enzymatische charakterisierung der alkoholdehydrogenase ... · 2.5 die ph‐abhängigkeit der...
TRANSCRIPT

3
- 1 -
Enzymatische Charakterisierung der Alkoholdehydrogenase aus Saccharomyces cerevisiae
Praktikumsvorschrift für
Schülerinnen und Schüler
Name: __________________
Dr. F. Schaller Waldring 71
44789 Bochum T.: 0234/9304411
E-Mail: [email protected] Homepage: www.schiller-schule.de

3
- 2 -
Einleitung
1.1 Allgemeine Hinweise und Ablaufsplan des Versuchstages
1.2 Gruppeneinteilung und Zuordnung zu dem jeweiligen Experiment
II Methoden
2.1 Übung zum Gebrauch von Mikroliterpipetten
2.2 Vorarbeiten und theoretischer Hintergrund ‐ Nachweis von Fleischsorten mittels
molekularbiologischer Methoden, aber wie?
2.3 DNA‐Isolierung aus Fleisch‐ und Dönerproben
2.3.1 Hintergrundinformation zum Thema: Zenntrifugation
2.4 Die Polymerase‐Kettenreaktion (PCR) – theoretischer Hintergrund
2.5 Die Polymerase‐Kettenreaktion (PCR) – praktischer Teil
2.6 Agarose‐Gelelektrophorese
III Ergebnisse
IV Anhang
4.1 Aufgabenstellungen
4.2 Materialien
4.2.1 DNA‐Präparation
4.2.2 Polymerase‐Kettenreaktion
4.2.3 Agarose‐Gelelektrophorese
4.3 Exemplarische Ergebnisse

3
- 3 -

3
- 4 -
Inhaltsverzeichnis
I Einleitung
1.1 Allgemeine Hinweise und Ablaufsplan des Versuchstages S. 2
1.2 Gruppeneinteilung und Zuordnung zu dem jeweiligen Experiment S. 2
1.3 Grundlegendes aus der Chemie ‐ Stoffmenge und Stoffmengenkonzentration S. 3
1.4 Einführung in die Grundlagen der Enzymologie am Beispiel der Alkoholdehydrogenase
aus Bäckerhefe (Saccharomyces cerevisiae) S. 4‐7
II Experimenteller Teil: Versuche und Methoden
2.1 Übung zum Gebrauch von Pipetten, Pipettierhilfen und Mikroliterpipetten S. 7‐8
2.2 Gewinnung des Hefe‐Rohextraktes S. 9
2.3 Proteinbestimmung: Eichreihe und Proteinkonzentration des Hefe‐Rohextraktes und die
Bedienung unserer Photometer und der Stoppuhr zur Zeitmessung S. 10
2.4 Aktivitätsbestimmung der Alkoholdehydrogenase im Heferohextrakt /
Abhängigkeit der Enzymaktivität von der Menge des eingesetzten Rohextrakts S. 11‐13
2.5 Die pH‐Abhängigkeit der Alkoholdehydrogenase‐Aktivität S. 13
2.6 Abhängigkeit der Alkoholdehydrogenase‐Aktivität von der Substratkonzentration S. 14
2.7 Berechnungen für Spezialisten S. 14‐16
III Ergebnisse / Auswertung
3.2 Gewinnung des Hefe‐Rohextraktes S. 17
3.3 Proteinbestimmung: Eichreihe und Proteinkonzentration des Hefe‐Rohextraktes S. 17‐18
3.4 Aktivitätsbestimmung der Alkoholdehydrogenase im Heferohextrakt /
Abhängigkeit der Enzymaktivität von der Menge des eingesetzten Rohextrakts S. 18‐19
3.5 Die pH‐Abhängigkeit der Alkoholdehydrogenase‐Aktivität S. 19‐20
3.6 Abhängigkeit der Alkoholdehydrogenase‐Aktivität von der Substratkonzentration S. 21‐22
IV Anhang 4.1 Aufgabenstellungen S. 24‐26 4.3 Materialien S. 27‐28 4.3.1 Chemikalien und Lösungen 4.3.2 Glaswaren und Verbrauchsmaterialien 4.3.3 Geräte 4.4 Literaturverzeichnis S. 28

3
- 5 -
I Einleitung
1.1 Allgemeine Hinweise und Ablaufsplan des Versuchstages
Ziel des hier durchgeführten Projektes ist der Nachweis und die Quantifizierung von
Enzymaktivitäten am Beispiel der
Alkoholdehydrogenase (ADH) aus Saccharomyces cerevisiae (Bäckerhefe)
über die Berechnung des photometrisch bestimmten Kosubstratverbrauchs.
Schematischer Ablauf des Versuchstages
I Begrüßung, Einleitung in die Thematik, Sicherheitsbelehrung, Gruppeneinteilung
II Lyse der Hefezellen, Vorbesprechung zur Pipettierübung
III Pipettierübung
IV Versuchsbesprechung Teil I: Einführung in die Photometrie am Beispiel der Chlorophyllbestimmung
V Versuche Teil I: Gewinnung des Heferohextraktes, Proteinbestimmung
VI Versuchsbesprechung Teil II: Einführung in die Photometrie (kinetische Analyse) am Beispiel der Katalase
VII Versuche Teil II: Aktivität im Rohextrakt, Abhängigkeit vom pH‐Wert
‐ PAUSE ‐
VIII Versuche Teil III: Abhängigkeit der ADH‐Aktivität von der Substratkonzentration
IX Fertigstellung der Auswertungen, Bearbeitung der Aufgabenstellungen, Aufräumen
X Versuchsbesprechung Teil IV (Auswertung und Diskussion)
1.2 Gruppeneinteilung und Zuordnung zu dem jeweiligen Experiment
Je nach Kursstärke werden unterschiedlich viele Gruppen aus 4 Schülerinnen und Schülern gebildet. Inner‐halb jeder Vierergruppe arbeiten allerdings jeweils zwei SuS zusammen. Die meisten Chemikalien und Ge‐räte sind für je zwei SuS vorhanden. Andere, wie z. B. das Photometer aber auch die 20‐200 µl Pipetten, müssen geteilt werden. Die Auswertungen der Versuche (Ergebnisse) sollen immer im Anschluss an das jeweilige Experiment er‐stellt werden. Diese werden dann innerhalb der nächsten Versuchsvorbesprechung gegenseitig vorgestellt bzw. ausgetauscht und besprochen.

