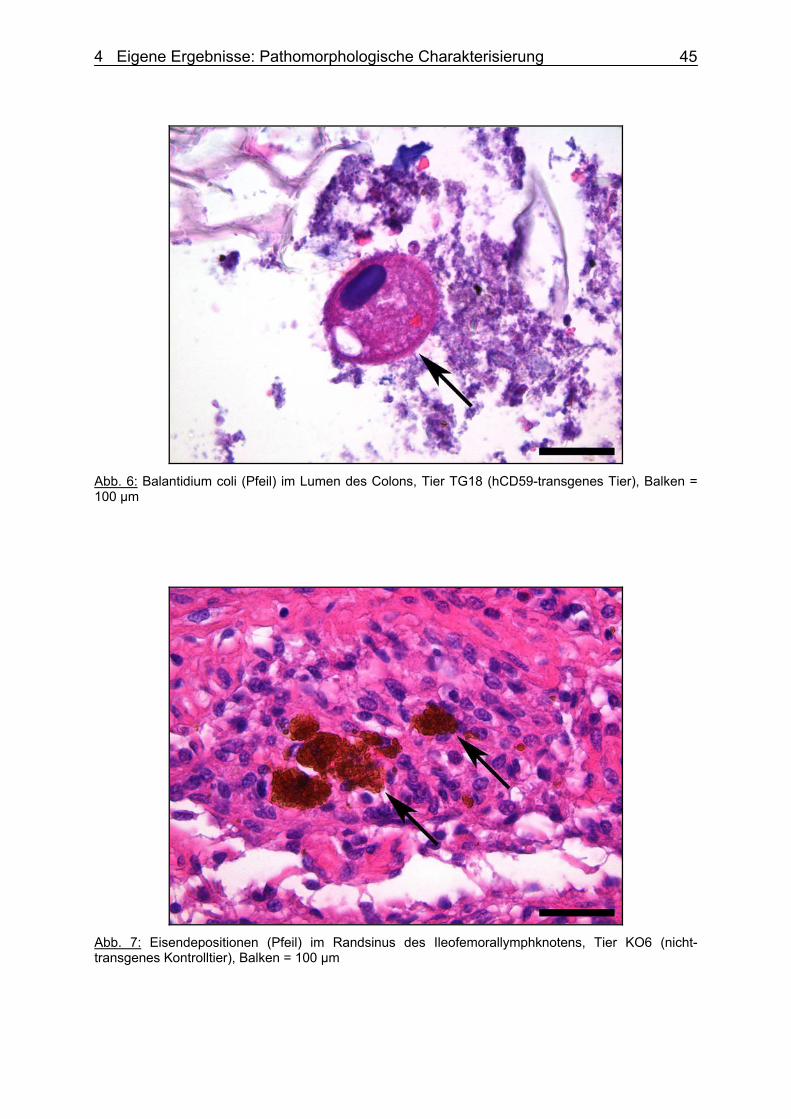
4 Eigene Ergebnisse: Pathomorphologische Charakterisierung 45
Abb. 6: Balantidium coli (Pfeil) im Lumen des Colons, Tier TG18 (hCD59-transgenes Tier), Balken = 100 µm
Abb. 7: Eisendepositionen (Pfeil) im Randsinus des Ileofemorallymphknotens, Tier KO6 (nicht-transgenes Kontrolltier), Balken = 100 µm

46
5 Eigene Ergebnisse: hCD59 - Proteinexpression 5.1 Hintergrund und Bedeutung Die transgenen Tiere tragen das hCD59-Konstrukt, um im Fall einer
Xenotransplantation die hyperakute Abstoßung über eine Hemmung der
Komplementkaskade zu verhindern. CD59 wird normalerweise auf Blutzellen,
Epithelzellen und Endothelzellen exprimiert. Um einen protektiven Effekt gegen die
hyperakute Abstoßung zu erreichen, muss das hCD59-Protein in den transgenen
Organen, vor allem im Endothel, exprimiert werden.
Die Untersuchung der hCD59-Proteinexpression wurde durchgeführt, um
festzustellen, ob das hCD59-Protein exprimiert wird, in welchen Organen und in
welchen Zellen eine Expression vorliegt und ob die Expression konstant in allen
Organen und Tieren aller Linien vorhanden ist.
Dabei war vor allem der letztgenannte Punkt von Interesse, da durch die Herstellung
der Tiere per Mikroinjektion die Möglichkeit bestand, dass jedes Tier einen anderen
Genotyp und damit ein anderes Expressionsmuster aufweist. Zudem war zu klären,
ob der gewählte CMV-Promotor zu einer ubiquitären Expression in allen Organen
führt, oder ob er eine Gewebsspezifität besitzt.
5.2 Experimentelles Design 5.2.1 Material Für die Expressionsanalyse des hCD59-Proteins wurde eine immunhistochemische
Untersuchung mit einem monoklonalen Maus-anti-Human CD59 – Antikörper (Best.-
Nr. 33861A, Klon p282 (H19), Fa. BD PharMingen, Heidelberg) durchgeführt. Alle 19
transgenen und 9 nicht-transgenen Kontrolltiere wurden untersucht. Als
Ausgangsmaterial diente formalin-fixiertes, paraffin-eingebettetes Material von Haut,
Skelettmuskulatur, Herz, Lunge, Milz, Leber, Niere, Harnblase, Pankreas, Gehirn,
Schilddrüse, Lymphknoten (Ln. pulmonalis, Ln. cervicalis superficialis, Lnn.

5 Eigene Ergebnisse: hCD59 - Proteinexpression 47
ileofemorales und Lnn. mesenteriales), Oesophagus, Magen, Duodenum, Jejunum,
Ileum, Colon und Aorta.
Als Positivkontrolle diente humanes Tonsillargewebe. Als Negativkontrolle diente
Gewebe von nicht-transgenen Kontrolltieren und dem hCD55-transgenen (TG23)
Tier.
5.2.2 Herstellung der Multiblöcke Für die immunhistochemische Expressionsanalyse wurden Multiblöcke hergestellt.
Dafür wurden aus den Paraffinblöcken aus vorher markierten Lokalisationen mit
einer Stanze 0,6 mm große Gewebestanzen entnommen und in eine
Paraffinschablone eingesetzt, die dann zu einem Paraffinblock eingeschmolzen
wurde (VON WASIELEWSKI et al., 2002). Pro Organ und Gewebe wurden 2
Stanzproben entnommen. In jedem Multiblock war ein komplettes Spektrum an
Organ- und Gewebeproben eines Tieres enthalten.
5.2.3 Durchführung der immunhistochemischen Färbung An den Multiblöcken wurde eine immunhistochemische Untersuchung mit einem
monoklonalen Maus-anti-Human CD59-Antikörper (Fa. BD PharMingen, Heidelberg)
nach der Tyramin-Amplifikations-Methode (MENGEL et al., 1999) durchgeführt.
Zu diesem Zweck wurden 4 µm dicke Paraffinschnitte von den Multiblöcken auf
Super Frost blau – Objektträger (Fa. Omnilab, Bremen) gezogen. Die Objektträger
wurden für 15 Minuten bei 60°C in einen Wärmeschrank (Heraeus UT 5042 E, Fa.
Heraeus, Hanau) verbracht. Anschließend erfolgte eine Entparaffinierung in Xylol für
2 mal 10 Minuten auf einem Schüttler, dann wurden die Schnitte in einer
absteigenden Alkoholreihe (2x100%ig, 2x95%ig, 1x 70%ig, 1x 50%ig, 1x 20%ig, 3
Minuten je Konzentration) dehydriert und einmalig mit entionisiertem Wasser gespült.
Die Antigendemaskierung erfolgte durch eine 10minütige Inkubation mit Protease
XXIV (21.4 mg auf 50 ml Aqua dest, Fa. Sigma-Aldrich, München) im Wärmeschrank
bei 37°C. Die endogene Peroxidase wurde durch eine 10minütige Inkubation mit
3%igem H2O2 (in Tris/HCl pH 7.5, Ansatz siehe Anhang) blockiert. Eine Hemmung
von unspezifischen Bindungen erfolgte durch eine 10minütige Behandlung mit

5 Eigene Ergebnisse: hCD59 - Proteinexpression 48
Casein (0.5% in Tris/HCl (Ansatz siehe Anhang) Fa. Sigma, München). Um eine
unspezifische Bindung an endogenes Biotin zu hemmen, wurden die Schnitte je 10
Minuten mit Avidin und Biotin (Avidin/Biotin Blocking Kit, Fa. Vector, Burlingame,
USA) inkubiert. Anschließend wurde der monoklonale Maus-anti-Human CD59-
Antikörper (Best.-Nr. 33861A, Klon p282 (H19), Fa. BD PharMingen, Heidelberg) in
der Verdünnung 1:200 (in Bovinem Serumalbumin, BSA/Tris pH 7.6, Ansatz siehe
Anhang) aufgetragen über Nacht bei 4°C inkubiert.
Als Sekundärantikörper wurde ein biotinylierter Ziege-anti-Maus-Antikörper (Fa.
Zytomed, Berlin) in der Verdünnung 1:900 (in Bovinem Serumalbumin, BSA/Tris pH
7.6, Ansatz siehe Anhang) für eine halbe Stunde bei 37°C appliziert. Anschließend
wurden die Schnitte für 30 Minuten mit Horseradish Peroxidase-Komplex (Fa.
Zytomed, Berlin) in der Verdünnung 1:500 in BSA/Tris pH 7.6 (Ansatz siehe Anhang)
inkubiert. Zur Verstärkung der Reaktion erfolgte daraufhin eine Applikation von
biotinyliertem Tyramin mit 0.003%igem H2O2 für 10 Minuten. Dafür wurde eine
Stammlösung aus 6.2 mg Tyramin hydrochlorid (Fa. Sigma, München), 8 ml 50 mM
Boratpuffer pH 8.0 und 20 mg Sulpho-NHS-LC-Biotin (Fa. Pierce, Rockford, USA)
hergestellt und aliquotiert. Von der Stammlösung wurde eine Gebrauchslösung aus
1800 µl Tris/HCl-Puffer pH 7.5 (Ansatz siehe Anhang), 10 µl Tyramin-Stammlösung
und 5.4 µl 3% H2O2 angesetzt und verwandt. Anschließend erfolgte eine 30minütige
Inkubation mit Avidin-biotinylierter alkalischer Phosphatase (Fa. Zytomed, Berlin) in
der Verdünnung 1:300 in Tris-Puffer, pH 7.8 (Ansatz siehe Anhang) im
Wärmeschrank bei 37°C. Zur Anfärbung wurde das Chromogen Fast Red (1 mg
Fast Red Salz auf 1ml Fast Red Puffer, Fa. Sigma-Aldrich, München) verwandt und
für 8 Minuten auf die Schnitte aufgetragen. Anschließend wurden die Schnitte mit
entionisiertem Wasser gewaschen, mit Mayer’s Hämalaun (1:3 verdünnt in
entionisiertem Wasser, Fa. Merck, Darmstadt) für 2 Minuten gegengefärbt, in
fließendem Leitungswasser für 10 Minuten gebläut und anschließend mit 37°C
warmer Kayser’s Glycerin Gelantine (Fa. Merck, Darmstadt) eingedeckt.
Zwischen den einzelnen Reaktionsschritten wurden die Schnitte mit Tris-Puffer pH
7.8 (Ansatz siehe Anhang) gewaschen.

5 Eigene Ergebnisse: hCD59 - Proteinexpression 49
5.2.4 Auswertung Die Auswertung der immunhistochemischen Färbung erfolgte semiquantitativ an
einem Lichtmikroskop (Axioskop, Fa. Zeiss, Göttingen). Dazu wurde ein Score von –
bis +++ benutzt, bei dem eine negative Reaktion mit – bewertet wurde, eine schwach
positive Reaktion in einzelnen (bis ca. 30%) Zellen mit +, eine mittelstark positive
Reaktion in ca. 30-60% der Zellen mit ++ und eine stark positive Reaktion in ca. 60-
100% der Zellen mit +++. Dabei wurde die Art der angefärbten Zellen sowie der
Anteil der positiven Zellen beurteilt. Zeigten Organe oder Gewebe in den
Multiblöcken eine positive Reaktion, so wurde die immunhistochemische Färbung
nochmals an größeren Organ- und Gewebeschnitten durchgeführt, um das
Verteilungsmuster der angefärbten Zellen besser beurteilen zu können. Alle
immunhistochemischen Färbungen und Untersuchungen wurden zweimal
durchgeführt, um eine Reproduzierbarkeit der Ergebnisse zu gewährleisten.
5.3 Ergebnisse und Interpretation 5.3.1 hCD59-Proteinexpression Von den 18 untersuchten, hCD59-transgenen Tieren zeigten 13 Tiere eine
Expression des hCD59-Proteins in verschiedenen Organen und Geweben. Dabei
war auffällig, dass keines der Tiere eine hCD59-Proteinexpression in allen Organen
aufwies. Vielmehr zeigten alle untersuchten Tiere die Proteinexpression nur in einem
Teil der Organe und Gewebe. Dabei zeigte sich eine starke Variation der
exprimierenden Organe zwischen den einzelnen Tieren.
Weiterhin ließ sich das bemerkenswerte Phänomen beobachten, dass in den
Kardiomyozyten, der Niere, den Skelettmuskelzellen und der Tunica muscularis des
Ösophagus die hCD59-Proteinexpression ein so genanntes „patchy pattern“ aufwies.
Dieses Muster war dadurch gekennzeichnet, dass sich innerhalb des Gewebes nur
einzelne Zellen anfärbten, während direkt benachbarte Zellen keine Anfärbung
zeigten.

