inhaltsverzeichnis · material und methoden kommt es zu temperaturanomalien im oberflächenwasser....
TRANSCRIPT


Inhaltsverzeichnis
1 Einleitung..........................................................................................................1
2 Material und Methoden..................................................................................4
2.1 Probennahmegebiet...............................................................................................4
2.2 Pobennahme..........................................................................................................7
2.3 Fauna.....................................................................................................................8
2.3.1 Vorbereitung der Faunaproben.................................................................................8
2.3.2 Auszählen und Bestimmen der Fauna......................................................................9
2.4 Biomassebestimmung...........................................................................................9
2.5 Chlorophyll a.......................................................................................................10
2.5.1 Bearbeitung der Sedimentproben für die Chlorophyll-Bestimmung......................10
2.5.2 Kalibrierung des Fluorometers mittels eines Photometers.....................................10
2.5.3 Messen der Chlorophyll-Proben im Fluorometer...................................................11
2.5.4 Ermittlung der Zerfallsskonstante und der Halbwertzeit von Chlorophyll a.........13
2.6 Bioturbation.........................................................................................................14
2.6.1 Zur Theorie der Bioturbation..................................................................................14
2.6.2 Beschreibung der Bioturbationsmodelle................................................................17
3 Ergebnisse.......................................................................................................21
3.1. Fauna..................................................................................................................21
3.1.1 Verteilung der Gesamtmakrofauna.........................................................................21
3.1.2 Verteilung der Benthischen Foraminiferen............................................................23
3.1.3 Verteilung der Polychaeta......................................................................................24
3.1.4 Einteilung der Polychaeta nach Ernährungstypen..................................................28
3.1.5 Die Polychaeta der einzelnen Stationen und ihre Ernährungstypen.......................30
3.1.6 Betrachtung der Gesamtmakrofauna (Ind. m-2 und Prozentuale Verteilung).........33

Inhaltsverzeichnis
3.2 Biomasse.............................................................................................................35
3.3 Chlorophyll a................................................................................................................36
3.4 Bioturbation..................................................................................................................39
4 Diskussion........................................................................................................45
4.1 Der Kontinentalhang der Iberischen Westküste..................................................45
4.2 Charakterisierung des Vigo-Profils.....................................................................46
4.3 Charakterisierung des Nazaré-Canyons..............................................................55
4.4 Vergleich der bearbeiteten Gebiete im Rahmen dieser Arbeit...........................62
5 Zusammenfassung..........................................................................................68
6 Literatur..........................................................................................................70

Inhaltsverzeichnis

Material und Methoden
2 Material und Methoden 2.1 Probennahmegebiet Das Probennahmegebiet am nordwestlichen Kontinentalhang der Iberischen Halbinsel ist
eines der Untersuchungsgebiete des europäischen Forschungsprojekts OMEX II (Ocean
Margin Exchange) im Rahmen des MAST Forschungsvorhaben (Marine Science and
Technology Programm) der EU. Die Probennahme erfolgte mit dem deutschen
Forschungsschiff METEOR im Dezember/Januar 98-99 in zwei unterschiedlichen Gebieten
am westlichen Kontinentalhang der Iberischen Halbinsel. Es wurde ein Transekt über den
gesamten Kontinentalhang vor Vigo (OMEX II-Box) sowie ein Canyonprofil im OMEX-
Canyon-Gebiet nördlich von Lissabon (Nazaré-Canyon) bearbeitet.
Das Probennahmegebiet ist durch eine Vielzahl von Besonderheiten gekennzeichnet. Die
westlich der kontinentalen Landmassen gelegenen Seegebiete sind Auftriebsgebiete, in denen
nährstoffreiches Tiefenwasser an die Oberfläche transportiert wird. Das Phänomen Auftrieb
ist ein Wind getriebener Prozess.
An der Westküste der Iberischen Halbinsel- zwischen Galizien im Norden und dem Kap Sao
Vicente im Süden- kommt es zwischen April und Oktober zu Auftrieb. Die im Sommer durch
das Wanderungsmuster der Azoren-Hochs hervorgerufenen vorherrschenden nördlichen
küstenparallelen Winde, verursachen eine nach Süden gerichtete Hauptwasserströmung
(LÓPEZ-JAMAR et al., 1992). Diese wird durch den Ekman-Transport im rechten Winkel
zur Windrichtung abgelenkt. Durch die Coriolis-Kraft ist der Ekman-Transport auf der
Nordhalbkugel nach rechts, auf der Südhalbkugel nach links gerichtet. Hierdurch entsteht ein
Wasserdefizit in Küstennähe, welches durch nachströmendes nordatlantisches Zentralwasser
(NACW => North Atlantic Central Water) ausgeglichen wird (FRAGA et al., 1981). Das
NACW kommt aus einer Tiefe von ca. 70-500 m und führt in Küstennähe zu
Temperaturunterschieden von 2-5,5°C im Vergleich zu Oberflächentemperaturen im offenen
Ozean (WOOSTER et al., 1976). Herrschen dagegen vorwiegend südliche Winde, so kehrt
sich die Ekman-Drehung um. Dann kommt es vor allem im Dezember und Januar, zu einer
auflandigen Hauptwasserströmung, welche einen in die Tiefe gerichteten Wassertransport
(Downwelling) zur Folge hat.
Die Topographie des Kontinentalhanges spielt eine große Rolle für die Auftriebsereignisse in
dieser Region. Die 200m Tiefenlinie liegt im Mittel 15-30 km von Land entfernt (PREGO &
BAO, 1996). Hierdurch gelangt das nährstoffreiche Tiefenwasser bis direkt vor die Küste.
Durch die unterseeischen Canyons und die großen nördlichen Kaps der Iberischen Halbinsel 4