3
- 6 -
1.3 Grundlegendes aus der Chemie ‐ Stoffmenge und Stoffmengenkonzentration
Die Konzentration eines Stoffes wird als Stoffmengenkonzentration in Mol/Liter (molare Konzentration, [M]) angegeben. Die Konzentrationsangabe erfolgt also nicht in Prozent [%], wie z. B. beim Alkohol, was im Weitesten einem Mischungsverhältnis entspricht, sondern als Angabe der Anzahl von Teilchen pro Liter.
Denn: Ein Mol eines Stoffes entspricht einer festgelegten Teilchenanzahl und zwar: 6,022 x 1023 Teilchen. Löse man ein Mol eines Stoffes in Wasser und füllt auf einen Liter auf, so hat man demnach eine Lösung, die eine Stoffmengenkonzentration von 1 Mol/Liter hat und demnach 1 molar (1 M) ist.
Da man die Atome oder Moleküle eines zu lösenden Stoffes nicht abzählen kann – so gut sind unsere Au‐gen nicht ‐, erhält man die Teilchenanzahl, die man für das Ansetzten einer Lösung einer bestimmten Stoffmengenkonzentration braucht, indem man die entsprechende Teilchenanzahl abwiegt. Jedes Atom bzw. Molekül hat ja ein charakteristisches Gewicht (besser: ein charakteristische Masse).
Erinnerung: Diese Masse nennt man Atom‐ bzw. Molekülmasse. Die Einheit hier ist das „u“. So hat das Was‐serstoffatom eine Atommasse von 1u, das Sauerstoffatom eine Atommasse von 16u – das Wassermolekül (H2O) demnach eine Molekülmasse von 18u.
Problem: Einzelne Atome oder Moleküle sind zu leicht, um sie auf normalen Wagen abwiegen zu können (hierfür bräuchte man ein Massenspektrometer). Hat man aber eine große Anzahl der entsprechenden Ato‐me oder Moleküle – zum Beispiel 1 Mol, d. h. 6,022 x 1023 Teilchen, funktioniert das schon, da man hier Massen (Gewichte) erreicht, die auch mit „normalen“ Wagen zu messen sind.
Es hat sich daher der Begriff der Molaren Masse (Mr), der angegeben wird in Gramm/Mol [g/mol], in der Chemie etabliert. Die Molare Masse sagt aus, welche Masse (Gewicht) ein Mol eines Stoffes hat.
Das Schöne: Die Molare Masse gemessen in g/mol entspricht der Atom‐ bzw. Molekülmasse. So wiegen ein Mol Wasserstoffatome 1g, ein Mol Sauerstoffatome 16g und ein Mol Wasser 18g. Die Molaren Mas‐sen von Wasserstoff, Sauerstoff und Wasser sind: 1 g/mol, 16 g/mol und 18 g/mol.
Soll man nun eine Lösung einer bestimmten Konzentration (z. B. eines der hier verwendeten Puffer) her‐stellen, so benötigt man demnach folgende Angaben: 1. Die molare Masse [Mr] des zu lösenden Stoffes 2. Die molare Konzentration [M] der gewünschten Lösung 3. Das gewünschte Volumen [V] der Lösung einer bestimmten Konzentration.
Mit folgender Formel lässt sich dann die abzuwiegende Menge des Stoffes leicht errechnen:
Menge des abzuwiegenden Stoffes [g] = (Molare Masse [g/mol] x Molare Konzentration [mol/Liter]) x Volumen [Liter]
Ein Beispiel: Es sollen 500mL einer 0,15molaren Dikaliumhydrogenphosphat‐Lösung (K2HPO4‐Lsg.) herge‐stellt werden. Mr (K2HPO4) = 174,18 g/mol; M = 0,15 mol/Liter; V = 0,5 Liter Rechnung: (174,18 g/mol x 0,15 mol/L) x 0,5L = 13,06g Vorgehen: Abwiegen der 13,06g K2HPO4, Lösen in ca. 400mL Wasser, Auffüllen auf 500mL

3
- 7 -
1.4 Einführung in die Grundlagen der Enzymologie am Beispiel der Alkoholdehydrogenase aus Bäcker‐ hefe (Saccharomyces cerevisiae) Kenntnisse im Umgang mit Enzymen und deren Anwendungsmöglichkeiten sind schon lange keine Domäne der Biochemie mehr. Auch andere naturwissenschaftliche Disziplinen, wie z. B. Biologie, Lebensmittelche‐mie oder medizinische Diagnostik werden heute wesentlich durch die Enzymatik geprägt. Die vorliegend Versuchsvorschrift soll einen Einblick in einige grundlegende Aspekte dieses Gebietes vermitteln.
Enzyme sind die Katalysatoren der lebenden Zelle (Biokatalysatoren). Als solche können sie durch Verringe‐rung der Aktivierungsenergie (E) die Gleichgewichtseinstellung metastabiler Systeme beschleunigen. So verläuft z. B. die Spaltung von Wasserstoffperoxid nach Gleichung (1)
(1) H2O2 H2O + ½ O2
trotz einer exergonen Änderung der freien Energie (ΔG = ‐96kJ x s‐1), es handelt sich also um eine exergone Reaktion, bei der Energie frei wird, nur äußerst langsam ab, da die Aktivierungsenergie sehr hoch ist. Die Geschwindigkeitskonstant (k) als Maß für die Schnelligkeit der Reaktion liegt bei k = < 10‐5 [mol x L‐1 x s‐1]. In Gegenwart von kolloidalem Platin als Katalysator beträgt die Aktivierungsenergie nur noch 50kJ x mol‐1, und k steigt auf ca. 10‐2. Bei Verwendung eines Enzyms als Biokatalysator (Katalase) fällt die Aktivierungs‐energie sogar auf nur 23kJ x mol‐1, gleichzeitig nimmt k den enormen Wert von ca. 106 an.
Eines können Enzyme jedoch nicht, nämlich Wunder vollbringen, oder physiko‐chemisch ausgedrückt, eine thermodynamisch unmögliche Reaktion bewerkstelligen. Bei ihrer katalytischen Aktivität zeichnen sich En‐zyme durch eine meist bemerkenswerte Substrat‐ und Wirkungsspezifität aus. Ein Enzym kann, im Gegen‐satz zu den meisten Reaktionen der organischen Chemie, auch in einem Substanzgemisch gezielt eine Komponente (das Substrat) identifizieren, binden und in ein ganz bestimmtes Produkt (wirkungsspezifisch) umwandeln.
Die erwähnten Faktoren (hohe katalytische Aktivität, hohe Substrat‐ und Wirkungsspezifität) stellen die Grundlage der enzymatischen Analyse dar. Dabei lassen sich, je nach Problemstellung, zwei prinzipielle Messmethoden unterscheiden:
1. Die Endwert‐Analyse Ziel dieses Verfahrens ist die Bestimmung der Konzentration bestimmter Metabolite in der zu untersuchen‐den Probe. In Gegenwart eines Überschusses an Enzymen (zur Erzielung kurzer Analysezeiten) wird dabei das Substrat quantitativ (vollständig) umgesetzt. Nach Stillstand der Reaktion wird dann der Gesamtumsatz und daraus die enzymatische Aktivität ermittelt. 2. Die kinetische Analyse Bei dieser Methode wird der Gehalt eines Enzyms in einer Probe anhand dessen katalytischer Aktivität be‐stimmt, wobei als Maß die erzielte Umsatzgeschwindigkeit (Stoffumsatz pro Zeiteinheit) ermittelt wird. Neben einer Reihe willkürlicher Einheiten (z. B. ΔE pro min, d. h. der Änderung der Lichtabsorption über die Zeit, entsprechend der Zu‐ bzw. Abnahme der Konzentration gefärbter Substanzen) hatte sich ursprünglich als internationale Maßeinheit der enzymatischen Aktivität die Einheit Unit (U) durchgesetzt, die auf der praxisnahen Definition 1 µmol Stoffumsatz pro Minute beruht. Kleinere Enzymaktivitäten werden in Milli‐Units (mU) angegeben, entsprechend einem Umsatz von 1nmol pro Minute. Seit dem 1.1.1978 ist das SI‐System verbindlich, wobei seither (meistens) als Einheit der katalytischen Aktivität das Katal (kat) gilt, das