5 Eigene Ergebnisse: hCD59 - Proteinexpression 50
Folgende Zelltypen zeigten eine intrazytoplasmatische, positive Reaktion für das
hCD59-Protein:
In Haut, Herz (siehe Abb. 8), Lunge, Milz, Leber, Niere (siehe Abb. 10), Harnblase,
Gehirn (siehe Abb. 9), Skelettmuskulatur, Ösophagus, Duodenum, Jejunum, Ileum
und Colon war die hCD59-Proteinexpression im Zytoplasma von Endothelzellen
kleiner und mittelgroßer Gefäße nachweisbar.
Das Pankreas einiger Tiere wies eine positive Reaktion im Zytoplasma von Azinus-
Zellen des exokrinen Pankreas und Inselzellen des endokrinen Pankreas auf, wobei
die Azinus-Zellen des exokrinen Pankreas von Tier TG15 ein „patchy pattern“
aufwiesen (siehe Abb. 14).
Im Herz zeigte sich bei einem Tier (TG7) zudem eine Anfärbung des randständigen
Zytoplasmas einzelner Kardiomyozyten („patchy pattern“) (siehe Abb. 12).
In der Niere eines Tieres (TG1) zeigten einzelne Glomerula in den Endothelzellen
der Kapillarschlingen und afferenten Arterien eine positive Anfärbung (siehe Abb.
10). Dabei wiesen aber nur einzelne Glomerula eine Anfärbung auf („patchy
pattern“).
In der Skelettmuskulatur und der Tunica muscularis des Ösophagus einiger Tiere
wiesen die Myozyten ebenfalls eine zytoplasmatische Reaktion im bereits
beschriebenen „patchy pattern“ auf (siehe Abb. 13).
Die Lunge zeigte zudem eine positive Anfärbung der Alveolarepithelzellen Typ I und
Typ II (siehe Abb. 11).
Eine genaue Beschreibung der hCD59-Proteinexpression ist in Tabelle 5 dargestellt.
Bei allen untersuchten nicht-transgenen Kontrolltieren und dem hCD55-transgenen
Tier konnte immunhistochemisch keine hCD59-Proteinexpression nachgewiesen
werden. Dies zeigte, dass der gewählte Antikörper spezifisch für humanes hCD59 ist
und nicht mit porzinem CD59 kreuzreagierte.
Das humane Tonsillargewebe zeigte eine positive Reaktion.
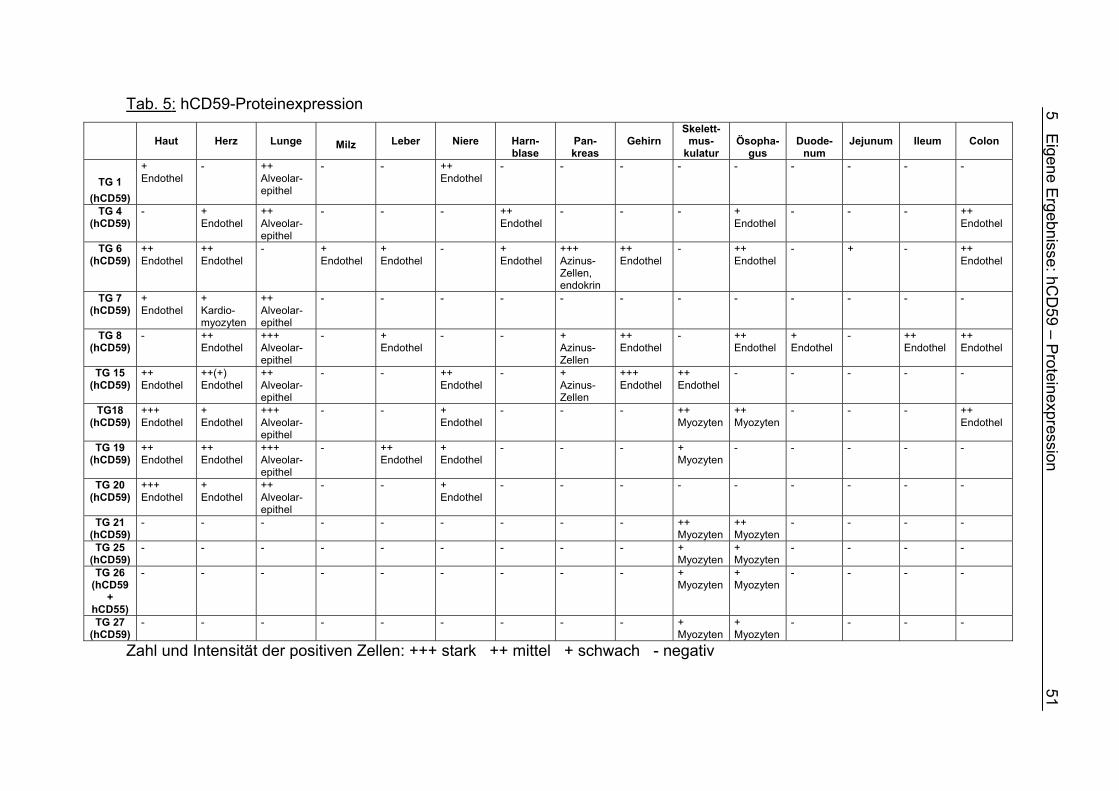
5 Eigene Ergebnisse: hCD59 - Proteinexpression 51
Tab. 5: hCD59-Proteinexpression 5 Eigene E
rgebnisse: hCD
59 – Proteinexpression 51
Haut
Herz
Lunge Milz
Leber
Niere
Harn-blase
Pan-kreas
Gehirn
Skelett-mus-
kulatur
Ösopha-
gus
Duode-
num
Jejunum
Ileum
Colon
TG 1 (hCD59)
+ Endothel
- ++Alveolar- epithel
- - ++Endothel
- - - - - - - - -
TG 4 (hCD59)
- +Endothel
++ Alveolar-epithel
- - - ++Endothel
- - - +Endothel
- - - ++Endothel
TG 6 (hCD59)
++ Endothel
++ Endothel
- + Endothel
+ Endothel
- +Endothel
+++ Azinus-Zellen, endokrin
++ Endothel
- ++Endothel
- + - ++Endothel
TG 7 (hCD59)
+ Endothel
+ Kardio- myozyten
++ Alveolar-epithel
- - - - - - - - - - - -
TG 8 (hCD59)
- ++ Endothel
+++ Alveolar-epithel
- +Endothel
- - +Azinus-Zellen
++ Endothel
- ++Endothel
+ Endothel
- ++Endothel
++ Endothel
TG 15 (hCD59)
++ Endothel
++(+) Endothel
++ Alveolar-epithel
- - ++Endothel
- + Azinus-Zellen
+++ Endothel
++ Endothel
- - - - -
TG18 (hCD59)
+++ Endothel
+ Endothel
+++ Alveolar-epithel
- - +Endothel
- - - ++Myozyten
++ Myozyten
- - - ++Endothel
TG 19 (hCD59)
++ Endothel
++ Endothel
+++ Alveolar-epithel
- ++Endothel
+ Endothel
- - - +Myozyten
- - - - -
TG 20 (hCD59)
+++ Endothel
+ Endothel
++ Alveolar-epithel
- - +Endothel
- - - - - - - - -
TG 21 (hCD59)
- - - - - - - - - ++Myozyten
++ Myozyten
- - - -
TG 25 (hCD59)
- - - - - - - - - + Myozyten
+ Myozyten
- - - -
TG 26 (hCD59
+ hCD55)
- - - - - - - - - + Myozyten
+ Myozyten
- - - -
TG 27 (hCD59)
- - - - - - - - - +Myozyten
+ Myozyten
- - - -
Zahl und Intensität der positiven Zellen: +++ stark ++ mittel + schwach - negativ
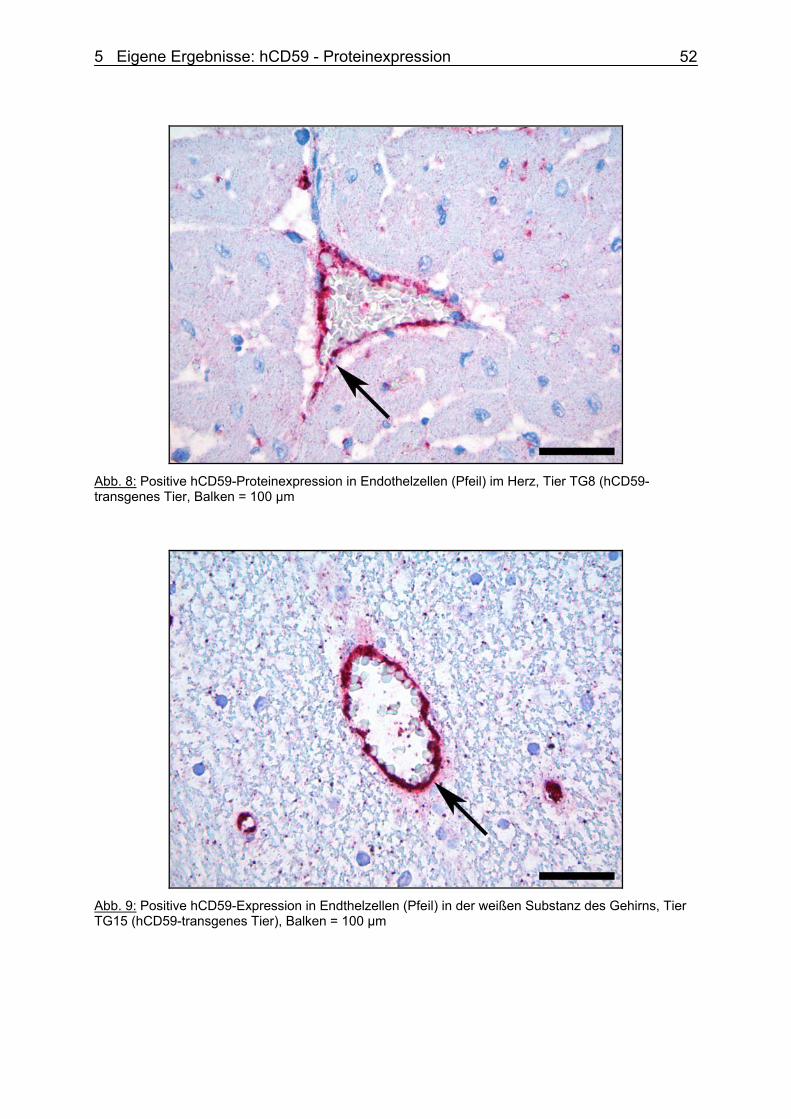
5 Eigene Ergebnisse: hCD59 - Proteinexpression 52
Abb. 8: Positive hCD59-Proteinexpression in Endothelzellen (Pfeil) im Herz, Tier TG8 (hCD59-transgenes Tier, Balken = 100 µm
Abb. 9: Positive hCD59-Expression in Endthelzellen (Pfeil) in der weißen Substanz des Gehirns, Tier TG15 (hCD59-transgenes Tier), Balken = 100 µm
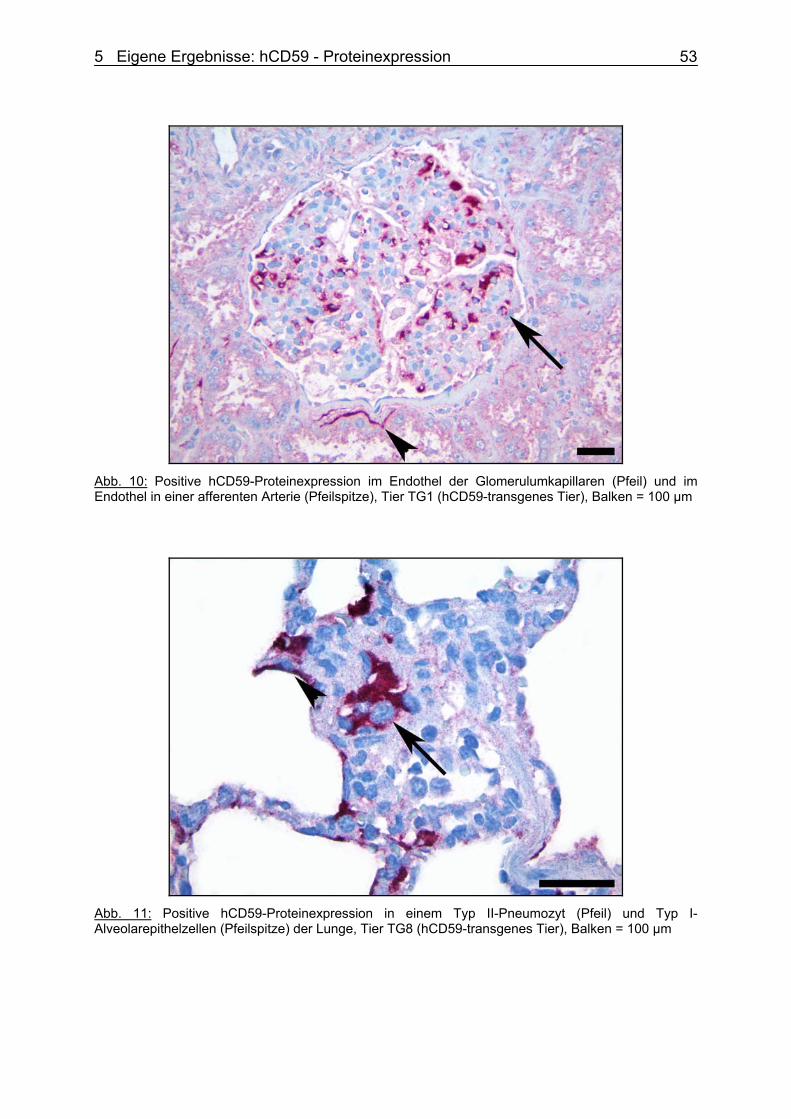
5 Eigene Ergebnisse: hCD59 - Proteinexpression 53
Abb. 10: Positive hCD59-Proteinexpression im Endothel der Glomerulumkapillaren (Pfeil) und im Endothel in einer afferenten Arterie (Pfeilspitze), Tier TG1 (hCD59-transgenes Tier), Balken = 100 µm
Abb. 11: Positive hCD59-Proteinexpression in einem Typ II-Pneumozyt (Pfeil) und Typ I-Alveolarepithelzellen (Pfeilspitze) der Lunge, Tier TG8 (hCD59-transgenes Tier), Balken = 100 µm
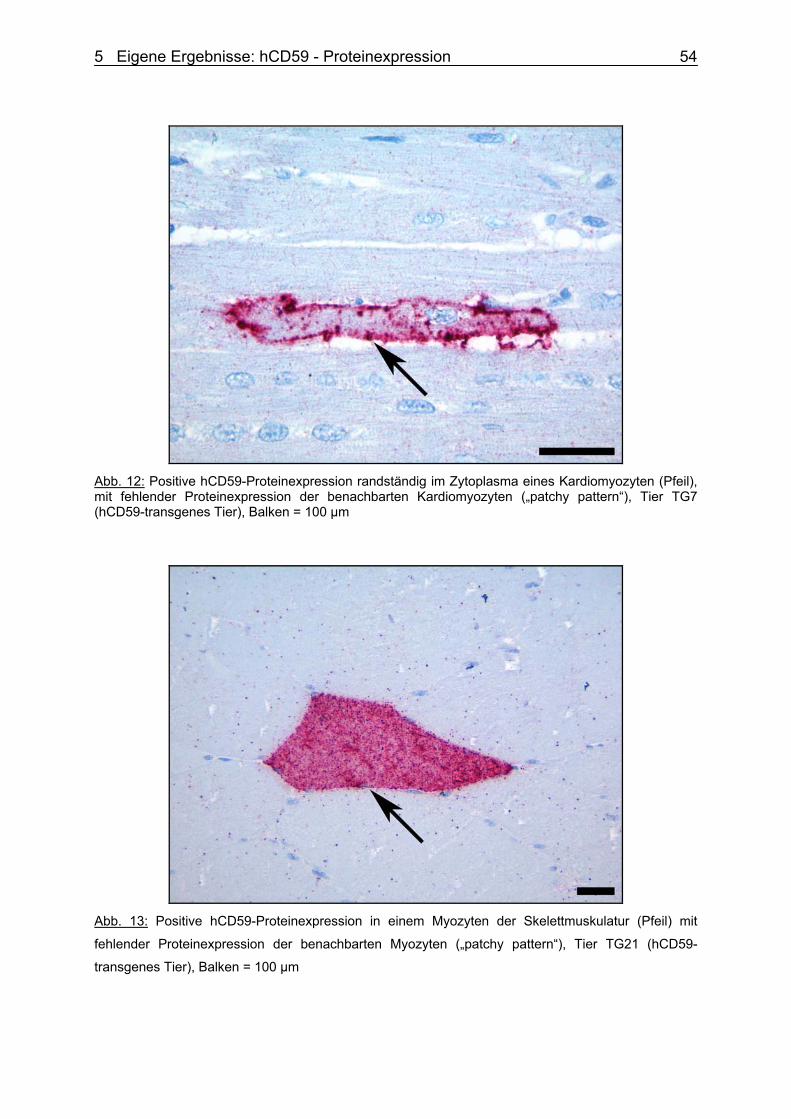
5 Eigene Ergebnisse: hCD59 - Proteinexpression 54
Abb. 12: Positive hCD59-Proteinexpression randständig im Zytoplasma eines Kardiomyozyten (Pfeil), mit fehlender Proteinexpression der benachbarten Kardiomyozyten („patchy pattern“), Tier TG7 (hCD59-transgenes Tier), Balken = 100 µm
Abb. 13: Positive hCD59-Proteinexpression in einem Myozyten der Skelettmuskulatur (Pfeil) mit
fehlender Proteinexpression der benachbarten Myozyten („patchy pattern“), Tier TG21 (hCD59-
transgenes Tier), Balken = 100 µm
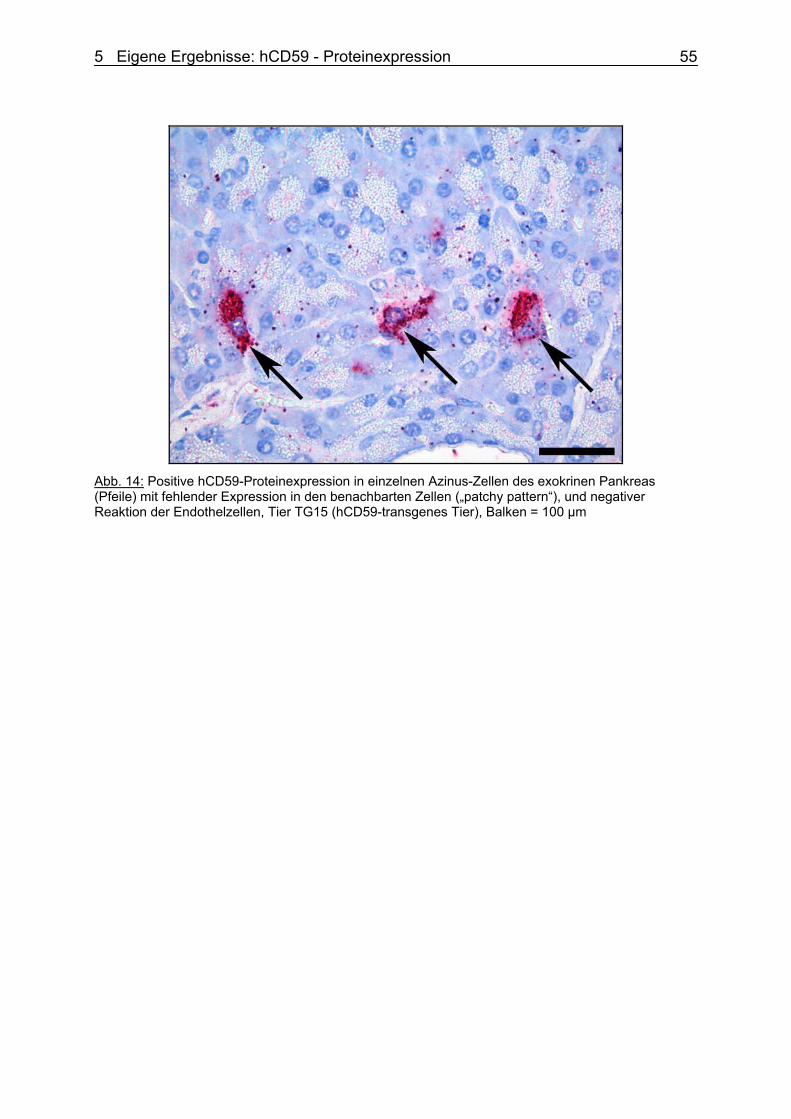
5 Eigene Ergebnisse: hCD59 - Proteinexpression 55
Abb. 14: Positive hCD59-Proteinexpression in einzelnen Azinus-Zellen des exokrinen Pankreas (Pfeile) mit fehlender Expression in den benachbarten Zellen („patchy pattern“), und negativer Reaktion der Endothelzellen, Tier TG15 (hCD59-transgenes Tier), Balken = 100 µm

56
6 Eigene Ergebnisse: hCD59 – mRNA – Expression 6.1 Hintergrund und Bedeutung Die immunhistochemische Untersuchung hatte ergeben, dass die hCD59-transgenen
Tiere ein sehr inhomogenes Proteinexpressionsmuster zeigten. Alle Tiere
exprimierten das hCD59-Protein nur in einem Teil ihrer Organe und Gewebe, wobei
die exprimierenden Organe und Gewebe zwischen den einzelnen Tieren stark
variierten. Zudem zeigten die Kardiomyozyten, die Skelettmuskelzellen und die
Zellen der Tunica muscularis des Ösophagus mit dem „patchy pattern“ ein sehr
ungewöhnliches Expressionsmuster.
Um zu klären, ob das uneinheitliche Expressionsmuster nur auf der Proteinebene
vorhanden war, oder ob es auf einer ebenso variierenden mRNA-Expression beruht,
wurde eine qualitative mRNA-Expressionsanalyse mittels Reverser Transkriptase -
Polymerase Kettenreaktion (RT-PCR) durchgeführt.
6.2 Experimentelles Design 6.2.1 Material Als Ausgangsmaterial für die hCD59-mRNA-Expressionsanalyse diente formalin-
fixiertes, paraffin-eingebettetes Material von Herz, Lunge, Milz, Leber, Niere,
Pankreas, Gehirn, Skelettmuskulatur und Ösophagus. Zum Zweck der hCD59-
mRNA-Expressionsanalyse wurde eine qualitative Reverse-Transkriptase-PCR
durchgeführt. Alle 18 hCD59-transgenen Tiere wurden untersucht. Dafür wurden aus
dem formalin-fixiertem, paraffin-eingebetteten Material eine RNA-Isolierung (BOCK
et al., 2001) und eine anschließende cDNA-Synthese durchgeführt.
Als Positivkontrollen dienten humanes Nierengewebe sowie Zellen, die mit dem in
die transgenen Schweine mikroinjizierten Konstrukt transfiziert waren. Als
Negativkontrollen dienten Proben, in die bei der cDNA-Synthese keine Reverse

6 Eigene Ergenisse: hCD59 – mRNA - Expression 57
Trankriptase pipettiert wurde (-RT), Proben von einem hCD55-transgenen Schwein
(TG23), Proben von einem nicht-transgenen Kontrolltier (KO5) sowie Proben, in die
keine cDNA gegeben wurde.
6.2.2 Durchführung der RNA-Isolierung Von den Paraffinblöcken wurden 8-10 15 µm dicke Paraffinschnitte angefertigt und
ohne Entparaffinierung oder Dehydrierung in ein 1.5 ml Eppendorf-Mikro-
Reaktionsgefäß überführt. Dazu wurden 750 µl Digestion solution (Ansatz siehe
Anhang), 5.25 µl beta-Mercaptoethanol (Fa. Merck, Darmstadt) und 300 µl
Proteinase K (20 mg/ml, Merck, Darmstadt) gegeben und im Thermoshaker
(Thermomixer compact, Fa. Eppendorf, Wesseling-Berzdorf) bei 55°C und 1400 rpm
verdaut.
Am nächsten Tag folgte eine Zentrifugation bei 4°C und 13000 g für 5 Minuten,
wodurch die Probe in eine Paraffinkappe, einen klaren Überstand und unverdautes
Material am Boden des Mikro-Reaktionsgefäßs getrennt wurde. 1000 µl Überstand
wurden in ein neues, 2 ml Eppendorf-Mikro-Reaktionsgefäß überführt und 100 µl 3M
Natrium-Acetat mit pH 5.2, 630 µl Roti-Aqua-Phenol (Fa. Roth, Karlsruhe) und 270 µl
Chloroform (Fa. J.T. Baker, Deventer) dazupipettiert. Daraufhin erfolgte eine
15minütige Phasentrennung auf Eis und eine anschließende Zentrifugation bei 4°C
und 13000 g für 20 Minuten. Nach der organischen RNA-Extraktion wurde der
wässrige Überstand in ein neues Eppendorf-Mikro-Reaktionsgefäß überführt und mit
1 Volumen Isopropanol und 1 µl Glykogen als RNA-Carrier versetzt. Die
anschließende Fällung erfolgte über Nacht bei -20°C.
Am darauf folgenden Tag erfolgte eine Zentrifugation bei 4°C und 13000 g für 20-30
Minuten. Der Überstand wurde verworfen und das RNA-Pellet mit eisgekühltem,
70%igem Ethanol gewaschen und anschließend unter dem Abzug getrocknet.
Anschließend wurde das RNA-Pellet in 40 µl H2O/DEPC gelöst, mit 4 µl (1/10
Volumen) 3M Natrium-Acetat mit pH 5.2 (Ansatz siehe Anhang) und 110 µl (2,5
Volumen) 100%igem Ethanol versetzt und über Nacht präzipitiert.
Am nächsten Tag erfolgte eine erneute Zentrifugation bei 4°C und 13000 g für 20-30
Minuten. Der Überstand wurde verworfen und das RNA-Pellet mit eisgekühltem,

6 Eigene Ergenisse: hCD59 – mRNA - Expression 58
70%igem Ethanol gewaschen und anschließend unter dem Abzug getrocknet.
Anschließend wurde das RNA-Pellet in 40 µl H2O/DEPC gelöst und 2 µl von dem
Ansatz in 98 µl in H2O/DEPC zur Messung der optischen Dichte verdünnt.
Anschließend erfolgte die Messung der optischen Dichte mit einem UV-DU®640
Spectrophometer (Fa. Beckmann) bei 260 und 280 nm.
Die nicht verwendete RNA wurde nochmals präzipitiert und bei -20°C gelagert.
6.2.3 Durchführung der cDNA-Synthese Die Konzentration an RNA für die cDNA-Synthese betrug 1 µg RNA / µl H2O/DEPC.
Für den DNAse-Verdau wurden 1 µl RQ1 10x DNAse-Puffer (400mM Tris/HCl,
100mM MgSO4, 10mM CaCl2, Fa. Promega, Mannheim) und 1 µl RQ1 DNAse (pro U
RQ1) (Fa. Promega, Mannheim) hinzugefügt und bei 37°C für 30 Minuten im
Biometra Personal Cycler (Version 3.26, Fa. Biometra, Göttingen) inkubiert.
Anschließend erfolgte die Zugabe von 1 µl RQ1 stop solution (20mM EGTA, Fa.
Promega, Mannheim) und 1 µl pd(N)6-Random Hexamer (Fa. Amersham, Freiburg)
und eine Inkubation bei 70°C für 10 Minuten im Biometra Personal Cycler. Danach
wurden die Proben für einige Minuten auf Eis gekühlt und je Mikro-Reaktionsgefäß 4
µl 5x1st strand buffer (250mM Tris/HCl, 375mM KCl, 15mM MgCl2, Fa. Invitrogen,
Karlsruhe), 2 µl 0.1M DTT (Dithiothreitol, Fa. Invitrogen, Karlsruhe), 1 µl 10mM dNTP
Mix (je 10 µmol dATP, dCTP, dGTP, dTTP pro 1.0 ml, Fa. Fermentas, St. Leon-Rot)
und 0.5 µl Ribonuclease Inhibitor (40u/µl, Fa. Fermentas, St. Leon-Rot)
hinzugegeben. Es folgte eine Inkubation im Biometra Personal Cycler bei 25°C für 10
Minuten. Anschließend wurde in jedes Mikro-Reaktionsgefäß 1 µl (= 200 U)
Superscript II RNase H- Reverse Transkriptase (Fa. Invitrogen, Karlsruhe) pipettiert
und im Biometra Personal Cycler bei 42°C für 50 Minuten und dann bei 70°C für 15
Minuten zur Inaktivierung des Enzyms belassen.
Anschließend wurde die cDNA bei -20°C gelagert.