Material und Methoden
kommt es zu Temperaturanomalien im Oberflächenwasser. Diese Anomalien nennt man
Kaltwasserfilamente, die von ihrer anfänglichen Größe von 20-30 km auf über 250 km Länge
heranwachsen können (HAYNES et al., 1993). Sie entstehen in der Regel im späten Juli,
Anfang August, als Folge von Mäandrierungen des südwärts gerichteten Stroms. Dessen
wärmeres Wasser und das kalte Auftriebwasser bilden eine Grenze, an der es zu einer
Advektion kommt, die das Auftriebwasser vom Land weg treibt. Hierdurch wird das kalte
Wasser immer mehr in den warmen Südstrom gedrückt und die Wasserkörper beginnen zu
mäandrieren (BATTEN et al., 1992). In diesen Kaltwasserfilamenten wurde an Hand von
Satellitenaufnahmen eine sehr hohe Pigmentkonzentration festgestellt (SOUSA & BRICAUD,
1992).
Durch das aufsteigende nährstoffreiche Tiefenwasser kommt es in Auftriebsgebieten zu den
höchsten Primärproduktionsraten im Ozean. Hierdurch werden auch erhöhte
Sekundärproduktionsraten von Zooplankton, sowie großer Fischreichtum bedingt. Diese
erhöhten Produktionsraten haben zur Folge, dass sehr viel mehr Material absinkt und am
Meeresboden dem Benthos als Nahrungsquelle dienen kann. Somit ist auch die benthische
Lebensgemeinschaft durch hohe Produktionsraten und Biomassen gekennzeichnet (LÓPEZ-
JAMAR et al., 1992).
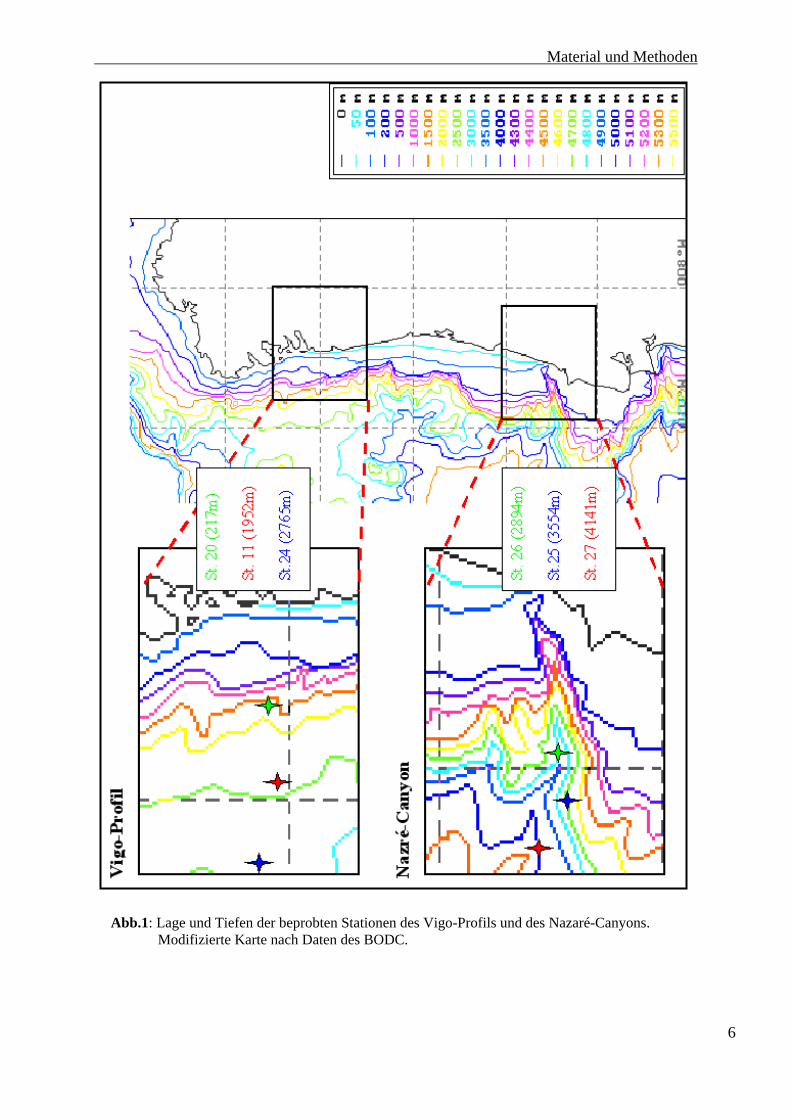
Im Rahmen dieser Arbeit wurden in den beiden Untersuchungsgebieten zwei hangnormale
Transekte bearbeitet (siehe Abb.1). Die Untersuchung des Vigo-Profils fand vom Schelf in
217 m Tiefe über den Kontinentalhang in 1951 m bis zu einem Plateau in 2763 m Tiefe statt.
Dieses Plateau dehnt sich vom beprobten Kontinentalhang bis zur Galicia-Bank etwas
nordwestlich aus.
Der Transekt vor Vigo steht unter dem Einfluss der sehr produktiven Rías. Diese sind
geflutete tektonische Täler der galizischen Küste.
Der Nazaré-Canyon wurde ebenfalls mit drei Stationen beprobt. Die flachste Station lag bei
2894 m, die beiden weiteren Stationen bei 3514 m sowie bei 4141 m. Eine Beprobung
flacherer Gebiete des Canyons war auf Grund seiner Topographie nicht möglich. Der obere
Abschnitt des Canyons ist steil und V-förmig. Starke Reflexionen im oberen Teil des Canyons
und schwere See verhinderten die exakte Tiefenbestimmung durch das Echolot und damit die
gefahrlose Probennahme. Das Canyonbett mäandriert stark, ist sehr schmal und von
Felswänden eingeschlossen. Den südlichen Hang des mittleren Abschnitts durchfurchen
steile, tiefe Einschnitte und Rinnen, die Hangkante ist gerundet. Der nördliche Hang im
mittleren Abschnitt hingegen ist weniger stark zerklüftet und weist eine kammartige
Hangkante auf. Der untere Canyonabschnitt ist abgeflacht und in die Iberische Tiefsee-Ebene 5
Material und Methoden
Abb.1: Lage und Tiefen der beprobten Stationen des Vigo-Profils und des Nazaré-Canyons.
Modifizierte Karte nach Daten des BODC.
6