3
- 8 -
als ein Umsatz von 1 mol pro Sekunde (mol x s‐1) definiert ist. Für kleinere Reaktionsraten gelten die be‐kannten Präfixe (also mkat, µkat, nkat, pkat). Für die Umrechnung der beiden Einheiten gilt (2): (2) 1U = 16,67 nkat Erinnerung: Sehr kleine und sehr große Bruchteile einer Grundeinheit (z. B. Liter, Meter, Gramm, Katal, Byte) werden durch Präfixe angegeben. Bekannt sind uns aus dem Alltag die Präfixe „Milli“ (s. Milligramm [mg], Millimeter [mm]) und „Mega“ sowie „Giga“ (s. Megabyte[Mb] und Gigabyte [Gb]). In den Naturwissen‐schaften sind v. a. noch kleine Bruchteile als „Milli“ (1/1000stel Gramm oder 1 x 10‐3 Gramm) gebräuchlich. 1/1000stel Milligramm entspricht somit einem Mikrogramm (µg oder 1 x 10‐6 Gramm), 1/1000stel Mikro‐gramm entspricht einem Nanogramm (ng oder 1 x 10‐9 Gramm), 1/1000stel Nanogramm entspricht einem Pikogramm (pg oder 1 x 10‐12 Gramm). Da die Aktivität eines Enzyms von einer Vielzahl von Parametern beeinfluss wird, müssen derartige Be‐stimmungen unter vergleichbaren Bedingungen durchgeführt werden (pH‐Wert, Substratsättigung, Tempe‐ratur, . . .). Ferner darf die eingesetzte Enzymmenge nicht zu hoch sein, da sonst die nur in der Anfangspha‐se der Reaktion bestehende Proportionalität zwischen Umsatzrate und Zeit nicht mehr zu beobachten ist, da das Substrat zu schnell verbraucht wird und damit nicht mehr im Überschuss vorhanden wäre (s. Abbil‐dung 2). Diese Unsicherheit, ob tatsächlich die lineare Anfangsgeschwindigkeit der Reaktion gemessen wird, tritt vor allem bei punktuellen Messungen auf, d. h. wenn die Reaktion nach einer bestimmten Inkubationszeit ab‐gebrochen und die Probe analysiert wird. Diese Zweifel beseitigt eine kontinuierliche Registrierung des Re‐aktionsverlaufs. Als geeignetes Mittel hierzu hat sich die Spektralphotometrie durchgesetzt, der sogenann‐te „optische Test“. Hierbei wird die Konzentrationsänderung einer lichtabsorbierenden Substanz (meist des Substrates oder des Produktes oder wie heute des Cosubstrates) erfasst. Den prinzipiellen Aufbau eines Photometers zeigt Abbildung 1.
Abb. 1 Aufbau und Strahlengang durch den Probenbehälter (Küvette) eines Photometers
Schematische Darstellung der Absorptionsmessungen mit dem Photometer DR 2800. Die Absportion beschreibt, welche
Wellenlängen des eingestrahlten bzw. von der Lichtquelle transmittierten Lichtstrahles eine Substanz aufnimmt. Die am
Detektor gemessene Extinktion bzw. Extinktionsänderung [ΔE] beschreibt die Menge des vom Küvetteninhalt absorbierten
Lichts.

3
- 9 -
Als Messgröße wird hier die Extinktion (E), die der Konzentration der lichtabsorbierenden Substanz linear proportional ist, verwendet. Das Lambert‐Beersche Gesetz gibt den Zusammenhang zwischen Extinktion einer Probe und der Substratkonzentration (c) [mol x L‐1] in der Probe an. Die Extinktion ist bei gegebener Schichtdicke (d) (meist 1cm entsprechend des Durchmessers der Küvette [Probenbehälter im Photometer]) direkt proportional zur Substratkonzentration: (3) E ~ c x d E = Extinktion; c = Substratkonzentration; d = Schichtdicke Der Proportionalitätsfaktor wird molarer Extinktionskoeffizient genannt und stellt anschaulich die Extink‐tion dar, die eine 1 molare Lösung (1mol x L‐1) der Substanz hätte. Der molare Extinktionskoeffizient ist da‐her eine für jede Wellenlänge charakteristische Stoffeigenschaft. Als Symbol wird der griechische Buchsta‐be Epsilon (ε) verwendet. Das Lambert‐Beersche Gesetz lautet vollständig: (4) E = ε x c x d Betrachtet man die Extinktionsdifferenzen pro Zeit, so ändert sich entsprechend die Substratkonzentration pro Zeit. Demnach kann man auch schreiben: (5) ΔE/Δt = ε x d x Δc/Δt, und entsprechend dach Kürzen von Δt: (6) ΔE = ε x Δc x d. Es ist zu beachten (s. o.) dass ΔE/Δt nur aus der linearen Anfangsphase der Reaktion entnommen werden darf (s. Abbildung 2). Zur genaueren Bestimmung wird hierzu eine Tangente angelegt und ein Steigungs‐dreieck eingezeichnet. Die Extinktionsänderung pro Minute kann dann durch Teilen des Extinktionswertes durch die entsprechende Anzahl an Minuten (Δt) ermittelt werden Abbildung 2: Verlauf einer enzymatischen Reaktion über die Zeit

3
- 10 -
In der biochemischen Literatur findet sich häufig die Angabe der spezifischen Aktivität von Enzymen, bei der die Enzymaktivität auf die Proteinmenge bezogen ist. Es gilt: spezifische Aktivität (sp. A.) = Enzymaktivität pro mg Protein (kat/mg bzw. U/mg) Diese Angabe ist vor allem wichtig, wenn man Proteine aus Zellhomogenaten reinigen will oder die spezifi‐sche Aktivität des reinen Proteins angeben will: Je höher die spezifische Aktivität einer Enzymprobe ist, desto weniger ist das betreffende Enzym mit Fremdproteinen verunreinigt. Die Alkoholdehydrogenase (ADH) Die sich anschließenden Versuche werden mit dem Enzym Alkoholdehydrogenase (ADH; Alkohol:NAD+‐Oxidoreduktase, EC 1.1.1.1*) durchgeführt, dass sich leicht aus Trockenhefe (luftgetrockneter Bäckerhefe, Saccharomyces cerevisiae) gewinnen lässt. Es katalysiert die reversible Reduktion von Acetaldehyd nach der Gleichung: (7) Acetaldehyd + NADH + H+ Ethanol + NAD+ Wobei das Reaktionsgleichgewicht weit auf der Seite der Alkoholbildung liegt. In vivo ist die ADH in der He‐fe das letzte Enzym bei der Vergärung von Glucose zu Ethanol (s. Hinreaktion). Im Menschen ist es das ers‐te Enzym, das am Abbau des Trinkalkohols (Ethanol) beteiligt ist (s. Rückreaktion). Nomenklatur von Enzymen Das „Nomenclature Committee“ der „International Union for Biochemistry“ hat ein Klassifikationssystem aufgestellt, in dem Enzyme systematisch entsprechend den Kategorien der von ihnen durchgeführten Re‐aktionen aufgelistet sind. Z. B. EC Enzyme Commisssion 1. Oxidoreduktase 1. Einwirken auf eine Aldehydgruppe 1. mit NAD oder NADP als Elektronenakzeptor 1 Alkoholdehydrogenase 1.5 Grundlegendes zum Aufbau von Proteinen
II Experimenteller Teil: Versuche und Methoden 2.1 Übungen zum Gebrauch von Pipetten, Pipettierhilfen und Mikroliterpipetten Mikroliterpipetten gibt es für unterschiedliche Voluminabereiche. Wir benutzen am Projekttag hauptsäch‐lich eine Mikroliterpipette, die im Bereich von 20 bis 200µL pipettiert und eine Pipette der Volumina von 100 bis 1000µL, wobei 1 µL einem tausendstel Milliliter (1 x 10‐3mL) also einem Millionstel Liter (1 x 10‐6L) entspricht. Andere Mikroliterpipetten umfassen die Voluminabereiche von z.B. 0,5 bis 10µL und 10 bis 100µL. Das zu pipettierende Volumen wird eingestellt, indem man am Einstellrad nach links bzw. rechts dreht; hierdurch wird die Kolbenhubhöhe verändert. Das eingestellte Volumen kann am „Sichtfenster“ abgelesen werden.