6 Eigene Ergenisse: hCD59 – mRNA - Expression 59
6.2.4 Auswahl der Primer Die Primerpaare für die hCD59-mRNA-Expressionsanalyse mittels RT-PCR wurden
basierend auf dem mikroinjizierten Konstrukt gewählt (siehe Anhang). Dabei
basierten die Sequenzen der Primerpaare auf der Region des Konstruktes, welches
für das hCD59 kodierte (Fa. Sigma-ARK, Steinheim):
Forward Primer hCD59: 5’ GGTCATAGCCTGCAGTGCTACA 3’ (bp 541-562 im
Konstrukt)
Reverse Primer hCD59: 5’ AAATCAGATGAACAATTGACGGC 3’ (bp 595-617 im
Konstrukt)
Das Produkt wies eine errechnete Größe von 77 Basenpaaren auf.
Als Housekeeping-Gen wurde EF1α gewählt (GRUBER und LEVINE, 1997), die
Primerpaare hatten folgende Sequenzen (Fa. Sigma-Genosys, Steinheim):
Forward Primer EF1 α: 5’ CAAAAAYGACCCACCAATGG 3’
Reverse Primer EF1 α: 5’ GGCCTGGATGGTTCAGGATA 3’
Das Produkt wies eine errechnete Größe von 68 Basenpaaren auf (GenBank
Accession number XM203909).
Die Primer wurden mit Hilfe der Primer Express software Version 1.0 (Applied
Biosystems, Foster City, USA) ausgewählt.
6.2.5 Durchführung der Reversen Transkriptase – PCR Für die RT-PCR wurde ein Mastermix mit 25 µl Reaktionsvolumen angesetzt. Der
Einsatz an cDNA betrug 3.0 µg für die hCD59-Analyse und 2.5 µg für die EF1α-
Analyse. Der Mastermix wurde folgendermaßen angesetzt:
6 Eigene Ergenisse: hCD59 – mRNA - Expression 60
Reagenz µl pro Mikro-Reaktionsgefäß
hCD59
µl pro Mikro-Reaktionsgefäß
EF1α
H2O-Ampuwa (dest. Wasser, Fa.
Fresenius Kabi, Bad Homburg)
16.4 µl 15.9 µl
10 x Puffer
(Invitrogen, Karlsruhe)
2.5 µl 2.5 µl
25 mmol MgCl2
(Fa. Invitrogen, Karlsruhe)
1.5 µl 2.5 µl
10 mM dNTP Mix
(Fa. Fermentas, St. Leon-Rot)
0.5 µl 0.5 µl
Primer F + R (10 pmol/ µl) 1.0 µl 1.0 µl
Platinum Taq DNA Polymerase 5U/ µl
(Fa. Invitrogen, Karlsruhe)
0.1 µl 0.1 µl
cDNA 3.0 µl 2.5 µl
Die Durchführung der RT-PCR erfolgte im Gene Amp Cycler PCR System 9700 (Fa.
PE Applied Biosystems, Foster City, USA) mit folgendem Programm:
94°C 5 Minuten (Aktivierung der Taq-Polymerase und initiale Denaturierung)
94°C 30 Sekunden (Denaturierung der DNA)
60°C 30 Sekunden (Annealing der Primer)
72°C 30 Sekunden (Elongation des Amplifikates)
72°C 5 Minuten (Finale Elongation)
4°C halten
Dabei wurde Schritt 2-4 in 40 Zyklen wiederholt.
Nach der Durchführung der RT-PCR im Cycler wurden die Proben mit Ladepuffer (6x
Loading Dye, 0.09% Bromphenolblau, 0.09% Xylen Cyanol FF, 60% Glycerol, 60mM
EDTA Fa. Fermentas, St. Leon-Rot) gemischt und auf ein 6%iges PAA-Gel
(Polyacrylamid-Gel, Rotiphorese-Gel, Fa. Roth, Karlsruhe, Ansatz siehe Anhang)

6 Eigene Ergenisse: hCD59 – mRNA - Expression 61
aufgetragen. Als Marker wurde pBR 322 DNA/BsuRI (HaeIII) Marker (Fa. Fermentas,
St. Leon-Rot) verwendet. An das Gel wurde eine Spannung von 160 V angelegt
(GIBCO BRL Electrophoresis Power Supply ST 606, Fa. GIBCO/Invitrogen,
Karlsruhe). Als Puffer wurde 1xTE-Puffer (Ansatz siehe Anhang) verwendet. Das Gel
wurde anschließend für 5 Minuten in Ethidiumbromid (1 ml 1%ige Stammlösung (10
mg/ml, Fa. Roth, Karlsruhe) in 1 Liter H2O) inkubiert, um die Produkte anschließend
auf einem UV-Tisch (Flourescent Table, Fa. Renner GmbH, Dannstadt)
dokumentieren zu können (CCD Module).
6.2.6 Auswertung Die untersuchten Organe und Gewebe wurden anhand der Banden (= PCR-
Produkte) hinsichtlich des qualitativen Expressionsmusters (Nachweis der hCD59-
mRNA in den untersuchten verschiedenen Organen und Geweben) sowie
hinsichtlich der Bandenstärke in den verschiedenen untersuchten Proben beurteilt.
Dabei wurden Organe und Gewebe ohne sichtbare Bande mit – beurteilt, schwache
Banden mit +, mittelstarke Banden mit ++ und starke Banden mit +++. Alle
Untersuchungen wurden zweimal durchgeführt, um eine Reproduzierbarkeit der
Ergebnisse zu gewährleisten.

6 Eigene Ergenisse: hCD59 – mRNA - Expression 62
6.3 Ergebnisse und Interpretation
6.3.1 EF1α – mRNA - Expression Alle untersuchten hCD59-transgenen Tiere, das hCD55-transgene Tier (TG23) und
das nicht-transgene Kontrolltier (KO5) zeigten eine deutliche Expression von EF1α in
allen untersuchten Organen (siehe Abb. 16). Dieses Ergebnis spricht dafür, dass
mRNA mit den eingesetzten Methoden in allen Organen und Geweben nachweisbar
war.
6.3.2 hCD59 - mRNA – Expression Von den 18 untersuchten hCD59-transgenen Tieren wiesen 3 Tiere (TG18, TG19,
TG25) eine Expression von hCD59-mRNA in allen untersuchten Organen auf.
12 Tiere (TG1, TG2, TG7, TG8, TG12, TG13, TG14, TG15, TG20, TG21, TG26,
TG27) zeigten, wie in der Proteinanalyse, eine hCD59–mRNA–Expression nur in
einem Teil der untersuchten Organe und Gewebe (siehe Abb. 16).
3 Tiere (TG4, TG5, TG6) wiesen in der RT-PCR keine hCD59-mRNA-Expression in
allen untersuchten Organen und Geweben auf.
Die genaue Verteilung der hCD59-mRNA-Expression in den verschiedenen Organen
und Geweben sowie die Bandenstärken sind in Tabelle 6 dargestellt.
Die humanen Nierenproben und die Proben aus den mit dem Konstrukt transfizierten
Zellinien zeigten eine deutliche hCD59-mRNA-Expression.
Das nicht-transgene Kontrolltier, das hCD55-transgene Tier, die Proben ohne
Reverse Transkriptase und die Proben ohne Template waren in allen untersuchten
Organen und Geweben negativ für hCD59-mRNA.
Dies spricht für die Spezifität der verwendeten Primerpaare (siehe Abb. 15).
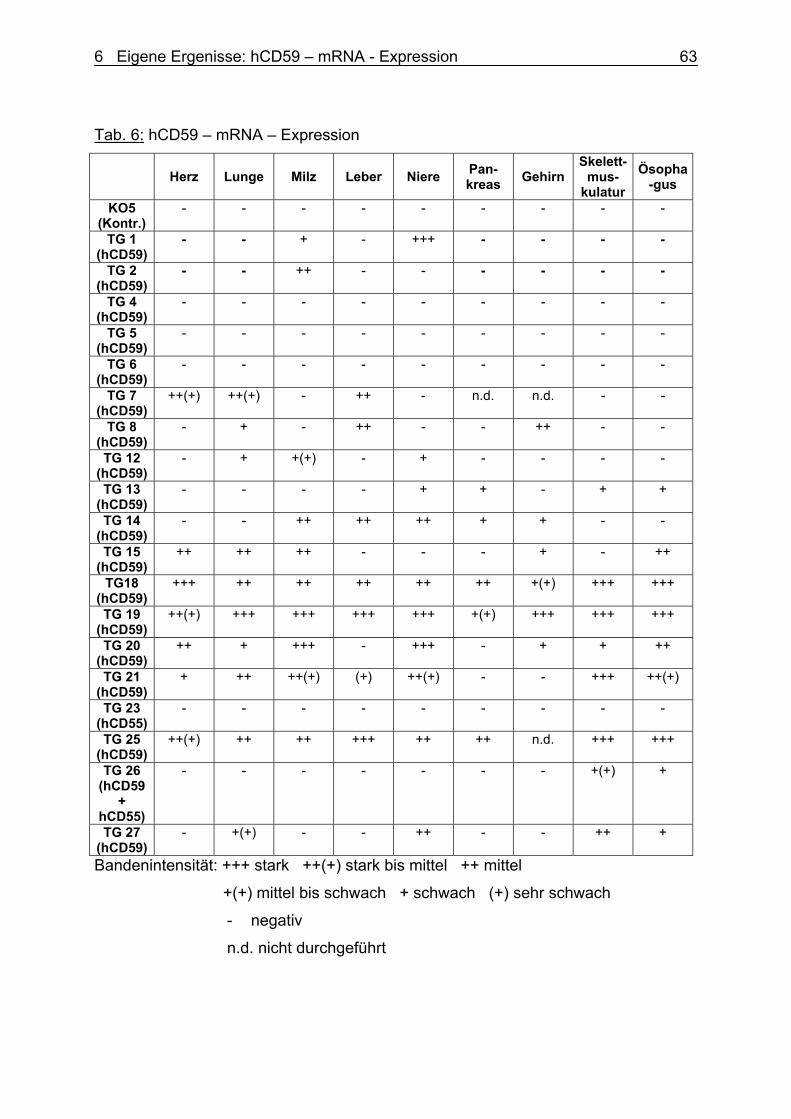
6 Eigene Ergenisse: hCD59 – mRNA - Expression 63
Tab. 6: hCD59 – mRNA – Expression
Herz Lunge Milz Leber Niere Pan-kreas Gehirn
Skelett-mus-
kulatur Ösopha
-gus
KO5 (Kontr.)
- - - - - - - - -
TG 1 (hCD59)
- - + - +++ - - - -
TG 2 (hCD59)
- - ++ - - - - - -
TG 4 (hCD59)
- - - - - - - - -
TG 5 (hCD59)
- - - - - - - - -
TG 6 (hCD59)
- - - - - - - - -
TG 7 (hCD59)
++(+) ++(+) - ++ - n.d. n.d. - -
TG 8 (hCD59)
- + - ++ - - ++ - -
TG 12 (hCD59)
- + +(+) - + - - - -
TG 13 (hCD59)
- - - - + + - + +
TG 14 (hCD59)
- - ++ ++ ++ + + - -
TG 15 (hCD59)
++ ++ ++ - - - + - ++
TG18 (hCD59)
+++ ++ ++ ++ ++ ++ +(+) +++ +++
TG 19 (hCD59)
++(+) +++ +++ +++ +++ +(+) +++ +++ +++
TG 20 (hCD59)
++ + +++ - +++ - + + ++
TG 21 (hCD59)
+ ++ ++(+) (+) ++(+) - - +++ ++(+)
TG 23 (hCD55)
- - - - - - - - -
TG 25 (hCD59)
++(+) ++ ++ +++ ++ ++ n.d. +++ +++
TG 26 (hCD59
+ hCD55)
- - - - - - - +(+) +
TG 27 (hCD59)
- +(+) - - ++ - - ++ +
Bandenintensität: +++ stark ++(+) stark bis mittel ++ mittel
+(+) mittel bis schwach + schwach (+) sehr schwach
- negativ
n.d. nicht durchgeführt
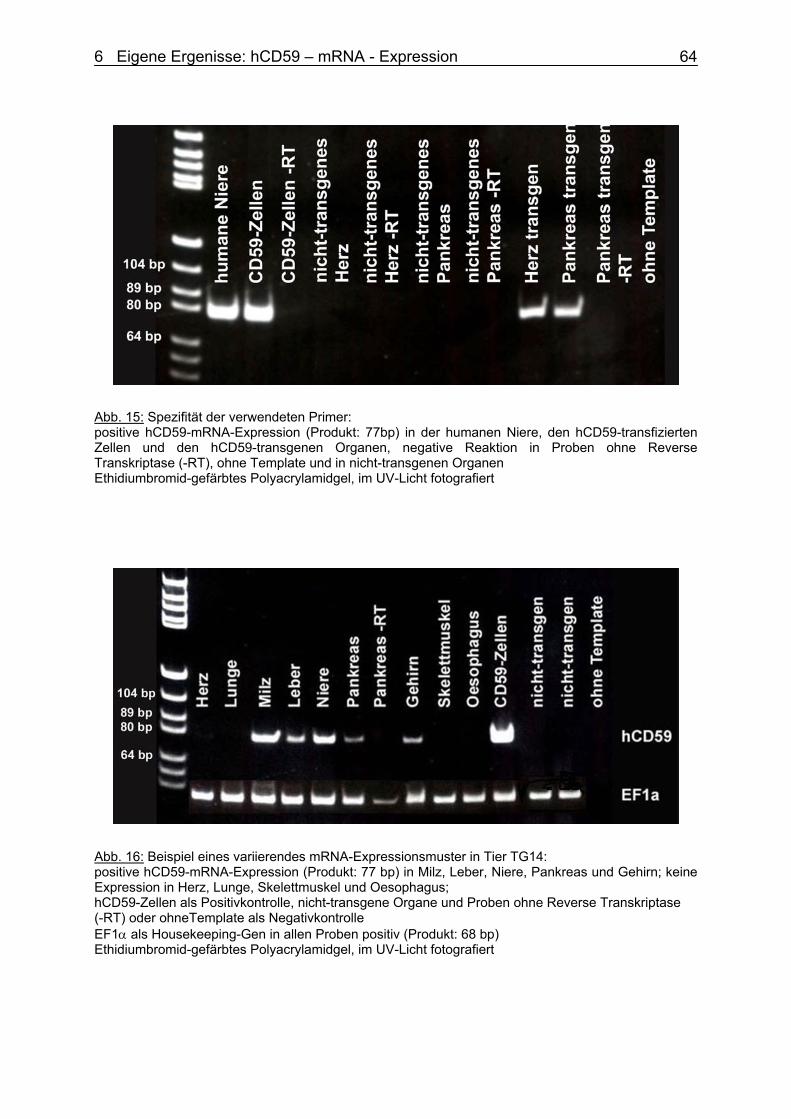
6 Eigene Ergenisse: hCD59 – mRNA - Expression 64
Abb. 15: Spezifität der verwendeten Primer: positive hCD59-mRNA-Expression (Produkt: 77bp) in der humanen Niere, den hCD59-transfizierten Zellen und den hCD59-transgenen Organen, negative Reaktion in Proben ohne Reverse Transkriptase (-RT), ohne Template und in nicht-transgenen Organen Ethidiumbromid-gefärbtes Polyacrylamidgel, im UV-Licht fotografiert
Abb. 16: Beispiel eines variierendes mRNA-Expressionsmuster in Tier TG14: positive hCD59-mRNA-Expression (Produkt: 77 bp) in Milz, Leber, Niere, Pankreas und Gehirn; keine Expression in Herz, Lunge, Skelettmuskel und Oesophagus; hCD59-Zellen als Positivkontrolle, nicht-transgene Organe und Proben ohne Reverse Transkriptase (-RT) oder ohneTemplate als Negativkontrolle EF1α als Housekeeping-Gen in allen Proben positiv (Produkt: 68 bp) Ethidiumbromid-gefärbtes Polyacrylamidgel, im UV-Licht fotografiert

6 Eigene Ergenisse: hCD59 – mRNA - Expression 65
6.3.3 Vergleich hCD59 – Protein - und mRNA - Expression Vergleicht man die Expression von dem hCD59-Protein mit der Expression der
hCD59-mRNA (Tabelle 7), so wird deutlich, dass es folgende Gruppen gibt:
1) Tiere, bei denen sowohl hCD59-Protein als auch hCD59-mRNA nachweisbar
waren
(TG1, TG7, TG8, TG15, TG18, TG19, TG20, TG21, TG25, TG26)
2) Tiere, bei denen hCD59-mRNA, aber kein hCD59-Protein nachweisbar war
(TG2, TG12, TG13, TG14, TG27)
3) Tiere, bei denen hCD59-Protein, aber keine hCD59-mRNA nachweisbar war
(TG4, TG6)
4) Tiere, bei denen weder hCD59-Protein noch hCD59-mRNA nachweisbar war
(TG5)
Tab. 7: Vergleich Expression von hCD59-Protein und hCD59-mRNA
Tier-Nummer hCD59 - Protein hCD59 - mRNA KO5 (Kontrolle) - - TG1 + Teil der Organe + Teil der Organe TG2 - + Teil der Organe TG4 + Teil der Organe - TG5 - - TG6 + Teil der Organe - TG7 + Teil der Organe + Teil der Organe TG8 + Teil der Organe + Teil der Organe TG12 - + Teil der Organe TG13 - + Teil der Organe TG14 - + Teil der Organe TG15 + Teil der Organe + Teil der Organe TG18 + Teil der Organe + alle Organe TG19 + Teil der Organe + alle Organe TG20 + Teil der Organe + Teil der Organe TG21 + Teil der Organe + Teil der Organe TG23 (hCD55) - - TG25 + Teil der Organe + alle Organe TG26 + Teil der Organe + Teil der Organe TG27 - + Teil der Organe

66
7 Eigene Ergebnisse: hCD59 – DNA – Detektion 7.1 Hintergrund und Bedeutung Die Untersuchung der hCD59-Protein- und hCD59-mRNA-Expression hatte gezeigt,
dass auch in der hCD59-mRNA-Expression ein sehr variables Muster vorherrschte.
Der nächste Schritt lag darin, die hCD59-DNA nachzuweisen, um festzustellen, ob
auch die Konstrukt-DNA nur in einem Teil der Organe nachweisbar war, was auf das
Vorliegen eines Chimärismus in den transgenen Tieren hindeuten könnte. Weiterhin
gab es Tiere, die die hCD59-mRNA, aber nicht das hCD59-Protein exprimierten, was
auf eine Störung, Hemmung oder Blockade der Translation hinweisen könnte. Es war
also von Interesse, ob es auch Tiere gab, in denen hCD59-DNA nachweisbar war,
obwohl die hCD59-mRNA in diesen Organen nicht nachgewiesen werden konnte.
Diese Fragestellung war besonders im Hinblick auf eine mögliche
Trankskriptionsstörung oder –hemmung, oder eine fehlende Transkription
interessant. Und zuletzt existierten auch Tiere, die das hCD59-Protein exprimierten,
obwohl die hCD59-mRNA nicht nachgewiesen werden konnte. Es war zu bedenken,
ob hier vielleicht die mRNA nur eine kürzere Halbwertszeit hatte, oder ob die
Immunhistochemie möglicherweise unspezifisch reagiert hatte.
Aus diesen Gründen wurde an 6 ausgewählten Tieren aus den 3 genannten Gruppen
ein Nachweis der hCD59-DNA durchgeführt, um zu klären, ob die Konstrukt-DNA in
allen Organen und Geweben integriert war, oder ob auch hier eine Variation bestand.
7.2 Experimentelles Design 7.2.1 Material Zum Zweck der hCD59-DNA-Detektion wurde eine qualitative PCR durchgeführt. Als
Ausgangsmaterial diente formalin-fixiertes, Paraffin-eingebettetes Material von Herz,
Lunge, Milz, Leber, Niere, Pankreas, Gehirn, Skelettmuskulatur und Ösophagus. Für

7 Eigene Ergebnisse: hCD59 – DNA - Detektion 67
die hCD59-DNA-Detektion wurden 6 hCD59-transgene Schweine (TG4, TG6, TG8,
TG14, TG18, TG19) exemplarisch untersucht. Dabei handelte es sich bei den Tieren
TG4 und TG6 um Tiere, die das hCD59-Protein exprimierten, aber keine Expression
der hCD59-mRNA gezeigt hatten. TG8 und TG14 waren Tiere, die in der Protein-
und in der mRNA-Analyse eine Expression von hCD59 in einem Teil der Organe
gezeigt hatten. TG18 und TG19 hatten in der Proteinanalyse nur in einem Teil der
Organe positiv reagiert, in der RT-PCR jedoch eine Expression von hCD59-mRNA in
allen Organen gezeigt.
Zunächst wurde DNA isoliert und anschließend eine qualitative PCR durchgeführt.
Als Positivkontrollen dienten Zellen, die mit dem in die transgenen Schweine
mikroinjizierten Konstrukt transfiziert waren. Als Negativkontrollen dienten Proben
von einem nicht-transgenen Kontrolltier (KO5) sowie Proben, in die kein Template
(DNA) gegeben wurde.
7.2.2 Durchführung der DNA – Isolierung Von den Paraffinblöcken wurden 8-10 15 µm dicke Paraffinschnitte angefertigt und
ohne Entparaffinierung oder Rehydrierung in ein 1.5 ml Eppendorf-Mikro-
Reaktionsgefäß verbracht. Dazu wurden 1000 µl Proteinase K-Puffer (Ansatz siehe
Anhang) und 50 µl Proteinase K (20 mg/ml, Merck, Darmstadt) pipettiert und im
Thermoshaker (Thermomixer compact, Fa. Eppendorf, Wesseling-Berzdorf) bei 55°C
und 1400 rpm verdaut.
Am nächsten Tag folgte eine Zentrifugation bei 4°C und 13000 g für 5 Minuten. 1000
µl Überstand wurden in ein neues 2 ml Eppendorf-Mikro-Reaktionsgefäß überführt
und 750 µl Phenol-Chloroform (Roti-Phenol/C/I, Fa. Roth, Karlsruhe) dazugegeben.
Daraufhin erfolgte eine Zentrifugation bei Raumtemperatur und 13000 g für 10
Minuten. Der wässrige Überstand wurde in ein neues Eppendorf-Mikro-
Reaktionsgefäß überführt und erneut mit 750 µl Phenol-Chloroform (Roti-Phenol/C/I,
Fa. Roth, Karlsruhe) versetzt und wieder bei Raumtemperatur und 13000 g für 10
Minuten zentrifugiert. Diese Phenol-Chloroform-Extraktion wurde wiederholt, bis
keine weiße Interphase mehr zu sehen war. Dann wurden zu dem klaren Überstand
5 µl RNAse A (10 mg/ml) gegeben und bei 37°C und 800 rpm im Thermoshaker für