Material und Methoden
aufgefächert. Hier treten harte Sedimenthorizonte zu Tage (VANNEY & MOUGENOT,
1990).
Die Stationen, ihre Koordinaten und Tiefenangaben sind in Tabelle 1 zusammengefaßt. Die
Lage der Stationen ist in Abbildung 1 dargestellt. Unterschiedliche Koordinaten und
Tiefenangaben an den Stationen resultieren aus der Verwendung unterschiedlicher
Probenahmegeräte, die separat voneinander eingesetzt wurden.
Tab.1 Stationen mit verwendeten Probenahmengeräten (Multicorer = MC, Großkastengreifer =
GKG), Datum, Koordinaten und Wassertiefen Transekt Station Gerät Datum Breiten Längen Tiefe
Vigo-Profil 20 MC 08.01.99 42° 09,0´ N 009° 18,7´ W 217m Vigo-Profil 20 GKG 08.01.99 42° 09,0´ N 009° 18,7´ W 217m Vigo-Profil 11 GKG 07.01.99 42° 10,5´ N 009° 36,0´ W 1945m Vigo-Profil 11 MC 07.01.99 42° 10,5´ N 009° 35,9´ W 1952m Vigo-Profil 24 GKG 09.01.99 42° 09,0´ N 010° 29,9´ W 2763m Vigo-Profil 24 MC 09.01.99 42° 09,1´ N 010° 30,0´ W 2765m
Nazaré-Canyon 26 GKG 12.01.99 39° 29,0´ N 009° 45,1´ W 2894m Nazaré-Canyon 26 MC 12.01.99 39° 29,0´ N 009° 45,1´ W 2878m Nazaré-Canyon 25 GKG 10.01.99 39° 29,7´ N 009° 55,4´ W 3514m Nazaré-Canyon 25 MC 10.01.99 39° 29,7´ N 009° 55,5´ W 3554m Nazaré-Canyon 27 GKG 12.01.99 39° 34,0´ N 010° 10,3´ W 4141m Nazaré-Canyon 27 MC 12.01.99 39° 33,9´ N 010° 10,0´ W 4121m
2.2 Pobennahme Für die Probennahme wurden ein Großkastengreifer (GKG, 0,5m x 0,5m)
(HESSLER/SANDIA USNEL box-corer), sowie ein Multicorer (MC) modifiziert nach
BARNETT et al., (1984) eingesetzt. Die Proben aus dem Großkastengreifer dienten zur
Bestimmung der Fauna, die Sedimentkerne (∅=10 cm) aus dem Multicorer zur Bestimmung
des Sedimentchlorophyll a Gehaltes und der Zerfallskonstante von Chlorophyll a. Auf Grund
stürmischer See und starken Seegangs wurde die Probennahme sehr erschwert. Von sechs
Stationen pro Transekt wurden nur drei möglich. Die Sedimentproben aus dem
Großkastengreifer wurden an Deck mit Seewasser aus der Schiffspumpe aufgeschlämmt und
durch ein 500µm Sieb geseit. Je nach Probenausbeute wurde entweder ein ganzer
Kastengreifer oder eine Unterprobe aus einem Kastengreifer bearbeitet.
Nachdem der Multicorer geborgen war, wurden die Sedimentkerne für die Chlorophyll a
Bestimmung in einen Kühlraum (4°C) überführt. Die Sedimentkerne für die Bestimmung der
Zerfallskonstante von Chlorophyll a wurden in einer Hälterungstruhe bei 4°C für 2-3 Tage
eingelagert (GRAF et al., 1995).
7
Material und Methoden
2.3 Fauna 2.3.1 Vorbereitung der Faunaproben Die Probennahme erfolgte mit dem oben beschriebenen Großkastengreifer (GKG). Nachdem
das Gerät an Deck war, wurde das Oberflächenwasser zu Analysezwecken abgesaugt. Danach
wurde entweder der gesamte Inhalt des GKG’s, oder nur ein bestimmter Teil des Sediments
aufgeschlämmt und durch ein 500µm-Sieb gegeben. Der Siebrückstand wurde in 500ml-
Kautex-Flaschen gefüllt und mit gepuffertem Formalin (4%) fixiert. Aus Zeitgründen und
bedingt durch schweres Wetter, mußten einige Kastengreifer unter mehreren Wissenschaftlern
aufgeteilt werden. Teilweise wurde der Inhalt des GKG’s geviertelt, oder aber es wurden mit
Plexiglasrohren Unterproben genommen. Bei Station 25 mit einem Plexiglasrohr, welches
einen Durchmesser von 14cm und eine Höhe von 36cm hatte, bei Station 26 und 27 hatte das
Rohr 19cm Durchmesser und eine Höhe von 45,5cm. Hierdurch sind verschiedene Mengen
von Sediment gesiebt worden. In Tabelle 2 sind die gesiebten Probenanteile des GKG’s
angegebene, da die ausgelesene Fauna später auf den Quadratmeter hochgerechnet wird.
Da die Proben der Stationen 26 und 27 einen sehr hohen Anteil an terrigenem Material
aufwiesen, war eine Durchsicht der gesamten Probe zu zeitaufwendig. Hier wurden noch
einmal Unterproben genommen, die in Tabelle 2 als Splitt angegeben sind.
Tab.2:Gesiebte Probenfläche des jeweiligen GKG’s in cm² und die für die Probenbearbeitung genommenen Splitts in %.
Transekt Stationen Probenfläche
[cm²] Splitt [%]
Vigo 11 1250 -- Vigo 20 2500 -- Vigo 24 216 --
Nazaré 25 153 -- Nazaré 26 568 3% Nazaré 27 284 2%
Im Labor des Institutes für Aquatische Ökologie i.Gr. in Rostock wurden die Proben für die
Analysen gespült, um das Formalin auszuwaschen. Die Proben wurden dann mit einer Lösung
aus 2g Bengalrot (C.I. 45440) und 1 Liter unverdünntem Methanol nachfixiert. Diese
Methode (LUTZE&ALTENBACH, 1991) wurde modifiziert nach WALTON et al. (1952)
und wird eigentlich zur Färbung von Foraminiferen eingesetzt. Um zu gewährleisten, daß
auch die bei der Probennahme noch lebenden Foraminiferen angefärbt werden, müssen die
Proben 5 Wochen im Dunkeln stehen. Wird dies nicht beachtet, ist zwar die Makrofauna in
den Proben gefärbt, die Foraminiferen jedoch nicht.
8