3
- 11 -
Achtung: Pipetten dürfen niemals überdreht oder unterdreht werden, d.h. bei der 20‐200µL Pipette darf niemals unter 020 bzw. über 200 eingestellt und bei der 1000der Pipette niemals über 100 bzw. unter 010µL eingestellt werden. Übung: Mikroliterpipetten werden niemals ohne Einwegspitze benutzt, haben zwei Druckpunkte und meist einen weiteren Druckknopf zum Abwerfen der Spitze. Versuche zuerst die Druckpunkte der Pipetten zu identifizieren. Nehme dann die passende Pipette, um auf den Filterpapierstreifen die angezeigten Volumina innerhalb der Kreise zu pipettieren. Gehe folgenderma‐ßen vor: 1. Einstellen des zu pipettierenden Volumens am Stellrad und Aufstecken der passenden
Pipettenspitze durch Andrücken der Spitze. 2. Drücken des Kolbens bis zum ersten Druckpunkt. 3. Eintauchen der Pipettenspitze mit gedrücktem Kolben in die zu pipettierende Flüssig‐
keit. 4. Den Druck vom Kolben langsam lösen, so dass dieser nach oben geht und die entspre‐
chende Menge an Flüssigkeit angesaugt wird. 5. Mittig über die Kreismarkierung des Filterpierstreifen gehen und durch langsames aber
kontinuierliches Drücken bis zum zweiten Druckpunkt sämtliche Flüssigkeit aus der Spitze entleeren. Pipette vom Filterpapier entfernen während der Kolben noch ge‐drückt bleibt.
6. Finger vom Kolben nehmen und den Spitzenabwurf über dem Abfallgefäß betätigen. Muster: Für größere Volumina werden i. d. R. sogenannte Glaspipetten in Verbindung mit einem Peleusball oder einer Pipettierhilfe verwendet. Pipettierhilfen werden durch leichten Druck unter Drehbewegung auf die Glaspipette aufgesetzt. Überführt man anschließend die Pipette in die abzumessende Flüssigkeit und dreht an dem Rad der Pipettierhilfe, wird eine entsprechende Menge Flüssigkeit aufgesogen. Diese kann entlas‐sen werden, indem man das Rad zurückdreht oder auf den Auslassknopf drückt. Es gibt unterschiedliche Ausführungen an Pipettierhilfen: Die roten sind für Glaspipetten von 10mL bis 25 mL, die Grünen für Pipetten von 5,0mL bis 10,0mL und die Blauen für Pipetten mit einem Volumen klei‐ner 1,0mL.
1 µL
2 µL
4 µL
8 µL
20 µL
40 µL
Auftrag in µL
erwartete G
röße
Pipettentest80 µL
1 µL
2 µL
4 µL
8 µL
20 µL
40 µL
Auftrag in µL
erwartete G
röße
Pipettentest80 µL

3
- 12 -
2.2 Gewinnung des Hefe‐Rohextraktes Bäckerhefe (Supermarkt) wird fein zerkrümelt und für ca. 1 Woche auf Fließpapier an der Luft getrocknet. Versuch: 1,5g Trockenhefe werden abgewogen und mit ca. 7,5mL 0,15 M K2HPO4‐Lösung in einem 15mL Falkontube gelöst und im Überkopfschüttler (bzw. durch ständiges Schütteln) bei Raumtemperatur für 30 min suspen‐diert und extrahiert. Die K2HPO4‐Lösung hierzu einfach bis zur 8‐9mL‐Markierung auf die Hefe geben. Die Suspension wird anschließend mit einer Pasteurpipette auf vier 2,2mL Eppis gleichmäßig aufgeteilt und die Zellbestandteile abzentrifugiert (15 min, max. speed). Der Überstand wird mittels eines Faltenfilters in ein frisches Falkontube filtriert, das Volumen abgelesen ( Ergebnisse) und die Proteinlösung auf Eis gela‐gert. Dieser sogenannte „Rohextrakt“, der u. a. die ADH enthält, ist die in den folgenden Experimenten verwendete Enzymlösung. 2.3 Proteinbestimmung: Eichreihe und Proteinkonzentration des Hefe‐Rohextraktes und die Bedienung unserer Photometer und der Stoppuhr zur Zeitmessung Für die quantitative Analyse von Proteinen gibt es eine Vielzahl von Methoden. Wir bestimmen heute den Proteingehalt des Rohextraktes nach der Methode von Bradford, bei der sich die Farbstoffmoleküle an die positiven Ladungen der Proteine binden und einen Komplex bilden, der im Photometer bei einer Wellen‐länge von 595nm sein Absorptionsmaximum (λmax = 595nm) besitzt. Wir messen also die Lichtabsorbtion eines Gemisches von Bradford‐Lösung und Proteinen. Je mehr Proteine vorhanden sind, desto mehr Farb‐stoffmoleküle können gebunden werden und desto größer ist die Extinktion des Lichts. Um die Protein‐menge unseres Rohextraktes quantitativ messen zu können, müssen wir aber wissen, welche Extinktion bei einer bestimmten Proteinmenge entsteht – wir müssen also mit bekannten Proteinmengen zunächst eine Extinktions‐Eichkurve erstellen. Als Eichsubstanz zur Berechnung der Proteinkonzentration dient uns eine Rinderserumalbumin‐Lösung bekannter Konzentration (RSA bzw. BSA (bovine serum albumine)). Versuch ‐ Proteineichreihe: 0, 20, 40, 60, 80 und 100µl der Rinderserumalbuminlösung (0,1mg/mL) entsprechen einer Proteinmenge von 0µg, 2µg, 4µg, …, 10µg werden in einem 1,5mL Eppi auf 800µL mit A. dest. aufgefüllt und 200µL Brad‐ford‐Reagens dazugegeben. Pipettierschema
RSA (0,1mg/mL)
0µL 20 µL 40 µL 60 µL 80 µL 100 µL
Wasser / H2O / A. bidest.
800 µL 780 µL 760 µL 740 µL 720 µL 700 µL
erst danach in alle: Bradford‐Reag. 200 µL 200 µL 200 µL 200 µL 200 µL 200 µL Nach Durchmischen (Eppis verschließen und mehrfach invertieren) werden die Ansätze 15min stehengelas‐sen und anschließend die Extinktion im Photometer bei 595nm gemessen. Der Ansatz mit 0µL RSA dient als Nullabgleich (blank / reference). Hierfür die Probe in die Küvette füllen und am Photometer „Null“ drücken,