7 Eigene Ergebnisse: hCD59 – DNA - Detektion 68
45 Minuten inkubiert. Anschließend wurde eine erneute Phenol-Chloroform-
Extraktion durchgeführt. Danach wurde der wässrige Überstand mit 750 µl
Chloroform (Fa. J.T. Baker, Deventer) versetzt und bei Raumtemperatur und 13000 g
für 3 Minuten zentrifugiert. Zu dem Überstand wurden anschließend 1/10 Volumen
3M Natrium-Acetat mit pH 7.0 / 100 µg/ml Dextran (Ansatz siehe Anhang) und 2.5
Volumen 100%iges Ethanol pipettiert und über Nacht bei -20°C eine Fällung
durchgeführt.
Am nächsten Tag erfolgte eine erneute Zentrifugation bei 4°C und 13000 g für 20-30
Minuten. Der Überstand wurde abgenommen und das DNA-Pellet mit 70%igem
Ethanol gewaschen und anschließend unter dem Abzug getrocknet. Anschließend
wurde das DNA-Pellet in 100 µl 1x TE-Puffer (Ansatz siehe Anhang) gelöst und 2 µl
von dem Ansatz in 98 µl in H2O/DEPC zur Messung der optischen Dichte verdünnt.
Anschließend erfolgte die Messung der optischen Dichte mit einem UV-DU®640
Spectrophometer (Fa. Beckmann) bei 260 und 280 nm. Die DNA wurde mit 1x TE-
Puffer verdünnt, bis sie eine Konzentration von 50 ng/µl aufwies.
Die nicht verwendete DNA wurde anschließend bei -20°C gelagert.
7.2.3 Auswahl der Primer Für die hCD59-DNA-Detektionsanalyse wurden die gleichen Primer wie für die
hCD59-mRNA-Expressionsanalyse benutzt, da das Konstrukt im Gegensatz zum
humanen CD59-Gen keine Introns aufweist. (Fa. Sigma-ARK, Steinheim):
Forward Primer hCD59: 5’ GGTCATAGCCTGCAGTGCTACA 3’ (bp 541-562 im
Konstrukt)
Reverse Primer hCD59: 5’ AAATCAGATGAACAATTGACGGC 3’ (bp 595-617 im
Konstrukt)
Das Produkt wies eine errechnete Größe von 77 Basenpaaren auf.
Als Housekeeping-Gen wurde TNF gewählt (GRIERSON et al., 2004), die
Primerpaare hatten folgende Sequenzen (Fa. Sigma-Genosys, Steinheim):
Forward Primer TNF: 5’ ATAGTATGAGGAGGACTCGCTGAGC 3’
Reverse Primer TNF: 5’ ACATCCCATCTGTCCATGAGGTT 3’

7 Eigene Ergebnisse: hCD59 – DNA - Detektion 69
Das Produkt wies eine errechnete Größe von 94 Basenpaaren auf (GenBank
Accession number X54001).
Die Primer wurden mit Hilfe der Primer Express software Version 1.0 (Applied
Biosystems, Foster City, USA) ausgewählt.
7.2.4 Durchführung der PCR Für die PCR wurde ein Mastermix mit 25 µl Reaktionsvolumen angesetzt. Der
Einsatz an DNA betrug 250 ng für die hCD59-Analyse und 250 ng für die TNF-
Analyse. Der Mastermix wurde folgendermaßen angesetzt:
Reagenz µl pro Mikro-Reaktionsgefäß
hCD59
µl pro Mikro-Reaktionsgefäß
TNF
H2O-Ampuwa (dest. Wasser, Fa.
Fresenius Kabi, Bad Homburg)
15.5 µl 14.5 µl
10 x Puffer
(Fa. Invitrogen, Karlsruhe)
2.5 µl 2.5 µl
25 mmol MgCl2
(Fa. Invitrogen, Karlsruhe)
1.5 µl 2.5 µl
10 mM dNTP Mix
(Fa. Fermentas, St. Leon-Rot)
0.5 µl 0.5 µl
Primer (10 pmol/ µl) 1.0 µl 1.0 µl
Platinum Taq DNA Polymerase 5 U/ µl
(Fa. Invitrogen, Karlsruhe)
0.05 µl 0.05 µl
DNA (50 ng/µl) 5.0 µl 5.0 µl

7 Eigene Ergebnisse: hCD59 – DNA - Detektion 70
Die Durchführung der PCR erfolgte im Gene Amp Cycler PCR System 9700 (Fa. PE
Applied Biosystems, Foster City, USA) mit folgendem Programm:
94°C 5 Minuten (Aktivierung der Taq-Polymerase und initiale Denaturierung)
94°C 30 Sekunden (Denaturierung der DNA)
60°C 30 Sekunden (Annealing der Primer)
72°C 30 Sekunden (Elongation des Amplifikates)
72°C 5 Minuten (Finale Elongation)
4°C halten
Dabei wurden Schritt 2-4 in 40 Zyklen wiederholt.
Nach der Durchführung der PCR im Cycler wurden die Proben mit Ladepuffer (6x
Loading Dye, 0.09% Bromphenolblau, 0.09% Xylen Cyanol FF, 60% Glycerol, 60mM
EDTA, Fa. Fermentas, St. Leon-Rot) gemischt und auf ein 6%iges PAA-Gel
(Polyacrylamid-Gel, Rotiphorese-Gel, Fa. Roth, Karlsruhe, Ansatz siehe Anhang)
aufgetragen. Als Marker wurde pBR 322 DNA/BsuRI (HaeIII) Marker (Fa. Fermentas,
St. Leon-Rot) verwendet. An das Gel wurde eine Spannung von 160 V angelegt
(GIBCO BRL Electrophoresis Power Supply ST 606, Fa. GIBCO/Invitrogen,
Karlsruhe). Als Puffer wurde 1xTE-Puffer verwendet (Ansatz siehe Anhang). Das Gel
wurde anschließend für 5 Minuten in Ethidiumbromid (1 ml 1%ige Stammlösung (10
mg/ml, Fa. Roth, Karlsruhe) in 1 Liter H2O) verbracht, um die PCR-Produkte
anschließend auf einem UV-Tisch (Flourescent Table, Fa. Renner GmbH,
Dannstadt) dokumentieren zu können (CCD Module).
7.2.5 Auswertung Die untersuchten Organe und Gewebe wurden anhand der Banden (= PCR-
Produkte) hinsichtlich der qualitativen Detektion (Nachweis der hCD59-DNA in den
untersuchten verschiedenen Organen und Geweben) beurteilt. Dabei wurden
negative Reaktionen (keine Bande) mit – und positive Reaktionen mit + beurteilt. Alle
Untersuchungen wurden zweimal durchgeführt, um eine Reproduzierbarkeit der
Ergebnisse zu gewährleisten.

7 Eigene Ergebnisse: hCD59 – DNA - Detektion 71
7.3 Ergebnisse und Interpretation
7.3.1 TNF – Expression In allen untersuchten Tieren konnte in allen untersuchten Organen und Geweben
TNF (siehe Abb. 17) nachgewiesen werden. Dies zeigt, dass die isolierte DNA
vorhanden war.
7.3.2 hCD59 – DNA – Expression Die qualitative PCR zeigte in allen untersuchten Organen und Geweben der 6 Tiere
einen deutlichen Nachweis von hCD59-DNA (siehe Abb. 17).
Das nicht-transgene Kontrolltier zeigte keine hCD59-DNA-Detektion, was für eine
Spezifität der gewählten Primerpaare spricht.
Abb. 17: deutlicher hCD59-DNA-Nachweis (Produkt: 77 bp) in allen untersuchten Organen und Geweben in Tier TG14 (gleiches Tier wie in Abb. 12) TNF als Housekeeping-Gen in allen Proben positiv (Produkt: 94 bp) Ethidiumbromid-gefärbtes Polyacrylamidgel, im UV-Licht fotografiert.
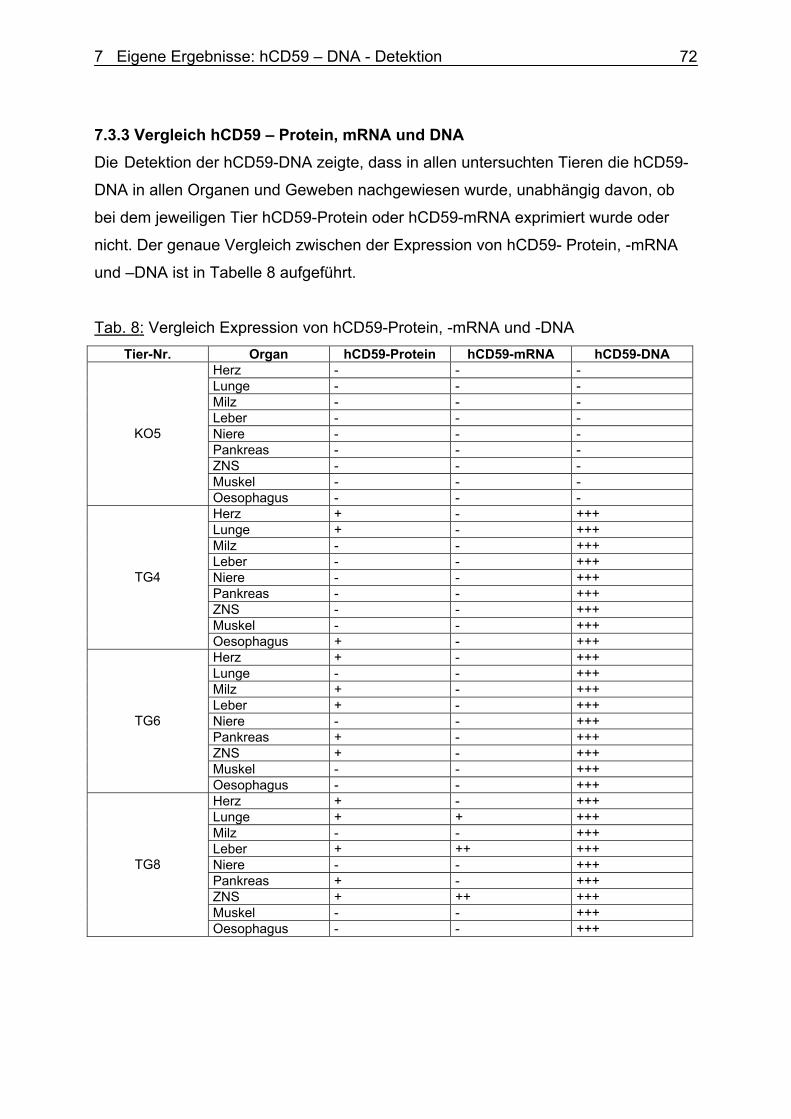
7 Eigene Ergebnisse: hCD59 – DNA - Detektion 72
7.3.3 Vergleich hCD59 – Protein, mRNA und DNA Die Detektion der hCD59-DNA zeigte, dass in allen untersuchten Tieren die hCD59-
DNA in allen Organen und Geweben nachgewiesen wurde, unabhängig davon, ob
bei dem jeweiligen Tier hCD59-Protein oder hCD59-mRNA exprimiert wurde oder
nicht. Der genaue Vergleich zwischen der Expression von hCD59- Protein, -mRNA
und –DNA ist in Tabelle 8 aufgeführt.
Tab. 8: Vergleich Expression von hCD59-Protein, -mRNA und -DNA Tier-Nr. Organ hCD59-Protein hCD59-mRNA hCD59-DNA
Herz - - - Lunge - - - Milz - - - Leber - - - Niere - - - Pankreas - - - ZNS - - - Muskel - - -
KO5
Oesophagus - - - Herz + - +++ Lunge + - +++ Milz - - +++ Leber - - +++ Niere - - +++ Pankreas - - +++ ZNS - - +++ Muskel - - +++
TG4
Oesophagus + - +++ Herz + - +++ Lunge - - +++ Milz + - +++ Leber + - +++ Niere - - +++ Pankreas + - +++ ZNS + - +++ Muskel - - +++
TG6
Oesophagus - - +++ Herz + - +++ Lunge + + +++ Milz - - +++ Leber + ++ +++ Niere - - +++ Pankreas + - +++ ZNS + ++ +++ Muskel - - +++
TG8
Oesophagus - - +++
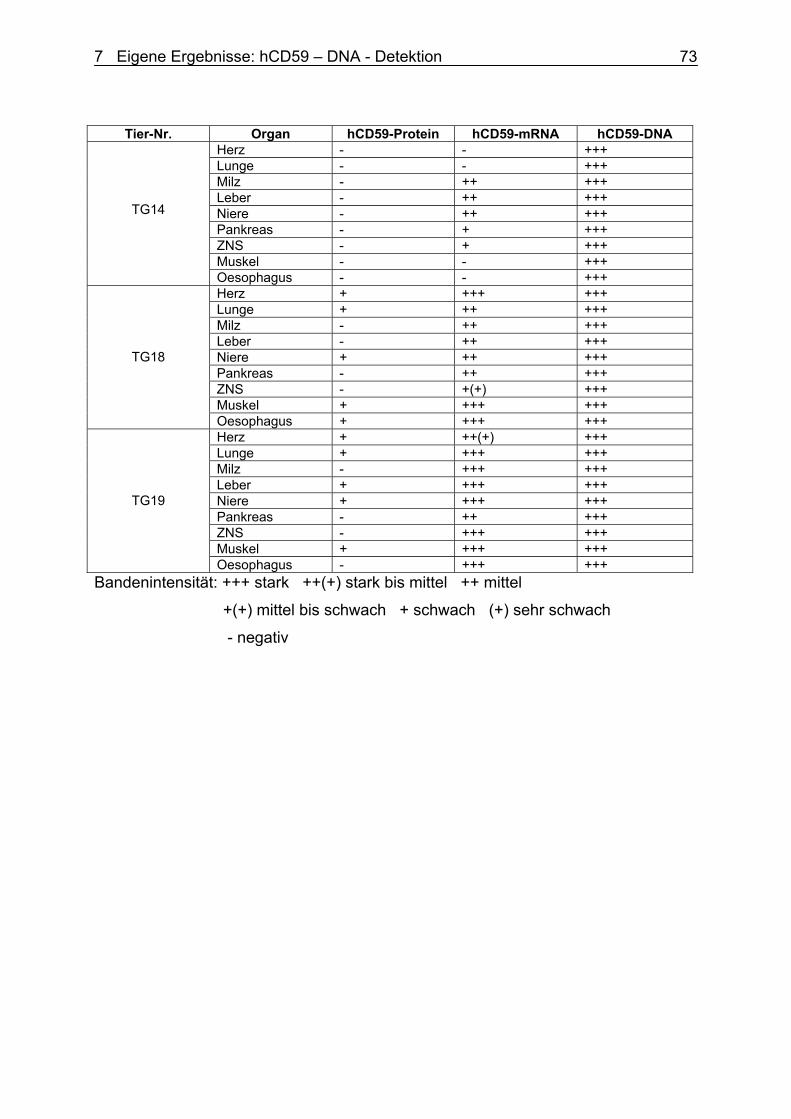
7 Eigene Ergebnisse: hCD59 – DNA - Detektion 73
Tier-Nr. Organ hCD59-Protein hCD59-mRNA hCD59-DNA Herz - - +++ Lunge - - +++ Milz - ++ +++ Leber - ++ +++ Niere - ++ +++ Pankreas - + +++ ZNS - + +++ Muskel - - +++
TG14
Oesophagus - - +++ Herz + +++ +++ Lunge + ++ +++ Milz - ++ +++ Leber - ++ +++ Niere + ++ +++ Pankreas - ++ +++ ZNS - +(+) +++ Muskel + +++ +++
TG18
Oesophagus + +++ +++ Herz + ++(+) +++ Lunge + +++ +++ Milz - +++ +++ Leber + +++ +++ Niere + +++ +++ Pankreas - ++ +++ ZNS - +++ +++ Muskel + +++ +++
TG19
Oesophagus - +++ +++ Bandenintensität: +++ stark ++(+) stark bis mittel ++ mittel
+(+) mittel bis schwach + schwach (+) sehr schwach
- negativ

74
8 Diskussion
Der Mangel an humanen Spenderorganen hat in den letzten Jahren zu einem
verstärkten Interesse an Xenotransplantaten geführt. Um die hyperakute Abstoßung
der Xenotransplantate zu verhindern, wurden transgene Schweine erzeugt, die den
humanen Komplementregulator hCD59 als Transgen tragen.
In der durchgeführten Studie wurden die hCD59-transgenen Schweine dahingehend
untersucht, ob sie einen spezifischen, pathomorphologischen Phänotyp aufweisen.
Zudem wurde eine Expressionsanalyse durchgeführt um zu klären, ob und in
welchen Organen und Geweben die hCD59-DNA, die hCD59-mRNA und das
hCD59-Protein nachweisbar ist. Da es sich bei den untersuchten transgenen
Schweinen um Tiere handelte, die zum Zweck der Xenotransplantation generiert
wurden, ist eine starke und möglichst ubiquitäre Expression des Transgens von
entscheidender Bedeutung um eine hyperakute Abstoßung des Xenotransplantates
zu verhindern.
Da das porzine Circovirus Typ II (PCV II) mittlerweile in großen Teilen der
Schweinepopulation verbreitet ist (KRAKOWKA et al., 2001) und zu
pathomorphologischen Läsionen in betroffenen Tieren führen kann, wurden alle
transgenen Schweine und Kontrolltiere mittels PCR auf PCV II und auf das Porzine
Respiratorische und Reproduktionssyndrom Virus (PRRSV) untersucht. 75% der
Kontrolltiere und 65% der transgenen Tiere waren positiv für PCV II, während in
keinem Tier PRRSV nachgewiesen werden konnte. PCV II wurde das erste Mal 1991
in Kanada nachgewiesen und hat sich in großen Teilen der Schweinepopulation
verbreitet. Während die meisten Tiere das Virus tragen, ohne klinische Symptome
oder pathomorphologische Läsionen zu zeigen, kann PCV II unter bestimmten
Bedingungen zu verschiedenen Krankheitsbildern führen. Vor allem nach einer
Aktivierung des Immunsystems kann PCV II zum so genannten Post Weaning
Multisystemic Wasting Syndrome (PMWS), der wohl bekanntesten PCV II-
assoziierten Erkrankung, führen (KRAKOWKA et al., 2001). Das
pathomorphologische Bild des PMWS umfasst folgende Läsionen: Klinisch fallen die

8 Diskussion 75
betroffenen Tiere durch Kümmern und Ikterus auf. Die Lymphknoten sind
generalisiert vergrößert und zeigen histologisch eine lymphozytäre Depletion mit
Nachweis von Histiozyten und Riesenzellen sowie intrazytoplasmatischen,
basophilen Einschlüssen. Gelegentlich weisen die Lymphknoten auch Nekrosen und
vaskuläre Thromben auf. In der Lunge findet sich eine interstitielle Pneumonie, teils
mit einer lymphatischen Depletion des bronchus-assoziierten lymphatischen
Gewebes (BALT). Die Leber weist lympho-histiozytäre Infiltrationen der Portalfelder,
Einzelzellnekrosen von Hepatozyten, Vakuolisierung von Hepatozyten und
Karyomegalie auf. In einigen Fällen kann es zu schweren Läsionen mit perilobulärer
Fibrose und massivem Verlust von Hepatozyten mit konsekutivem Ikterus kommen.
Zudem können gering- bis mittelgradige, lympho-histiozytäre Infiltrationen in nahezu
allen Organen vorkommen. In den Nieren wird gelegentlich eine interstitielle Nephritis
beobachtet. All diese Läsionen können einzeln oder gemeinsam bei dem PMWS
auftreten. Zu den weiteren Erkrankungen, die PCV II-assoziiert sind, gehören das
porzine Dermatitis und Nephropathie-Syndrom (PDNS), Störungen der Reproduktion,
die proliferative und nekrotisierende Pneumonie (PNP) und der kongenitale Tremor
(SEGALÉS et al., 2004). Die aufgeführten, PCV II-assoziierten Läsionen und
Erkrankungen sind zu berücksichtigen, wenn es um die Bewertung der
pathomorphologischen Untersuchung der transgenen Tiere geht.
Zur weiteren Abklärung eines spezifischen, pathomorphologischen Phänotyps wurde
eine pathologisch-anatomische und histologische Untersuchung der Organe und
Gewebe der transgenen Tiere durchgeführt. Dabei zeigten sich zwei Kategorien von
Veränderungen: Zum einen lagen Läsionen vor, die nur in einzelnen Tieren
diagnostiziert werden konnten, zum anderen gab es Befunde, die bei mehreren
Tieren festgestellt wurden.
Zu den Einzeltierläsionen (Tab. 3) gehören entzündliche Veränderungen, die
höchstwahrscheinlich durch virale oder bakterielle Infektionen verursacht wurden. Im
Vordergrund stehen hier die katarrhalisch-eitrige Bronchopneumonie, die interstitielle
Pneumonie sowie die Pleuritis und Epi- und Perikarditis. Diese entzündlichen
Veränderungen wurden sowohl in einem nicht transgenen Kontrolltier (KO7) als auch
in einem transgenen Tier (TG7) diagnostiziert. Derartige Veränderungen kommen

8 Diskussion 76
regelmäßig in Schweinebeständen vor und sind in der Regel durch Infektionen mit
bakteriellen Erregern bedingt. Bronchopneumonien können beim Schwein durch
verschiedene Bakterien verursacht werden. Pasteurella multiocida, Actinobacillus
pleuropneumoniae, Haemophilus spp., Acarnobacterium pyogenes, Bordetella
bronchiseptica und Salmonella cholerasuis gehören hier zu den am häufigsten
nachzuweisenden Pathogenen (JUBB et al., 1993). Dabei entstehen bakterielle
Infektionen oftmals erst sekundär nach einer primären, viralen Atemwegsinfektion.
Als lungenpathogene Erreger von interstitiellen Pneumonien spielen beim Schwein
vor allem Mycoplasma hyopneumoniae, Influenzaviren, porzines respiratorisches
Coronavirus, PRRSV und PCV II eine Rolle (DAHME, E. u. E. WEISS 1999). Auch
Pleuritis und Epi- und Perikarditis entstehen überwiegend durch bakterielle
Infektionen der Tiere. Hier sind vor allem Haemophilus- und Mycoplasmen-
Infektionen von Bedeutung. Um die genaue Ursache der entzündlichen
Veränderungen in den untersuchten Tieren zu klären, wäre eine mikrobiologische
Untersuchung von nativen Proben der betroffenen Organe nötig gewesen. Da die
Läsionen jedoch chronischer Natur waren und die ursprünglich verantwortlichen
Erreger in solch chronischen Prozessen häufig nicht mehr nachweisbar sind, wurde
auf die Durchführung einer mikrobiologischen Untersuchung verzichtet. Insgesamt
fanden sich jedoch keine Hinweise darauf, dass die transgenen Tiere anfälliger für
Infektionskrankheiten sind, da nur zwei transgene Tiere und auch ein Kontrolltier
fokale, entzündliche Veränderungen aufwiesen. Ein transgenes Tier (TG15) wies
eine fokale Fibrose der Leberkapsel auf. Auch diese Läsion werden in der
Schweinepopulation häufig beobachtet und sind meistens Folge von
Parasitenwanderungen, wie bespielsweise durch Ascaris suum. Derartige
Veränderungen können aber auch auftreten, ohne dass eine klare Ursache gefunden
wird. In den betroffenen Tieren fanden sich keine Parasiten oder andere Hinweise
auf die Ursache der Läsionen, aber auch hier handelte es sich um ein chronisches
Geschehen. Ein Tier (TG4) zeigte eine ulzerative Gastritis. Diese Veränderung stellt
einen regelmäßigen Befund in Schweinebeständen dar und die Ätiologie dieser
Erkrankung ist noch nicht geklärt. Stress und auch die Fütterung der Tiere scheinen
aber in einigen Fällen an der Ätiologie beteiligt zu sein (JUBB et al., 1993). Auch im

