forstliche forschungsberichte münchen nr. 215 /...
TRANSCRIPT


Forstliche Forschungsberichte München Nr. 215 / 2016
Schriftenreihe des
Zentrums Wald-Forst-Holz Weihenstephan
für
die Studienfakultät für Forstwissenschaft und Ressourcenmanagement der Techni-schen Universität München, die Fakultät Wald und Forstwirtschaft der Hochschule
Weihenstephan-Triesdorf und die Bayerische Landesanstalt für Wald und Forstwirt-schaft als Sonderbehörde der Bayerischen Forstverwaltung
Energiegehalt und Qualität der Nahrung von Rehen (Capreolus capreolus) im Jahresver-
lauf in zwei unterschiedlich geprägten Habitaten
Andreas König, Martina Scheingraber und Juliane Mitschke
Zentrum Wald Forst Holz Weihenstephan Hans-Carl-von-Carlowitz-Platz 1
D-85354 Freising

Abschlussbericht des Projektes „Energiegehalt der Rehnahrung“
Impressum
Der Druck dieses Berichts wurde ermöglicht mit finanzieller Unterstützung des Fördervereins Zentrum Wald Forst Holz Weihenstephan e.V.
ISSN 0174-1810 ISBN 3-933506-46-8
Alle Rechte, insbesondere das Recht der Vervielfältigung und Verbreitung sowie der Übersetzung
vorbehalten.
Titel Energiegehalt und Qualität der Nahrung von Rehen (Capreolus capreolus) im Jahresverlauf in unterschiedlich geprägten Habitaten
Analysis of annual changes of energy and quality in roe deer diet in two different habitats.
Schlüsselworte Reh, Nahrung, umsetzbare Energie, Qualität der Nahrung, Stress, Pansen-Mikrobiom, Bayern
Keywords Roe deer, diet energy, diet quality, stress, rumen microbiome, Ba-varia
Autoren PD Dr. Andreas König, MSc. Martina Scheingraber und MSc. Juliane Mitschke Arbeitsgruppe Wildbiologie und Wildtiermanagement, Technische Universität München
Herausgeber Zentrum Wald Forst Holz Weihenstephan Hans-Carl-von-Carlowitz-Platz 1 D-85354 Freising www.forstzentrum.de
Schriftleitung Dr. Joachim Hamberger
Dokumentation Forstliche Forschungsberichte München Nr. 215, 2016, 199 Seiten
Erscheinungsdatum Juli 2016
Druck Zuerl, München bzw. andere Druckerei
Umschlagseite grafik+design Gerd Rothe, Wang
Auflage 300
Zu beziehen über Förderverein Zentrum Wald Forst Holz Weihenstephan e.V. Hans-Carl-von-Carlowitz-Platz 1, 85354 Freising Tel.: 08161/71-4643; Fax: 08161/71-5404 E-Mail: [email protected]

Abschlussbericht des Projektes „Energiegehalt der Rehnahrung“
3
Stand des Projektes
Mit Schreiben Nr. F8-7904-1/26 vom 4.11.2011 bewilligte das Staatsministerium für
Ernährung, Landwirtschaft und Forsten Mittel für die Förderung eines Forschungsvor-
habens zu dem Thema „Analyse der Veränderung des Energiegehaltes und der Quali-
tät der Rehnahrung im Jahresverlauf“. Das Projekt hatte eine Laufzeit vom 01.01.2012
bis zum 31.12.2014.
Gesamtprojektleitung PD Dr. habil. Andreas König, TUM
Projektleitung Qualität der Rehnahrung Prof. Dr. Wilhelm Windisch, TUM Projektbearbeitung Martina Scheingraber M.Sc. Projektmitarbeit Juliane Mitschke M.Sc. Katrin Heckel B.Sc. Kooperation Jürgen Danier, Bioanalytik, TUM Ao. Univ.-Prof. Dr. Rupert Palme, Vetmed-Uni Wien

Danksagung
4
Danksagung
Besonderer Dank gilt dem Obersten Jagdbeirat sowie der Obersten Jagdbehörde in
Bayern für die Förderung des Projektes. Die Konrad Adenauer Stiftung übernahm die
Finanzierung der Doktorandin, wofür ihr gedankt sei. Dem Bayerischen Jagdverband
gilt unser Dank für konstruktive Gespräche und Anregungen in Feldkirchen und in den
Kreisgruppen. Einen wesentlichen Beitrag zum Gelingen des Projektes leisteten die
Revierverantwortlichen der Gemeinschaftsjagdreviere Angerstorf-Lohbruck, Vordersar-
ling und Rampoldstetten, in dem Sie uns sehr umfangreich mit Probenmaterial über die
Jahre versorgten und uns in ihren Jagdbetrieb integrierten. Eine ebenfalls umfassende
Unterstützung erfuhren wir durch den Forstbetrieb München sowie den Begehungs-
scheininhabern des Staatsjagdrevieres Baierbrunn. All diesen unermüdlichen Unter-
stützern gilt unser herzlicher Dank. In besonderem Maße möchten wir uns auch bei
den Kollegen der Technischen Universität München sowie der Veterinärmedizinischen
Universität Wien für die konstruktive Kooperation bedanken.

Zusammenfassung / Summary
5
Zusammenfassung
In einem Waldhabitat sowie in einem landwirtschaftlich geprägten Habitat würden von
2001 bis 2014 über 12 Monate hinweg von 220 Rehen Pansen gesammelt. Ziel war die
umsetzbare Energie und Qualität der Rehäsung anhand der tatsächlich aufgenommen
Äsung im gesamten Jahresverlauf zu ermitteln. Um sich ein umfassendes Bild von der
Energieversorgung der Rehe machen zu können, wurde im Rahmen eines wildbiologi-
schen Systemansatzes Konditionsparamter der Rehe sowie ihre Anpassung an die
lokale und jahreszeitlich bedingte Äsung erfasst. Weiterhin wurden Rahmenbedingun-
gen wie Verfügbarkeit und Qualität der lokalen Vegetation beurteilt sowie das Stressni-
veau der Rehe als Maß für menschliche Einflüsse aufgenommen. Für die Pansen-
Analysen wurden die Standardmethoden der Futtermittelanalyse verwendet. Den Re-
hen im Agrarhabitat steht mit durchschnittliche 6,29 MJ/kg TS aufgenommener Nah-
rung signifikant mehr Energie zur Verfügung als den Waldrehen mit durchschnittlich
5,43 MJ/ kg TS aufgenommener Nahrung. Waldrehe kompensieren den geringeren
Energiegehalt der Äsung durch höhere Äsungsmengen. Die Panseninhalte der Wal-
drehe waren vor allem im Herbst und Winter im Mittel etwa 300 g schwerer als die der
Landrehe. Im Schnitt lag der Fasergehalt der Äsung nicht unter 23 % TS. Im Vergleich
zu dem Raufutterfresser „Schaf“ konnten Rehe mit ihrem auf die lokale Vegetation an-
gepassten Pansensaft im Winter aus faserhaltiger Äsung mehr Energie gewinnen.
Aufgrund dessen sollte der Begriff „Konzentrat-Selektierer“ durch „Selektierer“ ersetzt
werden. In den drei Untersuchungsjahren ergab sich für beide Populationen kein Ener-
gieengpass. Energetisch gesehen, haben die Landrehe nicht gemerkt, dass es Winter
war.
Schlüsselworte: Reh, Nahrung, umsetzbare Energie, Qualität der Nahrung, Stress,
Pansen-Mikrobiom, Bayern

Zusammenfassung / Summary
6
Summary
From 2011 to 2014, the rumina of 220 roe deer and data on their condition were collec-
ted in both a forest habitat and an agricultural habitat. To permit the observation of se-
asonal influences, we gathered samples throughout the 12 months of each year. Our
aim was to measure the quality and energy content of the natural roe deer diet by
examining the actual content of their rumina. In order to get an overview of the total
energy budget of the roe deer, we used a wildlife systems approach to analyse the
condition and physiological adaption of roe deer to local and seasonal changes in diet.
In addition, the availability and quality of the local vegetation were evaluated, as well as
the stress of the deer as a measure of human influence. Standard methods of dietary
analysis were used. The fodder of roe deer in the agricultural habitat had a mean ener-
gy content of 6.29 MJ/ kg DM, which was significantly more digestible energy than was
available to roe deer in the forest habitat, where the mean was 5.43 MJ/kg DM. Forest
deer compensate for the lower energy content by browsing more. The rumen intake of
roe deer from the forest habitat was on average 300 g heavier than that of deer from
the agricultural habitat, particularly in autumn and winter. However, in both habitats the
proportion of fibre was not less than 23 % DM. As the roe deer’s ruminal fluid is adap-
ted to the local vegetation, they are able to extract more energy from a fibrous diet than
the domestic grazer sheep. The German term “concentrate selector” should therefore
be replaced by “selector” or “browser” to avoid misinterpretations. During the study
period, we did not observe any shortage of energy in either population. In terms of diet
energy, roe deer in our agricultural habitat did not notice that it was winter.
Keywords: Roe deer, diet energy, diet quality, stress, rumen microbiome, Bavaria

0 Kurzfassung
7
0 Kurzfassung (König)
0.1 Hintergrund
Das Reh (Capreolus capreolus) ist neben dem Wildschwein (Sus scrofa) die am häu-
figsten vorkommende Schalenwildart in Deutschland. Seine Verbreitung reicht von
sehr naturbelassenen Waldökosystemen bis hin zu intensiv genutzten Agrarlandschaf-
ten. Habitate wie die Hochlagen der bayerischen Alpen, die von Autoren wie Dietzel
(1921) als für Rehe ungeeignet beschrieben wurden, werden von dieser Schalenwildart
heute genauso genutzt wie suburbane und urbane Gebiete. Das Reh hat sich in den
letzten 100 Jahren als sehr anpassungsfähig erwiesen. Andererseits gibt es Stimmen,
die gerade die Lebensbedingungen von Rehen außerhalb von relativ naturnahen
Waldökosystemen problematisch sehen. So wird die These vertreten, dass die heutige
Landschaft, speziell die Agrarlandschaft, generell nicht mehr wildkonform ist und die
artgemäßen Ernährungsansprüche nicht mehr hinreichend erfüllt. Begründet wird dies
damit, dass Rehe als „Konzentrat-Selektierer“ auf leicht verdauliche, nährstoffreiche
Äsung angewiesen sind, da ihnen rohfaserspaltende Bakterien fast gänzlich fehlen.
Demnach sollte die Rehnahrung vor allem reich an Proteinen und leicht verdaulichen
Fasern bzw. Kohlehydraten sein.
Wie weit das Rehwild mit eher faserreicher Nahrung und weniger leicht verdaulichen
Nährstoffen zu Recht kommt, war und wird heftig diskutiert. So weisen Autoren wie
Dissen & Hartfiel (1985), Anke et al. (2007) oder Clauss (2010) gerade auf die Ver-
wertbarkeit von Rohfasern durch Rehe hin. Dagegen wird die Definition von Notzeit
unter anderem mit geringer Nahrungsqualität in Verbindung gebracht (Helm 2015). Es
gibt zwar eine Vielzahl von Untersuchungen zur Ernährung von Rehen, ihre Vergleich-
barkeit ist auf Grund der verwendeten Methoden und der unterschiedlichen Untersu-
chungsgebiete nur eingeschränkt möglich.
Was bei Durchsicht all dieser Arbeiten fehlt ist eine systemtheoretisch aufgebaute Ana-
lyse der Energie der von Rehen wirklich aufgenommenen Äsung unter Berücksichti-

0 Kurzfassung
8
gung der Qualität und Verdaulichkeit der Äsung. Die Anpassung des Rehes an die jah-
reszeitlichen Äsungsschwankungen sowie der Verfügbarkeit der Äsung für die Rehe
bis hin zum Stress der Tiere runden das Bild ab. Gerade der Stress ist von Bedeutung,
da es nicht ausgeschlossen werden kann, dass durch die „...stetig präsente Störkulis-
se,...“ (Herzog 2013, S. 29.) Rehe zwar viel Energie aufnehmen, aber durch Stress
einen viel höheren Energiebedarf aufweisen. Diese wissenschaftliche Lücke, deren
Schließung mit hohem praktischem Wert für den revierverantwortlichen Jäger verbun-
den ist, wird beispielhaft in der vorliegenden Untersuchung in zwei Modellgebieten
analysiert. Die hier gefundenen Ergebnisse gelten für die beiden unter Punkt 0.3 näher
erläuterten Untersuchungsgebiete. Sie lassen Schlüsse auf andere Gebiete zu, müs-
sen aber für weitere Bereiche in Bayern mit anderen Wald- und Agrarstrukturen über-
prüft werden.
0.2 Ziel und Fragestellung
Ziel der Studie war eine Schätzung des Energiegehaltes und der Qualität der Rehnah-
rung im Jahresverlauf.
Folgende Fragen sollten beantwortet werden:
• Wie stellt sich der verfügbare Energiegehalt der Äsung in den beiden Untersuchungsgebieten im Jahresverlauf dar?
• Wie setzt sich die Rehäsung in den Gebieten zusammen? • Wie stellt sich im Jahresverlauf die Nahrungsaufnahme in beiden Gebieten dar? • Können Rehe auf eine Veränderung der Äsungszusammensetzung und –qualität
reagieren? • Wie ist die Qualität und Verfügbarkeit der Äsung auf den landwirtschaftlichen
Flächen? • Wann haben Rehe im Jahresverlauf die geringsten Fettreserven und das
geringste Körpergewicht? • Unterschieden sich die beiden Gebiete hinsichtlich des Stressniveaus der Rehe?
Durch Schonzeitaufhebung in beiden Gebieten wurden die Proben in den drei Unter-
suchungsjahren 2012 bis 2014 jeweils über 12 Monate gesammelt.

0 Kurzfassung
9
0.3 Die wichtigsten Ergebnisse
Mit verwertbaren 220 Proben überragt dieses Projekt bisherige Projekte durch die ho-
he Probenzahl. Die Aussagekraft für beide Untersuchungsgebiete ist hierdurch wesent-
lich größer als in anderen Arbeiten, und eine Vielzahl von Detailergebnissen konnten
zum ersten Mal statistisch abgesichert werden. Zum ersten Mal wurde in einem Projekt
detailliert der Energiegehalt und die Qualität der Rehäsung im Jahresverlauf über 12
Monate erfasst, basierend auf der real aufgenommenen Äsung der Rehe. Im Sinne
eines systemtheoretischen Ansatzes wurden über den Zeitraum auch die Verfügbarkeit
von Nahrung, die Anpassung des Pansens an die Nahrung, Konditionswerte und
Stress erfasst und in die Bewertung mit aufgenommen. Ebenfalls wurde zum ersten
Mal mit wissenschaftlich modernsten Methoden das Mikrobiom der Rehe analysiert.
Die Untersuchung fand in einem naturnahen Waldökosystem südlich von München
sowie in einem Agrarökosystem bei Eggenfelden in Niederbayern statt. In beiden Ge-
bieten werden Rehe nicht gefüttert. Mit den Standardverfahren der Futtermittelanalytik
– Weender Verfahren, VanSoest Verfahren, Pansensaft Analyse und dem Hohenhei-
mer Futterwerttest, wurden der Energiegehalt und die Qualität der aufgenommenen
Nahrung untersucht. Die Panseninhaltsanalyse gab Aufschluss über die Zusammen-
setzung der Nahrung. Anhand des Gewichtes und des Nierenfettindexes wurde die
Kondition der Tiere bestimmt. Wie gestresst die Rehe in den Untersuchungsgebieten
sind, wurde über Glukokortikoidmetabolite im Rehkot gemessen. Die Anpassung des
Rehes an die Jahreszeiten wurde über das Pansenvolumen sowie die individuelle Art-
zusammensetzung der Pansenflora und -fauna der Tiere ermittelt. Abschließende Ve-
getationsaufnahmen in den Habitaten lieferten einen Einblick über die Nahrungsver-
fügbarkeit im Jahresverlauf.
Den Rehen in unserem landwirtschaftlich geprägten Untersuchungsgebiet steht mit
durchschnittliche 6,29 MJ/kg TS mindestens 0,6 MJ/ kg TS mehr Energie zur Verfü-
gung als den Waldrehen mit etwa durchschnittliche 5,43 MJ/kg TS. Die Unterschiede
sind signifikant. Der Energiegehalt der „Land“-Äsung liegt – mit Ausnahme im Mai –
das ganze Jahr über den Energiewerten des naturnahen Waldgebietes. Am größten
sind die Differenzen zwischen August und November sowie Januar bis April. Anhand

0 Kurzfassung
10
der vorhandenen wissenschaftlichen Literatur wurde ein Energieminimum der Re-
häsung im März/April erwartet. Die niedrigsten Energiegehalte der Äsung fand sich
im Agrarhabitat im Juli und im Waldhabitat im Februar. Anhand der Literatur hätten
wir jedoch ein Energieminimum im März oder April bzw. im Landhabitat im Herbst nach
der Ernte erwartet. Die Energiewerte liegen im Rahmen und über den Energiewer-
ten, die zur Erhaltung von Rehen notwendig sind.
Die Differenz zwischen dem naturnahen Waldgebiet und dem intensiv bewirtschafte-
tem Agrargebiet entsteht durch kohlehydrathaltigen Feld- und Baumfrüchte, die Land-
rehe vom Sommer bis zum Ende des Winters nutzen. Feldfrüchte und Süßgräser auf
den Ackerflächen stellen im Herbst eine wesentliche energiereiche Nahrungsquelle für
Rehe dar. Im Frühjahr stehen den Landrehen deutlich früher als den Waldrehen faser-
arme Kräuter und Süßgräser zur Verfügung. Masttragende Bäume wie Buchen und
Eichen sind in dem landwirtschaftlich geprägten Untersuchungsgebiet mit bäuerlichem
Wald charakteristisch. Die Folge ist, dass Landrehe zwischen Sommer und Frühjahr
signifikant mehr leichtlösliche Kohlehydraten und energiereiche Proteine aufnehmen.
Energetisch betrachtet, haben die Landrehe nicht gemerkt, dass es Winter war.
Dagegen steht den Rehen in naturnahen Waldökosystemen mehr faserreiche Äsung
zur Verfügung, und nur im Mai nehmen sie ähnlich wie die Landrehe durch hohe Roh-
proteinwerte leichter verdauliche Nahrung zu sich. Die Verdaulichkeit der gesamten
„Landäsung“ ist insgesamt für Rehe signifikant höher als die natürliche
Waldäsung. Damit verbunden ist auch der Energiegehalt der Landäsung deutlich
höher als jener der Waldäsung.
Der Rohfasergehalt der Äsung im Waldgebiet schwankte zwischen 24 % und 38 % TS,
jener im Pansen der Landrehe zwischen 21 % und 30 % TS. Diese hohen Rohfaseran-
teile werden in der Literatur mit den Äsungstypen „Intermediär“ oder „Raufutterfresser“
in Verbindung gebracht. Im Winter erreichten die Rohfasergehalte im Durchschnitt
einen Anteil zwischen 25 % TS und 34 % TS. Im naturnahen Waldhabitat sank ihr
Anteil nicht unter 30 % TS.
Waldrehe passen sich an die relativ geringere Nahrungsqualität mit einem 8 %
größeren Pansenvolumen an und durch 25 % mehr Nahrung, die sie aufnehmen.

0 Kurzfassung
11
Damit können sie die geringere Energieausstattung der Äsung kompensieren. Wäh-
rend im Sommer die Nahrungsmenge im Pansen von Land- und Waldrehen nahezu
gleich hoch ist, wiesen Pansen der Waldrehe im Frühjahr etwa 200 g und im Herbst
und Winter mehr als 300 g mehr Panseninhalt auf als jene der Landrehe. Die Unter-
schiede im Herbst und Winter sind signifikant. Schlechtere Nahrungsqualität gleichen
Waldrehe mit einer höheren Nahrungsaufnahme aus. Somit wurde bei den Waldrehen
der jahreszeitliche Gang der Nahrungsaufnahme nicht festgestellt, wie er von Gatter-
versuchen bekannt ist. Auch das Mikrobiom der Waldrehe passt sich an die schwer-
verdauliche Äsung an, indem signifikant mehr Mikroorganismen im Pansen vorkom-
men, die auf Faserzersetzung spezialisiert sind.
Die optimale Anpassung der Waldrehe an faserreiche Nahrung im Winter zeigt sich
auch in dem Vergleich der Energiewerte ermittelt mit Hammelpansensaft und mit Reh-
pansensaft. Der Pansensaft des Rehs wurde im Wald im Januar genommen. Während
mit dem Hammelpansensaft Nahrungsproben aus dem November und Dezember hö-
here Energiewerte lieferten, erbrachte der im Januar gewonnene Rehpansensaft bei
Pansenproben aus dem Januar signifikant höhere Energiewerte.
Unsere Rehe sind genauso wie das Rotwild an winterliche, faserreiche Äsung
besser angepasst als unsere Haustiere und können aus dieser mehr Energie
nutzten.
Auf Grund dieser Fähigkeiten und der häufig falschen Interpretation des Begrif-
fes „Konzentrat-Selektierer“ sollte auf diesen verzichtet werden und wie im eng-
lischen Sprachraum und international wissenschaftlich üblich in Bezug auf das
Reh besser von „Selektierer“ gesprochen werden.
Die hier gefundenen Energiewerte stellen einen Minimumwert für die aufgenommene
Rehäsung dar. Die Proben wurden aus Rehpansen gewonnen, der immer aus einer
Mischung von unverdautem bis hin zu fast vollkommen verdautem Pflanzenmaterial
besteht. Dennoch hat eine Gegenüberstellung der gefundenen Energiewerte mit Ener-
giebedarfswerten belegt, dass den hier analysierten Waldrehen genügend Energie

0 Kurzfassung
12
im gesamten Jahr zur Verfügung steht und Landrehe einen Energieüberschuss in
der Nahrung haben.
Diese gute Energieausstattung der Landrehe wird auch sichtbar in ihrem höheren Kör-
pergewicht (aufgebrochen: Land ∅15,4 kg, Wald 14,8 kg) sowie in einem höheren Nie-
renfettindex. In beiden Habitaten haben die Rehe die beste Kondition im Herbst und
Winter sowie die schlechteste im Sommer. Die niedrigsten Körpergewichte der aus-
gewachsenen Rehe fanden sich in unseren beiden Untersuchungsgebieten nach der
Brunft im September. Zwischen November und April liegen die Rehgewichte in einem
schmalen Rahmen zwischen 15,2 kg und 15,8 kg. Die niedrigeren Energiewerte der
Waldäsung im Februar werden neben der Mehraufnahme an Äsung auch durch
eine gute Kondition und hohes Körpergewicht ausgeglichen.
Anhand der vorhandenen Literatur wurde erwartet, dass Landrehe ein höheres Stress-
niveau haben, was sich in einer höheren Konzentration von Glukokortikoidmetaboliten
zeigen würde. Zwischen Land- und Waldhabitat konnten keine Unterschiede be-
züglich der Glukokortikoidmetabolit-Konzentration (GCM) festgestellt werden.
Geschlecht und Jahreszeit beeinflussen die GCM-Konzentration. So weisen
männliche Tiere höhere Werte auf. Zudem zeigen die Jahreszeiten Frühling und Som-
mer deutlich höhere Werte als Herbst und Winter, was durch das arttypische Verhalten
der Rehe zu erklären ist. Erhöhte Glukokortikoidmetabolit-Konzentration im Feb-
ruar, März oder April, die durch Energiemangel verursacht werden, konnten nicht
gefunden werden. Ein höherer Energiebedarf im Landhabitat auf Grund von
Stress kann nicht abgeleitet werden.
In den drei Projektjahren konnte in beiden Untersuchungsgebieten kein Engpass auf Grund von fehlender Energie in der Nahrung oder ungenü-gender körpereigener Energiereserven bei Rehen festgestellt werden.

Inhaltsverzeichnis
13
Inhalt
Zusammenfassung..........................................................................................................5 Summary.........................................................................................................................6 0 Kurzfassung (König) ................................................................................................... 70.1 Hintergrund ............................................................................................................ 70.2 Ziel und Fragestellung ........................................................................................... 80.3 Die wichtigsten Ergebnisse ................................................................................... 91 Einleitung ................................................................................................................ 171.1 Einführung in die Thematik (Scheingraber / König) ....................................................... 171.2 Stand der Forschung (Scheingraber) ......................................................................... 191.3 Zielsetzung (König, Scheingraber) ................................................................................ 211.4 Fragestellung (Scheingraber) ..................................................................................... 222 Material und Methode ............................................................................................. 232.1 Untersuchungsgebiete (Scheingraber) ...................................................................... 232.2 Untersuchungstier (Scheingraber) ............................................................................. 272.3 Datenerhebung und Probenahme (Scheingraber) ..................................................... 292.3.1 Untersuchungszeitraum. ................................................................................... 292.3.2 Probenahme in den Untersuchungsgebieten ................................................... 302.4 Anzahl der Proben (Scheingraber) ............................................................................. 322.4.1 Aufbereitung Pansen ........................................................................................ 332.5 Analysen zur Qualität der Nahrung (Scheingraber) ................................................... 352.5.1 Analyse fester Pansennhalt .............................................................................. 352.5.1.1 Weender Analyse ........................................................................................... 352.5.1.2 Bestimmung der Rohasche (XA) .................................................................... 362.5.1.3 Bestimmung des Rohfettes (XL) .................................................................... 372.5.1.4 Bestimmung des Rohproteins (XP) ................................................................ 372.5.1.5 Bestimmung der Rohfasern (XF) .................................................................... 392.5.1.6 Bestimmung der Stickstofffreien-Extraktstoffe (NfE) ...................................... 392.5.1.7 VanSoest Analyse .......................................................................................... 402.5.1.8 Bestimmung der Neutral-Detergenzien-Faser (NDF) ..................................... 412.5.1.9 Bestimmung der Säure-Detergenzien-Faser (ADF) ....................................... 422.5.1.10Bestimmung der Säure-Detergenzien-Lignins (ADL) ..................................... 422.5.1.11Bestimmung der Zellulose und Hemizellulose ............................................... 432.5.1.12Bestimmung der Nicht-Faser-Kohlenhydrate (NFC) ...................................... 442.5.2 Analyse Pansensaft .......................................................................................... 442.5.2.1 Ammoniak Analyse ......................................................................................... 472.5.2.2 Milchsäure Analyse ........................................................................................ 48

Inhaltsverzeichnis
14
2.5.2.3 pH-Wert .......................................................................................................... 502.6 Analyse zur Energie der Nahrung (Scheingraber) ..................................................... 512.6.1 Standard HFT (Hammelpansensaft) ................................................................. 522.6.2 HFT mit Rehpansensaft .................................................................................... 552.7 Konditionsbestimmung (Scheingraber) ...................................................................... 562.8 Volumenbestimmung Pansen (Scheingraber) ........................................................... 572.9 Analyse Mikrobiom (Scheingraber) ............................................................................ 582.10 Panseninhaltsanalyse (Mitschke) ........................................................................... 622.11 Pflanzensoziologische Aufnahmen (Mitschke) ....................................................... 642.12 Analyse zum Stress (Scheingraber) ......................................................................... 652.13 Datenauswertung (Scheingraber) ............................................................................ 663 Ergebnisse .............................................................................................................. 673.1 Energiegehalt der Rehnahrung (Scheingraber) ......................................................... 673.1.1 Energiegehalt der Rehnahrung auf der Basis des Standardverfahrens ........... 673.1.2 Vergleich der Energiewerte bei Verwendung von Hammelpansensaft und Rehpansensaft ............................................................................................................. 693.1.3 Fazit: Energiegehalt der Rehnahrung (König) ..................................................... 703.2 Qualität der Rehnahrung (Scheingraber) ................................................................... 713.2.1 Qualität des festen Panseninhaltes .................................................................. 713.2.1.1 Qualität des Raufutters ................................................................................... 713.2.1.2 Detailanalyse der Fasern nach VanSoest ...................................................... 743.2.1.3 Verdaulichkeit ................................................................................................. 773.2.2 Fazit Qualität des festen Panseninhaltes (König) ................................................ 793.2.3 Qualität des Rehpansensaft (Scheingraber) .................................................... 803.2.3.1 Flüchtige Fettsäuren ....................................................................................... 803.2.3.2 Verhältnis Essigsäure zu Propionsäure (C2/C3): ............................................ 823.2.3.3 Ammoniak ....................................................................................................... 833.2.3.4 Laktat .............................................................................................................. 863.2.3.5 pH-Wert .......................................................................................................... 863.2.4 Fazit Pansensaftanalyse (König) ......................................................................... 883.3 Vegetationsanalyse aufgenommener und vorhandener Äsungspflanzen (Mitschke) ................... ................................................................................................................... 903.3.1 Zusammensetzung der Rehnahrung ................................................................ 903.3.1.1 Nahrungsbestandteile im Pansen .................................................................. 903.3.1.2 Aufnahmehäufigkeit durch Rehe .................................................................... 953.3.1.3 Aufgliederung Nadel- und Laubholz ............................................................. 1003.3.2 Vegetationskartierung in den Untersuchungsgebieten ................................... 101

Inhaltsverzeichnis
15
3.3.2.1 Arten und ihre Auftretungsfrequenz in der Vegetation ................................. 1013.3.2.2 Qualität der kartierten Äsungspflanzen ........................................................ 1083.3.2.3 Bestockung der landwirtschaftlich genutzten Flächen im Untersuchungsgebiet „Land“ 1113.3.3 Fazit: Aufnahme der Pflanzen durch Rehe und Qualität sowie Verfügbarkeit der Äsungspflanzen .......................................................................................................... 1123.4 Anpassung des Pansens (Scheingraber) ................................................................. 1133.4.1 Mikrobiom ....................................................................................................... 1143.4.2 Pansenvolumen .............................................................................................. 1173.4.3 Panseninhaltsmenge ...................................................................................... 1183.4.4 Fazit: Anpassung des Pansens (König) ............................................................. 1203.5 Kondition der Untersuchungstiere (Scheingraber) ................................................... 1213.5.1 Gewicht der aufgebrochenen Rehe ................................................................ 1213.5.2 Nierenfettindex ............................................................................................... 1273.5.3 Fazit Kondition (König) ....................................................................................... 1293.6 Stress (Scheingraber) .............................................................................................. 1303.6.1 Konzentration von Glukokortikoidmetaboliten (GCM) in der Rehlosung ....... 1303.6.2 Fazit: Stress (König) .......................................................................................... 1324 Diskussion (König / Scheingraber) ................................................................................. 1334.1 Probennahme .................................................................................................... 1334.2 Die wichtigsten Ergebnisse ............................................................................... 1344.3 Energie der Rehnahrung ................................................................................... 1374.4 Qualität der Rehnahrung ................................................................................... 1434.4.1 Fester Panseninhalt ........................................................................................ 1434.4.2 Pansensaft ...................................................................................................... 1494.5 Nahrungsaufnahme & -Verfügbarkeit ................................................................ 1534.6 Pansenanpassung ............................................................................................. 1554.7 Gewicht und Kondition ....................................................................................... 1594.8 Stress ................................................................................................................ 1615 Literatur ................................................................................................................. 1636 Abbildungsverzeichnis .......................................................................................... 1767 Tabellenverzeichnis .............................................................................................. 1808 Formelverzeichnis ................................................................................................ 1829 Interdisziplinäre Zusammenarbeit und Austausch ................................................ 18310 Veröffentlichungen .............................................................................................. 184

Inhaltsverzeichnis
16
11 Studentische Abschlussarbeiten im Rahmen des Projektes .............................. 18512 Anhang ............................................................................................................... 18612.1 Auftretensfrequenz von Äsungspflanzen im Pansen nach Jahreszeiten und Habitaten .................................................................................................................... 18612.2 Anhang: Auftretensfrequenz von Äsungspflanzen nach Jahreszeiten und Habitaten. ................................................................................................................... 191

1 Einleitung
17
1 Einleitung
1.1 Einführung in die Thematik (Scheingraber / König)
Das Reh (Capreolus capreolus) ist eine der am häufigsten vorkommenden Wildtierar-
ten in unseren Breiten. Seinen ursprünglichen Lebensraum stellten hauptsächlich
Waldlichtungen und Waldrandzonen dar (Reimoser 2005). Die heute von Agrarflächen
durchzogenen oder umgebenden Waldgebiete bieten dem Reh nicht nur mehr Lebens-
raum, sondern führen auch zu einem stark veränderten Nahrungsspektrum. Vorliegen-
de Studien geben bereits Auskunft über den Energiebedarf des Rehs, weitere Unter-
suchungen beschäftigten sich intensiv mit der von Rehen aufgenommenen Vegetation
sowie deren qualitativen/quantitativen Unterschiede in verschiedenen Habitaten und
Jahreszeiten. In dieser Sammlung von Forschungsaktivitäten fehlen jedoch Ansätze,
die den Energiegehalt der tatsächlich aufgenommenen Rehnahrung im gesamten Jah-
resverlauf sowie im Vergleich zwischen intensiv landwirtschaftlichen Gebieten und Ge-
bieten mit überwiegenden Waldstrukturen analysieren, besonders in Anbetracht der
zunehmenden künstlichen Veränderung der Nahrungsnischen.
Die Fähigkeit eines Lebensraumes, Wildtiere zu tragen, ist maßgeblich von dem vor-
handenen Äsungsangebot beeinflusst. Besonders im Winter sinkt die Äsungsquantität
gegenüber Sommer oder Herbst ab (Onderscheka 1999). Dies führt zu einer Reduktion
der Tragfähigkeit des Lebensraums. Intensiv landwirtschaftlich geprägte Gebiete, die in
den vergangenen 20 Jahren flächenmäßig stark zu genommen haben (Fischer 1998),
stehen im Verdacht, speziell im Herbst und Winter Rehen unzureichend Äsung und
hingegen im Sommer unnatürlich viel Äsung zu bieten (Hofmann & Kirsten 1982, Bau-
er 2007, Helm 2015). Je größer die Differenz zwischen der tatsächlich für das Wild
verfügbaren Sommer- und Winteräsung ist, desto größer ist das Wildschadensrisiko im
Wald (Hofmann & Kirsten 1982, Ueckermann 1986, Reimoser 2004, Bauer 2007). Die-
ses wird besonders für das Reh angenommen, da dieses zu den „Konzentrat-
Selektierern“ gezählt wird (Drescher-Kaden 1976, 1984, Hofmann 1989, 2007) und ihm
eine faserarme, leichte verdauliche Äsung zugesprochen wird (Bauer 2007, Cornelis,

1 Einleitung
18
et al. 1999, Dissen & Hartfiel 1985, Drescher-Kaden & Seifelnasr 1976, Drescher-
Kaden & Seifelnasr 1977a, Drescher-Kaden & Seifelnasr 1977b, Drescher-Kaden
1984, Duncan et al. 1998, Fielitz & Albers 1996, Müller 1982, Onderscheka 1999).
Um die Qualität der Rehnahrung umfassend bewerten zu können, ist es nicht nur not-
wendig, Rehäsung auf seine verschiedenen Bestandteile (Rohproteine, -asche, -faser,
-fette, -wasser) hin zu analysieren, sondern den artspezifischen Energiegewinn für Re-
he aus der vorhanden Nahrung im Jahresverlauf zu erfassen. Ein Vergleich verschie-
dener Gebiete zu unterschiedlichen Jahreszeiten, die Berücksichtigung nur einer Jah-
reszeit oder die Analysen möglicher Nahrungskomponenten des Rehs durch künstliche
Selektion oder Fütterung führen unweigerlich zu Fehlinterpretationen. Eine fundierte
Aussage über das saisonale Nahrungs- und Energieangebot eines Habitats kann nur
im Zusammenhang und Vergleich mit dem Nahrungs- und Energieangebot der vorher-
gehenden sowie folgenden Jahreszeit hinsichtlich einer ausreichenden oder mangel-
haften Energieversorgung einer Wildart getroffen werden.
Speziell zum Energiebedarf oder zur Energieverwertung der Äsung durch Rehe wur-
den viele Untersuchungen mit Zoo- oder Gehegetieren durchgeführt (Drozdz & O-
siecki, 1973, Drozdz 1979, Eisfeld, 1974 a + b, Perzanowski 1978, Weiner 1977, Pa-
pageorgiou et al. 1981, Hartfiel et al. 1985, Oslage & Strothmann 1988, Enzinger &
Hartfiel 1998, Behrend 1999). Diese haben bei der Energieberechnung die Verdaulich-
keit von Hauswiederkäuern jener des Rehs gleichgestellt, künstlich selektierte Nahrung
verwendet oder die Nahrung mittels Bombenkalorimeter verbrannt (Papageorgiou et al.
1981), was auf Grund des Wiederkäuer-Verdauungssystems zu falschen und zu hohen
Energiewerten führt.
Diese Wissenslücke kann durch Anwendung des Hohenheimer Futterwerttests (HFT)
für die Analyse der tatsächlich aufgenommenen Nahrung der Rehe geschlossen wer-
den. Mit diesem kann die Energie einer Pansenfüllung eines erlegten Rehes gemessen
werden. Der HFT in Kombination mit dem Weender-/VanSoest Verfahren sowie der
Analyse der flüchtigen Fettsäuren, des Ammoniaks, des pH-Wertes und der Milchsäure
kann jahreszeitlich bedingte Defizite aufzeigen. Aufbauend auf dieser Defizitanalyse
können Rückschlüsse auf Energieengpässe des Rehwilds gezogen werden. Exempla-

1 Einleitung
19
risch sollen die Untersuchungen nicht nur für naturnahe Waldhabitate, sondern auch
für die in unserer Umwelt überwiegenden landwirtschaftlich geprägten Flächen erarbei-
tet werden. Um eine mögliche Differenz zwischen der Verdaulichkeit von Hauswieder-
käuern und Rehen zu aufzudecken, wird zur Energiemessung des Panseninhaltes er-
legter Rehe neben dem Standard-Hohenheimer-Futterwerttest auf Basis von Hammel-
Pansensaft zum ersten Mal dieser auch auf Basis von Reh-Pansensaft durchgeführt.
Für eine umfassende Interpretation der Energiewerte werden von den erlegten Rehen
der physiologische Zustand (Gewichts-, Konditions-, Stressbestimmung) und die sai-
sonale Anpassung des Pansens durch Pansenvolumenbestimmung und Mikrobiom-
analysen ergänzt. Abgerundet wird das Bild der Energieversorgung der Rehe in den
beiden Untersuchungsgebieten durch eine Kartierung der vorhandenen Vegetation,
ihrer Qualität und Verfügbarkeit in den vier Jahreszeiten. Der angesetzte systemtheo-
retische Ansatz umfasst den gesamten Themenkomplex „Ernährungsphysiologie der
Rehe in ihrem Ökosystem“ und beleuchtet diesen aus Sicht des Rehes, des Habitates
unter Einbeziehung menschlicher Rahmenfaktoren wie etwa Störungen.
1.2 Stand der Forschung (Scheingraber)
Auf Grund des weiten Verbreitungsareales dieser Tierart ist die Literatur zu Rehwild
sehr umfangreich. Allein zur Nahrungswahl des Rehes in Europa fanden Cornelis et al.
(1999) 37 Literaturquellen, wobei sie nur englischsprachige Literatur berücksichtigten.
Für die vorliegende Studie sind folgende Themenkomplexe relevant:
Nach Tixier & Duncan (1996) zeigte sich anhand verschiedenster Nahrungsanalysen,
dass Rehwild als Äsung europaweit 305 verschiedene Pflanzenarten aufnimmt. Die
Palette reicht von den verschiedensten Baum- und Straucharten über Zwergsträucher,
Pilze und Farne bis hin zu Gräsern und Kräutern. Landwirtschaftliche Nutzpflanzen
werden ebenso von Rehen angenommen. In vielen Gebieten bilden im Winter Brom-
beere und Himbeere einen wesentlichen Bestandteil der Rehnahrung (Anke et al.
2007, Barancekova et al. 2010, Bubenik 1984, Cornelis et al. 1999, Clauss 2010, Dis-
sen & Hartfiel 1985, Drescher-Kaden & Seifelnasr 1976, Duncan et al. 1998, Stubbe
1996, Tixier & Duncan 1996, Ueckermann 1996, Fielitz & Albers 1996, Klansek 2004,

1 Einleitung
20
2005). Im Vergleich zu anderen Wiederkäuern ist das Reh sehr flexibel in seiner Nah-
rungswahl. Es passt sich in der Regel bezüglich dieser an die vorhandene Vegetation
an (Anke et al. 2007, Drescher-Kaden & Seifelnasr 1976, Duncan et al. 1999, Onder-
scheka 1999, Tixier & Duncan 1996). Manche Autoren denken in diesem Zusammen-
hang sogar über die Bezeichnung „Konzentrat-Selektierer“ als unzutreffend nach (Anke
et al 2007, Tixier & Duncan 1996). Entsprechend der sehr unterschiedlichen Nutzung
der Vegetation durch Rehe variieren die Anteile an Rohproteinen, -fasern, -fetten, -
asche und -wasser.
Dabei stellt sich die Frage, inwieweit die aufgenommene Äsung durch das Reh auch in
Energie umgesetzt werden kann, da allein die Zusammensetzung der Pansenflora und
-fauna dem Reh eine weitestgehende Aufspaltung der Rohfasern (Zellulose) ermög-
licht. Studien zur Pansenflora und –fauna des Rehwildes sind nur rudimentär vorhan-
den und entsprechen nicht den neuesten molekularbiologischen Standards (Enzinger
1989, Drescher-Kaden & Seifelnasr 1977a). Gerade die Differenzierung des Pansen-
mikrobioms auf Basis von genetischen Methoden wird wichtige Einblicke und Erklärun-
gen über die Rehwildverdauung liefern.
Der Energiebedarf der Rehe wurde in einer Vielzahl von Fütterungsversuchen an Zoo-
und/oder Gattertieren durchgeführt (Drozdz & Osiecki 1973, Drozdz 1979, Eisfeld
1974a, b, Perzanowski 1978, Weiner 1977, Papageorgiou et al. 1981, Hartfiel et al.
1985, Oslage & Strothmann 1988, Enzinger & Hartfiel1998, Behrend 1999). Aus die-
sen geht hervor, dass Rehe je nach Jahreszeit und Aktivität einen Erhaltungsenergie-
bedarf zwischen ca. 4,3 ME MJ / Reh / Tag im Winter und ca. 5,1 ME MJ / Reh / Tag
im Sommer (Drozdz & Osiecki 1973, Weiner 1977, Hartfiel et al. 1985, Oslage &
Strothmann 1988, Stubbe 1997, Onderscheka 1999, Kamphues et al. 2008) aufweisen.
Der Energiebedarf steigt bei Bewegung um 100 % und bei Stress sogar um 200 % an
(Weiner 1977, Hartfiel et al. 1985, Bubenik 1984, Oslage & Strothmann 1988, Stubbe
1997, Onderscheka 1999). In all diesen Untersuchungen erfolgte die Energiewertmes-
sungen anhand zahmer Gehegetiere, denen unbegrenzt energiereiches Futter zur Ver-
fügung stand, die Angaben für den Energiebedarf bei abweichendem Verhalten beruh-
ten auf gutachterlichen Schätzungen. Auch wurde stets ein Futtermittel oder künstli-
che, durch die Autoren selektierte Nahrung zur Energiemessung verwendet und nicht

1 Einleitung
21
die real von Rehen aufgenommene Äsung. In all diesen Arbeiten wurde übereinstim-
mend festgestellt, dass unser Wild auch bei unbegrenztem Futterangebot hinsichtlich
der Futteraufnahme einem jahreszeitlichen Rhythmus folgt (Bubenik & Lochmann
1956, Drozdz 1979, Dissen & Hartfiel 1985).
Dieser jahreszeitliche Rhythmus der Futteraufnahme und Futterselektion könnte be-
sonders gut bei wildlebenden Tieren ohne Fütterung untersucht werden, was in dieser
hier durchgeführten Arbeit der Fall ist.
1.3 Zielsetzung (König, Scheingraber)
Ziel dieser Arbeit ist es, Veränderungen des Energiegehaltes und der Qualität der
Rehnahrung im Jahresverlauf über 12 Monate zu erfassen. Basis hierfür wird frisches
Pansenmaterial sein, um die tatsächlich von Rehen aufgenommene Äsung mit ihrem
Energiegehalt zu analysieren.
Diese Analyse wird in zwei Untersuchungsgebieten durchgeführt, wobei eines durch
forstliche Flächen und das andere durch moderne intensive Landwirtschaft geprägt ist.
Hierzu werden aus beiden Gebieten über das Jahr verteilt Pansenproben gewonnen
und mit den Standardverfahren der Futtermittelanalysen und der Molekularbiologie
untersucht. Auf Grund der relativ kurzen Verweildauer des Panseninhaltes beim Reh
(Reimoser 2005), wird mittels der Pansenproben die erst kurz zuvor aufgenommene
Nahrung erfasst. Zusätzliche Mikrobiomanalysen, Konditions- und Stressbestimmung
der Tiere sowie die Erfassung der Nahrungsverfügbarkeit in den Habitaten mittels Ve-
getationsaufnahmen ermöglichen einen größeren und übergeordneten Einblick in den
Verfassungszustand der Untersuchungstiere und den damit in Zusammenhang ste-
henden abiotischen Faktoren. Diese können mögliche Ab- oder Zunahme des Nah-
rungs-/ Energiebedarfs der Tiere bewirken.

1 Einleitung
22
1.4 Fragestellung (Scheingraber)
In der durchgeführten Studie werden folgende Fragen bearbeitet:
• Wie unterscheiden sich der Energiegehalt und die Qualität der Nahrung von
Rehen in unterschiedlich geprägten Habitaten im Jahresverlauf?
• Wie setzt sich die Nahrung von Rehen in den unterschiedlich geprägten Habita-
ten zu verschiedenen Jahreszeiten zusammen?
• Wie ist die Nahrungsverfügbarkeit in den unterschiedlich geprägten Habitaten
im Jahresverlauf?
• Findet eine physiologische Anpassung des Pansens an die verschiedenen Ha-
bitate und Jahreszeiten statt?
• Wie ist die Kondition der Untersuchungstiere im Jahresverlauf in den verschie-
denen Habitaten?
• Gibt es Unterschiede zwischen den beiden Habitaten hinsichtlich der Beunruhi-
gung und somit unterschiedliche Niveaus der Stresssignale der Untersuchungs-
tiere?

2 Material und Methode
23
2 Material und Methode
2.1 Untersuchungsgebiete (Scheingraber) Für die Durchführung der Studie wurden zwei unterschiedlich geprägte Untersu-
chungsgebiete ausgewählt. Ein Gebiet sollte überwiegend forstlich geprägte Waldflä-
chen repräsentieren, das zweite Gebiet ein stark landwirtschaftlich genutztes Habitat.
Abbildung 1 zeigt die Lage der ausgewählten Flächen. Die Untersuchungsflächen wur-
den nach folgenden Kriterien ausgewählt:
• Hoher Waldanteil (FO) oder hoher Anteil landwirtschaftlicher Flächen (LA)
• Rehwildvorkommen ohne starke Nahrungskonkurrenz
• Keine Fütterung
• Etwa vergleichbare Region, Fläche und Klima
• Gute Erreichbarkeit
• Unterstützung vor Ort
Abbildung 1: Übersichtskarte Studienflächen (links rot: Waldflächen, rechts rot: Landflä-chen), Quelle: Bayern Maps, bearb. Scheingraber

2 Material und Methode
24
Gebiet „Wald“
Das Studiengebiet „Wald“ (im weiteren Verlauf als FO bezeichnet) beinhaltet folgende
drei Flächen: Wadlhausergräben (448 ha, 47°59'3.41"N, 11°24'24.23"E), Breit-
wies/Schorn (136 ha, 48° 0'22.14"N, 11°28'0.11"E) und den Forstenrieder Park (2088
ha, 48° 1'14.27"N, 11°26'19.29"E). Die Gebiete (Abbildung 2) liegen südlich von Mün-
chen auf einer Höhe von 635 m bis 665 m. Mehr als 90 % Flächen sind mit Wald be-
stockt und weisen einen Baumartenanteil von 70 % Fichte (Picea abies), 20 % Buche
(Fagus sylvatica) und 2 % Tanne (Abies alba) auf. Die restliche Fläche teilt sich auf in
Wildäcker und Forststraßen. Die Teilflächen befinden sich im Wuchsbezirk 13.2
Münchner Schotterebene und 14.4. Oberbayer. Jungmoräne und Molassevorberge.
Die Jahresmitteltemperatur beträgt 8 °C (Arbeitskreis Standortserkundung 1985). Die
Flächen sind Eigenjagden der Bayerischen Staatsforsten AöR.
Abbildung 2: Untersuchungsgebiet FO, GIS Karte, Bayern Atlas, Quelle: bearb. Schein-graber.

2 Material und Methode
25
Gebiet „Land“
Das zweite Studiengebiet „Land“ (folgend als „LA“) beinhaltet drei Gemeinschaftsjag-
den mit einer Gesamtfläche von 2240 ha im Landkreis Rottal-Inn in Ostbayern
(Abbildung 3). Dieses beinhaltet die Gemeinschaftsjagdreviere Rampoldstetten (730
ha, 48°30'23.25"N, 12°32'37.73"E), Vordersarling (755 ha, 48°24'16.29"N,
12°40'38.47"E) und Angerstorf - Lohbruck (755 ha, 48°22'31.19"N, 12°49'17.26"E).
Abbildung 3: Untersuchungsgebiet LA, GIS Karte, Bayern Atlas, Quelle: bearb. Schein-graber.
Die Gebiete liegen 450-500 m über dem Meeresspiegel und haben einen Waldanteil
von 27 – 34 %. Das dortige Habitat ist hauptsächlich von landwirtschaftlichen Flächen
geprägt und wird von einzelnen Waldparzellen mit Flächen von 22 - 30 ha durchzogen.
Die Waldparzellen sind charakterisiert durch 75 - 80 % Fichte (Picea abies), 5 -15 %
Rampoldstetten Vordersarling
Angerstorf-Lohbruck
2 Material und Methode
26
Kiefer (Pinus sylvestris), 2 - 5 % Tanne (Abies alba), 1 - 2 % Buche (Fagus sylvatica)
sowie einem kleinen Anteil an Edellaubhölzern.
Der überwiegende landwirtschaftliche Anbau während der Untersuchungszeit bestand
in Grünwiesen (33 - 40 %), Winterweizen (16 - 21 %), Silomais (11 - 27 %), Körnermais
(3 – 10 %), Gerste (5 - 8 %) und Kleegras (3 - 10 %). Die Reviere liegen im Wuchsbe-
zirk 12.9 Niederbayerisches Tertiärhügelland mit einer Jahresmitteltemperatur von 8,5
°C (Arbeitskreis Standortserkundung 1985).
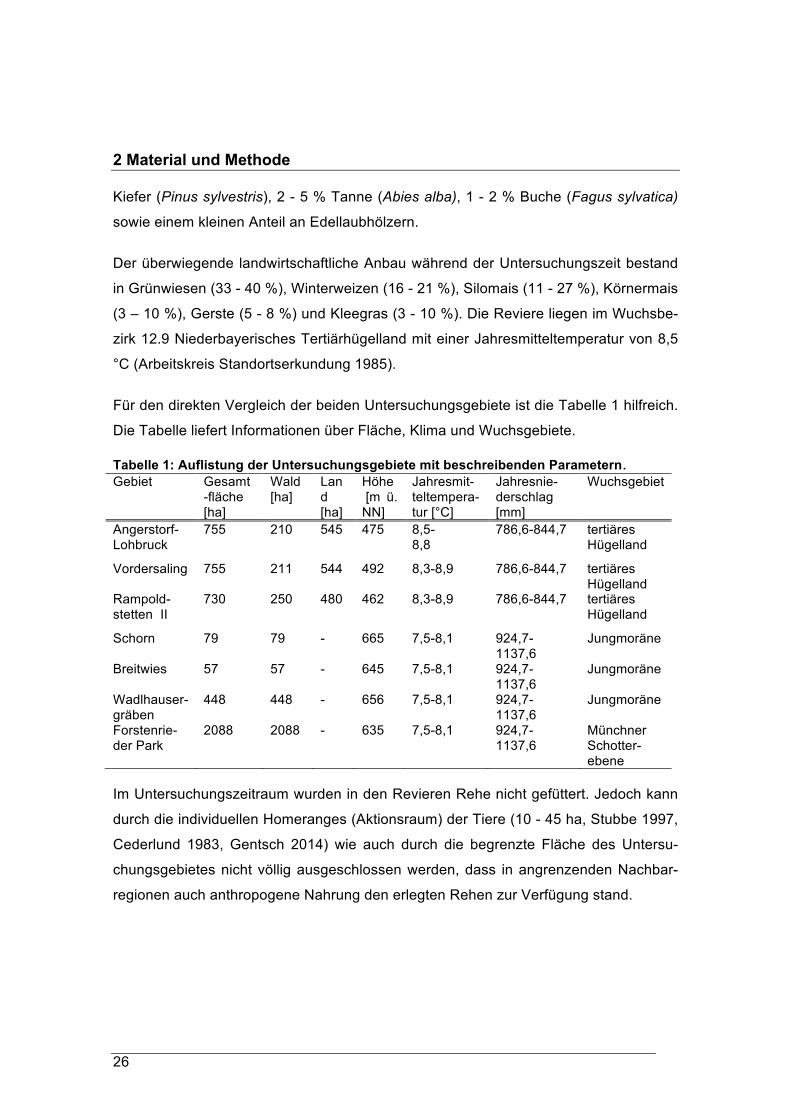
Für den direkten Vergleich der beiden Untersuchungsgebiete ist die Tabelle 1 hilfreich.
Die Tabelle liefert Informationen über Fläche, Klima und Wuchsgebiete.
Tabelle 1: Auflistung der Untersuchungsgebiete mit beschreibenden Parametern. Gebiet Gesamt
-fläche [ha]
Wald [ha]
Land [ha]
Höhe [m ü. NN]
Jahresmit-teltempera-tur [°C]
Jahresnie-derschlag [mm]
Wuchsgebiet
Angerstorf-Lohbruck
755 210 545 475 8,5-8,8
786,6-844,7 tertiäres Hügelland
Vordersaling
755 211 544 492 8,3-8,9 786,6-844,7 tertiäres Hügelland
Rampold-stetten II
730 250 480 462 8,3-8,9 786,6-844,7 tertiäres Hügelland
Schorn
79 79 - 665 7,5-8,1 924,7-1137,6
Jungmoräne
Breitwies
57 57 - 645 7,5-8,1 924,7-1137,6
Jungmoräne
Wadlhauser- gräben
448
448 - 656 7,5-8,1 924,7-1137,6
Jungmoräne
Forstenrie-der Park
2088 2088 - 635 7,5-8,1 924,7-1137,6
Münchner Schotter-ebene
Im Untersuchungszeitraum wurden in den Revieren Rehe nicht gefüttert. Jedoch kann
durch die individuellen Homeranges (Aktionsraum) der Tiere (10 - 45 ha, Stubbe 1997,
Cederlund 1983, Gentsch 2014) wie auch durch die begrenzte Fläche des Untersu-
chungsgebietes nicht völlig ausgeschlossen werden, dass in angrenzenden Nachbar-
regionen auch anthropogene Nahrung den erlegten Rehen zur Verfügung stand.

2 Material und Methode
27
2.2 Untersuchungstier (Scheingraber)
Das Europäische Reh (Capreolus capreolus) gehört zur Ordnung der Paarhufer (Artio-
dactyla), zur Unterordnung der Wiederkäuer (Ruminantia) und zur Familie der Hirsche
(Cervidae). Als eine von neun Gattungen (Capreolus) der Unterfamilie der Trughirsche
(Odocoilinae) kommt das Reh als kleinster und häufigster Vertreter der Wildwieder-
käuer in Europa vor. Die Gattung Capreolus umfasst zwei Arten, Capreolus capreolus,
das europäische Reh, sowie Capreolus pygargus, das sibirische Reh. Das europäische
Reh ist in seiner West-Ost-Verbreitung/Ausdehnung in ganz Europa bis zum Uralge-
birge anzutreffen (Stubbe 1997). Das Reh gehört zu den frühen ursprünglichen Hirsch-
arten, den Drückern und Schlüpfern, was sich in seinem Körperbau und Verhalten äu-
ßert.
Das Reh bevorzugt besonders gebüsch- und unterwuchsreiche lichte Laub- bis Laub-
Nadelholzbestände. Grenzlinien sind für Rehe wichtige Habitatelemente. An die Ver-
änderung seiner Habitate in Nadelholz dominierte Wälder und intensiv bewirtschaftete
Agrarlandschaften hat sich das Reh gut angepasst (Andersen et al. 1998).
Auf Grund der selektiven Nahrungswahl und des häufig hohen Proteingehalts der
Äsungspflanzen zählen die meisten Autoren Rehe zu den „Konzentrat-Selektierern“
(Drescher-Kaden 1984, Hofmann 1982, 1989, 2007). Andere wie Anke et al. (2007)
ziehen auf Grund der Fähigkeit von Rehen, auch relativ hohe Faseranteile gut verdau-
en zu können, den Begriff „Selektierer“ vor, wie er auch im englischsprachigen Raum
verwendet wird. Im Vergleich zu anderen Wildwiederkäuern und entsprechend seiner
Körpergröße weist das Rehwild mit einem durchschnittlichen Pansen-Fassungs-
vermögen von ca. 3 – 6 Liter (Feustel 1967, Hofmann 1976) einen der kleinsten Wie-
derkäuerpansen auf. Durch das relativ geringe Fassungsvermögen folgt das Reh ei-
nem über den Tag verteilten Äsungsrhythmus mit ca. 8 bis 11 Perioden (Bubenik 1960,
Klötzli 1965, Berg 1978). Mithilfe des Wiederkäuer-Verdauungssystems ist es dem
Rehwild möglich, auch schwer lösliche Pflanzenstoffe wie Zellulose aufzuspalten. Dies
gelingt ihnen mithilfe einer darin enthaltener Vielzahl von Mikroorganismen mit der Fä-
higkeit Zellulose aufzuspalten (Sommer 2004).
2 Material und Methode
28
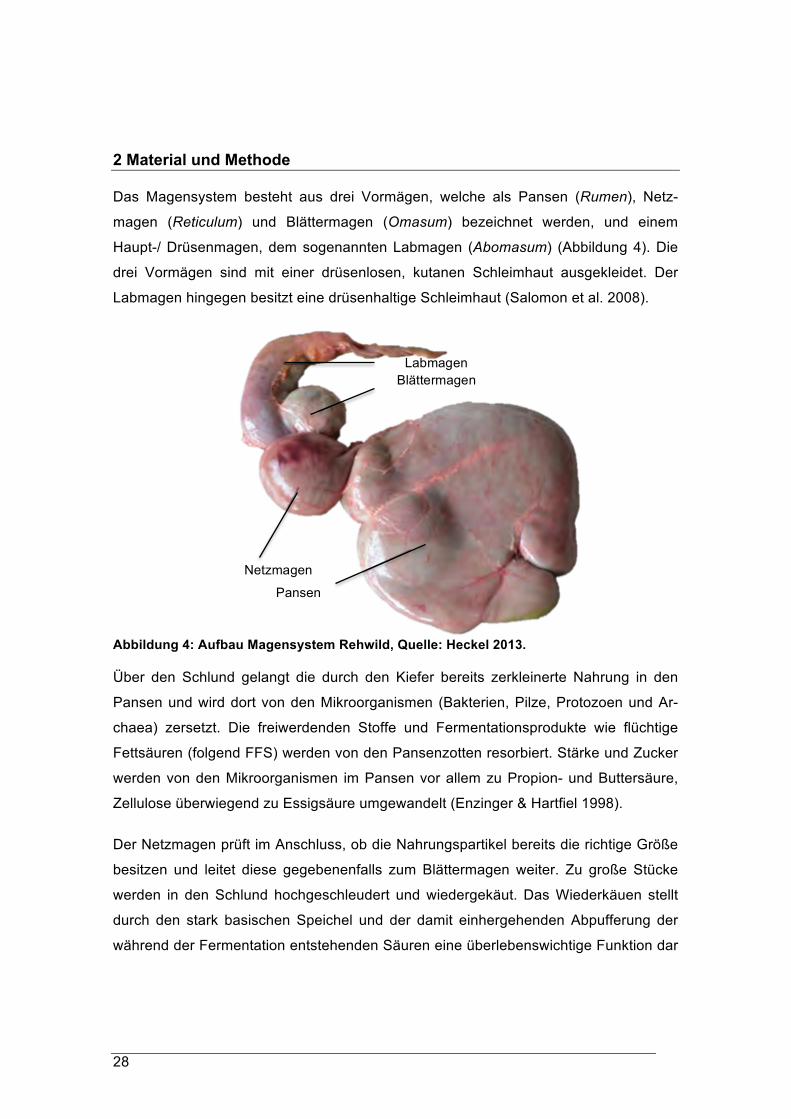
Das Magensystem besteht aus drei Vormägen, welche als Pansen (Rumen), Netz-
magen (Reticulum) und Blättermagen (Omasum) bezeichnet werden, und einem
Haupt-/ Drüsenmagen, dem sogenannten Labmagen (Abomasum) (Abbildung 4). Die
drei Vormägen sind mit einer drüsenlosen, kutanen Schleimhaut ausgekleidet. Der
Labmagen hingegen besitzt eine drüsenhaltige Schleimhaut (Salomon et al. 2008).
Abbildung 4: Aufbau Magensystem Rehwild, Quelle: Heckel 2013.
Über den Schlund gelangt die durch den Kiefer bereits zerkleinerte Nahrung in den
Pansen und wird dort von den Mikroorganismen (Bakterien, Pilze, Protozoen und Ar-
chaea) zersetzt. Die freiwerdenden Stoffe und Fermentationsprodukte wie flüchtige
Fettsäuren (folgend FFS) werden von den Pansenzotten resorbiert. Stärke und Zucker
werden von den Mikroorganismen im Pansen vor allem zu Propion- und Buttersäure,
Zellulose überwiegend zu Essigsäure umgewandelt (Enzinger & Hartfiel 1998).
Der Netzmagen prüft im Anschluss, ob die Nahrungspartikel bereits die richtige Größe
besitzen und leitet diese gegebenenfalls zum Blättermagen weiter. Zu große Stücke
werden in den Schlund hochgeschleudert und wiedergekäut. Das Wiederkäuen stellt
durch den stark basischen Speichel und der damit einhergehenden Abpufferung der
während der Fermentation entstehenden Säuren eine überlebenswichtige Funktion dar
Labmagen Blättermagen
Netzmagen
Pansen

2 Material und Methode
29
(Deutz 2006). Um den Vorgang des Wiederkäuens zu gewährleisten, benötigt die Nah-
rung des Rehwildes einen ebenso großen Anteil an Fasern wie an Protein (Kurt 1970).
Gelangen die Nahrungspartikel in den Blättermagen, so resorbiert dieser die restlichen
Fermentationsprodukte und entzieht dem Nahrungsbrei Wasser. Der Rest der Digesta
gelangt in den Labmagen. Dort setzt die Drüsenschleimhaut körpereigene Enzyme zur
Aufspaltung bisher unverdauter Nahrungsbestandteile frei. Die mit der Digesta mit-
transportierten Mikroorganismen werden im Labmagen durch die vorhandene Salzsäu-
re abgetötet, das Mikrobeneiweiß wird aufgespalten und resorbiert.
Ein weiteres und wichtiges Merkmal des Rehpansens ist die schnelle Durchflussrate
und kurze Retentionszeit / Verweildauer (ca. 12 bis 14 Std) der Ingesta (Behrend 1999,
Onderscheka 1999). Dadurch wird es zu einem oszillierenden „Fresszyklus“ gezwun-
gen (Hofmann 1989). Hierdurch befindet sich im Pansen der Rehe eine relativ homo-
gene Zusammensetzung von wenig bis stark anverdauten Nahrungsbestandteilen.
2.3 Datenerhebung und Probenahme (Scheingraber)
2.3.1 Untersuchungszeitraum.
Die Probenahme wurde im November 2011 begonnen und im Oktober 2014 abge-
schlossen. Die Untersuchungstiere wurden je nach Zeitpunkt ihrer Erlegung den Mona-
ten und einer Jahreszeit zugeordnet. Es wurde hierfür die meteorologische Einteilung
(Tabelle 2) der Jahreszeiten verwendet.
Tabelle 2: Meteorologische Einteilung der Jahreszeiten zur Einordnung der Untersu-chungstiere. Meteorologische Jahreszeit Monate der Probenherkunft
Frühling März, April, Mai
Sommer Juni, Juli, August
Herbst September, Oktober, November
Winter Dezember, Januar, Februar

2 Material und Methode
30
2.3.2 Probenahme in den Untersuchungsgebieten
Alle Rehe wurden im Rahmen des behördlich festgesetzten Abschussplanes erlegt.
Im Rahmen dieser fand die Erlegung der Untersuchungstiere durch jagdausübungsbe-
rechtigte Personen der Arbeitsgruppe Wildbiologie und Wildtiermanagement der TUM,
Privatpersonen oder Berufsjäger statt. Der Hauptanteil der Tiere wurde während der
regulären Jagdzeit auf Ansitz-, Pirsch oder Drückjagden erlegt. Um den gesamten Jah-
resverlauf berücksichtigen zu können, wurden Rehe auch innerhalb der Schonzeit auf
Grund einer Schonzeitaufhebung der entsprechenden Landratsämter (Landratsamt
Starnberg, Landratsamt Rottal-Inn, Schonzeitaufhebung, Bayerisches Jagdgesetz,
Artikel 33) erlegt.
Unmittelbar nach der Erlegung wurden die Rehe aufgebrochen und die Organe ent-
nommen. Verwendet wurden nur Pansen, die keinerlei Verletzung durch den Schuss
oder das Aufbrechen aufweisen. Auf Grund von Krankheit musste keines der Tiere als
Proband ausgeschlossen werden. Den Rehen wurde der Pansen, beide Nieren mit
umliegendem Fettgewebe und Kot aus dem Rektum entnommen. Dabei wurde vor
Entnahme des Verdauungstraktes aus dem Tierkörper oberhalb des Pansens die
Speiseröhre (Oesophagus/Schlund) sowie der Übergang vom Labmagen (Abomasum)
zum Zwölffingerdarm (Duodenum) mittels Kabelbinder abgebunden, um das Heraus-
fließen/Verlust von Digesta zu vermeiden. Das Gewicht der Rehe wurde aufgebrochen
und unaufgebrochen vor Ort ermittelt. Für die Auswertungen wurde nur das aufgebro-
chene Gewicht verwendet.
Die Proben wurden unmittelbar nach der Entnahme bei -20 °C in mobilen Gefriertruhen
tiefgefroren und bis zur weiteren Verarbeitung bei -20 °C im Labor gelagert. Von jedem
Tier wurden die Parameter Datum, Habitat, Geschlecht, Alter (Juvenil = Kitze, Subadul-
te = Einjährige, Adulte = ab Vollendung des zweiten Lebensjahres), Schütze, Erle-
gungsort, Jagdart, Uhrzeit der Erlegung, Zeitpunkt der Probenahme und Zeitpunkt des
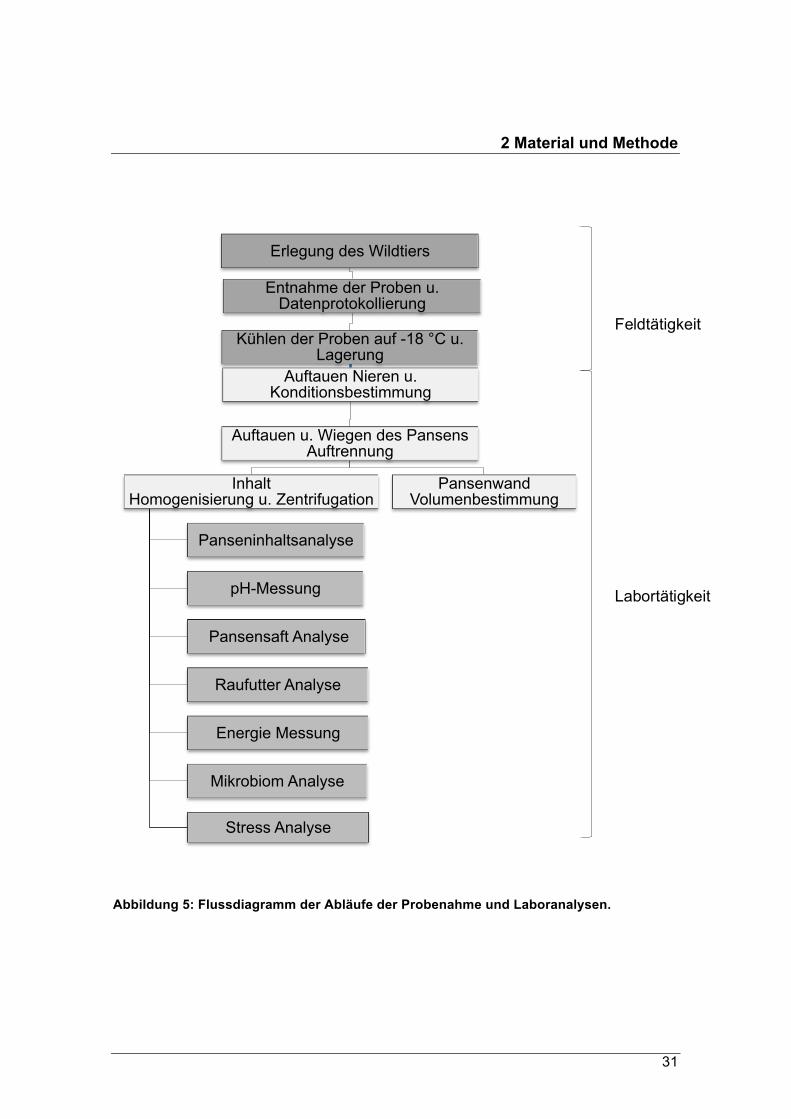
Einfrierens auf Protokollblättern (siehe Protokollblatt im Anhang) festgehalten. Abbil-
dung 5 stellt den genauen Ablauf der Vorgehensweise des Probenahme- und Analy-
senverfahrens dar.
2 Material und Methode
31
Abbildung 5: Flussdiagramm der Abläufe der Probenahme und Laboranalysen.
Erlegung des Wildtiers
Entnahme der Proben u. Datenprotokollierung
Kühlen der Proben auf -18 °C u. Lagerung
Auftauen Nieren u. Konditionsbestimmung
Auftauen u. Wiegen des Pansens Auftrennung
Inhalt Homogenisierung u. Zentrifugation
Panseninhaltsanalyse
pH-Messung
Pansensaft Analyse
Raufutter Analyse
Energie Messung
Mikrobiom Analyse
Stress Analyse
Pansenwand Volumenbestimmung
Feldtätigkeit
Labortätigkeit
2 Material und Methode
32
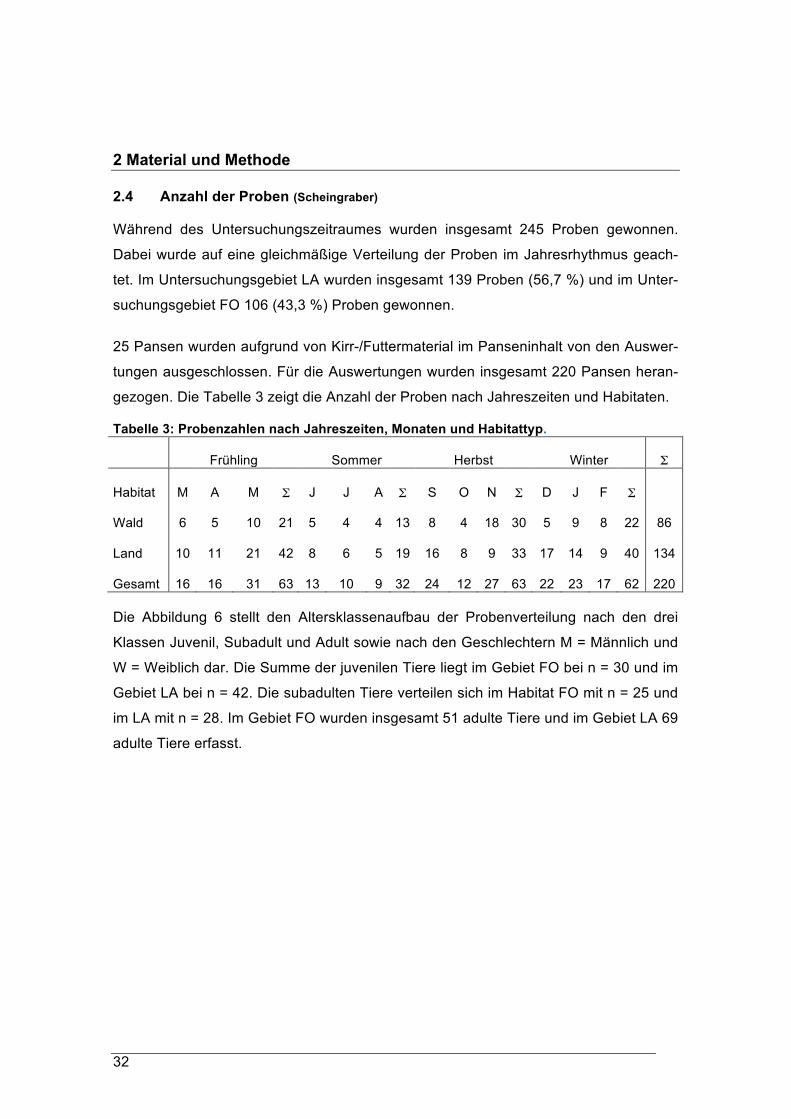
2.4 Anzahl der Proben (Scheingraber)
Während des Untersuchungszeitraumes wurden insgesamt 245 Proben gewonnen.
Dabei wurde auf eine gleichmäßige Verteilung der Proben im Jahresrhythmus geach-
tet. Im Untersuchungsgebiet LA wurden insgesamt 139 Proben (56,7 %) und im Unter-
suchungsgebiet FO 106 (43,3 %) Proben gewonnen.
25 Pansen wurden aufgrund von Kirr-/Futtermaterial im Panseninhalt von den Auswer-
tungen ausgeschlossen. Für die Auswertungen wurden insgesamt 220 Pansen heran-
gezogen. Die Tabelle 3 zeigt die Anzahl der Proben nach Jahreszeiten und Habitaten.
Tabelle 3: Probenzahlen nach Jahreszeiten, Monaten und Habitattyp.
Frühling Sommer
Herbst
Winter Σ
Habitat M A M Σ J J A
Σ S O N
Σ D J F
Σ
Wald 6 5 10
21 5 4 4
13 8 4 18
30 5 9 8
22 86
Land 10 11 21
42 8 6 5
19 16 8 9
33 17 14 9
40 134
Gesamt 16 16 31
63 13 10 9
32 24 12 27
63 22 23 17
62 220
Die Abbildung 6 stellt den Altersklassenaufbau der Probenverteilung nach den drei
Klassen Juvenil, Subadult und Adult sowie nach den Geschlechtern M = Männlich und
W = Weiblich dar. Die Summe der juvenilen Tiere liegt im Gebiet FO bei n = 30 und im
Gebiet LA bei n = 42. Die subadulten Tiere verteilen sich im Habitat FO mit n = 25 und
im LA mit n = 28. Im Gebiet FO wurden insgesamt 51 adulte Tiere und im Gebiet LA 69
adulte Tiere erfasst.