Material und Methoden
2.3.2 Auszählen und Bestimmen der Fauna Die Proben wurden in kleine Mengen aufgeteilt und unter einem Binokular (Olympus® SZ-
40) in einer Petri- oder Zählschale ausgelesen. Die ausgelesenen Tiere wurden nach
Hauptgruppen sortiert und bis zum Bestimmen in 70% Äthanol fixiert aufbewahrt. Aus
Zeitgründen und wegen der Möglichkeit, daß sich unbekannte Spezies unter den gefundenen
Tieren befinden, war es im Rahmen dieser Arbeit nicht möglich, die Fauna bis auf Artebene
zu bestimmen. Die Polychaeta wurden bis auf Familienebene bestimmt, und als Hauptgruppe
zur Charakterisierung der beprobten Seegebiete herangezogen. Bestand bei den anderen
Gruppen die Möglichkeit, sie bis auf Familienebene zu bestimmen, so wurde dies auch getan.
Für die Bestimmung der Fauna wurden ebenfalls ein Binokular (Olympus® SZ 40) sowie ein
Mikroskop(OLYMPUS® BH-2) verwendet. Die Bestimmung der Polychaeta wurde mit
HARTMANN-SCHRÖDER, (1996) durchgeführt.
Für die Bestimmung der restlichen Fauna wurde mit RIEDL, (1983), gearbeitet. Weitere
Spezialliteratur war nicht verwendet worden, da nicht bis auf Artebene bestimmt wurde.
2.4 Biomassebestimmung Für die Biomassebestimmung wurden die bestimmten und gezählten Tiere, nach ihren
Familien geordnet, in kleine Alufolie-Schiffchen überführt. Die Schiffchen wurden aus
einfacher Alufolie geformt. Ein ca. 2cm x 2cm großer Streifen wurde über eine
abgeschnittene 5cm³ Injektionsspritze gestreift, und zu einem runden Gefäß geformt (ca.
1,5cm hoch, ∅=1cm). Dies war nötig, da die Aluschalen, die für gewöhnlich zur
Biomassebestimmung verwendet werden, nicht in die für die Gewichtsbestimmung
verwendete Analysenwaage (Satorius® M2P, Genauigkeit: 0,001mg) paßten. Nachdem die
Leergewichte der Aluschiffchen ermittelt waren, wurden die Feuchtgewichte der einzelnen
Proben bestimmt. Die Proben wurden dann für 18h in eine Trockenschrank (60°C) überführt.
Nach Ablaufen der Zeit wurden die Proben erneut gewogen und das Trockengewicht
festgestellt. Hiernach wurden die Proben in den Muffeloffen verbracht, wo die Tiere bei
500°C für 18h verglüht wurden. Zur Bestimmung des aschefreien Trockengewichtes wurden
die Proben wieder gewogen.
50% vom aschefreien Trockengewicht werden als organischer Kohlenstoffanteil (mg Corg.)
angesehen.
9

Material und Methoden
2.5 Chlorophyll a 2.5.1 Bearbeitung der Sedimentproben für die Chlorophyll-Bestimmung Von jeweils drei Sedimentkernen (∅=10 cm) wurden die ersten 9 cm abgenommen und in die
folgenden 7 Unterproben aufgeteilt:
0,0 – 0,5 cm 0,5 – 1,0 cm 1,0 – 2,0 cm 2,0 – 3,0 cm 3,0 – 5,0 cm 5,0 – 7,0 cm 7,0 – 9,0 cm
Die so entstandenen Unterproben wurden mittels eines Spatels homogenisiert. Von jeder
Schicht wurden je drei Sedimentproben zu 1cm³ entnommen. Hierfür wurde eine 5 cm³
Injektionsspritze verwendet, welche am unteren Ende aufgeschnitten wurde. Das Sediment
wurde in 10 ml PE-Röhrchen überführt und mit 1cm³ Glaskügelchen, sowie 8 ml 90% Aceton
versehen. Die Glaskügelchen werden verwendet um die Pflanzenzellen in der Zellmühle
aufzuknacken, und das in ihnen befindliche Chlorophyll im Aceton zu lösen. Die Zugabe der
Glaskugeln geschieht auf Grund der Feinheit von Tiefseesedimenten.
Die Proben wurden hiernach in einer Zellmühle 3 Minuten lang gemahlen. Zur Beseitigung
von Sedimentpartikeln in der Aceton-Lösung, wurden die Proben 10 Minuten bei 4000 U/min
zentrifugiert. Nach dem Zentrifugieren erhält man eine grün-gelbe Flüssigkeit und einen
Sedimentniederschlag. Die Chlorophyll-Lösung wird in eine Küvette dekantiert, welche dann
in einem Turner Fluorometer TD 700 der Firma TURNER DESIGNS® gemessen wird.
2.5.2 Kalibrierung des Fluorometers mittels eines Photometers Für eine exakte Messung mit dem Fluorometer muß dieses zuvor kalibriert werden. Für die
Kalibrierung muß eine Chlorophyll a Standardlösung angesetzt werden. Diese wird mittels
1 mg reinem Chlorophyll a (aus Spinat extrahiert) und 100ml 90% Aceton hergestellt.
Nachdem das Chlorophyll gänzlich im Aceton gelöst ist, wird die genaue Chlorophyll a
Konzentration mit einem Photometer ( Spectronic® Genesys 5) der Firma SPECTRONIC
INSTRUMENTS bestimmt (JEFFREY & HUMPHREY 1975). Hierbei wird die Extinktion
der Standard-Lösung bei 4 verschiedenen Wellenlängen gemessen. Die Wellenlängen 647nm
und 663 nm, sind die Absorptionsmaxima von Chlorophyll a, und-b. Die Wellenlänge 630nm
ist das Absorptionsmaximum von Chlorophyll c1 und -c2(RICHTER, 1988). Die Wellenlänge
10

Material und Methoden
von 750 nm wird verwendet, um Spuren von Trübstoffen zu messen und diese durch
Subtraktion von den übrigen Meßwerten zu eliminieren. Der Chlorophyll a Gehalt errechnet
sich dann wie folgt [Gl.1]:
Gl.1
( ) ( ) ( )( ) [ ][ ] [ ]cmngeKüvettenlämlProbenvol
mlAcetonvol75063008,075064754,175066385,11 )ml(µg a Chl 1-
⋅⋅−⋅−−⋅−−⋅
=⋅EEEEEE
Nachdem die Konzentration des Chlorophyll-Standards ermittelt wurde, kann nun eine
Eichreihe angelegt werden. Hierbei wurde in dieser Arbeit von einer Sedimentchlorophyll-
Konzentration von 1µg oder 1000ng ausgegangen. Die Lösung mit der Konzentration von
1000ng wird durch Verdünnen der Standardlösung hergestellt. Da in Tiefseesedimenten zum
Teil sehr geringe Chlorophyll a Gehalte vorkommen, müssen Eichreihen für geringere
Konzentrationen erstellt werden. Hierfür werden die zuerst hergestellten Eichlösungen 1:10
verdünnt, so daß man am Ende drei Eichreihen mit den Konzentrationen von 1000ng – 200ng,
100ng – 20ng, und 10ng – 2ng erhält. Die genauen Konzentrationen der Standards werden
dann noch einmal photometrisch bestimmt.
Die gewonnenen Werte werden bei der Kalibrierung des Fluorometers in das
Kalibrierungsprogramm eingegeben. Das Fluorometer speichert die eingegebene
Konzentration und die gemessene Fluoreszenz der Probe. Nach dem Messen eines Aceton-
Blindwertes errechnet das Fluorometer eine Eichgerade.
2.5.3 Messen der Chlorophyll-Proben im Fluorometer Die gewonnenen Sediment-Chlorophyll-Lösungen können nun gemessen werden. Anhand der
Intensität der Färbung wird auf die zu verwendende Eichreihe geschlossen und das
Fluorometer wie oben beschrieben calibriert. Die einzelnen Chlorophyll-Lösungen werden im
Fluorometer gemessen, mit 250µl 1N HCl angesäuert und erneut gemessen. Durch das
Ansäuern wird das Mg-Zentralatom aus dem Porphyrin-Ring des Chlorophyll a Moleküls
herausgetrennt. Hierdurch wird das Chlorophyll in Phaeopigmente umgewandelt.
Phaeopigmente fluoreszieren bis zu 58% weniger als Chlorophyll (HOLM-HANSEN et al.,
1965). Diese Abnahme der Fluoreszenz wird mit dem Fluorometer ermittelt. Über die
Differenz kann später rechnerisch die Konzentration des Chlorophyll a und der
Phaeopigmente bestimmt werden. Nachdem die Lösung fertig gemessen wurde, wird sie
11