8 Diskussion 77
vorliegenden Fall konnte die Ursache der Läsion nicht geklärt werden. Ein
transgenes Schwein (TG23) wies im Mesenterium bandartige Verknöcherungen auf.
Diese mesenterialen Ossifikationen sind ein regelmäßiger Befund im Sektionsgut von
Schweinen. Die Verknöcherung ist wahrscheinlich metaplastischer Natur und die
Ursache für die Metaplasie ist noch nicht geklärt, ein Zusammenhang mit chronisch-
reparativen Prozessen, insbesondere mit Narbenbildungen nach Laparotomien oder
Kastrationen ist jedoch wahrscheinlich (DAHME, E. u. E. WEISS 1999). Das
betroffene Tier war weder kastriert, noch ist bei ihm eine Laparotomie durchgeführt
worden, so dass hier die Ursache der metaplastischen, mesenterialen
Verknöcherungen unklar bleibt. Ein Kontrolltier (KO3) zeigte einen beidseitigen,
abdominalen Kryptorchismus. Dieser Zustand, der durch einen ausbleibenden
Hodenabstieg nach der Geburt entsteht, ist ebenfalls ein regelmäßiger
Spontanbefund in Schweinebeständen. Die Ursache für das Ausbleiben des
Hodenabstiegs ist ebenfalls noch nicht geklärt.
Insgesamt repräsentieren die in der Studie diagnostizierten Einzeltierläsionen das
normale Spektrum der Befunde in Schweinebeständen. Dabei zeigten sowohl nicht
transgene Kontrolltiere als auch transgene Schweine diese Läsionen. Interessant ist
die Frage, warum die Tiere diese Läsionen, besonders die offenbar infektiös
bedingten Veränderungen aufwiesen, obwohl es sich um spezifiziert-pathogen-freie
(SPF) Tiere handelt. Das erklärt sich dadurch, dass SPF-Tiere nicht keimfrei
aufgezogen werden, wie z.B. Gnotobioten, sondern lediglich frei von
anzeigepflichtigen Erkrankungen und spezifischen Keimen sind. Bei den in der
Studie diagnostizierten Läsionen handelt es sich jedoch um Infektionskrankheiten
durch Erreger, die in nahezu allen Schweinepopulationen vorhanden sind und nicht
zu Seuchen oder anzeigepflichtigen Erkrankungen führen. Daher ist es erklärbar,
dass die Tiere, obwohl sie spezifisch-pathogen-frei sind, trotzdem
Infektionskrankheiten aufweisen können. Diese Situation ist dahingehend
problematisch, dass die durchgeführte Studie nicht standardisierbar ist und
undefinierten Bedingungen unterlag. Andererseits spiegelt die Studie dadurch
Feldbedingungen wieder, und zudem liegt durch die Infektionskrankheiten ein
natürlicher Challenge des Phänotyps der Tiere vor. Es ist durchaus denkbar, dass

8 Diskussion 78
ein Phänotyp in einem Tier unter sterilen Bedingungen nicht auftritt, während er bei
einer körperlichen Belastung des Tieres, wie beispielsweise einer
Infektionserkrankung, zum Ausdruck kommt.
Außer den Einzeltierläsionen existierten zudem pathomorphologische Befunde, die in
mehreren Tieren nachgewiesen wurden (Tab. 4).
Dazu zählen unter anderem geringgradige, multifokale, überwiegend lympho-
plasmazelluläre Infiltrationen in der Haut, in der Leber und in der Niere. Derartige
Infiltrationen in diversen Organen werden unter anderem beim Post-weaning-
multisystemic-wasting-syndrome nach einer PCV II-Infektion beobachtet. Die
betroffenen Tiere sind allerdings nur zu einem Teil positiv für PCV II. Auch die PCV
II-negativen Tiere zeigen die herdförmigen Infiltrationen. Derartige geringgradige
Infiltrationen werden allerdings häufig bei histologischen Untersuchungen festgestellt.
Sie haben keine klinische Relevanz und die Ursache ist zumeist unklar. Auch in den
vorliegenden Fällen konnte die Ätiologie der Infiltrate nicht geklärt werden. Weiterhin
zeigten 6 Tiere einen Befall mit Balantidium coli im Colon. Bei Balantidien handelt es
sich um Protozoen, die als Kommensalen im Caecum oder Colon der Tiere leben.
Bei einer Schädigung der Gewebe durch andere Einflüsse können Balantidien
allerdings auch zu einer opportunistischen Invasion führen (JUBB et al., 1993). In
allen betroffenen Tieren der Studie lagen die Balantidien allerdings reaktionslos im
Colonlumen, so dass ihnen höchstwahrscheinlich keine pathogene Bedeutung
zukommt. In der Milz sowie in allen untersuchten Lymphknoten mehrerer Tiere fand
sich eine follikuläre Hyperplasie mit Follikelzentrumsreaktion. Die follikuläre
Hyperplasie der Milz ist Ausdruck der immunbiologischen Leistung dieses Organs
und die Follikelzentrumsreaktion spricht für die Synthese von Antikörpern. Dabei tritt
die follikuläre Hyperplasie zumeist bei chronischen, infektiösen Prozessen auf
(DAHME, E. u. E. WEISS 1999). Einige der Tiere wiesen chronische, entzündliche
Veränderungen in der Lunge oder dem Herzbeutel auf und erklären somit die
lymphatische Aktivierung. Die meisten Tiere jedoch zeigten keinerlei Heinweise auf
infektiöse Prozesse, so dass die follikuläre Hyperplasie in diesen Tieren
wahrscheinlich eher die Reaktion auf allgemeine, unspezifische Reize darstellt. Da
die untersuchten Tiere nicht keimfrei gehalten wurden, hatten sie jederzeit Kontakt zu

8 Diskussion 79
ubiquitären Keimen. Ein weiterer Befund in den untersuchten Lymphknoten war ein
Ödem und eine Sinushistiozytose. Diese Befunde können bei einer PCV II-Infektion
auftreten, aber auch Ausdruck einer unspezifischen Immunstimulation sein. Da die
betroffenen Tiere wieder nur zu einem Teil PCV II-positiv waren, sprechen die
Befunde hier eher für eine allgemeine, unspezifische Immunantwort. In den
Ileofemorallymphknoten von einigen Tieren fanden sich multifokal Siderophagen
(Berliner-Blau positiv) in den Randsinus. Dieses Phänomen resultiert daraus, dass
Ferkeln in den ersten Lebenstagen eine Eisensubstitution in die Kniefalte verabreicht
wird, um einer Anämie vorzubeugen. Bei den Siderophagen in den Lymphknoten
handelt es sich also um Makrophagen, die das injizierte Eisen phagozytiert haben.
Die Siderophagen in Lymphknoten werden regelmäßig angetroffen und stellen einen
Nebenbefund ohne klinische Relevanz dar. Weiterhin fanden sich in den
Lymphknoten mehrerer untersuchter Tiere multifokale, frische Blutungen. Da es sich
um ein akutes Geschehen ohne Hinweise auf Blutresorption handelte, ist dieser
Befund wahrscheinlich agonal im Rahmen der Tötung der Tiere entstanden.
Obwohl mehr als die Hälfte der untersuchten Tiere positiv für PCV II waren, fanden
sich keine Hinweise auf ein klinisch oder pathologisch manifestes PMWS. In Bezug
auf Xenotransplantationen muss jedoch berücksichtigt werden, dass PCV II eine
potentielle Zoonose für den Transplantatempfänger darstellt (HATTERMANN et al.,
2004). Daher sollten alle potentiellen Organspender auf mögliche Zoonoseerreger,
wie z.B. PCV II untersucht werden.
Insgesamt zeigen sowohl die Einzeltierbefunde als auch die Befunde in mehreren
Tieren kein bestimmtes Muster, so dass keine Hinweise auf einen spezifischen,
pathomorphologischen Phänotyp vorliegen. Dafür spricht auch, dass nicht-transgene
Kontrolltiere und transgene Tiere in gleicher Anzahl und im gleichen Ausmaß von
den Läsionen betroffen sind.
Die Ergebnisse der Expressionsanalyse der transgenen Tiere zeigten, dass die
hCD59-DNA in allen untersuchten Tieren nachweisbar war. Die hCD59-mRNA und
das hCD59-Protein hingegen zeigten eine starke Variabilität in ihrer Expression. Die
Expression variierte dabei sowohl zwischen den verschieden Linien als auch

8 Diskussion 80
zwischen verschiedenen Tieren der gleichen Linie. Auch innerhalb eines Tieres
wiesen die einzelnen Organe eine stark unterschiedliche Expression auf.
Die Tatsache, dass in allen untersuchten transgenen Tieren hCD59-DNA
nachgewiesen werden konnte, spricht für eine erfolgreiche Integration des
Konstruktes in das porzine Genom. Dass die Negativkontrollen und das untersuchte
nicht-transgene Kontrolltier keine DNA-Detektion zeigten, spricht für die Spezifität der
Primer und lässt falsch-positive Ergebnisse in den transgenen Tieren sehr
unwahrscheinlich erscheinen. Das wirft die Frage auf, warum das Konstrukt zwar
erfolgreich in das porzine Genom integriert wurde und zu einem ubiquitären hCD59-
DNA-Nachweis führt, aber nicht zu einer ebensolchen ubiquitären RNA- und Protein-
Expression.
Eine mögliche Erklärung für die inkonsistente Expression des Transgens ist der
Einfluss von Positionseffekten. Die in der Studie untersuchten Schweine wurden
mittels Mikroinjektionstechnik generiert. Bei dieser Methode erfolgt die Integration
des Konstrukts in das porzine Genom rein zufällig und daher ist nicht
vorauszusehen, ob benachbarte Gene einen Einfluss auf das integrierte Transgen
und seine Expression haben. Positionseffekte können eine Expression des
Transgens völlig hemmen, aber auch zu einem unerwarteten Expressionsmuster
führen. Weiterhin ist es möglich, dass bestimmte Regionen des Genoms eher zu
einer Transgenexpression führen als andere (CLARK et al., 1994). Um zu klären, ob
Positionseffekte eine Rolle bei der Expression des benutzten hCD59-Transgens
spielen, müsste in jedem Tier analysiert werden, an welchen Lokalisationen des
porzinen Genoms das Transgen integriert wurde. Da dies bislang nicht untersucht
wurde, kann eine Beteiligung von Positionseffekten an dem inkonsistenten
Expressionsmuster nicht ausgeschlossen werden. FESTENSTEIN et al. konnten
1996 zwei Arten von Positionseffekten unterscheiden: Einerseits eine verringerte
Expression in allen Geweben, andererseits eine variierende Expression, bei der das
Gen nur in einem Teil der Zellen exprimiert wird. Das zuletzt genannte Phänomen ist
in der Literatur bereits wiederholt als „Variegation“ beschrieben worden. Die
Variegation wurde bereits in Drosophila und Hefen beschrieben, wo eine Variation
der Genexpression mit einer Translokation von einem normalen, euchromatischen

8 Diskussion 81
Gen zu einem im Centromer gelegenen Heterochromatin korreliert war
(EISSENBERG, J. C., 1989; ALLSHIRE et al., 1994). FESTENSTEIN et al. konnten
1996 mittels Floureszenz in situ Hybridisierung in transgenen Mäusen zeigen, dass
in Mauslinien mit einer variierenden Expression das Transgen in
heterochromatischen Regionen nahe des Centromers lokalisiert war. Weiterhin
konnte diese Arbeitsgruppe zeigen, dass eine Inaktivierung von Genen mit einer
geschlossenen Chromatin-Konfiguration korrelierte. Dies spricht dafür, dass eine
Integration des Transgens im heterochromatischen Centromer, sowie eine
geschlossene Chromatin-Konfiguration zu einer gestörten Expression und damit zur
Variegation führen. Eine Lösung dieses Problems könnten Locus control regions
(LCR) sein. Dabei handelt es sich um Sequenzen, die zu einer starken und
gewebsspezifischen Expression des Transgens, unabhängig von Positionseffekten
führen. FESTENSTEIN et al. zeigten 1996, dass transgene Mäuse, die zusätzlich zu
dem Transgen eine flankierende LCR trugen, keine Variegation zeigten, auch wenn
das Transgen im Centromer lokalisiert war. Dabei führte die LCR wahrscheinlich zu
einer offenen statt einer geschlossenen Chromatin-Konfiguration. Ob die variierende
Expression in der Studie mit einer Lokalisation des Transgens im Heterochromatin
verbunden war und es sich bei der Variation um eine echte Variegation handelt,
wurde nicht untersucht. Es ist aber denkbar, dass derartige Positionseffekte eine
Rolle bei der inkonsistenten mRNA- und Proteinexpression spielen.
Ein weiterer Mechanismus, der die Transgenexpression negativ beeinflussen kann
ist die DNA-Methylierung. Durch eine Methylierung von bestimmten Regionen in der
DNA kommt es zu einer reversiblen Ausschaltung dieser Gene (TYCKO, 1997).
Mehrere Studien konnten bereits zeigen, dass die Methylierung mit der integrierten
Kopienzahl des Transgens zunimmt. So könnte die Anzahl der Kopien über eine
verstärkte Methylierung einen Einfluss auf die Expressionsstärke oder das
Expressionsmuster haben. Ob dieser Mechanismus im vorliegenden Fall
verantwortlich ist, konnte jedoch nicht geklärt werden, da eine Methylierungsanalyse
nicht durchgeführt wurde. Die Anzahl der Transgenkopien könnte auch auf rein
quantitativen Weg einen positiven Einfluss auf die Expression haben, indem z.B.
mehr integrierte Transgenkopien zu einer stärkeren Expression führen (CLARK et al.,

8 Diskussion 82
1994). Durch die erhöhte Anzahl von Kopien wäre auch der Transktivator mehrfach
integriert, und da dieser zu einer verstärkten Expression führt, wäre auch eine
Beeinflusung der Expression auf diesem Weg denkbar. Eine quantitative Analyse der
hCD59-mRNA ist geplant, es liegen jedoch noch keine konkreten Ergebnisse vor.
In transgenen Mäusen wurden Locus control regions (LCR) identifiziert, die zu einer
starken und gewebsspezifischen Expression von Transgenen in den Mäusen geführt
haben. Durch die LCRs ist die Expression dabei unabhängig von der
Integrationsstelle des Transgens und kann so Positionseffekten entgegenwirken.
Weiterhin ist bekannt, dass der Promotor einen großen Einfluss auf die Expression
des Transgens hat. Viele Promotoren besitzen eine Gewebsspezifität und führen nur
in bestimmten Organen zu einer Expression des Transgens (FODOR et al., 1994,
COWAN et al., 1998). Der in dieser Studie gewählte CMV-Promotor induzierte in
früheren Studien eine starke und ubiquitäre Expression in Zelllinien und transgenen
Mäusen (BOSHART et al., 1985; FURTH et al., 1991), und auch NIEMANN et al.
wiesen 2001 mit dem CMV-Promotor eine Expression von hCD59 in vielen, wenn
auch nicht in allen Organen und Geweben nach. Ein weiterer Grund, der gegen eine
Gewebsspezifität des CMV-Promotors als alleinige Ursache der variierenden
Expression spricht, ist die Tatsache, dass die Expression der hCD59-mRNA und des
Proteins nicht auf bestimmte Organe oder Gewebe begrenzt ist, sondern variierend
in allen untersuchten Organen und Geweben nachgewiesen werden konnte. Dies
lässt auf eine ubiquitäre Expression des CMV-Promotors schließen, erklärt aber nicht
die starken Variationen zwischen den einzelnen Tieren und den verschiedenen
Organen und Geweben.
Eine weitere denkbare Ursache für die uneinheitliche Expression ist die Ausbildung
eines Chimärismus in den transgenen Tieren in dem Sinne, dass das Konstrukt nicht
in Zellen aller Organe und Gewebe integriert wurde, sondern nur ein einem Teil der
Zellen vorhanden ist. Diese Hypothese ist jedoch äußerst unwahrscheinlich, da die
hCD59-DNA in allen untersuchten Organen und Geweben nachgewiesen wurde, was
für eine erfolgreiche Integration des Konstruktes spricht. Zudem wären von einem
Chimärismus nur Tiere der F1-Generation betroffen, da sich bei erneuter Kreuzung
der Tiere der Chimärismus aufheben würde. In der Studie fand sich das variierende

8 Diskussion 83
Expressionsmuster jedoch sowohl in Tieren der F1-Generation als auch in Tieren der
F2-Generation.
Einige Tiere wiesen in einem Teil der Organe zwar eine Expression der hCD59-
mRNA auf, exprimierten in diesen Organen jedoch nicht immer das hCD59-Protein.
Eine genaue Ursache hierfür wurde nicht gefunden, aber es ist denkbar, dass die
mRNA zwar transkribiert, aber nicht translatiert wurde.
Da die hCD59-DNA in allen untersuchten Tieren nachgewiesen wurde, die
Expression der mRNA und des Proteins jedoch stark schwankte, liegt wahrscheinlich
eine Störung der Transkription und/oder der Translation in den Tieren vor. Eine
genaue Ursache oder ein Mechanismus dieser Störung konnte jedoch in den
untersuchten Tieren bislang nicht geklärt werden.
Bei zwei Tieren (TG4, TG6) wurde immunhistochemisch zwar hCD59-Protein
nachgewiesen, aber keine hCD59-mRNA. Auch in den Tieren mit einer variierenden
mRNA- und Proteinexpression zeigten einige Organe eine Proteinexpression ohne
den Nachweis von mRNA. Die Synthese des Proteins ist jedoch nur möglich, wenn
entsprechende mRNA vorhanden ist.
Eine mögliche Erklärung für diese widersprüchlichen Ergebnisse ist, dass die
hCD59-mRNA und das hCD59-Protein möglicherweise unterschiedliche
Halbwertszeiten haben. Wenn das Protein eine längere Halbwertszeit besitzt, wäre
es möglich, dass das Protein noch nachweisbar ist, obwohl die mRNA bereits
abgebaut ist. Da bislang nichts über die Halbwertszeiten von hCD59-mRNA und -
Protein bekannt ist, kann diese Möglichkeit nicht ausgeschlossen werden.
Eine weitere Möglichkeit ist, dass die Immunhistochemie zum Teil falsch positive
Ergebnisse geliefert hat. Während die Anfärbung der Kardiomyozyten und der
Myozyten im Skelettmuskel eindeutig positiv war, war die Anfärbung des Endothels
auf Grund einer unspezifischen Anfärbung des Blutplasmas nicht immer eindeutig.
Da es sich bei den Organen und Geweben, in denen das Protein, aber nicht die
mRNA nachgewiesen wurde, immer um endotheliale Proteinexpression handelte,
kann die Möglichkeit einer falsch positiven Anfärbung nicht ausgeschlossen werden.