2 Material und Methode
33
Abbildung 6: Probenverteilung nach Habitaten, Altersklassen und Geschlecht, W = Weib-lich, M = Männlich.
Das Geschlechterverhältnis des gesamten Datensatzes liegt bei 50,9 % männlichen zu
49,1 % weiblichen Rehen (♂ 1: ♀ 0,96). Davon stammen 63 männliche Tiere und 71
Weibchen aus dem LA Gebiet sowie 49 Männchen und 37 Weibchen aus dem FO Ge-
biet.
2.4.1 Aufbereitung Pansen
Zur Aufbereitung wurden die Pansen ca. 6 – 8 Std. vor der Bearbeitung aufgetaut. Im
aufgetauten Zustand wurden vom Pansen Lab- und Blättermagen entfernt. Pansen und
Netzmagen wurde mittels einer Analysewaage im vollen Zustand gewogen. Danach
wurde die Ingesta entnommen. Hierfür wurde der Pansen vom Schlund bis zum Pan-
sensack aufgeschnitten und umgestülpt. Um den gesamten Ingesta inklusive dem in
den Zotten verbleibenden Pansensaft zu gewinnen, wurde die Pansenwand ausge-
drückt. Danach wurde der Pansen ohne Inhalt gewogen, um mittels der Differenz den
Inhalt [g] zu berechnen. Die Digesta wurden in eine Schüssel gegeben, um sie durch
Verrühren zu homogenisieren. Des Weiteren wurden 150 g des Panseninhaltes für
eine Inhaltsanalyse und ca. 250 g als Rückstellprobe entnommen, in Weithalsflaschen
gefüllt und bis zur weiteren Verarbeitung eingefroren.
Altersklassen
Adult
Subadult
Juvenil
Anzahl60 40 20 0
Altersklassen
Adult
Subadult
Juvenil
6 040200
Geschlecht
MW
44
25
39
65
23
24
Seite 1

2 Material und Methode
34
Für die weitere Aufbereitung wurden jeweils acht Zentrifugen-Röhrchen zu maximal 70
g befüllt und bei 4460 g für 15 Minuten zentrifugiert. Die Befüllung der Röhrchen erfolg-
te auf 0,01 g genau. Anschließend wurde eine Trennung der festen und flüssigen Pan-
senbestandteile durchgeführt (Abbildung 7 ).
Abbildung 7: Trennung der Bestandteile durch Zentrifugation, rechts: Zentrifugenröhren nach der Zentrifugation, links: Überstand/Pansensaft in Weithalsgefäß, Raufutteranteil in Petrischale ausgestrichen, Quelle: Scheingraber
Verwendet wurde hierfür die Zentrifuge Heraeus Suprafuge (Typ 22). Der Über-
stand/Pansensaft wurde in eine beschriftete Weithalsflasche gefüllt und bis zur weite-
ren Analyse bei -20 °C gelagert.
Abbildung 8: Panseninhalte Rohmasse (links) und in Petrischalen gefriergetrocknet, beim Rückwiegen und vor dem Mahlen (rechts) , Quelle: Scheingraber
Der sich am Boden der Röhrchen absetzende Raufutteranteil wurde in Petrischalen
ausgebreitet, um eine optimale Gefriertrocknung zu gewährleisten. Die Gefriertrock-
nung erfolgte bei einer Eiskondensatortemperatur von -52 °C und einer Stellflächen-
temperatur von +30 °C für 72 Stunden in einer Gefriertrocknungsanlage. Nach der Ge-
friertrocknung wurden ebenfalls die Petrischalen mit dem Raufutteranteil zurückgewo-
gen, um den Wasserverlust des Rohstoffes bestimmen zu können (Abbildung 8). Der

2 Material und Methode
35
gefriergetrocknete Raufutteranteil wurde für die weiterführenden Analysen mittels einer
elektrischen Mühle auf 1 mm ∅ Kerngröße gemahlen.
2.5 Analysen zur Qualität der Nahrung (Scheingraber)
Die Analysen fanden überwiegend am Wissenschaftszentrum Weihenstephan der
Technischen Universität Münchens statt und wurden in den Laborräumen der Arbeits-
gruppe Wildbiologie und Wildtiermanagement des Lehrstuhls für Tierernährung und
der Bioanalytik durchgeführt.
2.5.1 Analyse fester Pansennhalt
Um die Qualität der Nahrung zu untersuchen, ist es unabdingbar, die einzelnen Nah-
rungskomponenten bzw. Rohnährstoffe, ihre Zusammensetzung und ihre mengenmä-
ßige Verteilung zu ermitteln. Die vorliegende Studie bediente sich hierfür der Weender
und VanSoest Analysen (nach VDLUFA Norm).
2.5.1.1 Weender Analyse
Die von Henneberg und Strohmann entwickelte Weender Analyse ist eine der ältesten
Verfahren zur Untersuchung der Inhaltsstoffe von Futtermitteln (Kirchgessner et al.
2008) und stellt eine sogenannte Konventionsanalyse dar (Schuldt und Dinse 2010).
Bestimmt werden die Trockensubstanz (TS) sowie Nährstoffgruppen Rohasche (XA),
Rohfaser (XF), Rohprotein (XP), Rohfett (XL) und stickstofffreie-Extraktstoffe (NfE). Die
Rohnährstoffe werden prozentual im Bezug zur Trockensubstanz angegeben und ana-
lytisch ermittelt. Ausnahme bilden die stickstofffreien-Extraktstoffe. Sie werden rechne-
risch ermittelt.
Nach Definition ist die Trockenmasse der nicht flüchtige Anteil eines Futtermittels, wel-
cher nach einer Wärmetrocknung bei 103 °C im Trockenschrank bis zur Massenkons-
tanz erhalten wird. Dabei verliert die Nahrung bzw. das Futtermittel sein Rohwasser,
welches sowohl Wasser aber auch Alkohole oder ätherische Öle beinhalten kann. Im
Labor wird für die Trockenmassebestimmung das sogenannte Wäge-

2 Material und Methode
36
Trocknungsverfahren mittels einer Analysewaage eingesetzt. Mittels der folgenden
Formel 1 kann der Trockengehalt jeder Probe berechnet werden.
Formel 1: Berechnung des prozentualen Trockensubstanzgehaltes nach VDLUFA (2013).
𝑇 𝑖𝑛 % = ( 𝑚! −𝑚! ∗ 100) ÷𝑚!
T = Trockenmasse in %
m1 = Leermasse der Petrischale in g
m2 = Probeneinwaage in g
m3 = Masse von Petrischale und Probe nach der Trocknung in g
2.5.1.2 Bestimmung der Rohasche (XA)
In der Rohnährstoffgruppe Rohasche werden alle Mineralstoffe in der Nahrung zu-
sammengefasst. Bei der Veraschung wird die getrocknete, gemahlene Probe auf 1 mg
genau in eine vorher geglühte und tarierte Veraschungsschale eingewogen. Anschlie-
ßend wird die Probe bei 103 °C über Nacht bis zum Verkohlen erhitzt und dann in ei-
nen auf 550 +/- 5 °C eingestellten Muffelofen gegeben. In diesem wird die Probe so
lange erwärmt, bis sich eine weiße, hellgraue oder rötliche Asche gebildet hat, die völ-
lig frei von Kohlepartikeln ist. Zuletzt wird die Probe in einen Exsikkator gestellt und
sofort nach dem Abkühlen gewogen. So lässt sich der Aschegehalt mittels Diffe-
renzwägung ermitteln, nachdem das Gewicht des Rückstandes (Angabe in Prozent)
aus der Differenz des Leergewichtes der Veraschungsschale und des Gesamtgewich-
tes errechnet wurde (siehe Formel 2).
Formel 2: Berechnung der Rohasche nach VDLUFA (2013).
𝑋𝐴 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑚! − 𝑚! ∗ 100 ∗ 100 ÷ (𝐸 ∗ 𝑇)
XA = Rohasche
M1 = Masse der leeren Veraschungsschale in g
M2 = Masse von Veraschungsschale und Probe nach der Veraschung in g
E = Probeneinwaage in g
T = Trockensubstanz der Probe in % (analytische T)

2 Material und Methode
37
2.5.1.3 Bestimmung des Rohfettes (XL)
Je nach Art der Nahrung/Futtermittels gibt es verschiedene Testvarianten, um den Ge-
halt des Rohfetts zu bestimmen. Bei dieser Arbeit wurde das Testverfahren mit vorher-
gehender Hydrolyse gewählt. Die Stoffgruppe Rohfett umfasst eine stark heteroge
Stoffgruppe, bestehend aus Triglyceriden (eigentliches Fett), Phospholipide, Glycolipi-
de, Wachse und Polyisoprenoiede (Kirchgessner et al. 2008). Bei diesem Verfahren
erfolgt erst eine Säureaufspaltung (Hydrolyse) mit einer anschließenden Extraktion der
Fette mittels Phenolether. Der prozentuale Rohfettgehalt wird wie folgt berechnet (sie-
he Formel 3).
Formel 3: Berechnung Rohfettgehalt nach VDLUFA (2013).
𝑋𝐿 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑚! − 𝑚! ∗ 100 ∗ 100 ÷ (𝐸 ∗ 𝑇)
XL = Rohfettgehalt
m1 = Masse des leeren getrockneten Extraktionsbechers in g
m2 = Masse des Extraktionsbechers mit Fett in g
E = Probeneinwaage in g
T = Trockensubstanz der Probe in % (analytische T)
2.5.1.4 Bestimmung des Rohproteins (XP)
Durch diese Methode kann der Rohproteingehalt von Futtermitteln anhand des mit
dem Kjeldahl-Verfahrens ermittelten Stickstoffgehalts bestimmt werden. In diesem Ver-
fahren werden alle stickstoffhaltigen Verbindungen erfasst. Anschließend wird das Er-
gebnis des gemessenen Stickstoffs mit dem Faktor multipliziert, der den reziproken
Wert des typischen N-Gehaltes von Rohprotein widerspiegelt. Im Fall von pflanzlichem
Protein beträgt der Faktor 6,25. Zur Ermittlung des Rohproteins werden u.a. auf das
Versetzen mit 0,1 molarer Schwefelsäure und Natronlauge sowie die Methode der Tit-
ration zurückgegriffen. Für die genaue Berechnung siehe den nachfolgenden Formel-
verlauf:

2 Material und Methode
38
Formel 4: Ermittlung der verbrauchten Schwefelsäure.
𝑉 50,0𝑚𝑙 0,1𝑛 𝐻!𝑆𝑂! −𝑊! 𝑚𝑙 0,1𝑛 𝑁𝑎𝑂𝐻) = 𝑊! 𝑚𝑙 0,1𝑛 𝐻!𝑆𝑂!
V = Vorlage
W1 = Titrationsergebnis
W2 = Menge der verbrauchten Schwefelsäure
Formel 5: 1ml Schwefelsäure 0,1n entspricht 1,4 mg Stickstoff.
1 𝑚𝑙 0,1𝑛 𝐻!𝑆𝑂! = 1,4 𝑚𝑔 𝑁
→ 𝑊! ∗ 1,4 𝑚𝑔 𝑁 = 𝐺𝑒𝑠𝑎𝑚𝑡 𝑁 𝑑𝑒𝑟 𝐹𝑢𝑡𝑡𝑒𝑟𝑚𝑖𝑡𝑡𝑒𝑙𝑒𝑖𝑛𝑤𝑎𝑎𝑔𝑒
N = Stickstoff
Formel 6: Berechnung des Gesamtstickstoffes.
𝐺𝑒𝑠𝑎𝑚𝑡 𝑁 𝑑𝑒𝑟 𝐹𝑢𝑡𝑡𝑒𝑟𝑚𝑖𝑡𝑡𝑒𝑙𝑒𝑖𝑛𝑤𝑎𝑎𝑔𝑒 ∗ 6,25 = 𝑋𝑃 𝑖𝑛 𝑚𝑔
N = Stickstoff
XP = Rohprotein
Faktor pflanzliches Protein = 6,25
Formel 7: Berechnung des Rohproteins auf die Trockenmasse.
𝑋𝑃 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑋𝑃 𝑖𝑛 𝑚𝑔 ∗ 100 ∗ 100 ÷ (𝐸 ∗ 𝑇)
XP = Rohprotein
E = Probeneinwaage in mg
T = Trockensubstanz der Probe in % (analytische T)

2 Material und Methode
39
2.5.1.5 Bestimmung der Rohfasern (XF)
Bei der Rohfaser handelt es sich um den organischen Anteil einer pflanzlichen Sub-
stanz, welcher für die Bestimmung nach einem hydrolytischen Aufschlussverfahren mit
Schwefelsäure und Kalilauge zurückbleibt. Die Rohfasern setzen sich aus Zellulose,
Lignin und Pentosane zusammen. Die Analyse der Rohasche erfolgte in Anlehnung an
den HCI-Aufschluss. Die Berechnung der Rohfaser nach Analyse geschieht wie in
Formel 8 ersichtlich.
Formel 8: Berechnung des Rohfaseranteils.
𝑋𝐹 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = ((𝑊! −𝑊!) ∗ 100 ∗ 100) ÷ (𝑊! ∗ 𝑇)
XF = Rohfaseranteil
W1 = Einwaage in g (Netto = Probeneinwaage ohne Synthetikbeutel + Celite)
W2 = getrocknete Probe in g (Brutto = Synthetikbeutel + Celite + Rückstand nach Probenauf-
schluss)
W3 = veraschte Probe in g (Brutto = Synthetikbeutel + Celite + Rückstand aus Veraschung)
T = Trockensubstanz der Probe in % (analytische T)
2.5.1.6 Bestimmung der Stickstofffreien-Extraktstoffe (NfE)
Zur Bestimmung der Stickstofffreien-Extraktstoffe genügt die Berechnung dieser aus
der Differenz der durch die Weender Analyse ermittelten anderen Rohnährstoffe. Unter
den NfE versteht man die Bestandteile Stärke, Zucker und lösliche Faseranteile. Dazu
gehören auch die Pektine, die Hemizellulose, Inulin, Glykogen wie auch lösliche Antei-
le von Zellulose, Pentosanen und Lignin. Die Berechnung der NfE erfolgt nach Formel
9.

2 Material und Methode
40
Formel 9: Berechnung der Stickstofffreien-Extraktstoffe.
𝑁𝑓𝐸 = 𝑇 − (𝑋𝐴 + 𝑋𝑃 + 𝑋𝐿 + 𝑋𝐹)
NfE = Stickstofffreie-Extraktstoffe
T = Trockensubstanz in % (nicht analytische T sondern = 100 %)
XA = Rohasche in % der T
XP = Rohprotein in % der T
XL = Rohfett in % der T
XF = Rohfaser in % der T
2.5.1.7 VanSoest Analyse
Die VanSoest Analyse ist eine Ergänzung der Weender Analyse, wurde 1967 von Pe-
ter J. van Soest entwickelt und stellt eine Detergenzienanalyse der Zellwandbestand-
teile dar. Sie dient der genaueren Auftrennung der Ergebnisse der Rohfaser- und NfE-
Bestimmung. Hierdurch können die Anteile an Hemizellulose, Zellulose und Lignin ge-
nau bestimmt werden. Auch können nach Durchführung der VanSoest Analyse durch
Errechnung die sogenannten Nicht-Faser-Kohlenhydrate (NFC) erfasst werden. Diese
Fraktion enthält die im Pansen schnell fermentierbaren Kohlenhydrate Stärke, Zucker
und die schnell abbaubaren Pektine. Die Abbildung 9 stellt die beiden Analysen mit
ihren ermittelten Parametern gegenüber.

2 Material und Methode
41
Abbildung 9: Weender Analyse und VanSoest Analyse grafische Darstellung, Quelle: verändert nach Kirchgeßner et al. 2008.
2.5.1.8 Bestimmung der Neutral-Detergenzien-Faser (NDF)
Unter der Neutral-Detergenzien-Faser (NDF) werden alle Bestandteile einer Nahrung
verstanden, die in einer neutralen Detergenzienlösung unter den angegebenen Bedin-
gungen nicht gelöst werden. Diese bestehen in der Summe hauptsächlich aus pflanzli-
chen Zellwandgerüstsubstanzen wie Hemizellulose, Zellulose, Lignin und Lignin-N-
Verbindungen. Die Durchführung erfolgte nach den Normen der VDLUFA. Die folgende
Formel 10 zeigt die Berechnung des NDF-Gehaltes auf.

2 Material und Methode
42
Formel 10: Berechnung des NDF-Gehaltes.
𝑁𝐷𝐹 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑊! −𝑊! ∗ 100 ∗ 100 ÷ (𝑊! ∗ 𝑇)
NDF = Neutral-Detergenzien-Faser
W1 = Einwaage der Probe in g
W2 = Gewicht des getrockneten Rückstanden einschließlich Glasfiltertiegel in g
W3 = Gewicht des leeren Glasfiltertiegels in g
T = Trockensubstanz der Probe in % (analytische T)
2.5.1.9 Bestimmung der Säure-Detergenzien-Faser (ADF)
Unter der Säure-Detergenzien-Faser werden alle Bestandteile einer Nahrung verstan-
den, die sich in einer sauren Detergenzienlösung unter den beschriebenen Bedingun-
gen nicht lösen. Durch den ADF-Gehalt lässt sich direkt auf den Gehalt der Zellwand-
gerüstsubstanzen Zellulose, Lignin und Lignin-N-Verbindungen schließen. Auch hier
erfolgte die Analyse nach den Richtlinien der VDLUFA. Die Formel 11 beschreibt die
Berechnung des ADF Gehaltes.
Formel 11: Berechnung des ADF-Gehaltes.
𝐴𝐷𝐹 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑊! −𝑊! ∗ 100 ∗ 100 ÷ (𝑊! ∗ 𝑇)
ADF = Säure-Detergenzien-Faser
W1 = Einwaage der Probe in g
W2 = Gewicht des getrockneten Rückstanden einschließlich Glasfiltertiegel in g
W3 = Gewicht des leeren Glasfiltertiegels in g
T = Trockensubstanz der Probe in % (analytische T)
2.5.1.10 Bestimmung der Säure-Detergenzien-Lignins (ADL)
Im Anschluss an die Bestimmung des ADF-Gehaltes folgte die Analyse des Säure-
Detergenzien-Lignins (auch Rohlignin genannt). Dieses wurde ebenfalls nach den
Richtlinien der VDLUFA ermittelt. Die Formel 12 zeigt die Berechnung des ADL-
Gehaltes auf.

2 Material und Methode
43
Formel 12: Berechnung des ADL-Gehaltes.
𝐴𝐷𝐿 𝑖𝑛 % 𝑑𝑒𝑟 𝑇 = 𝑊! −𝑊! ∗ 100 ∗ 100 ÷ (𝑊! ∗ 𝑇)
ADL = Säure-Detergenzien-Lignin
W1 = Einwaage der Probe in g
W2 = Gewicht des Glasfiltertiegel mit Inhalt nach Behandlung mit Schwefelsäure und Trocknung
in g
W3 = Gewicht des Glasfiltertiegels nach Veraschung in g
T = Trockensubstanz der Probe in % (analytische T)
2.5.1.11 Bestimmung der Zellulose und Hemizellulose
Durch die genaue Bestimmung des NDF-, ADF- und ADL-Gehaltes ist es nun möglich,
rechnerisch den exakten Gehalt von Zellulose und Hemizellulose zu bestimmen. Der
Formelverlauf zeigt den Vorgang.
Formel 13: Berechnung des Zellulose-Gehaltes.
𝑍𝑒𝑙𝑙𝑢𝑙𝑜𝑠𝑒 = 𝐴𝐷𝐹 − 𝐴𝐷𝐿
ADF = Säure-Detergenzien-Faser
ADL = Säure-Detergenzien-Lignin
Formel 14: Berechnung des Hemizellulose-Gehaltes.
𝐻𝑒𝑚𝑖𝑧𝑒𝑙𝑙𝑢𝑙𝑜𝑠𝑒 = 𝑁𝐷𝐹 − 𝐴𝐷𝐹
NDF = Neutral-Detergenzien-Faser
ADF = Säure-Detergenzien-Faser

2 Material und Methode
44
2.5.1.12 Bestimmung der Nicht-Faser-Kohlenhydrate (NFC)
Diese Fraktion der Kohlenhydrate enthält nur die schnell fermentierbaren Kohlenhydra-
te Zucker, Stärke und Pektine. Diese lassen sich nach Durchführung des Weender
Verfahrens wie auch des VanSoest Verfahrens durch die Formel 15 berechnen.
Formel 15: Berechnung der Nicht-Faser-Kohlenhydrate.
𝑁𝐹𝐶 = 𝑇 − (𝑋𝐴 + 𝑋𝑃 + 𝑋𝐿 + 𝑁𝐷𝐹)
NFC = Nicht-Faser-Kohlenhydrate
T = Trockensubstanz in % (nicht analytische T sondern = 100 %)
XA = Rohasche in % der T
XP = Rohprotein in % der T
XL = Rohfett in % der T
NDF = Neutral-Detergenzien-Faser in % der T
2.5.2 Analyse Pansensaft
Die Pansensaftanalyse dient zur Ermittlung der Qualität der aufgenommenen Nahrung.
Anhand der von Mikroorganismen aus der Nahrung gebildeten Fermentationsprodukte
lassen sich Rückschlüsse auf die Art und die Menge der Rohnährstoffgehalte in der
Nahrung treffen. Hierfür wurden folgende Analysen herangezogen. Tabelle 4 zeigt die
Analysen auf.
Tabelle 4: Parameter zur Analyse der Qualität der Nahrung von Rehen. Parameter Indikator für Analyseverfahren
Flüchtige Fettsäuren
Schwer- und leichtlösliche Fasern
Gaschromatographie
Ammoniak
Protein- und Kohlehydratgehalt
Photometrie
Milchsäure Kohlehydratgehalt Photometrie
Zur späteren Analyse des Pansensaftes bedarf es einer Aufbereitung, um eine mög-
lichst hohe Reinheit des Pansensaftes zu erlangen. Hierfür müssen alle Partikel und
Schwebstoffe entfernt werden. Der mittels Zentrifugation des Pansensaftes erhaltene
Überstand diente im späteren Verlauf zum einen für die Messung des Ammoniak- und
Milchsäuregehaltes und zum anderen für die Bestimmung des FFS. Für letztere wurde

2 Material und Methode
45
dieser mit 1,5 ml Metaphosphorsäure (25 %ig) und 0,5 ml Ameisensäure (98-100 %ig)
versetzt und dem nach erneutem Zentrifugieren erhaltenen Überstand für den Erhalt
als Säure ein Tropfen gesättigtes Quecksilberchlorid hinzugegeben. Für die Lagerung
und den späteren Einsatz zur Messung wurden die Proben der NH3- und Laktat-
Messung sowie der FFS-Messung mittels eines Spritzenfilters in Eppendorf Tubes
überführt und bei -18 °C gelagert. FFS-Analyse
Bei den flüchtigen Fettsäuren handelt es sich um kurzkettige, gesättigte Fettsäuren, die
sich in Essigsäure, Propionsäure, Buttersäure, Iso-Buttersäure, Valeriansäure und Iso-
Valeriansäure unterteilen lassen. Sie entstehen als Endprodukt bei der mikrobiellen
Fermentation von Kohlenhydraten, Stärken und schwerlöslichen Fasern wie z.B. Zellu-
lose im Pansen. Die Menge an gebildeten Fettsäuren ist hauptsächlich von der Nah-
rungszusammensetzung anhängig und nimmt zu, je energiereicher die aufgenommene
Nahrung ist (Sommer 2004). Das Verhältnis der beiden am häufigsten vorkommenden
Säuren Essigsäure und Propionsäure spiegelt den Anteil an leicht- und schwerlösli-
chen Fasern wieder (siehe Formel 29). Je mehr Essigsäure im Verhältnis zu Propions-
äure vorhanden ist, desto höher war der Anteil an schwerlöslichen Fasern in der Nah-
rung (Zellulose). Ein erhöhter Propionsäure-Wert im Verhältnis zur Essigsäure spiegelt
hingegen einen gesteigerten Anteil an leichtlöslichen Fasern (Zucker, Stärke) wider
(Enzinger & Hartfiel 1998). Die FFS-Analyse im Pansensaft gibt Auskunft über die Art
der Nahrungszusammensetzung.
Formel 29: Verhältnis Essigsäure zu Propionsäure.
𝑉!" = 𝐶! ÷ 𝐶!
VEP = Verhältnis C2 zu C3 (schwerlösliche Fasern zu leichtlösliche Fasern)
C2 = Essigsäure (mmol/l)
C3 = Propionsäure (mmol/l)
Die FFS-Analyse wurde mittels der Gaschromatographie durchgeführt, welche zur Auf-
spaltung komplexer Gemische dient. Die Lösung wurde in eine Trennsäule injiziert. In
der Trennsäule lösen sich die einzelnen Komponenten und werden mit der mobilen

2 Material und Methode
46
Phase (Stickstoff) transportiert. Je mehr C-Atome im Gas enthalten sind, desto länger
verbleibt die Substanz im Chromatographen (Kolb 2003).
Abbildung 10: Auftrennung der Flüchtigen Fettsäuren im Gaschromatograph, Probe Nr. 243, Quelle: Geritschke, Scheingraber
Die ermittelten FFS-Ergebnisse in g/l wurden, zur Vergleichbarkeit mit der Literatur, in
mmol/l umgerechnet. Dafür musste im ersten Schritt die Stoffmenge in Mol bestimmt
werden (siehe Formel 16).
Formel 16: Bestimmung der Stoffmenge nach Mentor (2003).
𝑀 =𝑚𝑛
n ≙ Stoffmenge [mol] m ≙ Stoffmasse [g] M ≙ Molare Masse [g/mol]

2 Material und Methode
47
Anschließend erfolgte eine Umstellung auf die aktuelle Fragestellung unter Hinzufü-
gung des Parameters „Liter“ (siehe Formel 17).
Formel 17: Formel 4 nach Stoffmenge pro Liter aufgelöst.
𝑛! = 𝑚!
𝑀
nl ≙ Stoffmenge [mol/l] ml ≙ Stoffmasse [g/l] M ≙ Molare Masse [g/mol]
2.5.2.1 Ammoniak Analyse
Mikroorganismen des Pansens nutzen als Stickstoffquelle sowohl das Nahrungsprotein
als auch in der Nahrung enthaltene Nicht-Protein-Stickstoff-(NPN)-Verbindungen. Am-
moniak (NH3) entsteht entweder bei der Aufspaltung solcher NPN-Verbindungen oder
als Endprodukt der sogenannten Desaminierung. Darunter versteht man den letzten
Schritt der Spaltung von Nahrungseiweißen durch die Pansenfauna und -flora. Der
Ammoniak dient nun den Mikroorganismen als Stickstoffquelle und wird zum Aufbau
von bakterien- und protozoeneigenen Proteinen verwendet (Sommer 2004). Hohe NH3-
Werte spiegeln einen hohen Eiweißgehalt der Nahrung und demnach eine hohe Ener-
giezufuhr wider. Die hier angewendete Methode dient der Erfassung des sich im Pan-
sen befindlichen Ammoniaks. Die Messung des NH3-Gehaltes des Pansensaftes er-
folgte mit Hilfe der Photometrie, einer Analysemethode, die dem Prinzip der Messung
von Lichtströmen unterliegt. Ziel bei dieser Methode ist es, die Transmission, d.h. die
Durchlässigkeit, des zu untersuchenden Mediums anhand von Messungen der Intensi-
täten des eingestrahlten und austretenden Lichtes zu berechnen. Diese ist abhängig
von der Anzahl der in der Lösung vorhandenen Teilchen. Durch die Nutzung der deka-
dischen Basis bei der Berechnung führt dies zum Begriff der Extinktion, welche auch
hier verwendet wird (Oesterle 1995). Für die Analyse reagiert der im Pansensaft ent-
haltene Ammoniak mit α-Ketoglutarsäure (KGA) und NADPH in Gegenwart von dem
Enzym L-Glutamatdehydrogenase (GDH) zu L-Glutamat und NADP+ (siehe Formel 18).

2 Material und Methode
48
Die Abnahme der Absorption bei 340 nm ist aufgrund der Oxidation des NADPH pro-
portional zur Ammoniak-Konzentration.
Formel 18: Reaktionsgleichung Bildung von NADP+ (Sigma 2012).
𝐾𝐺𝐴 + 𝑁𝐻!! + 𝑁𝐴𝐷𝑃𝐻!"#
𝐿 − 𝐺𝑙𝑢𝑡𝑎𝑚𝑎𝑡𝑒 + 𝑁𝐴𝐷𝑃! + 𝐻!𝑂
Formel 19: Berechnung der Endkonzentration NH3 (Sigma 2012), ∆A340 = Adsorption bei 340nm. ∆A340 = AAnfangswert – AEndwert
∆(∆A340) Test oder Standard = ∆A340(Test oder Standard) - ∆A340(Blank)
Die weitere Berechnung erfolgt nach der Formel des Ammonia Assay Kits (siehe For-
mel 20).
Formel 20: Berechnung des Ammoniakgehaltes (Sigma 2012).
𝑚𝑔 𝑁𝐻!/𝑚𝑙 𝑑𝑒𝑟 𝑂𝑟𝑖𝑔𝑖𝑛𝑎𝑙𝑝𝑟𝑜𝑏𝑒 = 𝐴 ×𝑇𝑉×𝑀𝑊 𝑜𝑓 𝐴𝑚𝑚𝑜𝑛𝑖𝑎 ×𝐹
𝜀 × 𝑑 ×𝑆𝑉×𝐶𝑜𝑛𝑣𝑒𝑟𝑠𝑖𝑜𝑛 𝐹𝑎𝑐𝑡𝑜𝑟 𝑓𝑜𝑟 𝜇𝑔 𝑡𝑜 𝑚𝑔
A = ∆(∆A340) Test oder Standard
TV = Total Assay Volume [ml]
MW of Ammonia = 17 g/mol oder entsprechend 17 µg/µmol
F = Dilution Factor from Sample Preparation
ε = Millimolar Extinktions-Koeffizient für NADPH bei 340 nm
d = Light path = 1 cm
SV = Sample Volume [ml]
2.5.2.2 Milchsäure Analyse
Neben FFS, NH3, Alkohol und Gasen (CO2, CH4) entsteht auch Milchsäure (Laktat,
C3H6O3) bei der Fermentation im Pansen. Laktat entsteht bei einem raschen Abbau
leichtlöslicher Kohlenhydrate (KH). Die Anzahl an Laktat-produzierenden Bakterien im
Pansen ist normalerweise gering, da sie einen sehr niedrigen pH-Wert präferieren. Bei
der Aufnahme ungewohnt großer Mengen leichtlöslicher Kohlenhydrate entsteht durch
deren raschen Abbau zu viel Laktat, welches nicht schnell genug zu Propionsäure um-
gewandelt werden kann. Resultat ist eine Absenkung des Pansen-pH-Wertes und so-

2 Material und Methode
49
mit ausgeprägte Veränderungen der Pansenmikroflora zugunsten milchsäurebildender
Bakterien, da die zellolytischen sowie laktatabbauenden Bakterien und Protozoen ab-
sterben (Abbildung 14). Dies fördert erneut eine Laktatbildung und Anreicherung. Eine
zu hohe Milchsäuregärung im Pansen führt nach längerer Zeit zu einer Pansenazido-
se. Der Laktatgehalt im Pansensaft ist daher ein guter Indikator für eine erhöhte Zufuhr
von kohlenhydratreicher und rohfaserarmer Nahrung (Brüggemann et al. 1967, 1968).
Der durchschnittliche Pansen-pH-Wert liegt zwischen 5,4 und 7,5 (Lohse 2000, Ritz et
al. 2013). In diesem pH-Bereich sollte der Propionsäurewert über dem des Milchsäu-
rewertes liegenDie Detektion der Milchsäure im Pansensaft dient als Hinweis auf eine
mögliche Übersäuerung und wird mittels Photometrie bei 340 nm durchgeführt. Die
Vorgehensweise des Tests erfolgt wie nach Roche beschrieben, Anleitungsbeschrei-
bung des Kits 2012. Während der stattfindenden Reaktionen wird Nicotinamid-adenin-
dinukleotid-hydrid (NADH) gebildet, welches als Messgröße dient.
Formel 21: Reaktionsgleichung Bildung von NADH (Roche 2012).
𝐿 − 𝐿𝑎𝑘𝑡𝑎𝑡 + 𝑁𝐴𝐷!!!!"#
𝑃𝑦𝑟𝑢𝑣𝑎𝑡 + 𝑁𝐴𝐷𝐻 + 𝐻!
𝐷 − 𝐿𝑎𝑘𝑡𝑎𝑡 + 𝑁𝐴𝐷!!!!"#
𝑃𝑦𝑟𝑢𝑣𝑎𝑡 + 𝑁𝐴𝐷𝐻 + 𝐻!
Für den Erhalt der Endkonzentration von Laktat müssen verschiedene Extinktionen der
Proben gemessen und im Anschluss Extinktionsdifferenzen nach Formel 22 gebildet
werden:
Formel 22: Berechnung der Endkonzentration von Laktat. E2-E1 (Ergebnis: ∆ED-Milchsäure) sowie E3-E2 (Ergebnis: ∆EL-Milchsäure)
Formel 23 stellt die allgemeine Berechnungsformel für die Bestimmung der Konzentra-
tion dar.

2 Material und Methode
50
Formel 23: Bestimmung der Laktat-Konzentration (Roche 2012).
𝑐 =𝑉×𝑀𝐺
𝜀×𝑑×𝑣×1000×∆𝐸 𝑔 𝑙
c = Laktat-Konzentration in %
V = Testvolumen [ml] → 2,260 für L-Milchsäure; 2,240 für D-Milchsäure
MG = Molekulargewicht der zu bestimmenden Substanz [g/mol]
ε = Extinktionskoeffizient von NADH bei 340 nm = 6,3 [l x mmol-1 x cm-1]
d = Schichtdicke [cm] → 1,0 cm
v = Probevolumen [ml] → 0,1 ml
E [g/l] = Energie Gramm pro Liter (g/l)
Daraus ergibt sich für L-Milchsäure folgende Formel 24:
Formel 24: Berechnung von L-Milchsäure mit ∆EL-Milchsäure (Roche 2012).
𝑐 =2,036𝜀
×∆𝐸 [𝑔 𝐿 −𝑀𝑖𝑙𝑐ℎ𝑠ä𝑢𝑟𝑒 /𝑃𝑟𝑜𝑏𝑒𝑙ö𝑠𝑢𝑛𝑔]
c = Laktat-Konzentration in %
ε = Extinktionskoeffizient von NADH bei 340 nm = 6,3 [l x mmol-1 x cm-1]
E [g/l] = Energie Gramm pro Liter (g/l)
Beziehungsweise für D-Milchsäure nach Formel 25:
Formel 25: Berechnung von D-Milchsäure mit ∆ED-Milchsäure (Roche 2012).
𝑐 =2,018𝜀
×∆𝐸 [𝑔 𝐷 −𝑀𝑖𝑙𝑐ℎ𝑠ä𝑢𝑟𝑒 /𝑃𝑟𝑜𝑏𝑒𝑙ö𝑠𝑢𝑛𝑔] c = Laktat-Konzentration in %
ε = Extinktionskoeffizient von NADH bei 340 nm = 6,3 [l x mmol-1 x cm-1]
E [g/l] = Energie Gramm pro Liter (g/l)
2.5.2.3 pH-Wert
Der pH-Wert des Pansensaftes ist für die Abläufe aller Verdauungsprozesse im Pan-
sen und somit für das Wohlbefinden und den physiologischen Zustand der Rehe von
großer Bedeutung. Er ist im Wesentlichen vom gefressenen Substrat abhängig. Leicht-
lösliche Kohlenhydrate (Einfach- und Zweifachzucker) sowie Stärke führen zu einer

2 Material und Methode
51
hohen und raschen Säurebildung. Schwerverdauliches Substrat mit hohem Zellulose-
oder Ligninanteil liefert sehr langsam entstehende Säure. Eine länger anhaltende Pan-
senübersäuerung führt zum Tod der Tiere (Deutz et al. 2009). Die Messung des pH-
Wertes des Pansensaftes gibt demnach Aufschluss über die Art des gefressenen Sub-
strates und dient als guter Weiser für ein intaktes Verdauungssystems. Nach vorange-
gangener Homogenisierung erfolgte die Messung des pH-Wertes mittels einer Sonde,
welche in die Weithalsflasche gegeben wurde. Nach 2 min permanenten leichten
Schwenkens konnten pH-Wert (0 - 14) und Temperatur (°C) abgelesen werden.
2.6 Analyse zur Energie der Nahrung (Scheingraber)
Der Hohenheimer Futterwerttest (HFT) oder auch Gastest (Steingaß & Menke 1986,
Menke & Steingass 1988) dient der Abschätzung des Energiegehaltes der Nahrung. Im
Standardverfahren wird hierfür der gefriergetrocknete und auf 1 mm Korngröße ge-
mahlene Raufaseranteil der Nahrung mit frischem Hammelpansensaft in vitro inkubiert.
Um die spezielle Energiegewinnung des Rehes zu erfassen, wurde neben dem Stan-
dardverfahren mit Hammelpansensaft auch frischer Rehpansensaft verwendet.
Die im Pansensaft enthaltenen Mikroorganismen fermentieren die Nahrungspartikel,
wobei die dabei entstehenden Gase (CO2, CH4) aufgefangen und volumetrisch gemes-
sen werden (Gasbildungswert). Da grundsätzlich eine hohe Korrelation zwischen Sub-
stratabbau und Endproduktbildung bei der Fermentation besteht, lässt sich mithilfe des
Gasbildungswertes sowie mit den im Weender Verfahren ermittelten Rohnährstoffen
über eine Regressionsgleichung die enthaltene Energie der Nahrung errechnen.
Zur Verdeutlichung der verschiedenen Energieformen siehe Abbildung 11.

2 Material und Methode
52
Abbildung 11: Energieformen und Energiekaskade
Es interessiert in erster Linie neben der Bruttoenergie der Nahrung (GE), welche auch
als Gesamtenergie der Nahrung verstanden wird, die Umsetzbare Energie (ME). Hier-
unter versteht man die Energie, die dem Organismus nach Verlust von Energie in Form
von Kotabgabe, Harnausscheidung und Darmgase zur Verfügung steht (Abbildung 11).
2.6.1 Standard HFT (Hammelpansensaft)
Zur Schätzung des Energiewertes eines Nahrungsmittels über den Standard Hohen-
heimer Futterwerttests wird Hammelpansensaft verwendet. Für die Durchführung ist
die Entnahme frischen Pansensaftes notwendig, welcher von zwei ausgewachsenen
Hammeln vor der Morgenfütterung via Pansenfistel entnommen wird. Um ein Absinken
der Temperatur des Pansensaftes zu verhindern, wird die Flasche vor Entnahme des
Pansensaftes auf 39 °C vorgewärmt und während der Entnahme mit CO2 begast, um
Sauerstoffeintritt zu verhindern. Der Transport ins Labor erfolgt ebenfalls begast im
vorgewärmten Thermogefäß (Abbildung 12). Die Durchführung des HFT erfolgte nach
dem Methodenhandbuch der Futtermittelanalytik III Stand 1997 der Bioanalytik der TU
München. Abbildung 12 gibt die Gewinnung des Pansensaftes wieder, in Abbildung 13
werden Teilschritte des HFT im Labor dargestellt.
Bruttoenergie GE
Verdauliche Energie DE
Umsetzbare Energie ME
Nettoenergie NE
Ø Verlust als Kot Energie
Ø Verlust als Harn Energie Ø Verlust als Energie der
Darmgase
Ø Verlust als thermische Energie

2 Material und Methode
53
Abbildung 12: Spendertier mit Pansenfistel (links), Woulff’sche Flasche mit Pansensaft (mittig), Thermobehälter mit Pansensaft und CO2 Begasung (rechts), Quelle: Scheingra-ber
Die durch die Reaktionen im Kolbenprober entstandenen Gase (CO2, CH4, N2, O2, H2
und H2S) bilden einen „Gasbildungswert“, mit welchem sich anhand einer Formel auf
die Energie der Nahrung schließen lässt. Eine vorherige Analyse der Rohnährstoffe
(wie dies im Weender Verfahren geschieht) ist notwendig, um die erforderlichen Para-
meter in die Gleichung einsetzen zu können.
Abbildung 13: Reagenzien zur Herstellung der Mediumlösung (links), Pansensaft-Filtration (mittig), Zugabe des Pansensaftes in Woulff’scher Flasche (rechts), Quelle: Scheingraber
Menke & Steingaß (1988) entwickelten Regressionsgleichungen zur Berechnung der
Verdaulichkeit der organischen Substanz (dOS), der Bruttoenergie (GE) und der Um-
setzbaren Energie (ME). Um die Formel anwenden zu können, ist zunächst eine Un-
terscheidung in Kraft- oder Raufutter nötig, da hierfür verschiedene Koeffizienten in die
Gleichung einfließen. In vorliegender Arbeit wurden die Werte mittels der Raufutterfor-
mel berechnet, da es sich um wildlebende Tiere ohne Fütterung handelt. Die folgende
Formelsammlung gibt die Berechnungen der Parameter Bruttoenergie, Umsetzbare

2 Material und Methode
54
Energie und Verdaulichkeitsfaktor mit hoher Genauigkeit wieder (nach Menke und
Steingaß 1988). Die gesamte Durchführung (Doppelbestimmung) des HFTs wird mit
zwei Laborwiederholungen an zwei verschiedenen Tagen durchgeführt. Somit erfahren
alle Proben eine Vierfach-Bestimmung. Es wurde jeweils der Mittelwert der vier GB24-
Werte einer Probe für die Berechnung der Energie verwendet.
Formel 26: Berechnung der Gasbildung GB24.
𝐺𝐵!" =𝑉!" − 𝑉! + 𝑉! − 30𝑚𝑙 − 𝐺𝐵!
𝐸𝑖𝑛𝑤𝑎𝑎𝑔𝑒 𝑖𝑛 𝑚𝑔∗ 200𝑚𝑔 ∗ 𝐹𝑎𝑘𝑡𝑜𝑟
GB24 = Gasbildung (ml/200mg)
V24 = Volumen nach 24 Stunden (ml)
V0 = Volumen nach Befüllen des Kolbens (ml)
V8 = Volumen nach 8 Stunden (ml)
GB0 = Blindwert (ml), d.h. mittlere Gasbildung, wenn nur Pansensaft inkubiert wird
Faktor = für Kontrollprobe 1 wird überprüft, wie weit der im konkreten Analysengang ermittelte
Wert vom Normwert der Bezugsserien abweicht
Formel 27: Berechnung der Umsetzbaren Energie ME.
𝑀𝐸 = 2,20 + 0,136 ∗ 𝐺𝐵24 + 0,057 ∗ 𝑋𝑃 + (0,00285 ∗ 𝑋𝐿 ∗ 𝑋𝐿)
ME = Umsetzbare Energie (MJ/kg T)
GB24 = Gasbildungswert (ml/200mg T in 24 Stunden)
XP = Rohprotein (in % der T)
XL = Rohfett (in % der T)

2 Material und Methode
55
Formel 28: Berechnung des Verdaulichkeitsfaktors dOS.
𝑑𝑂𝑆 = 15,38 + 0,845 ∗ 𝐺𝐵 + 0,595 ∗ 𝑋𝑃 + (0,675 ∗ 𝑋𝐴)
dOS = Verdaulichkeitsfaktor (%)
GB = Gasbildungswert (ml/200mg T in 24 Stunden)
XP = Rohprotein (in % der T)
XA = Rohasche (in % der T)
2.6.2 HFT mit Rehpansensaft
Wie in Punkt 2.6.1 beschrieben, wurde zuerst das Standard HFT Verfahren für die Ge-
samtheit aller Proben angewendet. Zusätzlich wurden ausgewählte Proben mit einem
spezialisierten HFT Verfahren behandelt, in dem statt mit herkömmlichem Hammel-
pansensaft mit Rehpansensaft gearbeitet wurde. Ziel war es, die Anpassung des rumi-
nalen Verdauungssystems des Rehwildes hinsichtlich der Jahreszeiten zu untersuchen
und mögliche Unterschiede herauszuarbeiten. Da die Durchführung dieses Versuches
mit erheblichen Schwierigkeiten und Aufwand verbunden war und energetisch der Win-
ter die entscheidende Jahreszeit darstellt, wurde der Versuchsansatz nur im Winter
durchgeführt. Zur Analyse des Winteransatzes wurden Proben aus folgenden Monaten
ausgewählt (siehe Tabelle 5).
Tabelle 5: Verwendete Probenanzahl und Herkunft für spezialisiertes HFT-Verfahren. Herkunftsmonat
Proben
Versuchsansatz Winter
Probenanzahl
November 5
Dezember 6
Januar 7
Gesamt 18
Um dem Standardverfahren so ähnlich wie möglich zu sein, wurde ein Pansensaftge-
misch von zwei beim Morgenansitz erlegten Rehen verwendet. Hierdurch werden indi-

2 Material und Methode
56
viduenspezifische Unterschiede minimiert und ein hoher Pansensaftanteil, ähnlich der
Nüchternheit der Schafe, erzielt.
Abbildung 14: Spendertier für HFT-Verfahren mit Rehpansensaft im Warmhalteprozess, Quelle: Kuwert, Scheingraber
2.7 Konditionsbestimmung (Scheingraber)
Die Konditionsbestimmung von Tieren ist in der Wildtierbiologie eine gängige Methode
zur Bestimmung des Verfassungszustandes von Tieren. Hierfür wird oft die Gewichts-
bestimmung mit einer Methode der Konditionsbestimmung kombiniert, da beide Para-
meter gute Indikatoren für den Gesundheits- und Allgemeinzustand eines Tieres dar-
stellen. Die Kondition eines Tieres spiegelt im Gegensatz zur Konstitution nicht allein
die genetisch vermittelten Eigenschaften eines Individuums wider, sondern in erster
Linie den aktuellen körperlichen Verfassungszustand eines Individuums, welcher durch
äußere Umwelteinflüsse gesteuert wird.
Die in dieser Studie verwendete Methode zur Konditionsbestimmung ist der Nierenfett-
index/Kidney Fat Index (NFI/KFI) nach Riney (1955). Der Nierenfettindex gibt das Ge-
wicht des die Niere umgebenden Fettes in Prozent (0 – 300 % Skala) des Nierenge-
wichtes an. Die Berechnung des Indexes erlaubt den Vergleich der Nierenfettwerte
zwischen verschiedenen Individuen. Der Nierenfettindex liefert Informationen über den
Ernährungszustand und die Kondition der Tiere und gibt somit auch einen Anhalts-
punkt auf die Nahrungsverfügbarkeit und Nahrungsqualität in den Habitaten. Zur Be-

2 Material und Methode
57
stimmung des NFI wurden beide Nieren mit umliegendem Fettgewebe entnommen.
Zur Standardisierung nach Riney (1955) wurde im Labor mittels Skalpells und Pinzette
seitlich an den Nierenpolen überstehendes Fett weggeschnitten, die Nieren aus dem
Fettgewebe sowie aus dem Nierenhäutchen freipräpariert und Nieren bzw. Fettgewebe
auf einer Präzisionswaage auf 0,01 g gewogen. Zudem wurden beide Nieren mit einer
digitalen Schublehre nach Länge, Breite und Tiefe (in mm) vermessen. Zur Berech-
nung des Nierenfettindexes wurde Formel 29 verwendet.
Formel 29: Berechnung des Nierenfettindexes nach Riney (1955). 𝐹𝑒𝑁𝑖
∗ 100
Fe ≙ Fettgewebe feucht [g]
Ni ≙ Niere feucht [g]
2.8 Volumenbestimmung Pansen (Scheingraber)
Die Messung des Pansenvolumens gibt Aufschluss über die physiologische Adaption
des Pansens der Rehe an die Jahreszeit und die Nahrungsverfügbarkeit in den Habita-
ten (Kamler et al. 2002), da Rehe je nach Notwendigkeit ihr Pansenvolumen reduzie-
ren oder ausdehnen können. Für diese Analyse wurde der für das Entleeren aufge-
schnittene Pansen mit Nadel und Faden wieder zugenäht, um eine möglichst wasser-
dichte und exakte Rekonstruktion des Ursprungsvolumens zu gewährleisten. Um das
Volumen des Pansens zu bestimmen, wurde dieser bis zur Schnittstelle Pila ruminore-
ticularis, an welcher der Netzmagen abgetrennt wurde, mit Wasser befüllt. Der gefüllte
Pansen wurde alsdann in eine Plastikwanne gesetzt und das Wasser aus dem Pansen
geleert sowie das restliche Wasser aus dem Pansen gedrückt. Mittels eines Messzy-
linders konnte im weiteren Verlauf die Menge an Wasser [in dm³] bestimmt werden.
Die Abbildung 15 beschreibt den Ablauf der Volumenmessung der Pansen.
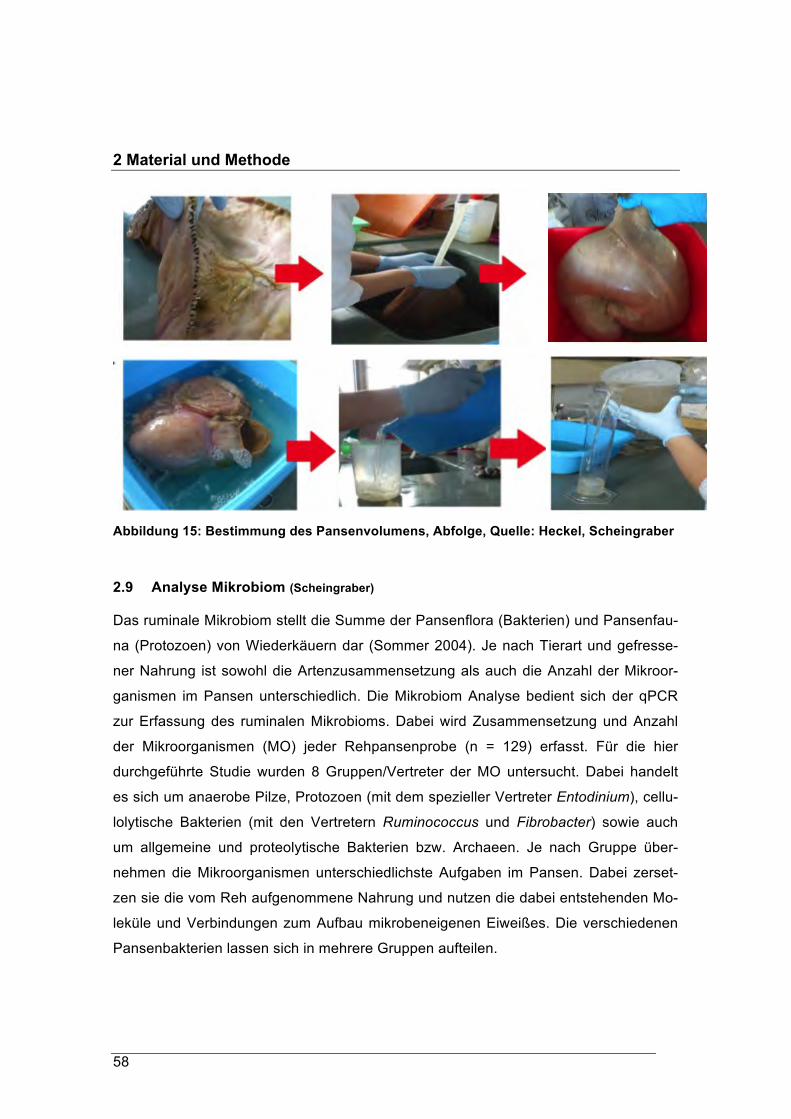
2 Material und Methode
58
Abbildung 15: Bestimmung des Pansenvolumens, Abfolge, Quelle: Heckel, Scheingraber
2.9 Analyse Mikrobiom (Scheingraber)
Das ruminale Mikrobiom stellt die Summe der Pansenflora (Bakterien) und Pansenfau-
na (Protozoen) von Wiederkäuern dar (Sommer 2004). Je nach Tierart und gefresse-
ner Nahrung ist sowohl die Artenzusammensetzung als auch die Anzahl der Mikroor-
ganismen im Pansen unterschiedlich. Die Mikrobiom Analyse bedient sich der qPCR
zur Erfassung des ruminalen Mikrobioms. Dabei wird Zusammensetzung und Anzahl
der Mikroorganismen (MO) jeder Rehpansenprobe (n = 129) erfasst. Für die hier
durchgeführte Studie wurden 8 Gruppen/Vertreter der MO untersucht. Dabei handelt
es sich um anaerobe Pilze, Protozoen (mit dem spezieller Vertreter Entodinium), cellu-
lolytische Bakterien (mit den Vertretern Ruminococcus und Fibrobacter) sowie auch
um allgemeine und proteolytische Bakterien bzw. Archaeen. Je nach Gruppe über-
nehmen die Mikroorganismen unterschiedlichste Aufgaben im Pansen. Dabei zerset-
zen sie die vom Reh aufgenommene Nahrung und nutzen die dabei entstehenden Mo-
leküle und Verbindungen zum Aufbau mikrobeneigenen Eiweißes. Die verschiedenen
Pansenbakterien lassen sich in mehrere Gruppen aufteilen.

2 Material und Methode
59
Die Essigsäurebildner oder auch cellulolytische Bakterien genannt, wie Fibrobacter
succinogenes und Ruminococcus flavefaciens, sind strikt anaerob, können Rohfasern
wie Zellulose und Hemizellulose zersetzen und bilden Essigsäure.
Die amylolytischen Bakterien wie Bacteroides amylophilus und Streptococcus bovis
gehören zu den Propion- und Buttersäurebildnern. Sie können vor allem Stärke so-
wie Einfach- und Zweifachzucker vergären und sind Schwankungen des pH-Werts ge-
genüber weniger empfindlich.
Eine weitere Bakteriengruppe sind die proteolytischen Pansenbakterien. Sie bauen
Proteine und Aminosäuren zu Ammoniak um. Darunter befinden sich zum Beispiel die
Gruppen Prevotella spp. und Ruminococcus amylophilus.
Neben den Bakterien findet man Protozoen, hauptsächlich Flagellaten und Cilliaten, im
Pansen von Wiederkäuern. Sie gehören zu den Einzellern und zersetzen überwiegend
Stärke und Proteine im Pansen. Bei der Proteinzersetzung greifen sie häufig auf ganze
Bakterien zurück. Protozoen sind zusammen mit den proteolytischen Bakterien für die
Hauptproduktion von Ammoniak im Pansen verantwortlich.
Anaerobe Pilze spielen bei der Verdauung im Pansen eine wichtige Rolle, da sie ha-
ben die Fähigkeit, mit ihren Hyphen Lignin aufzuspalten, und liefern hierdurch einen
wichtigen Beitrag bei der Faserverdauung (Theodorou et al. 1988). Jedoch weisen sie
nur einen recht geringen mengenmäßigen Anteil am Pansenmikrobiom auf.
Zuletzt findet man Archaeen im Pansen. Diese sind ebenfalls strikt anaerob und bilden
aus Essigsäure, Kohlendioxid und Wasserstoff mittels Methanogenese Methan. Als
Folge sinkt der Partialdruck im Pansen, wodurch eine übermäßige Bildung von Milch-
säure (Lactat) und somit eine Übersäuerung verhindert wird.
Die Ermittlung des ruminalen Mikrobioms findet mittels quantifizierter Echtzeit-
Polymerase Kettenreaktion (real-time qPCR) statt. Diese Methode der modernen Mo-
lekularbiologie dient der Feststellung eines bestimmten, artspezifischen DNA-
Fragmentes in der zuvor isolierten DNA-Probe. So lässt sich aus einer Vielzahl von
DNA-Stücken das gesuchte feststellen und das Vorkommen dieser Art nachweisen.
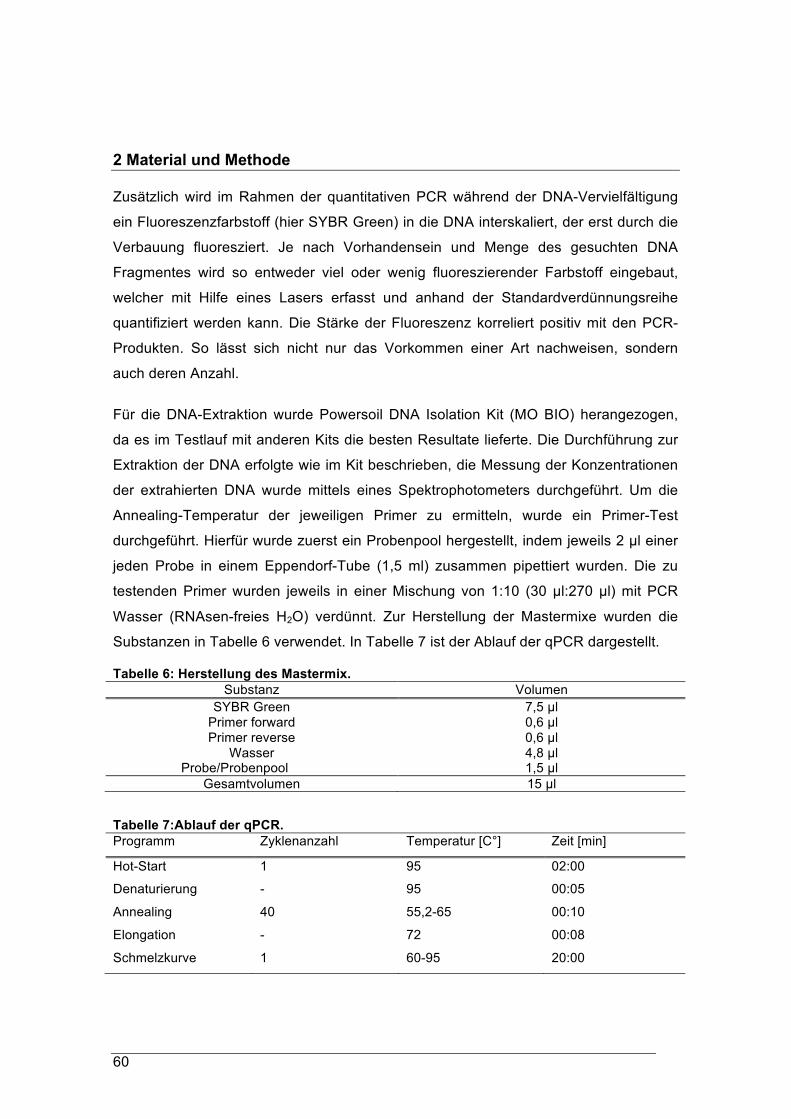
2 Material und Methode
60
Zusätzlich wird im Rahmen der quantitativen PCR während der DNA-Vervielfältigung
ein Fluoreszenzfarbstoff (hier SYBR Green) in die DNA interskaliert, der erst durch die
Verbauung fluoresziert. Je nach Vorhandensein und Menge des gesuchten DNA
Fragmentes wird so entweder viel oder wenig fluoreszierender Farbstoff eingebaut,
welcher mit Hilfe eines Lasers erfasst und anhand der Standardverdünnungsreihe
quantifiziert werden kann. Die Stärke der Fluoreszenz korreliert positiv mit den PCR-
Produkten. So lässt sich nicht nur das Vorkommen einer Art nachweisen, sondern
auch deren Anzahl.
Für die DNA-Extraktion wurde Powersoil DNA Isolation Kit (MO BIO) herangezogen,
da es im Testlauf mit anderen Kits die besten Resultate lieferte. Die Durchführung zur
Extraktion der DNA erfolgte wie im Kit beschrieben, die Messung der Konzentrationen
der extrahierten DNA wurde mittels eines Spektrophotometers durchgeführt. Um die
Annealing-Temperatur der jeweiligen Primer zu ermitteln, wurde ein Primer-Test
durchgeführt. Hierfür wurde zuerst ein Probenpool hergestellt, indem jeweils 2 µl einer
jeden Probe in einem Eppendorf-Tube (1,5 ml) zusammen pipettiert wurden. Die zu
testenden Primer wurden jeweils in einer Mischung von 1:10 (30 µl:270 µl) mit PCR
Wasser (RNAsen-freies H2O) verdünnt. Zur Herstellung der Mastermixe wurden die
Substanzen in Tabelle 6 verwendet. In Tabelle 7 ist der Ablauf der qPCR dargestellt.
Tabelle 6: Herstellung des Mastermix. Substanz Volumen
SYBR Green 7,5 µl Primer forward 0,6 µl Primer reverse 0,6 µl
Wasser 4,8 µl Probe/Probenpool 1,5 µl
Gesamtvolumen 15 µl Tabelle 7:Ablauf der qPCR. Programm Zyklenanzahl Temperatur [C°] Zeit [min]
Hot-Start 1 95 02:00
Denaturierung - 95 00:05
Annealing 40 55,2-65 00:10
Elongation - 72 00:08
Schmelzkurve 1 60-95 20:00

2 Material und Methode
61
Für das Annealing-Programm wurde ein Temperaturgradient von 55 - 65 °C verwen-
det, wobei sich die Wells absteigend innerhalb einer Spalte jeweils um 0,8 °C unter-
schieden. So konnte anhand der Schmelzkurven der verschiedenen Primer der Peak,
welcher am eindeutigsten war, ermittelt sowie die Temperatur des Peaks als Anneal-
ing-Temperatur des jeweiligen Primerpaares festgelegt werden. In Tabelle 6 sind die
getesteten Primer und deren jeweiligen Annealing-Temperaturen aufgelistet.
Tabelle 6: Verwendete Primer und ihre Annealing-Temperaturen. Primer Temperatur [C°]
Anaerobe Pilze 62
Protozoen 60
Fibrobacter succinogenes 55,3/61
Ruminococcus flavefaciens 55,3/60
Allgemeine Bakterien 64
Archaea 64
Proteolytische Bakterien α 61
Entodinium 61
Um eine quantitative Bestimmung der DNA zu ermöglichen, musste für jedes Primer-
paar ein Standard hergestellt werden. Dies erfolgte mit Hilfe des MinElute PCR Purifi-
cation Kits (Qiagen). Über den Algorithmus (http://www.uri.edu/ research/gsc/ re-
sources/ cndna.html) konnte die Anzahl der in den Proben enthaltenen Kopien berech-
net werden, welche daraufhin mittels Verdünnung auf eine Konzentration von 107 Ko-
pien gebracht werden mussten. Schließlich wurden die Verdünnungen auf einer 96
Well Reaktionsplatte durch die Zugabe von RNAse-freiem Wasser weiter auf eine
Standardreihe mit den Konzentrationen von 106 bis 100 eingestellt. Diese dienten bei
der Auswertung der qPCR einer absoluten Quantifizierung der amplifizierten DNA. Das
Programm der qPCR wurde wie in Tabelle 7 dargestellt mit einem Thermocycler (Ep-
pendorf realplex S Detektions-System) durchgeführt. Allgemein wurde die qPCR im
Doppelansatz durchgeführt, um fälschliche Messerwerte aufgrund von Primer-Wolken
zu eliminieren und einen Mittelwert bilden zu können. Mittels folgender Formel 30 wur-
de auf die Kopienanzahl pro 1 g Trockensubstanz zurückgerechnet:

2 Material und Methode
62
Formel 30: Berechnung der Kopienanzahl pro g Trockensubstanz.
𝑁 𝑞𝑃𝐶𝑅 ∗ 𝑉𝑑 ∗ 𝐸!𝐸𝑖𝑛𝑤𝑎𝑎𝑔𝑒 𝑇𝑆
= 𝐾𝑜𝑝𝑖𝑒𝑛𝑎𝑛𝑧𝑎ℎ𝑙
𝑔 𝑇𝑆
N qPCR = Anzahl Kopien der qPCR Vd = Verdünnung Ev = Elutionsvolumen g = Gramm TS = Trockensubstanz
2.10 Panseninhaltsanalyse (Mitschke)
Im Rahmen der Panseninhaltsanalyse (PIA) wurde der durch Öffnen des Pansens ge-
wonnene Inhalt auf seine inhaltlichen Bestandteile sowie deren prozentualen Anteil
untersucht. Ziel war es, Auskunft über die Art der aufgenommenen Äsung des Tieres
im letzten Retentionszeitraum zu erhalten. Für die Durchführung ist neben einer mak-
roskopischen auch eine mikroskopische Analyse möglich, wobei im Rahmen dieses
Projektes nur die erste Methode durchgeführt wurde. Deutlicher Vorteil dieser Vorge-
hensweise ist in der Größe der vorliegenden Pflanzenfragmente zu finden, da diese
beispielsweise im Vergleich zur Kotanalyse deutlich größer vorliegen und somit besser
bestimmbar sind. Das Vorgehen für die Untersuchung fand in Anlehnung an Onder-
scheka und Jordan (1976) statt. Für die Auswertung erfolgte eine Einteilung der gefun-
denen Äsungsarten in verschiedene Kategorien. Die Bestimmung wurde auf Gattungs-
und – sofern möglich – auf Artniveau durchgeführt (Tabelle 7). Für die Unterscheidung
der pflanzlichen Fragmente wurden morphologische Merkmale (Blattränder, Blattober-
und Blattunterflächen, Aderverlauf, Behaarung, Stacheln/Dornen) mittels Okular analy-
siert. Für die Bestimmung der Fragmente wurde verschiedene Bestimmungsliteratur
(Golte-Brechtle et al. 2008, Lüder 2013, Jäger 2011, Schmeil & Fitschen 2011) hinzu-
gezogen.

2 Material und Methode
63
Tabelle 7: Unterteilung der Äsungsarten nach logischen Kategorien in Anlehnung an Klansek et al. (1995). bzw. Klansek (2004).
Äsungsart Kategorie Weichäsung Kräuter
Sauergräser
Stauden
Süßgräser Zähäsung Laubhölzer
Nadelhölzer
Sträucher Anthropogen
Kirrung
Landwirtschaftliche Erzeugnisse Feldfrüchte
Kryptogamen Farne
Flechten
Moose
Pilze Baumfrüchte Früchte
Samen
Da die Inhaltsproben des Pansens Bestandteile verschiedener Größe beinhalteten,
mussten in einem ersten Schritt die festen Inhalte der Größe nach getrennt und der
noch vorhandene Pansensaft ausgewaschen werden. Dieser Schritt fand mit Hilfe von
drei Analysesieben statt, welche eine Maschenweite von 6,3 mm (Sieb 1), 3,55 mm
(Sieb 2) und 1,0 mm (Sieb 3) aufwiesen. Beim Ausspülen der Probe mit destilliertem
Wasser blieben die größten Pflanzenfragmente im obersten Sieb (Sieb 1), die übrigen
nahmen in ihrer Größe bis zum Sieb 3 ab. Die Probe wurde so lange durchgewaschen,
bis das aus Sieb 3 abfließende Wasser vollkommen klar war.
Die Nahrungsbestandteile wurden anhand der oben stehenden Kategorien unter Ver-
wendung eines Binokulars bestimmt und sortiert. Ferner wurden die sortierten Nah-
rungsbestandteile für 24 Stunden bei 60 °C im Trockenschrank bis zur Gewichts-
konstanz getrocknet. Im Anschluss wurden die Proben rückgewogen und der jeweilige
Anteil der einzelnen Kategorien an der Gesamtmenge des bestimmten Materiales be-
rechnet.