Material und Methoden
verworfen, und das Probengefäß mit dem Sedimentniederschlag wird erneut mit 8ml Aceton
gefüllt, in die Zellmühle gestellt und danach zentrifugiert. So entsteht eine neue, meist
geringer konzentrierte Chlorophyll a Lösung. Insgesamt werden diese Arbeitsschritte 3x
wiederholt. Sind die Meßwerte jedoch schon nach der ersten oder zweiten Messung unterhalb
1ng gesunken, so wurde hier nicht ein zweites- oder drittes Mal gemessen. Die Bearbeitung
der Stationen 11, 20, 24, 25, und 26 fand an Bord des Forschungsschiffes statt. Der Kern der
Station 27 wurde wie oben beschrieben in 7 Unterproben aufgeteilt und diese eingefroren. Die
Chlorophyll-Bestimmung fand im Institut für Aquatische Ökologie i.Gr. in Rostock statt. Die
Berechnung der Chlorophyll a Konzentration wurde mit den folgenden Formeln durchgeführt
(HOLM-HANSEN et al., 1965)[Gl.2 und 3]:
Gl.2
( )( )
Sv
Ef1-Sf
Sf1,03-Fn-Fv )ml(ng Chl 1-
Av a
⋅⋅⎟⎠⎞
⎜⎝⎛⋅
=⋅
Gl.3
( )( )
Sv
AvEf1-Sf
SfFvSf1,03Fn)ml(ng Phaep 1-
⋅⋅⎟⎠⎞
⎜⎝⎛⋅−⋅⋅
=⋅
Die Einheit der Konzentration entspricht immer der Angabe in der Eichreihe. Fv : Fluoreszenz vor dem Ansäuern Fn : Fluoreszenz nach dem Ansäuern 1,03 : Probenverdünnung durch Zugabe von 250µl 1N HCl SF : Säurefaktor (Fv/Fn). Quotient des Standards vor und nach dem
Ansäuern und dann gemittelt ⇒ Gerät- und Eichreihenabhängig EF : Eichfaktor. Reziproker Wert (1/a) der Eichreihensteigung. Sv : Sedimentvolumen (cm³) Av : Aufschlussvolumen⇒ 8ml Aceton + Sedimentvolumen
12

Material und Methoden
2.5.4 Ermittlung der Zerfallsskonstante und der Halbwertzeit von Chlorophyll a Die Ermittlung der Zerfallskonstante und der Halbwertzeit von Chl a ist für die spätere
Modellierung der Bioturbation nötig (siehe 2.6 Bioturbation).
Wie unter Punkt 2.2 beschrieben, werden die Sedimentkerne zur Bestimmung der
Chlorophyll-Zerfallskonstante für mindestens 48h in einer Hälterungstruhe bei in situ
Temperatur (4°C) inkubiert. Nach Ablaufen der Zeit werden die Kerne in oben genannte 7
Sedimenthorizonte aufgeteilt und eingefroren. Die Analyse der Proben fand im Labor statt.
Die Analyse erfolgte nach den unter Punkt 2.3 beschriebenen Arbeitsschritten. Die
Berechnung der Zerfallskonstante, sowie der Halbwertzeit erfolgte mit nachstehenden
Formeln:
Berechnung der Zerfallskonstante Gl.4
( )t
CC 0d
/lnK
−=
: Zerfallskonstante (d-1) Kd : Anfangskonzentration von Chlorophyll a (µg cm-3) C0
C : Konzentration von Chlorophyll a zur Zeit t (µg cm-3) t : Dauer der Inkubation (d) Berechnung der Halbwertzeit Gl.5
d2/1 K
ln2=t
Die Anfangskonzentration von Chlorophyll a ist die Menge, die direkt nach der Probennahme
in den Sedimenten ermittelt wurde. Die Konzentration zur Zeit t ist die Menge an Chlorophyll
a, die in den über 48h inkubierten Kernen ermittelt wurde. Diese Sedimentkerne wurden
jedoch, wie oben erwähnt, nicht mehr an Bord des FS Meteor analysiert.
Durch das Einfrieren der Proben konnte wesentlich mehr Chlorophyll a gefunden werden als
bei der direkten Analyse an Bord des Schiffes (SUN et al., 1991). Hierdurch wurde eine
zweite Inkubation nötig. Diese wurde mit den aufgetauten Sedimenten im Labor in Rostock
durchgeführt. Die aufgetauten Proben wurden 20 Tage bei 4°C im Kühlschrank stehen
13