8 Diskussion 84
Um diese Möglichkeit zu überprüfen, wurden die Organe von zwei Tieren (KO8,
TG25) direkt nach der Organentnahme perfundiert, um die Blutgefäße von
Blutplasma zu befreien und dadurch eine unspezifische Anfärbung zu verhindern.
Auf Grund der Organgröße war eine vollständige Perfusion, vor allem der kleineren
Blutgefäße jedoch nicht möglich, so dass die Möglichkeit einer unspezifischen
Anfärbung auch auf diesem Weg nicht völlig ausgeschlossen werden konnte.
Ein weiteres Phänomen, das bei der Expressionsanalyse beobachtet wurde, ist das
so genannte „patchy pattern“ der hCD59-Proteinexpression im Herz, Skelettmuskel
und der Niere von einigen Tieren. Dabei zeigten einige Zellen der Organe eine
deutliche hCD59-Proteinexpression, während direkt benachbarte Zellen keine
Proteinexpression aufwiesen (Abb. 12, Abb. 13, Abb. 14). Ein ähnliches
Expressionsmuster beobachteten auch CHARREAU et al. 1996 in hCD59-
transgenen Ratten. Die Ratten dieser Studie exprimierten das hCD59 ebenfalls unter
der Kontrolle eines CMV-Promotors. Insgesamt zeigten die transgenen Ratten von
CHARREAU et al. 1996 eine starke hCD59-Proteinexpression in vielen Organen und
Geweben, wie z.B. im Pankreas, Herz, Gehirn, Niere, Skelettmuskel und dem
Intestinaltrakt. Im Gegensatz zur vorliegenden Studie wiesen die Tiere jedoch kein
hCD59-Protein in Endothelzellen auf. Zudem wurde immunhistochemisch ein
„patchy pattern“ des Proteins im Herz und in der Niere nachgewiesen. Auch
BASKAR et al. wiesen ein sehr variables Expressionsmuster nach Einsatz eines
CMV-Promotors nach. In ihrer Studie konstruierten sie lacZ-transgene Mäuse, die
das Transgen unter der Kontrolle des HCMV (human cytomegalovirus) MIEP (major
immediate-early Promotor) Promotors exprimierten. Immunhistochemisch konnten
sie das lacZ-Protein nur in 19 von 29 untersuchten Organen und Geweben
nachweisen, wobei Herz und Muskel ein „patchy pattern“ aufwiesen (BASKAR et al.,
1996).
Diese Expressionsmuster ähneln dem Expressionsmuster des hCD59-Proteins
hinsichtlich des „patchy patterns“ in der vorliegenden Studie, wenn auch die
Verteilung der Proteinexpression in den verschiedenen Organen und Geweben nicht
identisch ist. Da in den aufgeführten Studien ebenfalls ein CMV-Promotor die

8 Diskussion 85
Expression des Transgens kontrollierte, ist es gut denkbar, dass dieser Promotor
eine gewisse Zellspezifität besitzt, die in dem eigentümlichen „patchy pattern“ zum
Ausdruck kommt. Bislang ist nicht bekannt, welche Kriterien die positiven Zellen von
den negativen Zellen unterscheiden, so dass ungeklärt bleibt, auf welche Zelltypen
sich die Spezifität bezieht. Um das Pänomen des „patchy patterns“ auszuschalten,
könnte die Verwendung eines anderen Promotors sinnvoll sein.
Auch die Verteilung der Transgenexpression nach Einsatz eines CMV-Promotors ist
in den verschiedenen Studien sehr unterschiedlich. Auch COWAN et al. konnten
1998 zeigen, dass das Expressionsmuster eines Transgens stark von dem
eingesetzten Promotor abhängt. Das humane Cytomegalovirus ist in der Lage, ein
großes Spektrum an Organen und Geweben zu infizieren (z.B. Zellen in
Speicheldrüse, Lunge, Leber, Pankreas, Niere, Auge, Ohr, Plazenta,
Verdauungstrakt, Herz, Ovarien, Nebennieren, Gehirn, Schilddrüse und Haut). Dabei
befällt das Cytomegalovirus die unterschiedlichsten Zelltypen, unter anderem auch
Endothelzellen und Epithelien (FIELDS 2001). Diese Fähigkeit des Cytomegalovirus,
ein großes Spektrum an Zelltypen, Organen und Geweben zu infizieren, macht man
sich zunutze, um mit Hilfe des Cytomegalovirus-Promotors eine möglichst ubiquitäre
Expression des Transgens zu erreichen. Obwohl die Untersuchungen zur
Transgenexpression nach Einsatz des CMV-Promotors unterschiedlicher
Arbeitsgruppen bislang unterschiedliche Verteilungsmuster aufwiesen, konnte in der
vorliegenden Studie eine Expression des hCD59-Proteins in nahezu allen
untersuchten Organen und Geweben nachgewiesen werden.
Die Expression des hCD59-Proteins in Endothelzellen ist von besonderer Bedeutung
für den protektiven Effekt des Transgens. NIEMANN et al. konnten 2001 bereits an
den in der Studie verwendeten transgenen Tieren nachweisen, dass das hCD59 in
Fibroblasten und Endothelzellen aus der Aorta der transgenen Schweine zu einer
erheblich herabgesetzten Sensitivität gegenüber humanen Antikörpern und
humanem Komplement führte. Weiterhin zeigten Nieren der transgenen Tiere bei
einer ex vivo Perfusion mit humanem Blut eine deutlich längere Überlebenszeit als
Nieren von nicht-transgenen Kontrolltieren. Diese Ergebnisse sprechen dafür, dass
das eingesetzte hCD59-Transgen einen protektiven Effekt bietet.

8 Diskussion 86
Zusammenfassend lässt sich sagen, dass die in der Studie untersuchten hCD59-
transgenen Schweine durch die Integration der humanen DNA keinen spezifischen,
pathomorphologischen Phänotyp entwickelten.
Die hCD59-DNA konnte in allen untersuchten Organen und Geweben nachgewiesen
werden, was für eine erfolgreiche Integration des eingesetzten Konstrukts in das
porzine Genom spricht.
Die Expression der hCD59-mRNA und des hCD59-Proteins wies jedoch eine starke
Variabilität zwischen den verschiedenen Linien, zwischen Tieren einer Linie sowie
zwischen verschiedenen Organen und Geweben jedes einzelnen Tieres auf. Zudem
wurde in der Skelettmuskulatur, im Herz und in der Niere ein so genanntes „patchy
pattern“ der Proteinexpression beobachtet.
Diese stark variierende Expression ist ein Problem, das vor dem Einsatz der Tiere für
Xenotransplantationen noch gelöst werden muss, da die Organe, die keine hCD59-
Proteinexpression aufweisen, wahrscheinlich hyperakut abgestoßen würden.
Das eingesetzte Transgen konnte in bereits durchgeführten Versuchen
Schweinenieren vor einer hyperakuten Abstoßung durch humanes Blut schützen
(NIEMANN et al., 2001), doch eine akute Abstoßung der Organe konnte nicht
verhindert werden.
Vor einem erfolgreichen Einsatz von transgenen Schweineorganen für die
Xenotransplantation ist noch viel Forschungsarbeit erforderlich, doch vielleicht kann
der Einsatz anderer Promotoren, die Etablierung von α-1-3-Galactosyltransferase
knockout-Schweinen oder der Einsatz mehrerer Transgene zu weiteren Fortschritten
auf dem Gebiet der Xenotransplantation führen.

87
9 Zusammenfassung
Stefanie Deppenmeier
Pathomorphologische Phänotypisierung und Expressionsanalyse von hCD59-transgenen Schweinen für Xenotransplantationen
Der Mangel an humanen Spenderorganen für Transplantationen hat in den letzten
Jahren zu einem gesteigerten Interesse an der Xenotranplantation von
Schweineorganen geführt. Um eine hyperakute Abstoßung der porzinen
Spenderorgane zu verhindern, wurden mittels Mikroinjektion transgene Schweine
konstruiert, die ein Konstrukt mit dem humanen Komplementregulator hCD59 unter
der Kontrolle eines Cytomegalovirus (CMV)-Promotors tragen. Dieses Konstrukt soll
die Abstoßung des Spenderorgans über eine Hemmung der Aktivierung der
Komplementkaskade verhindern.
Die durchgeführten Untersuchungen ergaben keinen Hinweis auf einen spezifischen,
pathomorphologischen Phänotyp der transgenen Tiere. Einige Tiere zeigten
pathomorphologische Läsionen, die jedoch als „unspezifische“ Hintergrundpathologie
interpretiert wurden. Alle untersuchten Tiere wiesen eine Detektion der hCD59-DNA
in allen untersuchten Organen und Geweben auf. Die hCD59-mRNA und das
hCD59-Protein zeigte eine breite Verteilung in den untersuchten Organen und
Geweben und konnte in Herz, Lunge, Milz, Leber, Niere, Pankreas, Gehirn,
Skelettmuskel und Verdauungstrakt nachgewiesen werden. Dabei variierte die
Expression der mRNA und des Proteins stark zwischen den verschiedenen Linien,
zwischen Tieren einer Linie sowie zwischen verschiedenen Organen und Geweben
jedes einzelnen Tieres. Im Herz, in der Niere und im Skelettmuskel zeigte die
Expression zudem ein „patchy pattern“, indem sich nur einzelne Zellen der Organe
distinkt positiv anfärbten. Es ist möglich, dass für dieses Expressionsmuster die so
genannte „Variegation“ verantwortlich ist, die durch eine Translokation eines Gens
verursacht wird und bereits in Hefen, Drosophila und Mäusen beschrieben wurde.

9 Zusammenfassung 88
Die ubiquitäre hCD59-DNA-Detektion spricht für eine erfolgreiche Integration des
hCD59-Konstrukts, während die mRNA- und Protein-Expression eine starke
Variation aufweist. Insgesamt führt der eingesetzte CMV-Promotor jedoch zu einer
weitreichenden Expression des Transgens in den verschiedenen Organen und
Geweben.
Insgesamt ist der Einsatz der transgenen Schweine für Xenotransplantationen
problematisch, solange das hCD59-Protein ein variierendes Expressionsmuster
aufweist, da Organe und Gewebe die das Protein nicht exprimieren wahrscheinlich
hyperakut abgestoßen würden.

89
10 Summary
Stefanie Deppenmeier
Pathomorphologic phenotyping and expression pattern analysis of hCD59-transgenic pigs designed for xenotransplantation
The shortage of human donor organs in recent years has led to an increasing interest
in the xenotransplantation of porcine organs. To prevent the hyperacute rejection of
the porcine donor organs, transgenic pigs were designed via microinjection of a
cytomegalovirus (CMV)-Promotor driven construct that contained the human
complement regulator hCD59. This construct should overcome the rejection of the
donor organ by inhibiting the activation of the complement cascade.
The accomplished examinations gave no hints to a specific pathomorphologic
phenotype in the transgenic animals. Some animals showed pathomorphologic
lesions, but these were interpreted as background pathology. All animals examined
had a detection of hCD59-DNA in all organs and tissues examined. The hCD59-
mRNA and the hCD59-protein showed a wide distribution in the organs and tissues
examined and could be detected in heart, lung, spleen, liver, kidney, pancreas, brain,
skeletal muscle and the intestinal tract. The mRNA und protein expression showed a
large variation between animals of different lines, between animals of one line and
between different organs and tissues of individual animals.
In addition, in heart, kidney and skeletal muscle the protein expression showed a
patchy expression pattern with only single positively staining cells within the organs.
It is possible, that the so called „variegation“, which is caused by a translocation of a
gene and was described already in yeast, Drosophila and mice, is responsible for
the described expression pattern.
The ubiquitous hCD59-DNA detection demonstrates the successful integration of the
hCD59-construct, whereas the expression of the mRNA and the protein shows a
large variation. All in all the CMV-Promotor leads to an extensive expression of the
transgene in different organs and tissues.

10 Summary 90
Taken together, the application of transgenic pigs is problematic as long as the
hCD59-protein has a varying expression pattern, since organs and tissues which do
not express the protein would probably be rejected hyperacutely.

91
11 Literaturverzeichnis
ADAMS, D. H., A. KADNER, R. H. CHEN u. R. S. FARIVAR (2001):
Human membrane cofactor protein (MCP, CD 46) protects transgenic pig hearts from
hyperacute rejection in primates
Xenotransplantation 8, S. 36-40
AKSENTIJEVICH, I., D. H. SACHS u. M. SYKES (1992):
Humoral tolerance in xenogeneic BMT recipients conditioned by a nonmyeloablative
regimen
Transplantation 53, S. 1108-1114
ALLSHIRE, R. C., J. P. JAVERZAT, N. J. REDHEAD u. G. CRANSTON (1994):
Position effects variegation at fission yeast centromeres
Cell 76, S. 157-169
ALWAYN, I. P. J., M. BASKER, L. BUHLER u. D. K. C. COOPER (1999):
The problem of anti-pig antibodies in pig-to-primate xenografting: current and novel
methods of depletion and/or suppression of production of anti-pig antibodies
Xenotransplantation 6, S. 157-168
ASHTON-CHESS, J., J.-C. ROUSSEL, R. MANEZ, C. RUIZ, A. MOREAU, E. COZZI,
D. MINAULT, J.-P. SOULILLOU u. G. BLANCHO (2003) :
Cellular participation in delayed xenograft rejection of hCD55 transgenic pig hearts
by baboons
Xenotransplantation 10, S. 446-453
BACH, F. H., H. WINKLER, C. FERRAN, W. W. HANCOCK u. S. C. ROBSON
(1996):
Delayed xenograft rejection
Immunol Today 17, S. 379-384

11 Literaturverzeichnis 92
BACH, F. H., M. A. TURMAN, G. M. VERCELLOTTI, J. L. PLATT u. A. P.
DALMASSO (1991):
Accomodation: A working paradigm for progressing toward clinical discordant
xenografting
Transplant Proc 23, S. 205-207
BASKAR, J. F., P. P. SMITH, G. NILAVER, R. A. JUPP, S. HOFFMANN, N. J.
PEFFER, D. J. TENNEY, A. M. COLBERG-POLEY, P. GHAZAL u. J. A. NELSON
(1996):
The enhancer domain of the human cytomegalovirus major immediate-early
Promotor determines cell type-specific expression in transgenic mice
J Virol 70, S. 3207-3214
BLUSCH, J. H., C. PATIENCE u. U. MARTIN (2002):
Pig endogenous retroviruses and xenotransplantation
Xenotransplantation 9, S. 242-251
BOCK, O., H. KREIPE u. U. LEHMANN (2001):
One-step extraction of RNA from archival biopsies
Anal Biochem 295, S. 116-117
BOSHART, M., F. WEBER, G. JAHN, K. DORSCH-HASLER, B. FLECKENSTEIN u.
W. SCHAFFNER (1985):
A very strong enhancer is located upstream of an immediate early gene of human
cytomegalovirus
Cell 41, S. 521-530

11 Literaturverzeichnis 93
BYRNE, G. W., K. R. McCURRY, M. J. MARTIN, S. M. McCLELLAN, J. L. PLATT u.
J. S. LOGAN (1997):
Transgenic pigs expressing humen CD59 and decay-accelerating factor produce an
intrinsic barrier to complement-mediated damage
Transplantation 63, S. 149-155
CAMPBELL, K. H. S., J. McWHIR, W. A. RITCHIE u. I. WILMUT (1996):
Sheep cloned by nuclear transfer from a cultured cell line
Nature 380, S. 64-66
CAPECCHI, M. R. (2000):
How close are we to implementing gene targeting animals in other than the mouse?
Proc Natl Acad Sci USA 97, S. 956-957
CARRINGTON, C. A. u. G. DOS SANTOS CRUZ (2001) :
Effect of cell surface concentration of human DAF on transgenic pig aortic endothelial
cells on the degree of protection afforded against human complement deposition
Xenotransplantation 8, S. 100-105
CARTER, D. B., L. LAI, K.-W. PARK, M. SAMUEL, J. C. LATTIMER, K. R. JORDAN,
D. M. ESTES, C. BESCH-WILLIFORD u. R. S. PRATHER (2002):
Phenotyping of transgenic cloned piglets
Cloning Stem Cells 4, S. 131-145
CHALINE, J., D. CARDOSO u. D. HOUSSIN (1994):
Organ xenografting between rodents: An evolutionary perspective
Transplant Int 7, S. 216-221

11 Literaturverzeichnis 94
CHARREAU, B., L. TESSON, J. BUSCAIL, J. P. SOULILLOU u. I. ANEGON (1996):
Analysis of human CD59 tissue expression directed by the CMV-IE-1 Promotor in
transgenic rats
Transgenic Res 5, S. 443-450
CHEN, G., Q.-Y. SUN, X.-M. WANG, S.-Q. SHEN, H. GUO, H. WANG, Y. WU, W.-Y.
WANG, Y.-L. XIONG u. S. CHEN (2004):
Improved suppression of circulating complement does not block acute vascular
rejection of pig-to-rhesus monkey cardiac transplants
Xenotransplanation 11, S. 123-132
CHEN, R. H., R. N. MITCHELL, A. KADNER u. D. H. ADAMS (1999):
Differential galactose α(1,3) galactose expression by porcine cardiac vascular
endothelium
Xenotransplantation 6, S. 169-172
CHEN, R. H., S. NAFICY, J. S. LOGAN, L. E. DIAMOND u. D. H. ADAMS (1999):
Hearts from transgenic pigs constructed with CD59/DAF genomic clones
demonstrate improved survival in primates
Xenotransplantation 6, S. 194-200
CIBELLI, J. B., S. L. STICE, P. J. GOLUEKE, J. J. KANE, J. JERRY, C.
BLACKWELL, F. ABEL PONCE DE LEÓN u. J. ROBI (1998):
Cloned transgenic calves produced from nonquiescent fetal fibroblasts
Science 280, S. 1256-1258
CLARK, A. J., P. BISSINGER, D. W. BULLOCK, S. DAMAK, R. WALLACE, C. B. A.
WHITELAW u. F. YULL (1994):
Chromosomal position effects and the modulation of transgene expression
Reprod Fertil Dev 6, S. 589-598

11 Literaturverzeichnis 95
CLARK, D. A., J. F. L. FRYER, A. W. TUCKER, P. D. MCARDLE, A. E. HUGHES, V.
C. EMERY u. P. D.GRIFFITHS (2003):
Porcine cytomegalovirus in pigs being bred for xenograft organs: progress towards
control
Xenotransplantation 10, S. 142-148
COLLINS, B. H., A. H. COTTERELL, K. R. McCURRY, C. G. ALVARADO, J. C.
MAGEE, W. PARKER u. J. L. PLATT (1995):
Cardiac xenografts between primate species provide evidence for the importance of
the α-galactosyl determinant in hyperacute rejection
J Immunol 154, S. 5500-5510
COOPER, D. K. C., B. GOLLACKNER u. D. H. SACHS (2002):
Will the pig solve the transplantation backlog?
Annu Rev Med 53, S. 133-147
COOPER, D. K. C., E. KEMP, J. L. PLATT u. D. J. WHITE (1996):
Xenotransplantation: The transplantation of organs and tissues between species
2. Auflage, Verlag Springer, Berlin
COSTA, C., L. ZHAO u. W. L. FODOR (1999):
Comparative analysis of three genetic modifications designed to inhibit human
serum-mediated cytolysis
Xenotransplantation 6, S. 6-16
COSTA, C., L. ZHAO, W. V. BURTON, C. ROSAS, K. R. BONDIOLI, B. L.
WILLIAMS, T. A. HOAGLAND, A. P. DALMASSO u. W. L. FODOR (2002):
Transgenic pigs designed to express human CD59 and H-transferase to avoid
humoral xenograft rejection
Xenotransplantation 9, S. 45-57

11 Literaturverzeichnis 96
COWAN, P. J., A. AMINIAN, H. BARLOW, A. A. BROWN, C.-G. CHEN, N.
FISICARO, D. M. A. FRANCIS, D. J. GOODMAN, W. HAN, M. KUREK, M. B.
NOTTLE, M. J. PEARSE, E. SALVARIS, T. A. SHINKEL, G. V. STAINSBY, A. B.
STEWART u. J. F. D’APICE (2000):
Renal xenografts from triple-transgenic pigs are not hyperacutely rejected but cause
coagulopathy in non-immunosuppressed baboons
Transplantation 69, S. 2504-2515
COWAN, P. J., C. A. SOMERVILLE, T. A. SHINKEL, M. KATERELOS, A. AMINIAN,
M. ROMANELLA, M. J. TANGE, M. J. PEARSE u. A. J. F. D’APICE (1998):
High-level endothelial expression of human CD59 prolongs heart function in an ex
vivo model of xenograft rejection
Transplantation 65, S. 826-831
COZZI, E., A. W. TUCKER, G. A. LANGFORD, G. PINO-CHAVEZ, L. WRIGHT, M.-J.
O’CONNELL, V. J. YOUNG, R. LANCASTER, M. McLAUGHLIN, K. HUNT, M. C.
BORDIN u. D. J. G. WHITE (1997):
Characterization of pigs transgenic for human decay-accelerating factor
Transplantation 64, S. 1383-1392
COZZI, E., F. BHATTI, M. SCHMOECKEL, G. CHAVEZ, K. G. C. SMITH, A. ZAIDI,
J. R. BRADLEY, S. THIRU, M. GODDARD, C. VIAL, D. OSTLIE, J. WALLWORK, D.
J. G. WHITE u. P. J. FRIEND (2000):
Long-term survival of nonhuman primates receiving life-supporting transgenic porcine
kidney xenografts
Transplantation 70, S. 15-21

11 Literaturverzeichnis 97
CZAPLICKI, J. u. B. BLONSKA (1992):
The lack of hyperacute xenogeneic heart trasnplant rejection in a human
J Heart Lung Transplant 11, S. 393-396
DAHME, E. u. E. WEISS (1999):
Grundriß der speziellen pathologischen Anatomie der Haustiere
5. Auflage, Ferdinand Enke Verlag, Stuttgart
DAI, Y., T. D. VAUGHT, J. BOONE, S.-H. CHEN, C. J. PHELPS, S. BALL, J. A.
MONAHAN, P. M. JOBST, K. J. McCREATH, A. E. LAMBORN, J. L. COWELL-
LUCERO, K.-D. WELLS, A. COLMAN, I. A. POLEJAEVA u. D. L. AYARES (2002):
Targeted disruption of the α1,3-galactosyltransferase gene in cloned pigs
Nature Biotechnol 20, S. 251-255
DALMASSO, A. P., G. M. VERCELLOTTI, R. J. FISCHEL, R. M. BOLMAN, F. H.
BACH u. J. L. PLATT (1992):
Mechanism of complement activation in the hyperacute rejection of porcine organs
transplanted into primate recipients
Am J Pathol 140, S. 1157-1166
DALMASSO, A. P., u. J. L. PLATT (1993):
Prevention of complement-mediated activation of xenogeneic endothelial cells in an
in vitro model of xenograft hyperacute rejection by C1 inhibitor
Transplantation 56, S. 1171-1176
DAVIES, A., D. L. SIMMONS, G. HALE, R. A. HARRISON, H. TIGHE, P. J.
LACHMANN u. H. WALDMANN (1989) :
CD59, an LY-6-like protein expressed in human lymphoid cells, regulates the action
of the complement membrane attack complex on homologous cells
J Exp Med 170, S. 637-654