3
- 13 -
es erscheint 0,000. Die Küvette entleeren (Flüssigabfall) und die nächste Probe einfüllen. Nun auf „messen“ drücken und den Extinktionswert nortieren. Auswertung: Die Extinktionswerte der übrigen Proben werden notiert ( Ergebnisse), in ein Koordinatensystem einge‐tragen und eine Eichkurve erstellt ( Ergebnisse) mit deren Hilfe im Anschluss die Proteinkonzentration des Rohextraktes bestimmt werden soll. Die Bedienung unserer Photometer
1) Am Druckknopf hinten links einschalten 2) Kalibrierung des Gerätes in Ruhe abwarten 3) „Einzel Wellenlänge“ am touch‐screen anwählen 4) Oben in der grauen Leiste die Wellenlänge (Zahl) anwählen 5) Gewünschte Wellenlänge eintippen (hier: 595nm) und mit „OK“ bestätigen 6) Küvettenschacht öffnen und Küvette für den Nullabgleich in die rechteckige Vertiefung ein‐
stellen. Hierbei müssen sich die klaren Seiten der Küvette links und rechts befinden. 7) Schließen des Küvettenschachts, Nullabgleich durch anwählen von „Null“ 8) Einstellen der Probenküvette und Messen der Extinktion durch anwählen von „Messen“ 9) Schritt 8) mit sämtlichen Proben durchführen, Extinktionsmesswerte notieren ( Ergebnisse) 10) Um die Wellenlänge zu verändern zurück ins Hauptmenü gehen und wieder durch anwählen von „Einzel Wellenlänge“ die entsprechende Wellenlänge einstellen (Schritte 4+5), weiter mit Schritt 6
Bedienung der Stoppuhr
1) „A“ drücken Stoppuhrbetrieb 2) „B“ drücken start und stopp 3) „C“ drücken stellt die Stoppuhr wieder auf „0“
Versuch – Bestimmung der Proteinmenge im Hefe‐Rohextrakt: Da die Proteinkonzentration in dem Rohextrakt wahrscheinlich so groß sein wird, dass die Messwerte pi‐pettierbarer Volumina außerhalb des linearen Bereichs unserer Eichkurve verlaufen würden, verdünnen wir (s. u.) einen Teil unseres Rohextraktes für die Proteinbestimmung. Herstellung einer 1:10 Verdünnung Hierzu werden allg. ein Teil der konzentrierten Lösung mit neun Teilen des Lösungsmittels gemischt (1 Teil + 9 Teile). Hier nehmen wir 50µL des Rohextraktes und 450µL Wasser und mischen diese in einem Eppendorfgefäß (Eppi). Je 0, 25, 50 und 100µl einer 1:10 Verdünnung des Rohextraktes werden in 1,5 mL Eppis auf 800µL mit A. dest. aufgefüllt und 200µL Bradford‐Reagens dazugegeben.
3
- 14 -
Pipettierschema
Rohextrakt (1:10 verdünnt)
0µL 25 µL 50 µL 100 µL
Wasser / H2O / A. bidest.
800 µL 775 µL 750 µL 700 µL
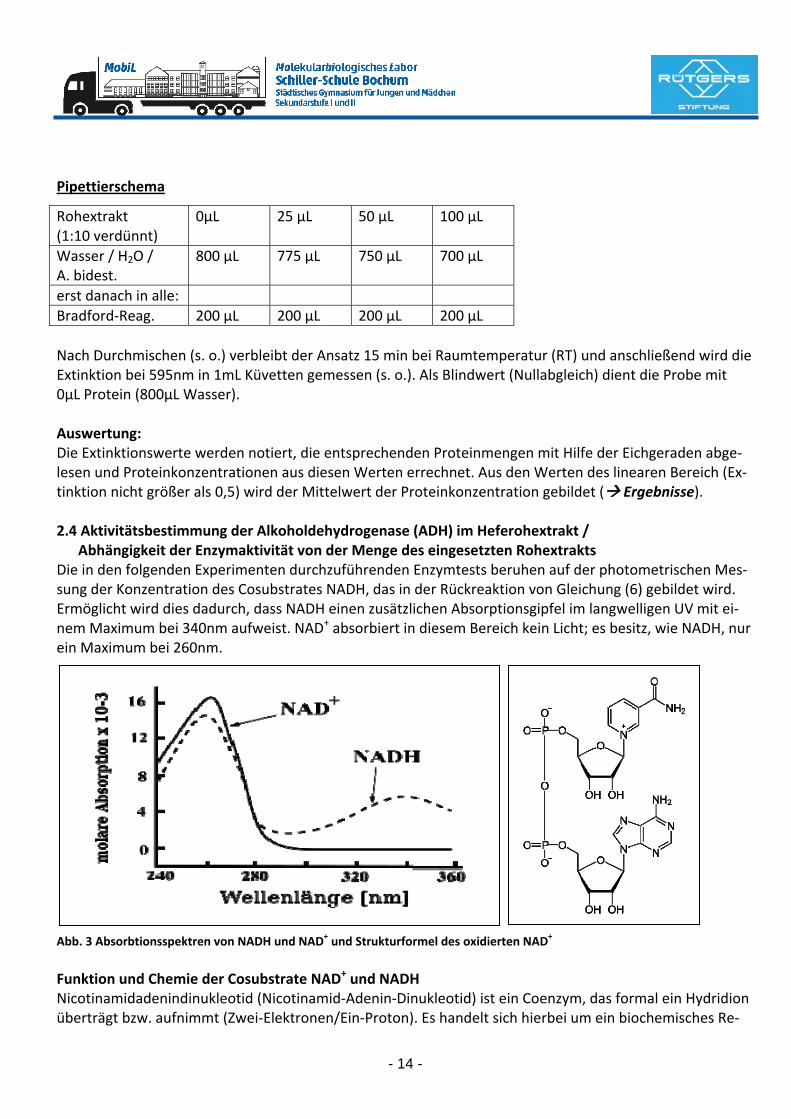
erst danach in alle: Bradford‐Reag. 200 µL 200 µL 200 µL 200 µL Nach Durchmischen (s. o.) verbleibt der Ansatz 15 min bei Raumtemperatur (RT) und anschließend wird die Extinktion bei 595nm in 1mL Küvetten gemessen (s. o.). Als Blindwert (Nullabgleich) dient die Probe mit 0µL Protein (800µL Wasser). Auswertung: Die Extinktionswerte werden notiert, die entsprechenden Proteinmengen mit Hilfe der Eichgeraden abge‐lesen und Proteinkonzentrationen aus diesen Werten errechnet. Aus den Werten des linearen Bereich (Ex‐tinktion nicht größer als 0,5) wird der Mittelwert der Proteinkonzentration gebildet ( Ergebnisse). 2.4 Aktivitätsbestimmung der Alkoholdehydrogenase (ADH) im Heferohextrakt / Abhängigkeit der Enzymaktivität von der Menge des eingesetzten Rohextrakts Die in den folgenden Experimenten durchzuführenden Enzymtests beruhen auf der photometrischen Mes‐sung der Konzentration des Cosubstrates NADH, das in der Rückreaktion von Gleichung (6) gebildet wird. Ermöglicht wird dies dadurch, dass NADH einen zusätzlichen Absorptionsgipfel im langwelligen UV mit ei‐nem Maximum bei 340nm aufweist. NAD+ absorbiert in diesem Bereich kein Licht; es besitz, wie NADH, nur ein Maximum bei 260nm.
Abb. 3 Absorbtionsspektren von NADH und NAD+ und Strukturformel des oxidierten NAD+
Funktion und Chemie der Cosubstrate NAD+ und NADH Nicotinamidadenindinukleotid (Nicotinamid‐Adenin‐Dinukleotid) ist ein Coenzym, das formal ein Hydridion überträgt bzw. aufnimmt (Zwei‐Elektronen/Ein‐Proton). Es handelt sich hierbei um ein biochemisches Re‐