2 Material und Methode
64
Um einen Vergleich des vorgefunden Materials im Pansen sowie dem im Untersu-
chungsgebiet vorhandenem Äsungsangebot vornehmen zu können, wurde die Auftre-
tungsfrequenz einzelner Nahrungsbestandteile im Pansen je Jahreszeit anhand der
Formel 31 berechnet. Formel 31: Berechnung der Auftretungsfrequenz einzelner Nahrungsbestandteile.
𝐴𝑛𝑧𝑎ℎ𝑙 𝑒𝑖𝑛𝑒𝑠 𝑣𝑜𝑟𝑔𝑒𝑓𝑢𝑛𝑑𝑒𝑛𝑒𝑛 𝑁𝑎ℎ𝑟𝑢𝑛𝑔𝑠𝑏𝑒𝑠𝑡𝑎𝑛𝑑𝑡𝑒𝑖𝑙𝑒𝑠𝐺𝑒𝑠𝑎𝑚𝑡𝑧𝑎ℎ𝑙 𝑎𝑙𝑙𝑒𝑟 𝑃𝑎𝑛𝑠𝑒𝑛
× 100
2.11 Pflanzensoziologische Aufnahmen (Mitschke)
Die Untersuchungsgebiete wurden nach ihren unterschiedlichen Flächen, deren Vor-
kommen und Anteilen charakterisiert. Anschließend wurde ein Gitternetz für jedes Ge-
biet erstellt. Anhand einer zufälligen Stichprobenziehung mittels ArcGIS wurde nach
Caspers 2013 eine Mindestanzahl an Kartierungspunkten für jedes Untersuchungsge-
biet ausgewählt. Diese Kartierungspunkte wurden in den Gebieten markiert und in al-
len zwölf Monaten kartiert. Die Kartierungspunkte wurden mittels GPS-Gerät aufge-
sucht. An jedem Punkt wurden im Radius von 0,5 m2 die vorkommenden Pflanzenar-
ten, Häufigkeiten und Vitalität/Qualität (1 = gering bis 4 = sehr vital) der Vegetation
aufgenommen. Dies geschah zur Prüfung der Nahrungsverfügbarkeit in den unter-
schiedlichen Habitaten der Untersuchungstiere und sollte ein besseres Verständnis
sowie eine bessere Einordung der makroskopischen Ergebnisse der Panseninhaltsan-
alysen ermöglichen. Für die Auswertung der pflanzensoziologischen Aufnahmen wurde
die meteorologische Einteilung der Jahreszeiten verwendet (Tabelle 2). Ferner erfolgte
eine Zusammenfassung der kartierten Vegetation in Anlehnung an die Äsungskatego-
rien der Panseninhaltsanalyse.
Um einen Vergleich der im Gebiet vorgefundenen Vegetation sowie der durch die Pan-
seninhaltsanalyse identifizierten Nahrungsbestandteile ziehen zu können, wurden be-
züglich der kartierten Pflanzen Auftretungsfrequenzen ermittelt. Diese Berechnung
wurde anhand der Formel 32 für jede Jahreszeit vorgenommen.
Formel 32: Berechnung der Auftretungsfrequenz vorgefundener Vegetation. 𝐴𝑛𝑧𝑎ℎ𝑙 𝑑𝑒𝑟 𝑣𝑜𝑟𝑔𝑒𝑓𝑢𝑛𝑑𝑒𝑛𝑒𝑛 𝑃𝑓𝑙𝑎𝑛𝑧𝑒𝑛𝑎𝑟𝑡𝐺𝑒𝑠𝑎𝑚𝑡𝑧𝑎ℎ𝑙 𝑎𝑙𝑙𝑒𝑟 𝑘𝑎𝑟𝑡𝑖𝑒𝑟𝑡𝑒𝑟 𝑃𝑓𝑙𝑎𝑛𝑧𝑒𝑛
× 100

2 Material und Methode
65
2.12 Analyse zum Stress (Scheingraber)
Ziel dieser Analysen war es, mögliche Stressbelastungen der beiden verschiedenen
Habitate zu erfassen. Stressreaktionen bei Tieren lassen sich über verschiedenste
Methoden messen. Die hier verwendet Methode ist die Messung von Glukokortikoid-
metaboliten im Kot der Untersuchungstiere. Erleiden Tiere Stress, schüttet ihr Körper
vermehrt Kortisol aus. Dieses dient zur Aktivierung kataboler Stoffwechselvorgänge.
Später gelangt das zu den Glucocorticoiden gehörende Hormon über den Blutkreislauf
in die Leber und wird dort verstoffwechselt. Die dabei entstehenden Metaboliten (Glu-
kokortikoidmetaboliten = GCM) werden über den Kot des Tieres ausgeschieden. Der
Ausscheidungsprozess dauert je nach Tierart mehrere Stunden. Beim Reh gelangen
die GCMs ca. binnen 10 - 14 Stunden in den Kot (Holand 1993, Behrend 1999, Weil-
nböck 2013). Über die Mengen der ausgeschiedenen Glukokortikoidmetaboliten im Kot
lassen sich Aussagen über die Intensitäten der verursachten Stressoren auf die Tiere
treffen. Der Kot wurde den Tieren unmittelbar nach Erlegung aus dem Rektum entfernt
und sofort bei -20 °C eingefroren, um eine mögliche Verfälschung der Werte durch
weitere Metabolisierung der GCMs durch Bakterien zu verhindern. Die GCM-Analysen
erfolgten mittels Enzymimmunoassays (EIA) an Kotproben von 130 Tieren gemäß des
EIA Protokolls von Palme und Möstl, Stand Juli 2014. Die Messung Glukokortikoidme-
taboliten erfolgte im Photometer mittels eines Messfilters bei 450 nm. Formel 33 gibt
die Berechnung der GCM-Konzentration wieder.
Formel 33: Berechnung der GCM-Konzentration von pg/Well in ng/g Kot.
𝑛𝑔 (𝑆𝑡𝑒𝑟𝑜𝑖𝑑)𝑔 𝐾𝑜𝑡 =
𝑝𝑔 𝑝𝑟𝑜 𝑊𝑒𝑙𝑙 × 𝐸! 𝜇𝑙 × 𝑉!𝑃! 𝑔 × 𝑃! 𝜇𝑙 × 1000
Ev = Extraktionsvolumen
Vf = Verdünnungsfaktor
PE = Probeneinwaage (Kot)
PV = Probenvolumen

2 Material und Methode
66
2.13 Datenauswertung (Scheingraber)
Alle Daten wurden mit Hilfe des Shapiro-Wilk-Test auf Normalverteilung sowie mit dem
Levené-Test auf Varianzhomogenität überprüft. Signifikanzen wurden je nach Vertei-
lung der Daten und ihrer Gruppengröße getestet. Hierfür wurde für normalverteilte Da-
ten der t-Test für unverbundene oder verbundene Stichproben und für mehr als zwei
Gruppen die einfaktorielle Varianzanalyse verwendet. Bei nicht normalverteilten unver-
bundenen Daten wurde der Mann-Whitney-U-Test und bei mehr als zwei Gruppen die
Rangvarianzanalyse verwendet. Bei Daten, die miteinander verbunden waren, wurde
der Wilcoxon Test und bei mehr als zwei Gruppen der Friedman Test angewendet. Bei
Prüfung auf signifikante Unterschiede zweier nominalskalierten Daten wurden für klei-
ne Stichproben der Exakte Fischer Test und bei größeren Stickproben der Chi-Quadrat
Test verwendet. Bei multiplen Testanwendungen wurden zur Vermeidung der Alpha-
Fehler-Kumulierung Post hoc Tests der U-Test und/oder der Tukey Test (je nach Ver-
teilung der Daten) mit anschließender Bonferroni Korrektur verwendet. Je nach Art der
Verteilung wurden Korrelationen nach Pearson oder Spearman Rho getestet. Als Sig-
nifikanzniveau wurde α = 1 % für Korrelationen und für alle übrigen statistischen Tests
α = 5 % angenommen. Alle Werte wurden als Mittelwerte (x̄) mit Standardabweichung
SD bei Normalverteilung oder als Mediane (x̂) bei nicht normalverteilten Daten mit 95%
Konfidenzintervallen angegeben.
Ausnahmen sind Grafiken und Beschreibungen der Daten mit Anzahl (n) oder in Pro-
zent (%).
Für die Darstellung der Mikrobiom-Ergebnisse wurden für eine einheitliche Skalierung
die Daten logarithmiert.
Zur Berechnungen und Erstellung der Grafiken wurde das Statistikprogramm SPSS
(Version 22.0.0.0) verwendet.

3 Ergebnisse
67
3 Ergebnisse
3.1 Energiegehalt der Rehnahrung (Scheingraber)
3.1.1 Energiegehalt der Rehnahrung auf der Basis des Standardverfahrens
Die Energieberechnungen auf der Basis von Hohenheimer Futterwerttest und Weender
Verfahren zeigen signifikante Unterschiede zwischen dem Energiegehalt der Nahrung
hinsichtlich der beiden Habitate Land und Wald (siehe Abbildung 16). Der durchschnitt-
liche umsetzbare Energiegehalt (ME) der Rehäsung liegt mit 6,29 MJ/kg TS im Land
sign. (p < 0,001) über dem durchschnittlichen Energiegehalt (ME) im Wald von 5,43
MJ/kg TS.
Abbildung 16: Median des umsetzbaren Energiegehaltes (ME) der Rehäsung [MJ/kg TS] beider Habitate (n = 220, p < 0,001)
Differenziert man die Energiewerte (ME) nach Monaten, finden sich außer im Mai, in
allen Monaten höhere Energiewerte im „Land“-Habitat im Vergleich zum „Wald“-Habitat
(Abbildung 17). Gerade die für die Populationsentwicklung wichtigen Monate Januar
LandWald
ME
[MJ/
kg T
S]
12.00
10.00
8.00
6.00
4.00
2.00
Seite 1
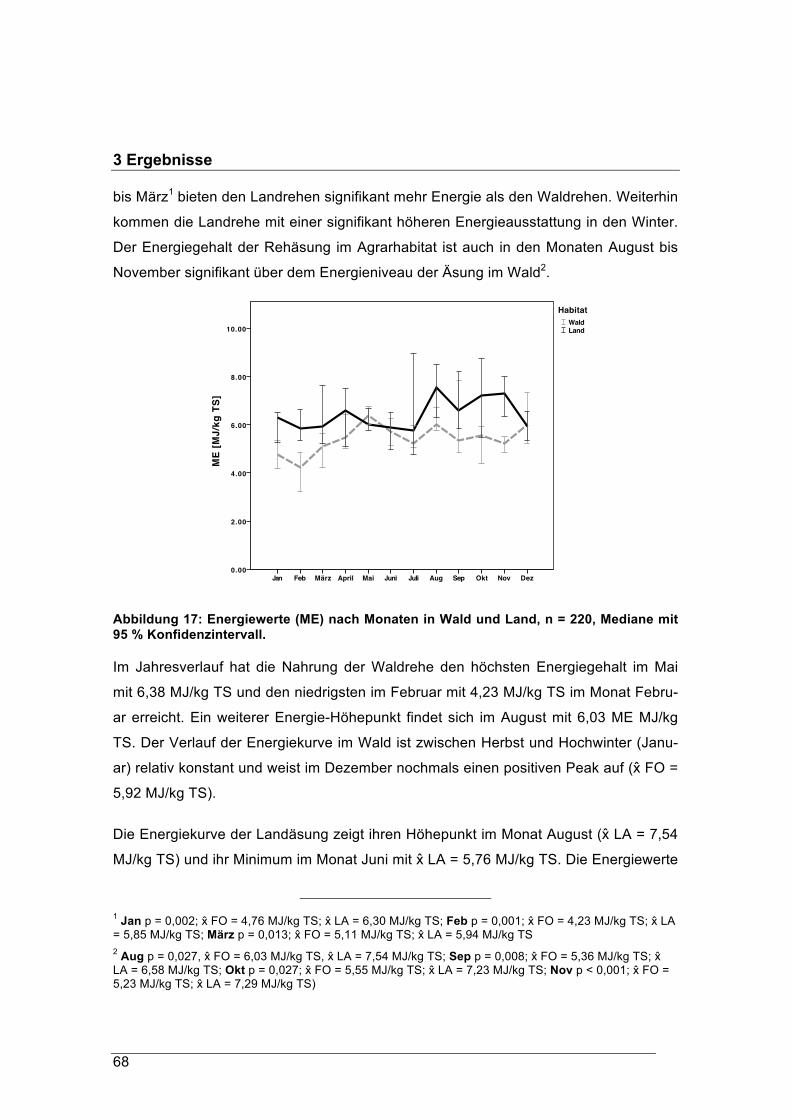
3 Ergebnisse
68
bis März1 bieten den Landrehen signifikant mehr Energie als den Waldrehen. Weiterhin
kommen die Landrehe mit einer signifikant höheren Energieausstattung in den Winter.
Der Energiegehalt der Rehäsung im Agrarhabitat ist auch in den Monaten August bis
November signifikant über dem Energieniveau der Äsung im Wald2.
Abbildung 17: Energiewerte (ME) nach Monaten in Wald und Land, n = 220, Mediane mit 95 % Konfidenzintervall.
Im Jahresverlauf hat die Nahrung der Waldrehe den höchsten Energiegehalt im Mai
mit 6,38 MJ/kg TS und den niedrigsten im Februar mit 4,23 MJ/kg TS im Monat Febru-
ar erreicht. Ein weiterer Energie-Höhepunkt findet sich im August mit 6,03 ME MJ/kg
TS. Der Verlauf der Energiekurve im Wald ist zwischen Herbst und Hochwinter (Janu-
ar) relativ konstant und weist im Dezember nochmals einen positiven Peak auf (x̂ FO =
5,92 MJ/kg TS).
Die Energiekurve der Landäsung zeigt ihren Höhepunkt im Monat August (x̂ LA = 7,54
MJ/kg TS) und ihr Minimum im Monat Juni mit x̂ LA = 5,76 MJ/kg TS. Die Energiewerte
1 Jan p = 0,002; x̂ FO = 4,76 MJ/kg TS; x̂ LA = 6,30 MJ/kg TS; Feb p = 0,001; x̂ FO = 4,23 MJ/kg TS; x̂ LA = 5,85 MJ/kg TS; März p = 0,013; x̂ FO = 5,11 MJ/kg TS; x̂ LA = 5,94 MJ/kg TS 2 Aug p = 0,027, x̂ FO = 6,03 MJ/kg TS, x̂ LA = 7,54 MJ/kg TS; Sep p = 0,008; x̂ FO = 5,36 MJ/kg TS; x̂ LA = 6,58 MJ/kg TS; Okt p = 0,027; x̂ FO = 5,55 MJ/kg TS; x̂ LA = 7,23 MJ/kg TS; Nov p < 0,001; x̂ FO = 5,23 MJ/kg TS; x̂ LA = 7,29 MJ/kg TS)
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
ME
[MJ/
kg T
S]
10.00
8.00
6.00
4.00
2.00
0.00
LandWald
Habitat
Seite 1
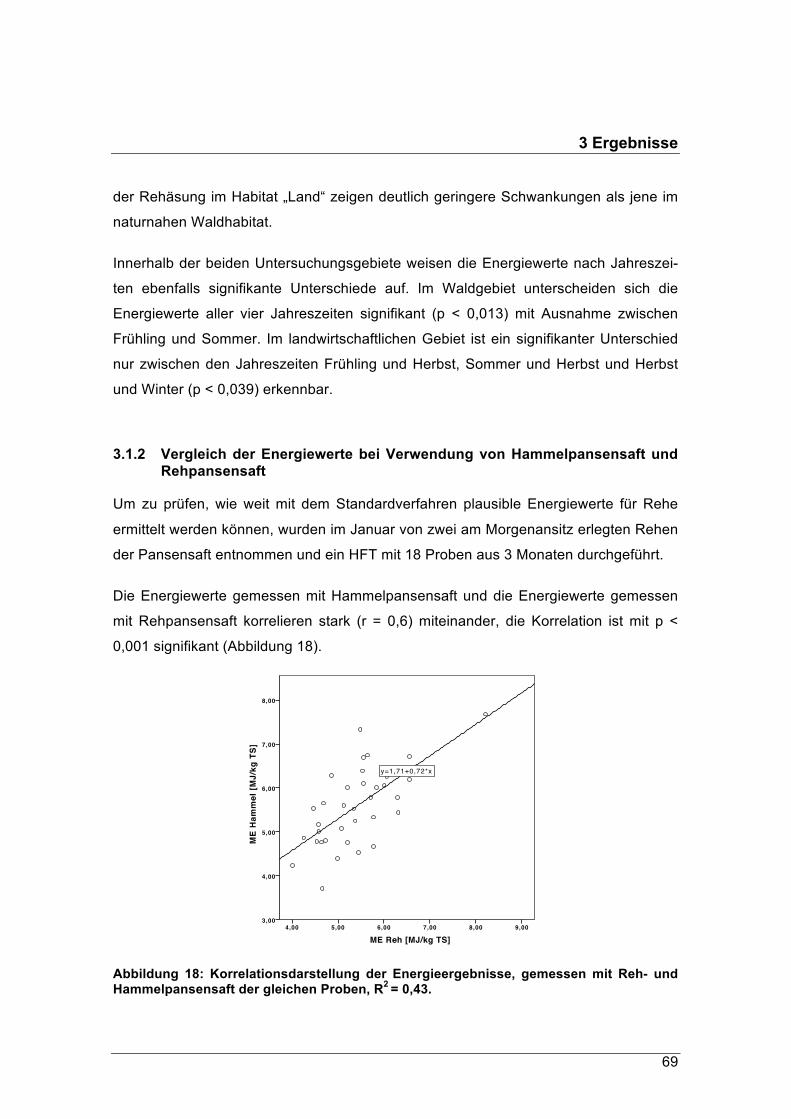
3 Ergebnisse
69
der Rehäsung im Habitat „Land“ zeigen deutlich geringere Schwankungen als jene im
naturnahen Waldhabitat.
Innerhalb der beiden Untersuchungsgebiete weisen die Energiewerte nach Jahreszei-
ten ebenfalls signifikante Unterschiede auf. Im Waldgebiet unterscheiden sich die
Energiewerte aller vier Jahreszeiten signifikant (p < 0,013) mit Ausnahme zwischen
Frühling und Sommer. Im landwirtschaftlichen Gebiet ist ein signifikanter Unterschied
nur zwischen den Jahreszeiten Frühling und Herbst, Sommer und Herbst und Herbst
und Winter (p < 0,039) erkennbar.
3.1.2 Vergleich der Energiewerte bei Verwendung von Hammelpansensaft und Rehpansensaft
Um zu prüfen, wie weit mit dem Standardverfahren plausible Energiewerte für Rehe
ermittelt werden können, wurden im Januar von zwei am Morgenansitz erlegten Rehen
der Pansensaft entnommen und ein HFT mit 18 Proben aus 3 Monaten durchgeführt.
Die Energiewerte gemessen mit Hammelpansensaft und die Energiewerte gemessen
mit Rehpansensaft korrelieren stark (r = 0,6) miteinander, die Korrelation ist mit p <
0,001 signifikant (Abbildung 18).
Abbildung 18: Korrelationsdarstellung der Energieergebnisse, gemessen mit Reh- und Hammelpansensaft der gleichen Proben, R2 = 0,43.
ME Reh [MJ/kg TS]9,008,007,006,005,004,00
ME
Ham
mel
[MJ/
kg T
S]
8 ,00
7,00
6,00
5,00
4,00
3,00
y=1,71+0,72*x
R2 Linear = 0,433
Seite 1
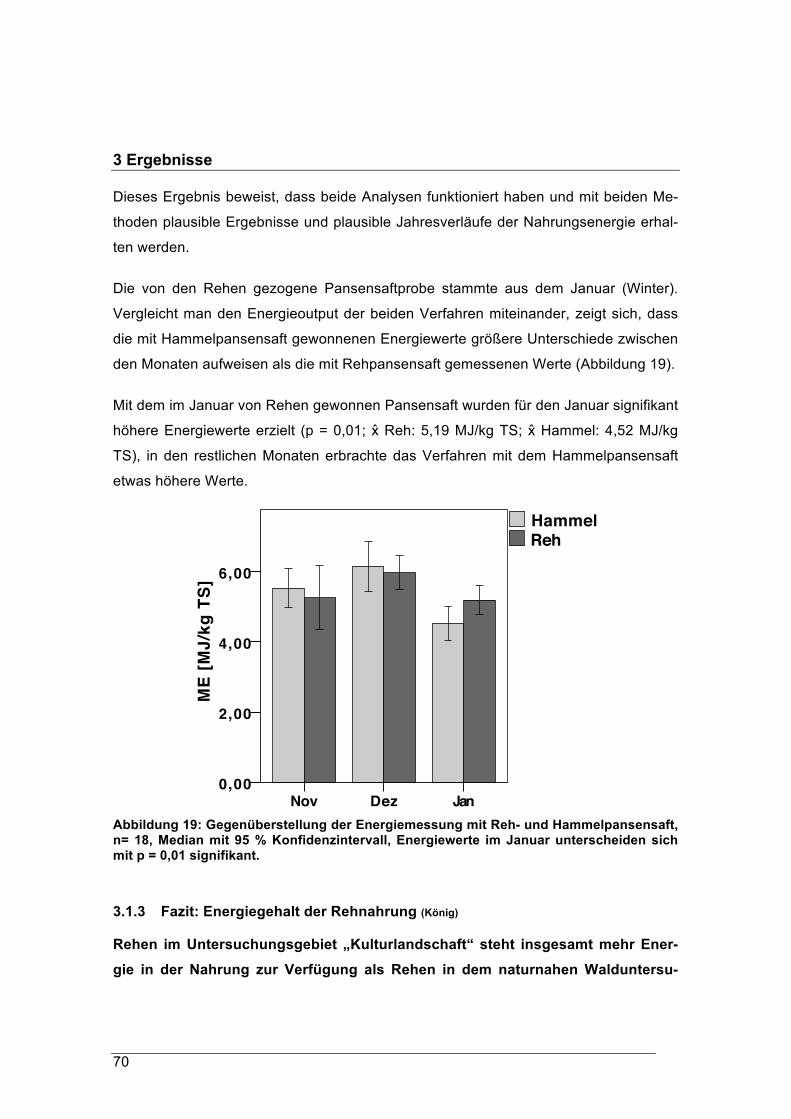
3 Ergebnisse
70
Dieses Ergebnis beweist, dass beide Analysen funktioniert haben und mit beiden Me-
thoden plausible Ergebnisse und plausible Jahresverläufe der Nahrungsenergie erhal-
ten werden.
Die von den Rehen gezogene Pansensaftprobe stammte aus dem Januar (Winter).
Vergleicht man den Energieoutput der beiden Verfahren miteinander, zeigt sich, dass
die mit Hammelpansensaft gewonnenen Energiewerte größere Unterschiede zwischen
den Monaten aufweisen als die mit Rehpansensaft gemessenen Werte (Abbildung 19).
Mit dem im Januar von Rehen gewonnen Pansensaft wurden für den Januar signifikant
höhere Energiewerte erzielt (p = 0,01; x̂ Reh: 5,19 MJ/kg TS; x̂ Hammel: 4,52 MJ/kg
TS), in den restlichen Monaten erbrachte das Verfahren mit dem Hammelpansensaft
etwas höhere Werte.
Abbildung 19: Gegenüberstellung der Energiemessung mit Reh- und Hammelpansensaft, n= 18, Median mit 95 % Konfidenzintervall, Energiewerte im Januar unterscheiden sich mit p = 0,01 signifikant.
3.1.3 Fazit: Energiegehalt der Rehnahrung (König)
Rehen im Untersuchungsgebiet „Kulturlandschaft“ steht insgesamt mehr Ener-
gie in der Nahrung zur Verfügung als Rehen in dem naturnahen Walduntersu-
JanDezNov
ME
[MJ/
kg T
S]
6 ,00
4,00
2,00
0,00
Reh Hammel
Page 1

3 Ergebnisse
71
chungsgebiet. Die größten Energieunterschiede zwischen den beiden Untersu-
chungsgebieten finden sich im Herbst und im Winter / Frühjahr. Die geringsten
Energiewerte wurden im Land im September und im Wald im Februar ermittelt.
Für die Berechnung der Energie von Rehäsung eignet sich das Standardverfah-
ren des Hohenheimer Futterwerttests basierend auf Hammelpansensaft gut.
Scheinbar können Rehe ebenso wie Rotwild gerade in der energiearmen Zeit im
Hochwinter durch ihre Anpassungsfähigkeit an die natürliche Äsung aus dieser
mehr Energie gewinnen als domestizierte Wiederkäuer. Im Winter wird mit den
Standardverfahren der Futtermittelanalyse der mögliche Energiewert der Nah-
rung für Rehe und Rotwild unterschätzt.
3.2 Qualität der Rehnahrung (Scheingraber)
3.2.1 Qualität des festen Panseninhaltes
3.2.1.1 Qualität des Raufutters
Die Weender-Analyse zeigt die Rohnährstoffe der sich im Pansen befindlichen Nah-
rung auf. Bei Betrachtung der beiden Habitate sind signifikante Unterschiede der
Rohnährstoffgehalte ersichtlich (Abbildung 20). Die schwerverdaulichen Anteile an Ro-
hasche (XA; FO: 8,34 %; LA: 7,47; p = 0,001) und Rohfasern (XF: x̂ FO = 29,61 %; x̂
LA = 24,99 %; p < 0,001) sind im Wald signifikant höher als im Land.
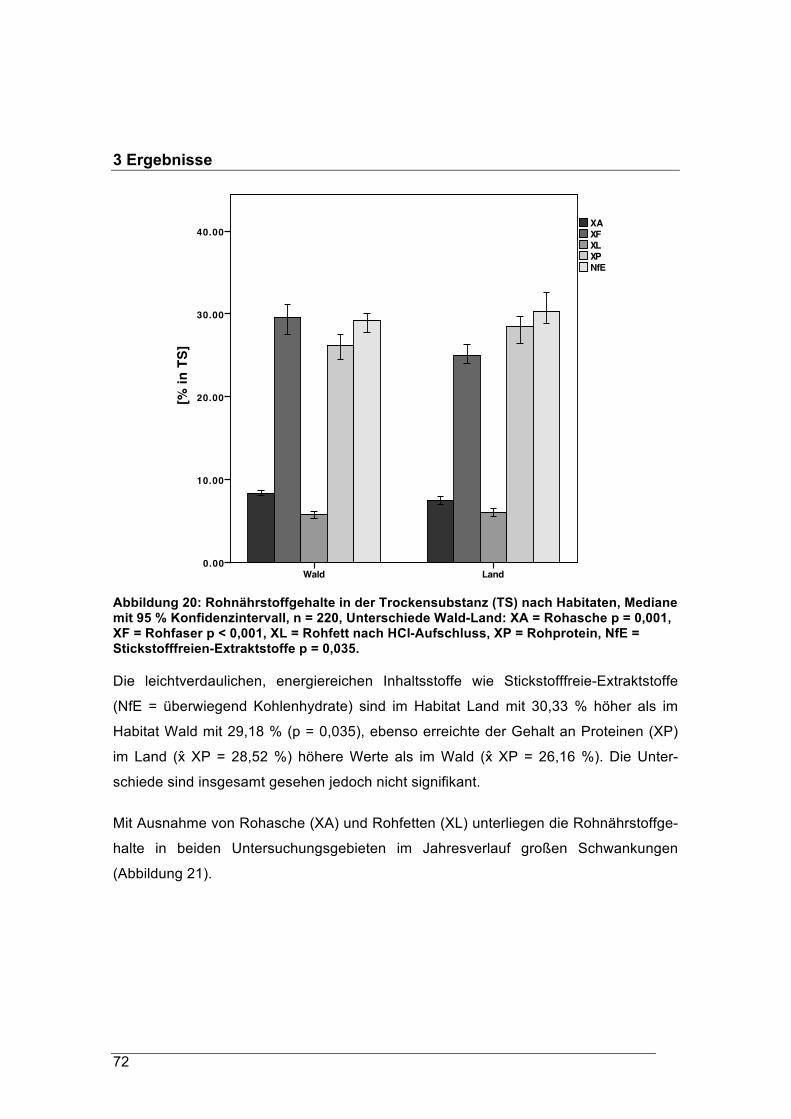
3 Ergebnisse
72
Abbildung 20: Rohnährstoffgehalte in der Trockensubstanz (TS) nach Habitaten, Mediane mit 95 % Konfidenzintervall, n = 220, Unterschiede Wald-Land: XA = Rohasche p = 0,001, XF = Rohfaser p < 0,001, XL = Rohfett nach HCl-Aufschluss, XP = Rohprotein, NfE = Stickstofffreien-Extraktstoffe p = 0,035.
Die leichtverdaulichen, energiereichen Inhaltsstoffe wie Stickstofffreie-Extraktstoffe
(NfE = überwiegend Kohlenhydrate) sind im Habitat Land mit 30,33 % höher als im
Habitat Wald mit 29,18 % (p = 0,035), ebenso erreichte der Gehalt an Proteinen (XP)
im Land (x̂ XP = 28,52 %) höhere Werte als im Wald (x̂ XP = 26,16 %). Die Unter-
schiede sind insgesamt gesehen jedoch nicht signifikant.
Mit Ausnahme von Rohasche (XA) und Rohfetten (XL) unterliegen die Rohnährstoffge-
halte in beiden Untersuchungsgebieten im Jahresverlauf großen Schwankungen
(Abbildung 21).
LandWald
[% in
TS]
40.00
30.00
20.00
10.00
0.00
NfEXPXLXFXA
Page 1
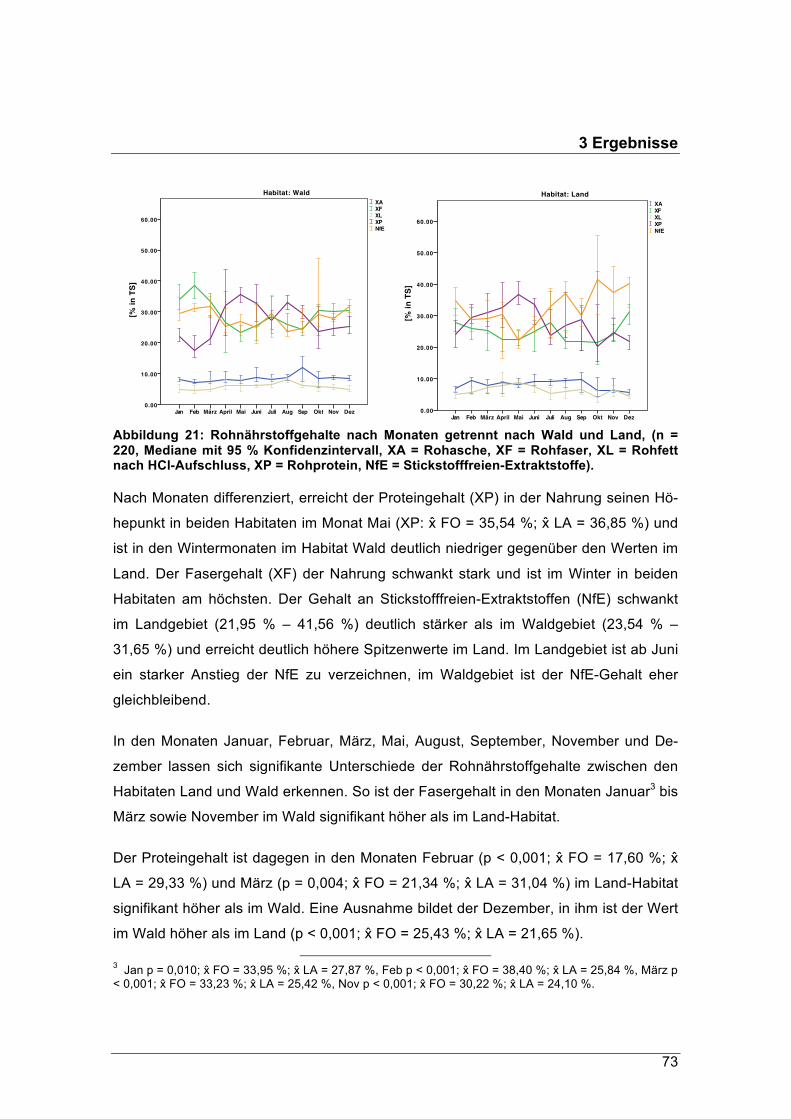
3 Ergebnisse
73
Abbildung 21: Rohnährstoffgehalte nach Monaten getrennt nach Wald und Land, (n = 220, Mediane mit 95 % Konfidenzintervall, XA = Rohasche, XF = Rohfaser, XL = Rohfett nach HCl-Aufschluss, XP = Rohprotein, NfE = Stickstofffreien-Extraktstoffe).
Nach Monaten differenziert, erreicht der Proteingehalt (XP) in der Nahrung seinen Hö-
hepunkt in beiden Habitaten im Monat Mai (XP: x̂ FO = 35,54 %; x̂ LA = 36,85 %) und
ist in den Wintermonaten im Habitat Wald deutlich niedriger gegenüber den Werten im
Land. Der Fasergehalt (XF) der Nahrung schwankt stark und ist im Winter in beiden
Habitaten am höchsten. Der Gehalt an Stickstofffreien-Extraktstoffen (NfE) schwankt
im Landgebiet (21,95 % – 41,56 %) deutlich stärker als im Waldgebiet (23,54 % –
31,65 %) und erreicht deutlich höhere Spitzenwerte im Land. Im Landgebiet ist ab Juni
ein starker Anstieg der NfE zu verzeichnen, im Waldgebiet ist der NfE-Gehalt eher
gleichbleibend.
In den Monaten Januar, Februar, März, Mai, August, September, November und De-
zember lassen sich signifikante Unterschiede der Rohnährstoffgehalte zwischen den
Habitaten Land und Wald erkennen. So ist der Fasergehalt in den Monaten Januar3 bis
März sowie November im Wald signifikant höher als im Land-Habitat.
Der Proteingehalt ist dagegen in den Monaten Februar (p < 0,001; x̂ FO = 17,60 %; x̂
LA = 29,33 %) und März (p = 0,004; x̂ FO = 21,34 %; x̂ LA = 31,04 %) im Land-Habitat
signifikant höher als im Wald. Eine Ausnahme bildet der Dezember, in ihm ist der Wert
im Wald höher als im Land (p < 0,001; x̂ FO = 25,43 %; x̂ LA = 21,65 %).
3 Jan p = 0,010; x̂ FO = 33,95 %; x̂ LA = 27,87 %, Feb p < 0,001; x̂ FO = 38,40 %; x̂ LA = 25,84 %, März p < 0,001; x̂ FO = 33,23 %; x̂ LA = 25,42 %, Nov p < 0,001; x̂ FO = 30,22 %; x̂ LA = 24,10 %.
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
[% in
TS]
60.00
50.00
40.00
30.00
20.00
10.00
0.00
NfEXP XL XF XA
Habitat: Wald
Seite 1
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan[%
in T
S]
60.00
50.00
40.00
30.00
20.00
10.00
0.00
NfE XP XL XFXA
Habitat: Land
Seite 1

3 Ergebnisse
74
Der Gehalt an Stickstofffreien-Extraktstoffen ist im Wald in den Monaten Februar (p =
0,028; x̂ FO = 31,01 %; x̂ LA = 28,75 %) und Mai (p = 0,031; x̂ FO = 26,78 %; x̂ LA =
21,96 %) signifikant höher als im Land. Umgekehrt ist dieser Wert in den Monaten Ja-
nuar4, August, November und Dezember im Land signifikant höher.
Die Werte der Rohasche (XA) und des Rohfetts (XF) zeigen geringe Schwankungen
und Unterschiede zwischen den Habitaten, allerdings liegt der Rohfettgehalt in den
Monaten Februar (p = 0,010; x̂ FO = 4,42 %; x̂ LA = 5,59 %) und Mai (p < 0,001; x̂ FO
= 6,07 %; x̂ LA = 8,64 %) im Landhabitat signifikant über dem im Waldhabitat.
Bei der Untersuchung signifikanter Unterschiede zwischen den vier Jahreszeiten in-
nerhalb der beiden Habitate zeigte das Wald-Habitat bei den Rohnährstoffen Roha-
sche (p < 0,001), Rohfaser (p < 0,001), Rohfett (p = 0,001), Rohprotein (p < 0,001) und
NfE (p = 0,013) zwischen den Jahreszeiten statistische Unterschiede. Ähnliche Unter-
schiede fanden sich in den Rohnährstoffen im Land-Habitat. Auch hier ließen sich hin-
sichtlich der vier Jahreszeiten bei Rohasche (p < 0,001), Rohfaser (p = 0,001), Rohfett
(p < 0,001), Rohprotein (p < 0,001) und NfE (p < 0,001) signifikante Unterschiede zwi-
schen allen Rohnährstoffgruppen feststellen.
3.2.1.2 Detailanalyse der Fasern nach VanSoest
Die VanSoest-Analyse erlaubt eine über das Weender-Verfahren hinausgehende de-
tailliertere Auftrennung der verschiedenen Faseranteile der aufgenommenen Nahrung.
Zellulose- (p = 0,042, x̂ FO = 17,79 %; x̂ LA = 15,58 %) und Lignin- (p < 0,001; x̂ FO =
16,45 %; x̂ LA = 12,81 %) Anteile der Nahrung sind signifikant höher bei den Proben im
Wald-Habitat gegenüber denen des Land-Habitats (Abbildung 22). Die Anteile der He-
mizellulose sind über den gesamten Untersuchungszeitraum betrachtet zwischen den
beiden Untersuchungsgebieten nicht signifikant unterschiedlich.
4 Jan p = 0,031; x̂ FO = 29,51 %; x̂ LA = 34,80 %, Aug p = 0,027; x̂ FO = 23,55 %; x̂ LA = 37,14 %, Nov p=0,006; x̂ FO = 27,78 %; x̂ LA = 37,15 %, Dez p = 0,014; x̂ FO = 31,36 %; x̂ LA = 40,15 %.
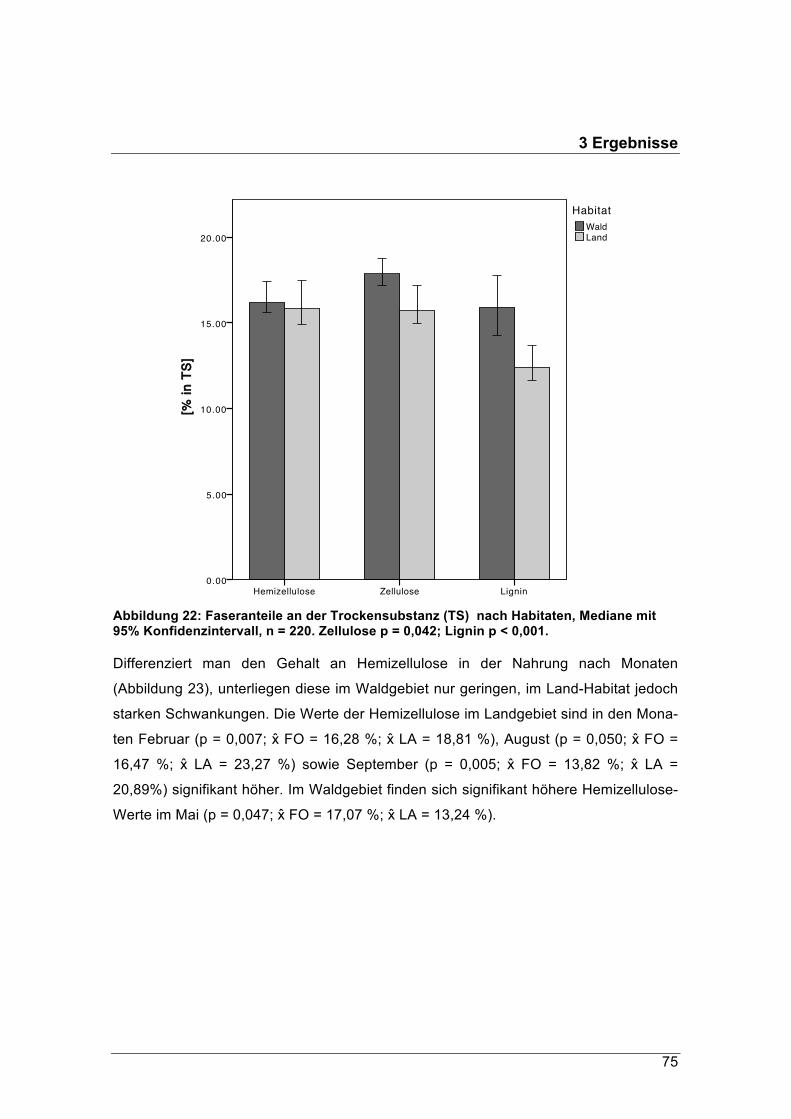
3 Ergebnisse
75
Abbildung 22: Faseranteile an der Trockensubstanz (TS) nach Habitaten, Mediane mit 95% Konfidenzintervall, n = 220. Zellulose p = 0,042; Lignin p < 0,001.
Differenziert man den Gehalt an Hemizellulose in der Nahrung nach Monaten
(Abbildung 23), unterliegen diese im Waldgebiet nur geringen, im Land-Habitat jedoch
starken Schwankungen. Die Werte der Hemizellulose im Landgebiet sind in den Mona-
ten Februar (p = 0,007; x̂ FO = 16,28 %; x̂ LA = 18,81 %), August (p = 0,050; x̂ FO =
16,47 %; x̂ LA = 23,27 %) sowie September (p = 0,005; x̂ FO = 13,82 %; x̂ LA =
20,89%) signifikant höher. Im Waldgebiet finden sich signifikant höhere Hemizellulose-
Werte im Mai (p = 0,047; x̂ FO = 17,07 %; x̂ LA = 13,24 %).
Lignin Zellulose Hemizellulose
[% in
TS]
20.00
15.00
10.00
5.00
0.00
LandWald
Habitat
Seite 1
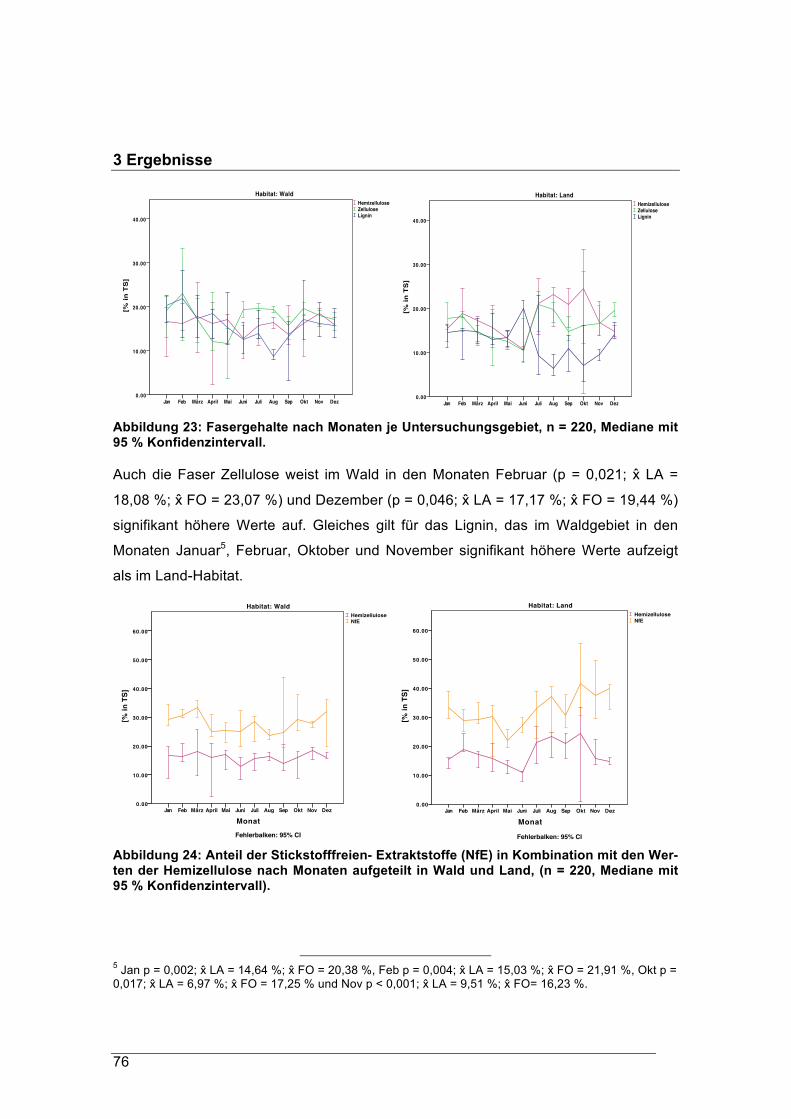
3 Ergebnisse
76
Abbildung 23: Fasergehalte nach Monaten je Untersuchungsgebiet, n = 220, Mediane mit 95 % Konfidenzintervall.
Auch die Faser Zellulose weist im Wald in den Monaten Februar (p = 0,021; x̂ LA =
18,08 %; x̂ FO = 23,07 %) und Dezember (p = 0,046; x̂ LA = 17,17 %; x̂ FO = 19,44 %)
signifikant höhere Werte auf. Gleiches gilt für das Lignin, das im Waldgebiet in den
Monaten Januar5, Februar, Oktober und November signifikant höhere Werte aufzeigt
als im Land-Habitat.
Abbildung 24: Anteil der Stickstofffreien- Extraktstoffe (NfE) in Kombination mit den Wer-ten der Hemizellulose nach Monaten aufgeteilt in Wald und Land, (n = 220, Mediane mit 95 % Konfidenzintervall).
5 Jan p = 0,002; x̂ LA = 14,64 %; x̂ FO = 20,38 %, Feb p = 0,004; x̂ LA = 15,03 %; x̂ FO = 21,91 %, Okt p = 0,017; x̂ LA = 6,97 %; x̂ FO = 17,25 % und Nov p < 0,001; x̂ LA = 9,51 %; x̂ FO= 16,23 %.
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
[% in
TS
]
40.00
30.00
20.00
10.00
0.00
Lignin Zellulose Hemizellulose
Habitat: Wald
Seite 1
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan[%
in T
S]
40.00
30.00
20.00
10.00
0.00
LigninZelluloseHemizellulose
Habitat: Land
Seite 1
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
[% in
TS]
60.00
50.00
40.00
30.00
20.00
10.00
0.00
Fehlerbalken: 95% CI
NfE Hemizellulose
Habitat: Wald
Seite 1
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
[% in
TS]
60.00
50.00
40.00
30.00
20.00
10.00
0.00
Fehlerbalken: 95% CI
NfE Hemizellulose
Habitat: Land
Seite 1

3 Ergebnisse
77
Bei der Kombination (Abbildung 24) der Werte der Stickstofffreien-Extraktstoffe aus
dem Weender Verfahren und den Hemizellulosewerten des VanSoest Verfahrens wird
deutlich, dass beide Kurven im jeweiligen Habitat relativ synchron bzw. parallel verlau-
fen. Dies weist darauf hin, dass ein großer Anteil der Stickstofffreien-Extraktstoffe
durch den Abbau von Hemizellulose zustande kommt.
Wie bei den Rohnährstoffen zeigen sich auch bei den Faseranteilen signifikante Unter-
schiede zwischen den vier Jahreszeiten innerhalb der beiden Untersuchungsgebiete.
So findet man signifikante Schwankungen aller drei Faseranteile zwischen den vier
Jahreszeiten im Landhabitat mit Hemizellulose (p = 0,008), Zellulose (p = 0,001) und
Lignin (p = 0,001).
Im Waldgebiet ließen sich nur bei Zellulose (p < 0,001) und Lignin (p < 0,001) signifi-
kante Schwankungen zwischen den Jahreszeiten feststellen.
3.2.1.3 Verdaulichkeit
Die Verdaulichkeit wird mithilfe des HFTs und der Fasergehalte durch das VanSoest
Verfahren berechnet und gibt Auskunft über die Verdaulichkeit der vom Reh aufge-
nommenen organischen Masse. Anhand des Balkendiagramms (Abbildung 25) wird
deutlich, dass die Werte der Gesamtverdaulichkeit der organischen Masse im landwirt-
schaftlichen Gebiet mit 52,37 % signifikant höher (p < 0,001) liegen als im naturnahen
forstlich geprägten Habitat mit nur 46,86 %.
Eine Differenzierung der Verdaulichkeitswerte nach Monaten in den beiden Untersu-
chungsgebieten zeigt (Abbildung 26), dass in den Monaten6 Januar bis März, August
und Oktober sowie November die Verdaulichkeit der Äsung im Landhabitat jeweils sig-
nifikant höher ist als im Waldhabitat. Die maximale Verdaulichkeit erreicht Waldrehnah-
rung im Mai (54,80 %) und die Landrehnahrung im August (59,56 %). Die schlechteste
Verdaulichkeit weist die Äsung im Februar mit 37,22 % im Waldgebiet und im Landha-
bitat im Dezember mit 47,20 % auf. Dieser „schlechteste“ Verdaulichkeitswert im
6 Januar p = 0,004; x̄ FO: 41,69 %, x̄ LA: 49,32 %; Februar p = 0,001; x̄ FO: 37,22 %, x̄ LA: 50,64 %; März p = 0,013; x̄ FO: 43,23 %, x̄ LA: 51,79 %; August p = 0,050; x̄ FO: 52,65 %, x̄ LA: 59,56 %; Oktober p = 0,027; x̄ FO46,17 %, x̄ LA: 56,15 %; und November p < 0,001; x̄ FO: 45,58 %, x̄ LA: 56,30 %.
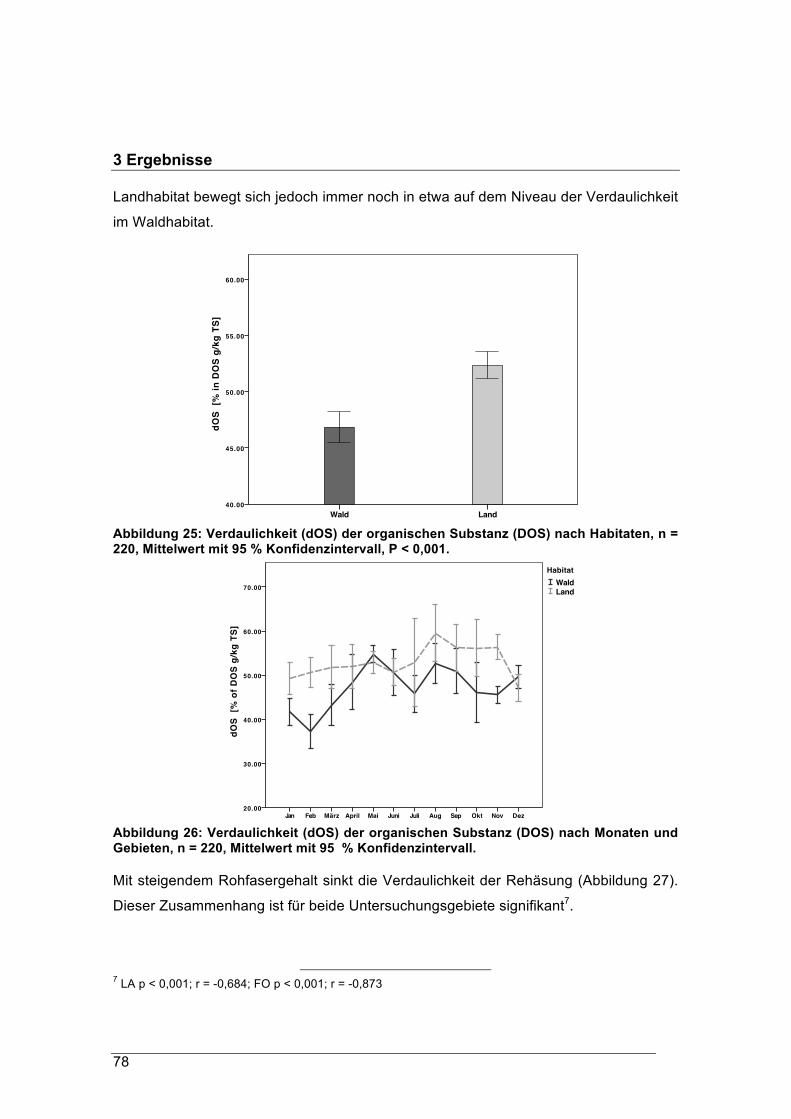
3 Ergebnisse
78
Landhabitat bewegt sich jedoch immer noch in etwa auf dem Niveau der Verdaulichkeit
im Waldhabitat.
Abbildung 25: Verdaulichkeit (dOS) der organischen Substanz (DOS) nach Habitaten, n = 220, Mittelwert mit 95 % Konfidenzintervall, P < 0,001.
Abbildung 26: Verdaulichkeit (dOS) der organischen Substanz (DOS) nach Monaten und Gebieten, n = 220, Mittelwert mit 95 % Konfidenzintervall.
Mit steigendem Rohfasergehalt sinkt die Verdaulichkeit der Rehäsung (Abbildung 27).
Dieser Zusammenhang ist für beide Untersuchungsgebiete signifikant7.
7 LA p < 0,001; r = -0,684; FO p < 0,001; r = -0,873
LandWald
dO
S [%
in D
OS
g/kg
TS]
60.00
55.00
50.00
45.00
40.00
Seite 1
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
dOS
[% o
f DO
S g/
kg T
S]
70.00
60.00
50.00
40.00
30.00
20.00
LandWald
Habitat
Page 1
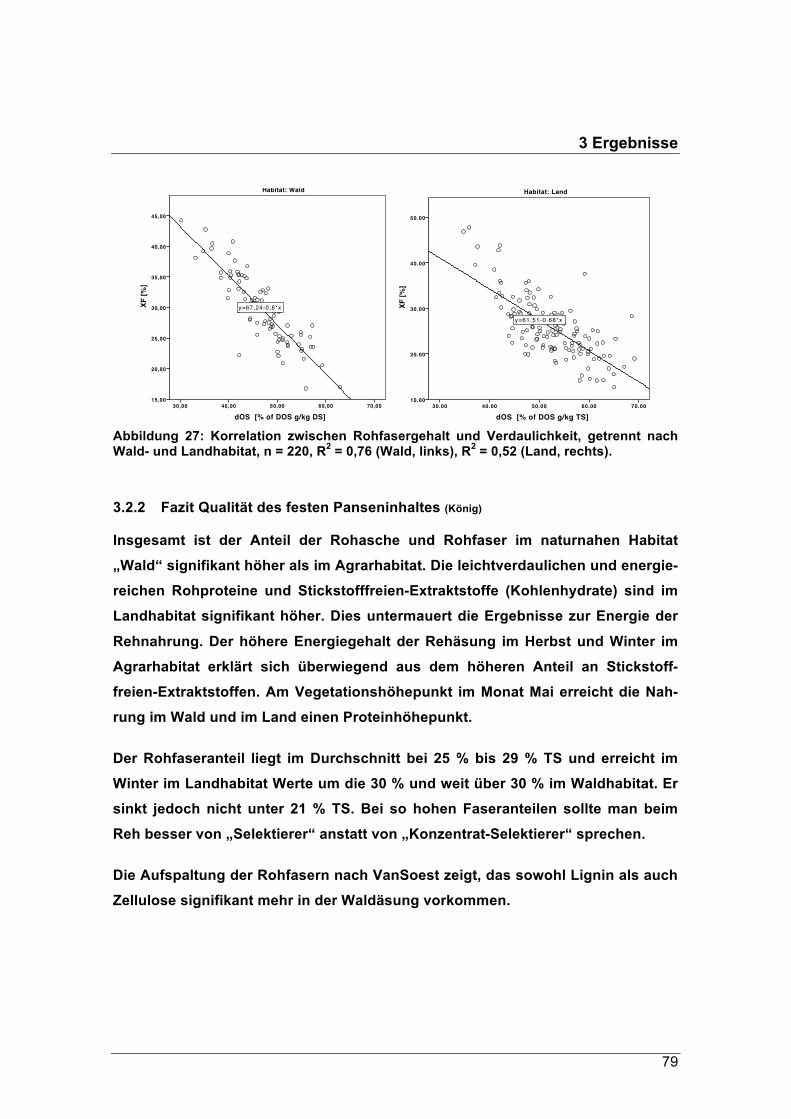
3 Ergebnisse
79
Abbildung 27: Korrelation zwischen Rohfasergehalt und Verdaulichkeit, getrennt nach Wald- und Landhabitat, n = 220, R2 = 0,76 (Wald, links), R2 = 0,52 (Land, rechts).
3.2.2 Fazit Qualität des festen Panseninhaltes (König)
Insgesamt ist der Anteil der Rohasche und Rohfaser im naturnahen Habitat
„Wald“ signifikant höher als im Agrarhabitat. Die leichtverdaulichen und energie-
reichen Rohproteine und Stickstofffreien-Extraktstoffe (Kohlenhydrate) sind im
Landhabitat signifikant höher. Dies untermauert die Ergebnisse zur Energie der
Rehnahrung. Der höhere Energiegehalt der Rehäsung im Herbst und Winter im
Agrarhabitat erklärt sich überwiegend aus dem höheren Anteil an Stickstoff-
freien-Extraktstoffen. Am Vegetationshöhepunkt im Monat Mai erreicht die Nah-
rung im Wald und im Land einen Proteinhöhepunkt.
Der Rohfaseranteil liegt im Durchschnitt bei 25 % bis 29 % TS und erreicht im
Winter im Landhabitat Werte um die 30 % und weit über 30 % im Waldhabitat. Er
sinkt jedoch nicht unter 21 % TS. Bei so hohen Faseranteilen sollte man beim
Reh besser von „Selektierer“ anstatt von „Konzentrat-Selektierer“ sprechen.
Die Aufspaltung der Rohfasern nach VanSoest zeigt, das sowohl Lignin als auch
Zellulose signifikant mehr in der Waldäsung vorkommen.
dOS [% of DOS g/kg DS]70,0060,0050,0040,0030,00
XF [%
]
45,00
40,00
35,00
30,00
25,00
20,00
15,00
y=67,24-0,8*x
Habitat: WaldR2 Linear = 0,761
Seite 1
dOS [% of DOS g/kg TS]70.0060.0050.0040.0030.00
XF [%
]
50.00
40.00
30.00
20.00
10.00
y=61.51-0.68*x
Habitat: LandR2 Linear = 0.521
Seite 1
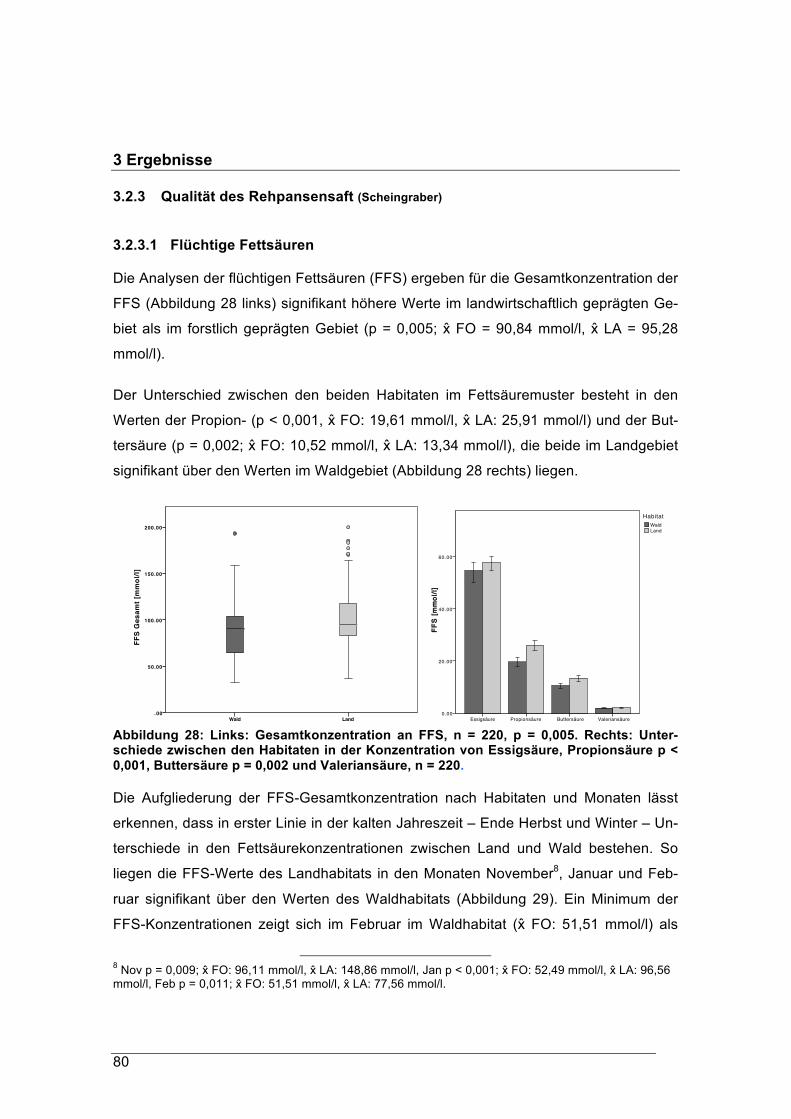
3 Ergebnisse
80
3.2.3 Qualität des Rehpansensaft (Scheingraber)
3.2.3.1 Flüchtige Fettsäuren
Die Analysen der flüchtigen Fettsäuren (FFS) ergeben für die Gesamtkonzentration der
FFS (Abbildung 28 links) signifikant höhere Werte im landwirtschaftlich geprägten Ge-
biet als im forstlich geprägten Gebiet (p = 0,005; x̂ FO = 90,84 mmol/l, x̂ LA = 95,28
mmol/l).
Der Unterschied zwischen den beiden Habitaten im Fettsäuremuster besteht in den
Werten der Propion- (p < 0,001, x̂ FO: 19,61 mmol/l, x̂ LA: 25,91 mmol/l) und der But-
tersäure (p = 0,002; x̂ FO: 10,52 mmol/l, x̂ LA: 13,34 mmol/l), die beide im Landgebiet
signifikant über den Werten im Waldgebiet (Abbildung 28 rechts) liegen.
Abbildung 28: Links: Gesamtkonzentration an FFS, n = 220, p = 0,005. Rechts: Unter-schiede zwischen den Habitaten in der Konzentration von Essigsäure, Propionsäure p < 0,001, Buttersäure p = 0,002 und Valeriansäure, n = 220.
Die Aufgliederung der FFS-Gesamtkonzentration nach Habitaten und Monaten lässt
erkennen, dass in erster Linie in der kalten Jahreszeit – Ende Herbst und Winter – Un-
terschiede in den Fettsäurekonzentrationen zwischen Land und Wald bestehen. So
liegen die FFS-Werte des Landhabitats in den Monaten November8, Januar und Feb-
ruar signifikant über den Werten des Waldhabitats (Abbildung 29). Ein Minimum der
FFS-Konzentrationen zeigt sich im Februar im Waldhabitat (x̂ FO: 51,51 mmol/l) als
8 Nov p = 0,009; x̂ FO: 96,11 mmol/l, x̂ LA: 148,86 mmol/l, Jan p < 0,001; x̂ FO: 52,49 mmol/l, x̂ LA: 96,56 mmol/l, Feb p = 0,011; x̂ FO: 51,51 mmol/l, x̂ LA: 77,56 mmol/l.
LandWald
FFS
Ges
amt [
mm
ol/l]
200.00
150.00
100.00
50.00
.00
Page 1
Valeriansäure ButtersäurePropionsäureEssigsäure
FFS
[m
mol
/l]
60.00
40.00
20.00
0.00
LandWald
Habitat
Page 1
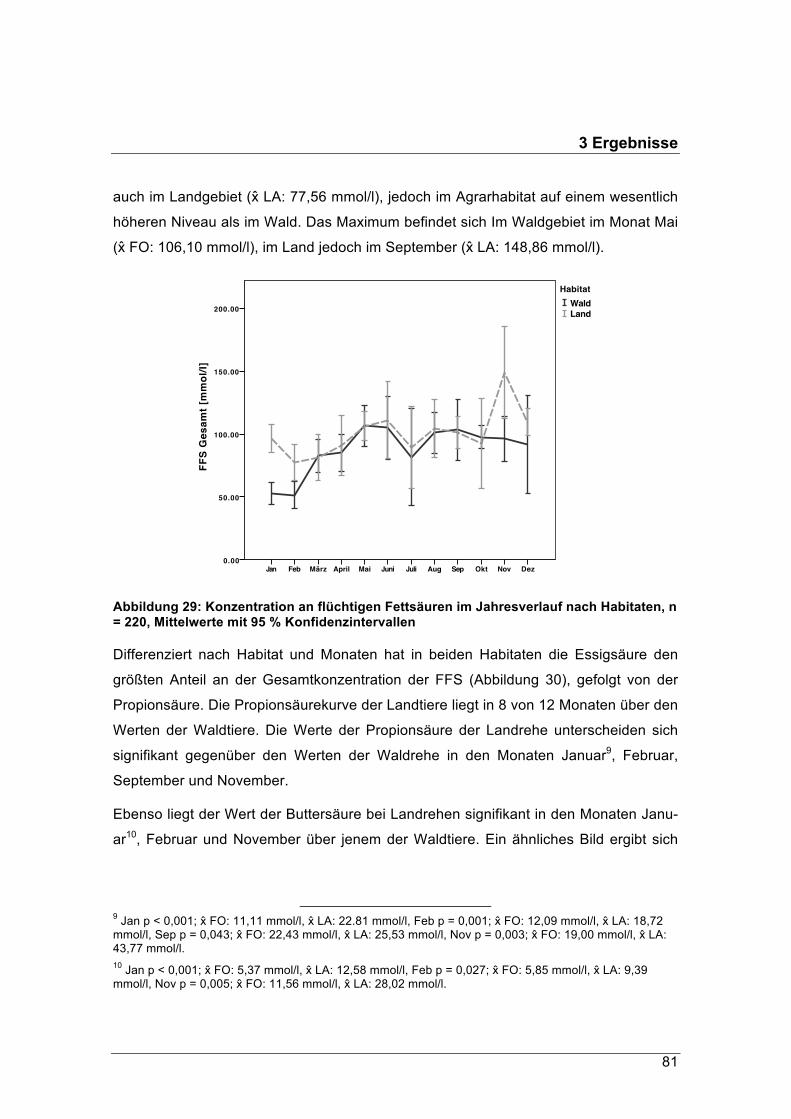
3 Ergebnisse
81
auch im Landgebiet (x̂ LA: 77,56 mmol/l), jedoch im Agrarhabitat auf einem wesentlich
höheren Niveau als im Wald. Das Maximum befindet sich Im Waldgebiet im Monat Mai
(x̂ FO: 106,10 mmol/l), im Land jedoch im September (x̂ LA: 148,86 mmol/l).
Abbildung 29: Konzentration an flüchtigen Fettsäuren im Jahresverlauf nach Habitaten, n = 220, Mittelwerte mit 95 % Konfidenzintervallen
Differenziert nach Habitat und Monaten hat in beiden Habitaten die Essigsäure den
größten Anteil an der Gesamtkonzentration der FFS (Abbildung 30), gefolgt von der
Propionsäure. Die Propionsäurekurve der Landtiere liegt in 8 von 12 Monaten über den
Werten der Waldtiere. Die Werte der Propionsäure der Landrehe unterscheiden sich
signifikant gegenüber den Werten der Waldrehe in den Monaten Januar9, Februar,
September und November.
Ebenso liegt der Wert der Buttersäure bei Landrehen signifikant in den Monaten Janu-
ar10, Februar und November über jenem der Waldtiere. Ein ähnliches Bild ergibt sich
9 Jan p < 0,001; x̂ FO: 11,11 mmol/l, x̂ LA: 22.81 mmol/l, Feb p = 0,001; x̂ FO: 12,09 mmol/l, x̂ LA: 18,72 mmol/l, Sep p = 0,043; x̂ FO: 22,43 mmol/l, x̂ LA: 25,53 mmol/l, Nov p = 0,003; x̂ FO: 19,00 mmol/l, x̂ LA: 43,77 mmol/l. 10 Jan p < 0,001; x̂ FO: 5,37 mmol/l, x̂ LA: 12,58 mmol/l, Feb p = 0,027; x̂ FO: 5,85 mmol/l, x̂ LA: 9,39 mmol/l, Nov p = 0,005; x̂ FO: 11,56 mmol/l, x̂ LA: 28,02 mmol/l.
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
FFS
Ges
amt [
mm
ol/l]
200.00
150.00
100.00
50.00
0.00
LandWald
Habitat
Page 1
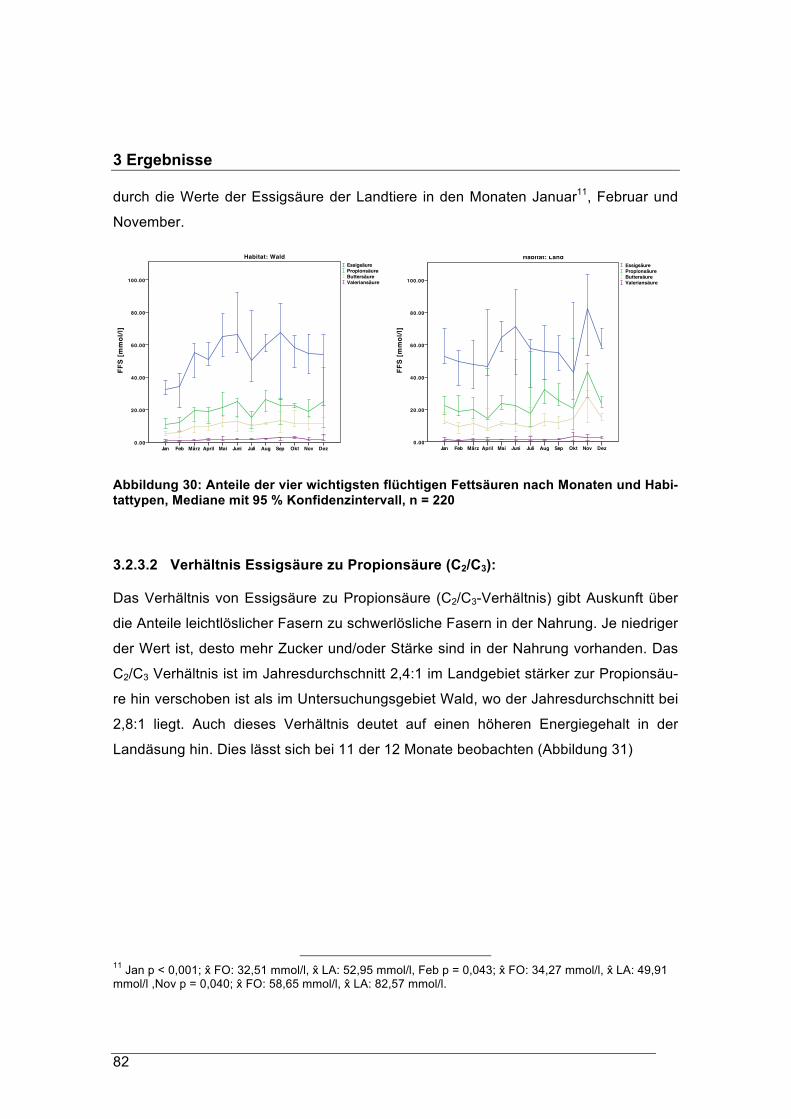
3 Ergebnisse
82
durch die Werte der Essigsäure der Landtiere in den Monaten Januar11, Februar und
November.
Abbildung 30: Anteile der vier wichtigsten flüchtigen Fettsäuren nach Monaten und Habi-tattypen, Mediane mit 95 % Konfidenzintervall, n = 220
3.2.3.2 Verhältnis Essigsäure zu Propionsäure (C2/C3):
Das Verhältnis von Essigsäure zu Propionsäure (C2/C3-Verhältnis) gibt Auskunft über
die Anteile leichtlöslicher Fasern zu schwerlösliche Fasern in der Nahrung. Je niedriger
der Wert ist, desto mehr Zucker und/oder Stärke sind in der Nahrung vorhanden. Das
C2/C3 Verhältnis ist im Jahresdurchschnitt 2,4:1 im Landgebiet stärker zur Propionsäu-
re hin verschoben ist als im Untersuchungsgebiet Wald, wo der Jahresdurchschnitt bei
2,8:1 liegt. Auch dieses Verhältnis deutet auf einen höheren Energiegehalt in der
Landäsung hin. Dies lässt sich bei 11 der 12 Monate beobachten (Abbildung 31)
11 Jan p < 0,001; x̂ FO: 32,51 mmol/l, x̂ LA: 52,95 mmol/l, Feb p = 0,043; x̂ FO: 34,27 mmol/l, x̂ LA: 49,91 mmol/l ,Nov p = 0,040; x̂ FO: 58,65 mmol/l, x̂ LA: 82,57 mmol/l.
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
FFS
[mm
ol/l]
100.00
80.00
60.00
40.00
20.00
0.00
ValeriansäureButtersäurePropionsäureEssigsäure
Habitat: Wald
Seite 1
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
FFS
[mm
ol/l]
100.00
80.00
60.00
40.00
20.00
0.00
ValeriansäureButtersäurePropionsäureEssigsäure
Habitat: Land
Seite 1
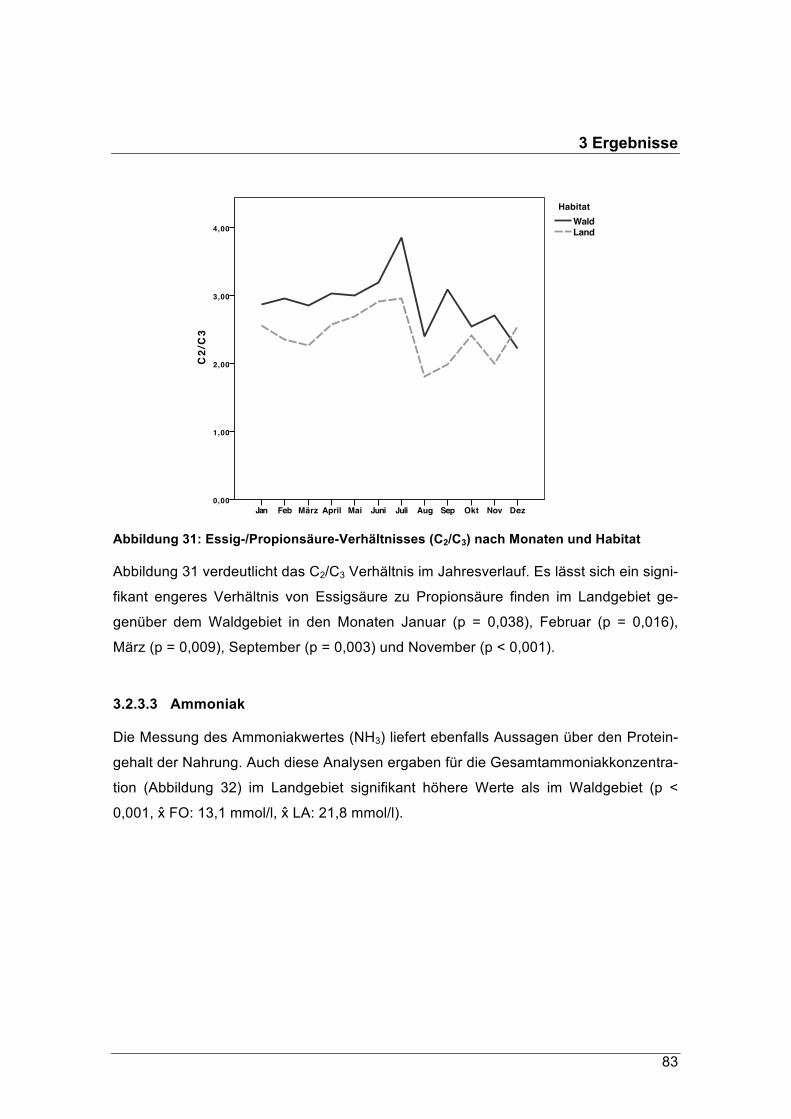
3 Ergebnisse
83
Abbildung 31: Essig-/Propionsäure-Verhältnisses (C2/C3) nach Monaten und Habitat
Abbildung 31 verdeutlicht das C2/C3 Verhältnis im Jahresverlauf. Es lässt sich ein signi-
fikant engeres Verhältnis von Essigsäure zu Propionsäure finden im Landgebiet ge-
genüber dem Waldgebiet in den Monaten Januar (p = 0,038), Februar (p = 0,016),
März (p = 0,009), September (p = 0,003) und November (p < 0,001).
3.2.3.3 Ammoniak
Die Messung des Ammoniakwertes (NH3) liefert ebenfalls Aussagen über den Protein-
gehalt der Nahrung. Auch diese Analysen ergaben für die Gesamtammoniakkonzentra-
tion (Abbildung 32) im Landgebiet signifikant höhere Werte als im Waldgebiet (p <
0,001, x̂ FO: 13,1 mmol/l, x̂ LA: 21,8 mmol/l).
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
C2/C3
4 ,00
3,00
2,00
1,00
0,00
LandWald
Habitat
Page 1