11 Literaturverzeichnis 98
DIAMOND, L. E., C. M. QUINN, M. J. MARTIN, J. LAWSON, J. L. PLATT u. J. S.
LOGAN (2001):
A human CD46 transgenic pig model system for the study of discordant
xenotransplantaion
Transplantation 71, S. 132-142
DIAMOND, L. E., K. R. McCURRY, M. J. MARTIN, S. B. McCLELLAN, E.
R.OLDHAM, J. L. PLATT u. J. S. LOGAN (1996):
Characterization of transgenic pigs expressing functionally active human CD59 on
cardiac endothelium
Transplantation 61, S. 1241-1249
DOR, F. J. M. F., J. CHENG, A. ALT, D. K. C. COOPER u. H.-J. SCHUURMAN
(2004):
Gal α 1,3Gal expression on porcine pancreatic islets, testis, spleen and thymus
Xenotransplantation 11, S. 101-106
EISSENBERG, J. C. (1989):
Position effect variegation in Drosophila: towards a genetics of chromatin assembly
Bioassays 11, S. 14-17
ERICSSON, T. A., Y. TAKEUCHI, C. TEMPLIN, G. QUINN, S. F. FARHADIAN, J. C.
WOOD, B. A. OLDMIXON, K. M. SULING, J. K. ISHII, Y. KITAGAWA, T.
MIYAZAWA, D. R. SALOMON, R. A. WEISS u. C. PATIENCE (2003):
Identification of receptors for pig endogenous retrovirus
Proc Natl Acad Sci USA 100, S. 6759-6764
FESTENSTEIN, R., M. TOLAINI, P. CORBELLA, C. MAMALAKI, J. PARRINGTON,
M. FOX, A. MILIOU, M. JONES u. D. KIOUSSIS (1996):
Locus control region function and heterochromatin-induced position effect variegation
Science 271, S. 1123-1125

11 Literaturverzeichnis 99
FIELDS (2001):
Virology
4. Auflage, Bd. 2, Verlag Lippincott Williams & Wilkins, Philadelphia, USA
FISCHEL, R. J., R. M. BOLMANN III, J. L.PLATT, J. S. NAJARIAN, F. H. BACH u. A.
J. MATAS (1990):
Removal of IgM anti-endothelial antibodies result in prolonged cardiac xenograft
survival
Transplant Proc 22, S. 1077-1078
FODOR, W. L., B. L. WILLIAMS, L. A. MATIS, J. A. MADRI, S. A. ROLLINS, J. W.
KNIGHT, W. VELANDER u. S. P. SQUINTO (1994):
Expression of a functional human complement inhibitor in a transgenic pig as a
model for the prevention xenogeneic hyperacute organ reaction
Proc Natl Acad Sci USA 91, S. 11153-11157
FURTH, P. A., L. HENNIGHAUSEN, C. BAKER, B. BEATTY u. R. WOYCHICK
(1991):
The variability in activity of the universally expressed human cytomegalovirus
immediate early gene 1 enhancer/Promotor in transgenic mice
Nucleic Acids Res 19, S. 6205-6208
FURTH, P. A., L. ST. ONGE, H. BOGER, P. GRUSS, M. GOSSEN, A. KISTNER, H.
BUJARD u. L. HENNIGHAUSEN (1994):
Temporal control of gene expression in transgenic mice by a tetracycline-responsive
Promotor
Proc Natl Acad Sci USA 91, S. 9302-9306
GALILI, U., D. C. LATEMPLE, A. W. WALGENBACH u. K. R. STONE (1997):
Porcine and bovine cartilage transplants in cynomolgus monkey
Transplantation 63, S. 646-651

11 Literaturverzeichnis 100
GALILI, U., E. A. RACHMILEWITZ, A. PELEG u. I. FLECHNER (1984):
A unique natural human IgG antibody with anti-α-galactosyl specifity
J Exp Med 160, S. 1519-1531
GALILI, U., M. R. CLARK, S. B. SHOHET, J. BUEHLER u. B. A. MACHER (1987):
Evolutionary relationship between the natural anti-Gal antibody and the Galα1-3Gal
epitopes in primates
Proc Natl Acad Sci USA 84, S. 1369-1373
GALILI, U., R. E. MANDRELL, R. M. HAMADEH, S. B. SHOHET u. J. Mc LEOD
GRIFFISS (1988):
Interaction between human natural anti-α-galactosyl immunoglobulin G and bacteria
of the human flora
Infect Immun 56, S. 1730-1737
GALILI, U., S. B. SHOHET, E. KOBRIN, C. L. M. STULTS u. B. A. MACHER (1988):
Man, apes and old world monkeys differ from other mammals in the expression of α-
Galactosyl epitopes on nucleated cells
J Biol Chem 263, S. 17755-17762
GALILI, U. u. K. L. MATTA (1996):
Inhibition of anti-Gal IgG binding to porcine endothelial cells by synthetic
oligosaccharides
Transplantation 62, S. 256-262
GODDARD, M. J., J. J. DUNNING, J. HORSELEY, C. ATKINSON, G. PINO-
CHAVEZ u. J. WALLWORK (2002):
Histopathology of cardiac xenograft rejection in the pig-to-baboon model
J Heart Lung Transplant 21, S. 474-484

11 Literaturverzeichnis 101
GRIERSON, S. S., D. P. KING, T. SANDVIK, D. HICKS, Y. SPENCER, T. W. DREW
u. M. BANKS (2004):
Detection and genetic typing of type 2 porcine circoviruses in archived pig tissues
from the UK
Archives Virology 149, S. 1171-1183
GRUBER, A. D. u. R. A. LEVINE (1997):
In situ assessment of mRNA accessibility in heterogenous tissue samples using
elongation factor-1α (EF-1α)
Histochem Cell Biol 107, S. 411-416
HAMMER, C., R. LINKE, F. WAGNER u. M. DIEFENBECK (1998):
Organs from animals for man
Int Arch Allergy Immunol 116, S. 5-21
HANNA, S. M., G. T. WILLIAMS, C. W. VAN DEN BERG u. B. P. MORGAN (1998):
Characterization in vitro and in vivo of the pig analogue of human CD59 using new
monoclonal antibodies
Immunology 95, 450-459
HATTERMANN, K., C. ROEDNER, C. SCHMITT, T. FINSTERBUSCH, T.
STEINFELDT u. A. MANKERTZ (2004):
Infection studies on human cell lines with porcine circovirus type 1 and porcine
circovirus type 2
Xenotransplanation 11, S. 184-294

11 Literaturverzeichnis 102
HENEINE, W., A. TIBELL, W. M. SWITZER, P. SANDSTROM, G. V. ROSALES, A.
MATHEWS, O. KORSGREN, L. E. CHAPMAN, T. M. FOLKS u. C. G. GROTH
(1998):
No evidence of infection with porcine endogenous retrovirus in recipients of porcine
islet-cell xenografts
Lancet 352, S. 695-699
HERRING, C. D. A. CUNNINGHAM, A. J. WHITTAM, X. M. FERNANDEZ-SUAREZ
u. G. A. LANGFORD (2001):
Monitoring xenotransplant recipients for infection by PERV
Clin Biochem 34, S. 23-27
HILL, J. R., A. J. ROUSSEL, J. B. CIBELLI, J. F. EDWARDS, N. L. HOOPER, M. W.
MILLER, J. A. THOMPSON, C. R. LOONEY, M. E. WESTHUSIN, J. M. ROBB u. S.
L. STICE (1999):
Clinical and pathologic features of cloned transgenic calves and features (13 case
studies)
Theriogenology 51, S. 1451-1465
IERINO, F. L., T. KOZLOWSKI, J. B. SIEGEL, A. SHIMIZU, R. B. COLVIN, P. T.
BANERJEE, D. K. C. COOPER, A. B. COSIMI, F. H. BACH, D. H. SACHS u. S. C.
ROBSON (1998):
Disseminated intravascular coagulation in association with the delayed rejection of
pig-to-baboon renal xenografts
Transplantation 66, S. 1439-1450
JUBB, K. V. F., P. C. KENNEDY u. N. PALMER (1993):
Pathology of domestic animals
4. Auflage, Bd. 2, Verlag Academic Press, San Diego, USA

11 Literaturverzeichnis 103
KOBAYASHI, T., S. TANIGUSHI, F. A. NEETHLING, A. G. ROSE, W. W.
HANCOCK, Y. YE, M. NIEKRASZ; S. KOSANKE, L. J. WRIGHT, D. J. G. WHITE u.
D. K. C. COOPER (1997):
Delayed xenograft rejection of pig-to-baboon cardiac transplants after cobra venom
factor therapy
Transplantation 64, S. 1255-1261
KOZLOWSKI, T., A. SHIMIZU, D. LAMBRIGTS, K. YAMADA, Y. FUCHIMOTO, R.
GLASER, R. MONROY, Y. XU, M. AWWAD, R. B. COLVIN, A. B. COSIMI, S. C.
ROBSON, J. FISHMAN, T. R. SPITZER, D. K. C: COOPER u. D. H. SACHS (1999):
Porcine Kidney and heart transplantation in baboons undergoing a tolerance
induction regimen and antibody adsorption
Transplantation 67, S. 18-30
KRAKOWKA, S., J. A. ELLIS, F. McNEILLY, S. RINGLER, D. M. RINGS u. G.
ALLAN (2001):
Activation of the immune system is the pivotal event in the production of wasting
disease in pigs infected with porcine circovirus-2 (PCV-2)
Vet Pathol 38, S. 31-42
LA VECCHIO, J. A., A. D. DUNNE u. A. S. B. EDGE (1995):
Enzymatic removal of alpha-galactosyl epitopes from porcine endothelial cells
diminishes the cytotoxic effect of natural antibodies
Transplantation 60, S. 841-847
LAI, L., D. KOLBER-SIMONDS, K.-W. PARK, H.-T. CHEONG, J. L. GREENSTEIN,
G.-S. IM, M. SAMUEL, A. BONK, A. RIEKE, B. N. DAY, C. N. MURPHY, D. B.
CARTER, R. J. HAWLEY u. R. S. PRATHER (2002):
Production of α-1,3-galactosyltransferase knockout pigs by nuclear transfer cloning
Science 295, S. 1089-1092

11 Literaturverzeichnis 104
LANGFORD, G. A., D. GALBRAITH, A. J. WHITTAM, P. McEWAN, X. M.
FERNÀNDEZ-SUÀREZ, J. BLACK, A. SHEPERD u. D. ONIONS (2001):
In vivo analysis of porcine endogenous retrovirus expression in transgenic pigs
Transplantation 72, S. 1996-2000
LE TISSIER, P., J. P. STOYE, Y. TAKEUCHI, C. PATIENCE u. R. A. WEISS (1997) :
Two sets of human-tropic pig retrovirus
Nature 389, S. 681-682
LEVENTHAL, J. R., A. P. DALMASSO, J. W. CROMWELL, C. J. MANIVEL, R. M.
BOLMAN III u. A. J. MATAS (1993):
Complement depletion prolongs discordant cardiac xenograft survival in rodents and
non-human primates
Transplant Proc 25, S. 398-399
LEVENTHAL, J. R., A. P. DALMASSO, J. W. CROMWELL, J. L. PLATT, C. J.
MANIVEL, R.M. BOLMAN III u. A. J. MATAS (1993):
Prolongation of cardiac xenograft survival by depletion of complement
Transplantation 55, S. 857-866
LEVY, M. F., J. CRIPPIN, S. SUTTON, G. NETTO, J. McCORMACK, T. CURIEL, R.
M. GOLDSTEIN, J. T. NEWMAN, T. A. GONWA, J. BANCHEREAU, L. E. DIAMOND,
G. BYRNE, J. LOGAN u. G. B. KLINTMAN (2000):
Liver allotranplantation after extracorporeal hepatic support with transgenic
(hCD55/hCD59) porcine livers
Transplantation 69, S. 272-280

11 Literaturverzeichnis 105
LIN, S. S., B. C. WEIDNER, G. W. BYRNE, L. E. DIAMOND, J. H. LAWSON, C. W.
HOOPES, L. J. DANIELS, C. W. DAGGETT, W. PARKER, R. C. HARLAND, R. D.
DAVIS, R. R. BOLLINGER, J. S. LOGAN u. J. L. PLATT (1998):
The role of antibodies in acute vascular rejection of pig-to-baboon cardiac transplants
J Clin Invest 101, S. 1745-1756
LOSS, M., B. VANGEROW, J. SCHMIDTKO, R. KUNZ, A. JALALI, H. ARENDS, M.
PRZEMECK, H. RÜCKHOLT, M. LEUWER, F. J. KAUP, S. RENSING, E. COZZI, D.
J. G. WHITE, J. KLEMPNAUER u. M. WINKLER (2000):
Acute vascular rejection is associated with systemic complement activation in a pig-
to-primate kidney xenograft model
Xenotransplantation 7, S. 186-196
LOVELAND, B. E., J. MILLAND, P. KYRIAKOU, B. R. THORLEY, D.
CHRISTIANSEN, M. B. LANTERI, M. VAN REGENSBURG, M. DUFFIELD, A. J.
FRENCH, L. WILLIAMS, L. BAKER, M. R. BRANDON, P-X. XING, D. KAHN u. I. F.
C. MCKENZIE (2004):
Characterization of a CD46 transgenic pig and protection of transgenic kidneys
against hyperacute rejection in non-immunosuppressed baboons
Xenotransplantation 11, S.171-183
LUO, Y., G. LEVY, B. M. GARCIA, H. YANG, J. PHILLIPS, L. NOBLE, S.
CHAKRABARTI, D. GRANT u. R. ZHONG (2003):
Ex vivo and extracorporeal perfusion with hDAF pig kidneys
Xenotransplantation 10, S. 410-421
MAHER, S. E., D. L. PHLUG, N. J. NIELS, M. F. ROTHSCHILD u. A. L. M.
BOTHWELL (1998):
Structure/function characterization of porcine CD59: Expression, chromosomal
mapping, complement inhibition, and costimulatory activity
Transplantation 66 (8), S. 1094-1100

11 Literaturverzeichnis 106
MANEZ, R., F. CRESPO, E. GONZALEZ, A. CENTENO, A. JUFFE, F. ARNAL,
E.COZZI, D. J. G. WHITE, R. DUTHALER, W. KINZY, R. OEHRLEIN u. A.
KATOPODIS (2000):
Neutralization of anti-αgalactosyl antibodies without immunosuppression prevents
hyperacute rejection but not acute vascular rejection of pig organs transplanted into
baboons
Transplant Proc 32, S. 888-889
MARTIN, U., V. KIESSIG, J. H. BLUSCH, A. HAVERICH, K. VON DER HELM, T.
HERDEN u. G. STEINHOFF (1998):
Expression of pig endogenous retrovirus by primary porcine endothelial cells and
infection of human cells
Lancet 352, S. 692-694
MASSOUD, M., J. ATTAL, D. THEPOT, H. POINTU, M. G. STINNAKRE, M. C.
THERON, C. LOPEZ u. L. M. HOUDEBINE (1996):
The deleterious effects of human erythropoetin gene driven by the rabbit whey acidic
protein gene Promotor in transgenic rabbits
Reprod Nutr Dev 36, S. 555-563
McCREATH, K. J., J. HOWCROFT, K. H. S. CAMPBELL, A. COLMAN, A. E.
SCHNIEKE u. A. J. KIND (2000):
Production of gene-targeted sheep bynuclear transfer from cultured somatic cells
Nature 405, S. 1066-1069
MENGEL, M., M. WERNER u. R. VON WASIELEWSKI (1999):
Concentration dependent and adverse effects in immunohistochemitry using the
tyramine amplification technique
Biochem J 31, S. 195-200

11 Literaturverzeichnis 107
MERCIER, D., B. CHARREAU, A. WIERINCKX, R. KEIJSER, L. ADRIAENSENS, R.
VAN DEN BERG u. D. H. JOZIASSE (2002):
Regulation of α1,3galactosyltransferase expression in pig endothelial cells
Eur J Biochem 269, S. 1464-1473
MERI, S., B. P. MORGAN, A. DAVIES, R. H. DANIELS, M. G. OLAVESEN u. H.
WALDMANN (1990):
Human protectin (CD59), an 18,000-20,000 MW complement lysis restricting factor,
inhibits C5b-8 catalysed insertion of C9 into lipid bilayers (1990):
Immunology 71, S. 1-9
MERI, S., H. WALDMANN u. P. J. LACHMANN (1991):
Distribution of Protectin (CD59), a complement membrane attack inhibitor, in normal
human tissues
Lab Invest 65, S. 532-537
NIEMANN, H., E. VERHOEYEN, K. WONIGEIT, R. LORENZ, J. HECKER, R.
SCHWINZER, H. HAUSER, W. A. KUES, R. HALTER, E. LEMME, D. HERRMANN,
D. WIRTH u. D. PAUL (2001):
Cytomegalovirus early Promotor induced expression of hCD59 in porcine organs
provides protection against hyperacute rejection
Transplantation 72, S. 1898-1906
NIEMANN, H., R. HALTER, J. W. CARNWATH, D. HERMANN, E. LEMME u. D.
PAUL (1999):
Expression of human blood clotting factor VIII in the mammary gland of transgenic
sheep
Transgenic Res 8, S. 237-247

11 Literaturverzeichnis 108
NIEMANN, H., u. D. RATH (2001):
Progress in reproductive biotechnology in swine
Theriogenology 56, S. 1291-1304
NIEMANN, H., u. W. A. KUES (2000):
Transgenic livestock: premises and promises
Anim Reprod Sci 60-61, S. 277-293
NORIN, A. J., R. J. BREWER, N. LAWSON, G. A. GRIJALVA, M. VAYNBLATT, W.
BURTON, S. P. SQUINTO, S. L. KAMHOLZ u. W. L. FODOR (1996):
Enhanced survival of porcine endothelial cells and lung xenografts expressing human
CD59
Transplant Proc 28, S. 797-798
OGLE, B. M., K. A. BUTTERS, T. B. PLUMMER, K. R. RING, B. E. KNUDSEN, M. R.
LITZOW, M. CASCALHO u. J. L. PLATT (2004):
Spontanous fusion of cells between species yields transdifferation and retroviral
transfer in vivo
FASEB J 18, S. 548-550
ONIONS, D., D. K. C. COOPER, T. J. L. ALEXANDER, C. BROWN, E. CLAASSEN,
J. E. FOWERAKER, D. L. HARRIS, B. W. J. MAHY, P. D. MINOR, A. D. M. E.
OSTERHAUS, P.-P. PASTORET u. K. YAMANOUCHI (2000):
An approach to the control of disease transmission in pig-to-human
xenotransplantation
Xenotransplantation 7, S. 143-155
ONISHI, A., M. IWAMOTO, T. AKITA, S. MIKAWA, K. TAKEDA, T. AWATA, H.
HANADA u. A. C. F. PERRY (2000):
Pig cloning by microinjection of fetal fibroblast nuclei
Science 289, S. 1188-1190

11 Literaturverzeichnis 109
OSMAN, N., I. F. C. McKENZIE, K. OSTENRIED, Y. A. IOANNOU, R. J. DESNICK u.
M. S. SANDRIN (1997):
Combined transgenic expression of α-galactosidase and α1,2-fucosyltransferase
leads to optimal reduction in the major xenoepitope Galα(1,3)Gal
Proc Natl Acad Sci USA 94, S. 14677-14682
PARK, K. W., H.-T. CHEONG, L. LAI, G.-S. IM, B. KÜHNHOLZER, A. BONK, M.
SAMUEL, A. RIEKE, B. N. DAY, C. N. MURPHY, D. B.CARTER u. R. S. PRATHER
(2001):
Production of nuclear transfer-derived swine that express the enhanced gree
flourescent protein
Anim Biotechnol 12, S. 173-181
PARKER, W., D. BRUNO, Z. E. HOLZKNECHT u. J. L. PLATT (1994):
Characterization and affinity isolation of xenoreactive human natural antibodies
J Immunol 153, S. 3791-3803
PHELPS, C. J., C. KOIKE, T. D. VAUGHT, J. BOONE, K. D. WELLS, S.-H. CHEN, S.
BALL, S. M. SPECHT, I. A. POLEJAEVA, J. A. MONAHAN, P. M. JOBST, S. B.
SHARMA, A. E. LAMBORN, A. S. GARST, M. MOORE, A. J. DEMETRIS, W. A.
RUDERT, R. BOTTINO, S. BERTERA, M. TRUCCO, T. E. STARZL, Y. DAI u. D. L.
AYARES (2003):
Production of α1,3-Galactosyltransferase-deficient pigs
Science 299, S. 411-414
PIEDRAHITA, J. A. u. B. MIR (2004):
Cloning and transgenesis in mammals: Implications for xenotransplantation
Am J Transplant 4, S. 43-50

11 Literaturverzeichnis 110
PLATT, J. L., R. J. FISCHEL, A. J. MATAS, S. A. REIF, R. M. BOLMAN u. F. H.
BACH (1991):
Immunopathology of hyperacute xenograft rejection in a swine-to-primate model
Transplantation 52, S. 214-220
PLATT, J. L., S. S. LIN u. C. G. A. McGREGOR (1998):
Acute vascular rejection
Xenotransplantation 5, S. 169-175
PRUITT, S. K., A. D. KIRK, R. R. BOLLINGER, H. C. MARSH JR., B. H. COLLINS, J.
L. LEVIN, J. R. MAULT, J. S. HEINLE, S. IBRAHIM, A. R. RUDOLPH, W. M.
BALDWIN III u. F. SANFILIPPO (1994):
The effect of soluble complement receptor type 1 on hyperacute rejection of porcine
xenografts
Transplantation 57, S. 363-370
PRUITT, S. K., R. R. BOLLINGER, B. H. COLLINS, H. C. MARSH, Jr., J. L. LEVIN,
A. R. RUDOLPH, W. M. BALDWIN, III u. F. SANFILIPPO (1997):
Effect of continous complement inhibition using soluble complement receptor type 1
on survival of pig-to-primate cardiac xenografts
Transplantation 63, S. 900-914
QIAN, Y.-M., X. QIN, T. MIWA, X. SUN, J. A. HALPERIN u. W.-C. SONG (2000):
Identification and functional characterization of a new gene encoding the mouse
terminal complement inhibitor CD59
J Immunol 165, S. 2528-2534