3
- 15 -
doxpaar. Die reduzierte Form ist das NADH, die oxidierte Form das NAD+. Beide Formen sind an zahlreichen Redoxreaktionen des Stoffwechsels der Zelle beteiligt.
Abb. 4 NAD+ kann durch Aufnahme von zwei Elektronen (e−) und einem Proton (H+) zu NADH reduziert werden. Versuch: Sämtliche nachfolgenden Testes werden im HachLange Photometer bei einer Wellenlänge von 366nm in 3mL Plastikküvetten durchgeführt. Die Tests selber werden in Reagenzgläsern angesetzt. Wichtig ist, dass der Hefe‐Rohextrakt erst direkt vor der Messung hinzugegeben wird, da ansonsten die en‐zymatische Reaktion bereits abgelaufen ist, bevor sich die Küvette im Photometer befindet. Pipettierschema (Volumenangaben in mL) Proben‐Nr. 1 2 3 4 Glycin‐NaPP‐Puffer pH 9.0 2,5 2,5 2,5 2,5 Ethanol (EtOH, absolut) 0,1 0,1 0,1 0,1 NAD (15mg / mL) 0,1 0,1 0,1 0,1 Aqua bidest. 0,28 0,25 0,225 0,2 [280µL 250µL 225µL 200µL] Proben durch Schwenken im Reagenzglas vermischen. Probe eins dann in eine Küvette geben und den Nul‐labgleich am Photometer durchführen. Danach wird der Rohextrakt (s. u.) in die Küvette pipettiert (20µL) und der Küvetteninhalt nach Auflegen eines Stückes Parafilm durch 2‐maliges Umdrehen zwischen Daumen und Zeigefinger gemischt. Die Küvette wird anschließend sofort ins Photometer verbracht und gleichzeitig die Stoppuhr gestartet. Die Extinktionswerte werden sodann in Abständen von 15 Sekunden über einen Zeitraum von 2 Minuten abgelesen und notiert ( Ergebnisse).
Messungen jetzt alle bei 366nm !!!!
3
- 16 -
Anschließend wird mit den Proben 2‐4 gleichermaßen verfahren, d. h. der Rohextrakt darf erst zu Küvette 2 gegeben werden, wenn die erste Messung abgeschlossen ist (usw.). Rohextrakt 0,02 0,05 0,075 0,1 [20µL 50µL 75µL 100µL] Auswertung: Trage die Messwerte in die vorbereitete Tabelle ein, trage die Werte in ein Diagramm ein und bestimme aus der Steigung der Kurven das ΔE/min (Extinktionsänderung pro Minute) für die Proben 1‐4 ( Ergebnis‐se). 2.5 Die pH‐Abhängigkeit der Alkoholdehydrogenase‐Aktivität Ein entscheidender Parameter bei enzymatischen Analysen ist die Wahl des geeigneten pH‐Wertes der In‐kubationslösung. pH‐Änderungen beeinflussen die Stabilität und die Reaktivität der beteiligten Enzyme. Darüber hinaus greift die H+‐Ionenkonzentration vielfach auch in das Reaktionsgleichgewicht ein. Das ist z.B. bei allen NAD(P)‐abhängigen Reaktionen der Fall (vgl. oben). Aus diesem Grund sind enzymkatalysierte Reaktionen grundsätzlich in ausreichend gepufferten Lösungen mit eingestelltem pH‐Wert durchzuführen. Der Einfluss verschiedener pH‐Werte auf die Aktivität der ADH soll im folgenden Experiment demonstriert werden. Pipettierschema (Volumenangaben in mL) Proben‐Nr. 1 2 3 4 5 6 Puffer (1‐6) 2,7 2,7 2,7 2,7 2,7 2,7 Ethanol (absolut) 0,1 0,1 0,1 0,1 0,1 0,1 NAD (15mg / mL) 0,1 0,1 0,1 0,1 0,1 0,1 Rohextrakt (Starter) 0,1 0,1 0,1 0,1 0,1 0,1 Puffer 1: Kaliumphosphat / KPi pH 6,0 Puffer 2: Kaliumphosphat / KPi pH 7,0 Puffer 3: Kaliumphosphat / KPi pH 8,0 Puffer 4: Glycin‐NaOH pH 9,0 Puffer 5: Glycin‐NaOH pH 10,0 Puffer 6: Glycin‐NaOH pH 11,0 Die einzelnen Tests werden wie in der unter 2.4 beschriebenen Weise durchgeführt und ausgewertet. An‐schließend wird der pH‐Wert der Proben 1‐6 mittels pH‐Indiktorpapier nachgemessen ( Ergebnisse).

3
- 17 -
2.6 Abhängigkeit der Alkoholdehydrogenase‐Aktivität von der Substratkonzentration Da bei Enzymreaktionen Enzym und Substrat aufeinander treffen, ist zu vermuten, dass auch eine Steige‐rung der Substratkonzentration zu einer erhöhten Umsatzrate und damit zu einer höheren Enzymatischen Aktivität führt. Folgender Versuch soll diese Hypothese prüfen: Pipettierschema (Volumenangaben in mL) Proben‐Nr. 1 2 3 4 5 6 7 8 Glycin‐NaPP‐Puffer pH 9.0 2,0 2,0 2,0 2,0 2,0 2,0 2,0 2,0 Ethanol (1:100 verd.) 0,0 0,02 0,03 0,05 0,1 0,2 0,3 0,5 NAD (15mg / mL) 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 Aqua bidest. 0,8 0,78 0,77 0,75 0,7 0,6 0,5 0,3 Rohextrakt 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 Die einzelnen Tests werden wie in der unter 2.4 beschriebenen Weise durchgeführt. Bestimme die Reakti‐onsgeschwindigkeit „v“, ausgedrückt als ΔE/min. Falls im Ansatz 1 eine Reduktion stattfindet, ist dieser Wert von den anderen Werten abzuziehen ( Ergebnisse). 2.7 Berechnungen für Spezialisten: Für Berechnungen der Enzymaktivitäten in U bzw. kat und spezifische Enzymakivitäten in U bzw. kat pro mg‐Protein: Es gilt Gleichung (4): E = ε x c d E ist dimensionslos, c wird in mol/mL (mol/cm3) gemessen und d in cm. Dann hat ε die Einheit cm2/mol. Für NADH gilt: ε366 = 6,22 x 106cm2/mol, d.h. wenn 1 mol NADH/mL gebildet wird und d=1cm beträgt, nimmt die Extinktion um 6,22x106 zu (1µmol/mL/E = 6,22; 1nmol/mL/E = 0,0062 Zu beachten ist weiterhin, das im Photometer Konzentrationen gemessen werden (d.h. bei der ADH: Um‐satz/mL). Beispielrechnung: Es sei: ΔE/min = 0,124; es wurden 0,1mL Rohextrakt (5mg Protein/mg) eingesetzt, das Ansatzvolumen betrug 3mL.