3 Ergebnisse
84
Abbildung 32: NH3-Gesamtkonzentration in Wald und Land, n = 220, p < 0,001.
Bei der Auswertung nach Monaten wird deutlich, dass der Ammoniakgehalt im Land-
gebiet in folgenden Monaten signifikant höher ist als im Waldgebiet: Februar12, März,
Mai, September und November.
Aus Abbildung 33 geht hervor, dass die beiden Maxima im Landhabitat in den Monaten
Mai und September zu finden sind. Die beiden Maxima im Waldgebiet liegen hingegen
im April (x̂ FO: 27,06 mmol/l) und Mai. Der Tiefpunkt der Ammoniakkonzentration findet
sich in beiden Gebieten im Februar, jedoch liegt dieser niedrigste Wert im Landhabitat
immer noch über dem mittleren Ammoniakgehalt im Wald. Mit Ausnahme des Aprils
weist der Pansensaft von Rehen im Agrarhabitat immer einen höheren Ammoniakgeh-
alt (NH3) auf.
12 Feb p < 0,006; x̂ FO: 5,61 mmol/l, x̂ LA: 12,78 mmol/l, März p < 0,005; x̂ FO: 9,62mmol/l, x̂ LA: 23,94 mmol/l, Mai p < 0,031; x̂ FO: 23,50 mmol/l, x̂ LA: 29,82 mmol/l, Sep p < 0,023; x̂ FO: 28,13 mmol/l, x̂ LA: 27,17 mmol/l, Nov p < 0,015; x̂ FO: 11,01 mmol/l, x̂ LA: 18,00 mmol/l.
LandWald
Am
mon
iak
[mm
ol/l]
60,00
40,00
20,00
,00
Page 1
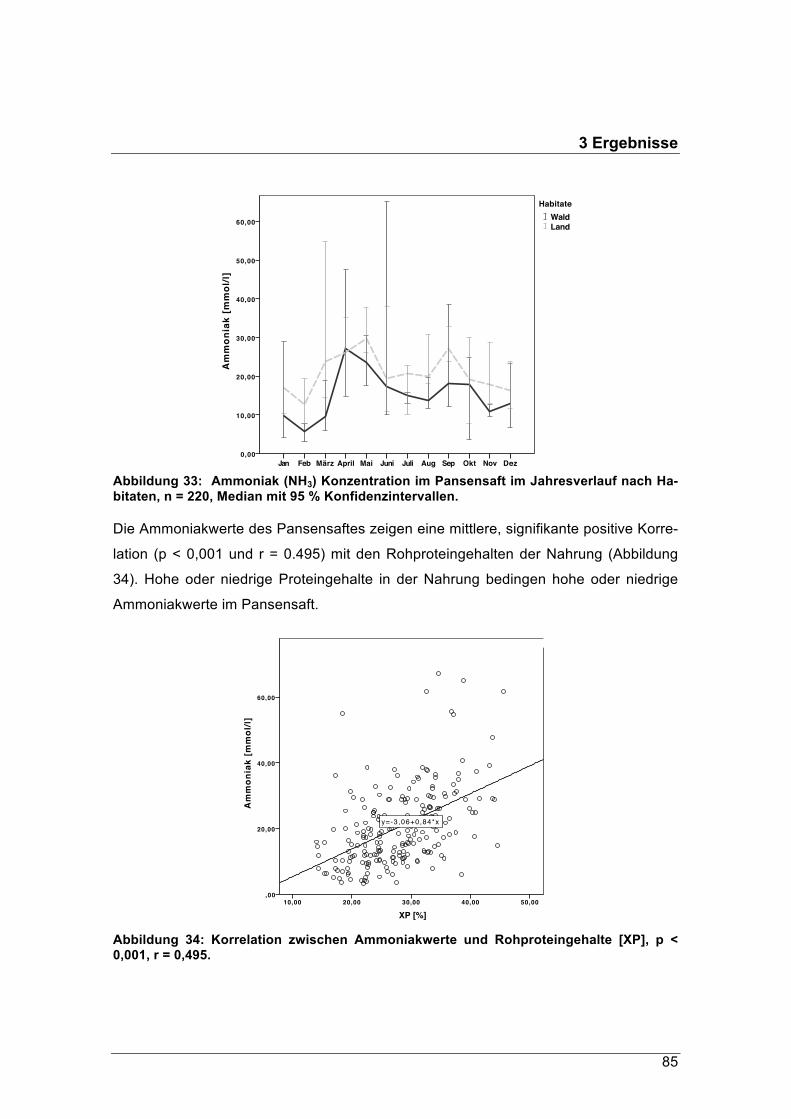
3 Ergebnisse
85
Abbildung 33: Ammoniak (NH3) Konzentration im Pansensaft im Jahresverlauf nach Ha-bitaten, n = 220, Median mit 95 % Konfidenzintervallen.
Die Ammoniakwerte des Pansensaftes zeigen eine mittlere, signifikante positive Korre-
lation (p < 0,001 und r = 0.495) mit den Rohproteingehalten der Nahrung (Abbildung
34). Hohe oder niedrige Proteingehalte in der Nahrung bedingen hohe oder niedrige
Ammoniakwerte im Pansensaft.
Abbildung 34: Korrelation zwischen Ammoniakwerte und Rohproteingehalte [XP], p < 0,001, r = 0,495.
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Am
mon
iak
[mm
ol/l]
60,00
50,00
40,00
30,00
20,00
10,00
0,00
LandWald
Habitate
Page 1
XP [%]50,0040,0030,0020,0010,00
Am
mon
iak
[mm
ol/l]
60,00
40,00
20,00
,00
y=-3,06+0,84*x
R2 Linear = 0,238
Seite 1
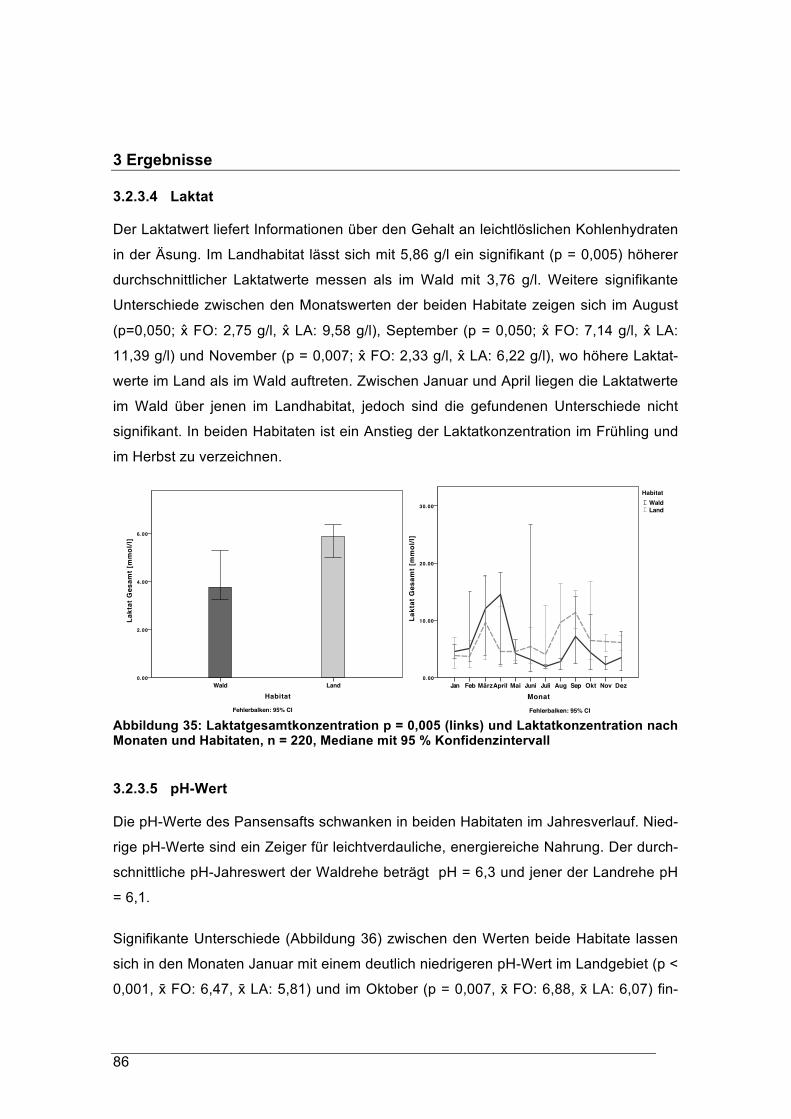
3 Ergebnisse
86
3.2.3.4 Laktat
Der Laktatwert liefert Informationen über den Gehalt an leichtlöslichen Kohlenhydraten
in der Äsung. Im Landhabitat lässt sich mit 5,86 g/l ein signifikant (p = 0,005) höherer
durchschnittlicher Laktatwerte messen als im Wald mit 3,76 g/l. Weitere signifikante
Unterschiede zwischen den Monatswerten der beiden Habitate zeigen sich im August
(p=0,050; x̂ FO: 2,75 g/l, x̂ LA: 9,58 g/l), September (p = 0,050; x̂ FO: 7,14 g/l, x̂ LA:
11,39 g/l) und November (p = 0,007; x̂ FO: 2,33 g/l, x̂ LA: 6,22 g/l), wo höhere Laktat-
werte im Land als im Wald auftreten. Zwischen Januar und April liegen die Laktatwerte
im Wald über jenen im Landhabitat, jedoch sind die gefundenen Unterschiede nicht
signifikant. In beiden Habitaten ist ein Anstieg der Laktatkonzentration im Frühling und
im Herbst zu verzeichnen.
Abbildung 35: Laktatgesamtkonzentration p = 0,005 (links) und Laktatkonzentration nach Monaten und Habitaten, n = 220, Mediane mit 95 % Konfidenzintervall
3.2.3.5 pH-Wert
Die pH-Werte des Pansensafts schwanken in beiden Habitaten im Jahresverlauf. Nied-
rige pH-Werte sind ein Zeiger für leichtverdauliche, energiereiche Nahrung. Der durch-
schnittliche pH-Jahreswert der Waldrehe beträgt pH = 6,3 und jener der Landrehe pH
= 6,1.
Signifikante Unterschiede (Abbildung 36) zwischen den Werten beide Habitate lassen
sich in den Monaten Januar mit einem deutlich niedrigeren pH-Wert im Landgebiet (p <
0,001, x̄ FO: 6,47, x̄ LA: 5,81) und im Oktober (p = 0,007, x̄ FO: 6,88, x̄ LA: 6,07) fin-
HabitatLandWald
Lak
tat
Ges
amt
[mm
ol/l]
6 .00
4.00
2.00
0.00
Fehlerbalken: 95% CI
Page 1
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Lak
tat
Ges
amt
[mm
ol/l]
30.00
20.00
10.00
0.00
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
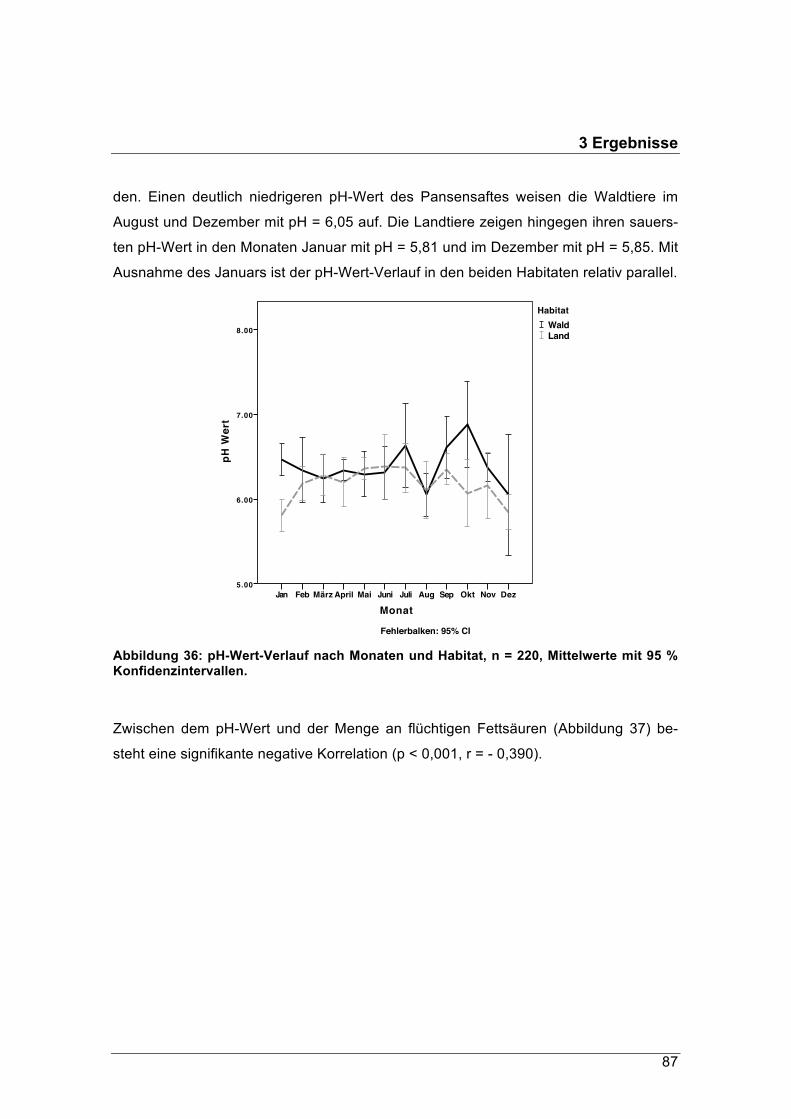
3 Ergebnisse
87
den. Einen deutlich niedrigeren pH-Wert des Pansensaftes weisen die Waldtiere im
August und Dezember mit pH = 6,05 auf. Die Landtiere zeigen hingegen ihren sauers-
ten pH-Wert in den Monaten Januar mit pH = 5,81 und im Dezember mit pH = 5,85. Mit
Ausnahme des Januars ist der pH-Wert-Verlauf in den beiden Habitaten relativ parallel.
Abbildung 36: pH-Wert-Verlauf nach Monaten und Habitat, n = 220, Mittelwerte mit 95 % Konfidenzintervallen.
Zwischen dem pH-Wert und der Menge an flüchtigen Fettsäuren (Abbildung 37) be-
steht eine signifikante negative Korrelation (p < 0,001, r = - 0,390).
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
pH W
ert
8 .00
7.00
6.00
5.00
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
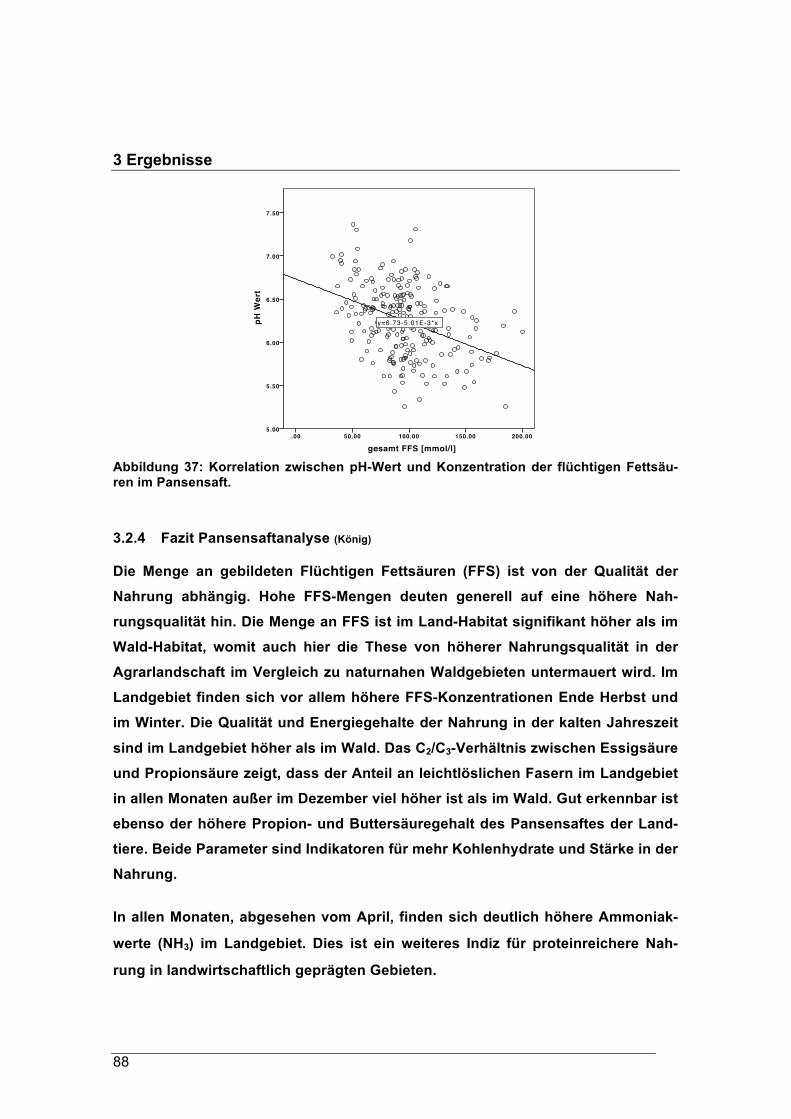
3 Ergebnisse
88
Abbildung 37: Korrelation zwischen pH-Wert und Konzentration der flüchtigen Fettsäu-ren im Pansensaft.
3.2.4 Fazit Pansensaftanalyse (König)
Die Menge an gebildeten Flüchtigen Fettsäuren (FFS) ist von der Qualität der
Nahrung abhängig. Hohe FFS-Mengen deuten generell auf eine höhere Nah-
rungsqualität hin. Die Menge an FFS ist im Land-Habitat signifikant höher als im
Wald-Habitat, womit auch hier die These von höherer Nahrungsqualität in der
Agrarlandschaft im Vergleich zu naturnahen Waldgebieten untermauert wird. Im
Landgebiet finden sich vor allem höhere FFS-Konzentrationen Ende Herbst und
im Winter. Die Qualität und Energiegehalte der Nahrung in der kalten Jahreszeit
sind im Landgebiet höher als im Wald. Das C2/C3-Verhältnis zwischen Essigsäure
und Propionsäure zeigt, dass der Anteil an leichtlöslichen Fasern im Landgebiet
in allen Monaten außer im Dezember viel höher ist als im Wald. Gut erkennbar ist
ebenso der höhere Propion- und Buttersäuregehalt des Pansensaftes der Land-
tiere. Beide Parameter sind Indikatoren für mehr Kohlenhydrate und Stärke in der
Nahrung.
In allen Monaten, abgesehen vom April, finden sich deutlich höhere Ammoniak-
werte (NH3) im Landgebiet. Dies ist ein weiteres Indiz für proteinreichere Nah-
rung in landwirtschaftlich geprägten Gebieten.
gesamt FFS [mmol/l]200.00150.00100.0050.00.00
pH W
ert
7 .50
7.00
6.50
6.00
5.50
5.00
y=6.73-5.01E-3*x
R2 Linear = 0.151
Seite 1

3 Ergebnisse
89
Im Landhabitat lässt sich deutlich mehr Laktat finden, was ebenfalls auf einen
höheren Gehalt an leichtlöslichen Kohlenhydraten hinweist.
Der pH-Wert zeigt deutlich saure Werte im Landgebiet in den Wintermonaten De-
zember, Januar und im Sommer im August. Dies deckt sich mit den Ergebnissen
der ME-Berechnung und zeigt, dass sich in diesen Monaten viele FFS im Pansen
befunden haben, was darauf hinweist, dass sich hier ausreichend KH/Stärke und
Zellulose in der Nahrung befand. Diese Peaks im August und Dezember lassen
sich auch im Waldgebiet finden. Dies könnte aber hauptsächlich an den hohen
Mengen gebildeter Essigsäure liegen. Diese Säure ist die am häufigsten und
mengenmäßig am stärksten gebildete Säure im Pansen und wird hauptsächlich
aus Zellulose gebildet.
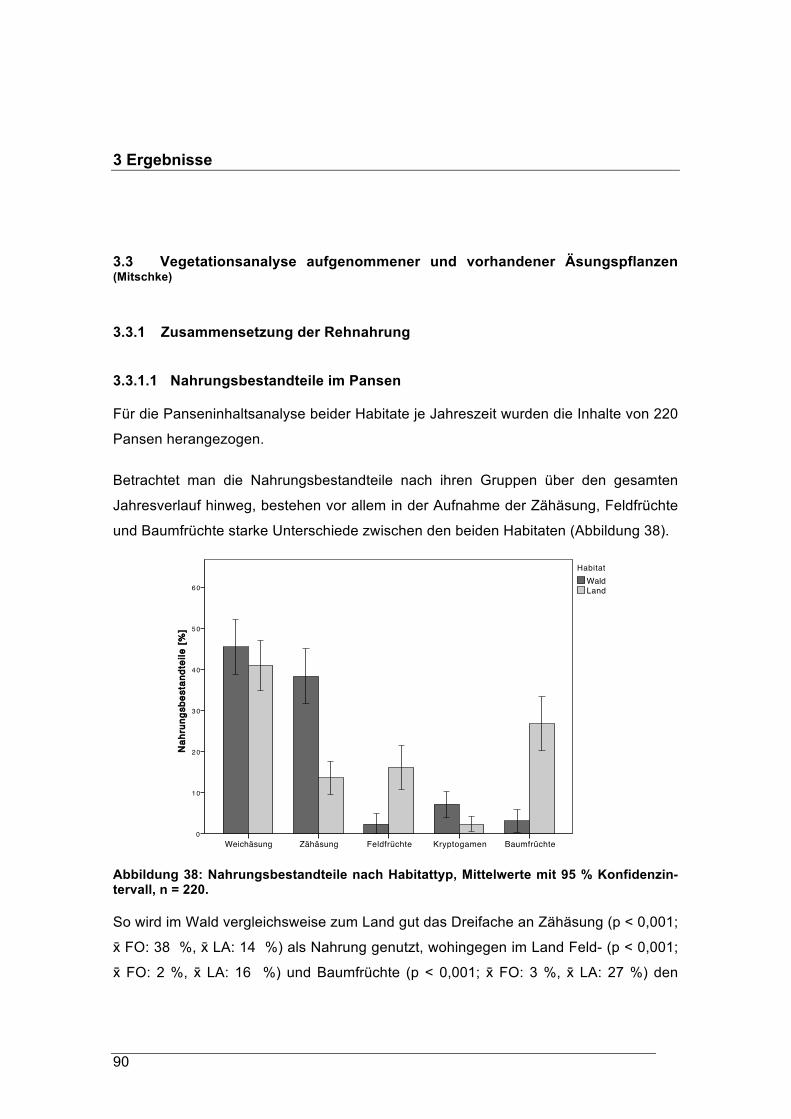
3 Ergebnisse
90
3.3 Vegetationsanalyse aufgenommener und vorhandener Äsungspflanzen (Mitschke)
3.3.1 Zusammensetzung der Rehnahrung
3.3.1.1 Nahrungsbestandteile im Pansen
Für die Panseninhaltsanalyse beider Habitate je Jahreszeit wurden die Inhalte von 220
Pansen herangezogen.
Betrachtet man die Nahrungsbestandteile nach ihren Gruppen über den gesamten
Jahresverlauf hinweg, bestehen vor allem in der Aufnahme der Zähäsung, Feldfrüchte
und Baumfrüchte starke Unterschiede zwischen den beiden Habitaten (Abbildung 38).
Abbildung 38: Nahrungsbestandteile nach Habitattyp, Mittelwerte mit 95 % Konfidenzin-tervall, n = 220.
So wird im Wald vergleichsweise zum Land gut das Dreifache an Zähäsung (p < 0,001;
x̄ FO: 38 %, x̄ LA: 14 %) als Nahrung genutzt, wohingegen im Land Feld- (p < 0,001;
x̄ FO: 2 %, x̄ LA: 16 %) und Baumfrüchte (p < 0,001; x̄ FO: 3 %, x̄ LA: 27 %) den
BaumfrüchteKryptogamenFeldfrüchteZähäsungWeichäsung
Nah
rung
sbes
tand
teile
[%]
6 0
5 0
4 0
3 0
2 0
1 0
0
LandWald
Habitat
Page 1
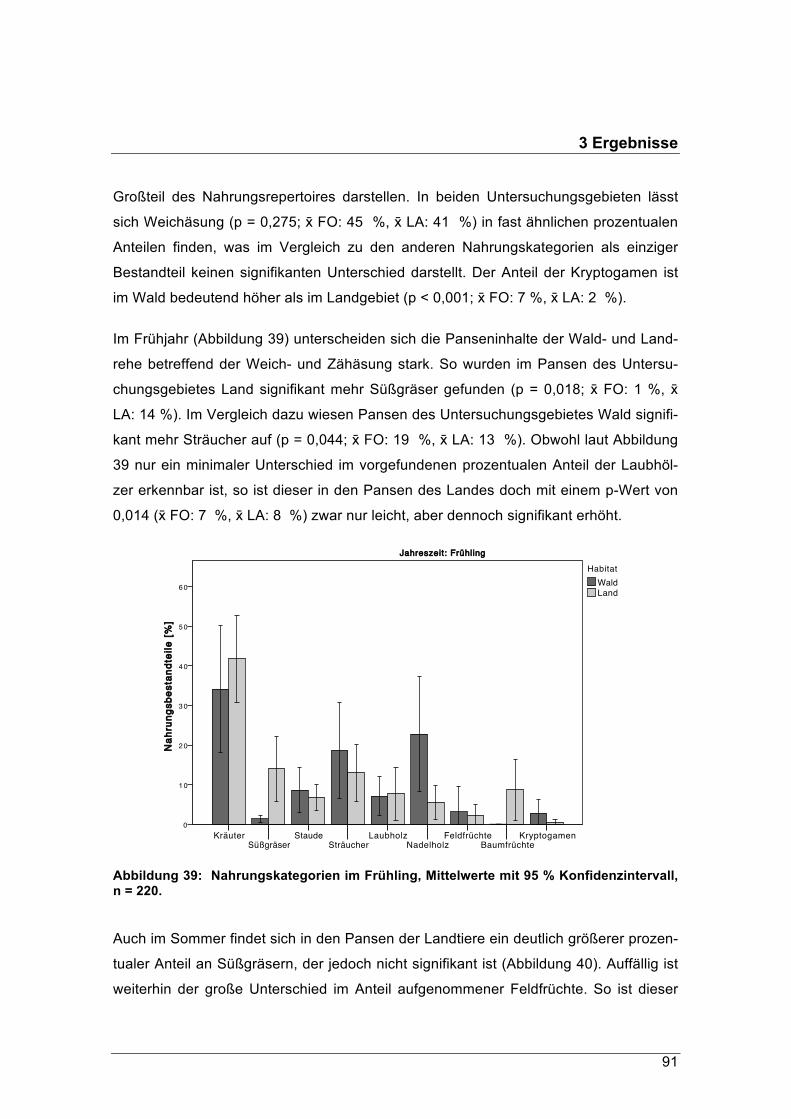
3 Ergebnisse
91
Großteil des Nahrungsrepertoires darstellen. In beiden Untersuchungsgebieten lässt
sich Weichäsung (p = 0,275; x̄ FO: 45 %, x̄ LA: 41 %) in fast ähnlichen prozentualen
Anteilen finden, was im Vergleich zu den anderen Nahrungskategorien als einziger
Bestandteil keinen signifikanten Unterschied darstellt. Der Anteil der Kryptogamen ist
im Wald bedeutend höher als im Landgebiet (p < 0,001; x̄ FO: 7 %, x̄ LA: 2 %).
Im Frühjahr (Abbildung 39) unterscheiden sich die Panseninhalte der Wald- und Land-
rehe betreffend der Weich- und Zähäsung stark. So wurden im Pansen des Untersu-
chungsgebietes Land signifikant mehr Süßgräser gefunden (p = 0,018; x̄ FO: 1 %, x̄
LA: 14 %). Im Vergleich dazu wiesen Pansen des Untersuchungsgebietes Wald signifi-
kant mehr Sträucher auf (p = 0,044; x̄ FO: 19 %, x̄ LA: 13 %). Obwohl laut Abbildung
39 nur ein minimaler Unterschied im vorgefundenen prozentualen Anteil der Laubhöl-
zer erkennbar ist, so ist dieser in den Pansen des Landes doch mit einem p-Wert von
0,014 (x̄ FO: 7 %, x̄ LA: 8 %) zwar nur leicht, aber dennoch signifikant erhöht.
Abbildung 39: Nahrungskategorien im Frühling, Mittelwerte mit 95 % Konfidenzintervall, n = 220.
Auch im Sommer findet sich in den Pansen der Landtiere ein deutlich größerer prozen-
tualer Anteil an Süßgräsern, der jedoch nicht signifikant ist (Abbildung 40). Auffällig ist
weiterhin der große Unterschied im Anteil aufgenommener Feldfrüchte. So ist dieser
KryptogamenBaumfrüchte
FeldfrüchteNadelholz
LaubholzSträucher
StaudeSüßgräser
Kräuter
Nah
rung
sbes
tand
teile
[%]
6 0
5 0
4 0
3 0
2 0
1 0
0
LandWald
Habitat
Jahreszeit: Frühling
Page 1
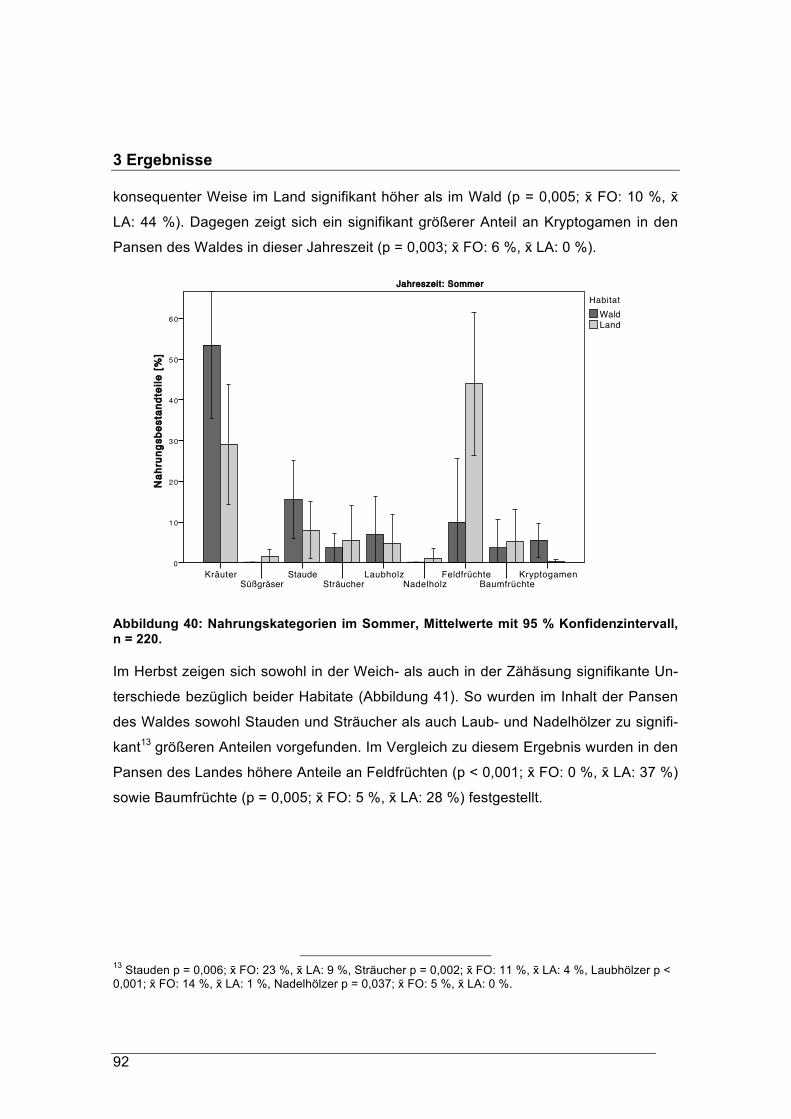
3 Ergebnisse
92
konsequenter Weise im Land signifikant höher als im Wald (p = 0,005; x̄ FO: 10 %, x̄
LA: 44 %). Dagegen zeigt sich ein signifikant größerer Anteil an Kryptogamen in den
Pansen des Waldes in dieser Jahreszeit (p = 0,003; x̄ FO: 6 %, x̄ LA: 0 %).
Abbildung 40: Nahrungskategorien im Sommer, Mittelwerte mit 95 % Konfidenzintervall, n = 220.
Im Herbst zeigen sich sowohl in der Weich- als auch in der Zähäsung signifikante Un-
terschiede bezüglich beider Habitate (Abbildung 41). So wurden im Inhalt der Pansen
des Waldes sowohl Stauden und Sträucher als auch Laub- und Nadelhölzer zu signifi-
kant13 größeren Anteilen vorgefunden. Im Vergleich zu diesem Ergebnis wurden in den
Pansen des Landes höhere Anteile an Feldfrüchten (p < 0,001; x̄ FO: 0 %, x̄ LA: 37 %)
sowie Baumfrüchte (p = 0,005; x̄ FO: 5 %, x̄ LA: 28 %) festgestellt.
13 Stauden p = 0,006; x̄ FO: 23 %, x̄ LA: 9 %, Sträucher p = 0,002; x̄ FO: 11 %, x̄ LA: 4 %, Laubhölzer p < 0,001; x̄ FO: 14 %, x̄ LA: 1 %, Nadelhölzer p = 0,037; x̄ FO: 5 %, x̄ LA: 0 %.
KryptogamenBaumfrüchte
FeldfrüchteNadelholz
LaubholzSträucher
StaudeSüßgräser
Kräuter
Nah
rung
sbes
tand
teile
[%]
6 0
5 0
4 0
3 0
2 0
1 0
0
LandWald
Habitat
Jahreszeit: Sommer
Page 1
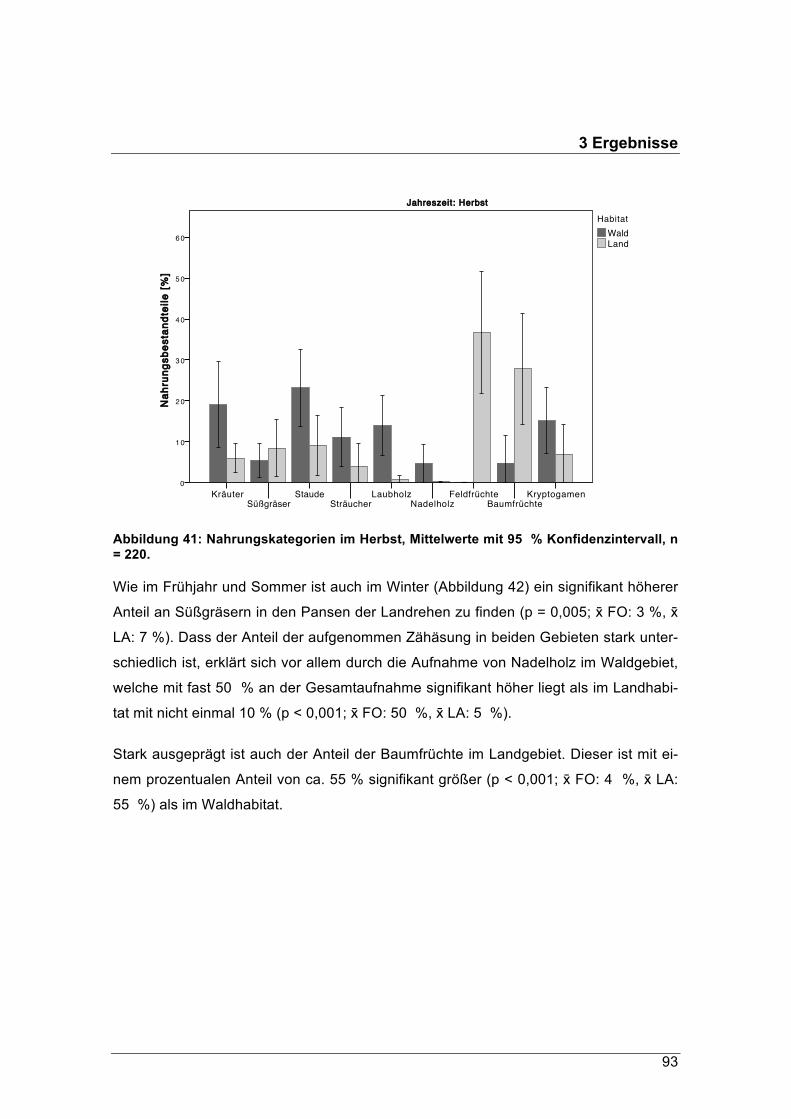
3 Ergebnisse
93
Abbildung 41: Nahrungskategorien im Herbst, Mittelwerte mit 95 % Konfidenzintervall, n = 220.
Wie im Frühjahr und Sommer ist auch im Winter (Abbildung 42) ein signifikant höherer
Anteil an Süßgräsern in den Pansen der Landrehen zu finden (p = 0,005; x̄ FO: 3 %, x̄
LA: 7 %). Dass der Anteil der aufgenommen Zähäsung in beiden Gebieten stark unter-
schiedlich ist, erklärt sich vor allem durch die Aufnahme von Nadelholz im Waldgebiet,
welche mit fast 50 % an der Gesamtaufnahme signifikant höher liegt als im Landhabi-
tat mit nicht einmal 10 % (p < 0,001; x̄ FO: 50 %, x̄ LA: 5 %).
Stark ausgeprägt ist auch der Anteil der Baumfrüchte im Landgebiet. Dieser ist mit ei-
nem prozentualen Anteil von ca. 55 % signifikant größer (p < 0,001; x̄ FO: 4 %, x̄ LA:
55 %) als im Waldhabitat.
KryptogamenBaumfrüchte
FeldfrüchteNadelholz
LaubholzSträucher
StaudeSüßgräser
Kräuter
Nah
rung
sbes
tand
teile
[%]
6 0
5 0
4 0
3 0
2 0
1 0
0
LandWald
Habitat
Jahreszeit: Herbst
Page 1
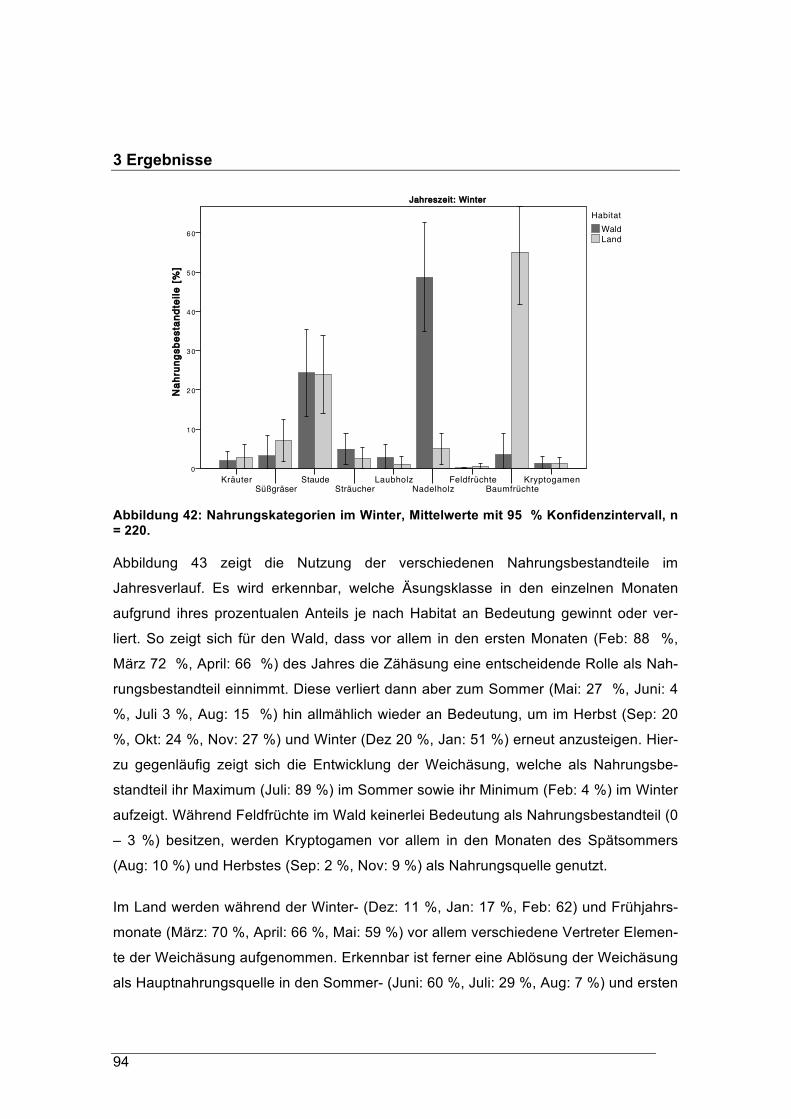
3 Ergebnisse
94
Abbildung 42: Nahrungskategorien im Winter, Mittelwerte mit 95 % Konfidenzintervall, n = 220.
Abbildung 43 zeigt die Nutzung der verschiedenen Nahrungsbestandteile im
Jahresverlauf. Es wird erkennbar, welche Äsungsklasse in den einzelnen Monaten
aufgrund ihres prozentualen Anteils je nach Habitat an Bedeutung gewinnt oder ver-
liert. So zeigt sich für den Wald, dass vor allem in den ersten Monaten (Feb: 88 %,
März 72 %, April: 66 %) des Jahres die Zähäsung eine entscheidende Rolle als Nah-
rungsbestandteil einnimmt. Diese verliert dann aber zum Sommer (Mai: 27 %, Juni: 4
%, Juli 3 %, Aug: 15 %) hin allmählich wieder an Bedeutung, um im Herbst (Sep: 20
%, Okt: 24 %, Nov: 27 %) und Winter (Dez 20 %, Jan: 51 %) erneut anzusteigen. Hier-
zu gegenläufig zeigt sich die Entwicklung der Weichäsung, welche als Nahrungsbe-
standteil ihr Maximum (Juli: 89 %) im Sommer sowie ihr Minimum (Feb: 4 %) im Winter
aufzeigt. Während Feldfrüchte im Wald keinerlei Bedeutung als Nahrungsbestandteil (0
– 3 %) besitzen, werden Kryptogamen vor allem in den Monaten des Spätsommers
(Aug: 10 %) und Herbstes (Sep: 2 %, Nov: 9 %) als Nahrungsquelle genutzt.
Im Land werden während der Winter- (Dez: 11 %, Jan: 17 %, Feb: 62) und Frühjahrs-
monate (März: 70 %, April: 66 %, Mai: 59 %) vor allem verschiedene Vertreter Elemen-
te der Weichäsung aufgenommen. Erkennbar ist ferner eine Ablösung der Weichäsung
als Hauptnahrungsquelle in den Sommer- (Juni: 60 %, Juli: 29 %, Aug: 7 %) und ersten
KryptogamenBaumfrüchte
FeldfrüchteNadelholz
LaubholzSträucher
StaudeSüßgräser
Kräuter
Nah
rung
sbes
tand
teile
[%]
6 0
5 0
4 0
3 0
2 0
1 0
0
LandWald
Habitat
Jahreszeit: Winter
Page 1
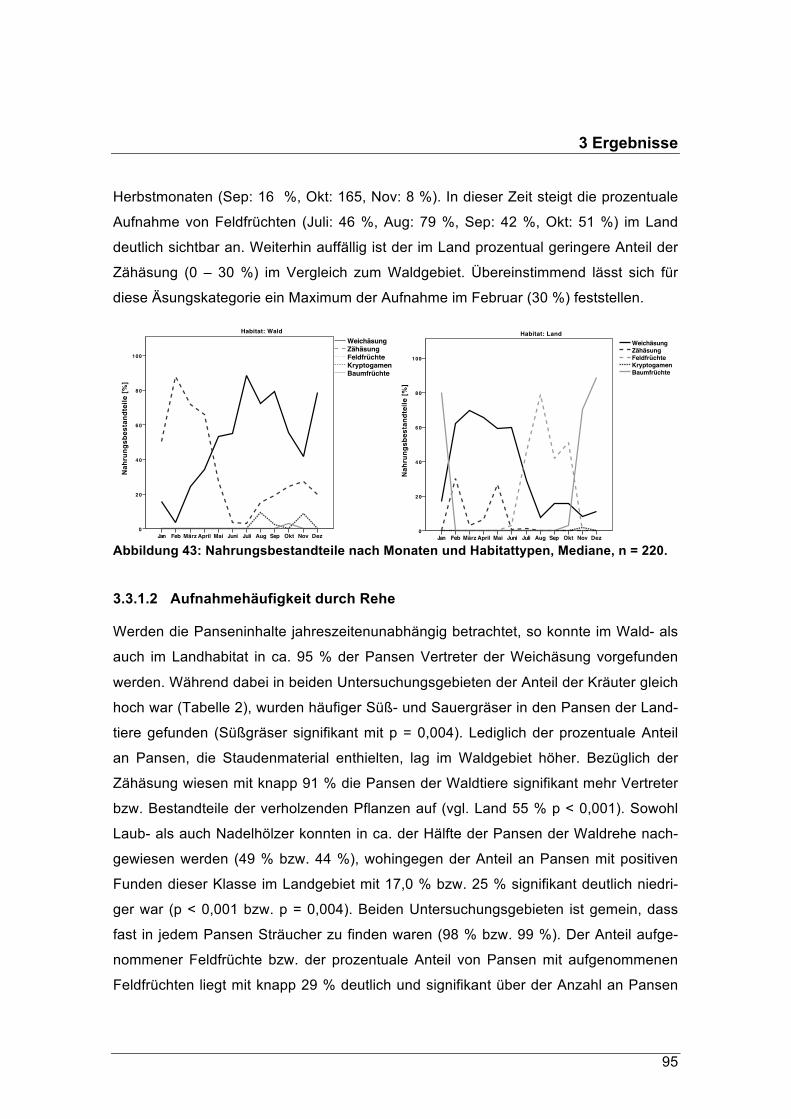
3 Ergebnisse
95
Herbstmonaten (Sep: 16 %, Okt: 165, Nov: 8 %). In dieser Zeit steigt die prozentuale
Aufnahme von Feldfrüchten (Juli: 46 %, Aug: 79 %, Sep: 42 %, Okt: 51 %) im Land
deutlich sichtbar an. Weiterhin auffällig ist der im Land prozentual geringere Anteil der
Zähäsung (0 – 30 %) im Vergleich zum Waldgebiet. Übereinstimmend lässt sich für
diese Äsungskategorie ein Maximum der Aufnahme im Februar (30 %) feststellen.
Abbildung 43: Nahrungsbestandteile nach Monaten und Habitattypen, Mediane, n = 220.
3.3.1.2 Aufnahmehäufigkeit durch Rehe
Werden die Panseninhalte jahreszeitenunabhängig betrachtet, so konnte im Wald- als
auch im Landhabitat in ca. 95 % der Pansen Vertreter der Weichäsung vorgefunden
werden. Während dabei in beiden Untersuchungsgebieten der Anteil der Kräuter gleich
hoch war (Tabelle 2), wurden häufiger Süß- und Sauergräser in den Pansen der Land-
tiere gefunden (Süßgräser signifikant mit p = 0,004). Lediglich der prozentuale Anteil
an Pansen, die Staudenmaterial enthielten, lag im Waldgebiet höher. Bezüglich der
Zähäsung wiesen mit knapp 91 % die Pansen der Waldtiere signifikant mehr Vertreter
bzw. Bestandteile der verholzenden Pflanzen auf (vgl. Land 55 % p < 0,001). Sowohl
Laub- als auch Nadelhölzer konnten in ca. der Hälfte der Pansen der Waldrehe nach-
gewiesen werden (49 % bzw. 44 %), wohingegen der Anteil an Pansen mit positiven
Funden dieser Klasse im Landgebiet mit 17,0 % bzw. 25 % signifikant deutlich niedri-
ger war (p < 0,001 bzw. p = 0,004). Beiden Untersuchungsgebieten ist gemein, dass
fast in jedem Pansen Sträucher zu finden waren (98 % bzw. 99 %). Der Anteil aufge-
nommener Feldfrüchte bzw. der prozentuale Anteil von Pansen mit aufgenommenen
Feldfrüchten liegt mit knapp 29 % deutlich und signifikant über der Anzahl an Pansen
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Nah
rung
sbes
tand
teile
[%]
100
8 0
6 0
4 0
2 0
0
BaumfrüchteKryptogamenFeldfrüchteZähäsungWeichäsung
Habitat: Wald
Page 1
DezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Nah
rung
sbes
tand
teile
[%]
100
8 0
6 0
4 0
2 0
0
BaumfrüchteKryptogamenFeldfrüchteZähäsungWeichäsung
Habitat: Land
Page 1
3 Ergebnisse
96
mit Feldfrüchten im Waldgebiet (ca. 5 %, p < 0,001). Gleiches zeigt sich für Baumfrüch-
te (vgl. Wald 12 % bzw. Land 37 %, p < 0,001). Kryptogamen wurden mit 41% signifi-
kant häufiger in Pansen der Waldtiere nachgewiesen (p < 0,001).
Tabelle 8: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen über alle Jahreszeiten, Mittelwerte, n = 220.
Kategorie Wald in % Land in %
Weichäsung 95,30 94,80
Kräuter 60,50 61,50
Süßgräser 33,70 54,50
Sauergräser 1,20 2,20
Stauden 69,80 57,50
Zähäsung 90,70 54,50
Laubhölzer 48,80 17,20
Nadelhölzer 44,20 25,40
Sträucher 97,70 99,30
Feldfrüchte 4,70 29,10 Kryptogamen 40,70 15,70
Farne 22,10 7,50
Moose 3,50 1,50
Pilze 22,10 8,20
Baumfrüchte 11,60 36,60 Sonstiges 5,80 0,00
Sowohl im Waldgebiet als auch im Landgebiet wurde in fast 100 % der Pansen des
Frühjahres Weichäsung gefunden (Tabelle 9). Dabei gestaltet sich der prozentuale
Anteil der Pansen in Bezug auf Kräuter und Stauden fast annähernd gleich. Lediglich
der Anteil der Pansen mit Süßgräsern war im Land im Vergleich zum Wald erhöht (62
%). Sowohl in der Zähäsung Gesamt, als auch in den Unterkategorien der Laub- und
Nadelhölzer konnten in mehreren Pansen der Waldtiere und der Landtiere Nachweise
erbracht werden. Dabei wurden in Pansen der Waldtiere nicht nur häufiger Nadelhölzer
gefunden, sondern ebenfalls auch signifikant häufiger Laubhölzer (p=0,045). Lediglich
der Anteil der Pansen mit aufgenommenen Sträuchern ist im Landgebiet höher (vgl.
Waldtiere 91% bzw. Landtiere 98 %). Während Feldfrüchte in gleichem Umfang im
Wald- wie im Landhabitat aufgenommen wurden, wurden mehr Pansen mit Kryptoga-
men im Waldgebiet festgestellt.
3 Ergebnisse
97
Tabelle 9: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen im Frühling, Mittelwerte, N=220.
In Anlehnung an das Frühjahr konnte auch im Sommer in 100 % der Pansen Vertreter
der Weichäsung aufgefunden werden. Wie in Tabelle 10 ersichtlich, wurden im Wald-
habitat vor allem krautige Vertreter und Stauden identifiziert. Im Landgebiet wurden in
den Pansen alle Untergruppen der Weichäsung festgestellt. So beinhalteten 90 % der
Pansen der Landtiere Kräuter, 47 % Stauden, 32 % Süß- und 11 % Sauergräser. Wie
bereits im Frühjahr so ist auch im Sommer der Anteil der Pansen mit aufgenommener
Zähäsung im Waldgebiet mit 77 % höher als im Landgebiet (hier 53 % der Pansen mit
Zähäsung). Während sich mit 46 % mehr Pansen mit Laubhölzern im Untersuchungs-
gebiet Wald finden lassen, ist der Anteil an Pansen mit aufgenommenen Nadelhölzern
mit 11 % im Untersuchungsgebiet Land deutlich höher. Der Anteil der Pansen mit auf-
genommenen Feldfrüchten steigt im Sommer im Landhabitat deutlich auf 74 % an,
wohingegen die Anzahl der Pansen im Waldgebiet bezüglich der Feldfrüchte nur eine
leichte Anhebung auf 15 % erfährt. Diese Differenz des prozentualen Anteils zwischen
Land- und Waldgebiet kann mit einem p-Wert von 0,003 als signifikant verschieden
angesehen werden. Mit ca. 54 % der Pansen der Waldtiere, die Kryptogamen enthiel-
ten, ist dieser Anteil im Sommer um das Doppelte größer als im Frühjahr. Ähnlich wie
Klasse Wald Land
Weichäsung 100,00 97,60
Kräuter 85,70 81,00
Süßgräser 38,10 61,90
Sauergräser 0,00 0,00
Stauden 52,40 47,60
Zähäsung 90,50 69,00
Laubhölzer 52,40 23,80
Nadelhölzer 47,60 31,00
Sträucher 90,50 97,60
Feldfrüchte 9,50 9,50 Kryptogamen 23,80 7,10
Farne 23,80 7,10
Moose 0,00 0,00
Pilze 0,00 0,00
Baumfrüchte 0,00 14,30 Sonstiges 0,00 0,00
3 Ergebnisse
98
im Frühjahr bleibt dieser Wert auch im Sommer für die Pansen des Landes annähernd
gleich niedrig. Hervorzuheben ist an dieser Stelle vor allem der Unterschied bezüglich
des Anteils an Pansen mit Pilzen. So ist dieser im Wald signifikant größer als auf dem
Land (p = 0,006). Generell lassen sich mehr Pansen mit Kryptogamen im Wald finden
(vgl. Waldtiere 54 %, Landtiere 5 %, p = 0,003). Baumfrüchte lassen sich in dieser Zeit
in Pansen beider Untersuchungsgebiete finden.
Tabelle 10: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen im Sommer, Mittelwerte.
Kategorie Wald Land
Weichäsung 100,00 100,00
Kräuter 100,00 89,50
Süßgräser 0,00 31,60
Sauergräser 0,00 10,50
Stauden 76,90 47,40
Zähäsung 76,90 52,60
Laubhölzer 46,20 26,30
Nadelhölzer 0,00 10,50
Sträucher 100,00 100,00
Feldfrüchte 15,40 73,70 Kryptogamen 53,80 5,30
Farne 30,80 5,30
Moose 7,70 0,00
Pilze 38,50 0,00
Baumfrüchte 15,40 10,50 Sonstiges 0,00 0,00
Im Herbst nimmt in beiden Gebieten der prozentuale Anteil an Pansen mit Weichäsung
ab (Tabelle 11). So sinkt in beiden Gebieten vor allem der Anteil der Pansen mit Kräu-
tern, wohingegen mehr Süßgräser festgestellt werden. Auch die Anzahl der Pansen mit
Stauden nimmt prozentual ab, wobei im Waldgebiet noch signifikant höhere Anteile an
Pansen mit dieser Form der Weichäsung auftreten (p = 0,044). Auch die Anzahl der
Pansen mit Zähäsung erweist sich im Wald- als signifikant höher als im Landgebiet (p
< 0,001). Vor allem die große Differenz zwischen den prozentualen Anteilen der Pan-
sen mit Laubholz (Wald 67 % bzw. Land 9 %) ist signifikant (p < 0,001). Im Gegensatz
zu diesem Fund steht der Unterschied des Anteils an Pansen mit Feldfrüchten im
Landhabitat (58 %), welcher aufgrund der fehlenden Pansen mit Feldfrüchten im
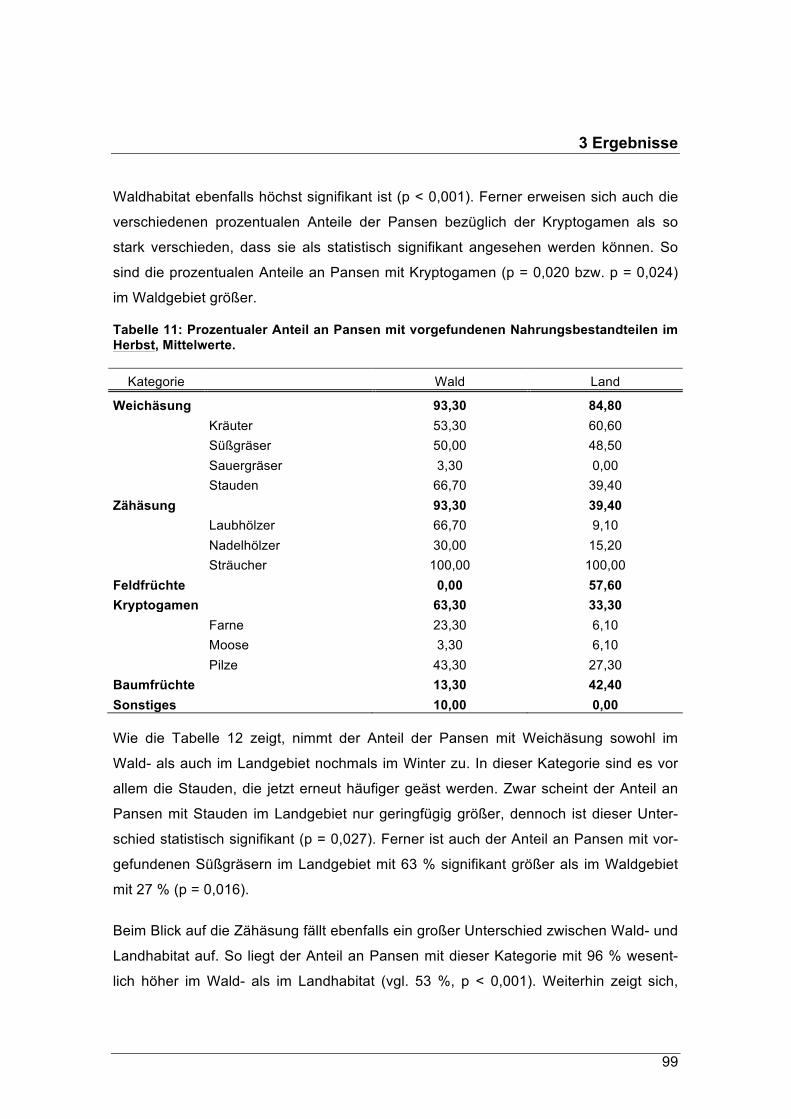
3 Ergebnisse
99
Waldhabitat ebenfalls höchst signifikant ist (p < 0,001). Ferner erweisen sich auch die
verschiedenen prozentualen Anteile der Pansen bezüglich der Kryptogamen als so
stark verschieden, dass sie als statistisch signifikant angesehen werden können. So
sind die prozentualen Anteile an Pansen mit Kryptogamen (p = 0,020 bzw. p = 0,024)
im Waldgebiet größer.
Tabelle 11: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen im Herbst, Mittelwerte.
Kategorie Wald Land
Weichäsung 93,30 84,80
Kräuter 53,30 60,60
Süßgräser 50,00 48,50
Sauergräser 3,30 0,00
Stauden 66,70 39,40
Zähäsung 93,30 39,40
Laubhölzer 66,70 9,10
Nadelhölzer 30,00 15,20
Sträucher 100,00 100,00
Feldfrüchte 0,00 57,60 Kryptogamen 63,30 33,30
Farne 23,30 6,10
Moose 3,30 6,10
Pilze 43,30 27,30
Baumfrüchte 13,30 42,40 Sonstiges 10,00 0,00
Wie die Tabelle 12 zeigt, nimmt der Anteil der Pansen mit Weichäsung sowohl im
Wald- als auch im Landgebiet nochmals im Winter zu. In dieser Kategorie sind es vor
allem die Stauden, die jetzt erneut häufiger geäst werden. Zwar scheint der Anteil an
Pansen mit Stauden im Landgebiet nur geringfügig größer, dennoch ist dieser Unter-
schied statistisch signifikant (p = 0,027). Ferner ist auch der Anteil an Pansen mit vor-
gefundenen Süßgräsern im Landgebiet mit 63 % signifikant größer als im Waldgebiet
mit 27 % (p = 0,016).
Beim Blick auf die Zähäsung fällt ebenfalls ein großer Unterschied zwischen Wald- und
Landhabitat auf. So liegt der Anteil an Pansen mit dieser Kategorie mit 96 % wesent-
lich höher im Wald- als im Landhabitat (vgl. 53 %, p < 0,001). Weiterhin zeigt sich,
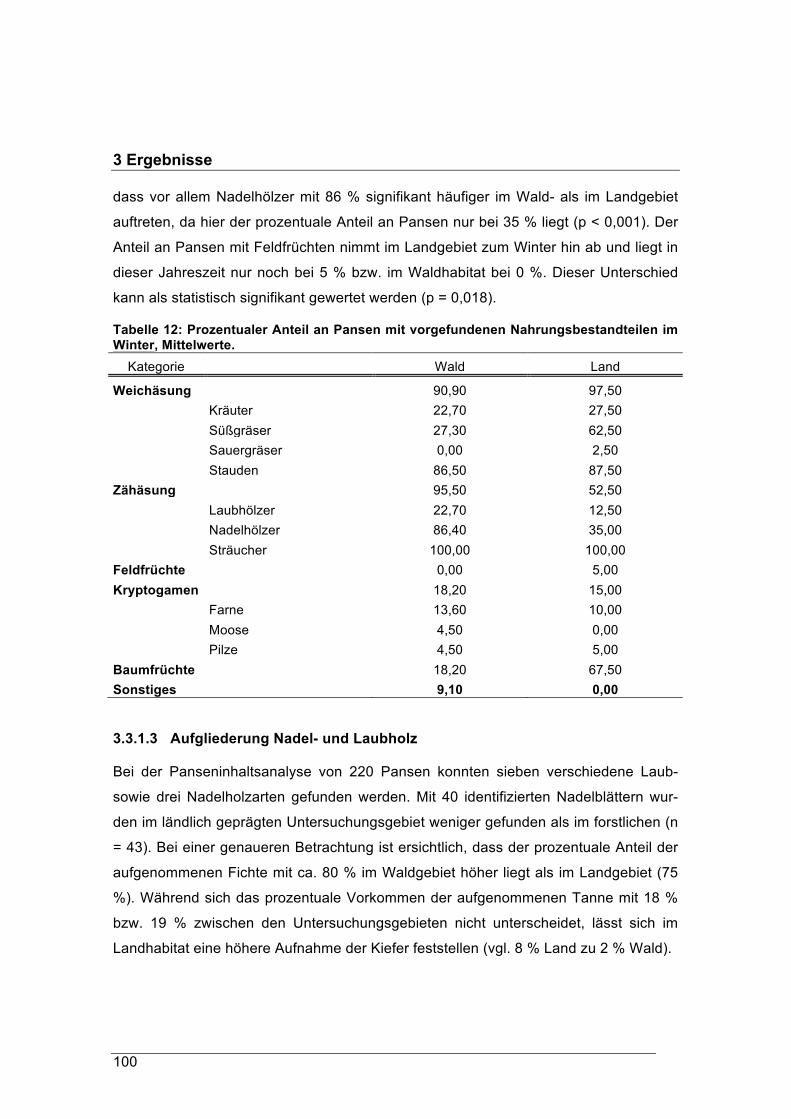
3 Ergebnisse
100
dass vor allem Nadelhölzer mit 86 % signifikant häufiger im Wald- als im Landgebiet
auftreten, da hier der prozentuale Anteil an Pansen nur bei 35 % liegt (p < 0,001). Der
Anteil an Pansen mit Feldfrüchten nimmt im Landgebiet zum Winter hin ab und liegt in
dieser Jahreszeit nur noch bei 5 % bzw. im Waldhabitat bei 0 %. Dieser Unterschied
kann als statistisch signifikant gewertet werden (p = 0,018).
Tabelle 12: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen im Winter, Mittelwerte.
Kategorie Wald Land
Weichäsung 90,90 97,50
Kräuter 22,70 27,50
Süßgräser 27,30 62,50
Sauergräser 0,00 2,50
Stauden 86,50 87,50
Zähäsung 95,50 52,50
Laubhölzer 22,70 12,50
Nadelhölzer 86,40 35,00
Sträucher 100,00 100,00
Feldfrüchte 0,00 5,00 Kryptogamen 18,20 15,00
Farne 13,60 10,00
Moose 4,50 0,00
Pilze 4,50 5,00
Baumfrüchte 18,20 67,50 Sonstiges 9,10 0,00
3.3.1.3 Aufgliederung Nadel- und Laubholz
Bei der Panseninhaltsanalyse von 220 Pansen konnten sieben verschiedene Laub-
sowie drei Nadelholzarten gefunden werden. Mit 40 identifizierten Nadelblättern wur-
den im ländlich geprägten Untersuchungsgebiet weniger gefunden als im forstlichen (n
= 43). Bei einer genaueren Betrachtung ist ersichtlich, dass der prozentuale Anteil der
aufgenommenen Fichte mit ca. 80 % im Waldgebiet höher liegt als im Landgebiet (75
%). Während sich das prozentuale Vorkommen der aufgenommenen Tanne mit 18 %
bzw. 19 % zwischen den Untersuchungsgebieten nicht unterscheidet, lässt sich im
Landhabitat eine höhere Aufnahme der Kiefer feststellen (vgl. 8 % Land zu 2 % Wald).