Material und Methoden
gelassen und dann gemessen. Somit war C0 die Anfangskonzentration nach dem Auftauen und
die Endkonzentration C die Menge an Chlorophyll a, die nach 20 Tagen ermittelt wurde.
2.6 Bioturbation 2.6.1 Zur Theorie der Bioturbation Durch die Aktivität benthischer Organismen, unterliegen Sedimente einer intensiven
Durchmischung. Dieser Effekt wird als Bioturbation bezeichnet. Frisch sedimentiertes
Material kann innerhalb weniger Tage in bis zu 10cm tiefe Sedimentschichten verfrachtet
werden.(GRAF&RITZRAU, 1997). Um diesen Mischungseffekt zu quantifizieren benötigt
man einen Tracer. Tracer sind reaktive oder nichtreaktive Substanzen, welche die
Sedimentbewegung markieren und ihr folgen ohne diese zu beeinflussen (BOUDREAU,
1986a). Reaktive Tracer sind z.B. Radionuclide oder auch Chlorophyll. Sie unterliegen einem
Zerfall, der im Falle von Radionucliden bekannt und konstant ist. Im Falle von Chlorophyll
kann der Zerfall experimentell bestimmt werden, er variiert jedoch geringfügig. Nichtreaktive
Tracer sind z.B. Luminophoren (mit Fluoreszenzfarbstoff gefärbte Sandkörner), die zur
Quantifizierung der Bioturbation als eine definierte Menge auf das Sediment gegeben werden.
Ihre Verteilung im Sediment kann dann unter UV-Licht beobachtet werden.
Die eigentliche Information über die Durchmischung des Sediments erlangt man durch
statistisch angepaßtes Auflösen der Transportgleichungen (Gl.6) in Bezug auf die beobachtete
Tracerverteilung. Im Falle dieser Arbeit wurde Chlorophyll a als Tracer verwendet.
Der klassische Ansatz die Verteilung des Tracers zu bestimmen, ist nach Berner (1980):
idi
2i
2i CK-
zδCδω
zδCδDb
tδCδ
−=Gl. 6
C = : Konzentration des Tracers i (g cm-3) Db = : Bioturbationskoeffizient (cm²d-1) z = : Sedimenttife (cm) t = : Zeit (d) Kd = : Zerfallskonstante (d-1) ω = : Sedimentationsrate (cm² d-1)
2. Fick’sches Diffusionsgesetz Reaktion oder Zerfall
Sedimentation
14

Material und Methoden
Die Fick´schen Gesetze beschreiben die Diffusion, d.h., daß im Falle des klassischen
Ansatzes von einem diffusionsanalogen Mischungsprozeß ausgegangen wird (BOUDREAU,
1986a). Es wird angenommen, daß bei diffusionsanalogen Mischungsprozessen Partikel über
sehr kleine Strecken transportiert werden. Außerdem wird angenommen, daß sie dabei mit der
gleichen Wahrscheinlichkeit in alle Richtungen transportiert werden. Dies ist aber der
Idealfall, und trifft selten zu. Im Falle des diffusionsanalogen Mischen kann die Infauna als
sehr klein angenommen werden. Sie beeinflusst das Sediment nur soweit (lockert es auf), dass
diffusionsanloges Mischen leichter abläuft. Es gibt jedoch größere Tiere im Sediment, wie die
Conveyor-Belt-Feeder, die Material Förderbandartig in das Sediment hinein oder
heraustransportiern. Durch sie werden größere Mengen Material transportiert als durch
Diffusion.
Man gehe von einer idealisierten vierschichtigen Sedimentsäule aus (siehe Abb.2,
BOUDREAU und IMBODEN, 1987). Die oberen drei Schichten sind biologisch aktiv, und
die vierte unterste ist eine historisch nicht durchmischte Schicht, die unter der
Bioturbationstiefe liegt
Gelangt Sediment auf die Oberfläche an der
Sediment-Wasser-Grenzschicht, so wird es durch
eine Scheingeschwindigkeit, w, nacheinander in
die erste, zweite, dritte und schließlich in die
historische Schicht verfrachtet. Wenn nun
vorhandene Fauna, ausreichend groß ist, so kann
sie Material in der Schicht 1 aufnehmen und als
Fecal-Pellets in Schicht 2 oder 3 abgeben. Ebenso
kann es umgekehrt sein, so daß Material in
Schicht 3 aufgenommen wird, und in Schicht 1
oder 2 abgegeben wird. Es kann also Material in
nicht angrenzende Schichten (1 ⇒ 3 oder 3 ⇒ 1,
siehe Abb.2, Doppelpfeile) verfrachtet werden.
Diese Transportprozesse werden als nonlocal
bezeichnet.
w 1
2
3
Historische Schicht
Um die Tracerverteilung bei nonlocal-Ereignissen zu quantifizieren, muß in (Gl.6) ein
nonlocal-Term eingefügt werden.
Abb.2: Schematische Darstellung der möglichen Austauschprozeße in einer 3-schichtigen Sedimentsäule. Es kann lokalen Austausch zwischen benachbarten Schichten 2 und 3 geben, sowie nicht lokalen Austausch zwischen Schicht 1 und 3. Lokaler Transport durch Begraben ist angezeigt durch w
Bei Stady-State-Bedingungen herrscht eine konstante Zufuhr von Chl.a, die durch Prozesse,
die Chl.a aus dem System entfernen (Transport, Abbau) ausgeglichen wird. Außerdem wird
15

Material und Methoden
angenommen, daß kein Porositäts-Gradient und kontinuierliches „non-local-mixing“
herrschen. Nach SOETAERT et al.,, (1996) läßt sich die Tracerverteilung bei diesen
Bedingungen wie folgt beschreiben (Gl.7):
Gl. 7
0SpQrC-CK-zδ
CδωzδCδDb iiid
i2
i2
=++−
C = : Konzentration des Tracers i (g cm-3) Db = : Bioturbationskoeffizient (cm² d-1) z = : Sedimenttife (cm) t = : Zeit (s) K = : Zerfallskonstante (d-1) d
ω = : Sedimentationsrate (cm² d-1) r = : Ingestionsrate (d-1)
= : nonlocal-Eintarag in Schicht n Qi-1Sp= : Produktion des Tracers (d )
Die Ingestionsrate r beschreibt die Konsumierung von Material durch die Infauna in das
Sediment. Qn ist die nonlocal-Konstante, die den direkten Eintrag in eine Schicht n
beschreibt. Die Produktion des Tracers (Sp) kann bei der Verwendung von Chl.a zur
Untersuchung von Tiefseesedimenten vernachlässigt werden, da Chlorophyll nur in der
euphotischen Zone produziert wird. Somit wird Sp aus den folgenden Gleichungen
herausgenommen.
Möchte man nun die Konzentration des Tracers i in der Tiefe x unter Stady-state-
Bedingungen berechnen, so löst man Gleichung 7 unter Annahme, daß Db>0 ist, wie folgt
auf:
( ) ( )rKQeEeAxCd
ixi
xii
21
+++= ⋅⋅ θθGl. 8
Mit:
( )2Db
DbrK4ωωθ d
2
2
+++=
( )2Db
DbrK4ωωθ d
2
1
++−= und
A und Ei i sind Integrationskonstanten, die durch die Randbedingungen festgelegt werden. Die
Randbedingungen beschreiben die Zustände zwischen den einzelnen Grenzen am und im
Sediment. Die Randbedingungen werden im folgenden Abschnitt (Punkt 2.6.2) genauer
beschrieben.
16