3
- 18 -
Daraus folgt: Δc/Δt x ε d = 0,124/6,2 x 106 x 1 (mol/cm2 x cm x min) = 20x10‐9 mol/min x mL Küvettenvolumen = 60x10‐9 mol/min x 3mL Küvettenvolumen 3 mL Küvettenvolumen enthielten 0,1mL Rohextrakt, der Rohextrakt hat somit eine Enzymaktivität von = 600x10‐9 mol/min x mL Rohextrakt oder = 0,6µmol/min x mL Extrakt = 0,6U/mL Rohextrakt Die spezifische Aktivität der ADH errechnet sich daraus bei einer Proteinkonzentration von 5mgProtein/mL Rohextrakt mit: = 0,6 U/5mg Protein = 0,12 U/mg Protein = 2nkat/mg Protein Für Super‐Strategen: Für jede enzymkatalysierte Reaktion gilt: (8) Enzym + Substrat Enzym‐Substrat‐Komplex Produkt + Enzym Kurzform: E + S ES P + E Für die weiteren Überlegungen gilt die in der Regel zutreffende Voraussetzung, dass die 2. Teilreaktion we‐sentlich! langsamer verläuft als die Bildung von ES. Dann ist die messbare Geschwindigkeit der Gesamtre‐aktion (V) proportional der Konzentration an ES, d.h. „V“ ergibt ein Maß für die Konzentration an ES. Die Anwendung des Massenwirkungsgesetzes für die 1. Teilreaktion aus der Gleichung (8) ergibt: (9) [E] x [S]
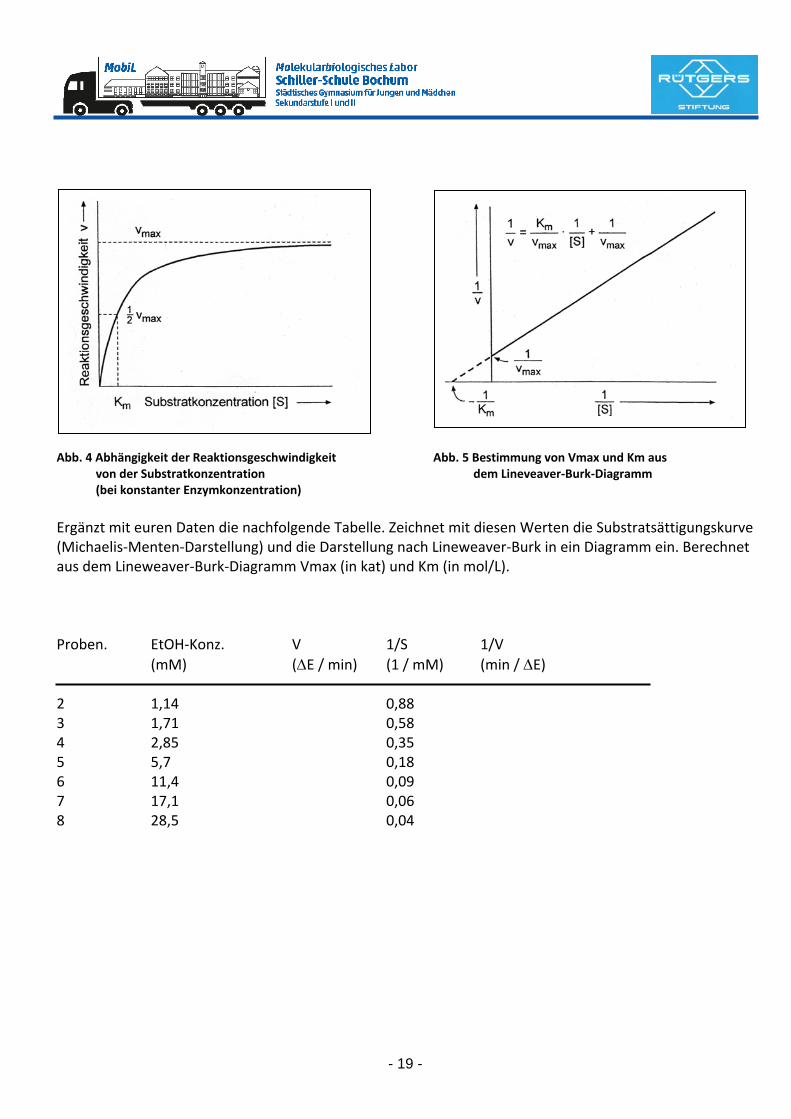
= K [ES] Daraus folgt, dass bei einer gegebenen Menge an E und steigenden Mengen an S immer mehr ES gebildet wird. Gleichzeitig steigt die Reaktionsgeschwindigkeit asymptotisch bis zum Grenzwert Vmax an, bei dem praktisch alles E als ES vorliegt (= Substratsättigung). Eine weitere Steigerung von V ist bei gegebener En‐zymkonzentration nicht möglich. Wie Abb. 5 zeigt, lässt sich die Sättigungskonzentration, im Gegensatz zu Vmax nur schlecht aus der Kurve ablesen. Definierte Verhältnisse ergeben sich jedoch, wenn ½ Vmax als Bezugspunkt gewählt wird. In diesem Fall liegt E zur Hälfte als ES vor, da V proportional ES sein sol. Glei‐chung (9) vereinfacht sich für diesen Fall zu (10) [S] = K d.h. die Substratkonzentration, bei der halbmaximale Reaktionsgeschwindigkeit erreicht ist, ist gleich der Dissoziationskonstanten von ES. Nach den Begründern der Theorie wird sie als Michaelis‐Menten‐Konstante (Km) genannt.
3
- 19 -
Abb. 4 Abhängigkeit der Reaktionsgeschwindigkeit Abb. 5 Bestimmung von Vmax und Km aus von der Substratkonzentration dem Lineveaver‐Burk‐Diagramm (bei konstanter Enzymkonzentration) Ergänzt mit euren Daten die nachfolgende Tabelle. Zeichnet mit diesen Werten die Substratsättigungskurve (Michaelis‐Menten‐Darstellung) und die Darstellung nach Lineweaver‐Burk in ein Diagramm ein. Berechnet aus dem Lineweaver‐Burk‐Diagramm Vmax (in kat) und Km (in mol/L). Proben. EtOH‐Konz. V 1/S 1/V (mM) (ΔE / min) (1 / mM) (min / ΔE) 2 1,14 0,88 3 1,71 0,58 4 2,85 0,35 5 5,7 0,18 6 11,4 0,09 7 17,1 0,06 8 28,5 0,04
3
- 20 -
III Ergebnisse / Auswertung
3.2 Gewinnung des Hefe‐Rohextraktes
Ergebnis: Volumen des Rohextraktes: ______ mL
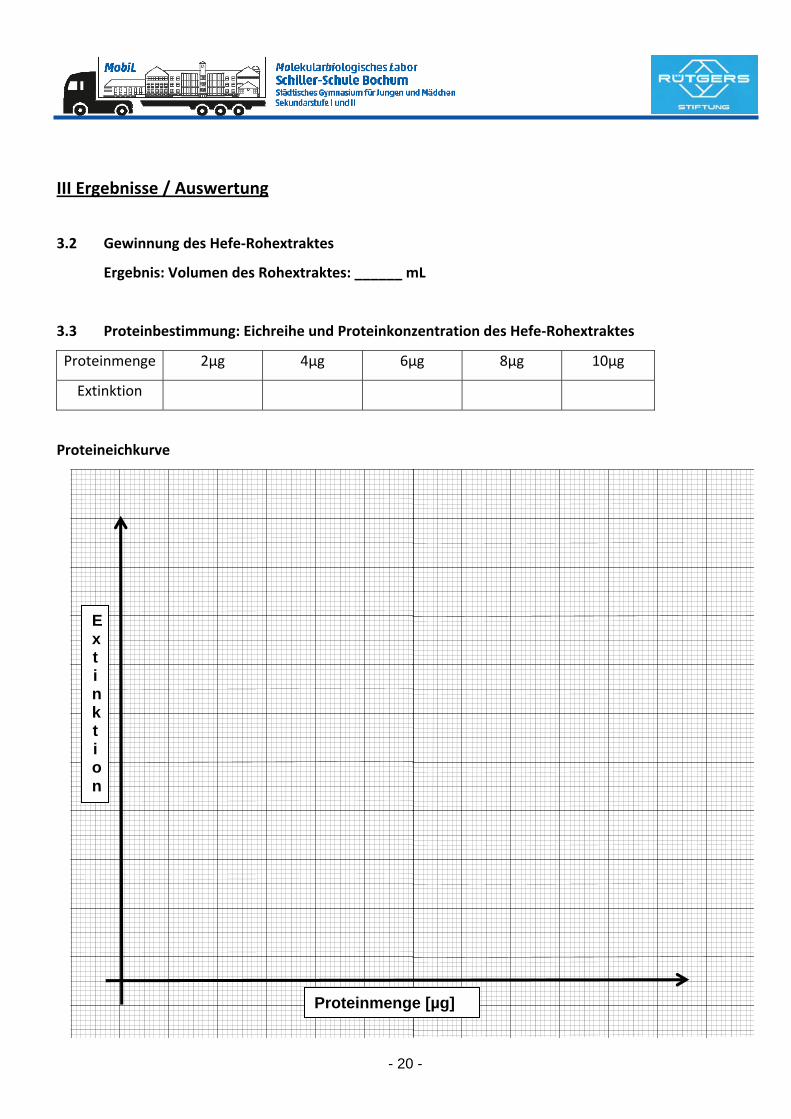
3.3 Proteinbestimmung: Eichreihe und Proteinkonzentration des Hefe‐Rohextraktes
Proteinmenge 2µg 4µg 6µg 8µg 10µg
Extinktion
Proteineichkurve
Proteinmenge [µg]
E x t i n k t i o n
3
- 21 -
Proteinmengen und Proteinkonzentration des Rohextraktes
Eingesetztes Volumen einer 1:10 Verdünnung
Extinktion Protein/Volumen der 1:10 Verdünnung
Proteinkonzentration des Rohextraktes
25µL µg/25µL µg/µL
50µL µg/50µL µg/µL
100µL µg/100µL µg/µL
Proteinkonzentration: _______________ mg/mL
Gesamt‐Proteinmenge: _______________ mg
3.4 Aktivitätsbestimmung der Alkoholdehydrogenase im Heferohextrakt /
Abhängigkeit der Enzymaktivität von der Menge des eingesetzten Rohextrakts
Zeit (sec)
Extinktion des Rohextraktes
20µl 50µL 75µL 100µL
15
30
45
60
75
90
105
120
Aus der Steigung der jeweiligen Kurve ergibt sich ein ΔE/min von:
Rohextrakt ΔE/min
20µL
50µL
75µl
100µL
3
- 22 -
Aktivitätsbestimmung der Alkoholdehydrogenase (ADH) im Heferohextrakt 3.5 Die pH‐Abhängigkeit der Alkoholdehydrogenase‐Aktivität
Zeit (sec) E bei pH 6 E bei pH 7 E bei pH 8 E bei pH 9 E bei pH 10 E bei pH 11
15
30
45
60
75
90
105
120