3 Ergebnisse
101
Tabelle 13: Prozentuale Anteile gefundener Nadel- und Laubhölzer nach Habitaten, Mit-telwerte, n = 220.
Fund Land Wald
n % n %
Laubhölzer 25
100,0
38
100
Ahorn 0
0,0
5
13,2
Buche 2
8,0
14
36,8
Eiche 6
24,0
2
5,26
Erle 1
4,0
0
0,0
Hainbuche 1
4,0
0
0,0
Linde 1
4,0
0
0,0
Pappel 0
0,0
1
2,63
unbek. Laubhölzer 14
56,0
16
42,1
Nadelhölzer 40
100,0
43
100
Fichte 30
75,0
34
79,1
Kiefer 3
7,5
1
2,3
Tanne 7 17,5 8 18,6
In Bezug auf Laubhölzer konnten 25 Funde im Land- sowie 38 im Waldhabitat getätigt
werden. Dabei wurden sowohl Buche als auch Eiche in beiden Untersuchungsgebieten
mit jeweils 8 bzw. 24 % (Land) sowie 33 bzw. 5 % (Wald) identifiziert (Tabelle 13).
Während Ahorn und Pappel mit 12 % bzw. 2 % nur in Pansen der Waldtiere gefunden
wurden, konnten Erle, Hainbuche und Linde mit jeweils 4 % nur in Proben des Land-
gebietes festgestellt werden. Während 56 % der vorgefundenen Laubblätter im LA
nicht genauer bestimmt werden konnten, lag dieser Anteil im FO bei 38 %.
3.3.2 Vegetationskartierung in den Untersuchungsgebieten
3.3.2.1 Arten und ihre Auftretungsfrequenz in der Vegetation
Die verfügbare Vegetation ist im Landgebiet deutlich hin zu Feldfrüchten und Weich-
äsung verschoben. Das Äsungsangebot ist im Wald deutlich breiter als im Agrarhabitat
(Abbildung 44).
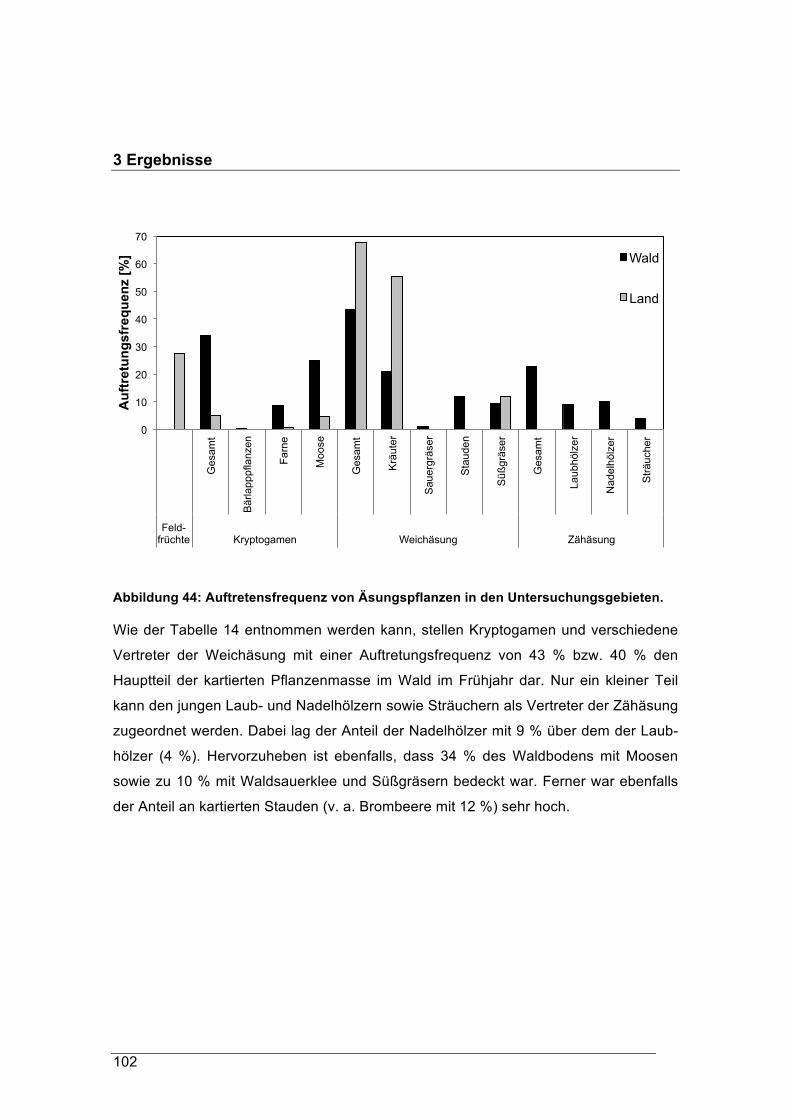
3 Ergebnisse
102
Abbildung 44: Auftretensfrequenz von Äsungspflanzen in den Untersuchungsgebieten.
Wie der Tabelle 14 entnommen werden kann, stellen Kryptogamen und verschiedene
Vertreter der Weichäsung mit einer Auftretungsfrequenz von 43 % bzw. 40 % den
Hauptteil der kartierten Pflanzenmasse im Wald im Frühjahr dar. Nur ein kleiner Teil
kann den jungen Laub- und Nadelhölzern sowie Sträuchern als Vertreter der Zähäsung
zugeordnet werden. Dabei lag der Anteil der Nadelhölzer mit 9 % über dem der Laub-
hölzer (4 %). Hervorzuheben ist ebenfalls, dass 34 % des Waldbodens mit Moosen
sowie zu 10 % mit Waldsauerklee und Süßgräsern bedeckt war. Ferner war ebenfalls
der Anteil an kartierten Stauden (v. a. Brombeere mit 12 %) sehr hoch.
0
10
20
30
40
50
60
70
Ges
amt
Bär
lapp
pfla
nzen
Farn
e
Moo
se
Ges
amt
Krä
uter
Sau
ergr
äser
Sta
uden
Süß
gräs
er
Ges
amt
Laub
hölz
er
Nad
elhö
lzer
Strä
uche
r
Feld- früchte
Kryptogamen
Weichäsung
Zähäsung
Auf
tret
ungs
freq
uenz
[%] Wald
Land

3 Ergebnisse
103
Tabelle 14: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Waldgebiet.
Klasse
Auftretungsfrequenz
[%]
Kryptogamen 43,14
Farne
9,38
Moose
33,75
Weichäsung 40,20
Kräuter
18,91
Sauergräser
0,14
Stauden
12,04
Süßgräser
9,10
Zähäsung 16,67
Laubhölzer
3,50
Nadelhölzer 9,10
Sträucher 4,06
Im Untersuchungsgebiet Land konnten bezüglich der Vegetation im Frühjahr nur zwei
Äsungsklassen kartiert werden, wovon die Weichäsung mit diversen Kräutern sowie
Süßgräsern mit einer Gesamtauftretungsfrequenz von knapp 68 % den größeren Anteil
besitzt (Tabelle 15). Dabei ist in Bezug auf die Weichäsung vor allem der vergleichs-
weise hohe Anteil an Rot- und Weißklee (knapp 7 % bzw. 8 %) und diversen Löwen-
zahnarten (ca. 8 %) zu erwähnen. Im Hinblick auf die gezielt anthropogen ausgebrach-
te Vegetation stellt vor allem das Wintergetreide mit ca. 24 % bei den Feldfrüchten die
Vegetationsart mit der höchsten Auftretungsfrequenz dar.
Tabelle 15: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Landgebiet.
Klasse
Auftretungsfrequenz [ %]
Feldfrüchte 32,29 Weichäsung 67,71
Kräuter 51,78
Süßgräser 15,93
Im Vergleich zum Frühjahr sinkt im Sommer im Untersuchungsgebiet Wald die Auftre-
tungsfrequenz von Kryptogamen drastisch auf nur noch ca. 10 %, wohingegen die
Weichäsung um ca. 20 % auf insgesamt 61 % zunimmt (Tabelle 16). Vor allem der
Anteil des Springkrautes steigt von unter 1 % im Frühjahr auf 6 % der kartierten Vege-

3 Ergebnisse
104
tation im Sommer. Während der Anteil der Brennnessel ebenfalls eine leichte Steige-
rung in der Auftretungsfrequenz zeigt, bleibt die des Waldsauerklees ungefähr mit
knapp 9 % auf dem gleichen Niveau wie im Frühjahr. Bezugnehmend auf die
Zähäsung ist eine Steigerung in der Auftretungsfrequenz auf 29 % ersichtlich. Dies ist
mit der Zunahme an kartierten Laubhölzern von 4 % im Frühjahr auf 14 % im Sommer
zu erklären.
Tabelle 16: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Waldgebiet.
Klasse
Auftretungsfrequenz
[%]
Kryptogamen 10,34
Farne
5,75
Moose
4,60
Weichäsung 60,92
Kräuter
33,91
Sauergräser
1,15
Stauden
13,79
Süßgräser
12,07
Zähäsung 28,74
Laubhölzer
14,37
Nadelhölzer 9,77
Sträucher 4,60
Im Sommer steigt der Anteil der Auftretungsfrequenz anthropogener Pflanzen im Un-
tersuchungsgebiet Land. Während das Wintergetreide noch nicht abgeerntet ist und
daher immer noch gut 25 % der kartierten Pflanzen darstellt (Tabelle 17), erfolgt in die-
ser Zeit ebenfalls der Maisanbau auf brachen Flächen. Für diese Kulturpflanze ergibt
sich zusammenfassend eine Auftretungsfrequenz von ca. 15 %. Im Vergleich zum
Frühjahr sinkt der Anteil der Weichäsung auf rund 58 %. Vor allem Weißklee und Lö-
wenzahn bleiben in dieser Klasse die Pflanzen mit dem häufigsten Auftreten.
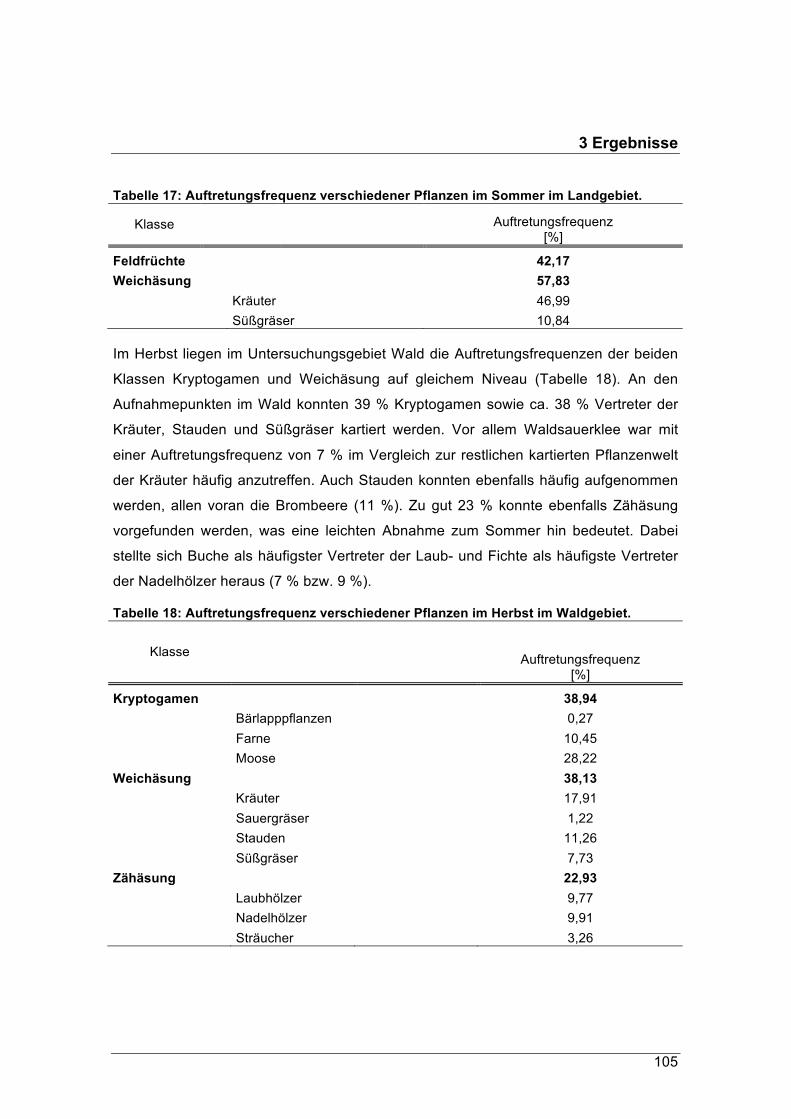
3 Ergebnisse
105
Tabelle 17: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Landgebiet.
Klasse
Auftretungsfrequenz [%]
Feldfrüchte 42,17 Weichäsung 57,83
Kräuter 46,99
Süßgräser 10,84
Im Herbst liegen im Untersuchungsgebiet Wald die Auftretungsfrequenzen der beiden
Klassen Kryptogamen und Weichäsung auf gleichem Niveau (Tabelle 18). An den
Aufnahmepunkten im Wald konnten 39 % Kryptogamen sowie ca. 38 % Vertreter der
Kräuter, Stauden und Süßgräser kartiert werden. Vor allem Waldsauerklee war mit
einer Auftretungsfrequenz von 7 % im Vergleich zur restlichen kartierten Pflanzenwelt
der Kräuter häufig anzutreffen. Auch Stauden konnten ebenfalls häufig aufgenommen
werden, allen voran die Brombeere (11 %). Zu gut 23 % konnte ebenfalls Zähäsung
vorgefunden werden, was eine leichten Abnahme zum Sommer hin bedeutet. Dabei
stellte sich Buche als häufigster Vertreter der Laub- und Fichte als häufigste Vertreter
der Nadelhölzer heraus (7 % bzw. 9 %).
Tabelle 18: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Waldgebiet.
Klasse
Auftretungsfrequenz
[%]
Kryptogamen 38,94
Bärlapppflanzen 0,27
Farne
10,45
Moose
28,22
Weichäsung 38,13
Kräuter
17,91
Sauergräser
1,22
Stauden
11,26
Süßgräser
7,73
Zähäsung 22,93
Laubhölzer
9,77
Nadelhölzer 9,91
Sträucher 3,26

3 Ergebnisse
106
Tabelle 19 zeigt im Vergleich zu den Auftretungsfrequenzen des Sommers eine klare
Senkung des Anteils der anthropogenen Pflanzen im Herbst im Landgebiet um ca. 25
%. Im Gegenzug steigt der Anteil der Weichäsung auf 80 % an. Während noch Rest-
bestände des Maises im frühen sowie die Aussaat des Wintergetreides im späten
Herbst die vergleichsweise hohen Auftretungsfrequenzen dieser beiden Kulturformen
innerhalb der Feldfrüchte erklären können, stellen Löwenzahn und verschiedene Klee-
arten den Großteil der Weichäsung dar.
Tabelle 19: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Landgebiet.
Klasse
Auftretungsfrequenz [%]
Feldfrüchte 17,99 Kryptogamen 1,90
Farne
1,90
Weichäsung 80,10
Kräuter 61,59
Süßgräser 18,51
In den Monaten des Winters steigt die prozentuale Auftretungsfrequenz von Kryptoga-
men im Waldgebiet weiter an (Tabelle 20). Dieser Anstieg ist dem vermehrten Vor-
kommen von Moosen um ca. 5 % auf 33 % zuzuschreiben. Während der Anteil der
Zähäsung gleichbleibend bei 23 % liegt, sinkt die Auftretungshäufigkeit von Vertretern
der Weichäsung. Dabei bleiben sowohl Brombeeren als auch der Waldsauerklee mit
10 % und 7 % die häufigsten kartierten Pflanzen dieser Klasse. Jedoch ist eine Ab-
nahme der Laubhölzer auf 8 % und eine Zunahme der Nadelhölzer auf 12 % festzu-
stellen.
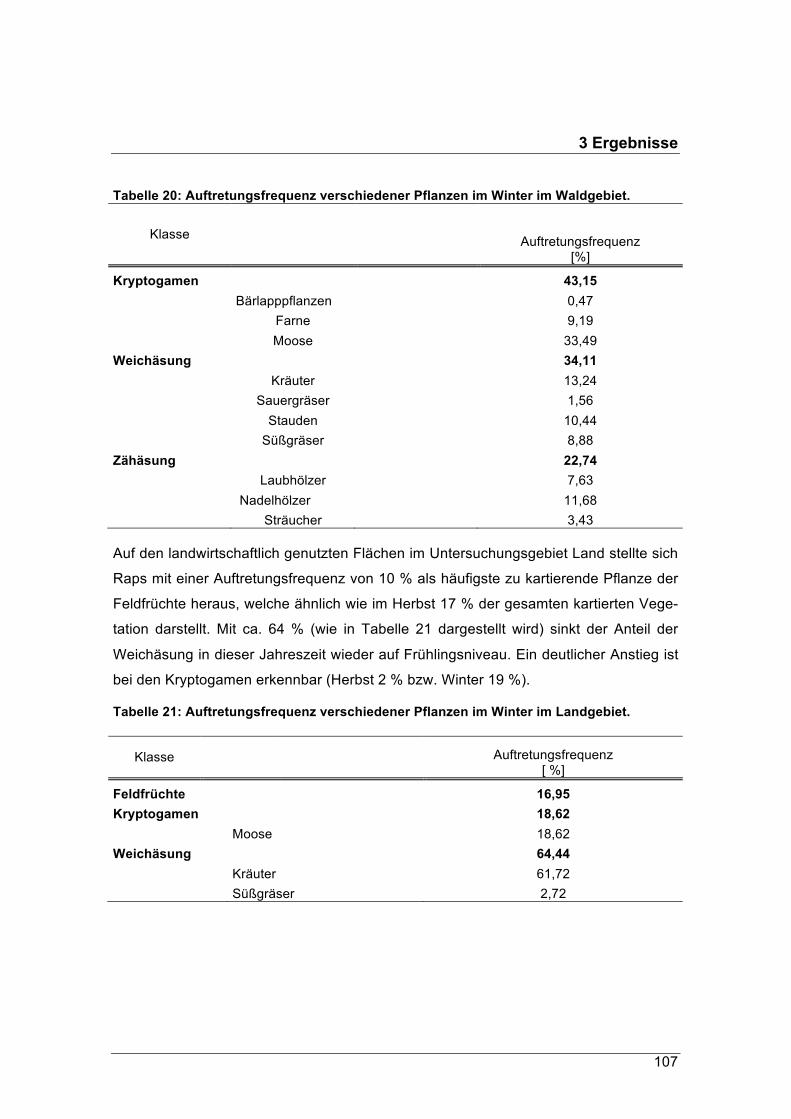
3 Ergebnisse
107
Tabelle 20: Auftretungsfrequenz verschiedener Pflanzen im Winter im Waldgebiet.
Klasse
Auftretungsfrequenz
[%]
Kryptogamen 43,15
Bärlapppflanzen 0,47
Farne
9,19
Moose
33,49
Weichäsung 34,11
Kräuter
13,24
Sauergräser
1,56
Stauden
10,44
Süßgräser
8,88
Zähäsung 22,74
Laubhölzer
7,63
Nadelhölzer 11,68
Sträucher 3,43
Auf den landwirtschaftlich genutzten Flächen im Untersuchungsgebiet Land stellte sich
Raps mit einer Auftretungsfrequenz von 10 % als häufigste zu kartierende Pflanze der
Feldfrüchte heraus, welche ähnlich wie im Herbst 17 % der gesamten kartierten Vege-
tation darstellt. Mit ca. 64 % (wie in Tabelle 21 dargestellt wird) sinkt der Anteil der
Weichäsung in dieser Jahreszeit wieder auf Frühlingsniveau. Ein deutlicher Anstieg ist
bei den Kryptogamen erkennbar (Herbst 2 % bzw. Winter 19 %).
Tabelle 21: Auftretungsfrequenz verschiedener Pflanzen im Winter im Landgebiet.
Klasse
Auftretungsfrequenz [ %]
Feldfrüchte 16,95 Kryptogamen 18,62
Moose 18,62
Weichäsung 64,44
Kräuter 61,72
Süßgräser 2,72
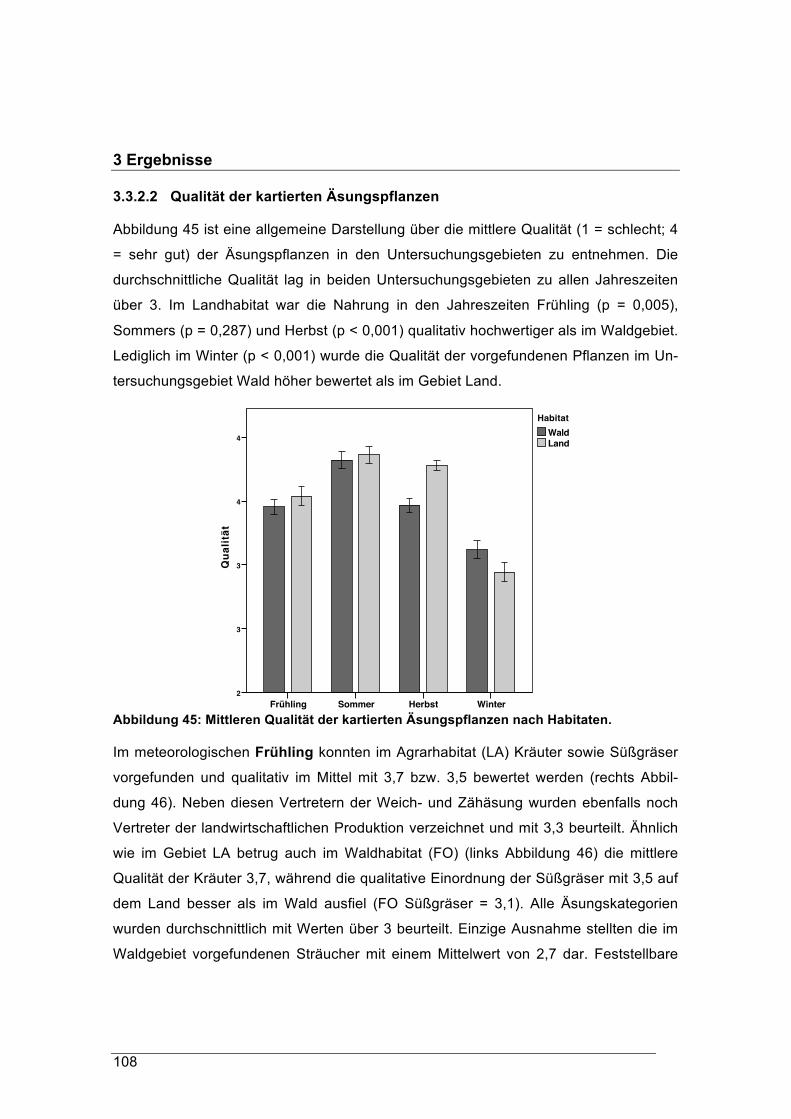
3 Ergebnisse
108
3.3.2.2 Qualität der kartierten Äsungspflanzen
Abbildung 45 ist eine allgemeine Darstellung über die mittlere Qualität (1 = schlecht; 4
= sehr gut) der Äsungspflanzen in den Untersuchungsgebieten zu entnehmen. Die
durchschnittliche Qualität lag in beiden Untersuchungsgebieten zu allen Jahreszeiten
über 3. Im Landhabitat war die Nahrung in den Jahreszeiten Frühling (p = 0,005),
Sommers (p = 0,287) und Herbst (p < 0,001) qualitativ hochwertiger als im Waldgebiet.
Lediglich im Winter (p < 0,001) wurde die Qualität der vorgefundenen Pflanzen im Un-
tersuchungsgebiet Wald höher bewertet als im Gebiet Land.
Abbildung 45: Mittleren Qualität der kartierten Äsungspflanzen nach Habitaten.
Im meteorologischen Frühling konnten im Agrarhabitat (LA) Kräuter sowie Süßgräser
vorgefunden und qualitativ im Mittel mit 3,7 bzw. 3,5 bewertet werden (rechts Abbil-
dung 46). Neben diesen Vertretern der Weich- und Zähäsung wurden ebenfalls noch
Vertreter der landwirtschaftlichen Produktion verzeichnet und mit 3,3 beurteilt. Ähnlich
wie im Gebiet LA betrug auch im Waldhabitat (FO) (links Abbildung 46) die mittlere
Qualität der Kräuter 3,7, während die qualitative Einordnung der Süßgräser mit 3,5 auf
dem Land besser als im Wald ausfiel (FO Süßgräser = 3,1). Alle Äsungskategorien
wurden durchschnittlich mit Werten über 3 beurteilt. Einzige Ausnahme stellten die im
Waldgebiet vorgefundenen Sträucher mit einem Mittelwert von 2,7 dar. Feststellbare
WinterHerbstSommerFrühling
Qua
lität
4
4
3
3
2
LandWald
Habitat
Page 1
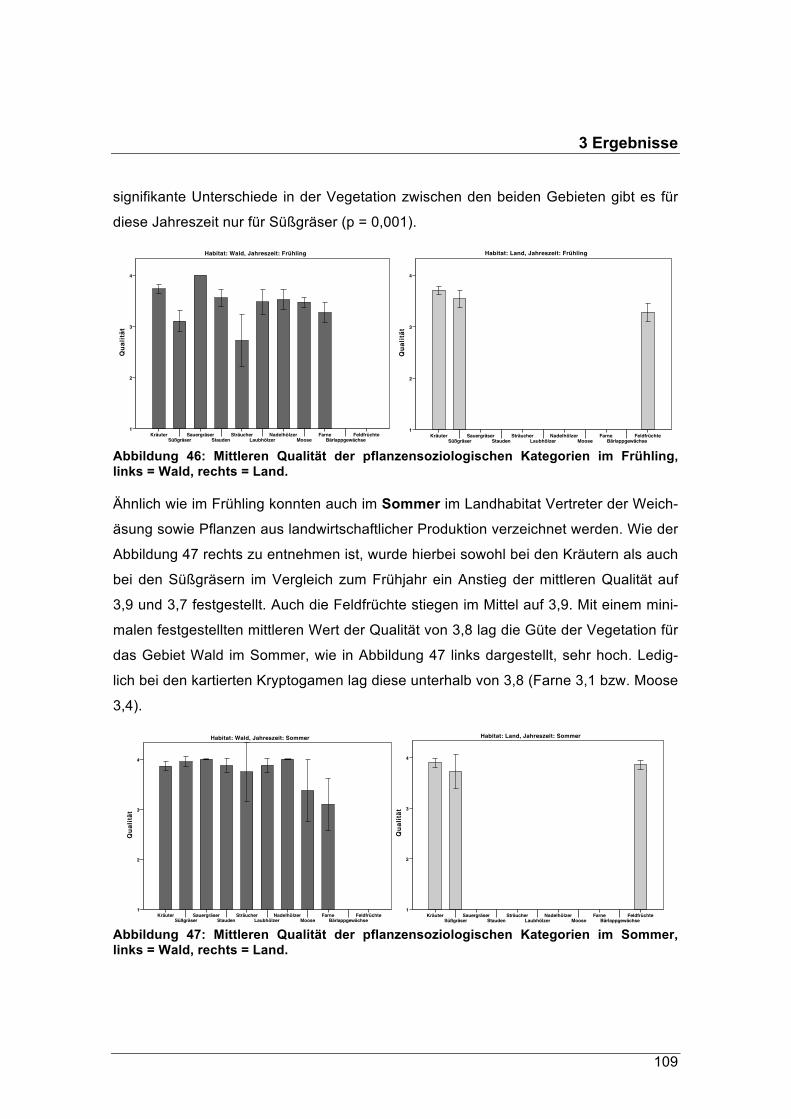
3 Ergebnisse
109
signifikante Unterschiede in der Vegetation zwischen den beiden Gebieten gibt es für
diese Jahreszeit nur für Süßgräser (p = 0,001).
Abbildung 46: Mittleren Qualität der pflanzensoziologischen Kategorien im Frühling, links = Wald, rechts = Land.
Ähnlich wie im Frühling konnten auch im Sommer im Landhabitat Vertreter der Weich-
äsung sowie Pflanzen aus landwirtschaftlicher Produktion verzeichnet werden. Wie der
Abbildung 47 rechts zu entnehmen ist, wurde hierbei sowohl bei den Kräutern als auch
bei den Süßgräsern im Vergleich zum Frühjahr ein Anstieg der mittleren Qualität auf
3,9 und 3,7 festgestellt. Auch die Feldfrüchte stiegen im Mittel auf 3,9. Mit einem mini-
malen festgestellten mittleren Wert der Qualität von 3,8 lag die Güte der Vegetation für
das Gebiet Wald im Sommer, wie in Abbildung 47 links dargestellt, sehr hoch. Ledig-
lich bei den kartierten Kryptogamen lag diese unterhalb von 3,8 (Farne 3,1 bzw. Moose
3,4).
Abbildung 47: Mittleren Qualität der pflanzensoziologischen Kategorien im Sommer, links = Wald, rechts = Land.
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Wald, Jahreszeit: Frühling
Page 1
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Land, Jahreszeit: Frühling
Page 1
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Wald, Jahreszeit: Sommer
Page 1
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Land, Jahreszeit: Sommer
Page 1

3 Ergebnisse
110
Im Herbst konnten wie bereits im Frühling und Sommer diverse Kräuter, Süßgräser
und Feldfrüchte im Landgebiet kartiert werden (Abbildung 48 rechts). Während die
mittlere Qualität der Äsungskategorie der Kräuter und Feldfrüchte im Vergleich zum
Sommer auf 3,8 bzw. 3,7 leicht abnahm, stieg die der Süßgräser minimal auf 3,8. Fer-
ner wurden erstmals in dieser Zeit für das Landgebiet Farne kartiert. Diese wiesen eine
mittlere Qualität von 3,4 auf. Mit Ausnahme der Äsungskategorie der Kryptogamen
wurden für das Waldgebiet für alle Äsungsarten Abnahmen in der mittleren Qualität
festgestellt (Abbildung 48). Mit Ausnahme der Sträucher (2,6) lagen dennoch alle
Pflanzenarten im Mittel über einer Qualität von 3,0. Die zu den Kryptogamen gehören-
den Farne und Moose konnten Qualitäten von durchschnittlich 3,4 und 3,8 protokolliert
werden. Zwischen beiden Gebieten konnten signifikante Unterschiede zwischen der
Qualität der Kräuter und Süßgräser errechnet werden (jeweils p < 0,001).
Abbildung 48: Mittlere Qualität der pflanzensoziologischen Kategorien im Herbst, links = Wald, rechts = Land.
Im Winter folgte eine weitere Abnahme der mittleren Qualität der im Landgebiet kar-
tierten Agrarpflanzen, Kräuter und Süßgräser. So betrug diese für die erste und zweite
Äsungsart nur noch für 2,9 sowie für die dritte 3,2. Wie in der Abbildung 49 rechts dar-
gestellt, konnten auch in dieser Jahreszeit keine Farne kartiert werden. Jedoch wurden
im Landgebiet Moose vorgefunden, welche im Mittel eine Qualität von 3,0 aufwiesen.
Auch im Waldhabitat konnte eine Senkung in der Qualität der Weichäsung im Ver-
gleich zur vorhergehenden Jahreszeit diagnostiziert werden (links Abbildung 49). Wäh-
rend die Sträucher als Vertreter der Zähäsung ebenfalls in ihrer Qualität abnahmen,
wurde für die zur gleichen Äsungskategorie gehörenden Laub- und Nadelbäume ein
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Wald, Jahreszeit: Herbst
Page 1
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Land, Jahreszeit: Herbst
Page 1
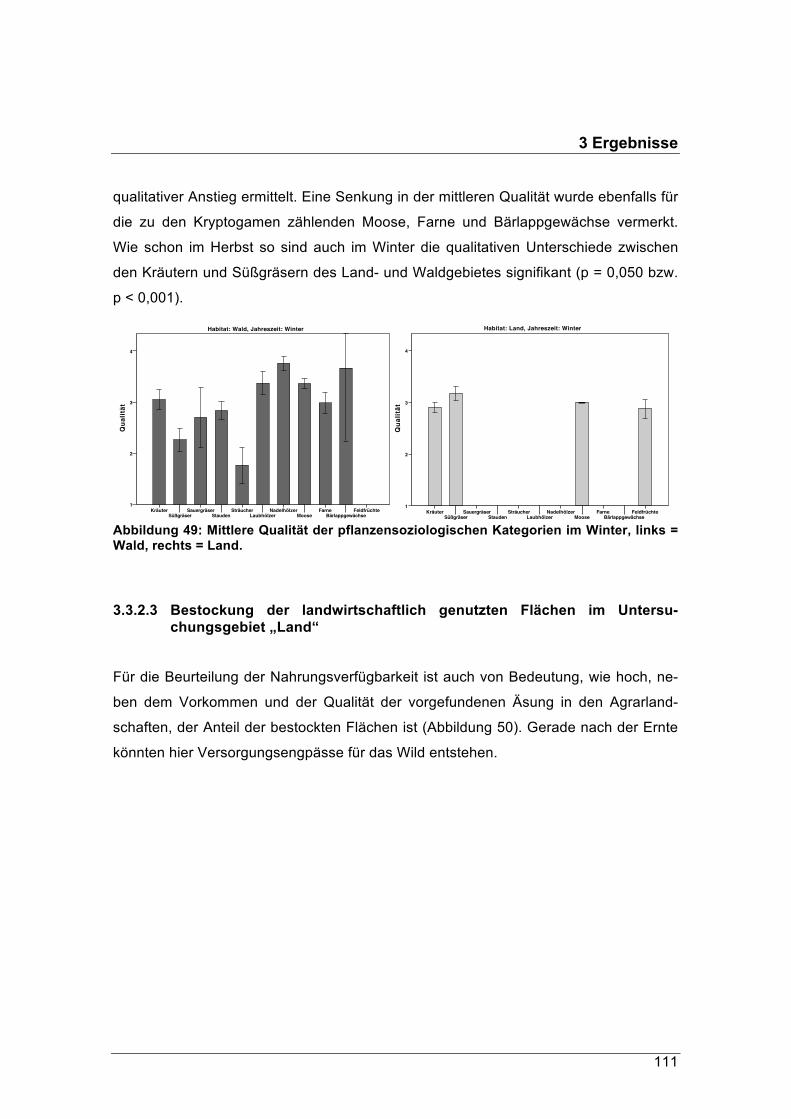
3 Ergebnisse
111
qualitativer Anstieg ermittelt. Eine Senkung in der mittleren Qualität wurde ebenfalls für
die zu den Kryptogamen zählenden Moose, Farne und Bärlappgewächse vermerkt.
Wie schon im Herbst so sind auch im Winter die qualitativen Unterschiede zwischen
den Kräutern und Süßgräsern des Land- und Waldgebietes signifikant (p = 0,050 bzw.
p < 0,001).
Abbildung 49: Mittlere Qualität der pflanzensoziologischen Kategorien im Winter, links = Wald, rechts = Land.
3.3.2.3 Bestockung der landwirtschaftlich genutzten Flächen im Untersu-chungsgebiet „Land“
Für die Beurteilung der Nahrungsverfügbarkeit ist auch von Bedeutung, wie hoch, ne-
ben dem Vorkommen und der Qualität der vorgefundenen Äsung in den Agrarland-
schaften, der Anteil der bestockten Flächen ist (Abbildung 50). Gerade nach der Ernte
könnten hier Versorgungsengpässe für das Wild entstehen.
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Wald, Jahreszeit: Winter
Page 1
FeldfrüchteBärlappgewächse
FarneMoose
NadelhölzerLaubhölzer
SträucherStauden
SauergräserSüßgräser
Kräuter
Qua
lität
4
3
2
1
Habitat: Land, Jahreszeit: Winter
Page 1
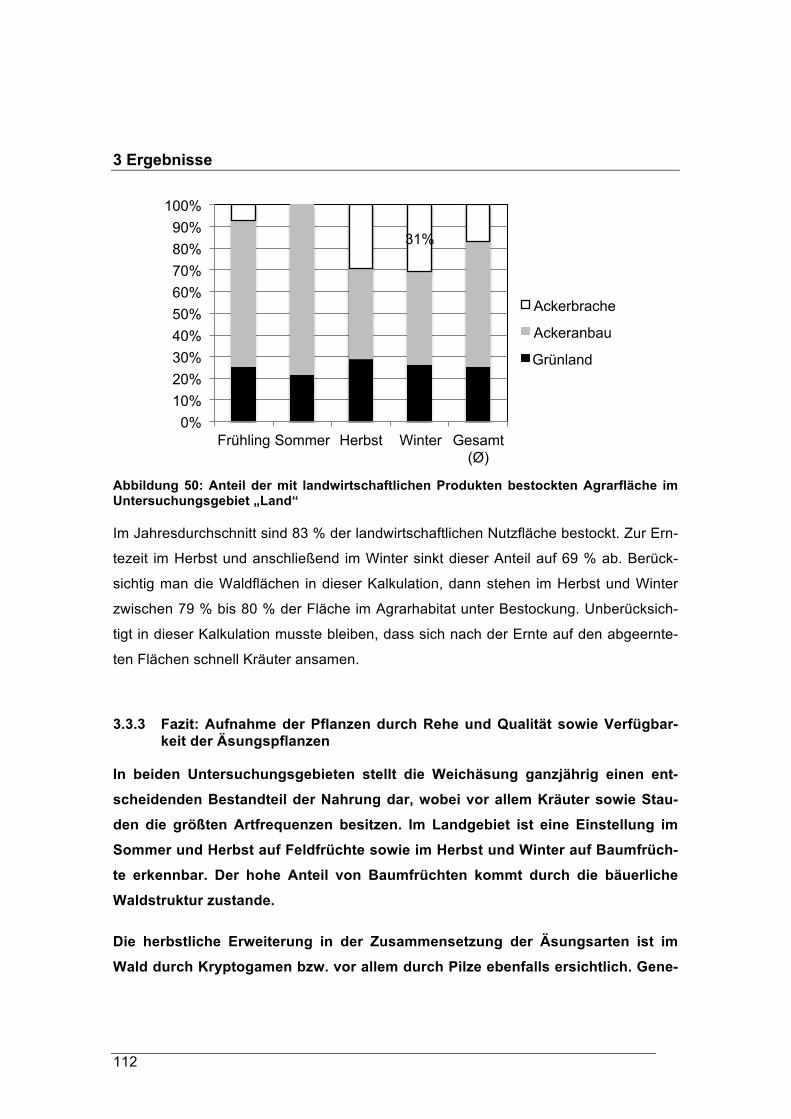
3 Ergebnisse
112
Abbildung 50: Anteil der mit landwirtschaftlichen Produkten bestockten Agrarfläche im Untersuchungsgebiet „Land“
Im Jahresdurchschnitt sind 83 % der landwirtschaftlichen Nutzfläche bestockt. Zur Ern-
tezeit im Herbst und anschließend im Winter sinkt dieser Anteil auf 69 % ab. Berück-
sichtig man die Waldflächen in dieser Kalkulation, dann stehen im Herbst und Winter
zwischen 79 % bis 80 % der Fläche im Agrarhabitat unter Bestockung. Unberücksich-
tigt in dieser Kalkulation musste bleiben, dass sich nach der Ernte auf den abgeernte-
ten Flächen schnell Kräuter ansamen.
3.3.3 Fazit: Aufnahme der Pflanzen durch Rehe und Qualität sowie Verfügbar-keit der Äsungspflanzen
In beiden Untersuchungsgebieten stellt die Weichäsung ganzjährig einen ent-
scheidenden Bestandteil der Nahrung dar, wobei vor allem Kräuter sowie Stau-
den die größten Artfrequenzen besitzen. Im Landgebiet ist eine Einstellung im
Sommer und Herbst auf Feldfrüchte sowie im Herbst und Winter auf Baumfrüch-
te erkennbar. Der hohe Anteil von Baumfrüchten kommt durch die bäuerliche
Waldstruktur zustande.
Die herbstliche Erweiterung in der Zusammensetzung der Äsungsarten ist im
Wald durch Kryptogamen bzw. vor allem durch Pilze ebenfalls ersichtlich. Gene-
31%
0% 10% 20% 30% 40% 50% 60% 70% 80% 90%
100%
Frühling Sommer Herbst Winter Gesamt (Ø)
Ackerbrache
Ackeranbau
Grünland

3 Ergebnisse
113
rell werden bezüglich der Zähäsung mehr Laub- und Nadelhölzer im Wald- als im
Landgebiet aufgenommen. Innerhalb der Laubhölzer scheint eine Präferenz für
die Aufnahme von Buchen zu bestehen. Die am häufigsten aufgenommene Na-
delbaumart ist die Fichte, die am meisten bevorzugte ist die Tanne, wie sich aus
dem Vergleich von Aufnahmehäufigkeit und Verfügbarkeit zeigt. Zu jeder Jahres-
zeit lassen sich in beiden Gebieten Heidelbeeren in den Pansen nachweisen.
Anhand der Kartierung zeigt sich die dominante Prägung des Landes durch
landwirtschaftliche Ackerpflanzen, die ganzjährig zu einer qualitativ guten und in
den verschiedenen Jahreszeiten stets wechselhaften Äsung führen. Selbst auf
brachliegenden Flächen lassen sich für das Rehwild wertvolle Ackerunkräuter
finden. Ferner bietet die Vielzahl an Wiesen und Weiden ebenfalls zu jeder Jah-
reszeit ein in der Qualität sehr gut bewertetes Äsungsangebot mit verschiedens-
ten Gräsern und Kräutern.
Im Untersuchungsgebiet Wald stehen den Rehen ganzjährig vor allem Vertreter
der Zähäsung zur Verfügung. So sind nicht nur Laub- und Nadelbäume im Gebiet
kartiert worden, sondern großflächig auch die zu den Sträuchern zählende Hei-
delbeere. Ebenfalls zeigt sich fast ganzjährig auf Lichtungen, Wegen und Rücke-
gassen eine Vielfalt von Kräutern und Gräsern, welche durch optimale Bedin-
gungen im Sommer sowie aufgrund des isolierenden Schutzes des Schnees im
Winter jederzeit in Bezug auf ihre Qualität mit hohen Werten beurteilt werden
konnten.
Beide Untersuchungsgebiete können bezüglich ihres Äsungsangebotes als ar-
tenreich und qualitativ wertvoll eingestuft werden.
Beide Untersuchungsgebiete bieten das ganze Jahr hinweg dem Wild Äsung.
Eine bedeutende Einschränkung des Angebots durch die Ernte findet nicht statt.
3.4 Anpassung des Pansens (Scheingraber)
Um die Frage der physiologischen Anpassung des Pansens hinsichtlich der Jahreszei-
ten, aber auch an das jeweilige Habitat zu beantworten, wurden Pansenvolumen [l] (n
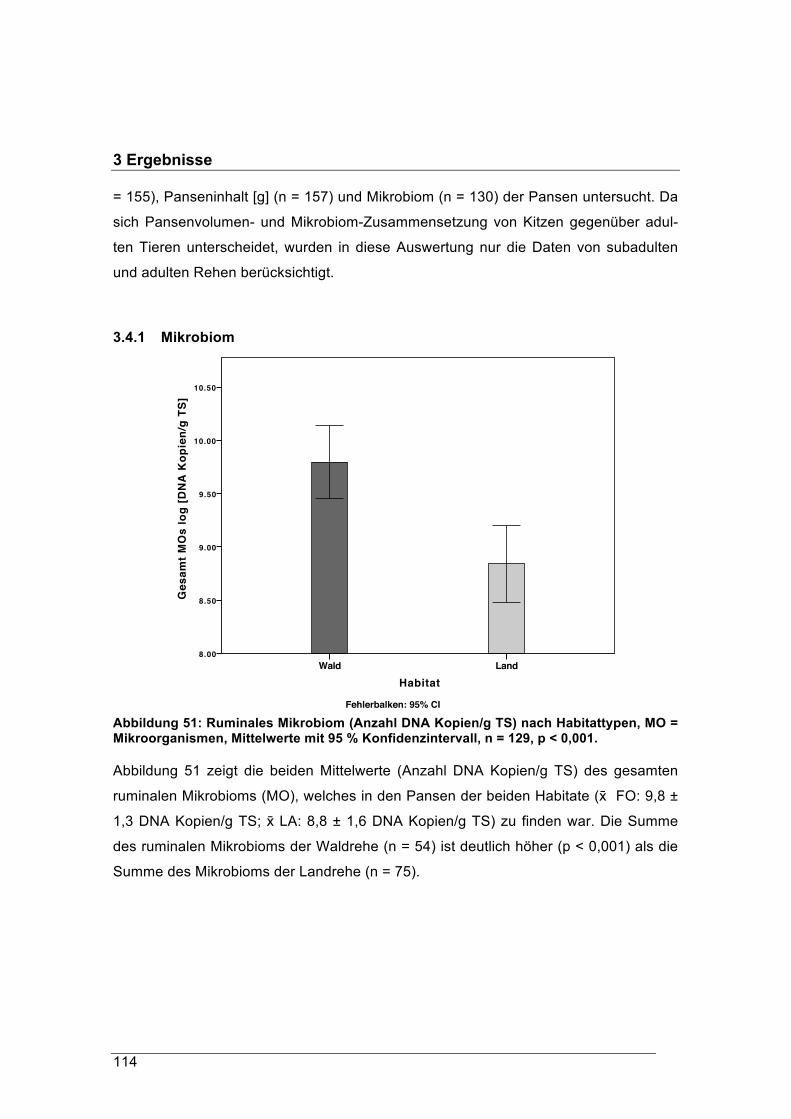
3 Ergebnisse
114
= 155), Panseninhalt [g] (n = 157) und Mikrobiom (n = 130) der Pansen untersucht. Da
sich Pansenvolumen- und Mikrobiom-Zusammensetzung von Kitzen gegenüber adul-
ten Tieren unterscheidet, wurden in diese Auswertung nur die Daten von subadulten
und adulten Rehen berücksichtigt.
3.4.1 Mikrobiom
Abbildung 51: Ruminales Mikrobiom (Anzahl DNA Kopien/g TS) nach Habitattypen, MO = Mikroorganismen, Mittelwerte mit 95 % Konfidenzintervall, n = 129, p < 0,001.
Abbildung 51 zeigt die beiden Mittelwerte (Anzahl DNA Kopien/g TS) des gesamten
ruminalen Mikrobioms (MO), welches in den Pansen der beiden Habitate (x̄ FO: 9,8 ±
1,3 DNA Kopien/g TS; x̄ LA: 8,8 ± 1,6 DNA Kopien/g TS) zu finden war. Die Summe
des ruminalen Mikrobioms der Waldrehe (n = 54) ist deutlich höher (p < 0,001) als die
Summe des Mikrobioms der Landrehe (n = 75).
HabitatLandWald
Ges
amt M
Os
log
[DN
A K
opie
n/g
TS]
10.50
10.00
9.50
9.00
8.50
8.00
Fehlerbalken: 95% CI
Page 1
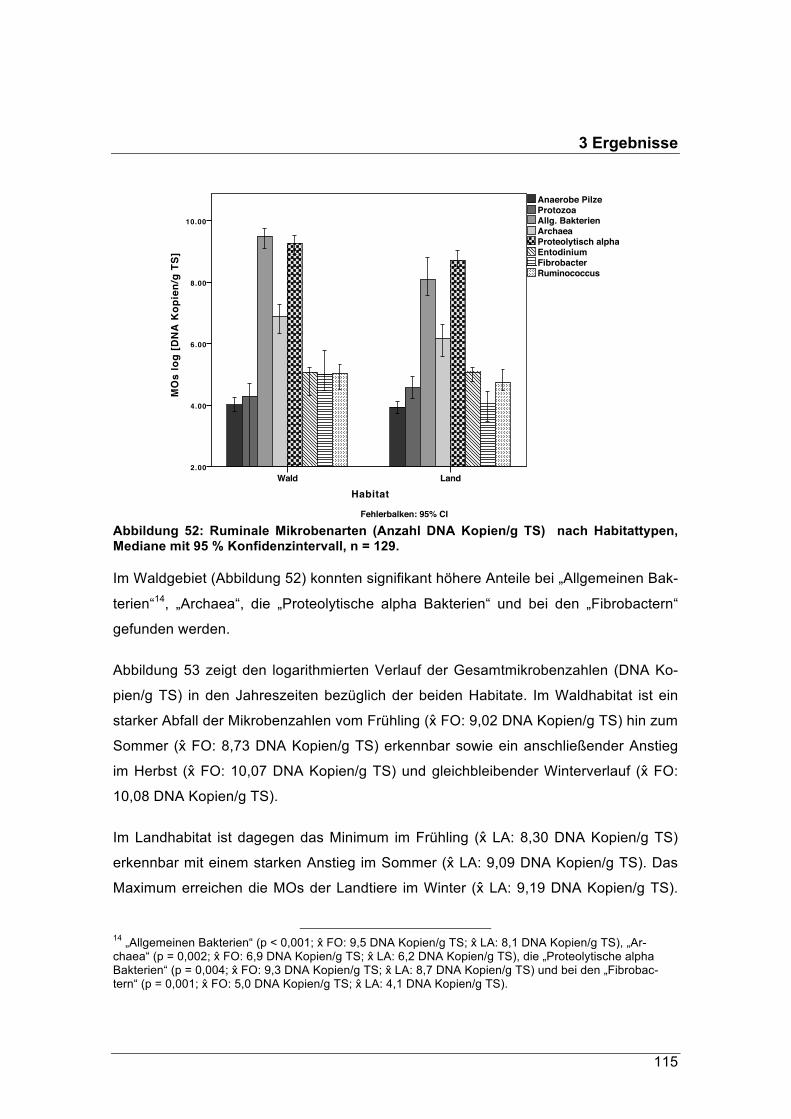
3 Ergebnisse
115
Abbildung 52: Ruminale Mikrobenarten (Anzahl DNA Kopien/g TS) nach Habitattypen, Mediane mit 95 % Konfidenzintervall, n = 129.
Im Waldgebiet (Abbildung 52) konnten signifikant höhere Anteile bei „Allgemeinen Bak-
terien“14, „Archaea“, die „Proteolytische alpha Bakterien“ und bei den „Fibrobactern“
gefunden werden.
Abbildung 53 zeigt den logarithmierten Verlauf der Gesamtmikrobenzahlen (DNA Ko-
pien/g TS) in den Jahreszeiten bezüglich der beiden Habitate. Im Waldhabitat ist ein
starker Abfall der Mikrobenzahlen vom Frühling (x̂ FO: 9,02 DNA Kopien/g TS) hin zum
Sommer (x̂ FO: 8,73 DNA Kopien/g TS) erkennbar sowie ein anschließender Anstieg
im Herbst (x̂ FO: 10,07 DNA Kopien/g TS) und gleichbleibender Winterverlauf (x̂ FO:
10,08 DNA Kopien/g TS).
Im Landhabitat ist dagegen das Minimum im Frühling (x̂ LA: 8,30 DNA Kopien/g TS)
erkennbar mit einem starken Anstieg im Sommer (x̂ LA: 9,09 DNA Kopien/g TS). Das
Maximum erreichen die MOs der Landtiere im Winter (x̂ LA: 9,19 DNA Kopien/g TS).
14 „Allgemeinen Bakterien“ (p < 0,001; x̂ FO: 9,5 DNA Kopien/g TS; x̂ LA: 8,1 DNA Kopien/g TS), „Ar-chaea“ (p = 0,002; x̂ FO: 6,9 DNA Kopien/g TS; x̂ LA: 6,2 DNA Kopien/g TS), die „Proteolytische alpha Bakterien“ (p = 0,004; x̂ FO: 9,3 DNA Kopien/g TS; x̂ LA: 8,7 DNA Kopien/g TS) und bei den „Fibrobac-tern“ (p = 0,001; x̂ FO: 5,0 DNA Kopien/g TS; x̂ LA: 4,1 DNA Kopien/g TS).
HabitatLandWald
MO
s lo
g [D
NA
Kop
ien/
g TS
]
10.00
8.00
6.00
4.00
2.00
Fehlerbalken: 95% CI
RuminococcusFibrobacter Entodinium Proteolytisch alphaArchaea Allg. BakterienProtozoa Anaerobe Pilze
Page 1
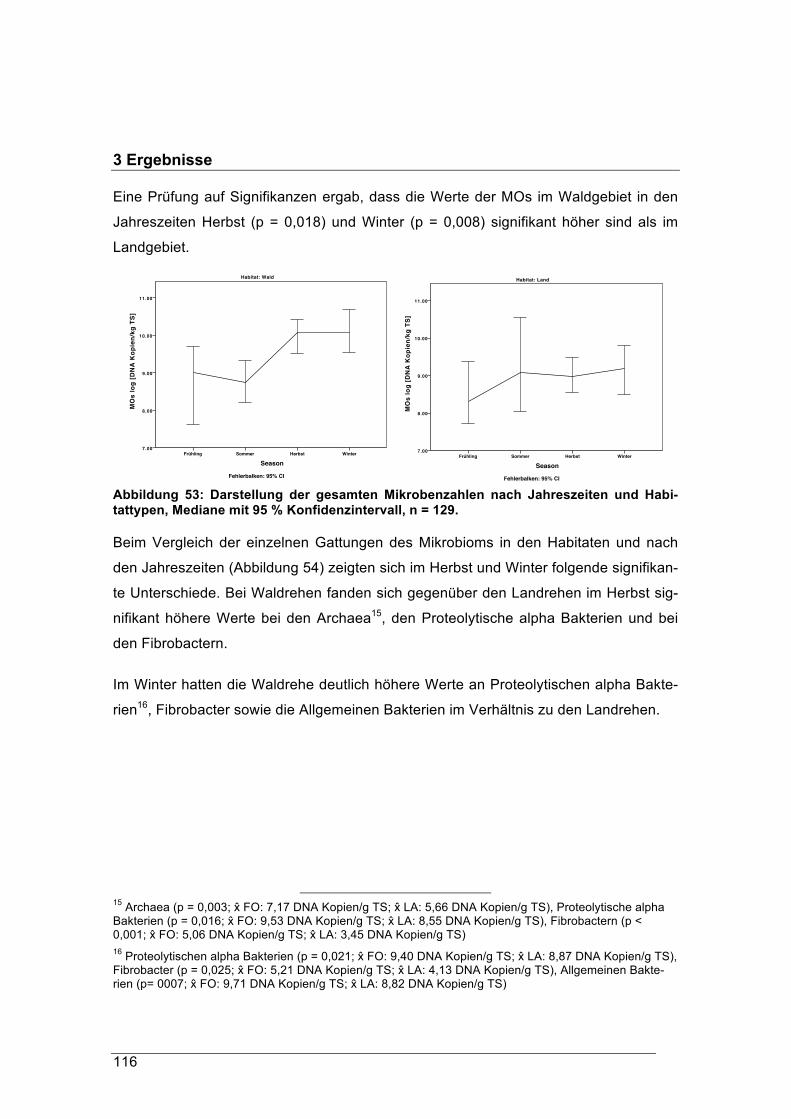
3 Ergebnisse
116
Eine Prüfung auf Signifikanzen ergab, dass die Werte der MOs im Waldgebiet in den
Jahreszeiten Herbst (p = 0,018) und Winter (p = 0,008) signifikant höher sind als im
Landgebiet.
Abbildung 53: Darstellung der gesamten Mikrobenzahlen nach Jahreszeiten und Habi-tattypen, Mediane mit 95 % Konfidenzintervall, n = 129.
Beim Vergleich der einzelnen Gattungen des Mikrobioms in den Habitaten und nach
den Jahreszeiten (Abbildung 54) zeigten sich im Herbst und Winter folgende signifikan-
te Unterschiede. Bei Waldrehen fanden sich gegenüber den Landrehen im Herbst sig-
nifikant höhere Werte bei den Archaea15, den Proteolytische alpha Bakterien und bei
den Fibrobactern.
Im Winter hatten die Waldrehe deutlich höhere Werte an Proteolytischen alpha Bakte-
rien16, Fibrobacter sowie die Allgemeinen Bakterien im Verhältnis zu den Landrehen.
15 Archaea (p = 0,003; x̂ FO: 7,17 DNA Kopien/g TS; x̂ LA: 5,66 DNA Kopien/g TS), Proteolytische alpha Bakterien (p = 0,016; x̂ FO: 9,53 DNA Kopien/g TS; x̂ LA: 8,55 DNA Kopien/g TS), Fibrobactern (p < 0,001; x̂ FO: 5,06 DNA Kopien/g TS; x̂ LA: 3,45 DNA Kopien/g TS) 16 Proteolytischen alpha Bakterien (p = 0,021; x̂ FO: 9,40 DNA Kopien/g TS; x̂ LA: 8,87 DNA Kopien/g TS), Fibrobacter (p = 0,025; x̂ FO: 5,21 DNA Kopien/g TS; x̂ LA: 4,13 DNA Kopien/g TS), Allgemeinen Bakte-rien (p= 0007; x̂ FO: 9,71 DNA Kopien/g TS; x̂ LA: 8,82 DNA Kopien/g TS)
SeasonWinterHerbstSommerFrühling
MO
s lo
g [D
NA
Kop
ien/
kg T
S]
11.00
10.00
9.00
8.00
7.00
Fehlerbalken: 95% CI
Habitat: Wald
Seite 1
SeasonWinterHerbstSommerFrühling
MO
s lo
g [D
NA
Kop
ien/
kg T
S]
11.00
10.00
9.00
8.00
7.00
Fehlerbalken: 95% CI
Habitat: Land
Seite 1
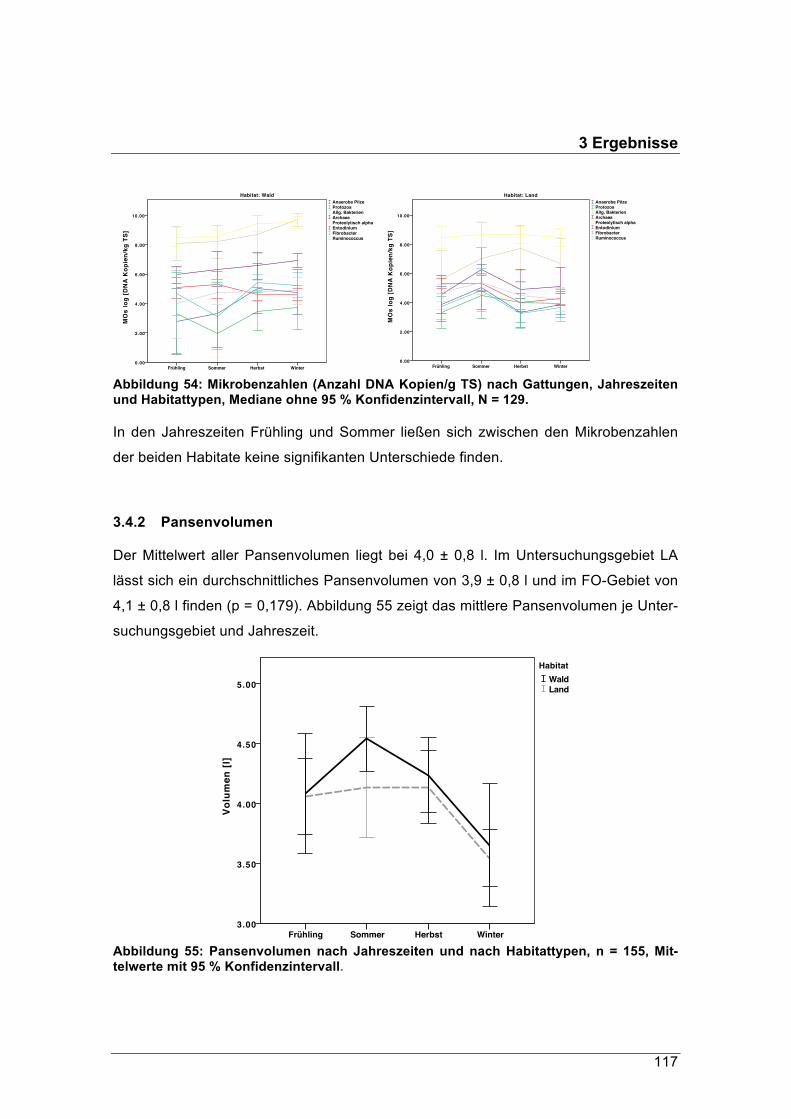
3 Ergebnisse
117
Abbildung 54: Mikrobenzahlen (Anzahl DNA Kopien/g TS) nach Gattungen, Jahreszeiten und Habitattypen, Mediane ohne 95 % Konfidenzintervall, N = 129.
In den Jahreszeiten Frühling und Sommer ließen sich zwischen den Mikrobenzahlen
der beiden Habitate keine signifikanten Unterschiede finden.
3.4.2 Pansenvolumen
Der Mittelwert aller Pansenvolumen liegt bei 4,0 ± 0,8 l. Im Untersuchungsgebiet LA
lässt sich ein durchschnittliches Pansenvolumen von 3,9 ± 0,8 l und im FO-Gebiet von
4,1 ± 0,8 l finden (p = 0,179). Abbildung 55 zeigt das mittlere Pansenvolumen je Unter-
suchungsgebiet und Jahreszeit.
Abbildung 55: Pansenvolumen nach Jahreszeiten und nach Habitattypen, n = 155, Mit-telwerte mit 95 % Konfidenzintervall.
WinterHerbstSommerFrühling
MO
s lo
g [D
NA
Kop
ien/
kg T
S]
10.00
8.00
6.00
4.00
2.00
0.00
RuminococcusFibrobacterEntodiniumProteolytisch alphaArchaeaAllg. BakterienProtozoaAnaerobe Pilze
Habitat: Wald
Seite 1
WinterHerbstSommerFrühling
MO
s lo
g [D
NA
Kop
ien/
kg T
S]
10.00
8.00
6.00
4.00
2.00
0.00
RuminococcusFibrobacterEntodinium Proteolytisch alphaArchaeaAllg. BakterienProtozoa Anaerobe Pilze
Habitat: Land
Seite 1
WinterHerbstSommerFrühling
Vol
umen
[l]
5 .00
4.50
4.00
3.50
3.00
LandWald
Habitat
Page 1

3 Ergebnisse
118
Auch wenn zwischen Land und Wald hinsichtlich der Jahreszeiten keine Signifikanzen
erkennbar sind, so lässt sich ein Trend hin zu höheren Werten im Wald erkennen. So
zeigen sich im Frühling Mittelwerte im Wald mit 4,1 ± 0,9 l und im Land mit 4,0 ± 1,0 l.
Im Sommer ergibt sich ein Mittelwert des Pansenvolumens von 4,5 ± 0,5 l im Wald und
im Land von 4,1 ± 0,9 l. Im Wald liegen die Werte im Herbst bei 4,2 ± 0,7 l und im Land
bei 4,1 ± 0,6 l. Im Winter sinken die Werte sowohl im Wald (3,7 ± 0,8 l) als auch im
Land (3,6 ± 0,5 l) ab.
Das Pansenvolumen von Rehen unterscheidet sich in den vier Jahreszeiten im Wald
signifikant zwischen Sommer und Winter (p = 0,004) sowie zwischen Herbst und Win-
ter (p = 0,048).
Im Gebiet Land zeigt sich das gleiche Bild, jedoch lässt sich hier noch ein zusätzlicher
signifikanter Unterschied feststellen: Die Werte im Frühling (p = 0,011), Sommer (p =
0,016) und Herbst (p = 0,003) sind signifikant höher als die Land-Winterwerte des Vo-
lumens.
3.4.3 Panseninhaltsmenge
Die mittlere Panseninhaltsmenge aller Pansenproben umfasst 1182 ± 393 g. Differen-
ziert nach Untersuchungsgebieten beträgt der durchschnittliche Panseninhalt 1315 ±
423 g im Wald und 1077 ± 334 g im Land. Der Unterschied ist signifikant (p < 0,000).
Während der Verlauf der Pansenfüllmengen im Landhabitat etwa dem Verlauf des
Pansenvolumens entspricht (Abbildung 56), weicht die Pansenfüllmenge der Rehe im
Waldhabitat deutlich davon ab. Obwohl das Pansenvolumen abnimmt, steigt die Nah-
rungsmenge im Pansen an.
Beim Vergleich der Jahreszeiten innerhalb eines Habitats zeigt sich ein signifikanter
Unterschied im Gebiet Land zwischen den Jahreszeiten Winter (x̄ = 1002 ± 300 g) und
Sommer (x̄ = 1285 ± 282 g) mit p = 0,011.
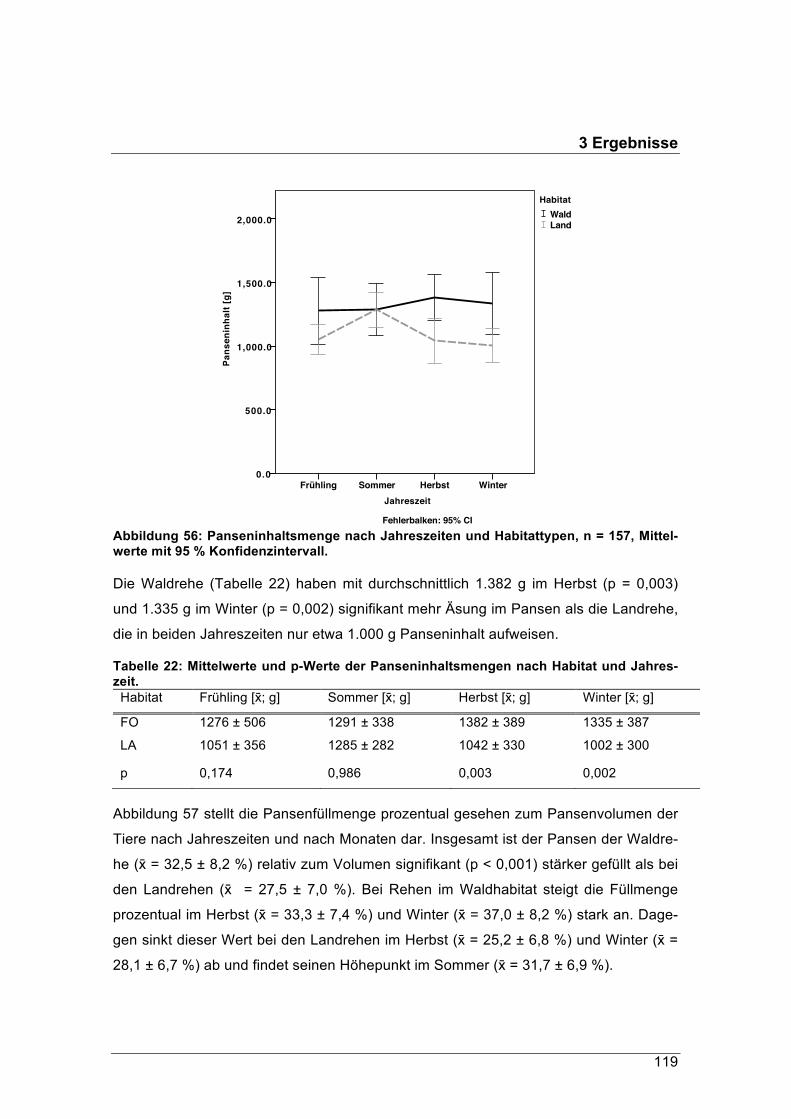
3 Ergebnisse
119
Abbildung 56: Panseninhaltsmenge nach Jahreszeiten und Habitattypen, n = 157, Mittel-werte mit 95 % Konfidenzintervall.
Die Waldrehe (Tabelle 22) haben mit durchschnittlich 1.382 g im Herbst (p = 0,003)
und 1.335 g im Winter (p = 0,002) signifikant mehr Äsung im Pansen als die Landrehe,
die in beiden Jahreszeiten nur etwa 1.000 g Panseninhalt aufweisen.
Tabelle 22: Mittelwerte und p-Werte der Panseninhaltsmengen nach Habitat und Jahres-zeit.
Habitat Frühling [x̄; g] Sommer [x̄; g] Herbst [x̄; g] Winter [x̄; g]
FO 1276 ± 506 1291 ± 338 1382 ± 389 1335 ± 387
LA 1051 ± 356 1285 ± 282 1042 ± 330 1002 ± 300
p 0,174 0,986 0,003 0,002
Abbildung 57 stellt die Pansenfüllmenge prozentual gesehen zum Pansenvolumen der
Tiere nach Jahreszeiten und nach Monaten dar. Insgesamt ist der Pansen der Waldre-
he (x̄ = 32,5 ± 8,2 %) relativ zum Volumen signifikant (p < 0,001) stärker gefüllt als bei
den Landrehen (x̄ = 27,5 ± 7,0 %). Bei Rehen im Waldhabitat steigt die Füllmenge
prozentual im Herbst (x̄ = 33,3 ± 7,4 %) und Winter (x̄ = 37,0 ± 8,2 %) stark an. Dage-
gen sinkt dieser Wert bei den Landrehen im Herbst (x̄ = 25,2 ± 6,8 %) und Winter (x̄ =
28,1 ± 6,7 %) ab und findet seinen Höhepunkt im Sommer (x̄ = 31,7 ± 6,9 %).
JahreszeitWinterHerbstSommerFrühling
Pans
enin
halt
[g]
2,000.0
1,500.0
1,000.0
500.0
0.0
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
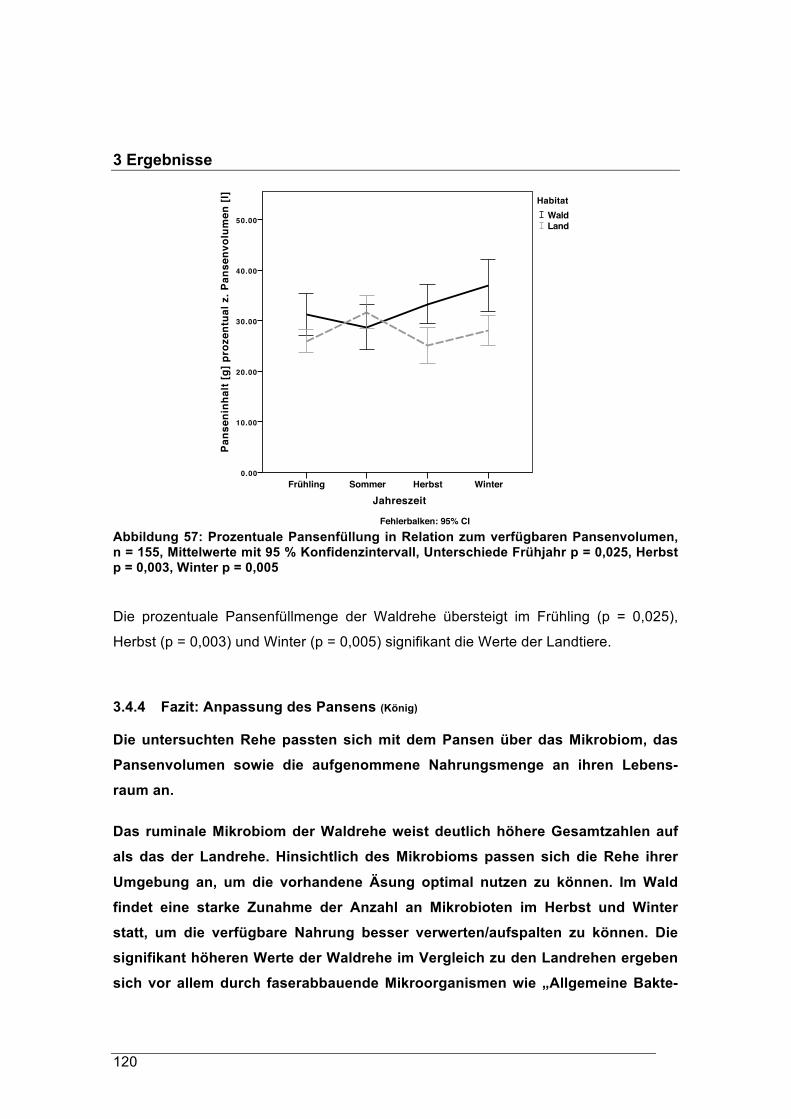
3 Ergebnisse
120
Abbildung 57: Prozentuale Pansenfüllung in Relation zum verfügbaren Pansenvolumen, n = 155, Mittelwerte mit 95 % Konfidenzintervall, Unterschiede Frühjahr p = 0,025, Herbst p = 0,003, Winter p = 0,005
Die prozentuale Pansenfüllmenge der Waldrehe übersteigt im Frühling (p = 0,025),
Herbst (p = 0,003) und Winter (p = 0,005) signifikant die Werte der Landtiere.
3.4.4 Fazit: Anpassung des Pansens (König)
Die untersuchten Rehe passten sich mit dem Pansen über das Mikrobiom, das
Pansenvolumen sowie die aufgenommene Nahrungsmenge an ihren Lebens-
raum an.
Das ruminale Mikrobiom der Waldrehe weist deutlich höhere Gesamtzahlen auf
als das der Landrehe. Hinsichtlich des Mikrobioms passen sich die Rehe ihrer
Umgebung an, um die vorhandene Äsung optimal nutzen zu können. Im Wald
findet eine starke Zunahme der Anzahl an Mikrobioten im Herbst und Winter
statt, um die verfügbare Nahrung besser verwerten/aufspalten zu können. Die
signifikant höheren Werte der Waldrehe im Vergleich zu den Landrehen ergeben
sich vor allem durch faserabbauende Mikroorganismen wie „Allgemeine Bakte-
JahreszeitWinterHerbstSommerFrühling
Pans
enin
halt
[g] p
roze
ntua
l z. P
anse
nvol
umen
[l]
50.00
40.00
30.00
20.00
10.00
0.00
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1