Material und Methoden
2.6.2 Beschreibung der Bioturbationsmodelle In dieser Arbeit wurde mit den Modellen nach (SOETAERT et al., 1996) gearbeitet. Es wurde
von den Autoren eine Familie aus 5 aufeinander aufbauenden Modellen gebildet. Von Modell
zu Modell sind alle Prozesse der vorhergehenden Modelle integriert, und es kommen jeweils
1-2 neue Parameter pro Modell hinzu. Für die Lösung der einzelnen Modelle in den
jeweiligen Schichten werden Integrationskonstatnten benötigt. Diese Integrationskonstatnten
werden durch die Randbedingungen (Abb. 3) bestimmt. Die Randbedingungen beschreiben
verschiedene Zustände an den Grenzschichten zwischen zwei benachbarten
Sedimentschichten oder der Sediment-Wasser-Grenze.
Unter Stady-state-Bedingungen herrschen folgende Randbedingungen:
BB0 :
BB1 : (nach GRAF&RITZRAU,1997) 01 ωCFlux =
Flussgrenze an der Sediment-Wasser-Grenzschicht
001 dxωCdCDbFlux +−=
B : B2
Kontinuität der Aktivität zwischen den Schichten i und j
Ci =Cjs s B : B3 Einfacher Fluß zwischen den Schichten
ssss ωCjdxdCjDbωCi
dxdCiDb +−=+−
BB4 :
BB5 :
Abb.3: BB0 – B5B beschreiben die Zustände der Grenzschichten, wobei i und j für eine Schichtnummer
stehen, 0 für die Sedimentoberfläche und S für eine Tiefe im Sediment. . Auf den folgenden Seiten werden die einzelnen Modelle kurz beschrieben
ss2ss ωCjdxdCjDbFluxωCi
dxdCiDb +−=++−
Fortbestand des Flusses zwischen den Schichten i und j
Fluß in eine definierte Schicht
0dxdC
=∞ „nicht-Gradienten-Grenze“ in der Tiefe ∞
17

Material und Methoden
Modell 1 Hier herrscht keine Durchmischung durch Tiere. Die Verteilung des Tracers ist nur durch die
Sedimentation (ω), und den Zerfall(K ) beeinflußt. d
BB0
0CKdxdCω d =−−
Modell 2: Zu den unter Modell 1 beschriebenen Bedingungen kommt hier ein Diffusionsterm hinzu. Es
besteht eine Flussgrenze B und eine „nicht-Gradientengrenze“ B1 5. Modell 2 beschreibt somit
„stady-state diffusions Mischen“.
Flux1
BB1
0CKdxdCω
dxCdDb d2
2
=−−
BB5
Modell 3:
Das Modell 2 wird durch den Flux 2 (hier die Menge Chl a die direkt in die tiefere
Sedimentschicht gelangt) in die Tiefe L modifiziert. Es sind somit 2 Schichten die betrachtet
werden müssen, (0-L) und (L-∞), mit gleichem Db und w. Hier wird einfaches non-local-
mixing angenommen.
Flux1 Flux2
BB1
0C1Kdx
dC1ωdx
C1dDb d2
2
=−−
BB2 und B4L
0C2Kdx
dC2ωdx
C2dDb d2
2
=−−
BB5
18

Material und Methoden
Modell 4a:
Das Modell 3 wird ergänzt mit der Schichtdicke 2δx in der Tiefe L.
Das heißt, daß der Flux 2 direkt in die Schicht L-δx bis L+δx gelangt. Es wird angenommen,
daß das Material homogen in die Schicht gelangt. Hier wird non-local-mixing in eine
definierte Schicht angenommen.
Modell 4b: Hier wird nicht der direkte Flux 2 aus der Wassersäule in eine definierte Schicht
angenommen, sondern ein Transport von Material aus der Schicht 0-X0 in eine Lage in der
Tiefe L. Der Flux 2 wird durch die Ratenkonstante (r) und C1 (resultiert aus Flux 1) gesteuert.
Der Fluß zur Tiefe L wird über Schicht 0-X0 integriert.
Modell 5: Hier wird nicht der direkte Flux 2 aus der Wassersäule in eine definierte Schicht
angenommen. Hier findet der Transport von Material aus der Schicht 0-X0 in eine Schicht in
der Tiefe L-δx bis L+δx. Also ein direkter Transport aus einer Schicht in eine tiefer gelegene
Schicht. Der Flux 2 wird durch die Ratenkonstante (r) und C1 (resultiert aus Flux 1) gesteuert.
Der Fluß zur Tiefe L wird über Schicht 0-X0 integriert.
BB1
BB5
0C1Kdx
dC1ωdx
C1dDb d2
2
=−−
0xδ2
Flux2C2Kdx
dC2ωdx
C2dDb d2
2
=+−−
BB2 und B3
Flux1
BB1 X0
L
BB2 und B3B2B und B4
0C3Kdxdx d2
dC3ωC3dDb2
=−−
0rC1-C1Kdx
dC1ωdx
C1dDb d2
2
=−−
0C2Kdx
dC2ωdx
C2dDb d2
2
=−−
0
∫=xo
02 rC1dxFlux
Flux1 Flux2
0CKdx
dC3ωdx
C3dDb d2
2
=−−
L-δx
L+δx
BB2 und B3
BB5
19