3
- 23 -
Ergebnisse / Auswertung 2.2 Gewinnung des Hefe‐Rohextraktes Ergebnis: Volumen des Rohextraktes: ______ mL Tabelle für Werte der Eichkurve Eichkurve Tabelle für Werte der Messungen der Proteinlösung Platz für Rechnung Proteinkonzentration: ________________ mg/mL Gesamt‐Proteinmenge: _______________ mg Tabelle und Koordinatensystem zu Spektren von NAD+ und NADH (Wellenlänge Kontrolle(Küvette mit Wasser) Probe (Küvette mit NAD/NADH) Aus der Steigung der jeweiligen Kurve ergibt sich ein ΔE/min von:
pH‐Wert ΔE/min Nachgemessener pH‐Wert
6,0
7,0
8,0
9.0
10,0
11,0
3
- 24 -
3.6 Abhängigkeit der Alkoholdehydrogenase‐Aktivität von der Substratkonzentration
S u b s t r a t : E t h a n o l 1 : 1 0 0 v e r d ü n n t Zeit (sec) 0µL 20µL 30µL 50µL 100µL 200µL 300µL 500µL
15
30
45
60
75
90
105
120

3
- 25 -
Aus der Steigung der jeweiligen Kurve ergibt sich ein ΔE/min von:
Substratmenge
EtOH 1:100
ΔE/min
0µL
20µL
30µL
50µL
100µL
200µL
300µL
500µL

3
- 26 -
Zusatzblatt: Ergebnisse / Auswertung 2.2 Gewinnung des Hefe‐Rohextraktes Ergebnis: Volumen des Rohextraktes: ______ mL Tabelle für Werte der Eichkurve Eichkurve Tabelle für Werte der Messungen der Proteinlösung Platz für Rechnung Proteinkonzentration: ________________ mg/mL Gesamt‐Proteinmenge: _______________ mg Tabelle und Koordinatensystem zu Spektren von NAD+ und NADH (Wellenlänge Kontrolle(Küvette mit Wasser) Probe (Küvette mit NAD/NADH)

3
- 27 -
IV Anhang 4.1 Aufgabenstellungen
1) Welche Stoffmengenkonzentration hat eine Zuckerlösung (C6H12O6), wenn du 90g Zucker in 500mL
Wasser löst?
2) Wie viel Mol Wasser sind in einem Liter Wasser enthalten?
3) Was verbindest du mit dem Namen: Lambert‐Beer?
4) Welche Stoffmenge an Wasserstoffatomen befinden sich in einem Mol Methan (CH4)?
5) Gib die Molekülmasse von Methan an.
6) Welche Masse hat ein Mol Methan?
7) Das Molekulargewicht von NaOH beträgt 40g/mol. Wie viel Gramm NaOH musst du abwiegen, um
500mL einer 0,25M NaOH‐Lsg. herzustellen?
Hinweise: Molare Massen: C = 12g/mol, H = 1g/mol, O = 16g/mol
8) Beschreibe die Wirkungsweise von Enzymen.
9) Was bedeutet im Zusammenhang von Enzymatik die Begriffe: Substratspezifität und Wirkungsspezi‐
fität?
10) Gib die international gültige Einheit der Enzymaktivität sowie deren Bedeutung an.
11) Beschreibe die Bedeutung des Begriffs: Cosubstrat
12) Was verbirgt sich hinter dem Begriff: Extinktionsänderung pro Zeit (ΔE/min)?
13) Erläutere: E = ε x c x d
14) Schreibe die Reaktionsgleichungen auf, die durch die ADH katalysiert werden.
15) Worauf beruht der Protein‐Quantifizierungstest nach Bradford?
16) Warum ist die spezifische Aktivität eines „reinen“ Proteins größer als die eines Rohextraktes?
17) Auf welchem physiko‐chemischen Vorgang beruht die hier durchgeführte Messung der ADH‐
Aktivität?
18) Nennen Gründe dafür, dass Enzyme in extrem saurem oder extrem basischen Lösungen nicht arbei‐
ten.
19) Enzyme sind Proteine. Beschreibe, woraus Proteine bestehen und erläutere deren Aufbau.

3
- 28 -
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________

3
- 29 -
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________
_______________________________________________________________________________________

3
- 30 -
4.2 Materialien (jeweils pro Vierergruppe)
4.2.1 Chemikalien und Lösungen Trockenhefe, 5g (1 kg Presshefe = ca. 250g Trockenhefe) Aqua dest. Eis Ethanol p.a., 10mL ADH aus Hefe (Sigma); 2800U in 2 mL A. dest. Rinderserumalbumin (RSA, 0,1mg / mL); 1,5mL Bradfordreagenz (80mg Coomassie Brillantblau G in 50mL Ethanol techn. gelöst, 2h gerührt, dann 100mL 85%ige Phosphor‐säure zugegeben und mit A. dest. auf 1L aufgefüllt. Nach einer weiteren Stunden unter Rühren, filtriert und im Dunkeln bei 4°C aufbewahrt) NAD (5x10‐5M); 3mL (1xpro Kurstag) NADH (5xs10‐5M); 3mL (1x pro Kurstag) NAD (15mg pro mL); 4,5mL K2HPO4 (0,15M); 70mL Kalium‐Phosphat‐Puffer (0,1M, pH 6,0); 5mL Kalium‐Phosphat‐Puffer (0,1M, pH 7,0); 5mL Kalium‐Phosphat‐Puffer (0,1M, pH 8,0); 5mL Glycin‐NaOH‐Puffer (0,1M, pH 9,0); 5mL Glycin‐NaOH‐Puffer (0,1M, pH 10,0); 5mL Glycin‐NaOH‐Puffer (0,1M, pH 11,0); 5mL Glycin‐Na‐Pyrophosphat‐Puffer (0,75M, pH 9,0); 50mL Glycin‐Na‐Pyrophosphat‐Semicarbazid‐Puffer (0,75M, pH 8,7); 100mL Bier, alkoholfreies Bier, Schnaps; je einige mL 4.2.2 Glaswaren und Verbrauchsmaterialien Enzymtestpipetten 10mL; 5x 40 Stück 5mL; 10x 80 Stück [1mL; 5x // 0,5mL; 10x // 0,1mL; 10x] 40 + 80 + 80 oder Mikroliterpipetten anstelle: Mikropipetten P‐1000 und P‐100 Pasteurpipetten mit Schnubbel; 10x pH‐Indikatorpapier (pH 4‐10) Parafilm Bechergläser (100mL); 1x Messzylinder (50mL und 25mL); je 1x Messkolben (100mL); 4x Glasstab; 1x Spritzflasche mit H2O; 2x Eisbox; 1x Plastikküvetten; 50x Reagenzglasgestell; 3x Wägeschälchen

3
- 31 -
4.2.3 Geräte Obeschalenwaage Überkopfschüttler Eppendorfzentrifuge Spektralphotometer Laborstoppuhren 4.3 Literaturverzeichnis Strasburger: Lehrbuch der Botanik, Teil Physiologie, 35.‐36. Auflage, Spektrum, 2002/2004 H.‐W. Heldt: Pflanzenbiochemie. 2. Auflage, Spektrum, 1999 G. Richter: Stoffwechselphysiologie der Pflanzen. 6. Auflage, Thieme, 1997 C. E. Mortimer: Chemie. Das Basiswissender Chemie. 6. Auflage, Thieme, 1996 B. K. Keppler, A. Ding: Chemie für Biologen. Spektrum, 1997 F. Schaller: Experimentelle Untersuchungen von stoffwechselphysiologischen Prozessen bei der Sonnenblume – Entwicklung einer Handreichung für die Fachkonferenz Biologie, Schriftliche Hausarbeit zur Zweiten Staatsprüfung, 2008 F. Schaller: Identifizierung von Lebensmittelskandalen. Ist Pferdefleisch im Döner?, Praktikumsvorschrift für Schülerinnen und Schüler, 2013