3 Ergebnisse
121
rien“ (Faserverwerter = Zellulose, Hemizellulose, KH und Stärke), „Fibrobactern“
(Zelluloseverwerter), „Archaea“ (Essigsäureverwerter).
Waldrehe haben ein größeres Pansenvolumen über das Jahr gesehen als die
Landrehe. Der Jahresverlauf des Pansenvolumens bei Wald- und Landrehe zeigt
einen ähnlichen Trend. Jedoch erreicht der Pansen im Wald sein größtes Volu-
men im Sommer, wohingegen der Pansen der Landrehe sein größtes Volumen im
Herbst erreicht. Bei allen Rehen zeigt sich das niedrigste Pansenvolumen im
Winter.
Die Füllmenge des Pansens der Waldrehe ist über das Jahr gesehen sowie diffe-
renziert nach Jahreszeiten höher als bei den Landrehen. Waldrehe passen sich
an den niedrigeren Energiegehalt ihres Habitats durch mehr Nahrungsaufnahme
an. Im Durchschnitt der subadulten und adulten Rehe war der Pansen im Wald
mit 1.315 g und mit 1.077 g im Land gefüllt. Die Landrehe zeigen die höchste rela-
tive Füllmenge im Sommer. Bei Waldrehen ist dagegen der Pansen absolut und
relativ zu seinem Volumen im Winter am stärksten gefüllt.
3.5 Kondition der Untersuchungstiere (Scheingraber)
Zur Feststellung der Kondition der Untersuchungstiere wurde der Nierenfettindex nach
Riney 1955 (n = 222) wie auch das Gewicht der aufgebrochenen Tiere (n = 245) erho-
ben und ausgewertet.
3.5.1 Gewicht der aufgebrochenen Rehe
Das durchschnittliche Körpergewicht, aufgebrochen und unmittelbar danach gewogen,
aller Untersuchungstiere mit Kitzen liegt bei 13,8 ± 2,8 kg und ohne Kitze bei 15,2 ± 1,9
kg (Tabelle 23). Das Gewicht der Landrehe liegt mit Kitzen im Durchschnitt 800 g über
jenem der Waldrehe und ohne Kitze etwa 600 g höher.
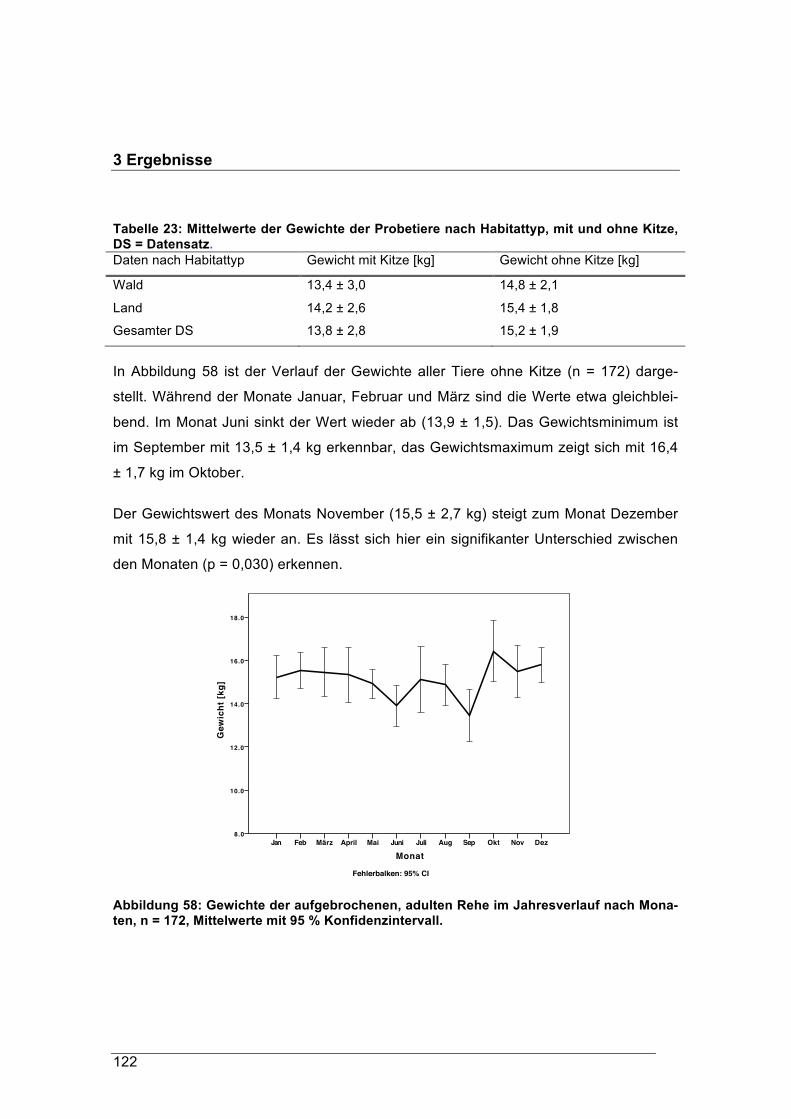
3 Ergebnisse
122
Tabelle 23: Mittelwerte der Gewichte der Probetiere nach Habitattyp, mit und ohne Kitze, DS = Datensatz. Daten nach Habitattyp Gewicht mit Kitze [kg] Gewicht ohne Kitze [kg]
Wald 13,4 ± 3,0 14,8 ± 2,1
Land 14,2 ± 2,6 15,4 ± 1,8
Gesamter DS 13,8 ± 2,8 15,2 ± 1,9
In Abbildung 58 ist der Verlauf der Gewichte aller Tiere ohne Kitze (n = 172) darge-
stellt. Während der Monate Januar, Februar und März sind die Werte etwa gleichblei-
bend. Im Monat Juni sinkt der Wert wieder ab (13,9 ± 1,5). Das Gewichtsminimum ist
im September mit 13,5 ± 1,4 kg erkennbar, das Gewichtsmaximum zeigt sich mit 16,4
± 1,7 kg im Oktober.
Der Gewichtswert des Monats November (15,5 ± 2,7 kg) steigt zum Monat Dezember
mit 15,8 ± 1,4 kg wieder an. Es lässt sich hier ein signifikanter Unterschied zwischen
den Monaten (p = 0,030) erkennen.
Abbildung 58: Gewichte der aufgebrochenen, adulten Rehe im Jahresverlauf nach Mona-ten, n = 172, Mittelwerte mit 95 % Konfidenzintervall.
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Gew
icht
[kg]
18.0
16.0
14.0
12.0
10.0
8.0
Fehlerbalken: 95% CI
Page 1
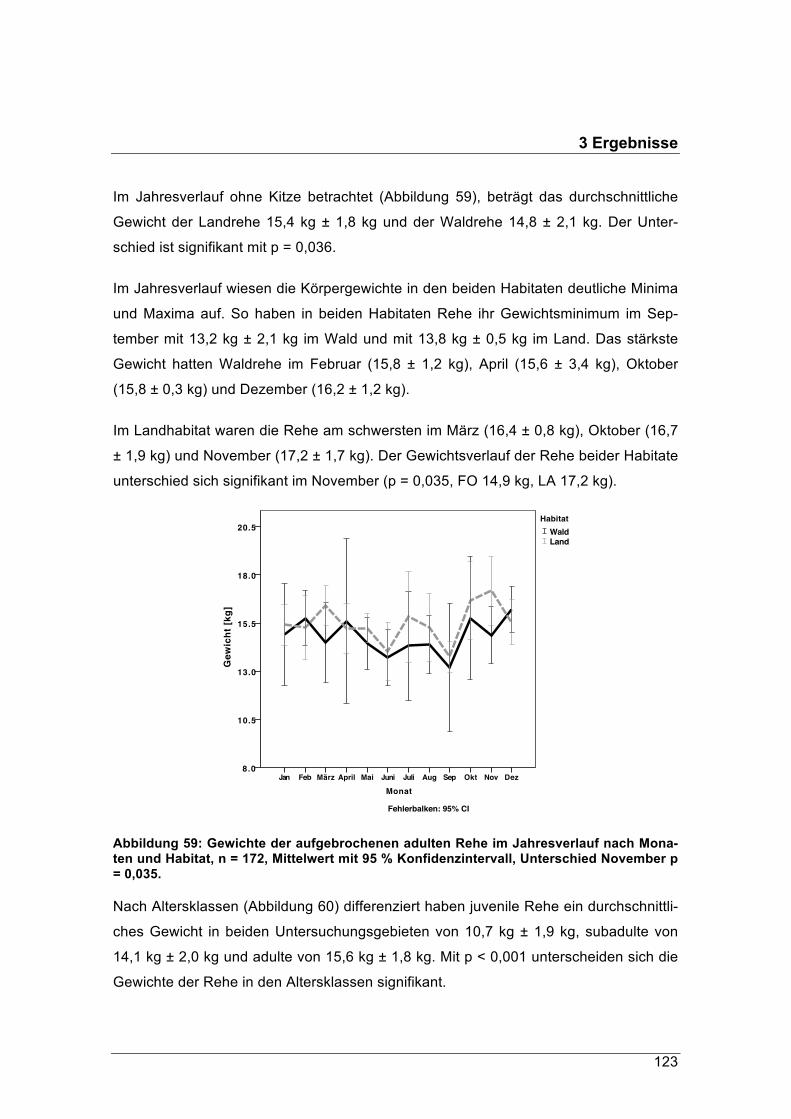
3 Ergebnisse
123
Im Jahresverlauf ohne Kitze betrachtet (Abbildung 59), beträgt das durchschnittliche
Gewicht der Landrehe 15,4 kg ± 1,8 kg und der Waldrehe 14,8 ± 2,1 kg. Der Unter-
schied ist signifikant mit p = 0,036.
Im Jahresverlauf wiesen die Körpergewichte in den beiden Habitaten deutliche Minima
und Maxima auf. So haben in beiden Habitaten Rehe ihr Gewichtsminimum im Sep-
tember mit 13,2 kg ± 2,1 kg im Wald und mit 13,8 kg ± 0,5 kg im Land. Das stärkste
Gewicht hatten Waldrehe im Februar (15,8 ± 1,2 kg), April (15,6 ± 3,4 kg), Oktober
(15,8 ± 0,3 kg) und Dezember (16,2 ± 1,2 kg).
Im Landhabitat waren die Rehe am schwersten im März (16,4 ± 0,8 kg), Oktober (16,7
± 1,9 kg) und November (17,2 ± 1,7 kg). Der Gewichtsverlauf der Rehe beider Habitate
unterschied sich signifikant im November (p = 0,035, FO 14,9 kg, LA 17,2 kg).
Abbildung 59: Gewichte der aufgebrochenen adulten Rehe im Jahresverlauf nach Mona-ten und Habitat, n = 172, Mittelwert mit 95 % Konfidenzintervall, Unterschied November p = 0,035.
Nach Altersklassen (Abbildung 60) differenziert haben juvenile Rehe ein durchschnittli-
ches Gewicht in beiden Untersuchungsgebieten von 10,7 kg ± 1,9 kg, subadulte von
14,1 kg ± 2,0 kg und adulte von 15,6 kg ± 1,8 kg. Mit p < 0,001 unterscheiden sich die
Gewichte der Rehe in den Altersklassen signifikant.
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Gew
icht
[kg]
20.5
18.0
15.5
13.0
10.5
8.0
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
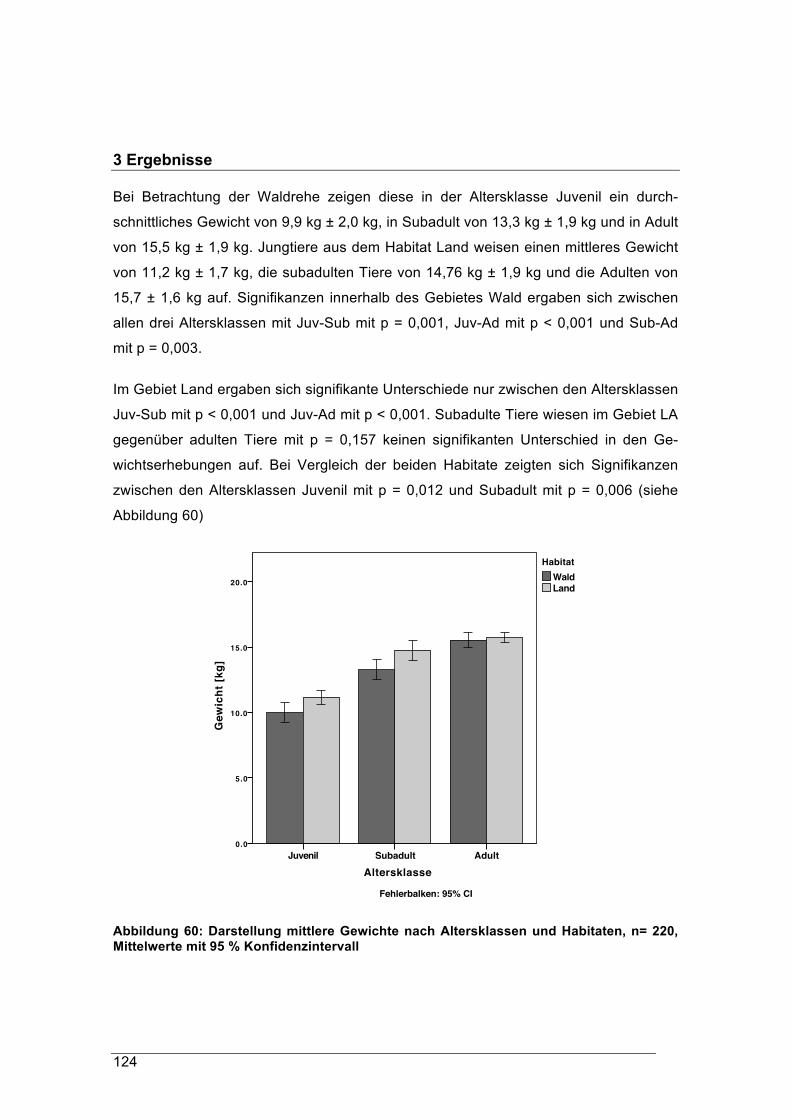
3 Ergebnisse
124
Bei Betrachtung der Waldrehe zeigen diese in der Altersklasse Juvenil ein durch-
schnittliches Gewicht von 9,9 kg ± 2,0 kg, in Subadult von 13,3 kg ± 1,9 kg und in Adult
von 15,5 kg ± 1,9 kg. Jungtiere aus dem Habitat Land weisen einen mittleres Gewicht
von 11,2 kg ± 1,7 kg, die subadulten Tiere von 14,76 kg ± 1,9 kg und die Adulten von
15,7 ± 1,6 kg auf. Signifikanzen innerhalb des Gebietes Wald ergaben sich zwischen
allen drei Altersklassen mit Juv-Sub mit p = 0,001, Juv-Ad mit p < 0,001 und Sub-Ad
mit p = 0,003.
Im Gebiet Land ergaben sich signifikante Unterschiede nur zwischen den Altersklassen
Juv-Sub mit p < 0,001 und Juv-Ad mit p < 0,001. Subadulte Tiere wiesen im Gebiet LA
gegenüber adulten Tiere mit p = 0,157 keinen signifikanten Unterschied in den Ge-
wichtserhebungen auf. Bei Vergleich der beiden Habitate zeigten sich Signifikanzen
zwischen den Altersklassen Juvenil mit p = 0,012 und Subadult mit p = 0,006 (siehe
Abbildung 60)
Abbildung 60: Darstellung mittlere Gewichte nach Altersklassen und Habitaten, n= 220, Mittelwerte mit 95 % Konfidenzintervall
AltersklasseAdultSubadultJuvenil
Gew
icht
[kg]
20.0
15.0
10.0
5.0
0.0
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
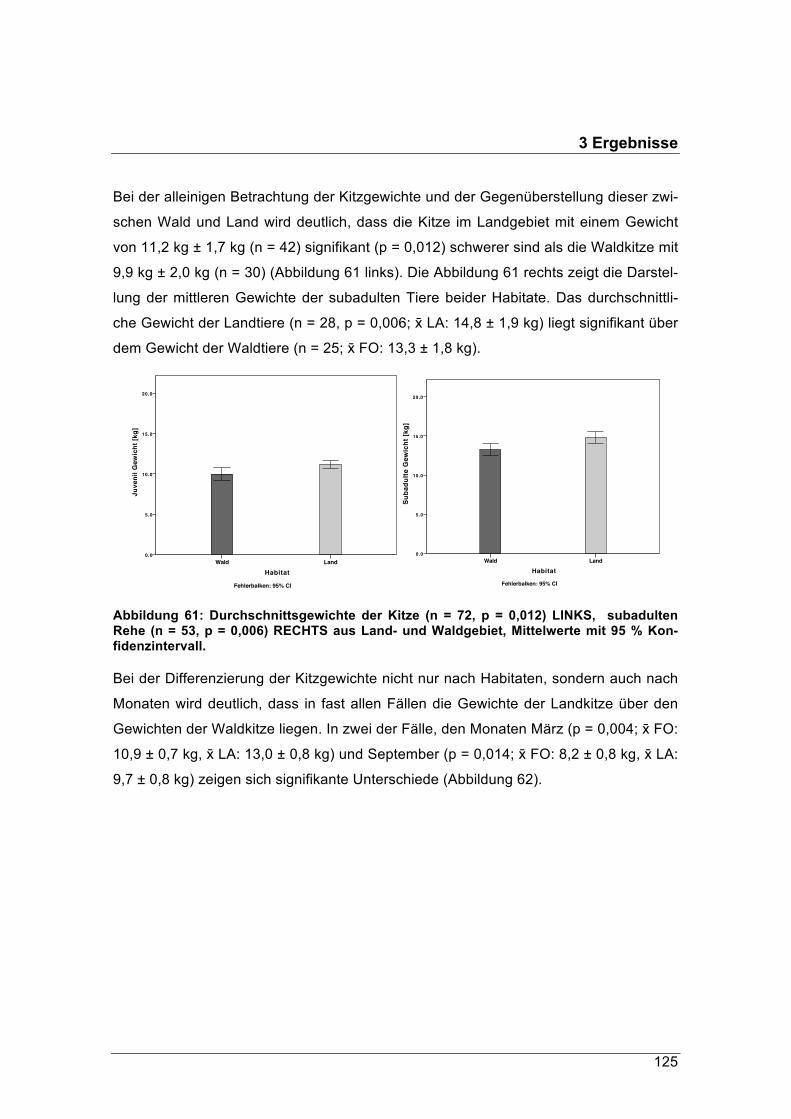
3 Ergebnisse
125
Bei der alleinigen Betrachtung der Kitzgewichte und der Gegenüberstellung dieser zwi-
schen Wald und Land wird deutlich, dass die Kitze im Landgebiet mit einem Gewicht
von 11,2 kg ± 1,7 kg (n = 42) signifikant (p = 0,012) schwerer sind als die Waldkitze mit
9,9 kg ± 2,0 kg (n = 30) (Abbildung 61 links). Die Abbildung 61 rechts zeigt die Darstel-
lung der mittleren Gewichte der subadulten Tiere beider Habitate. Das durchschnittli-
che Gewicht der Landtiere (n = 28, p = 0,006; x̄ LA: 14,8 ± 1,9 kg) liegt signifikant über
dem Gewicht der Waldtiere (n = 25; x̄ FO: 13,3 ± 1,8 kg).
Abbildung 61: Durchschnittsgewichte der Kitze (n = 72, p = 0,012) LINKS, subadulten Rehe (n = 53, p = 0,006) RECHTS aus Land- und Waldgebiet, Mittelwerte mit 95 % Kon-fidenzintervall.
Bei der Differenzierung der Kitzgewichte nicht nur nach Habitaten, sondern auch nach
Monaten wird deutlich, dass in fast allen Fällen die Gewichte der Landkitze über den
Gewichten der Waldkitze liegen. In zwei der Fälle, den Monaten März (p = 0,004; x̄ FO:
10,9 ± 0,7 kg, x̄ LA: 13,0 ± 0,8 kg) und September (p = 0,014; x̄ FO: 8,2 ± 0,8 kg, x̄ LA:
9,7 ± 0,8 kg) zeigen sich signifikante Unterschiede (Abbildung 62).
HabitatLandWald
Juve
nil G
ewic
ht [k
g]
20.0
15.0
10.0
5.0
0.0
Fehlerbalken: 95% CI
Page 1
HabitatLandWald
Suba
dulte
Gew
icht
[kg]
20.0
15.0
10.0
5.0
0.0
Fehlerbalken: 95% CI
Page 1
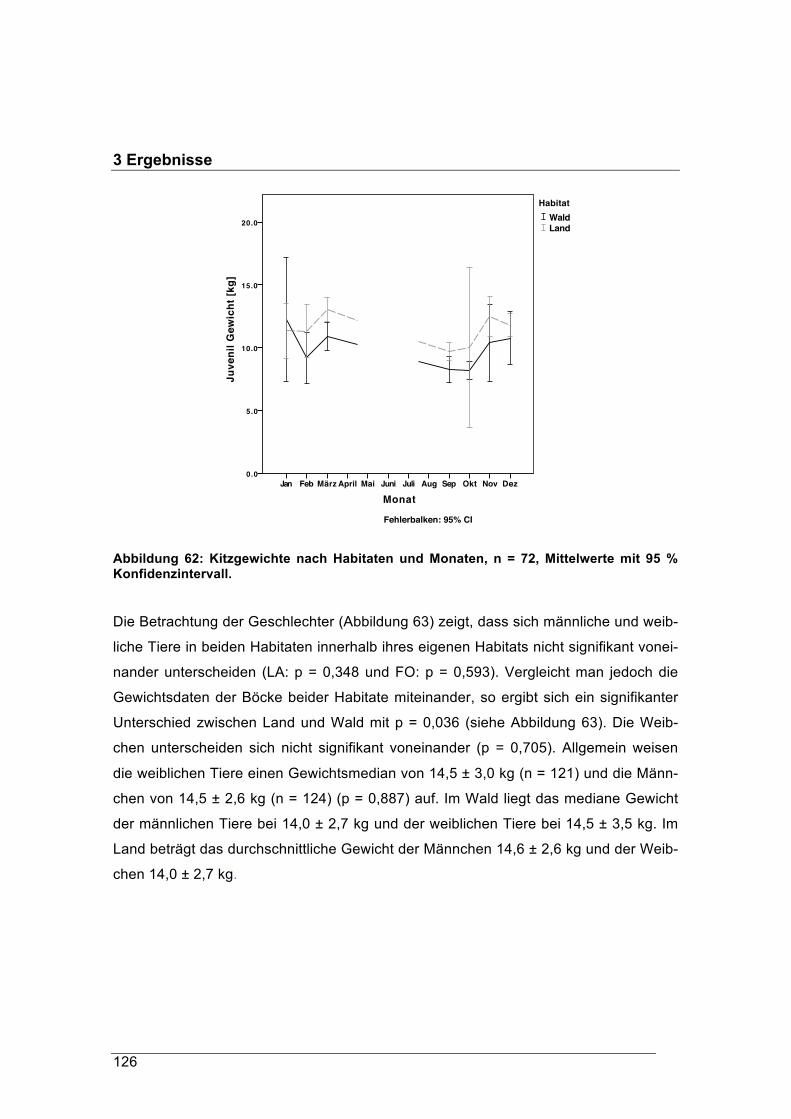
3 Ergebnisse
126
Abbildung 62: Kitzgewichte nach Habitaten und Monaten, n = 72, Mittelwerte mit 95 % Konfidenzintervall.
Die Betrachtung der Geschlechter (Abbildung 63) zeigt, dass sich männliche und weib-
liche Tiere in beiden Habitaten innerhalb ihres eigenen Habitats nicht signifikant vonei-
nander unterscheiden (LA: p = 0,348 und FO: p = 0,593). Vergleicht man jedoch die
Gewichtsdaten der Böcke beider Habitate miteinander, so ergibt sich ein signifikanter
Unterschied zwischen Land und Wald mit p = 0,036 (siehe Abbildung 63). Die Weib-
chen unterscheiden sich nicht signifikant voneinander (p = 0,705). Allgemein weisen
die weiblichen Tiere einen Gewichtsmedian von 14,5 ± 3,0 kg (n = 121) und die Männ-
chen von 14,5 ± 2,6 kg (n = 124) (p = 0,887) auf. Im Wald liegt das mediane Gewicht
der männlichen Tiere bei 14,0 ± 2,7 kg und der weiblichen Tiere bei 14,5 ± 3,5 kg. Im
Land beträgt das durchschnittliche Gewicht der Männchen 14,6 ± 2,6 kg und der Weib-
chen 14,0 ± 2,7 kg.
MonatDezNovOktSepAugJuliJuniMaiAprilMärzFebJan
Juve
nil G
ewic
ht [k
g]20.0
15.0
10.0
5.0
0.0
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
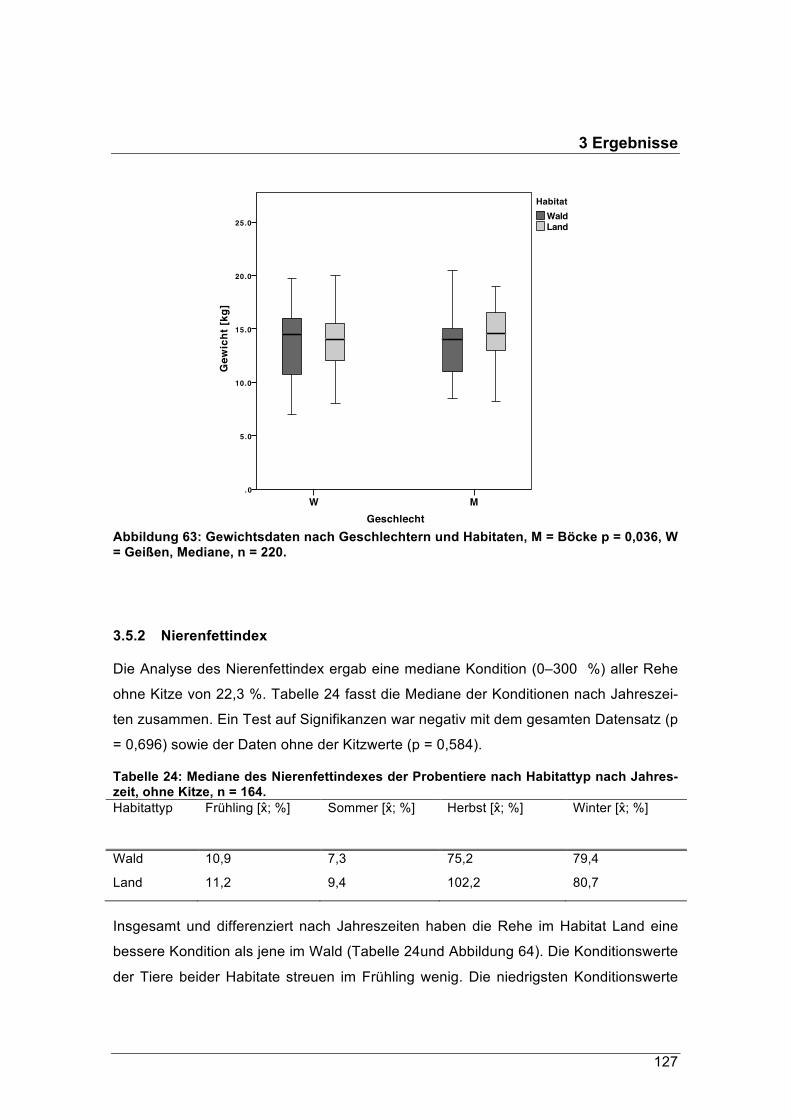
3 Ergebnisse
127
Abbildung 63: Gewichtsdaten nach Geschlechtern und Habitaten, M = Böcke p = 0,036, W = Geißen, Mediane, n = 220.
3.5.2 Nierenfettindex
Die Analyse des Nierenfettindex ergab eine mediane Kondition (0–300 %) aller Rehe
ohne Kitze von 22,3 %. Tabelle 24 fasst die Mediane der Konditionen nach Jahreszei-
ten zusammen. Ein Test auf Signifikanzen war negativ mit dem gesamten Datensatz (p
= 0,696) sowie der Daten ohne der Kitzwerte (p = 0,584).
Tabelle 24: Mediane des Nierenfettindexes der Probentiere nach Habitattyp nach Jahres-zeit, ohne Kitze, n = 164. Habitattyp Frühling [x̂; %] Sommer [x̂; %] Herbst [x̂; %] Winter [x̂; %]
Wald 10,9 7,3 75,2 79,4
Land 11,2 9,4 102,2 80,7
Insgesamt und differenziert nach Jahreszeiten haben die Rehe im Habitat Land eine
bessere Kondition als jene im Wald (Tabelle 24und Abbildung 64). Die Konditionswerte
der Tiere beider Habitate streuen im Frühling wenig. Die niedrigsten Konditionswerte
GeschlechtMW
Gew
icht
[kg]
25.0
20.0
15.0
10.0
5.0
.0
LandWald
Habitat
Page 1
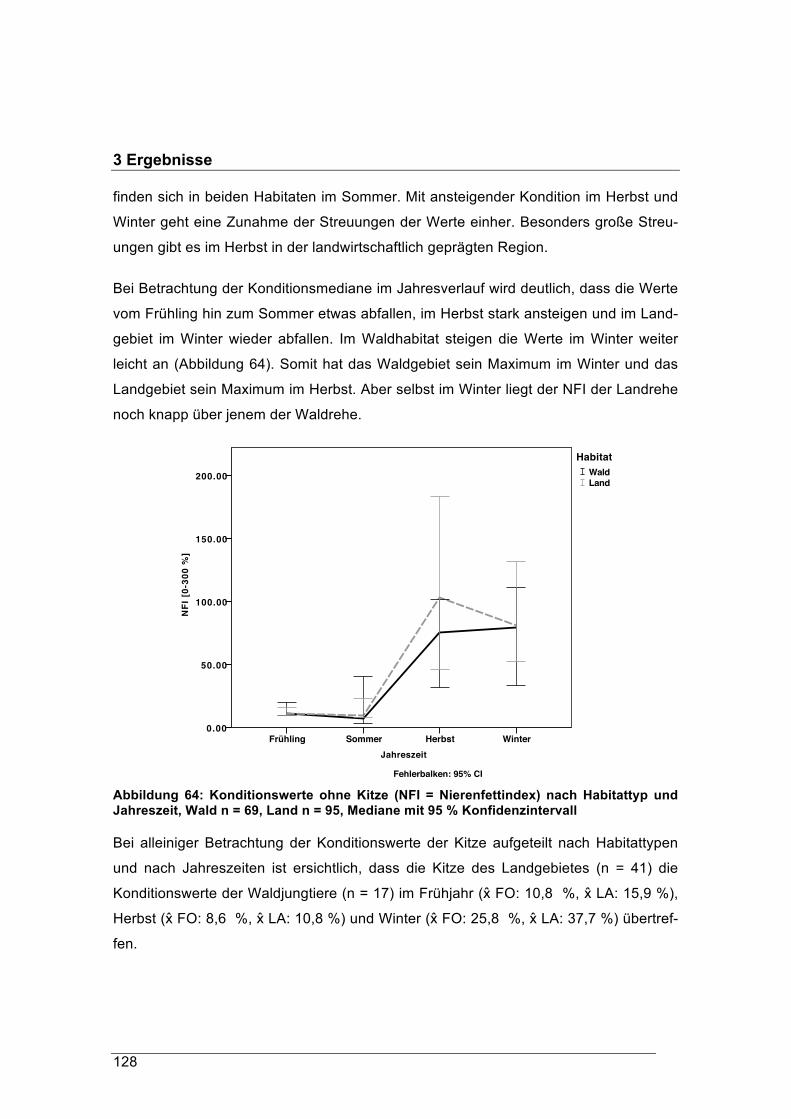
3 Ergebnisse
128
finden sich in beiden Habitaten im Sommer. Mit ansteigender Kondition im Herbst und
Winter geht eine Zunahme der Streuungen der Werte einher. Besonders große Streu-
ungen gibt es im Herbst in der landwirtschaftlich geprägten Region.
Bei Betrachtung der Konditionsmediane im Jahresverlauf wird deutlich, dass die Werte
vom Frühling hin zum Sommer etwas abfallen, im Herbst stark ansteigen und im Land-
gebiet im Winter wieder abfallen. Im Waldhabitat steigen die Werte im Winter weiter
leicht an (Abbildung 64). Somit hat das Waldgebiet sein Maximum im Winter und das
Landgebiet sein Maximum im Herbst. Aber selbst im Winter liegt der NFI der Landrehe
noch knapp über jenem der Waldrehe.
Abbildung 64: Konditionswerte ohne Kitze (NFI = Nierenfettindex) nach Habitattyp und Jahreszeit, Wald n = 69, Land n = 95, Mediane mit 95 % Konfidenzintervall
Bei alleiniger Betrachtung der Konditionswerte der Kitze aufgeteilt nach Habitattypen
und nach Jahreszeiten ist ersichtlich, dass die Kitze des Landgebietes (n = 41) die
Konditionswerte der Waldjungtiere (n = 17) im Frühjahr (x̂ FO: 10,8 %, x̂ LA: 15,9 %),
Herbst (x̂ FO: 8,6 %, x̂ LA: 10,8 %) und Winter (x̂ FO: 25,8 %, x̂ LA: 37,7 %) übertref-
fen.
JahreszeitWinterHerbstSommerFrühling
NFI
[0-
300
%]
200.00
150.00
100.00
50.00
0.00
Fehlerbalken: 95% CI
LandWald
Habitat
Page 1
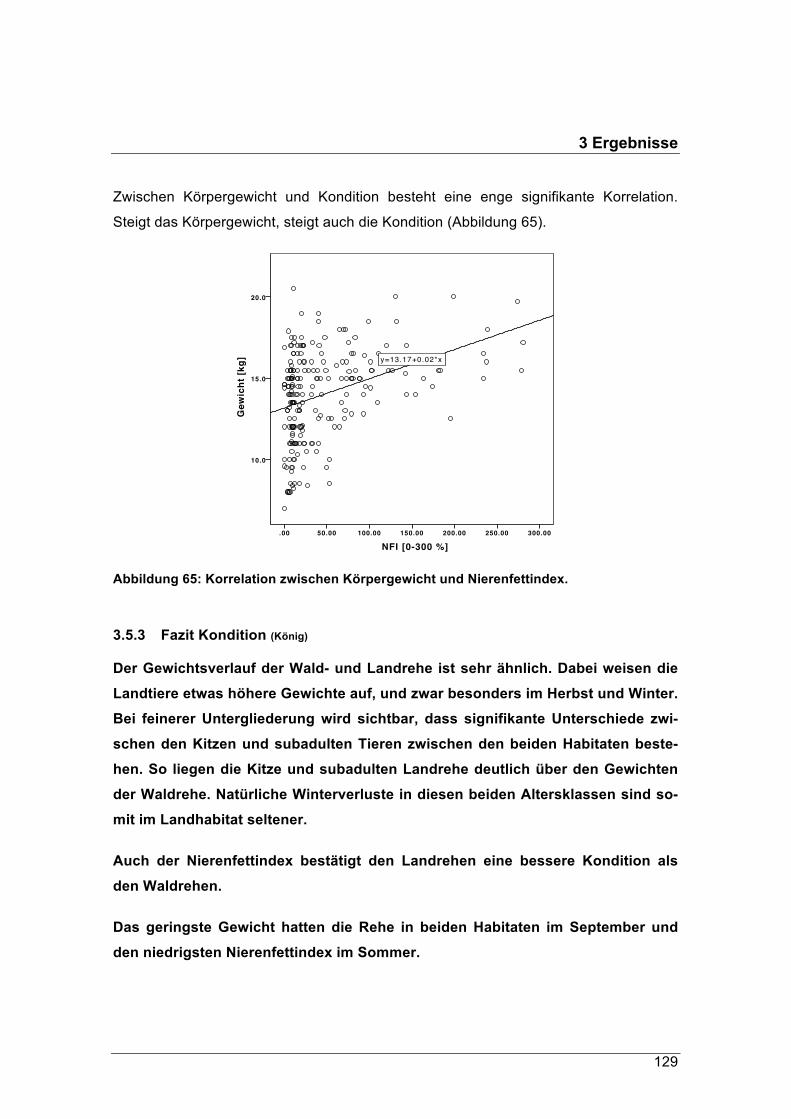
3 Ergebnisse
129
Zwischen Körpergewicht und Kondition besteht eine enge signifikante Korrelation.
Steigt das Körpergewicht, steigt auch die Kondition (Abbildung 65).
Abbildung 65: Korrelation zwischen Körpergewicht und Nierenfettindex.
3.5.3 Fazit Kondition (König)
Der Gewichtsverlauf der Wald- und Landrehe ist sehr ähnlich. Dabei weisen die
Landtiere etwas höhere Gewichte auf, und zwar besonders im Herbst und Winter.
Bei feinerer Untergliederung wird sichtbar, dass signifikante Unterschiede zwi-
schen den Kitzen und subadulten Tieren zwischen den beiden Habitaten beste-
hen. So liegen die Kitze und subadulten Landrehe deutlich über den Gewichten
der Waldrehe. Natürliche Winterverluste in diesen beiden Altersklassen sind so-
mit im Landhabitat seltener.
Auch der Nierenfettindex bestätigt den Landrehen eine bessere Kondition als
den Waldrehen.
Das geringste Gewicht hatten die Rehe in beiden Habitaten im September und
den niedrigsten Nierenfettindex im Sommer.
NFI [0-300 %]300.00250.00200.00150.00100.0050.00.00
Gew
icht
[kg]
20.0
15.0
10.0
y=13.17+0.02*x
R2 Linear = 0.138
Seite 1
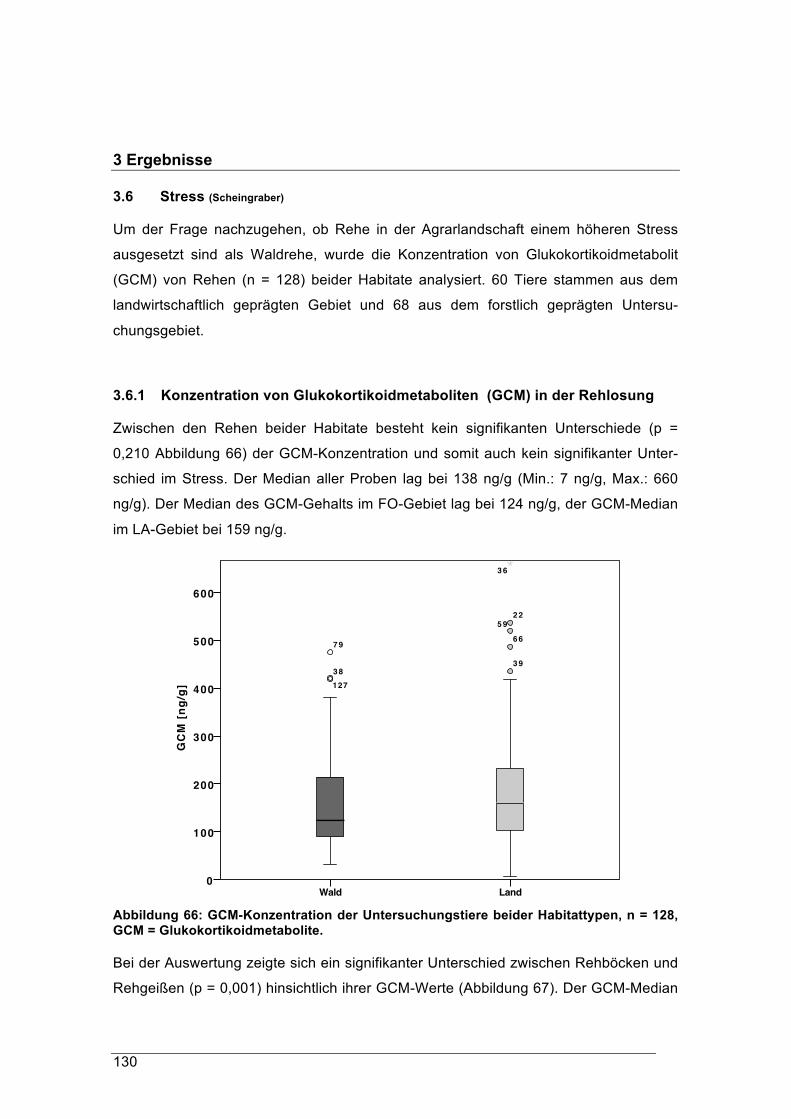
3 Ergebnisse
130
3.6 Stress (Scheingraber)
Um der Frage nachzugehen, ob Rehe in der Agrarlandschaft einem höheren Stress
ausgesetzt sind als Waldrehe, wurde die Konzentration von Glukokortikoidmetabolit
(GCM) von Rehen (n = 128) beider Habitate analysiert. 60 Tiere stammen aus dem
landwirtschaftlich geprägten Gebiet und 68 aus dem forstlich geprägten Untersu-
chungsgebiet.
3.6.1 Konzentration von Glukokortikoidmetaboliten (GCM) in der Rehlosung
Zwischen den Rehen beider Habitate besteht kein signifikanten Unterschiede (p =
0,210 Abbildung 66) der GCM-Konzentration und somit auch kein signifikanter Unter-
schied im Stress. Der Median aller Proben lag bei 138 ng/g (Min.: 7 ng/g, Max.: 660
ng/g). Der Median des GCM-Gehalts im FO-Gebiet lag bei 124 ng/g, der GCM-Median
im LA-Gebiet bei 159 ng/g.
Abbildung 66: GCM-Konzentration der Untersuchungstiere beider Habitattypen, n = 128, GCM = Glukokortikoidmetabolite.
Bei der Auswertung zeigte sich ein signifikanter Unterschied zwischen Rehböcken und
Rehgeißen (p = 0,001) hinsichtlich ihrer GCM-Werte (Abbildung 67). Der GCM-Median
LandWald
GC
M [
ng/g
]
600
500
400
300
200
100
0
3 6
6 6
5 9
3 9
2 2
127
7 9
3 8
Page 1
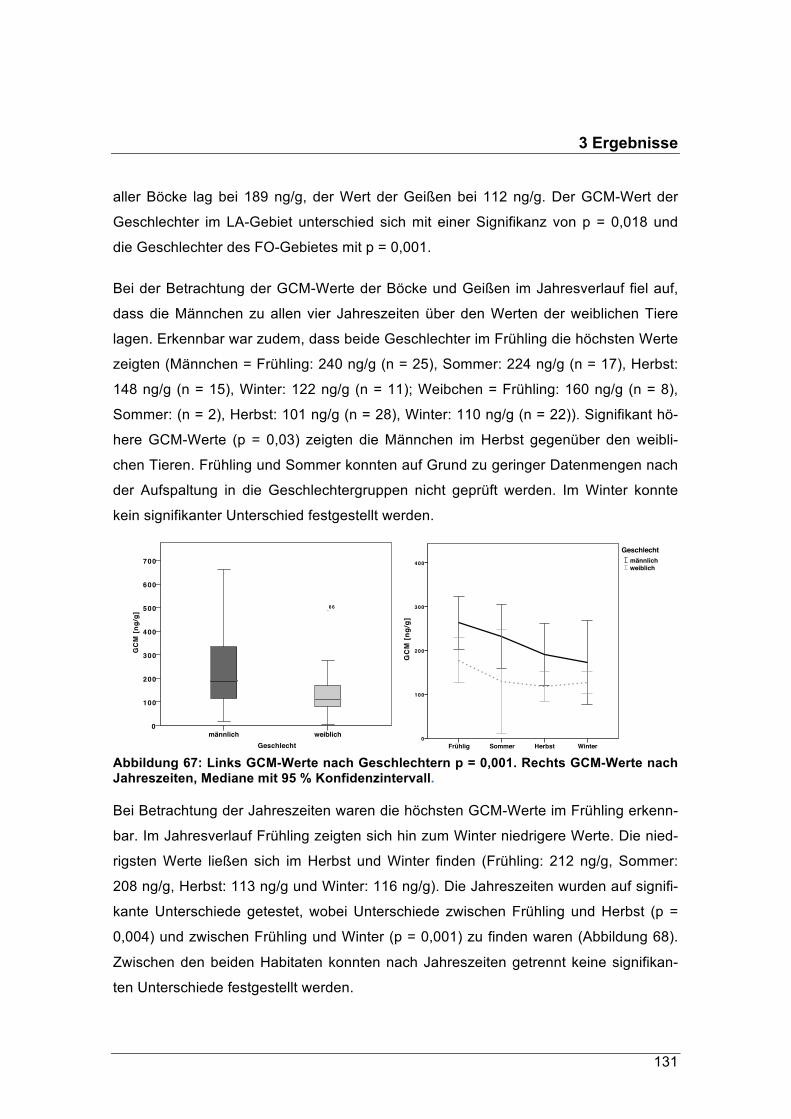
3 Ergebnisse
131
aller Böcke lag bei 189 ng/g, der Wert der Geißen bei 112 ng/g. Der GCM-Wert der
Geschlechter im LA-Gebiet unterschied sich mit einer Signifikanz von p = 0,018 und
die Geschlechter des FO-Gebietes mit p = 0,001.
Bei der Betrachtung der GCM-Werte der Böcke und Geißen im Jahresverlauf fiel auf,
dass die Männchen zu allen vier Jahreszeiten über den Werten der weiblichen Tiere
lagen. Erkennbar war zudem, dass beide Geschlechter im Frühling die höchsten Werte
zeigten (Männchen = Frühling: 240 ng/g (n = 25), Sommer: 224 ng/g (n = 17), Herbst:
148 ng/g (n = 15), Winter: 122 ng/g (n = 11); Weibchen = Frühling: 160 ng/g (n = 8),
Sommer: (n = 2), Herbst: 101 ng/g (n = 28), Winter: 110 ng/g (n = 22)). Signifikant hö-
here GCM-Werte (p = 0,03) zeigten die Männchen im Herbst gegenüber den weibli-
chen Tieren. Frühling und Sommer konnten auf Grund zu geringer Datenmengen nach
der Aufspaltung in die Geschlechtergruppen nicht geprüft werden. Im Winter konnte
kein signifikanter Unterschied festgestellt werden.
Abbildung 67: Links GCM-Werte nach Geschlechtern p = 0,001. Rechts GCM-Werte nach Jahreszeiten, Mediane mit 95 % Konfidenzintervall.
Bei Betrachtung der Jahreszeiten waren die höchsten GCM-Werte im Frühling erkenn-
bar. Im Jahresverlauf Frühling zeigten sich hin zum Winter niedrigere Werte. Die nied-
rigsten Werte ließen sich im Herbst und Winter finden (Frühling: 212 ng/g, Sommer:
208 ng/g, Herbst: 113 ng/g und Winter: 116 ng/g). Die Jahreszeiten wurden auf signifi-
kante Unterschiede getestet, wobei Unterschiede zwischen Frühling und Herbst (p =
0,004) und zwischen Frühling und Winter (p = 0,001) zu finden waren (Abbildung 68).
Zwischen den beiden Habitaten konnten nach Jahreszeiten getrennt keine signifikan-
ten Unterschiede festgestellt werden.
Geschlechtweiblichmännlich
GC
M [
ng/g
]
700
600
500
400
300
200
100
0
6 6
Page 1
WinterHerbstSommerFrühlig
GC
M [
ng/g
]
400
300
200
100
0
weiblichmännlich
Geschlecht
Page 1
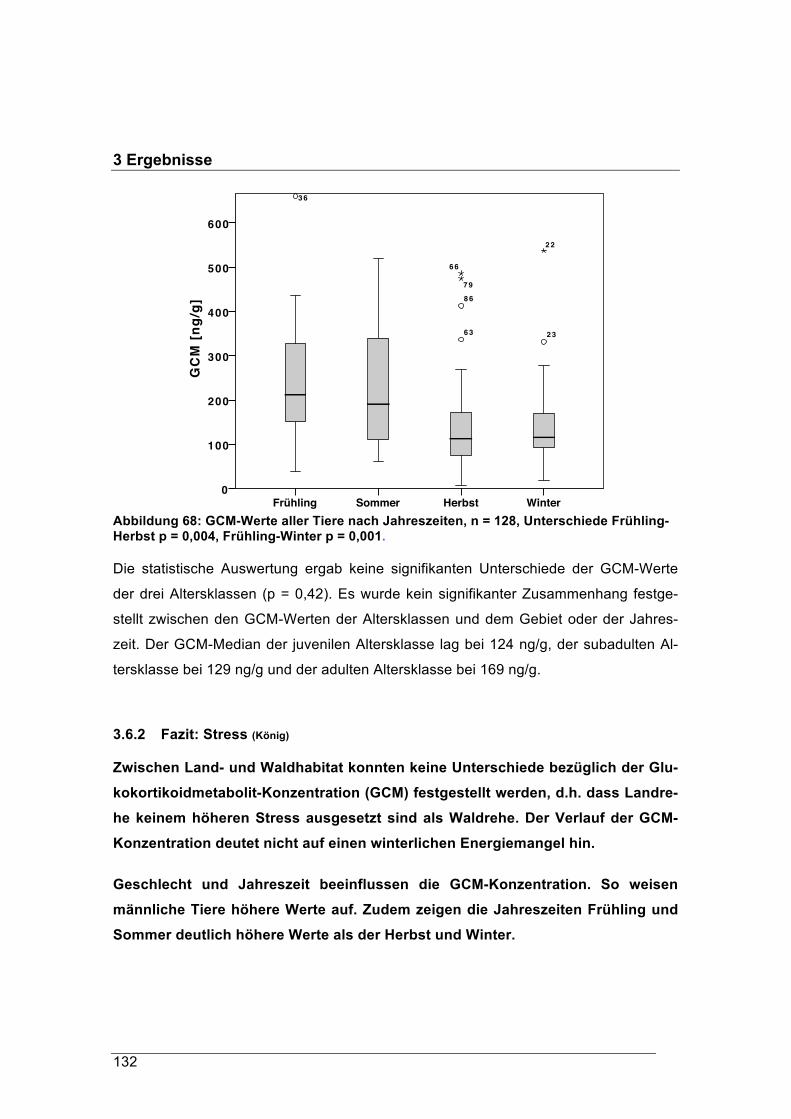
3 Ergebnisse
132
Abbildung 68: GCM-Werte aller Tiere nach Jahreszeiten, n = 128, Unterschiede Frühling-Herbst p = 0,004, Frühling-Winter p = 0,001.
Die statistische Auswertung ergab keine signifikanten Unterschiede der GCM-Werte
der drei Altersklassen (p = 0,42). Es wurde kein signifikanter Zusammenhang festge-
stellt zwischen den GCM-Werten der Altersklassen und dem Gebiet oder der Jahres-
zeit. Der GCM-Median der juvenilen Altersklasse lag bei 124 ng/g, der subadulten Al-
tersklasse bei 129 ng/g und der adulten Altersklasse bei 169 ng/g.
3.6.2 Fazit: Stress (König)
Zwischen Land- und Waldhabitat konnten keine Unterschiede bezüglich der Glu-
kokortikoidmetabolit-Konzentration (GCM) festgestellt werden, d.h. dass Landre-
he keinem höheren Stress ausgesetzt sind als Waldrehe. Der Verlauf der GCM-
Konzentration deutet nicht auf einen winterlichen Energiemangel hin.
Geschlecht und Jahreszeit beeinflussen die GCM-Konzentration. So weisen
männliche Tiere höhere Werte auf. Zudem zeigen die Jahreszeiten Frühling und
Sommer deutlich höhere Werte als der Herbst und Winter.
WinterHerbstSommerFrühling
GC
M [
ng/g
]
600
500
400
300
200
100
0
2 2
2 3
7 9
6 6
8 6
6 3
3 6
Page 1

4 Diskussion
133
4 Diskussion (König / Scheingraber)
Ziel der vorliegenden Studie war die ganzjährige Erfassung der Energie und Qualität
der Rehnahrung in einem Wald- und Agrarhabitat. Großes Augenmerk lag auf der
Energie und Qualität der Rehnahrung außerhalb der regulären Jagdzeiten, da diese
Zeiten besonders wichtig für das Überleben von Wildtieren im Winter sind und einen
entscheidenden Einfluss auf die Zuwachsraten von Populationen haben. Im Sinne ei-
nes wildbiologischen Systemansatzes wurden Parameter zum Habitat sowie der Ein-
fluss von menschlichen Störungen in Form des Stressniveaus der Tiere in die Betrach-
tungen mit einbezogen.
4.1 Probennahme
Die Erlegung fand im Rahmen des behördlich festgelegten Abschussplanes statt. Für
die Erlegung von Rehen außerhalb der regulären Jagdzeiten lagen Schonzeitaufhe-
bungen der entsprechenden Landratsämter vor. Insgesamt wurden von 245 Rehen
Proben genommen. Somit liegt den Ergebnissen dieser Untersuchung eine wesentlich
höhere Stichprobenzahl und damit Aussagekraft zugrunde als allen anderen zuvor
durchgeführten vergleichbaren Studien wie z.B. von Brüggemann et al. (1967), Dre-
scher-Kaden (1976), Schmidl (1996), Djordjevic (2006), Reimoser (2006), Popovic
(2009), die mit einem Viertel bis Hälfte an Proben arbeiteten.
Von den 245 Rehen wurden 139 Tieren im landwirtschaftlich geprägten Habitat, 106 im
forstlich geprägten Habitat erlegt. 25 Proben wurden bei der Auswertung nicht berück-
sichtig, da sie Kirr- bzw. Futtermaterial beinhalteten. Geplant war, aus jedem Monat
pro Habitat 5 verwertbare Proben zu bekommen, weshalb 8 Proben pro Monat ange-
peilt wurden. Dieses Ziel wurde insgesamt weit überschritten, die Sichtbarkeit der Re-
he und der damit verbundene Jagderfolg folgen den von Ellenberg (1976) beschriebe-
nen Aktivitätszyklen des Rehwildes. Dieser Aktivitätszyklus zeigt deutliche Aktivitäts-
einbußen in den Monaten Juni bis August und im Oktober. In diesen Monaten gelang
es nur die angestrebte Mindestzahl von 5 bis 8 Untersuchungstieren pro Habitat und

4 Diskussion
134
Monat zu erlegen. Die Zahl an Untersuchungstieren ist ausreichend für statistisch ab-
gesicherte Aussagen.
In den Monaten mit hoher Aktivitätsrate (September, November, Dezember, Januar
und Mai) ist ein Datenüberschuss vorhanden (Tabelle 3).
Das Geschlecht betreffend wurde im Waldgebiet ein ausgeglichenes Geschlechterver-
hältnis bei den adulten Tieren erreicht. Im Landgebiet wurden vermehrt männliche Tie-
re geschossen. Berücksichtigt man neben dem Geschlecht auch die Altersklasse,
wurden bei den subadulten und juvenilen Rehen im Land mehr weibliche Tiere und im
Wald mehr männliche Tiere erlegt. Wird der Datensatz als Ganzes betrachtet, ist das
Geschlechterverhältnis mit ♂ 1: ♀ 0,96 in etwa ausgewogen.
Um von Anfang an eine konstant hohe Qualität der Probengewinnung zu garantieren,
wurde 3 Monate vor dem Beginn des Projektes eine Vorstudie zur optimalen Proben-
nahme begonnen. Neben der Probennahme wurde auch die bestmöglichste Proben-
aufbereitung und Probenanalyse getestet und erprobt (Schmid 2013, Schneider 2012).
Dieses Vorgehen vermied den Verlust von Proben durch unsachgemäße Behandlung
und hat insgesamt zu einer Qualitätssteigerung bei der Probennahme bis hin zur Ana-
lysemethode erbracht.
Die Laboranalysen wurden entsprechend den neuesten internationalen wissen-
schaftlichen Standards mit Doppel- oder Vierfachbestimmungen ausgeführt.
4.2 Die wichtigsten Ergebnisse
Die Studie wurde vor dem Hintergrund einer Vielzahl von Veröffentlichungen und For-
schungsarbeiten entwickelt. Anhand dieser Literatur wurden folgende Ergebnisse er-
wartet:
1. Als Konzentrat-Selektierer kann das Reh aus den landwirtschaftlichen Kultur-
pflanzen nur wenig Energie gewinnen (Hofmann & Kirsten 1982, Bauer 2007).

4 Diskussion
135
2. Niedrige Energieausstattung der Rehe im März / April, d.h. wenig Energie in der
Äsung sowie abgebaute Reserven (Ellenberg 1978, Hofmann & Kirsten 1982,
Oslage & Strohtmann 1988, Arnold 2004, Sommer 2004, Reimoser 2006).
3. Rohfasergehalte in der Nahrung, die deutlich unter dem Niveau der Äsungsty-
pen „Intermediär“ und „Raufutter“ liegen, wie sie z.B. bei Rothirsch und Mufflon
zu finden sind. (Drescher-Kaden & Seifelnasr 1976, Hofmann 1982, Müller
1982, Drescher-Kaden 1984, Hofmann 1989).
4. Im Agrarhabitat steht im Herbst für die notwendige Feistbildung keine energe-
tisch ausreichende Äsung zur Verfügung (Hofmann & Kirsten 1982, Hofmann
1982, Bauer 2007, Helm 2015). Es ist zu erwarten, dass das Energieniveau in
diesem Zeitraum sinkt und vermehrt Waldvegetation geäst wird.
5. Das größte Volumen besitzt der Pansen im Herbst und das niedrigste im Winter
und Frühjahr, parallel hierzu wird der Panseninhalt verringert (Hofmann 1978 +
1981, Hofmann & Kirsten 1982). Das Volumen stellt eine Kapazitätsgrenze für
die aufnehmbare Äsung dar.
6. Landrehe besitzen auf Grund von Störungen ein höheres Stressniveau als
Waldrehe (Dehnhard et al. 2001, Arlettaz et al. 2007, Thiel et al. 2008, Herzog
2013, Rehnus et al. 2014). Im März / April steigt das Stressniveau der Rehe auf
Grund des Abbaus von Fettreserven an (Huber et al. 2003; Dalmau et al.
2007).
Im Gegensatz zu den Thesen unter Punkt 1 fanden gerade die Rehe im Agrarhabitat
signifikant mehr Energie in der Äsung als im Waldhabitat (Kap. 3.1.1 und Abbildung
16). Niedrige Energiewerte fanden sich im Übergang Winter-Frühjahr nur im Waldhabi-
tat im Februar, einen Monat früher als erwartet, während gleichzeitig die Waldrehe ein
hohes Körpergewicht (Abbildung 59) sowie hohe Fettreserven (Abbildung 64) hatten
(These 2). Auch Hoffmann (1978) fand die niedrigsten Darmfett-Reserven im August
und nicht im Februar. Im Herbst stand den Landrehen nicht nur mehr Energie in ihrer
Agraräsung als den Rehen im naturnahen Waldhabitat zur Verfügung, sondern es wa-
ren auch die höchsten Werte im Jahresverlauf. Gleichzeitig stiegen Körpergewicht und

4 Diskussion
136
Nierenfettindex an, so dass in diesem Zeitraum Reserven für den Winter bei den Land-
rehen angelegt werden konnten (These 4). Das Gleiche gilt für Waldrehe, wenn auch
auf einem etwas niedrigeren Niveau. Im März und April wurden in beiden Habitaten
bereits relativ hohe Energiewerte in der Äsung gefunden. Landrehe haben zwischen
Februar und März sogar an Gewicht zugelegt. In den drei Projektjahren konnte in bei-
den Untersuchungsgebieten auf Grund der Gewichts- und Feistbildung sowie der ver-
fügbaren Energie aus der Äsung in den kritischen Monaten März und April kein Eng-
pass bei Rehen festgestellt werden (These 2).
In unseren Untersuchungsgebieten zeigten alle Panseninhalte relativ hohe Faserantei-
le, die nicht unter 23 % Trockensubstanz sanken (These 3). Diese hohen Faseranteile
sind vor allem von Intermediären Äsungstypen und Raufutterfressern bekannt (Dre-
scher-Kaden & Seifelnasr 1976, Hofmann 1982 + 1989, Müller 1982, Drescher-Kaden
1984, Stubbe 1988). Allerdings zeigten auch schon andere Studien, dass Rehwild gut
mit faserreicher Äsung umgehen kann (Drescher-Kaden & Seifelnasr 1976, Anke et al.
2007, Clauss 2010), nur wurde dies von vielen nicht zur Kenntnis genommen. Auch
der Hohenheimer Futterwerttest mit dem Rehpansensaft zeigte, dass Rehe optimal an
faserhaltige Äsung im Winter angepasst sind und aus dieser mehr Energie ziehen kön-
nen als ein an Fasern angepasster Raufutterfresser. Eine bessere Ausnutzung von
faserhaltiger Äsung konnten auch Arnold et al. (2015) in ihren Fütterungsversuchen bei
Rotwild nachweisen.
Auf Grund dieser Fähigkeiten und der häufig falschen Interpretation des Begrif-
fes „Konzentrat-Selektierer“ sollte auf diesen verzichtet werden und wie im eng-
lischen Sprachraum und international wissenschaftlich üblich besser „Selektie-
rer“ verwendet werden. Futtermittel für Rehe sollten im Winter mindestens 20 %
TS Fasergehalt aufweisen, besser wäre ein Gehalt von nicht unter 25 % TS. Feh-
lende Fasern in Futtermitteln holen sich die Rehe aus der Waldvegetation.
Bei den Landrehen wurde erwartet (These 4), dass sie während und nach der Ernte
weniger Energie im Pansen haben und vermehrt Waldvegetation äsen. Gerade im
Herbst haben Landrehe aber ihre höchsten Energiewerte im Pansen (Abbildung 17).
Gerade in den Monaten September bis November fanden sich in den Pansen der Lan-

4 Diskussion
137
drehe besonders viele Feldfrüchte, Kräuter und Süßgräser, während Waldvegetation
eine deutlich untergeordnete Rolle spielte (Tabelle 11).
Im Gegensatz zur bekannten Literatur (These 5) haben Waldrehe im Sommer ein Ma-
ximum an Pansenvolumen und senken im Herbst und Winter, trotz abnehmenden
Pansenvolumens, nicht die aufgenommene Äsungsmenge. Intensive Nahrungsauf-
nahme im Winter beobachtet Hofmann (1981) auch bei Rehen an Fütterungen und in
Gattern, verneint dies aber bei Rehen in freier Wildbahn. Waldrehe können geringere
Energie der vorhandenen Äsung durch ein Mehr an Nahrungsaufnahme ausgleichen.
Anders als erwartet (These 6) fand sich kein Unterschied zwischen dem Stressniveau
der Waldrehe und der Landrehe. Weder bei den Waldrehen noch bei den Landrehen
stieg die GCM-Konzentration im Spätwinter bis Frühjahr durch Katabolismus an.
4.3 Energie der Rehnahrung
Die Analyse des Energiegehaltes der aufgenommenen Äsung bildet das Kernstück der
Untersuchung. Zur Energiemessung und Berechnung wurde der Hohenheimer Futter-
werttest (HFT) herangezogen, der den aktuellen wissenschaftlichen Stand in der Fut-
termittelanalyse darstellt (Kirchgessner et al. 2008).
Die Entscheidung, für die metabolische Energieberechnung nicht das Weender Verfah-
ren und eine anschließende Regressionsgleichung zu verwenden, sondern ein In-Vitro
Verfahren anzuschließen, basierte auf Ergebnissen früherer Studien und der Idee einer
wissenschaftlichen Weiterentwicklung dieser Verfahren.
Frühere Studien (Papageorgiou & Neophytou 1981, Drozdz & Osiecki 1973, Drozdz
1979, Baumgärtl 2012) errechneten allein mit den Angaben der Rohnährstoffe von
Nahrungspflanzen oder Futtermitteln die metabolisch verfügbare Energiemenge für
Rehwild. Dissen & Hartfiel (1985) nahmen sich des Problems der rehwildspezifischen
Verdauung an. Sie ermittelten für zwei verschiedene Futterkomponenten (Alleinfutter,
Wildsilage) für alle vier Jahreszeiten Verdauungsquotienten für das Rehwild. Sie erar-
beiteten ihre Verdauungskoeffizienten an gefütterten, im Gehege gehaltenen Rehen.
Da in dieser Studie die tatsächlich aufgenommene Äsung verwendet und analysiert

4 Diskussion
138
wurde, erwarteten wir durch den Hohenheimer Futterwerttest bessere und genauere
Werte für die tatsächlich von Rehen aufgenommene Äsung.
Zur Kontrolle wurden ergänzend die Verfahren nach Dissen & Hartfiel (1985) verwen-
det. Es zeigte sich, dass mit dem Hohenheimer Futterwerttest (HFT) deutlichere Unter-
schiede zwischen den Jahreszeiten und Habitaten gefunden wurden als mit dem Ver-
fahren von Dissen & Hartfiel (1985). Das Verfahren nach Dissen & Hartfiel (1985) be-
rechnet höhere, aber von Jahreszeit zu Jahreszeit weniger differenzierte Werte. Zieht
man zusätzlich die Veränderung der Qualität der Rehnahrung im Jahresverlauf in Be-
tracht, sind die mit dem HFT gefundenen Werte plausibler.
Der Hohenheimer Futterwerttest (HFT) arbeitet mit einem standardisierten Hammel-
pansensaft, das Probenmaterial ist aber die real zu jeder Jahreszeit aufgenommene
Äsung und nicht die für diese Jahreszeit theoretisch angenommene Äsung von Rehen.
Um etwaige Unterschiede durch die Verwendung von Hammelpansensaft gegenüber
dem natürlichen Rehpansensaft zu ermitteln, wurden 18 Proben aus dem Zeitraum
November bis Januar zusätzlich mit Rehpansensaft aus dem Januar ausgewertet
(Abbildung 18). Zwischen den mit Hammelpansensaft und Rehpansensaft gewonne-
nen Ergebnissen besteht eine signifikante starke Korrelation (p < 0,001; r = 0,6). Hier-
bei zeigten sich im Januar signifikante höherer Gasbildungswerte beim Rehpansensaft
im Vergleich zum Hammelpansensaft.
Das Reh konnte als „Konzentrat-Selektier“ besser die raufaserhaltige Januaräsung
energetisch ausnutzen als das Haustier „Schaf“ als Raufutterfresser. Auch Rehbinder
& Ciszuk (1985) und Cederlund (2006) fanden in ihren In-Vitro-Verdauungsstudien,
dass Rehe speziell im Winter besser mit raufaserreicher Äsung umgehen können als
Schafe oder Elche. Arnold et al. (2015) untermauert dies mit Ergebnissen an Rotwild,
in denen sie zeigen konnten, dass die relative Ausbeute an Rohproteinen im Winter
aus der Nahrung deutlich gesteigert war. Sie führen das auf die optimale Anpassung
von Wildwiederkäuern an den Winter zurück.
Eine Anpassung des Rehes an die Äsungsqualität in den verschiedenen Jahreszeiten
fanden neben Rehbinder & Ciszuk (1985) und Cederlund (2006) auch Holand (1993)
und Chen (2006).

4 Diskussion
139
Bei der Analyse von Panseninhalt zur Bestimmung der Energiewerte muss bei der In-
terpretation beachtet werden, dass die realen Energiewerte der aufgenommenen Nah-
rung unterschätzt werden. Die gefundenen Werte stellen Mindestwerte dar.
• Der Inhalt des Rehpansens beinhaltet aufgrund der regelmäßigen Äsungszyk-
len immer ein Gemisch an frischem und bereits vor- und verdautem Nahrungs-
material. Das vorverdaute Pflanzenmaterial hat bereits Energie abgegeben, die
Energiewerte sind niedriger als bei frisch geästen Nahrungspflanzen.
• Es reichern sich vermehrt schwerlösliche/-verdauliche Bestandteile (Fasern) im
Pansen an (Norris 1943, Klötzli 1965, Esser, 1954, Mcinnis 1983, Meyer 1988),
welche beim In-Vitro-Versuch (HFT) geringe Gasbildungswerte und somit nied-
rigere Energiewerte liefern (Giger-Reverdin 1998, Hummel et al. 2006).
• Durch die Tätigkeit des Mikrobioms befindet sich der Nahrungsbrei unter konti-
nuierlicher Verdauung und ein Teil der Nahrungsenergie ist im Pansensaft ge-
löst oder absorbiert. Der Pansensaft wurde bei der Aufbereitung abzentrifugiert
und gesondert analysiert. Sein Energiegehalt geht nicht in die Energieberech-
nung des HFT ein.
• Sehr leichtverdauliche und energiereiche Pflanzensäfte der Äsung können Re-
he direkt über die Pansenrinne am Pansen vorbei in den Blätter- und Lab-
magen schleusen (Rowell-Schäfer et al 2000, Ditchkoff 2000, Djordjevic 2006)
und verdauen. Diese Form der Energieaufnahme kann nicht durch Analysen
des Panseninhaltes erfasst werden.
• Die Analysen liefern den Energiewert der Nahrung als eine Momentaufnahme.
In Wirklichkeit liegt die aufgenommene Energiemenge pro Tag deutlich höher,
da Rehe aufgrund ihres kleinen Pansens (Hofmann 1976) und der damit ver-
bundenen kurzen Retentionszeit (Behrend 1999) gezwungen sind, sehr häufig
und regelmäßig, je nach Jahreszeit 6–14 mal pro Tag, Nahrung aufzunehmen
(Berg 1978, Stubbe 1997). Somit nehmen Rehe fortwährend neues, frisches
Pflanzenmaterial zu sich, welches durch eine Momentaufnahme nicht miterfasst
werden kann.