11 Literaturverzeichnis 111
RAMSOONDAR, J. J., Z. MACHÁTY, C. COSTA, B. L. WILLIAMS, W. L. FODOR u.
K. R. BONDIOLI (2003):
Production of α1,3-Galactosyltransferase-knockout cloned pigs expressing human
α1,2-Fucosyltransferase
Biol Reprod 69, S. 437-445
REEMTSMA, K., B. H. McCRACKEN u. J. U. SCHLEGEL (1964):
Renal heterotransplantation in man
Ann Surg 160, S. 384-410
ROLLINS, S. A. u. P. J. SIMS (1990):
The complement-inhibitory activity of CD59 resides in its capacity to block
incorporation of C9 into membrane C5b-9
J Immunol 144, S. 3478-3483
ROSE, A. G. u. D. K. C. COOPER (2000):
Venular thrombosis is the key event in the pathogenesis of antibody-mediated
cardiac rejection
Xenotransplantation 7, S. 31-41
ROSE, A. G., D. K. C. COOPER, P. A. HUMAN, H. REICHENSPURNER u. B.
REICHART (1991):
Histopathology of hyperacute rejection of the heart: Experimental and clinical
observations in allografts and xenografts
J Heart Lung Transplant 10, S. 223-234

11 Literaturverzeichnis 112
SABLINSKI, T., P. R. GIANELLO, M. BAILIN, K. S. BERGEN, D. W. EMERY, J. A.
FISHMAN, A. FOLEY, T. HATCH, R. J. HAWLEY, T. KOZLOWSKI, T. LORF, S.
MEEHAN, R. MONROY, J. A. POWELSON, R. B. COLVIN, A. B. COSIMI u. D. H.
SACHS (1997):
Pig to monkey bone marrow and kidney xenotransplantation
Surgery 121, S. 381-391
SANDRIN, M. S., H. A. VAUGHAN, P. L. DABKOWSKI u. F. C. MCKENZIE (1993):
Anti-pig IgM antibodies in human serum react predominantly with Gal(α1-3)Gal
epitopes
Proc Natl Acad Sci USA 90, S. 11391-11395
SCHMOEKEL, M., F. N. K. BHATTI, A. ZAIDI, E. COZZI, P. D. WATERWORTH, M.
J. TOLAN, M. GODDARD, R. G. WARNER, G. A. LANGFORD, J. J. DUNNING, J.
WALLWORK u. D. J. G. WHITE (1998):
Orthotopic heart transplantation in a transgenic pig-to-primate model
Transplantation 65, S. 1570-1577
SCHUURMAN, H.-J., G. PINO-CHAVEZ, M. J. PHILLIPS, L. THOMAS, D. J. G.
WHITE u. E. COZZI (2002):
Incidence of hyperacute rejection in pig-to-primate transplantation using organs from
hDAF-transgenic donors
Transplantation 73, S. 1146-1151
SCHUURMAN, H.-J., J. CHENG u. T. LAM (2003):
Pathology of xenograft rejection: a commentary
Xenotransplantation 10, S. 293-299

11 Literaturverzeichnis 113
SEGALÉS, J., C. ROSELL u. M. DOMINGO (2004):
Pathological findings associated with naturally acquired porcine circovirus type 2
associated disease
Vet Microbiol 98, S. 137-149
SHAMAY, A., V. G. PURSEL, E. WILKINSON, R. J. WALL u. L. HENNIGHAUSEN
(1992):
Expression of the whey acid protein in transgenic pigs impairs mammary
development
Transgenic Res 1, S. 124-132
SHARMA, A., J. OKABE, P. BIRCH, S. B. McCLELLAN, M. J. MARTIN, J. L. PLATT
u. J. S. LOGAN (1996):
Reduction in the level of Gal(α1,3)Gal in transgenic mice and pigs by the expression
of an α(1,2)fucosyltransferase
Proc Natl Acad Sci USA 93, S. 7190-7195
SHIMIZU, A., S. M. MEEHAN; T. KOZLOWSKI, T. SABLINSKI, F. L. IERINO, D. K.
C. COOPER, D. H. SACHS u. R. B. COLVIN (2000):
Acute humoral xenograft rejection: Destruction of the microvascular capillary
endothelium in pig-to-nonhuman primate renal grafts
Lab Invest 80, S. 815-830
SIMON, P. M., F. A. NEETHLING, S. TANIGUCHI, P. L. GOODE, D. ZOPF, W. H.
HANCOCK u. D. K. C. COOPER (1998):
Intravenous infusion of Galα1-3Gal oligosaccharides in baboons delays hyperacute
rejection of porcine heart xenografts
Transplantation 65, S. 346-353

11 Literaturverzeichnis 114
SMITH, G. P. u. R. A. G. SMITH (2001):
Membrane-targeted complement inhibitors
Mol Immunol 38, S. 249-255
SOMERVILLE, C. A., A. G. KYRIAZIS, A. McKENZIE, J. ALLISON, M. J. PEARSE u.
A. J. F. D’APICE (1994) :
Functional expression of human CD59 in transgenic mice
Transplantation 58, S. 1430-1435
SPECKE, V., S. RUBANT u. J. DENNER (2001):
Productive infection of human primary cells and cell lines with porcine endogenous
retroviruses
Virology 285, S. 177-180
STONE, K. R., G. AYALA, J. GOLDSTEIN, R. HURST, A. WALGENBACH u. U.
GALILI (1998):
Porcine cartilage transplants in the cynomolgus monkey
Transplantation 65, S. 1577-1583
TANAKA, M., D.LATINNE, P. GIANELLO, T. SABLINSKI, T. LORF, M. BAILIN, V.
NICKELEIT, R. COLVIN, E. LEBOWITZ, M. SYKES, A. B. COSIMI u. D. H. SACHS
(1994):
Xenotransplantation from pig to cynomolgus monkey: The potential for overcoming
xenograft rejection through induction of chimerism
Transplant Proc 26, S. 1326-1327
TANIGUCHI, S. u. D. K. C. COOPER (1997)
Clinical xenotransplantation – past, present and future
Ann R Coll Surg Engl 79, S. 13-19

11 Literaturverzeichnis 115
TANIGUCHI, S., F. A. NEETHLING, E. Y. KORCHAGINA, N. BOVIN, Y. YE, T.
KOBAYASHI, M. NIEKRASZ, S. LI, E. KOREN, R. ORIOL u. D. K. C. COOPER
(1996):
In vivo immunoadsorption of antipig antibodies in baboons using a specific Galα1-
3Gal column
Transplantation 62, S. 1379-1384
TEARLE, R. G., M. J. TANGE, Z. L. ZANNETTINO, M. KATERELOS, T. A.
SHINKEL, B. J. W. VAN DENDEREN, A. J. LONIE, I. LYONS, M. B. NOTTLE, T.
COX, C. BECKER, A. M. PEURA, P. L. WIGLEY, R. J. CRAWFORD, A. J. ROBINS,
M. J. PEARSE u. A. J. F. D’APICE (1996):
The α-1,3-Galactosyltransferase knockout mouse
Transplantation 61, S. 13-19
TYCKO, B. (1997):
DNA methylation in genomic imprinting
Mutat Res 386, S. 103-105
VAN DEN BERG, C. W., J. M. PÈREZ DE LA LASTRA, D. LLANES u. B. P.
MORGAN (1997) :
Purification and characterization of the pig analogue of human membrane cofactor
protein (CD46/MCP)
J Immunol 158, S. 1703-1709
VAN DEN BERG, C. W., u. B. P. MORGAN (1994):
Complement-inhibiting activities of human CD59 and analogues from rat, sheep and
pig are not homologously restricted
J Immunol 152, S. 4095-4101

11 Literaturverzeichnis 116
VAN REENEN, C. G., T. H. E. MEUWISSEN, H. HOPSTER, K. OLDENBROEK, T.
A. M. KRUIP u. H. J. BLOKHUIS (2001):
Transgenesis may affect farm animal welfare: A case for systemic risk assessment
J Anim Sci 79, S. 1763-1779
VON WASIELEWSKI, R., M. MENGEL, B. WIESE, T. RUDIGER, H. K. MULLER-
HERMELINK u. H. KREIPE (2002):
Tissue array technology for testing interlaboratory and interobserver reproducibility of
immunohistochemical estrogen receptor analysis in a large multicenter trial
Am J Clin Pathol 118, S. 675-682
WATERWORTH, P. D., J. DUNNING, M. TOLAN, E. COZZI, G. LANGFORD, G.
CHAVEZ, D. WHITE u. J. WALLWORK (1998):
Life-supporting pig-to-baboon heart xenotransplantation
J Heart Lung Transplant 17, S. 1201-1207
WATIER, H., J.-M. GUILLAUMIN, F. PILLER, M. LACORD, G. THIBAULT, Y.
LEBRANCHU, M. MONSIGNY u. P. BARDOS (1996) :
Removal of terminal α-Galactosyl residues from xenogeneic porcine endothelial cells
Transplantation 62, S. 105-113
WILSON, C. A., S. WONG, J. MULLER, C. E. DAVIDSON, T. M. ROSE u. P. BURD
(1998):
Type C retrovirus released from porcine primary peripheral blood mononuclear cells
infects human cells
J Virol 72, S. 3082-3087

11 Literaturverzeichnis 117
XU, Y., T. LORF, T. SABLINSKI, P. GIANELLO, M. BAILIN, R. MONROY, T.
KOZLOWSKI, M. AWWAD, D. K. C. COOPER u. D. H. SACHS (1998):
Removal of anti-porcine natural antibodies from human and nonhuman primate
plasma in vitro and in vivo by a Galα1-3Galβ1-4βGlc-X immunoaffinity column
Transplantation 65, S. 172-179
YOO, D., u. A. GIULIVI (2000):
Xenotranplantation and the potential risk of xenogeneic transmission of porcine
viruses
Can J Vet Res 64, S. 193-203
YOUNG, L. E., K. D. SINCLAIR u. I. WILMUT (1998):
Large offspring syndrome in cattle and sheep
Rev Reprod 3, S. 155-163
ZAIDI, A., M. SCHMOECKEL, F. BHATTI, P. WATERWORTH, M. TOLAN, E. COZZI,
G. CHAVEZ, G. LANGFORD, S. THIRU, J. WALLWORK, D. WHITE u. P. FRIEND
(1998):
Life-supporting pig-to-primate renal xenotransplantation using genetically modified
donors
Transplantation 65, S. 1584-1590
ZHOU, C.-Y., E. MCINNES, N. PARSON, G. LANGFORD, R. LANCASTER, A.
RICHARDS, G. PINO-CHAVEZ, G. DOS SANTOS CRUZ, L. COPEMAN, C.
CARRINGTON u. S. THOMPSON (2002) :
Production and characterization of a pig line transgenic for human membrane
cofactor protein
Xenotransplantation 9, S. 183-190

11 Literaturverzeichnis 118
ZHU, A. (2000):
Binding of human natural antibodies to non-α-gal xenoantigens on porcine
erythrocytes
Transplantation 69, S. 2422-2428

119
12 Anhang 9.1 Tris-Puffer Stammlösung:
121.14 g Trishydroxymethyl-aminomethan (Fa. Merck, Darmstadt)
in 1700 ml entionisiertem Wasser lösen
pH 7.6 einstellen
9 ml Tween20 hinzufügen
auf 2000 ml mit entionisiertem Wasser auffüllen
Gebrauchslösung:
350g NaCl pro analysis (Fa. Merck, Darmstadt)
+ 16 l entionisiertem Wasser
+ 2 l Tris-Stammlösung
auf 20 l auffüllen
9.2 Tris/HCl-Puffer 8.16 g NaCl pro analysis (Fa. Merck, Darmstadt)
+ 12.1 g Trishydroxymethyl-aminomethan (Fa. Merck, Darmstadt)
+ 700 ml Aqua dest.
pH 7.5 einstellen mit 37%iger HCl
auf 1000 ml auffüllen
9.3 Digestion solution 100 g Guanidin Thiocyanat (Fa. Fluka, Seelze)
+ 6 ml 1M Tris/HCl (pH 7.6)
+ 4.2 ml 97%iges Natrium-N-Lauryl-Sarcosine
auf 200 ml mit H2O/DEPC auffüllen

12 Anhang 120
9.4 3M Na-Acetat pH 5,2 24.6 g Na-Acetat in 80 ml H2O/DEPC lösen
pH 5.2 mit konzentrierter Essigsäure einstellen
mit H2O/DEPC auf 100 ml auffüllen
9.5 3M Na-Acetat pH7 /100 µg/ml Dextran 50 ml 3M Na-Acetat pH7 (Fa. Sigma, Seelze)
+ 25 µl 20%iges Dextran T500000
9.6 Proteinase K-Puffer 25 ml Trishydroxymethyl-aminomethan (Fa. Merck, Darmstadt) pH 8.1 100 mM
+ 24.65 ml H2O
+ 100 µl 0.5 M EDTA
+ 250 µl Tween20
9.7 1xTE-Puffer 270 g Trishydroxymethyl-aminomethan (Fa. Merck, Darmstadt)
+ 137 g Borsäure
+ 100 ml 0.5 M EDTA pH 8
auf 20 l mit entionisiertem Wasser auffüllen
9.8 6%iges PAA-Gel 6%ige PAA-Stammlösung:
78 ml 38% Acrylamid-Stocksolution (Rotiphorese-Gel 40, Fa. Roth, Karlsruhe)
+ 50 ml 10xTBE für Polyacrylamid-Gele
+ 371 ml entionisiertes Wasser

12 Anhang 121
Gel:
30 ml 6%ige PAA-Stammlösung
+ 375 µl 10%iges Ammoniumpersulfat (Fa. AppliChem, Darmstadt)
+ 15 µl Tetramethylethylendiamin (Fa. Fluka, Seelze)
15 Minuten polymerisieren lassen
Füllung der Gelkammer mit 10xTBE-Puffer für Acrylamidgele
9.9 10xTBE-Puffer für Acrylamidgele 54 g Tris
+ 27.5 g Borsäure
+ 20 ml 0.5M EDTA pH8
mit destiliertem Wasser auf 500 ml auffüllen
9.10 BSA/Tris-Puffer 12.1 g Tris
+ 35 g NaCl
+ 9 ml Tween
+ 20 g Bovines Serumalbumin (Fa. Biomol, Hamburg)
mit entionisiertem Wasser auf 2 l auffüllen
pH 7.6 einstellen

12 Anhang 122
9.11 Ausschnitt aus der hCD59/NTA-Konstruktsequenz CTCGAGTTTA CCACTCCCTA TCAGTGATAG AGAAAATGGA AAGTCGAGTT TACCACTCCC 60
TATCAGTGAT AGAGAAAAGT GAAAGTCGAG TTTACCACTC CCTATCAGTG ATAGAGAAAA 120
GTGAAAGTCG AGTTTACCAC TCCCTATCAG TGATAGAGAA AAGTGAAAGT CGAGTTTACC 180
ACTCCCTATC AGTGATAGAG AAAAGTGAAA GTCGAGTTTA CCACTCCCTA TCAGTGATAG 240
AGAAAAGTGA AAGTCGAGTT TACCACTCCC TATCAGTGAT AGAGAAAAGT GAAAGTCGAG 300
CTCGGTACCC GGGTCGAGTA GGCGTGTACG GTGGGAGGCC TATATAAGCA GAGCTCGTTT 360
AGTGAACCGT CAGATCGCCT GGAGACGCCA TCCACGCTGT TTTGACCTCC ATAGAAGACA 420
CCGGGACCGA TCCAGCCTCC GCGGCCCCGA ATTCAGGTTC TGTGGACAAT CACAATGGGA 480
ATCCAAGGAG GGTCTGTCCT GTTCGGGCTG CTGCTCGTCC TGGCTGTCTT CTGCCATTCA 540
GGTCATAGCC TGCAGTGCTA CAACTGTCCT AACCCAACTG CTGACTGCAA AACAGCCGTC 600
AATTGTTCAT CTGATTTTGA TGCGTGTCTC ATTACCAAAG CTGGGTTACA AGTGTATAAC 660 hCD59
AAGTGTTGGA AGTTTGAGCA TTGCAATTTC AACGACGTCA CAACCCGCTT GAGGGAAAAT 720
GAGCTAACGT ACTACTGCTG CAAGAAGGAC CTGTGTAACT TTAACGAACA GCTTGAAAAT 780
GGTGGGACAT CCTTATCAGA GAAAACAGTT CTTCTGCTGG TGACTCCATT TCTGGCAGCA 840
GCCTGGAGCC TTCATCCCTA AGTCAACACC AGGAGAGCTT CTCCCAAACT CCCCGTTCCT 900
GCGTAGTCCG CTTTCTCTTG CTGCCACATT CTAAAGGCTT GATATTTTCC AAGCTTTTAA 960
AACAGCTCTG GGGTTGTACC CACCCCAGAG GCCCACGTGG CGGCTAGTAC TCCGGTATTG 1020
CGGTACCCTT GTACGCCTGT TTTATACTCC CTTCCCGTAA CTTAGACGCA CAAAACCAAG 1080
TTCAATAGAA GGGGGTACAA ACCAGTACCA CCACGAACAA GCACTTCTGT TTCCCCGGTG 1140
ATGTCGTATA GACTGCTTGC GTGGTTGAAA GCGACGGATC CGTTATCCGC TTATGTACTT 1200
CGAGAAGCCC AGTACCACCT CGGAATCTTC GATGCGTTGC GCTCAGCACT CAACCCCAGA 1260 IRES-Sequenz
GTGTAGCTTA GGCTGATGAG TCTGGACATC CCTCACCGGT GACGGTGGTC CAGGCTGCGT 1320
TGGCGGCCTA CCTATGGCTA ACGCCATGGG ACGCTAGTTG TGAACAAGGT GTGAAGAGCC 1380
TATTGAGCTA CATAAGAATC CTCCGGCCCC TGAATGCGGC TAATCCCAAC CTCGGAGCAG 1440
GTGGTCACAA ACCAGTGATT GGCCTGTCGT AACGCGCAAG TCCGTGGCGG AACCGACTAC 1500
TTTGGGTGTC CGTGTTTCCT TTTATTTTAT TGTGGCTGCT TATGGTGACA ATCACAGATT 1560
GTTATCATAA AGCGAATTGG ATTGCGGCCG CATGTCTAGA TTAGATAAAA GTAAAGTGAT 1620
TAACAGCGCA TTAGAGCTGC TTAATGAGGT CGGAATCGAA GGTTTAACAA CCCGTAAACT 1680
CGCCCAGAAG CTAGGTGTAG AGCAGCCTAC ATTGTATTGG CATGTAAAAA ATAAGCGGGC 1740
TTTGCTCGAC GCCTTAGCCA TTGAGATGTT AGATAGGCAC CATACTCACT TTTGCCCTTT 1800
AGAAGGGGAA AGCTGGCAAG ATTTTTTACG TAATAACGCT AAAAGTTTTA GATGTGCTTT 1860
ACTAAGTCAT CGCGATGGAG CAAAAGTACA TTTAGGTACA CGGCCTACAG AAAAACAGTA 1920 Transaktivator-Gen
TGAAACTCTC GAAAATCAAT TAGCCTTTTT ATGCCAACAA GGTTTTTCAC TAGAGAATGC 1980
ATTATATGCA CTCAGCGCTG TGGGGCATTT TACTTTAGGT TGCGTATTGG AAGATCAAGA 2040
GCATCAAGTC GCTAAAGAAG AAAGGGAAAC ACCTACTACT GATAGTATGC CGCCATTATT 2100
ACGACAAGCT ATCGAATTAT TTGATCACCA AGGTGCAGAG CCAGCCTTCT TATTCGGCCT 2160
TGAATTGATC ATATGCGGAT TAGAAAAACA ACTTAAATGT GAAAGTGGGT CCGCGTACAG 2220
CCGCGCGCGT ACGAAAAACA ATTACGGGTC TACCATCGAG GGCCTGCTCG ATCTCCCGGA 2280
CGACGACGCC CCCGAAGAGG CGGGGCTGGC GGCTCCGCGC CTGTCCTTTC TCCCCGCGGG 2340
ACACACGCGC AGACTGTCGA CGGCCCCCCC GACCGATGTC AGCCTGGGGG ACGAGCTCCA 2400
CTTAGACGGC GAGGACGTGG CGATGGCGCA TGCCGACGCG CTAGACGATT TCGATCTGGA 2460 Poly-A
CATGTTGGGG GACGGGGATT CCCCGGGTCC GGGATTTACC CCCCACGACT CCGCCCCCTA 2520
…

123
14 Danksagung
Zunächst möchte ich mich bei Prof. Dr. Hans Kreipe für die Bereitstellung des
interessanten Themas bedanken sowie für die fachliche Unterstützung und die
hilfreichen Anregungen.
Ich danke auch den anderen Mitgliedern meiner Betreuungsgruppe, Prof. Dr. Achim
Gruber, Prof. Dr. Edda Töpfer-Petersen und Prof. Dr. Heiner Niemann für ihr
Interesse an meinem Projekt und die anregenden Diskussionen in netter
Atmosphäre.
Bei Prof. Dr. Achim Gruber möchte ich mich außerdem für seine tiermedizinisch-
fachliche Unterstützung bedanken sowie für die Möglichkeit, in seiner Arbeitsgruppe
mitzuarbeiten.
Ich bedanke mich auch bei Prof. Dr. Oliver Bock für die geduldige Hilfe und fachliche
Beratung bei der PCR und für die Unterstützung im Endspurt dieser These.
Dr. Michael Mengel danke ich für die Hilfe bei der Etablierung der
Immunhistochemie.
Christina Koop, Sanja Galgoci und Martina Erbs möchte ich für die geduldige und
unermüdliche Unterstützung im Labor und die nette Arbeitsatmosphäre danken.
Ich bedanke mich bei meinen Mädels für eine tolle Zeit im Studium, die mir
unvergesslich bleiben wird.
Ich danke meiner Familie für die jahrelange liebevolle Unterstützung und Ermutigung
und das Verständnis bei allen Herausforderungen auf meinem bisherigen Weg.
Und last but not least bedanke ich mich bei Olli für seine große Geduld und
Hilfsbereitschaft und dafür, dass er immer da ist um mich wieder aufzumuntern.