4 Diskussion
140
Das von uns gewählte Verfahren eignet sich besser für die Nahrungsanalyse wildle-
bender Rehe mit natürlicher Nahrung, die dem saisonalen Wechsel und anderen bioti-
schen/abiotischen Einflüssen unterliegt. So wird in der klassischen Panseninhaltsana-
lyse nicht jede wirklich aufgenommene Pflanze erfasst (Klansek 2004). Künstlich zu-
sammengestellte Äsungspflanzensortimente können nur grobe Näherungswerte der
Quantität und Zahl der Arten liefern. Dies wird deutlich beim Vergleich verschiedener
Studien (Oslage & Strothmann 1988, Dissen & Hartfiel 1985), in denen mit künstlich
selektiertem frischen Pflanzenmaterial oder Futtermitteln gearbeitet wurde. Allerdings
ist ein direkter Vergleich der hier vorgestellten Ergebnisse mit anderen Studien, die mit
abweichenden Methoden gearbeitet haben, nur teilweise möglich.
Der metabolische Energiegehalt der aufgenommenen Nahrung wurde nach Habitat
und Jahresverlauf unterschieden. Es ergeben sich große Unterschiede zwischen den
Energiewerten der Nahrung von Land- und Waldtieren. Der Grundenergiewert der
Waldtiere liegt bei 5,4 MJ/kg TS, der Wert der Landtiere bei 6,3 MJ/kg TS. In sieben
der zwölf Monate liegt der metabolische Energiegehalt der Landrehe signifikant über
den Werten der Waldtiere. Die Nahrung der Landrehe ist um einiges energiereicher.
Im Hinblick auf den Jahresverlauf stellt sich im Waldgebiet der Höhepunkt der Ener-
giewerte im Mai und August sowie das Minimum im Februar ein (Abbildung 17). Dieser
Verlauf ist identisch mit dem Jahresverlauf der Rohproteine im Waldgebiet (Abbildung
21, links), weshalb naheliegt, dass ein wesentlicher Teil der Energie im Wald durch
Rohproteine in der Äsung geliefert wird. Im Landgebiet erreicht das Energieniveau sei-
nen Höhepunkt in den Monaten August, Oktober und November. Im August und Okto-
ber fanden sich bei den Landrehen die höchsten Werte an Stickstoff-freien-
Extraktstoffen (Kohlenhydrate). Im landwirtschaftlich geprägten Gebiet werden die gro-
ßen Energiemaxima vor allem durch hohe Kohlenhydratgehalte in der Nahrung er-
reicht. Die im Pansen gefundene Äsung der Landrehe ist nicht nur energiereiche-
re als jene im Wald, sondern die Äsung unterscheidet sich auch in Hinsicht ihrer
Energiequellen.
Die unterschiedlichen Nahrungsquellen bestätigt auch die Panseninhaltsanalyse. Das
Energiemaximum der Waldtiere befindet sich im Frühling. Zu dieser Zeit nehmen die

4 Diskussion
141
Waldrehe sehr hohe Anteile an Kräutern (Kap. 3.3.1.1) zu sich, die für höhere Protein-
anteile bekannt sind (Esser 1954).
Das Energiemaximum der Landrehe im Sommer und Herbst lässt sich durch die Auf-
nahme großer Mengen an Feld- und Baumfrüchten erklären (Kap. 3.3.1.1), die vor al-
lem Kohlehydrate (Zuckern, Stärken usw.) sowie Rohproteine aufweisen (Anke et al.
2007). Das Vorkommen im Agraruntersuchungsgebiet von relativ vielen masttragenden
Baumarten wie Buche und Eiche ist typisch für bäuerliche Waldbesitzstrukturen.
Im Frühjahr steigt der Energiegehalt in der Äsung mit der Vegetationsentwicklung an.
Dieser Anstieg der in der Vegetation enthaltenen Energie ist im Landhabitat be-
reits im April und im Waldhabitat erst im Mai anzutreffen. Das Phänomen lässt sich
mit Hilfe der pflanzensoziologischen Aufnahmen erklären (Kap.3.3). Die Vegetation auf
freien, sonnenexponierten, landwirtschaftlichen Flächen befindet sich in einem frühe-
ren und rascheren Wachstum als die Vegetation des Waldes. In Wäldern herrscht ein
waldtypisches Mikroklima vor, welches widerstandsfähiger gegenüber Schwankungen,
aber auch gegenüber raschen Erwärmungen ist. Somit erreichen Wiesen und Felder
mit Wintersaat ihren vegetativen Höhepunkt deutlich rascher (Schmidt 2004). In dem
durch Landwirtschaft geprägten Gebiet ist zudem der Einfluss der Jahreszeiten abge-
schwächt. Die Energiewerte schwanken dort wesentlich geringer im Jahresverlauf als
sie es im forstlich dominierten Gebiet tun. Diese Ergebnisse werden durch Arbeiten
Schmidt (2004) und Zweifel-Schielly (2005) bestätigt, wonach in Wäldern je nach Jah-
reszeit das Äsungsangebot schwankt, jedoch Wiesen, Felder und landwirtschaftliche
Flächen diese saisonalen Schwankungen stark nivellieren.
Energetisch und qualitativ wird der Winter von vielen Autoren als Engpass für die Er-
nährung von Wildwiederkäuern gesehen (Sommer 2004, Clauss 2005, Arnoldt 2013).
Dieser Engpass konnte in dieser Studie im Land-Untersuchungsgebiet nicht be-
stätigt werden. Der Gehalt an Proteinen und Stickstofffreien-Extraktstoffen ist hier in
diesen Monaten nicht im Minimum. Der Energiegehalt liegt im Winter im Land zwi-
schen 5,85 MJ/kg TS und 6,30 MJ/kg TS sowie im Durchschnitt bei 6,02 MJ/kg TS. Im
Vergleich zu einem Jahresdurchschnitt von 6,29 MJ/kg TS im Land und einem Jahres-

4 Diskussion
142
durchschnitt im naturnahen Waldhabitat von 5,43 MJ/kg TS bedeutet dies keine Ver-
schlechterung der Ernährungssituation im Land im Winter!
Zur Beurteilung des Energiestatus wurde die Studie von Weiner (1977) herangezogen.
Weiner (1977) untersuchte bei zahmen, in Gehegen gehaltenen Rehen durch respira-
torische Stoffwechselversuche den Energiebedarf im Dezember bis Februar und Mai
bis August. Die in Weiners (1977) Studie angegebenen Energiebedarfswerte
[kcal/kg0,75/Tag] wurden zum Vergleich mit unseren Werten in [MJ/20 kg Reh0,75/ Tag]
umgerechnet. Er arbeitet zudem mit dem gemessenen Bruttoenergiebedarf, wir dage-
gen mit der umsetzbaren metabolischen Energie von bereits vorverdautem Pansenma-
terial, die unter dem gesamten Energiebedarf liegt (vgl. Abbildung 11). Trotzdem un-
terscheiden sich Weiners Bruttoenergiebedarfswerte und die metabolischen Energie-
werte des Panseninhaltes im Jahresverlauf nur geringfügig. Im Fall des Waldgebietes
liegen Weiners Werte (5,7–7,3 MJ/20 kg Reh0,75/ Tag) auf gleichem Niveau mit den von
uns gemessenen metabolischen Mindestenergiewerten der Waldrehe (4,7–6,4 MJ/kg
TS) und leicht unter den gemessenen Mindestenergiemengen im Pansen der Landre-
he (5,8–7,5 MJ/kg TS). In beiden Gebiete deckt die verfügbare Nahrung den Energie-
bedarf der Rehe in den entsprechenden Monaten, im landwirtschaftlich geprägten Ge-
biet sogar mehr als ausreichend. Oslage & Strohtmann (1988) berechneten anhand
der Ergebnisse von Weiner (1977) einen Erhaltungsenergiebedarf von Rehen in den
Wintermonaten von 4,5 ME MJ/Rehe/Tag. Bobek et al. (1974) gehen von 3,23
MJ/Tier/Tag–4,0 MJ/ Tier/Tag aus und Onderscheka (1999) gibt eine Wert von 4,3 ME
MJ/Rehe/Tag. Für Bewegung wird dieser um 40 % bis 60 % erhöht, bei Stress bis 150
%. In den Wintermonaten lagen im Durchschnitt der Waldrehe die gefundenen um-
setzbaren Energiewerte bei 4,97 ME MJ/kg TS und bei den Landrehen bei 6,02 ME
MJ/kg TS. Bei Verwendung des Rehpansensaftes stieg der Energiewert sogar auf 5,19
MJ/kg TS im Waldhabitat an. Obwohl unsere Werte den Energiegehalt der Äsung un-
terschätzen und die Fettreserven nicht in Ansatz gebracht werden, übersteigen unsere
Werte den Erhaltungsbedarf deutlich, wie er von Oslage & Strohtmann (1988), Bobek
et al. (1974) oder Onderscheka (1999) berechnet wurde. Zusammen mit den Fettre-
serven haben in beiden Habitaten Rehe genügend Energie, um den Winter und

4 Diskussion
143
das beginnende Frühjahr zu überleben. Dies wird auch durch die Stressanalysen
untermauert.
Nach Sommer (2004) werden die niedrigeren Energiewerte der Waldtiere durch den
höheren Faseranteil (Kap. 3.2) in der Nahrung kompensiert, da dieser durch gesteiger-
te Fermentation für eine erhöhte Wärmeabstrahlung des Pansens sorgt, womit weniger
Energie zur Aufrechterhaltung der Körperkerntemperatur (Hartfiel et al. 1985, Sommer
2004) aufgewendet werden muss und eine höhere Nettoenergie resultiert.
Ein Grund für das gefundene Energieminimum im Februar und nicht im März/April
könnte in der Verschiebung der phänologischen Phasen begründet sein. Die Verschie-
bung der Phasen geht auf die beobachteten Temperaturanstiege in den letzten Jahr-
zehnten zurück (Badecke et al. 2004, Menzel et al. 2006). Die Phasenverschiebung im
Frühling nach vorne wird mit ein Zeitraum zwischen -2,8 bis -5,1 Tage/Jahrzehnt an-
gegeben (Stahl 2016). Zwischen den Arbeiten von Hofmann & Kirsten (1982) und heu-
te haben sich die Phänologischen Phasen um -11,8 Tage nach vorne verschoben
(Stahl 2016). Somit könnte sich das Energieminimum in der Äsung im März sich nach
vorne verlagert haben.
4.4 Qualität der Rehnahrung
4.4.1 Fester Panseninhalt
Die chemische Analyse des Panseninhaltes zur Bestimmung der Rohnährstoffe mittels
des Weender und VanSoest Verfahren wurde zur Beurteilung der Qualität der Rehnah-
rung herangezogen. Diese Methoden gelten als Stand der Forschung zur Ermittlung
der chemischen Zusammensetzung der aufgenommenen Nahrung bzw. der Pflanzen
(Esser 1954, Schmidl 1996, Djordjevic et al. 2006, Kirchgessner et al. 2008). Durch die
direkte Entnahme und Analyse der Panseninhaltsmasse erreicht man die exakte che-
mische Zusammensetzung der vom Rehwild selektierten Pflanzenarten.
Weiterhin wurde das Weender Verfahren zur Bestimmung der Qualität der Rehnahrung
bereits in früheren Studien verwendet (u.a. Esser 1954, Brüggemann et al. 1967, On-

4 Diskussion
144
derscheka 1972, Drescher-Kaden & Seifelnasr 1976, Enzinger & Hartfiel 1989,
Schmidl 1996, Latham et al. 1999, Kamler & Homolka 2005, Djordjevic et al. 2006,
Reimoser 2006, Popovic et al.2009) und bietet somit eine breite Vergleichsmöglich-
keit.
Die Ergebnisse der Analysen der Rohnährstoffe zeigten, dass sich der Fasergehalt der
aufgenommenen Nahrung zwischen Land- und Waldgebiet stark unterscheidet. So ist
der Fasergehalt im forstlich geprägten Gebiet mit durchschnittlich 30 % signifikant hö-
her als im Landgebiet mit einem Anteil von 25 % an der Trockensubstanz. Speziell in
den Herbst-, Winter- und frühen Frühlingsmonaten lagen die Faserwerte der
Waldäsung deutlich höher als im Landgebiet. Im Winter und Frühling besteht der
Hauptteil der Äsung im Wald aus Nadelbäumen, Stauden und Sträuchern, welche für
ihren hohen Fasergehalt und stark verholzte Anteile bekannt sind (Tabelle 8, Tabelle 9,
Tabelle 11, Tabelle 12, Esser 1954, Melchiar 1960, Fred 1970, Albers 1990, Anke et
al. 2007, Djordjevic et al. 2006, Barancekova et al. 2010). Dies erklärt, weshalb nach
der detaillierteren Faseranalyse (3.2.1.2VanSost Analyse) der Anteil an Lignin und Zel-
lulose im Waldhabitat signifikant über den Werten des Landes liegt und die Hemizellu-
lose in beiden Gebieten auf etwa dem gleichen Niveau lag (Abbildung 22).
Das gleiche Bild zeigt sich auch mit niedrigeren Werten im landwirtschaftlichen Gebiet.
Dort lässt sich der Höhepunkt der Rohfaserwerte in einigen Wintermonaten (Dezem-
ber, Januar) verzeichnen. Zu gleichen Ergebnissen kamen Redjadj et al. (2014), wel-
che die Faseranteile in Rehnahrung von September bis Januar untersuchten. Auch
hier stiegen die Lignin- und Zelluloseanteile in der aufgenommenen Nahrung stark an,
wohingegen die Anteile der Hemizellulose abnahmen. Diese Zunahme der Fasergehal-
te lässt sich durch die natürliche Reifung/Alterung der Vegetation erklären. So nimmt
der Zellulose- und Ligningehalt einer Pflanze mit zunehmendem Vegetationsstadium
zu, während der Proteingehalt abnimmt (Esser 1954, Drozdz & Osiecki 1973, Drozdz
1979, Buchgraber 2005, Djordjevic et al. 2006, Reimoser 2006, Baumgärtl 2012).
Der höhere Faseranteil im Wald ist jedoch nicht nur negativ zu werten (Kurt 1970). Ho-
he Faseranteile weisen nicht gleich auf energetisch mindere Nahrung hin, da sich bei
rohfaserhaltiger Nahrung die schwerabbaubaren Fasern im Pansen anreichern (Klötzli

4 Diskussion
145
1965, Meyer 1988). Dissen & Hartfiel 1985 zeigten durch Fütterungsversuche, dass
Rehe gerne Heu aufnehmen, sobald es für sie kleingeschnitten ist oder sie es abrupfen
können (vgl. auch Deutz et al. 2009). Mit ihren Stoffwechselversuchen fanden sie her-
aus, dass Rehe eine recht hohe Verdaulichkeit von Fasern aufweisen, die fast so hoch
war wie jene des „Raufutterfressers“ Ziege. Dies deckt sich mit unserem Vergleich des
Hohenheimer Futterwerttests an Pansenproben, die einmal mit Hammelpansensaft
und einmal mit Rehpansensaft behandelt wurden. Mit dem Rehpansensaft konnte aus
faserhaltiger Äsung signifikant mehr Energie gewonnen (Kap. 3.1.2, Abbildung 19).
Dissen & Hartfiel (1985) fanden eine Verdaulichkeit von Fasern beim Rehwild von 50–
78 %. Für Haustiere leichtverdauliche Futtersorten, wie etwa Luzerne, können vom
Rehwild ohne schwer- oder sogar unverdauliche ligninreiche Rohfasern kaum verdaut
werden. Sie vertreten weiterhin die Meinung, dass ein plötzliches Ausbleiben grobfase-
riger Äsung zu Verdauungsstörungen und unter Umständen zu einem lebensbedrohli-
chen Gewichtssturz führen kann. Dieser Meinung ist auch Buchgraber (2005), der die
Äsung im Frühjahr und Herbst speziell im Grünland und Kulturarten auf Äckern als zu
schnell verdaulich einstuft, sowie ihr einen zu hohen Rohproteingehalt und zu niedri-
gen Rohfasergehalt (unter 20 %) zuschreibt und feststellen musste, dass diese Fut-
teraufnahme zu Durchfall bei den Rehen oder verstärktem Verbiss an der Waldvegeta-
tion führt. Unsere Ergebnisse der chemischen Analysen zeigen, dass bei den Landre-
hen im Frühling und Herbst die hohen NfE- und Proteingehalte der Nahrung mit der
Aufnahme großer Mengen von Feld- und Baumfrüchte zusammenhängen. Gleichzeitig
äsen sie in diesen Jahreszeiten auch große Mengen Süßgräser, um damit den fehlen-
den Fasergehalt zu erhöhen. Auch Abbas et al. (2013) fanden bei chemischen und
makroskopischen Pansenanalysen von Rehwild heraus, dass es zu einer vermehrten
Grasaufnahme bei diesen Tieren in Offenlandschaften/landwirtschaftlichen Flächen im
Herbst und Winter kommt.
Die energiereichen Inhaltsstoffe Proteine und Stickstofffreie-Extrakstoffe haben bei den
Landrehen über das gesamte Jahr gesehen einen höheren Anteil als bei den Waldre-
hen. Während die Gehalte an Rohproteinen in den beiden Habitaten sich nicht signifi-
kant unterschieden, liegt bei den Stickstofffreien-ExtrakstoffeN ein signifikanter Unter-
schied vor. Differenziert man die Werte nach Monaten, ergeben sich größere Unter-

4 Diskussion
146
schiede. Zwischen Januar und Juni beinhaltete die Nahrung der Landrehe deutlich
mehr Proteine als jene der Waldrehe. Die Werte für Februar und März unterschieden
sich signifikant. Ab Juli lagen die Proteinwerte im Wald höher. Fast gegenläufig zu den
Rohproteinen verlaufen im Landhabitat die Stickstofffreien- Extraktstoffe: Ab Juli liegen
ihre Anteile im Land bis zum Januar weit über den Rohproteinen. Im Landhabitat wird
zwischen März und Juni die Energie überwiegend durch Proteine geliefert, ab Juli bis
Januar basieren diese vor allem auf Kohlehydraten und Stärke. Im Waldhabitat be-
kommen erst ab Oktober bis Ende März Stickstofffreie Extraktstoffe für den Energie-
haushalt eine größere Bedeutung. In den anderen Monaten wird auch hier die Energie
überwiegend durch Proteine bereitgestellt.
Diese Ergebnisse decken sich mit Buchgraber (2005) und Zweifel-Schielly (2005), die
in Ackerbaugebieten durch die Wiesen, Weiden und Feldfutterflächen ein kontinuierli-
ches ganzjähriges qualitativ hochwertiges Nahrungsangebot für Rehe vorfanden. So
ließ sich speziell im Winter auf Talwiesen eine deutlich bessere Nahrungsqualität im
Vergleich zur Nahrung im Wald und Bergwiesen finden (Zweifel-Schielly 2005). Zwei-
fel-Schielly (2005) führt dies auf die höhere Verfügbarkeit der Biomasse sowie auf die
Düngung der Wiesen zurück. Ab Juni steigt der Anteil der Feldfrüchte am Gesamtanteil
der aufgenommenen Nahrung in unserem Landgebiet stark an. Feldfrüchte wie Getrei-
de und Mais zeichnen sich durch ihren hohen Kohlenhydrat- und Stärkegehalt sowie
niedrige Fasergehalte aus (Buchgraber 2005), was sich im Verlauf der Stickstoffreien
Extraktstoffe im Landhabitat widerspiegelt (Abbildung 21).
Das Proteinmaximum in beiden Gebieten findet sich im Mai mit 35,5 % TS im Wald
und 36,9 % TS im Land. Dieses Ergebnis lässt sich durch das Vegetationswachstum
erklären, da mit Beginn der Vegetationszeit parallel zum zeitlichen Entwicklungsverlauf
der Jungpflanzen auch der Proteingehalt der Jungpflanzen ansteigt (Buchgraber
2005). In weiteren Studien (Onderscheka 1991, Schmidl 1996, Reimoser 2006,
Djordevic et al. 2006, Popovic et al. 2009) wurden je nach Jahreszeit Rohproteinanteile
im Panseninhalt der Rehe zwischen 20,8 % und 37,4 % TS gefunden. Der Rohprotein-
gehalt unserer Rehe des Waldgebietes schwankte zwischen 17,6 und 35,5 % TS so-
wie im landwirtschaftlich geprägten Gebiet zwischen 20,1 und 36,9 % TS, womit diese
Werte im Rahmen der bekannten Literatur liegen. Betrachtet man den Winter, haben

4 Diskussion
147
Brüggemann et al. (1967) und Anke et al. (1984) 22 % TS Rohproteine vorgefunden,
wogegen wir im Februar im Land auf einen höheren Median von 29 % TS Rohprotein
kamen.
Die von vielen Autoren angegebenen Rohproteinbedarfsmengen des Rehwildes pro
Tag (20 kg Reh) von 5,5 % XP (Eisfeld 1974b), 11 % XP (Ueckermann 1986), 10 – 12
% XP (Enzinger & Hartfiel 1989) oder 9 %–17 % XP (Oslage & Strothmann 1988), lie-
gen weit unter den von uns gefundenen Werten.
Stellt man die Rohproteinwerte der Pflanzen jenen gegenüber, die im Pansen gefun-
den werden, liegen die Pansenwerte um ein Vielfaches über den Pflanzenwerten. Die
Rohproteinanteile (XP) von Waldbäumen liegen bei etwa 12,9 % TS (XP), Laubtriebe
bei ca. 14,3 % TS (XP), Sträucher bei ca. 17,3 % TS (XP), Kräuter bei ca. 19,4 % TS
(XP), Bucheckern bei 14,3 % TS (XP), Gräser bei 3 – 12,6 % TS (XP), Luzerne bei 7 –
17 % TS (XP), Heidelbeere bei ca. 13,1 % TS (XP) und Geranium robertianum bei ca.
14,5 % TS (XP) (Esser 1954, Klötzli 1965, Anke et al. 1978, Veith 1992, Baumgärtl
2012). Drozdz (1973) sammelte Rehnahrung zu verschiedenen Jahreszeiten und fand
wie wir den höchsten Rohproteingehalt der Vegetation im Monat Mai mit 19,7 % XP
(Sommer 16 – 19 % XP) und den geringsten im Winter mit durchschnittlich 8,5 % XP.
Dagegen ist der Proteingehalt im Pansen das Resultat aller gewählten Nahrungsbe-
standteile, der Gesamtmenge an Mikroorganismen und des Stickstoffs, der durch den
ruminohepatischen Zyklus über den Speichel dem Pansen wieder zugesetzt wird (Klein
& Schönheyden 1970, Eisfeld 1975, VanSoest 1982, Meyer 1988, Veith 1992, Djord-
jevic 2006, Deutz 2009). Die großen Bakterienmenge im Pansen können bis zu 65 %
des Rohproteins in der Trockensubstanz ausmachen (Veith 1992).
Geringe Proteingehalte der Nahrung kann das Reh aufgrund des ruminohepati-
schen Zyklus abpuffern (Drozdz 1973), weshalb sich ein relativer Eiweißmangel
bei Rehen, z.B. im Winter, nicht so stark negativ auswirkt (Deutz 2009).
Vergleicht man vorhandene Studien zu Rohnährstoffen in der Rehäsung, finden sich
zwar Unterschiede in der Höhe der Werte, im Jahresverlauf ergeben sich ähnliche
Tendenzen (Tabelle 25).
4 Diskussion
148
Tabelle 25: Ergebnisse der Rohnährstoffanalyse im Vergleich zu Ergebnissen früherer Studien, Mittelwerte der Rohnährstoffe angegeben, n = Anzahl, XP = Rohprotein, XL= Rohfett, XF = Rohfaser, NfE= Stickstofffreie-Extraktstoffe, Rohnährstoffe in [% Trocken-substanz].
Arbeitsgruppe Wildbiologie und Wildtiermanagement Jahreszeit XP XL XF NfE Frühling 29,6 6,0 27,7 27,9
Waldgebiet (n = 106) Sommer 31,0 6,9 26,8 26,0 Herbst 26,0 6,1 28,3 27,3 Winter 21,7 5,0 34,4 30,7 Landgebiet (n = 139) Frühling 33,5 8,3 18,3 26,8 Sommer 27,9 6,3 24,8 32,4 Herbst 24,4 5,7 22,4 36,3 Winter 25,0
4,8 25,1 34,6,
Popovic et al. (2009) Frühling 36,1 11,6 17,0 22,7 Barajevska Raka (n = 43) Sommer 28,8 9,6 23,8 25,6 Herbst 24,7 9,7 28,1 25,1
Winter 26,9 9,7 32,3 15,2 Djordjevicet al. (2006) Frühling 37,4 10,8 15,2 23,5 Barajevska Raka (n = 20) Sommer 29,8 8,5 19,7 28,1 Herbst 24,1 7,5 27,4 25,9 Winter 27,3 7,5 28,3 18,8 Reimoser (2006) Mai/Juli 34,9 7,4 12,6 32,7 Anlauftal (n = 39) Aug/Sep 27,5 6,3 16,1 37,8 Okt/Dez 20,2 4,7 19,7 44,3 Kötschachtal (n = 30) Mai/Juli 30,6 6,7 14,5 36,3 Aug/Sep 27,5 7,7 16,3 35,8 Okt/Dez 20,8 6,0 20,1 41,4 Schmidl (1996) Frühling 33,2 5,6 13,7 33,8 Manhartdberg (n = 42) Sommer 27,9 5,6 15,0 38,7 Herbst 23,6 5,2 18,1 42,1 Winter 23,2 5,2 16,9 44,7 Maissau (n = 46) Frühling 33,7 5,6 12,4 34,0 Sommer 27,7 5,7 15,5 38,1 Herbst 27,2 5,9 16,9 37,9 Winter 26,4 5,8 18,3 38,1 Onderscheka (1991) Frühling 33,6 16,0 Achenkirch (n = 104) Sommer 28,4 17,4 Herbst 21,7 25,2 Winter 23,8 22,7 Drescher-Kaden & Seifelnasr (1976) Jura (n = 13) Herbst 25,5 8,8 24,2 28,1 München Ost (n = 21) Herbst 29,3 7,1 20,1 29,0

4 Diskussion
149
Tendenziell nehmen die Rohproteine (XP) im Jahresverlauf ab, während Rohfasern
(XF) und Kohlehydrate (NfE) zunehmen. Auch die Rohfette nehmen von Frühjahr zum
Winter hin eher ab, nur bei Schmidl (1996) bleiben sie eher konstant bzw. nehmen
teilweise auch leicht zu. Die Rohfasergehalte lagen in den Untersuchungen nie unter
13 % TS, stiegen aber in mehreren Studien auf ähnliche Höhen an, wie wir sie vorge-
funden haben. Sehr geringe Rohfaserwerte im Winter (<20 % TS), wie sie bei
Schmidl (1996) und Reimoser (2006) zu finden sind, die gleichzeitig mit hohen
Kohlehydraten verbunden sind, deuten auf einen Fütterungsbetrieb hin, was bei
Reimoser (2006) im Anlauf- und Kötschachtal der Fall war. Diese Werte sind mit
unseren Ergebnissen, die ohne Fütterung entstanden sind, nur eingeschränkt
vergleichbar und liefern ein vollkommen falsches Bild über den Faser- und Koh-
lehydratbedarf von Rehen speziell im Winter. Dagegen fanden Djordjevicet al.
(2006) und Popovic et al. (2009) sehr ähnliche Fasergehalte in der Äsung, ihre Werte
des Kohlehydratanteils lagen jedoch unter unseren.
Für Damhirsche gaben Drescher-Kaden & Seifelnasr (1976) Rohfasergehalte von 24 –
25 % TS sowie für Mufflon von 22 – 34 % TS an. Reimoser (2006) fand bei Rotwild im
Gasteinertal Rohfasergehalte zwischen Mai und Dezember zwischen 18 % und 25 %
TS. Bei Gams fand er teilweise noch niedrigere Werte von 15 – 21 % TS Rohfaser-
gehalt. Vergleicht man den Rahmen der in Tabelle 25 aufgeführten Rohfasergehalt in
den Pansen von Rehen mit 13 % bis 34 % TS, dann decken sich diese Werte nicht
nur mit den Werten der „Intermediären Äser“ Rothirsch und Gams, sondern erreichen
Fasergehalte, wie sie von Raufutterfresser wie dem Mufflon bekannt sind.
Die Nährstoffgehalte sind nicht nur stark von der Vegetationsentwicklung im
Jahresverlauf und dem Habitat abhängig, unsere Rehe können sich in einem
sehr weiten Spektrum diesem anpassen. Der Begriff „Konzentrat-Selektierer“ ist
für das Reh unzutreffend.
4.4.2 Pansensaft
Parallel zu den Rohnährstoffanalysen und den Energieanalysen wurden Pansen-
saftanalysen vorgenommen. Dies wurde als sinnvoll angesehen, da wir mit vorverdau-

4 Diskussion
150
tem Pansenmaterial arbeiteten und somit ein Teil der Nährstoffe und Energie bereits in
gelöster Form im Pansensaft vorlag. Die Analysen des Pansensaftes erfassten die
bereits entstandenen, im Pansensaft gelösten Fermentationsprodukte. Die Parameter
des Pansensaftes wie flüchtigen Fettsäuren, Ammoniakgehalt, Laktat- und pH-Wert
geben außerdem Hinweise auf die Qualität der aufgenommenen Nahrung ((Brügge-
mann & Giesecke 1968), Brüggemann et al. 1968, Drescher-Kaden & Seifelnasr
1977b, Enzinger & Hartfiel 1998, Sommer 2004).
• Hohe Konzentrationen an flüchtigen Fettsäuren (FFS) allgemein, an Laktat und
Ammoniak (Abbildung 34) sind Zeiger für einen hohen Energiegehalt der Nah-
rung.
• Die Fettsäuren „Essigsäure“ und „Propionsäure“ geben einen Hinweis auf
leicht- bzw. schwerverdauliche Fasern. Das Verhältnis von Essig- zu Propions-
äure zeigt, ob mehr oder weniger schwer- bzw. leichtverdauliche Äsung aufge-
nommen wurde.
• Buttersäure entsteht durch Stärke in der Nahrung.
• Der pH-Wert mit zunehmenden Anteilen an Proteinen und / oder Kohlehydrate
in der Äsung (Kamphues et al. 2008, Kirchgessner et al. 2008).
Die Ergebnisse der Pansensaftparameteranalyse decken sich mit den Ergebnissen der
Rohnährstoffanalyse und dem Energiegehalt der Äsung. Die Landrehe haben niedrige-
re pH-Werte und Essigsäure-Propionsäureverhältnisse (Abbildung 31), dafür liegen
alle anderen Parameter – angefangen mit der Menge an FFS (Abbildung 28), Anteil an
Propion- und Buttersäure (Abbildung 30) sowie Laktat (Abbildung 35) und Ammoniak
(Abbildung 32) – über jenen der Waldrehe.
Das enge Verhältnis zwischen FFS und Energie zeigt sich im Wald, wo das Maximum
der FFS im Monat Mai zum Zeitpunkt der höchsten Rohprotein- und Energiewerte er-
reicht wird. Umgekehrt findet sich im Wald das Minimum der FFS im Februar mit den
niedrigsten Rohprotein- und Energiewerten. Gleiches kann im Landgebiet beobachtet
werden. Hier liegen die Maxima der FFS-Konzentrationen im November, Juni und Au-

4 Diskussion
151
gust. In den gleichen Monaten steigen die Energiewerte oder auch NfE-Werte der Nah-
rung im Landgebiet stark an. Das FFS-Minimum stellt sich hier im Februar ein.
Im Jahresmittel erreichen die Waldtiere FFS-Werte von 90,8 mmol/l und die Land-
tiere Werte von 95,3 mmol/l. Behrend (1999) gibt als Jahresdurchschnittswert bei im
Gatter gehaltenen Rehen 60,3 mmol/l an. Drescher-Kaden & Seifelnasr (1977b) analy-
sierten die FFS-Werte von verschiedenen Wildwiederkäuer im Herbst und fanden FFS
Werte von 140- 156 mmol/l bei Rehen. Da der Herbst allerdings als Hochzeit leichtfer-
mentierbarer Nahrung angesehen werden kann, überraschen diese hohen Werte nicht.
Zu dieser Zeit erreichen auch unsere Untersuchungstiere ihre Maximalwerte im Land-
gebiet mit 148,9 mmol/l. Die Tiere des Waldgebietes weisen einen durchschnittlichen
Wert von 106,1 mmol/l im Herbst auf.
Die Landtiere weisen in elf von zwölf Monaten ein engeres Essigsäure-
Propionsäure (C2/C3)-Verhältnis von 2,4:1 auf als die Waldtiere mit einem Verhät-
lnis von 2,8:1. Behrend (1999) erhielt bei Gatterrehen eine C2/C3-Verhältnis von 2,3:1
und bei Mufflons von 3,4:1, was auf eine deutlich energiereiche Nahrung der Rehe
hinweist. Unsere Rehe im Landhabitat hatten ohne Fütterung energiereichere Nahrung
als die gefütterten Gatterrehe von Behrend (1999). Drescher-Kaden & Seifelnasr
(1977b) gingen beim Reh generell von einem C2/C3-Verhältnis zwischen 2,5:1 bis 3:1
aus. Diese Werte entsprachen in etwa den Verhältniszahlen von C2/C3 von Damhir-
schen (3,2:1; 3,2:1) sowie Mufflons (3,8:1 und 4,0:1). Zwar hatten die Rehe von Dre-
scher-Kaden & Seifelnasr (1977b) das engere Essigsäure/Propionsäure-Verhältnis als
Damhirsch und Mufflon, jedoch zeigen die Werte, dass genauso wie unsere Waldrehe
die Rehe im Ebersberg Wildpark (Drescher-Kaden & Seifelnasr 1977b) gut mit faser-
haltiger Äsung zurechtkommen.
Der pH-Wert steht in enger Verbindung mit den Mengen gebildeter Pansensäuren. So
wird bei übermäßigem Konsum leichtfermentierbarer Nahrungsbestandteile auch der
pH-Wert sinken. Dies ist die Folge zu rascher Fermentation, Bildung großer Mengen
an Säuren sowie fehlender Faserstruktur, die das Wiederkäuen und damit die Abpuffe-
rung durch alkalischen Speichel begünstigt (Giesecke 1963, Enzinger 1989). Die Land-
rehe unserer Studie weisen speziell im Herbst (pH= 6,07) und Winter (pH=5,81) einen

4 Diskussion
152
niedrigen pH-Wert auf. Entsprechend der natürlichen Vegetationsentwicklung und ei-
nem zunehmenden Faseranteil in der Äsung (Tabelle 25) müssen beim Reh die pH-
Werte im Pansen zum Herbst und Winter hin ansteigen (Djordevic et al. 2006). Dieser
Trend ist bei unseren Waldtieren erkennbar.
Die Erklärung, warum sich der pH-Wert der Landrehe anders verhält, ist die Aufnahme
von kohlenhydratreichen und faserarmen Material im Herbst und Winter (Abbildung 21,
Abbildung 41). Auch Enzinger (1989) und Ritz et al. (2013) stellten fest, dass durch
Kraftfuttergabe oder durch zusätzliche Fütterung im Winter der pH-Wert beim Rehwild
stark sank. Im Falle von Enzinger (1989) sank der pH-Wert von 6,4 auf 5,9 und im Fal-
le von Ritz et al. (2013) wiesen die Tiere ohne Fütterung einen pH-Wert von 5,7 und
Tiere mit Winterfütterung einen pH-Wert von 5,5 auf. Im Jahresdurchschnitt decken
sich die pH-Werte im Waldgebiet von pH 6,3 mit den Werten von Enzinger (1989) für
normale Nahrungsaufnahme mit pH 6,4 und im Landgebiet mit pH 6,1 tendieren sie zu
Enzingers (1989) pH Wert für gesteigerter Kraftfutteraufnahme von pH 5,9. Die sehr
niedrigen pH-Werte von Ritz et al. (2013) konnten hier nicht gefunden werden, jedoch
liegen unsere Rohfaseranteile auch im oberen Rahmen verglichen mit anderen Arbei-
ten (Tabelle 25).
Wie der pH-Wert so lassen auch hohe Laktatwerte auf eine protein- und kohlenhyd-
ratreiche und rohfaserarme Nahrung schließen (Brüggemann & Giesecke 1968). Die
Waldtiere weisen einen Laktatdurchschnittswert von 3,76 g/l auf, die Landtiere hinge-
gen einen Durchschnittswert von 5,86 g/l. Entsprechend den hohen Proteinwerte im
Frühjahr und hohen Kohlehydratwerte im Herbst finden sich in beiden Gebieten die
beiden Verlaufsspitzen im Frühling und Herbst.
Ammoniak entsteht bei der mikrobiellen Zersetzung des Nahrungsproteins und dient
den Mikroorganismen zum Aufbau ihrer eigenen Proteine. Je mehr Ammoniak sich im
Pansen befindet, desto mehr Proteinbiosynthese der Mikroorganismen ist möglich und
umso mehr Mikrobenprotein steht dem Reh bei der späteren Verdauung im Lab-
magen/Dünndarm zur Verfügung. Bei zu vielen Nahrungsproteinen wird jedoch Am-
moniak schneller gebildet, als er von den Mikroorganismen verarbeitet werden kann.
Es kommt zu einer Anflutung von Ammoniak im Pansen, der durch die Pansenwand in

4 Diskussion
153
die Blutbahn und zur Leber transportiert wird (Schröder 2003). In der Leber erfolgt die
Entgiftung des Ammoniaks durch Umwandlung in Harnstoff (Enzinger & Hartfiel 1989).
Bei großen Mengen des Ammoniaks schränkt dies die Tiergesundheit durch Leber-
und Stoffwechselbelastung sowie durch eine Insulinabsenkung im Rehkörper ein. Als
Folge daraus resultiert eine erhöhte Fettabbaurate und ein starker Energiemangel (En-
zinger & Hartfiel 1998).
Der Ammoniakgehalt ist somit ein sehr sensibler Parameter, welcher stets im ausge-
wogenen Verhältnis stehen sollte. Als Richt- und Grenzwerte des Ammoniakgehaltes
schreiben Piatkowski & Nagel (1975) von einem optimalen Sollwert im Pansen von
8,8–17,6 mmol/l Pansensaft. Sie sehen einen Wert von ca. 58 mmol/l im Pansensaft
als oberste Grenze an. Deutz (mündlich) spricht von einem Richtwert im Pansen von
Rehen zwischen 5–15 mmol/l Ammoniak. Er sieht Werte im Bereich 30–35 mmol/l als
kritisch und 60 mmol/l im Pansensaft als hoch toxisch an.
Die Ammoniakwerte der Waldrehe weisen einen Jahresmedian von 13,1 mmol/l auf.
Die Landtiere liegen mit 21,8 mmol/l signifikant höher als die Waldtiere. Der Wert der
Waldtiere liegt genau im Optimalbereich. Der Ammoniakwert der Landtiere liegt über
den Sollwerten. Drescher-Kaden & Seifelnasr (1977b) fanden bei Rehen im Jura, die
sich proteinreicher ernährten, einen Ammoniakwert im Pansensaft von 18,8 mmol/l.
Rehe mit hohen Fasergehalten in der Äsung hatten bei ihnen einen Wert von 10,6
mmol/l. Diese Ergebnisse decken sich mit unseren Ergebnissen. Enzinger & Hartfiel
(1989) konnten bei ihren Fütterungsversuchen mit Rehwild zeigen, dass mit zuneh-
mender proteinreicher und energiereicher Nahrung der NH3-Wert von 17 mmol/l auf
29,9 mmol/l anstieg. Tataruch & Onderscheka (1993) fanden Werte von 15,8 mmol/l
(Median) und 19,4 mmol/l (Mittelwert) bei Rehwild im Pansensaft.
4.5 Nahrungsaufnahme & -Verfügbarkeit
In einer Vielzahl von Studien wurde das Nahrungsspektrum der Rehe untersucht (Es-
ser 1954, Bubenik 1959, Möhring 1963, Klötzli 1965, Schäfer 1965, Ueckermann 1971,
Klein 1972, Treichler 1972, Ramin 1975, Koch 1976, Kurt 1978, Raesfeld et al. 1978,
Cederlund et al. 1980, Jackson 1980, Dittrich & Groppel 1980, Kioroglanidis 1981, Pa-

4 Diskussion
154
pageorgiou et al. 1981, Kaluzinski 1982, Meyer 1988, Albers 1990, Veith 1992, Fielitz
& Albers 1996, Schmidl 1996, Latham et al. 1999, Reimoser 2006, Storm et al. 2008,
Barancekova et al. 2010, Argunow & Stepanova 2011, Barta 2012, Abbas et al. 2013,
Redjadj et al. 2014). Die chemischen Analysen liefern zwar die Rohnährstoffwerte der
aufgenommenen Nahrung und spiegeln somit die Qualität der Nahrung wider, die mak-
roskopische Inhaltsanalyse liefert die Information, woher diese Werte bzw. durch wel-
che Pflanzen oder Nahrungskomponenten diese Werte verursacht werden (Anthony &
Smith 1974, Mcinnis 1983, Cornelis 1999, Osborn et al. 1997, Tixier & Duncan 1996).
Einige Studien konnten jahreszeitliche oder geschlechtsspezifische Unterschiede bei
der Nahrungswahl bei Rehen erkennen. Aber fast alle kamen zu dem Schluss, dass
das jeweilige Habitat und die dortige Nahrungsverfügbarkeit ausschlaggebend sind für
die Nahrungsselektion des Rehwildes.
So fällt auf, dass die Waldrehe und Landrehe sich bevorzugt von Weichäsung wie
Kräutern, Gräsern und Stauden ernähren. Die Waldrehe griffen hauptsächlich zu Kräu-
tern und Stauden, die Landrehe eher zu Gräsern und Kräutern. Diese Selektion ist
auch in anderen Studien erkennbar (Klötzli 1965, Veith 1992, Storm et al. 2008). Dies
liegt an der unterschiedlichen vegetativen Beschaffenheit der beiden Habitate und wird
durch die pflanzensoziologischen Aufnahmen bestätigt. Der höhere Grasanteil ist den
offenen Flächen, Wiesen und Weiden des landwirtschaftlich geprägten Habitats zuzu-
ordnen. Die Aussage, dass Rehwild keine Gräser zu sich nimmt und auch nicht ver-
dauen kann (Hofmann 1976), kann nicht bestätigt werden und wurde bereits durch
zahlreiche Studien widerlegt (Ramin 1975, Dissen & Hartfiel 1985, Argunow & Ste-
pamova 2011). Abbas et al. (2013) konnten eine vermehrte Aufnahme von Gras im
Winter verzeichnen sowie deutlich höhere Grasprozentwerte im Pansen von Feldrehen
im Vergleich zu Waldrehen finden. Kioroglanidis (1981) fand bei 100 % seiner Unter-
suchungstiere Gräser im Pansen.
Neben Weichäsung ist allerdings auch deutlich zu sehen, dass weitere Schwer-
punkte der Nahrung der Waldrehe bei der Zähäsung liegen. Bei den Landrehen
hingegen liegt der Schwerpunkt bei der Aufnahme von Feldfrüchten und Baum-
früchten. Koch (1967) stellte in landwirtschaftlichen Gebieten fest, dass kein Bedürfnis
des Wildes zu sehr artenreich zusammengesetzter Nahrung besteht, sondern dass das

4 Diskussion
155
Rehwild eine relativ einförmige Auswahl des örtlichen Angebotes wahrnahm. Koch
(1967) und Kaluzinski (1982) fanden in den Pansen einen 85 – 88 % Anteil landwirt-
schaftlicher Nutzpflanzen, welcher sich im Winter noch erhöhte Barta (2012). Cornelis
et al. (1999) analysierten eine Zusammenfassung und Bewertung verschiedener Pan-
seninhaltsstudien zum Reh und kamen zu dem Schluss, dass die Nahrungszusam-
mensetzung von Rehen habitatabhängig ist und nach Jahreszeiten variiert (vgl. Esser
1954, Klötzli 1965, Schäfer 1965, Latham et al. 1999 Barancekova et al. 2010). Im De-
tail wird die Äsungswahl durch das Alter (Holisova et al. 1984, Schmidl 1996, Djord-
jevic 2006), Geschlecht oder persönlichen Präferenzen (Esser 1954, Klötzli 1965) mo-
difiziert, womit die Variationsbereite der gefundenen Ergebnisse in den Studien erklärt
werden kann. Damit werden die Variationen der Energiegehalte, Rohnächstofanteile
((?)) und des flüssigen Pansenmaterials durch die Vegetationsaufnahme des Pansens
und Untersuchungsgebietes gestützt und ergänzt.
4.6 Pansenanpassung
Das Mikrobiom des Pansens gibt Aufschluss über die saisonale Anpassung des Pan-
sens bei Rehwild. Es ist ein empfindlicher Mikrokosmos, welcher schnell auf Verände-
rungen wie energiereiche oder energiearme Nahrung reagiert, und zwar durch Verän-
derung der Zusammensetzung oder Anzahl der Mikroorganismen (Enzinger 1989,
Sommer 2004). Die Mikrobiota der Land- und Waldrehe wurden mit molekularbiologi-
schen Methoden qualitativ und quantitativ analysiert. Es gibt derzeit keine weitere Stu-
die, die mit Hilfe von molekularbiologischen Methoden das Mikrobiom des Rehwildes
untersucht hat.
Frühere Studien wie die von Brüggemann et al. (1967), Prins & Geelen (1971), Kamler
(1999), Drescher-Kaden & Seifelnasr (1977), Enzinger (1989), Imai et al. (1981) und
Teichler (1972) untersuchten die Pansenflora und -Fauna mikroskopisch und kamen
daher oft nur zu unzureichenden Aussagen.
Bisher gibt es auch keine Studien, die das Mikrobiom zweier Tierpopulationen, die in
stark unterschiedlich geprägten Habitaten leben, miteinander verglichen haben. So fällt
auf, dass die Anzahl der Mikrobiota im Pansen der Waldtiere viel höher (Abbildung 53)

4 Diskussion
156
ist als bei den Landtieren. Dies betrifft alle Mikroorganismen-Stämme mit Ausnahme
der Protozoen. Die auch hier gefundene höhere Anzahl an Bakterienstämmen im Pan-
sen der Waldrehe (Abbildung 54) führt Buchgräber (2005) auf die größere Vielfalt,
stärkere Struktur und protein- und stärkeärmere Nahrung im Waldgebiet (Abbildung
21) zurück, die das mikrobielle Wachstum der Bakterien begünstigt. Die ebenfalls hö-
here Anzahl an anaeroben Pilze bei den Waldtieren ist auf den höheren Ligningehalt in
der Nahrung der Waldtiere zurückzuführen (Abbildung 22). Das höhere Aufkommen
der Fibrobacter sowie der Ruminococcen ist ebenfalls durch den höheren Fasergehalt
der Waldnahrung erklärbar (Abbildung 20, Abbildung 21), denn die aufgenommenen
Fasern werden von den Fibrobactern und Ruminococcen zu flüchtigen Fettsäuren,
genauer zur Essigsäure, fermentiert (Kirchgeßner et al. 2008).
Dass die generelle Anzahl der Mikroorganismen im Pansen der Waldrehe deutlich hö-
her ist, könnte durch die geringeren Anzahlen an Protozoen begründet sein (Abbildung
52). Dies würde auch mit den höheren Protozoenzahlen und generell niedrigeren Bak-
terienzahlen der Landtiere übereinstimmen, denn Protozoen werden durch energie-,
eiweiß- und kohlenhydratreiche Nahrung stark begünstigt (Dennis 1983, Enzinger &
Hartfiel 1989, Sommer 2004). Diese energie-, eiweiß- und kohlenhydratreichere Nah-
rung konnte bei den Landtieren nachgewiesen werden und lässt die Anzahl der Proto-
zoen steigen. Diese besitzen die Fähigkeit, Bakterien zu verdauen und somit zur Re-
duktion der Bakterienpopulation beizutragen (Enzinger & Hartfiel 1989).
Enzinger (1989) konnte zeigten, dass durch Abtötung oder starker Verringerung der
Protozoen die Bakterienanzahl um das 2- bis 3-fache ansteigt. Diese Annahme wird
bestätigt durch höheren NH3-Werte im Pansensaft der Landtiere (Abbildung 32). Bei
der Zersetzung von Pansenbakterien durch Protozoen wird NH3 freigesetzt und verur-
sacht höhere NH3-Werte. Höhere NH3-Werte wie auch höhere Protozoenanzahlen las-
sen sich besonders bei den Tieren des Landhabitats finden. Brüggemann et al. (1967)
geht weiterhin davon aus, dass die bei Rehen entdeckten Protozoenarten weniger
empfindlich gegenüber sauren pH-Werten sind, was auch wiederum die hohen Werte
erklärt.

4 Diskussion
157
Diese Aussage deckt sich durch die weitgehend gleichen Anzahlen an Entodinium in
den Pansen der Tiere beider Habitate. Ein weiterer Grund für deutlich geringere Mikro-
organismen der Landtiere könnten die größeren Mengen flüchtiger Fettsäuren und der
niedrigere pH-Wert im Pansen sein. Dieses saure Milieu vermindert das Wachstum
nützlicher Bakterienarten. Die Vermutung wird durch die Laktatmessung unserer Tiere
bestätigt: Zu hohe Mengen an Säure fördern die Laktatbildner und reduziert die Laktat-
verwerter (Kirchgessner et al.2008).
Ein weiterer Punkt der Mikrobiomanalyse ist die Entwicklung des Mikrobioms in den
verschiedenen Jahreszeiten. Während des Frühlings und Sommers zeigen die Wal-
drehe relativ gleiche Gesamtzahlen an Mikroorganismen. Ihren Höhepunkt erreichen
die Mikroorganismen-Anzahlen im Waldgebiet im Herbst und Winter. Bei den Landtie-
ren sind ein Tief im Frühling und sehr ähnliche und gleichbleibende Werte der Anzah-
len im Sommer bis Winter zu verzeichnen. Allgemein lässt sich sagen, dass die Mikro-
biomwerte der Landtiere weniger stark fluktuieren. Dies könnte auf die geringeren qua-
litativen Unterschiede der Nahrung zwischen den Jahreszeiten sowie auf die größere
Einseitigkeit der Nahrung in Ackerbaugebieten zurückzuführen sein (Buchgraber
2005).
Brüggemann et al. (1967) untersuchten die Mikroorganismenzahlen beim Rehwild im
Jahresverlauf. Die Ergebnisse von Brüggemann et al. (1967) und die hier vorgefunde-
nen stimmen weitestgehend überein. Auch bei Brüggemann et al. (1967) nehmen die
Mikroorganismen im Sommer etwas ab und steigen im Herbst stark an. Der Unter-
schied zwischen den beiden Untersuchungen besteht darin, dass bei Brüggemann et
al. (1967) die Werte im Winter stark absinken. Die MO-Werte der Waldtiere hingegen
bleiben im Winter konstant. Ursache für die unterschiedlichen Ergebnisse im Winter in
beiden Untersuchungsgebieten im Vergleich zu Brüggemann et al. (1967) kann in der
hier anderen Äsungsgrundlage in den Habitaten begründet sein (Brüggemann et al.
1967, Prins & Geelen 1970).
Das Pansenvolumen ist ebenfalls ein Weiser für die Anpassung des Pansens an die
Jahreszeiten und die zu dieser Zeit verfügbare Nahrung im Habitat (Hofmann 1989).
Deutlich ist bei den Tieren beider Habitate eine jahreszeitliche Anpassung zu sehen.

4 Diskussion
158
Das Pansenvolumen-Maximum ist bei den Waldtieren im Sommer, bei den Landtieren
im Sommer und Herbst zu sehen. In beiden Fällen fällt das Pansenvolumen im Winter
stark ab: bei den Waldtieren von 4,5 l im Sommer auf 3,7 l im Winter; bei den Landre-
hen von 4,1 l im Sommer auf 3,6 l im Winter. Dies entspricht einer Reduktion des Pan-
senvolumens der Tiere im Waldgebiet um 18 % und um 13 % bei den Landrehen. So-
mit weisen die Waldrehe nicht nur ein größeres Volumenmaximum, sondern auch eine
stärkere Reduktion des Volumens auf. Hofmann (1989) und Kamler et al. (2002) bestä-
tigen den Verlauf der Reduktion des Pansenvolumens bei Rehwild vom Sommer zum
Winter hin. Hofmann (1989) schreibt von einer Reduktion zwischen 20 und 30 %. Eine
derart starke Reduktion lässt sich nur bei den Waldtieren beobachten.
Eine Nivellierung des Pansenvolumens der Landtiere spricht somit für ein eher gleich-
bleibendes Nahrungsangebot und -Qualität der Rehnahung, da dort die Reduktion
auch die Schwankungen des Pansenvolumens deutlich geringer ausfällt. Auch wird
eine solche Reduktion durch faserreiche Nahrung wie im Waldhabitat begünstigt. Auch
bei den untersuchten Rehen von Kamler et al. (2002) ist eine Abnahme des Pansenvo-
lumens in der Vegetationszeit von 7,2 l auf 6,0 l im Winter zu beobachten. Diese Re-
duktion von 17 % deckt sich fast exakt mit der unserer Waldrehe.
Im Jahresmittelwerten lag im Waldhabitat das Pansenvolumen bei 4,1 l und im Agrar-
habitat bei 3,9 l. Feustel (1967) sowie Hofmann (1976) fanden ähnliche Werte. Sie ge-
ben Pansenvolumen-Durchschnittswerte von 3,5 l (Feustel 1967) bis 4,2–5,5 l (Hof-
mann 1976) an. Der Rückgang der Werte von Kamler et al. (2002) von 7,2 l auf 6,0 l
liegt jedoch weit über den hier gefunden Werten.
Die Panseninhaltsmenge wird durch die Intensität der Nahrungsaufnahme und das
verfügbare Pansenvolumen bestimmt, sie passt sich ebenfalls dem Jahresverlauf und
der Vegetation an. Mehrere Autoren wie Dissen & Hartfiel (1985) und Klötzli (1965)
belegen, dass die Nahrungsaufnahme selbst bei ad libitum Nahrungsverfügbarkeit im
Winter abnimmt. Bei den Landrehen war dies ebenfalls der Fall, jedoch nicht bei den
Rehen des Waldgebietes. Selbst bei Betrachtung des Verhältnisses der prozentualen
Füllmenge zum Pansenvolumen ist ein starker Anstieg vom Sommer zum Winter hin

4 Diskussion
159
bei den Waldrehen zu verzeichnen. Die Waldrehe haben entgegen der bisherigen
gängigen Meinung durch mehr Äsung die niedrigeren Energiewerte ausgeglichen.
Bei den Landrehen wurde der Höhepunkt der Panseninhaltsmenge und der Pansenfül-
lung im Sommer gemessen. Kamler et al. (2002), Argunow (2011) sowie Drozdz
(1973) konnten eine Abnahme des Pansengewichts bzw. der Nahrungsaufnahme zum
Winter hin nachweisen. Drozdz (1973) zeigte, dass die Nahrungsaufnahme mit 600 g
TS Biomasse pro Tag im September am höchsten und im Winter mit nur noch 350 g
TM Biomasse am geringsten ist. Auch Argunow & Stepamova (2011) zeigten mit ihrer
Studie, dass das durchschnittliche Panseninhaltsgewicht zwischen den Monaten Mai
und August bei 2,9 kg liegt sowie in den Monaten Oktober bis März nur bei 2,7 kg. Bei
Kamler et al. (2002) nahm das durchschnittliche Panseninhaltsgewicht mit 1,45 kg in
der Vegetationsperiode bis zum Winter hin ab; zu diesem Zeitpunkt lag es nur noch bei
1,1 kg. Die Werte von Kamler et al. (2002) decken sich mit den unseren. Die Werte der
Rehe im Landgebiet schwanken von 1,2 kg im Sommer und 1,0 kg im Winter. Im
Waldgebiet steigen sie von 1,2 kg im Frühling und Sommer bis auf 1,4 kg im Herbst
an. Eine mögliche Erklärung, warum die Rehe des Waldgebietes trotz Abnahme des
Pansenvolumens im Winter mehr Nahrung zu sich nehmen, könnte die stark faserhal-
tige Nahrung sein.
4.7 Gewicht und Kondition
Das mittlere Gewicht der aufgebrochenen Waldrehe lag bei 14,8 kg und das der Land-
rehe bei 15,4 kg. Der Gewichtsverlauf der Rehe beider Habitate zeigt im Jahresverlauf
jeweils einen Einbruch im Juni und einen im September.. Das Absenken der Gewichte
im Juni ist als Folge des geringeren Energiegehaltes der Nahrung durch Einlagerung
von Lignin und Cellulose sowie der Territorialkämpfe der Böcke (Stubbe 1997) zu se-
hen. Das geringere Gewicht im September ist auf das Brunftgeschehen zurückzufüh-
ren (Ellenberg 1978, Wotschikowsky 1994, Hewison 1996).
Ellenberg (1978) und Reimoser (2006) sprechen von einem weiteren Gewichtsein-
bruch bei Rehen in freier Wildbahn. Demnach wäre ein Gewichtsminimum im Spätwin-
ter zu erwarten. Dies kann durch unsere Ergebnisse nicht bestätigt werden. Beide

4 Diskussion
160
Rehpopulationen zeigten recht konstante Gewichte im Winterverlauf. Zu diesem Er-
gebnis kam auch Hewison (1996), der für die Waldrehe zwar einen leichten Abfall ihrer
Gewichte im Monat März verzeichnete, bei den Feldrehen hingegen aber sogar einen
Anstieg feststellte. Das konstante Halten der Gewichte während des Winters weist auf
eine gute und ausreichende Nahrungsverfügbarkeit und -Qualität hin. Das Absinken
des Gewichtes der Waldrehe im März liegt völlig im Normalbereich und liegt deutlich
über den beiden Gewichtsminima im Juni und September. Der Gewichtsanstieg der
Landrehe im Monat März zeigt, dass dort deutlich früher Nahrungsquellen verfügbar
sind als im Waldhabitat. Das größte Gewichtsmaximum erreichen die Tiere in beiden
Habitaten im Herbst. Dies lässt sich durch das spezifische Nahrungsangebot des
Herbstes erklären wie Eicheln, Bucheckern oder andere Früchte sowie die gesteigerte
Intensität der Nahrungsaufnahme (Hofmann & Herzog 1980, Deipenbrock 1985). Der
Gewichtsanstieg ist jedoch bei den Landtieren signifikant höher als bei den Waldtieren.
Dies ist auf die Aufnahme der Feld- und Baumfrüchte zurückzuführen, welche deutlich
mehr Energie liefern und deren Aufnahme durch die Panseninhaltsanalyse bestätigt
wurde. Durch diese gesteigerte Nahrungsaufnahme und die energiereiche Nahrung im
Herbst können sich die Rehe Fettreserven für den Winter anfressen (Deipenbrock
1985).
Insgesamt lagen die Gewichte der Landtiere fast immer über den Gewichten der Wald-
tiere liegen. Die Landtiere kamen zudem mit höheren Gewichten aus dem Winter in
den Frühling und konnten deutlich früher und schneller wieder an Gewicht zunehmen
als die Waldtiere, was an der höheren Energie in der Nahrung lag (vgl. Enzinger &
Hartfiel 1989).
Von Andersen (2000) ist bekannt, dass weibliches Rehwild mit guten Körpergewichten
eine 40 % höhere Reproduktionserfolg aufweist als weibliche Tiere mit schlechten Kör-
pergewichten. Diese Tatsache lässt die Schlussfolgerung zu, dass in Gebieten mit aus-
reichender, qualitativ hochwertiger Nahrung, wie sie in unseren Habitaten gegeben ist,
deutlich höhere Reproduktionsraten aufgrund guter körperlicher Verfassung des weib-
lichen Rehwildes zustande kommen. Dabei fallen besonders die höheren Gewichte der
Kitze und subadulten Tiere des Landgebietes unserer Studie ins Auge. Die weiblichen
Tiere des Landgebietes zeigen höhere Gewichte in den Monaten Oktober und Novem-

4 Diskussion
161
ber auf. Sie starten somit mit höheren Körpergewichten in den Winter und weisen im
Frühjahr eine bessere Kondition auf. Hinzu kommt das frühere Nahrungsangebot im
Landgebiet im Frühling. All diese Gründe führen zu fitteren Geißen, welche mehr oder
stärkere Kitze hervorbringen. Bei einer sehr guten Nahrungsverfügbarkeit und -Qualität
steigt die Reproduktion einer Rehpopulation (Ellenberg 1978, Andersen 2000).
Neben dem Körpergewicht der aufgebrochenen Rehe wurde zur Beurteilung der Kon-
dition der Untersuchungstiere der Nierenfettindex nach Riney (1955) herangezogen.
Wie auch bei den Gewichten der Rehe aus den zwei Gebieten zeigt sich auch beim
Nierenfettindex ein Unterschied. Dieser ist aber weniger deutlich ausgeprägt. So gibt
es nur einen geringfügigen Unterschied in den Jahreszeiten Frühling, Sommer und
Winter. Hier scheinen die Tiere beider Habitate nahezu gleiche Nierenfettindizes zu
haben. Allein im Herbst weisen die Rehe des Landgebietes höhere Nierenfettindizes
auf. Dieses Ergebnis deckt sich mit den Gewichtserhebungen. Auch hier zeigen die
Rehe der beiden Habitate im Herbst den größten Gewichtsunterschied, was mit der
höheren Nahrungsqualität und Energiegehalt der Nahrung zusammenhängt, wie Fütte-
rungsversuche zeigten (Warren & Kirkpatrick 1982, Santos 2013). Den Höhepunkt des
Nierenfettindexes fanden auch Deipenbrock (1985), Enghofer (1990) und Georgii
(1973) im Herbst. Bei Georgii (1973) stieg das Nierenfett von 30 g auf 80 g zwischen
September und Dezember an. Bei Enghofer (1990) hob sich der Nierenfettindex der
Rehe von September mit 35 % bis November auf 125 % an. Der Nierenfettindex unse-
rer Landrehe steigt von September bis November auf 102 %, jener der Waldrehe auf
75 %, jedoch bis Ende Februar auf 78 % an. Beide Rehpopulationen erreichen im
Frühjahr den gleichen Ausgangswert. Das absolute Konditionstief ist im Sommer zu
verzeichnen und sicher auf das Brunftgesehen zurückzuführen.
4.8 Stress
Die Stressanalysen sollten den Gesamteindruck der beiden untersuchten Rehpopulati-
onen dahingehend abrunden, dass die höheren Energiewerte der Landrehe nicht etwa
durch höheren Stress aufgebraucht werden. Habitat-Fragmentierung, das Leben in der
Nähe von Autostraßen (Millspaugh et al. 2001, Navarro-Castilla et al. 2014) oder auch

4 Diskussion
162
menschliche Aktivitäten wie der Erholungstourismus (Thiel et al. 2011, Zwijacz-Koziac
et al. 2012, Rehnus et al. 2014) können die GCM-Gehalte im Kot von Tieren deutlich
erhöhen.
Unsere Rehe zeigten keine signifikanten Unterschiede zwischen den Habitaten. Zu
gleichen Ergebnissen kam auch Sauerwein et al. (2004), die GCM-Werte bei Rotwild in
sieben verschiedenen Gebieten untersuchten und auch dort keine Unterschiede im
GCM-Gehalt bezüglich der Habitatqualität feststellen konnten. Auch Dalmau et al.
(2007) konnten bei der Untersuchung von Gamswild keinen Effekt von menschlichen
Störungen in Habitaten finden. Auch nach Alter differenziert konnten keine signifikan-
ten Unterschiede zwischen den Individuen festgestellt werden. Dies deckt sich erneut
mit Sauerwein et al. (2004), die bei Rotwild keine GCM-Altersunterschiede entdecken
konnten. Corlatti et al. (2014) konnten hingegen bei Gamswild während der Brunftzeit
Unterschiede in den GCM-Werten bei männlichen Tieren messen. Hier zeigten ältere
Tiere weniger Stress als junge Böcke, dies führt Corlatti et al. (2014) auf die größere
Erfahrung der älteren Böcke bei Rangkämpfen zurück.

5 Literatur
163
5 Literatur
Abbas, F., Picot, D., Merlet, J., Cargnelutti, B., Lourtet, B., Angibault, J-M., Daufrense, T., Aulagnier, S., Verheyden, H. (2013): A typical browser, the roe deer, may consume substantial of grasses in open landscapes. Eur J Wildl Res. 59, 69-75.
Albers, U. (1990): Botanische Panseninhaltsanalyse bei Rehwild. Diplomarbeit der forstwissenschaftlichen Fakultät der Georg-August Universität Göttingen.
Anderson, R. (2000): Factors affecting maternal care in an income breeder, the European roe deer. Journal of Animal Ecology, Volume 69, Issue 4, pp 672-682.
Anderson, R., Duncan, P. & Linnel, J.D.C. (1998): The European Roe Deer: The Biology of Success. Scandinavian University Press, Oslo.
Anke, M., Dittrich, G., Groppel, B., Schäfer, U., Müller, R. & Hoppe, C. (2007): Zusammensetzung und Aufnahme von Winteräsung durch das Reh-, Muffel-, Dam- und Rotwild. Beiträge zur Jagd- und Wildforschung, 32, 379-398.
Anthony, R.G., Smith, N.S. (1974) Comparison of rumen and fecal analysis to describe deer diets. J Wildl Manage 38:535–540
Arlettaz, R., Patthey, P., Baltic, M., Leu, T.,2007, Schraub, M., Palme, R. & Jenni-Eiermann, S. (2007): Spreading free-riding snow sports represent a novel serious threat for wildlife. Proc. Biol. Sci. B 274: 1219–1224.
Arnold, W. (2013): Jahreszeitliche Anpassungen bei Wildwiederkäuern – wo steht das Rehwild? In: Landesjagdverband Bayern (Hrsg.). Schriftenreihe des Landesjagdverbandes Bayern e.V. Bd. 20, 13-22.
Arnold, W., Beiglböck, C., Burmester, M., Guschlbauer, M., Lengauer, A., Schröer, B., Wilkens, M. & Breves, G. (2015): Conrary seasonal changes of rates of nutrient uptake, organ mass, and voluntary food intake in red deer (Cervus elaphus). American Journal of Physiology, 309, 3, DOI:10.1152/ajpregu.00084.2015
Argunow, A.V., Stepamova, V.V. (2011): Diet Structure of the Siberian Roe Deer in Yakutia. Russian Journal of Ecology, Vol.42, No. 2, 161-164.
Ashley, N.T., Barboza, P.S., Macbeth, B.J., Janz, D.M., Cattet, M.R.L., Booth, R.K. & Wasser, S.K. (2001): Glucocorticosteroid concentrations in feces and hair of captive caribou and reindeer following adrenocorticotropic hormone challenge. Genereal and Comperative Endocrinology 172, 382-391.
Badeck, F.W., Bondeau, A., Bottcher, K., Doktor, D., Lucht, W., Schaber, J. & Sitch, S. (2004): Responses of spring phenology to climate change. New Phytol 162 (2), 295-309.

5 Literatur
164
Barancekova, M., Krojerova-Prokesova, J., Sustr, P. & Heurich, M. (2010): Annual changes in roe deer (Capreolus capreolus L.) diet in the Bohemian Forest, Czech Republic / Germany. Eur. J. Wildl. Res. 56, 327-333.
Barta, T. (2012): The food selection of roe deer on plain habitats. University of Debrecen, PHD school of animal husbandry.
Barth, D. (1980): Untersuchungen über das jahreszyklische Verhalten der Blutspiegel von Thyroxin und Harnstoff beim Reh. MSD Research Laboratories Sharp & Dohme GmbH, D-8201.
Bauer, J. (2007) Rehwildfütterung und Waldschutz. In: Reddemann, J. (Hrsg.): Tierschutz in der Jagd. Schriftenreihe des Landesjagdverbandes Bayern e.V. Bd. 16, 67-71.
Baumgärtl, T. (2012): Heu und Silage in der Wildwiederkäuferfütterung. Fachtagung Landwirtschaftliche Wildhaltung. Tll, Ref. 530.
Behrend A. (1999): Kinetik des Ingestaflusses bei Rehen (Capreolus capreolus) und Mufflons (Ovis ammon musimon) im saisonalen Verlauf, Dissertation an Mathematisch-Naturwissenschaftlichen Fakultät Ι der Humboldt-Universität zu Berlin.
Berg, F.C. v. (1978): Zum Raum-Zeit-System des Rehes. Allg. Forstzeitschr. 3; 48-50. Brüggemann, J., Giesecke, D., Walser-Kärst, K. (1967): Mikroorganismen im Pansen
von Rothirsch und Reh. Zeitschrift für Tierphysiologie, Tierernährung und Futtermittelkunde. 23, 143-151.
Brüggemann, J., Giesecke, D., Walser-Kärst, K. (1968): Methods for studying microbial digestion in ruminants post mortem with special reference to wild specis. Journal of Wildlife Management, 32,1, 198-207.
Brüggemann, J., Giesecke, D. (1968): Milchsäuregehalt im Pansen und mikrobieller Abbau in vito. Zentralblatt für Veterinarmedizin 15/5, 470-476.
Bobek, B., Grodzinski, W., Weiner, J. (1974): Studies on Produktivity of the Roe Deer Population in Poland. Proc. Xith Int. Con. Game Biologist, 115-123
Bubenik, A. B., Lochman J. (1956): Futterverbrauch und Tagesrhythmus der Futteraufnahme bei Reh- und Rotwild, Zeitschrift für Jagdwissenschaften, Vol. 2. No.1, 112-118.
Bubenik, A. B. (1984): Ernährung, Verhalten und Umwelt des Schalenwildes. BLV Ver-lag, München.
Buchgraber K. (2005): Nahrungsangebot für Rehwild auf Grünland- und Ackerflächen, Rehwild in der Kulturlandschaft, 11. Österreichische Jägertagung, 27-32.
Caspers, S. (2013): Vegetationsökologische Untersuchungen zur Habitatqualität im E+E-Vorhaben „ Wisente im Rothaargebirge“, Diplomarbeit, Universität Bonn.

5 Literatur
165
Cederlund, G., Nyström A. (1981): Seasonal differences between moose and roe deer in ability to digest browse, Holarctic Ecology, Vol. 4/1, 59-65.
Cederlund, G. (1983): Home Range Dynamics and Habitat Selection by Roe Deer in a Boreal Area in Central Sweden, Acta Theriologica, Vol 28/20, 443-460.
Chen, Z., Bernand, H.U. (2006): Classification of papillomaviruses based on 189 PV types and proposal of taxonomic amendments. Virolog, Volume 401, Issue 1, pp 70-79.
Corlatti, L. (2011): Climatic cues and glucocorticoids in a free-ranging riparian popula-tion of red deer. Folia Zool. – 60(2): 176-180.
Corlatti, L. (2014): Hormones, parasites and male mating tactics in Alpine chamois: identifying the mechanisms of life history trade-offs. Elsevier, Animal Behaviour, 1-10.
Cornelis, J., Casaer, J & Hermy, M. (1999): Impact of season, habitat and research techniques on diet composition of roe deer (Capreolus capreolus): a review. J. Zool. London 248, 195-297.
Clauss, M. (2010): Artgerechte Fütterung der Wildtiere – was ist fachlich vertretbar? 16. Österreichische Jägertagung.
Dalmau, A., Ferret, A., Chacon, G. & Manteca, X. (2007): Seasonal changes in fecal cortisol metabolites in Pyrenean chamois. Journal of Wildlife Management, 71, 190-194
Dehnhard, M. Clauss, M. (2001): Noninvasive Monitoring of Adrenocortical Activity in Roe Deer by Measurement of Fecal Cortisol Metabolites. General and Comparative Endocrinology, Volume 123, Issue 1, pp 111-120.
Deipenbrock, P.-H. (1985): Morphological studies relating to growth and condition of roe deer in a game management community of Northern Rhineland-Palatinate, F.R Germany. AGRIS.
Dennis, S.M., Arambel, M.J., Bartley, E.E., Dayton, A.D. (1983): Effect of energy concentration and source of nitrogen on numbers and types of rumen protozoa. Journal of Dairy Science, 66, 1248-1254
Deutz, A. (2009): Untersuchungen zum Auftreten von Nierenveränderungen beim gefüttertem und ungefüttertem Rehwild. Gesellschaft für Wildtier und Lebensraum. St. Lambrecht, Mödling.
Deutz, A., Gasteiner, J. & Gressmann, G. (2009): Fütterung von Reh- und Rotwild. Ein Praxisratgeber. Leopold Stocker-Verlag, Graz.
Diezel, C.E. (1921): Erfahrungen aus dem Gebiete der Niederenjagd. Franckh’sche Verlagshandlung, Stuttgart.
Dissen, J., Hartfiel, W. (1985): Beobachtungen zum Äsungsverhalten sowie Untersuchungen zur Nährstoffverdaulichkeit von Rehwild. Z. Jagdwiss. 31, 83-91.

5 Literatur
166
Ditchkoff, S.S. (2000): A decade since ‘diversification of ruminants’: has our knowledge improved? Oecologia 125, 82–84
Dittrich, G., Groppel, B. (1980): Vergleichende Untersuchungen des Nährstoffgehaltes im Pansen von Reh-, Rot-, Dam- und Muffelwild. Beitrag Jagd- und Wildforsch, XI, 39–46
Djordjevic, N., Popovic, Z., Grubic, G. (2006): Chemical composition of the rumen contents in roe deer as potential quality indicator of their feeding. Journal of Agricultural Sciences, Volume 51, Issue 2, pages: 133-140.
Drescher-Kaden, U. (1976): Untersuchungen am Verdauungstrakt von Reh, Damhirsch und Mufflon. Mitteilung 1: Gewichtserhebungen und Kapazitätsmessungen am Verdauungstrakt, insbesondere am Pansen-Haubenraum von Reh, Damhirsch und Mufflon. Z. Jagdwiss. 22, 184-190.
Drescher-Kaden, U. (1984): Stellung des Rehs im System der Wiederkäuer. Rehwild-biologie. Arbeitstagung am 10.-12. März 1984 im Nationalpark Bayerischer Wald, 29-35.
Drescher-Kaden, U., Seifelnasr E.A. (1976): Untersuchungen am Verdauungstrakt von Reh, Damhirsch und Mufflon. Mitteilungen 2: Rohnährstoffe im Panseninhalt von Reh, Damhirsch und Mufflon. Z. Jagdwiss. 23, 6-11.
Drescher-Kaden, U., Seifelnasr E.A. (1977a): Untersuchungen am Verdauungstrakt von Reh, Damhirsch und Mufflon. Mitteilungen 3: Mikroorganismen im Pansen von Reh, Damhirsch und Mufflon. Z. Jagdwiss. 23, 64-89.
Drescher-Kaden, U., Seifelnasr E.A. (1977b): Untersuchungen am Verdauungstrakt von Reh, Damhirsch und Mufflon. Mitteilungen 4: Fermentationsprodukte im Pansen von Reh, Damhirsch und Mufflon. Z. Jagdwiss. 23, 126-131.
Drozdz, A., Osiecki, A. (1973): Intake and Digestibility of Natural Feeds by Roe-Deer. Acta Theriologica, 18, 81-91.
Drozdz, A. (1979): Seasonal intake and digestibility of natural foods by roe deer. Acta Theriologica, 24, 137-170.
Duncan, P, Tixier, H, Hofmann, R.R., Lechner-Doll, M. (1999): Feeding strategies and the physiology of digestion in roe deer. In: Anders, R, Duncan, P. & Linnell, J.D.C (Eds.) The European Roe Deer: The Biology of Success. Scandinavian University Press, Oslo.
Eisfeld, D. (1974a): Der Eiweiß- und Energiebedarf des Rehes (C. capreolus L.), diskutiert an Hand von Laborversuchen. Ges. f. Ökologie, 129-139.
Eisfeld, D. (1974b): Der Proteinbedarf des Rehes (Capreolus capreolus L.) zur Erhaltung. Z. Jagdwiss. 20, 43-48.
Ellenberg, H. (1978): Zur Populationsökologie des Rehes in Mitteleuropa. Spixiana, Supplement 2, ISSN 0343-5512.

5 Literatur
167
Enghofer, P. (1990): Über die Kondition beim Rehwild (Capreolus capreolusL.), Diplomarbeit im Bereich Wildbiologie und Jagdkunde, TU München
Enzinger, W., Hartfiel W. (1989): Auswirkungen gesteigerter Energie- und Proteingehalte des Futters auf Fermentationsprodukte, Fauna und Schleimhaut des Pansens von Wildwiederkäuern (Damhirsch / Reh) im Vergleich zu Hauswiederkäuern (Schaf / Ziege), Z. Jagdwiss. 44, 201-220.
Esser, W.(1954): Beitrag zur Untersuchung der Äsung des Rehwildes. Institut für Tierernährungslehre der Landw. Hochschule Hohenheim.
Feustel, G. (1967): Vergleichende Untersuchungen am Verdauungstrakt von Rothirsch (Cervus elaphus) und Reh (Capreolus capreolus) post mortem unter besonderern berücksichtigung der Gerüst-Kohlehydrate und des Ligningehaltes der Ingesta- Diss. Med. vet. München
Fielitz, U. Albers, U. (1996): Nahrungsspektrum von Rehen aus dem Bayerischen Wald. Z. Jagdwiss. 42, 195-202.
Fischer, K. (1998): Ernährungsphysiologie heimischer Schalenwildarten im Jahresverlauf. Fachseminar 6.2.1998 zu Fütterung von Schalenwild. Hessisches Ministerium des Inneren und für Landwirtschaft, Forsten und Naturschutz, 7-17.
Gentsch, R. (2014): Raumverhalten von Rehwild (Capreolus capreolus) in einem Privatjagdrevier des Bayerischen Waldes Homeranges, Tagesablauf und Habitatnutzung. BSc Thesis, LS Tierökologie, Arbeitsgruppe Wildbiologie und Wildtiermanagement.
Georgii, B. (1973): Untersuchungen über Stress und Fortpflanzung des Rehes (Capreolus capreolus L.), Diplomarbeit im Bereich Biologie, Universität Tübingen.
Georgii, B., Schroeder J., Schroeder W., (1991): A simulation model for roe deer management in Bavarian Forest National Park, 207-210.
Giesecke, D. (1966): Gaschromatographische Bestimmung flüchtiger Fettsäuren und ihre Produktion im Pansen. Blackwell Verlag, Vol. 22, Issue 1-5, 354-364.
Giger-Reverdin, S. (1998): Interest and limits of the HFT method to evalute the energy value of compound feeds for small ruminants. Laboratoire de Nutrition et Alimentation (INRA) de l`INA-PG. 39-42.
Golte-Bechtle, M., Aichele, D., Spohn, M. & Spohn, R. (2008): Was blüht denn da?, 58. Auflage, Verlag: Franckh-Kosmos
Hartfiel, W., Pheiffer, J., Dissen, J. (1985): Energetische Untersuchungen an Reh und Schaf mit Hilfe der quantitativen Thermographie zur Beurteilung des Energiebedarfs im Winter. Z. Jagdwiss. 31, 34-41.
Helm, G. (2015): Ernteschock und Verbiss. Volle Silos, leere Mägen. Jagd in Bayern, 10, 16-17.

5 Literatur
168
Herwig, N. (2010): Isotopenanalytik zur Bestimmung des Einflusses der Ernährung auf die Isotopenzusammensetzung in Rinderproben, BAM-Dissertationsreihe, Band 62, Berlin.
Herzog, S. (2013): Das Reh, der Wald und der Förster: Rehwild im Lichte unterschiedlicher Nutzungsinteressen. In: Landesjagdverband Bayern (Hrsg.). Schriftenreihe des Landesjagdverbandes Bayern e.V. Bd. 20, 13-22.
Hewison, A. (1996): Behavioural ecology of Siberian and European roe deer. Chapman & Halls´s Wildlife, Ecology and Behaviour Series, focuses on studies of both European and Siberian roe deer.
Hofmann, B., Barth, D., Gimenez, T. (1976): Testosteronkonzentration im peripheren Blut beim Rehbock. Zeitschrift für Jagdwissenschaft, Volume 22, Issue 3, pp 134-148.
Hofmann, A. (1972): Das jahreszeitliche Verteilungmuster und der Äsungsdruck von Alpensteinbock, Gemse, Rothirsch und Reh in einem begrenzten Gebiet im Oberengadin. Arbeitsgruppe für Wildforschung, Universität Zürich.
Hofmann R.R. (1978): Die Ernährung des Rehwildes im Jahresablauf nach dem Modell Wichselboden, Wildbiologische Information für den Jäger 2, Enke Stuttgart, 121-136.
Hofmann, R. R. (1989): Evolutionary steps of ecophysiological adaption and diversifi-cation of ruments: a comparative view of their digestive system. Oecologia, 78, 443-457.
Hofmann, RR. (1982): Die Stellung der europäischen Wildwiederkauer im System der Äsungstypen. (In: Hofmann, R.R.(Hrsg.): Wildbiologische Informationen für den Jäger, Bd. I, 9-19.
Hofmann, R.R. (2007): Wildtiere in Bildern zur vergleichenden Anatomie. M& H. Schaper, Hannover.
Hofmann, R.R, Geiger G. König R. (1976): Vergleichend-anatomische Untersuchungen an der Vormagenschleimhaut von Rehwild (Capreolus capreolus) und Rotwild (Cervus elaphus), 2. Säugetierkunde 41, 167-193.
Hofmann, R.R., Herzog, A. (1980): Jahreszeitliche Ernährungssituation beim Reh und Rotwild. Jagd in Tirol 9, 7-13
Hofmann, R.R. & Kirsten, N. (1982): Die Herbstmast-Simulation. Untersuchungs-ergebnisse und kritische Analyse eines praxisorientierten AKWJ-Projektes zur Problematik der Schalenwildfütterung. Schriften des Arbeitskreises für Wildbiologie und Jagdwissenschaft an der Justus-Liebig-Universität Gießen, H. 9, Ferdinand Enke Verlag, Stuttgart.
Holand, O. (1993): In vitro dry matter disappearance using roe deer inocula from summer and winter. Agricultural University of Norway, Department of Biology and Nature Conservation. P.O.Box 14, N-1432.

5 Literatur
169
Holisova, V., Kozena, I. & Obrtel, R. (1984): The summer diet of field roe bucks (Capreolus capreolus) in southern Moravia. Folia Zool. 33: 193±208.
Huber, S., Palme, R., Arnold, W. (2003): Effects of season, sex, and sample collection on concentration of fecal cortisol metabolites in red deer. Academic Press, General and Comparative Endocrinology 130, 48-54.
Hummel, J., Südekum, K-H., Streich, W.J. & Clauss, M. (2006): Forage fermentation patterns and their implications for herbivore ingesta retention times. Functional Ecology, 20, 989-1002.
Imai, S., Abe, M., Ogimoto, K. (1981): Ciliate Protozoa from the Rumen of the Japanese Serow, Capricornis crispus (Temminck), Jpn. J. Vet. Sci., 43: 359-367
Jackson, J. (1980): The annual diet of the roe deer (Capreolus capreolus) in the New Forest, Hampshire, as determined by rumen content analysis. J. Zool. Lond. 192: 71-83
Jäger, E. (2011): Rothmaler - Exkursionsflora von Deutschland. Bd. 2: Gefäßpflanzen: Grundband, 20. Auflage, Verlag: Spektrum Akademischer Verlag
Kaluzinski, J. (1982): Composition of the food on roe deer living in fields and the effects of their feeding on plant production. Acta Theriol 27:457–470
Kamler, J. (1999): Infusorial concentration in rumen fluid of red deer, fallow deer, roe deer and moufflon. Academy of Sciences of the Czech Republic, Brno. 68: 247-252.
Kamler, J., Dvorak, J. & Kamlerova, K. (2002): Morphological variability of stomach and concentration of rumen infusorial in red deer, fallow deer, roe deer and mouflon. Folia venatoria, 32, 15-24.
Kamler, J., Kamlerova, K. (2003): Differences in relative volume and weight of stomach among four free living ruminants, Acta Veterinaria Brno, 72/1.
Kamler, J., Homolka, M. (2005): Faecal nitrogen; a potential indicator of red and roe deer diet qulity in forest habitats. Folia Zool. – 54(1-2): 89-98.
Kamler, J., Homolka, M. (2011): Needles in faeces: an index of quality of wild ungulate winter diet, Folia Zool. 60/1, 63-69.
Kamler, J., Homolka,M., Heroldova M., Literákova, P. (2011): Feeding Strategy of Wild Herbivores in Habitats of Limited Food Resources, Wildl. Biol. Pract., 7(1): 46-55
Kamphues, J., Coenen, M., Iben, C. Kienzle, E., Pallauf, J., Simon, O., Wanner, M., & Zentek, J. ( 2008): Supplemente zu Vorlesung und Übung in der Tierernährung, M&H. Schaper, Hannover.
Keay, J.M (2006): fecal glucocorticoids and theit metabolites as indicators of stress in various mamalian species. a literature review. Journal of Zoo and Wildlife Medicine 37(3), 234-244.

5 Literatur
170
Kioroglanidis, J. (1981): Nahrungswahl und Überlegungen zur Nahrungsnische bei Reh, Rothirsch und Gemse. Dissertation durch die forstwirtschaftliche Fakultät der LMU München. 83/153.
Kirchgessner, M. (1997): Tierernährung, Leitfaden für Studium, Praxis und Beratung. Verlagsunion Agrar.
Kirchgessner, M., Roth, F.X., Schwarz, F.J. & Stangl, G.I. (2008): Tierernährung. DLG Verlag, Frankfurt a. Main.
Klansek, E., Vavra, I., Onderscheka, K. (1995): die Äsungszusammensetzung des Alpensteinwildes (Capra i. ibex L.) in Abhängigkeit von Jahreszeit, Alter und Äsungsangebot in Graubünden. Z. Jagdwiss 41, 171-181
Klansek, E. (2004): Nahrungszusammensetzung von Rot-, Reh-, und Gamswild im Berggebiet, Tagung für die Jägerschaft, 16-17 Februar 2004.
Klansek, E. (2005): Äsungswahl des Rehwildes in unterschiedlichen Lebensräumen, 11. Österreichische Jägertagung, 15-16 Februar 2005.
Klein, D. (1972): Factors affecting growth and body size of roe deer. The Journal of Wildlife Management. Vol. 36, No. 1.
Klötzli, F. (1965): Qualität und Quantität der Rehäsung. Promotionsarbeit der eidgenössischen technischen Hochschule in Zürich. Prom.-Nr 3516.
Konjevic, D. (2011): Non-invasive monitoring of adrenocortical activity in free-ranging fallow deer. Eur. J. Wildl. Res., 57:77-81.
Kreuzer, M. Kirchgessner, M. (1986a): The role of rumen ciliate protozoa in digestion of nutrients, N retention and partition of faecal and urinary nitrogen when feeding different amounts and sources of starch to wether. Journal of Animal Physiology and Animal Nutrition 56: 217-231
Kreuzer, M., Kirchgessner, M. (1986b): Die Bedeutung der Protozoen im Pansen yon Hammeln für Verdaulichkeit, N-Bilanz und N-Fraktionen im Kot und Harn bei unterschiedlichen Gehalten und Arten von Stärke in der Ration. Z. Tierphysiol. Tiererrähr. Futtermittelk. 56, 217-23
Latham, J. Staines, B.W., Gorman, M.L.(1999): Comparative feeding ecology of red and roe deer in Scottish plantation forests. Journal of Zoology, Volume 247, Issue 3, pages 409-418.
Lechner – Doll, M., Clauss, M. (2006): Differences in selective reticulo-ruminal particle retention as a key factor in ruminant diversification. Journal Oecologia, Vol.29, Issue 3, pp 321-327.
Lohse, K. (2000): Beiträge zur Ernährungsforschung bei Wiederkäuern. Dissertation durch das Institut für Tierernährung der Tierärztlichen Hochschule Hannover.
Lüder, R. (2006): Grundkurs Pflanzenbestimmung. Eine Praxisanleitung für Anfänger und Fortgeschrittene, 7. Auflage, Verlag: Quelle & Meyer

5 Literatur
171
Mattiello, S., Ardunio, D., Tosi, M.V., Carenzi, C. (2005): Survey on housing, management and welfare of dairy cattle in tie-stalls in western Italian Alps. Acta Agriculturae Scandinavica, Section A, Animal Science 55/1, 31-39.
Mcinnis, M., Vavra, M., Grueger, W. (1983): A comparison of four methods used to determine the diet of large herbivores. Journal of range management 36/3, 302-306
Menke K.H. & Steingass H. (1988): Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res. Dev. 28., 7-55
Menzel, A., Sparks, T.H., Estrella, N., Koch, E., Aasa, A., Ahas, R., Al-Kübler, K., Bissollo, P., Braslavska, O., Bride, A., Chmielewski, F.M., Crepensek, Z., Curnel, Y., Dahl, A., Defila, C., Donnelly, A., Filella, Y., Jatczak, K., Mage, F., Mestre, A., Nordli, O., Penuelas, J., Pirinen, P., Remisova, V., Scheifinger, H., Striz, M., Sunsik, A., Van Vliet, A., Wielgolaski, F.-E., Zach, S., & Zust, A. (2006):European phenological response to climate changematches the warming pattern. Global Change Biology (2006) 12, 1969–1976, doi: 10.1111/j.1365-2486.2006.01193.x
Meyer C. (1988): Nahrungswahl von Rehen in einem Hochgebirgsareal, Diplomarbeit im Bereich Wildbiologie und Jagdkunde, LMU München.
Millspaugh, J.J., Woods RJ, Hunt KE, Raedeke KJ, Brundige GC, Washburn BE, Wasser SK (2001): Using fecal glucocorticoid assays to study the physiological stress response of elk. Wildlife Soc. B. 29, 899–907.
Millspaugh, J.J. (2004): Use of fecal glucocorticoid metabolite measures in conservation biology research: considerations for application and interpretation. Elsevier, General and Comparative Endocrinology 138, 189-199.
Millspaugh, J.J., Washburn, B.E. (2004): Use of fecal glucocortocoid metabolite measures in conservation biology research: condiderations for application and interpretation. USDA National Wildlife Research Center- Staff Publications, Paper 395
Moen, A. N. (1973): Wildlife ecology: an analytical approach. W. H. Freeman and Co., San Francisco
Möhring, G. (1963): Zur Beerennahrung des Rehes. Waldhyg. 5, 68-69. Navarro-Castilla, A., Mata, C., Ruiz-Capillas, P., Palme, R., Malo, J.E., Barja, I. (2014):
Are motorways potential stressors of road-side wood mice populations (Apodemus sylvaticus). PLoS ONE 9, e91942
Norris, J.J. (1943): Botanical analysis of stomach contents as a method of determining forage consumption of range sheep. Ecology 24: 244-251

5 Literatur
172
Onderscheka, K. (1999): Das Rehwild – seine Ernährung und Fütterung. In. Reddemann, J. (Hrsg.) Rehwild in der Kulturlandschaft. Schriftenreihe des Landesjagdverbandes Bayern. e.V. 7, 37-61.
Onderdcheka, K. & Jordan H.R. (1976): Die Bodenkultur – Journal für Landwirtschaftliche Forschung. Österreichischer Agrarverlag Wien. 27 Band, Heft 2.
Osborn, R., Jenks, J., Jensen, W. (1997): Diet of north dakota elk determined from rumen and fecal analyses. Wildlife and Fisheries Sciences, Texas A&M University: 237-248
Oslage, H.J. & Strothmann, A. (1988): Zum Energie- und Proteinbedarf von Rehwild, Z. Jagdwiss. 34, 164-181.
Papageorgiou, N., Neophytou, C., Spais, A. & Vavalekas, C. (1981): Food preferences, and protein and energy requirements for maintenance of roe deer. J. Wild. Manag. 45, 728-733.
Perzanowski, K. (1978): The Effect of Winter Food Competition on Roe Deer Energy Budget. Acta Theriologica, 23, 451-467.
Piatrowski, B. & Nagel, S. (1975): Ergebnisse über die Kau- und Wiederkauaktivität von Kühen bei Rationen mit Getreidestroh verschiedener physikalischer Form und nach chemischer Behandlung mit Natronlauge. Archiv für Tierernährung 25/8: 575-582
Popovic, Z., Dordevic, N., Dordevic, M., Grubic, G. & Stojanovic, B. (2009): Estimation of the Quality of the Natution of Roe Deer based on Chemical Composition of the Rumen Content. Acta Veterinaria (Beograd), Vol.59, No. 5-6, 653-663.
Prins, R.A. & Geelen, M. (1970): Rumen Characteristic of Red Deer, Fallow Deer, and Roe Deer. Allen Press, the Journal of Wildlife Management, Vol.35, No.4, 673-680.
Purser, D.B., Moir, R.J. (1959): Ruminal flora studies in the sheep. IX. The effect pf pH on the ciliate population of the rumen in vivo. Australian Journal of Agricultural Research, 10(4), 555-564.
Raesfeld, F., Neuhaus, A.H., Schaich K. (1978): Das Rehwild – Verlag Parey, Hamburg
Ramin, L. (1975): Untersuchung über die Beäsung verschiedener Grünland- und Ackerplanzen durch Rehwild. 1001/Diss.13 1240.
Redjadj, C., Darmon, G., Maillard, D., Chevrier, T., Bastianelli, D., Verheyden, H., Loison, A. & Said, S. (2014): Intra- and Interspecific Differences in Diet Quality and Composition in large Herbivore Community. PLOS ONE, Vol.9, Issue 2.
Rehbinder, C., Ciszuk, P. (1985): Supplemental feeding of roe deer with late harvested hay. A pilot study. Rangifer 5/2: 6–14

5 Literatur
173
Rehnus, M., Wehrle, M., Palme, R. (2014): Mountain hares (Lepus timidus) and tour-ism: Stress events and reactions. J. Appl. Ecol. 51, 6-12
Reimoser, F. (2004): Äsungsverbesserung und Fütterung für Schalenwild in der mitteleuropäischen Kulturlandschaft, Grundsätzliche Aspekte aus wild-ökologischer Sicht unter besonderer Berücksichtigung der Wildschadensver-meidung im Wald. Tagung für die Jägerschaft, 16-17 Februar 2004.
Reimoser, F. (2005): Rehwild in der Kulturlandschaft - Biologie, Lebensraum, Hege, Bejagung. 11. Österreichische Jägertagung, 15-16 Februar 2005.
Reimoser, F. (2006): Begleitforschung zum Projekt Nationalparkjagden im Gasteinertal. Endbericht; Forschungsinstitut für Wildtierkunde und Ökologie, Veterinärmedizinische Universität.
Riney, T. (1955): Evaluating condition of free-ranging red deer (Cervus elaphus), with special reference to New Zealand. N.Z. J. Sci. Technol. 36, 429-463.
Ritz, J. Hofer, K., Hofer, E., Hackländer, K., Immekus, D., Codron, D., Clauss, M. (2013): Forestomach pH in hunted roe deer in relation to forestomach region, time of measurement and supplemental feeding and comparison among wild rumiant species. Eur J Wildl Res 59, 505-517.
Romero, L.M. (2002): Seasonal changes in plasma glucocorticoid concentrations in free-living vertebrates. Gen. Comp. Endocrinol. 128, 1–24
Romero, M.L., Meister, C.J., Cyr, N.E., Kenagy, G.J., Wingfield, J.C. (2007): Seasonal glucocorticoid responses to capture in wild free-living mammals. American Journal of Physiology - Regulatory, Integrative and Comparative Physiology, Vol. 294, No. 2, 614-622
Rowell-Schäfer, A. (2000): metabolic evidence of a rumen bypass of a ruminal escape of nutrients in roe deer. Elsevier, comparative biochemistry and physiology Part A128, 289-298.
Rusterholz, M., Turner, D.C. (1978): "Versuche über die" Nährstoffweisheit" beim Reh (Capreolus capreolus)." Rev. Suisse Zool. 85: 718-730
Santos, J.P.V., Fernandes-de-Mera J.G., Acevedo P., Boadella M., Fierro Y., Vicente J., Gortazar C. (2013): Optimizing the sampling effort to evaluate body condition in ungluates: A case study on red deer, Ecological Indicators.1472, 1-7.
Sauerwein, H. (2004): Establishing baseline values of parameters potentially indicative of chronic stress in red deer from different habitats in western Germany. European Journal of Wildlife Research. Volume 50, Issue 4, pp 168-172.
Schäfer, E. (1974): Hegen und Ansprechen von Rehwild. BLV-München Schmeil, O. & Fitschen, J. (2011): Die Flora Deutschlands und der angrenzenden
Länder, 95. Auflage, Verlag: Quelle &Meyer Schmid, M. 2013: Anwendung des Hohenheimer Futterwerttests & des Weender-
Verfahrens bei natürlicher Nahrung von Rehen (Capreolus capreolus). Bachelor

5 Literatur
174
Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München
Schmidl, C. (1996): Botanische und chemische Panseninhaltsanalyse, chemische Blutuntersuchung und Untersuchung der Schwermetallbelastund beim Rehwild in zwei unterschiedlichen Lebensräumen. Universität Wien.
Schmidt, M. (2004): Dispersal of vascular plants by game in northern Germany. Part 1: Roe deer and wild boar. European Journal of Forest Research, Volume 123, Issue 2, pp 167-176.
Schneider, H. 2012: Entwicklung eines Standardverfahrens zur Analyse der Qualität der Nahrung von Rehen (Capreolus capreolus). Bachelor Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München
Schröder, A. (2003): Ernährung und Management der Milchkuh vor dem Kalben. Fruchtbarkeitsseminare der RSH eG Rendsburg und Neumünster.
Sommer, A. (2004): Stoffwechsel der Wildwiederkäuer, Bundesanstalt für alpenländische Landwirtschaft Gumpenstein Tagung für die Jägerschaft.
Stahl, B. (2016): Die Verschiebung der phänologischen Phasen potentieller Nahrungsplfanzen des Rehs (Capreolus capreolus Linenaeus) während der seztzeit als Folge des Klimawandels und die damit verbundene zeitliche Änderung der Nahrungsquantität und –qualität. Masterarbeit an der Studienfakultät für Forstwissenschaft und Ressourcenmanagement der TU-München
Steingaß, H., Menke, K. H. (1986): Schätzungen des energetischen Futterwertes aus der in vitro mit Pansensaft bestimmten Gasbildung und der chemischen Analyse. 1. Mitteilungen: Untersuchungen zur Methode. Übers. Tierern. 14, 251-270.
Storm, D, Aubry, P., Hamann J.L. Said, S., Fritz, H., Stain-Andrieux C., Klein, F. (2008): Seaasonal variation in diet composition and similarity of sympatric red deer Cervus elaphus and roe deer Capreolus capreolus. Wildl. Biol. 14: 237-250
Stubbe, I. (1966): Growth and body size of the European roe deer. Der Zoologische Garten N.F. 33, 85-105.
Stubbe, C. (1997): Das Rehwild. Parey Buchverlag, Berlin. Tataruch, F., Onderscheka, K. (1996): Untersuchung zur Kondition des Steinwildes in
Graubünden. Blackwell Wissenschafts-Verlag, Berlin. Z.Jagdwiss. 42, 92-103. Tataruch, F., Onderscheka, K. (1993): Gehalt an Ammoniak und flüchtigen Fettsäuren
im Pansensaft von Rot-, Reh- und Gamswild. Veterinary medicine Austria, Wien. Tierärztl. Mschr. 80,9, 269-274.

5 Literatur
175
Thiel, D., Jenni-Eiermann, S., Palme, R., Jenni, L. (2011): Winter tourism increases stress hormone levels in endangered capercaillie (Tetrao urogallus). Ibis 153, 122–133
Tixier, H., Duncan, P. (1996): Are European roe deer browsers, a review of variations in the composition of their diets. Revue d´ecologie. 51, 3-17.
Toïgo, C., Gaillard, J.M., Van Laere, G., Hewison, M., Morellet, N. (2006): How does environmental variation influence body mass, body size, and body condition? Roe deer as a case study. Ecography, 29, 3: 301–308
Touma, C., Palme, R. (2005): Measuring Fecal Glucocorticoid Metabolites in Mammels and Birds: The Importance of Validation. Ann.N.Y. Acad. Sci. 1046: 54-74.
Turner, A.I., Canny, B.J., Hobbs, R.J., Bond, J.D., Clarke, I.J., Tilbrook, A.J. (2002): Influence of sex and gonadal status of sheep on cortisol secretion in response to ACTH and on cortisol and LH secretion in response to stress: importance of different stressors. J. Endocrinol. 173, 113-122
Ueckermann, E. (1986): Die Fütterung des Schalenwildes. Verlag Paul Parey, Ham-burg.
Van Lier, E., Perez-Clariget, R., Forstberg, M. (2007): Sex differences in cortisol secretion after administration of an ACTH analogue in sheep during the breeding and non-breeding season. Elsevier, Animal Reproduction Science, Volume 79, Issues 1-2, Pages 81-92
Van Soest, P.J. (1982): Nutritional Ecology oft he Ruminant. Cornell University Press Veith, S. (1992): Nahrungswahl von Rehen in einem Revier der Zentralalpen,
Diplomarbeit im Bereich Wildtierforschung und Wildtiermanagement, LMU München.
Warren, R.J., Kirkpatrick, R.L. (1982): Evaluating nutritional status of white-tailed deer using fat indices, Proc. Annual Confernece SEAFWA, 463-472.
Weiner, J. (1977): Energy metabolism of roe deer. Acta Theriologica, 22, 3.24. Weilnböck, G. (2013): Zur Stressbelastung des Rehwildes (Capreolus capreolus) beim
Fang mit der Kastenfalle. Inaugural-Dissertation zur Erlangung der tiermedizini-schen Doktorwürde der Tierärztlichen Fakultät der LMU München
Wotschikowsky U., Schwab G., (1994): Das Rehprojekt Hahnebaum, Schlußbericht, Wildbiologische Gesellschaft München e.V.
Zweifel-Schielly, B. (2005): Spatial and nutritional ecology of GPS-collared red deer in an alpine region: the role of forage availability and quality. Diss., Naturwissen-schaften, Eidgenössische Technische Hochschule ETH Zürich, Nr. 16055, 2005.
Zwijac-Kozica, T. (2012): Concentration of fecal cortisol metabolites in chamois in rela-tion to tourist pressure in Tatra National Park (South Portland). Polish Academy of Sciences, Poland.

6 Abbildungsverzeichnis
176
6 Abbildungsverzeichnis
Abbildung 1: Übersichtskarte Studienflächen (links rot: Waldflächen, rechts rot: Landflächen), Quelle: Bayern Maps, bearb. Scheingraber ............................... 23
Abbildung 2: Untersuchungsgebiet FO, GIS Karte, Bayern Atlas, Quelle: bearb. Scheingraber. .................................................................................................... 24
Abbildung 3: Untersuchungsgebiet LA, GIS Karte, Bayern Atlas, Quelle: bearb. Scheingraber. .................................................................................................... 25
Abbildung 4: Aufbau Magensystem Rehwild, Quelle: Heckel 2013. ............................. 28Abbildung 5: Flussdiagramm der Abläufe der Probenahme und Laboranalysen. ........ 31Abbildung 6: Probenverteilung nach Habitaten, Altersklassen und Geschlecht, W =
Weiblich, M = Männlich. .................................................................................... 33Abbildung 7: Trennung der Bestandteile durch Zentrifugation, rechts: Zentrifugenröhren
nach der Zentrifugation, links: Überstand/Pansensaft in Weithalsgefäß, Raufutteranteil in Petrischale ausgestrichen, Quelle: Scheingraber ................. 34
Abbildung 8: Panseninhalte Rohmasse (links) und in Petrischalen gefriergetrocknet, beim Rückwiegen und vor dem Mahlen (rechts) , Quelle: Scheingraber .......... 34
Abbildung 9: Weender Analyse und VanSoest Analyse grafische Darstellung, Quelle: verändert nach Kirchgeßner et al. 2008. ........................................................... 41
Abbildung 10: Auftrennung der Flüchtigen Fettsäuren im Gaschromatograph, Probe Nr. 243, Quelle: Geritschke, Scheingraber ............................................................. 46
Abbildung 11: Energieformen und Energiekaskade ..................................................... 52Abbildung 12: Spendertier mit Pansenfistel (links), Woulff’sche Flasche mit Pansensaft
(mittig), Thermobehälter mit Pansensaft und CO2 Begasung (rechts), Quelle: Scheingraber ..................................................................................................... 53
Abbildung 13: Reagenzien zur Herstellung der Mediumlösung (links), Pansensaft-Filtration (mittig), Zugabe des Pansensaftes in Woulff’scher Flasche (rechts), Quelle: Scheingraber ........................................................................................ 53
Abbildung 14: Spendertier für HFT-Verfahren mit Rehpansensaft im Warmhalteprozess, Quelle: Kuwert, Scheingraber ........................................... 56
Abbildung 15: Bestimmung des Pansenvolumens, Abfolge, Quelle: Heckel, Scheingraber ..................................................................................................... 58
Abbildung 16: Median des umsetzbaren Energiegehaltes (ME) der Rehäsung [MJ/kg TS] beider Habitate (n = 220, p < 0,001) .......................................................... 67
Abbildung 17: Energiewerte (ME) nach Monaten in Wald und Land, n = 220, Mediane mit 95 % Konfidenzintervall. .............................................................................. 68
Abbildung 18: Korrelationsdarstellung der Energieergebnisse, gemessen mit Reh- und Hammelpansensaft der gleichen Proben, R2 = 0,43. ........................................ 69

6 Abbildungsverzeichnis
177
Abbildung 19: Gegenüberstellung der Energiemessung mit Reh- und Hammelpansensaft, n= 18, Median mit 95 % Konfidenzintervall, Energiewerte im Januar unterscheiden sich mit p = 0,01 signifikant. ..................................... 70
Abbildung 20: Rohnährstoffgehalte in der Trockensubstanz (TS) nach Habitaten, Mediane mit 95 % Konfidenzintervall, n = 220, Unterschiede Wald-Land: XA = Rohasche p = 0,001, XF = Rohfaser p < 0,001, XL = Rohfett nach HCl-Aufschluss, XP = Rohprotein, NfE = Stickstofffreien-Extraktstoffe p = 0,035. .. 72
Abbildung 21: Rohnährstoffgehalte nach Monaten getrennt nach Wald und Land, (n = 220, Mediane mit 95 % Konfidenzintervall, XA = Rohasche, XF = Rohfaser, XL = Rohfett nach HCl-Aufschluss, XP = Rohprotein, NfE = Stickstofffreien-Extraktstoffe). .................................................................................................... 73
Abbildung 22: Faseranteile an der Trockensubstanz (TS) nach Habitaten, Mediane mit 95% Konfidenzintervall, n = 220. Zellulose p = 0,042; Lignin p < 0,001. .......... 75
Abbildung 23: Fasergehalte nach Monaten je Untersuchungsgebiet, n = 220, Mediane mit 95 % Konfidenzintervall. .............................................................................. 76
Abbildung 24: Anteil der Stickstofffreien- Extraktstoffe (NfE) in Kombination mit den Werten der Hemizellulose nach Monaten aufgeteilt in Wald und Land, (n = 220, Mediane mit 95 % Konfidenzintervall). .............................................................. 76
Abbildung 25: Verdaulichkeit (dOS) der organischen Substanz (DOS) nach Habitaten, n = 220, Mittelwert mit 95 % Konfidenzintervall, P < 0,001. .............................. 78
Abbildung 26: Verdaulichkeit (dOS) der organischen Substanz (DOS) nach Monaten und Gebieten, n = 220, Mittelwert mit 95 % Konfidenzintervall. ...................... 78
Abbildung 27: Korrelation zwischen Rohfasergehalt und Verdaulichkeit, getrennt nach Wald- und Landhabitat, n = 220, R2 = 0,76 (Wald, links), R2 = 0,52 (Land, rechts). .............................................................................................................. 79
Abbildung 28: Links: Gesamtkonzentration an FFS, n = 220, p = 0,005. Rechts: Unterschiede zwischen den Habitaten in der Konzentration von Essigsäure, Propionsäure p < 0,001, Buttersäure p = 0,002 und Valeriansäure, n = 220. .. 80
Abbildung 29: Konzentration an flüchtigen Fettsäuren im Jahresverlauf nach Habitaten, n = 220, Mittelwerte mit 95 % Konfidenzintervallen .......................................... 81
Abbildung 30: Anteile der vier wichtigsten flüchtigen Fettsäuren nach Monaten und Habitattypen, Mediane mit 95 % Konfidenzintervall, n = 220 ........................... 82
Abbildung 31: Essig-/Propionsäure-Verhältnisses (C2/C3) nach Monaten und Habitat 83Abbildung 32: NH3-Gesamtkonzentration in Wald und Land, n = 220, p < 0,001. ....... 84Abbildung 33: Ammoniak (NH3) Konzentration im Pansensaft im Jahresverlauf nach
Habitaten, n = 220, Median mit 95 % Konfidenzintervallen. ............................. 85Abbildung 34: Korrelation zwischen Ammoniakwerte und Rohproteingehalte [XP], p <
0,001, r = 0,495. ................................................................................................ 85Abbildung 35: Laktatgesamtkonzentration p = 0,005 (links) und Laktatkonzentration
nach Monaten und Habitaten, n = 220, Mediane mit 95 % Konfidenzintervall . 86Abbildung 36: pH-Wert-Verlauf nach Monaten und Habitat, n = 220, Mittelwerte mit 95
% Konfidenzintervallen. .................................................................................... 87

6 Abbildungsverzeichnis
178
Abbildung 37: Korrelation zwischen pH-Wert und Konzentration der flüchtigen Fettsäuren im Pansensaft. ................................................................................ 88
Abbildung 38: Nahrungsbestandteile nach Habitattyp, Mittelwerte mit 95 % Konfidenzintervall, n = 220. ............................................................................... 90
Abbildung 39: Nahrungskategorien im Frühling, Mittelwerte mit 95 % Konfidenzintervall, n = 220. ............................................................................... 91
Abbildung 40: Nahrungskategorien im Sommer, Mittelwerte mit 95 % Konfidenzintervall, n = 220. ............................................................................... 92
Abbildung 41: Nahrungskategorien im Herbst, Mittelwerte mit 95 % Konfidenzintervall, n = 220. ............................................................................................................. 93
Abbildung 42: Nahrungskategorien im Winter, Mittelwerte mit 95 % Konfidenzintervall, n = 220. ............................................................................................................. 94
Abbildung 43: Nahrungsbestandteile nach Monaten und Habitattypen, Mediane, n = 220. ................................................................................................................... 95
Abbildung 44: Auftretensfrequenz von Äsungspflanzen in den Untersuchungsgebieten. ........................................................................................................................ 102
Abbildung 45: Mittleren Qualität der kartierten Äsungspflanzen nach Habitaten. ...... 108Abbildung 46: Mittleren Qualität der pflanzensoziologischen Kategorien im Frühling,
links = Wald, rechts = Land. ............................................................................ 109Abbildung 47: Mittleren Qualität der pflanzensoziologischen Kategorien im Sommer,
links = Wald, rechts = Land. ............................................................................ 109Abbildung 48: Mittlere Qualität der pflanzensoziologischen Kategorien im Herbst, links
= Wald, rechts = Land. .................................................................................... 110Abbildung 49: Mittlere Qualität der pflanzensoziologischen Kategorien im Winter, links
= Wald, rechts = Land. .................................................................................... 111Abbildung 50: Anteil der mit landwirtschaftlichen Produkten bestockten Agrarfläche im
Untersuchungsgebiet „Land“ ........................................................................... 112Abbildung 51: Ruminales Mikrobiom (Anzahl DNA Kopien/g TS) nach Habitattypen,
MO = Mikroorganismen, Mittelwerte mit 95 % Konfidenzintervall, n = 129, p < 0,001. .............................................................................................................. 114
Abbildung 52: Ruminale Mikrobenarten (Anzahl DNA Kopien/g TS) nach Habitattypen, Mediane mit 95 % Konfidenzintervall, n = 129. ............................................... 115
Abbildung 53: Darstellung der gesamten Mikrobenzahlen nach Jahreszeiten und Habitattypen, Mediane mit 95 % Konfidenzintervall, n = 129. ........................ 116
Abbildung 54: Mikrobenzahlen (Anzahl DNA Kopien/g TS) nach Gattungen, Jahreszeiten und Habitattypen, Mediane ohne 95 % Konfidenzintervall, N = 129. ................................................................................................................. 117
Abbildung 55: Pansenvolumen nach Jahreszeiten und nach Habitattypen, n = 155, Mittelwerte mit 95 % Konfidenzintervall. ......................................................... 117
Abbildung 56: Panseninhaltsmenge nach Jahreszeiten und Habitattypen, n = 157, Mittelwerte mit 95 % Konfidenzintervall. ......................................................... 119

6 Abbildungsverzeichnis
179
Abbildung 57: Prozentuale Pansenfüllung in Relation zum verfügbaren Pansenvolumen, n = 155, Mittelwerte mit 95 % Konfidenzintervall, Unterschiede Frühjahr p = 0,025, Herbst p = 0,003, Winter p = 0,005 ................................. 120
Abbildung 58: Gewichte der aufgebrochenen, adulten Rehe im Jahresverlauf nach Monaten, n = 172, Mittelwerte mit 95 % Konfidenzintervall. ........................... 122
Abbildung 59: Gewichte der aufgebrochenen adulten Rehe im Jahresverlauf nach Monaten und Habitat, n = 172, Mittelwert mit 95 % Konfidenzintervall, Unterschied November p = 0,035. .................................................................. 123
Abbildung 60: Darstellung mittlere Gewichte nach Altersklassen und Habitaten, n= 220, Mittelwerte mit 95 % Konfidenzintervall .......................................................... 124
Abbildung 61: Durchschnittsgewichte der Kitze (n = 72, p = 0,012) LINKS, subadulten Rehe (n = 53, p = 0,006) RECHTS aus Land- und Waldgebiet, Mittelwerte mit 95 % Konfidenzintervall. ................................................................................. 125
Abbildung 62: Kitzgewichte nach Habitaten und Monaten, n = 72, Mittelwerte mit 95 % Konfidenzintervall. ........................................................................................... 126
Abbildung 63: Gewichtsdaten nach Geschlechtern und Habitaten, M = Böcke p = 0,036, W = Geißen, Mediane, n = 220. ........................................................... 127
Abbildung 64: Konditionswerte ohne Kitze (NFI = Nierenfettindex) nach Habitattyp und Jahreszeit, Wald n = 69, Land n = 95, Mediane mit 95 % Konfidenzintervall . 128
Abbildung 65: Korrelation zwischen Körpergewicht und Nierenfettindex. .................. 129Abbildung 66: GCM-Konzentration der Untersuchungstiere beider Habitattypen, n =
128, GCM = Glukokortikoidmetabolite. ........................................................... 130Abbildung 67: Links GCM-Werte nach Geschlechtern p = 0,001. Rechts GCM-Werte
nach Jahreszeiten, Mediane mit 95 % Konfidenzintervall. .............................. 131Abbildung 68: GCM-Werte aller Tiere nach Jahreszeiten, n = 128, Unterschiede
Frühling-Herbst p = 0,004, Frühling-Winter p = 0,001. ................................... 132

7 Tabellenverzeichnis
180
7 Tabellenverzeichnis
Tabelle 1: Auflistung der Untersuchungsgebiete mit beschreibenden Parametern. ..... 26Tabelle 2: Meteorologische Einteilung der Jahreszeiten zur Einordnung der
Untersuchungstiere. .......................................................................................... 29Tabelle 3: Probenzahlen nach Jahreszeiten, Monaten und Habitattyp. ....................... 32Tabelle 4: Parameter zur Analyse der Qualität der Nahrung von Rehen. .................... 44Tabelle 5: Verwendete Probenanzahl und Herkunft für spezialisiertes HFT-Verfahren.
.......................................................................................................................... 55Tabelle 6: Verwendete Primer und ihre Annealing-Temperaturen. .............................. 61Tabelle 7: Unterteilung der Äsungsarten nach logischen Kategorien in Anlehnung an
Klansek et al. (1995). bzw. Klansek (2004). ..................................................... 63Tabelle 8: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen
über alle Jahreszeiten, Mittelwerte, n = 220. .................................................... 96Tabelle 9: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen
im Frühling, Mittelwerte, N=220. ....................................................................... 97Tabelle 10: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen
im Sommer, Mittelwerte. ................................................................................... 98Tabelle 11: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen
im Herbst, Mittelwerte. ...................................................................................... 99Tabelle 12: Prozentualer Anteil an Pansen mit vorgefundenen Nahrungsbestandteilen
im Winter, Mittelwerte. .................................................................................... 100Tabelle 13: Prozentuale Anteile gefundener Nadel- und Laubhölzer nach Habitaten,
Mittelwerte, n = 220. ........................................................................................ 101Tabelle 14: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Waldgebiet.
........................................................................................................................ 103Tabelle 15: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Landgebiet.
........................................................................................................................ 103Tabelle 16: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Waldgebiet.
........................................................................................................................ 104Tabelle 17: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Landgebiet.
........................................................................................................................ 105Tabelle 18: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Waldgebiet. . 105Tabelle 19: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Landgebiet. . 106Tabelle 20: Auftretungsfrequenz verschiedener Pflanzen im Winter im Waldgebiet. . 107Tabelle 21: Auftretungsfrequenz verschiedener Pflanzen im Winter im Landgebiet. . 107Tabelle 22: Mittelwerte und p-Werte der Panseninhaltsmengen nach Habitat und
Jahreszeit. ....................................................................................................... 119Tabelle 23: Mittelwerte der Gewichte der Probetiere nach Habitattyp, mit und ohne
Kitze, DS = Datensatz. .................................................................................... 122Tabelle 24: Mediane des Nierenfettindexes der Probentiere nach Habitattyp nach
Jahreszeit, ohne Kitze, n = 164. ...................................................................... 127Tabelle 25: Werte der Rohnährstoffanalyse im direkten Vergleich zu Werten früherer
Studien, Mittelwerte der Rohnährstoffe angegeben, n = Anzahl, XP = Rohprotein, XL= Rohfett, XF = Rohfaser, NfE= Stickstofffreie-Extraktstoffe, Rohnährstoffe in [ % zu g/kg Trockensubstanz]. ............................................ 148

7 Tabellenverzeichnis
181
Tabelle 28: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Frühling, Mittelwerte, n = 220. ......................................................................... 186
Tabelle 29: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Sommer, Mittelwerte, n = 220. ........................................................................ 187
Tabelle 30: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Herbst, Mittelwerte, n = 220. ........................................................................... 188
Tabelle 31: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Winter, Mittelwerte, n = 220. ........................................................................... 190
Tabelle 32: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Waldgebiet. ........................................................................................................................ 191
Tabelle 33: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Landgebiet. ........................................................................................................................ 192
Tabelle 34: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Waldgebiet. ........................................................................................................................ 193
Tabelle 35: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Landgebiet. ........................................................................................................................ 194
Tabelle 36: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Waldgebiet. . 195Tabelle 37: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Landgebiet. . 196Tabelle 38: Auftretungsfrequenz verschiedener Pflanzen im Winter im Waldgebiet. . 197Tabelle 39: Auftretungsfrequenz verschiedener Pflanzen im Winter im Landgebiet. . 198

10 Veröffentlichungen
182
8 Formelverzeichnis
Formel 1: Berechnung des prozentualen Trockensubstanzgehaltes nach VDLUFA (2013). ................................................................................................................ 36
Formel 2: Berechnung der Rohasche nach VDLUFA (2013). ...................................... 36Formel 3: Berechnung Rohfettgehalt nach VDLUFA (2013). ....................................... 37Formel 4: Ermittlung der verbrauchten Schwefelsäure ................................................ 38Formel 5: 1ml Schwefelsäure 0,1n entspricht 1,4 mg Stickstoff ................................... 38Formel 6: Berechnung des Gesamtstickstoffes ............................................................ 38Formel 7: Berechnung des Rohproteins auf die Trockenmasse .................................. 38Formel 8: Berechnung des Rohfaseranteils. ................................................................ 39Formel 9: Berechnung der Stickstofffreien-Extraktstoffe. ............................................. 40Formel 10: Berechnung des NDF Gehaltes. ................................................................ 42Formel 11: Berechnung des ADF Gehaltes. ................................................................. 42Formel 12: Berechnung des ADL Gehaltes. ................................................................. 43Formel 13: Berechnung des Zellulose Gehaltes. ......................................................... 43Formel 14: Berechnung des Hemizellulose Gehaltes. ................................................. 43Formel 15: Berechnung der Nicht-Faser-Kohlenhydrate. ............................................. 44Formel 16: Bestimmung der Stoffmenge nach Mentor (2003). .................................... 46Formel 17: Formel 4 nach Stoffmenge pro Liter aufgelöst. .......................................... 47Formel 18: Reaktionsgleichung Bildung von NADP+ (Sigma 2012). ........................... 48Formel 19: Berechnung der Endkonzentration NH3 (Sigma 2012), ∆A340 = Adsorption
bei 340nm. ......................................................................................................... 48Formel 20: Berechnung des Ammoniakgehaltes (Sigma 2012). .................................. 48Formel 21: Reaktionsgleichung Bildung von NADH (Roche 2012). ............................. 49Formel 22: Berechnung der Endkonzentration von Laktat. .......................................... 49Formel 23: Bestimmung der Laktat-Konzentration (Roche 2012). ............................... 50Formel 24: Berechnung von L-Milchsäure mit ∆EL-Milchsäure (Roche 2012). ............ 50Formel 25: Berechnung von D-Milchsäure mit ∆ED-Milchsäure (Roche 2012). ........... 50Formel 26: Berechnung der Gasbildung GB24. ........................................................... 54Formel 27: Berechnung der Umsetzbaren Energie ME. ............................................... 54Formel 28: Berechnung des Verdaulichkeitsfaktors dOS. ............................................ 55Formel 29: Berechnung des Nierenfettindexes nach Riney (1955). ............................. 57Formel 30: Berechnung der Kopienanzahl pro g Trockensubstanz ............................. 62Formel 31: Berechnung der Auftretungsfrequenz einzelner Nahrungsbestandteile. .... 64Formel 32: Berechnung der Auftretungsfrequenz vorgefundener Vegetation. ............. 64Formel 33: Berechnung der GCM Konzentration von pg/Well in ng/g Kot. .................. 65

9 Interdisziplinäre Zusammenarbeit und wissenschaftlicher Austausch
183
9 Interdisziplinäre Zusammenarbeit und Austausch
• Forstwissenschaftliche Tagung Sep. 2012 im Wissenschaftszentrum Wei-
henstephan, Vorstellung und Diskussion des Projektes mit Fachleuten
• Tagung der Vereinigung der Wildbiologen und Jagdwissenschaftlern Oktober
2012 in Bonn, Vorstellung und Diskussion des Projektes mit Fachleuten
Infoveranstaltung für BJV Kreisgruppen Eggenfelden und Starnberg im Feb. 2013 • Statusseminar 11. März 2013, Ausführliche Darstellung des Themas und Stand
der Projektarbeit
• IUGB Tagung Aug. 2013 Brüssel, Vorstellung des Themas, Poster-Präsentation
Tagung der Vereinigung der Wildbiologen und Jagdwissenschaftlern November 2014 in Weihenstephan Freising, Forstfakultät, TU München. Abschlussseminar 11. März 2016, Ausführliche Darstellung der Ergebnisse dem Obersten Jagdbeirat sowie einem wissenschaftlichem Auditorium Tagung und 25jähriges Gründungsjubiläum der Gesellschaft für Wildtier- und Jagdfor-schung 21.4.24.4.2016; Vortrag: Energiehaushalt von Rehen in einem Wald- und Ag-rarökosystem Tagung der Vereinigung der Wildbiologen und Jagdwissenschaftler Deutschland in Trippstadt 21. Juni 2016; Vortrag: Energiehaushalt von Rehen im Jahresverlauf

10 Veröffentlichungen
184
10 Veröffentlichungen
König A., Scheingraber, M (2016): Energiehaushalt von Rehen in einem Wald- und Agrarökosystem. Beitrage zur Jagd- und Wildtierforschung, Bd. 41, im Druck
König, A., Scheingraber, M. (2016): Energiehaushalt von Rehen im Jahresverlauf. In: König, A., Hohmann, U., Ebert, C., Mitschke, J Wildbiologische Forschungsberichte Bd. 2, 197 – 207, Kessel-Verlag, Remagen-Oberwinter.
Scheingraber, M., Klobetz, E., Palme, R., König, A. (2016): Wie gestresst sind unsere Rehe? How stressed are our roe deer? In: König, A., Hohmann, U., Ebert, C., Mitschke, J Wildbiologische Forschungsberichte Bd. 2, 74 – 85, Kessel-Verlag, Remagen-Oberwinter

11 Studentische Abschlussarbeiten
185
11 Studentische Abschlussarbeiten im Rahmen des Projektes
Mayr, J. 2015: Untersuchungen des Pansen-Mikrobioms beim Reh (Capreolus capreo-lus). Bachelorarbeit Naturwissenschaftliche Bildung (Lehramt an Gymnasien) Fächerkombination Biologie/Chemie, AG Wildbiologie und Wildtiermanage-ment, LS Tierernährung, WZW, TU-München
Linhart, K 2014: Untersuchung des Nahrungsspektrums vom Rehwild (Capreolus cap-reolus) aus waldreichen und landwirtschaftlich geprägten Habitaten in Bayern anhand von Panseninhaltsanalysen. Bachelorarbeit an der Studienfakultät für Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wild-tiermanagement, LS Tierökologie, WZW, TU-München
Hudler, E. 2013: Molekulare Analyse von Borrelia burgdiorferi s.l., FSMW-Viren und Rickettisa spp. Beim Rehwild (Capreolus capreolus) und Zecken (Ixodes ricinus) in unterschiedlich geprägten Habitaten in Bayern. Master Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München
Häuslschmid, K. 2013: Nahrungswahl und Nahrungsqualität der Cerviden. Bachelor Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München
Heckel, K. 2013: Physiologische Untersuchung der Adaption des Rehpansens im Jahresverlauf. Bachelor Thesis Naturwissenschaftliche Bildung (Lehramt an Gymnasien) Fächerkombination Biologie/Chemie
Schmid, M. 2013: Anwendung des Hohenheimer Futterwerttests & des Weender-Verfahrens bei natürlicher Nahrung von Rehen (Capreolus capreolus). Bachelor Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München
Schneider, H. 2012: Entwicklung eines Standardverfahrens zur Analyse der Qualität der Nahrung von Rehen (Capreolus capreolus). Bachelor Thesis an der Stud.fak. Forstwissenschaft und Ressourcenmanagement, AG Wildbiologie und Wildtiermanagement, LS Tierökologie, WZW, TU-München

12 Anhang
186
12 Anhang
12.1 Auftretensfrequenz von Äsungspflanzen im Pansen nach Jahreszeiten und Habitaten Tabelle 26: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Früh-ling, Mittelwerte, n = 220.
Klasse Wald Anteil [%]
Land Anteil [%]
Baumfrüchte
1,18 3,45
Eicheln 1,18 3,45
Feldfrüchte
3,53 4,14
Mais 0,00 2,07
unbek. Ackerpflanze 3,53 2,07
Kirrung
2,35 0,00
Apfeltrester 2,35 0,00
Kryptogamen
8,24 2,07
Farne
5,88 1,38
unbek. Farn 5,88 1,38
Moose
1,18 0,69
unbek. Moos 1,18 0,69
Pilze
1,18 0,00
unbek. Pilz 1,18 0,00
Weichäsung
44,71 59,31
Kräuter
23,53 29,66
Brennnessel 7,06 3,45
Ferkelkraut 0,00 1,38
Hahnenfuß 0,00 2,07
Kräuter 0,00 9,66
Kriechender Günsel 0,00 0,69
Löwenzahn 0,00 0,69
Margerite 0,00 0,69
Rotklee 0,00 0,69
Sternmiere 0,00 0,69
unbek. Klee 9,41 9,66
unbek. Kräuter 4,71 0,00
Waldsauerklee 1,18 0,00
Wegerich 1,18 0,00
Stauden
12,94 13,79

12 Anhang
187
Brombeere 12,94 13,10
Himbeere 0,00 0,69
Süßgräser
8,24 15,86
Zähäsung
40,00 31,03
Laubhölzer
10,59 8,97
Buche 5,88 0,69
Eiche 0,00 2,07
Erle 0,00 0,69
Haselnuss 0,00 1,38
unbek. Laubhölzer 4,71 4,14
Nadelhölzer 12,94 10,34
Fichte 10,59 7,59
Tanne 2,35 2,76
Sträucher
16,47 11,72
Heidelbeere 16,47 10,34
Mistel 0,00 1,38
Tabelle 27: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Som-mer, Mittelwerte, n = 220.
Klasse Wald Anteil [%]
Land Anteil [%]
Baumfrüchte
4,55 2,86
Brombeere (Beere) 3,03 0,00
Eicheln 0,00 2,86
Himbeere 1,52 0,00
Feldfrüchte
3,03 21,43
Mais 0,00 14,29
unbek. Ackerpflanze 3,03 5,71
Weizen 0,00 1,43
Kirrung
1,52 0,00
Apfeltrester 1,52 0,00
Kryptogamen
18,18 1,43
Farne
9,09 1,43
unbek. Farn 9,09 1,43
Moose
1,52 0,00
unbek. Moos 1,52 0,00
Pilze
7,58 0,00
unbek. Pilz 7,58 0,00
12 Anhang
188
Weichäsung
50,00 54,29
Kräuter
34,85 28,57
Brennnessel 13,64 10,00
Doldenblütler 0,00 1,43
Hirtentäschel 0,00 1,43
Rotklee 0,00 1,43
unbek. Klee 10,61 8,57
unbek. Kräuter 6,06 5,71
Wegerich 3,03 0,00
Weißklee 1,52 0,00
Sauergräser
0,00 2,86
Stauden
15,15 14,29
Brombeere 15,15 12,86
Himbeere 0,00 1,43
Süßgräser
0,00 8,57
Zähäsung
22,73 20,00
Laubhölzer
9,09 8,57
Buche 1,52 0,00
Eichel 0,00 4,29
Linde 0,00 1,43
Pappel 1,52 0,00
unbek. Laubhölzer 6,06 2,86
Nadelhölzer 1,52 7,14
Fichte 1,52 5,71
Tanne 0,00 1,43
Sträucher
12,12 4,29
Heidelbeere 10,61 2,86
unbek. Sträucher 1,52 1,43
Tabelle 28: Auftretungsfrequenz der Nahrungsbestandteile nach Habitattypen im Herbst, Mittelwerte, n = 220.
Klasse Wald Anteil [%]
Land Anteil [%]
Baumfrüchte
2,42 11,30
Eicheln 2,42 11,30
Feldfrüchte
0,00 15,65
Mais 0,00 13,91

12 Anhang
189
unbek. Ackerpflanze 0,00 1,74
Kirrung
4,03 0,87
Apfeltrester 4,03 0,87
Kryptogamen
15,32 9,57
Farne
5,65 1,74
Ackerschachtelhalm 0,00 0,87
unbek. Farn 5,65 0,87
Moose
0,81 0,87
unbek. Moos 0,81 0,87
Pilze
8,87 6,96
unbek. Pilz 8,87 6,96
Weichäsung
42,74 47,83
Kräuter
16,13 23,48
Brennnessel 5,65 6,09
Doldenblütler 0,00 0,87
Löwenzahn 0,00 0,87
Rotklee 0,00 0,87
Storchenschnabel 0,81 0,00
unbek. Klee 4,03 6,09
unbek. Kräuter 4,84 6,96
unbek. Wegerich 0,00 1,74
Wicke 0,81 0,00
Sauergräser
0,81 0,00
Stauden
15,32 12,17
Brombeere 15,32 11,30
Himbeere 0,00 0,87
Süßgräser
10,48 12,17
Zähäsung
35,48 14,78
Laubhölzer
14,52 3,48
Ahorn 4,03 0,00
Buche 4,84 0,87
Eiche 1,61 0,00
unbek. Laubhölzer 4,03 2,61
Nadelhölzer 8,06 4,35
Fichte 6,45 4,35
Tanne 1,61 0,00
Sträucher
12,90 6,96
Heidelbeere 8,87 2,61
Mistel 0,00 0,87
unbek. Sträucher 4,03 3,48

12 Anhang
190
Tabelle 29: Auftretungsfrequenz der Nahrungsbestandteile (Winter), Mittelwerte, n = 220.
Klasse Wald [%] Land [%]
Baumfrüchte 2,70 17,95
Eicheln 2,70 17,95 Feldfrüchte 1,35 1,92
Mais 1,35 0,64
unbek. Ackerpflanze 0,00 0,64
Weizen 0,00 0,64 Kirrung 6,76 1,28
Apfeltrester 6,76 1,28 Kryptogamen 9,46 5,77
Farne 8,11 3,21
unbek. Farn 8,11 3,21
Moose 1,35 0,64
unbek. Moos 1,35 0,64
Pilze 0,00 1,92
unbek. Pilz 0,00 1,92 Weichäsung 36,49 52,56
Kräuter 6,76 9,62
Brennnessel 0,00 3,21
unbek. Klee 1,35 4,49
unbek. Kräuter 4,05 1,92
Waldsauerklee 1,35 0,00
Sauergras 0,00 0,64
Stauden 22,97 25,00
Brombeere 22,97 23,72
Himbeere 0,00 1,28
Süßgräser 6,76 17,31 Zähäsung 43,24 20,51
Laubhölzer 6,76 2,56
Buche 2,70 0,64
unbek. Laubhölzer 4,05 1,92
Nadelhölzer 28,38 9,62
Fichte 21,62 6,41
Kiefer 1,35 1,92
Tanne 5,41 1,28
Sträucher 8,11 8,33
Heidelbeere 6,76 5,77
Liguster 1,35 0,00
Mistel 0,00 2,56

12 Anhang
191
12.2 Anhang: Auftretensfrequenz von Äsungspflanzen nach Jahreszeiten und Habitaten. Tabelle 30: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Waldgebiet.
Kategorie
Auftretungsfrequenz [%]
Kryptogamen 43,14
Farne
9,38
Waldfrauenfarn 8,40
Wurmfarn 0,84
unbekannter Farn 0,14
Moose 33,75
Farnähnliches Starknervenmoos 10,78
Goldenes Frauenhaarmoos 7,00
Lebermoos 0,14
Sternmoos 2,80
Zypressenschlafmoos 13,03
Weichäsung 40,20
Kräuter 18,91
Brennnessel 4,06
Distel 0,14
Fingerhut 0,14
Frauenmantel 0,14
Großer Sauerampfer 0,28
Huflattich 0,28
Kleiner Sauerampfer 0,14
Klettenlabkraut 0,56
Löwenzahn 0,42
Pestwurz 0,42
Springkraut 0,28
Storchschnabel 0,98
Waldmeister 0,14
Waldrebe 0,28
Waldsauerklee 9,24
Waldveilchen 0,14
Wilde Erdbeere 1,26
Sauergräser 0,14
Stauden 12,04
Brombeere 11,90
Himbeere 0,14
Süßgräser 9,10

12 Anhang
192
Zähäsung 16,67
Laubhölzer 3,50
Ahorn 0,14
Buche 3,22
Kastanie 0,14
Nadelhölzer 9,10
Fichte 7,42
Kiefer 0,70
Lärche 0,14
Tanne 0,84
Sträucher 4,06
Heidelbeere 4,06
Tabelle 31: Auftretungsfrequenz verschiedener Pflanzen im Frühling im Landgebiet.
Kategorie
Auftretungsfrequenz [%]
Feldfrüchte 32,29
Hafer 0,21
Raps 2,10
Senf 4,40
Silagegras 1,89
Wintergetreide 23,69
Weichäsung 67,71
Kräuter 51,78
Breitwegerich 2,10
Ehrenpreis 1,68
Ferkelkraut 0,63
Frauenmantel 0,42
Gänseblümchen 4,61
Gewöhnliches Hornkraut 1,05
Großer Sauerampfer 1,26
Hahnenfußgewächs 1,89
Hirtentäschel 0,84
Kleiner Sauerampfer 2,94
Löwenzahn 8,39
Mittlerer Wegerich 1,05
Rotklee 6,50
Schafgarbe 1,68
Scharbockskraut 0,21

12 Anhang
193
Spitzwegerich 5,24
Storchschnabelgewächs 1,68
Taubnessel 0,63
Vergissmeinnicht 0,63
Vogelmiere 0,21
Weißklee 7,55
Wilde Möhre 0,63
Süßgräser 15,93
Tabelle 32: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Waldgebiet.
Kategorie
Auftretungsfrequenz [%]
Kryptogamen 10,34
Farne
5,75
Wurmfarn 0,57
unbekannter Farn 5,17
Moose 4,60
Farnähnliches Starknervenmoos 1,15
Zypressenschlafmoos 0,57
unbekanntes Moos 2,87
Weichäsung 60,92
Kräuter 33,91
Brennnessel 6,32
Distel 1,15
Doldenblütler 0,57
Frauenmantel 0,57
Gelber Klee 0,57
Hahnenfuß 1,72
Klettenlabkraut 2,30
Pestwurz 0,57
Rotklee 0,57
Springkraut 6,32
Storchschnabel 2,30
Vergissmeinnicht 0,57
Waldmeister 0,57
Waldsauerklee 8,62
Wilde Erdbeere 1,15
Sauergräser 1,15
Stauden 13,79

12 Anhang
194
Brombeere 13,79
Süßgräser 12,07
Zähäsung 28,74
Laubhölzer 14,37
Ahorn 3,45
Buche 6,32
Eberesche 4,02
Eiche 0,57
Nadelhölzer 9,77
Fichte 9,20
Lärche 0,57
Sträucher 4,60
Heidelbeere 4,60
Tabelle 33: Auftretungsfrequenz verschiedener Pflanzen im Sommer im Landgebiet.
Kategorie
Auftretungsfrequenz [%]
Feldfrüchte 42,17
Hafer 1,20
Mais 15,06
Raps 1,20
Wintergetreide 24,70
Weichäsung 57,83
Kräuter 46,99
Breitwegerich 4,22
Doldenblütergewächs 0,60
Gänseblümchen 3,01
Hahnenfußgewächs 4,22
Huflattich 0,60
Kleiner Sauerampfer 2,41
Löwenzahn 9,04
Rotklee 4,82
Schafgarbe 1,20
Spitzwegerich 6,02
Storchschnabelgewächs 3,01
Weißklee 7,83
Süßgräser 10,84

12 Anhang
195
Tabelle 34: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Waldgebiet.
Kategorie
Auftretungsfrequenz [%]
Kryptogamen 38,94
Bärlapppflanzen 0,27
Bärlappgewächs 0,27
Farne
10,45
Waldfrauenfarn 7,87
Wurmfarn 2,44
unbekannter Farn 0,14
Moose 28,22
Farnähnliches Starknervenmoos 9,91
Goldenes Frauenhaarmoos 0,54
Lebermoos 0,27
Sternmoos 6,38
Zypressenschlafmoos 11,13
Weichäsung 38,13
Kräuter 17,91
Alant 0,14
Brennnessel 2,31
Distel 0,27
Fingerhut 0,41
Frauenmantel 0,41
Froschlöffel 0,14
Geflecktes Johanniskraut 0,14
Hahnenfuß 0,41
Klettenlabkraut 0,27
Löwenzahn 0,14
Pestwurz 0,41
Rotklee 0,14
Springkraut 2,71
Storchschnabel 0,68
Waldmeister 0,41
Waldsauerklee 7,46
Weißklee 0,41
Wilde Erdbeere 1,09
Sauergräser 1,22
Stauden 11,26
Brombeere 11,13

12 Anhang
196
Himbeere 0,14
Süßgräser 7,73
Zähäsung 22,93
Laubhölzer 9,77
Ahorn 1,22
Birke 0,14
Buche 6,78
Eberesche 1,36
Eiche 0,27
Nadelhölzer 9,91
Fichte 8,68
Kiefer 0,41
Lärche 0,14
Tanne 0,68
Sträucher 3,26
Heidelbeere 3,26
Tabelle 35: Auftretungsfrequenz verschiedener Pflanzen im Herbst im Landgebiet.
Kategorie
Auftretungsfrequenz [%]
Feldfrüchte 17,99
Hafer 0,17
Mais 4,84
Markstammkohl 0,17
Raps 2,60
Senf 2,60
Silagegras 0,69
Wintergetreide 6,92
Kryptogamen 1,90
Farne
1,90
Ackerschachtelhalm 1,90
Weichäsung 80,10
Kräuter 61,59
Breitwegerich 3,29
Ehrenpreis 1,04
Ferkelkraut 1,56
Frauenmantel 0,87
Gänseblümchen 5,02
Großer Sauerampfer 2,08

12 Anhang
197
Hahnenfußgewächs 2,25
Hirtentäschel 0,52
Hornklee 0,17
Huflattich 0,52
Kamille 0,52
Kleiner Sauerampfer 1,21
Löwenzahn 11,94
Rotklee 7,09
Schafgarbe 2,94
Spitzwegerich 6,75
Storchschnabelgewächs 1,90
Taubnessel 0,69
Weißklee 11,25
Süßgräser 18,51
Tabelle 36: Auftretungsfrequenz verschiedener Pflanzen im Winter im Waldgebiet.
Kategorie
Auftretungsfrequenz [%]
Kryptogamen 43,15
Bärlapppflanzen 0,47
Bärlappgewächs 0,47
Farne
9,19
Waldfrauenfarn 6,39
Wurmfarn 2,80
Moose 33,49
Farnähnliches Starknervenmoos 11,21
Lebermoos 0,47
Sternmoos 8,10
Zypressenschlafmoos 13,71
Weichäsung 34,11
Kräuter 13,24
Brennnessel 1,71
Distel 0,47
Fingerhut 0,31
Frauenmantel 0,16
Springkraut 0,31
Storchschnabel 0,47
Waldmeister 0,62
Waldsauerklee 7,48

12 Anhang
198
Weißklee 0,16
Wilde Erdbeere 1,56
Sauergräser 1,56
Stauden 10,44
Brombeere 10,44
Süßgräser 8,88
Zähäsung 22,74
Laubhölzer 7,63
Buche 7,48
Eiche 0,16
Nadelhölzer 11,68
Fichte 10,12
Kiefer 0,78
Tanne 0,78
Sträucher 3,43
Heidelbeere 3,43
Tabelle 37: Auftretungsfrequenz verschiedener Pflanzen im Winter im Landgebiet.
Kategorie
Auftretungsfrequenz [%]
Feldfrüchte 16,95
Markstammkohl 2,72
Raps 0,42
Senf 9,83
Silagegras 3,56
Wintergetreide 0,42
Kryptogamen 18,62
Moose 18,62
Zypressenschlafmoos 18,62
Weichäsung 64,44
Kräuter 61,72
Breitwegerich 0,42
Ehrenpreis 2,51
Ferkelkraut 7,53
Frauenmantel 1,05
Gänseblümchen 17,78
Hahnenfußgewächs 0,63
Hirtentäschel 0,84
Huflattich 0,42
Kleiner Sauerampfer 0,84

12 Anhang
199
Löwenzahn 3,56
Rotklee 0,42
Schafgarbe 0,84
Spitzwegerich 3,14
Storchschnabelgewächs 7,74
Taubnessel 0,21
Vogelmiere 0,42
Weißklee 13,39
Süßgräser 2,72