Material und Methoden Flux1
B1 X0
Für die Berechnung der Db-Werte mußten die ermittelten Chlorophyll a Werte und die
jeweiligen Tiefenhorizonte in vorgegebener Art in einen Texteditor eingegeben werden.
Außerdem werden für die Berechnung die Sedimentationsrate, sowie die Zerfallskonstante
von Chlorophyll a benötigt. Da hier in der Zeitdimension pro Tag gerechnet wurde und
Chlorophyll a Halbwertzeiten von 20-30 Tagen hat, kann die Sedimentationsrate = 0 gesetzt
werden. Die Zerfallskonstante wurde rechnerisch, wie unter Punkt 2.5.4 beschrieben,
ermittelt. Hiernach rechnet ein auf MS-DOS basierendes Programm alle fünf Modelle durch.
Die Ergebnisse werden für jedes Modell einzeln, sowie in einer Gesamtübersicht ausgegeben.
Neben dem Db-Wert werden von von den Modellen unter anderem der Oberflächenfluss (µg
cm-2 d-1), ein Injektionsfluss (µg cm-2 d-1), die Injektionstiefe (cm), die Hälfte des
Injektionsintervall dX (cm), die Rate der Nahrungsaufnahme r (d-1) und die Stärke des
Aufnahmeintervalls X0 (cm) berechnet. Der Oberflächenfluss gibt den Chlorophyll- Fluss in
das Sediment an, über den der Kohlenstofffluss berechnet werden kann. Hierfür werden
Corg/Chl.a-Verhältnisse benötigt. Über den Inventar (I), µg Chl.a cm-2 von 0-9cm
Sedimenttiefe und die Zerfallskonstatnte Kd kann nach SUN et al., (1994) unter Annahme von
stady-state ebenfalls der Chlorophyll a-Fluss (J) errechnet werden (J=I Kd). Dieser kann
dem durch die Modelle berechneten Fluss gegenübergestellt werden.
Die Autoren haben zum Testen der Signifikanz der Modelle zu den gemessenen Daten einen
einseitigen F-Test (SOKAL & ROHLF, 1995 aus SOETAERT et al.,1996) verwendet. Dieser
Test gibt für jede berechnete Station an welches Modell statistisch gesehen den besten Fit
ergibt. Für die hier verwendeten Chlorophyll a Profile ergab dieser Test meist keine
B5
L-δx
0rC1-C1Kdx
dC1ωdx
C1dDb d2
2
=−−0
∫=xo
02 rC1dxFlux
B2 und B3
0C2Kdx
dC2ωdx
C2dDb d2
2
=−−
B2 und B3
0C3Kdx
dC3ωdx
C4dDb d2
2
=−−
0xδ2
Flux2C3Kdx
dC3ωdx
C3dDb d2
2
=+−−
L+δx B2 und B3
20

Material und Methoden
schlüssigen Ergebnisse. Es wurde sich daher auf die graphische Auswertung gestützt, da dort
zu sehen ist, welcher Fit am besten auf die gemessenen Profile paßt.
21

Ergebnisse
3 Ergebnisse 3.1. Fauna
3.1.1 Verteilung der Gesamtmakrofauna
In Abbildung 4 sind die Individuenzahlen der einzelnen Stationen nach Stämmen, Klassen
oder Familien geordnet aufgeführt (je nach Möglichkeiten der Bestimmung). Hierbei werden
die Polychaeta und Foraminifera separat behandelt, da sie auf Grund ihrer hohen
Individuenzahlen nicht in die Diagramme aufgenommen werden konnten.
Es ist deutlich zu erkennen, dass sich die Stationen 20 und 11 des Vigo-Profils von den
anderen Stationen durch höhere Familienzahlen unterscheiden. Außerdem ist zu erkennen,
dass Nematoda und Bivalvia an allen Stationen des Vigo-Profils und des Nazaré-Canyons
vorgefunden wurden. Die Klasse der Bivalvia umfasst alle Tiere dieser Gruppe, die nicht
weiter bestimmt werden konnten.
An Station 20 (217m) bilden die Bivalvia mit 36 Ind. m-2 die größte Klasse. Amphipoda und
Anisopoda sowie Thalassinidae aus der Klasse der Crustacea sind mit 20-28 Ind m-2 ebenfalls
stark vertreten. Goneplacidae und Harpacticoida (Crustacea) sind mit 4 Ind. m-2 nicht
auffällig. Die Familie der Pennatulacea (Seefedern) aus der Klasse der Anthozoa wurde mit 4
Ind. m-2 nur an Station 20 gefunden. Die Nematoda sind an Station 20 mit 4 Ind. m-2 vertreten.
An Station 11 (1945m) stellen die Nematoda die größte Klasse mit 176 Ind. m-2. Die Klasse
der Bivalvia ist mit 120 Ind. m-2 vertreten. Der nächste große Stamm umfasst die Sipunculida.
Sie wurden nur an Station 11 mit 72 Ind. m-2 nachgewiesen. Die Klasse der Crustacea ist
durch die Familien der Anisopoda (64 Ind. m-2), Isopoda (32 Ind. m-2 ), Caprellidae (32 Ind.
m-2 ), Amphipoda und Cumacea (beide 24 Ind. m-2) sowie der Ostracoda ebenfalls mit 24 Ind.
m-2 vertreten. Die Klasse der Ophiuroidae aus dem Stamm der Echinodermata wurde nur an
Station 11 nachgewiesen und ist mit 16 Ind. m-2 vertreten. Auch die Klasse der Scaphopoda
(Grab-oder Kahnfüßer) aus dem Stamm der Mollusca ist mit 16 Ind. m-2 gefunden worden.
Auffallend ist, dass bei der Klasse der Crustacea und der Klasse der Bivalvia die
Individuenzahlen um den Faktor 3-5 höher sind als an Station 20. Außerdem wurden an
Station 11 zwei Faunengruppen mehr nachgewiesen als an Station 20. An Station 24 (2763m)
nimmt die Anzahl der verschiedenen Gruppen auf 5 ab. Auch hier sind die Nematoda die
dominante Klasse mit 417 Ind. m-2 . Die Individuenzahl der Bivalvia nimmt auf 93 Ind. m-2
21

Ergebnisse
ab. Die Familien der Isopoda und Anisopoda sowie die Klasse der Scaphopoda wurden mit
jeweils 46 Ind. m-2 nachgewiesen.
Vigo-Profil Nazaré-Canyon
A St.20 217m
0
5
10
15
20
25
30
35
40
Indi
vidu
en (n
/m²)
Nematoda
Bivalvia
Anisopoda
Amphipoda
Goneplacidae
Cumacea
Harpacticoida
Thalassinidae
Pennatulacea
DSt.26 2894m
0
5000
10000
15000
20000
25000
30000
Indi
vidu
e ([n
/m²)
Nematoda
Bivalvia
E St.25 3514m
0
200
400
600
800
1000
1200
1400
1600
Indi
vidu
en (n
/m²)
Nematoda
Bivalvia
Anisopoda
Porifera
Scaphopoda
Prochaetoder-matidae
B St. 11 1945m
0
20
40
60
80
100
120
140
160
180
200
Indi
vidu
en (n
/m²)
NematodaBivalviaAnisopodaIsopodaAmphipodaCumaceaSipunculidaCaprellidaeOphiuroidaeOstracodaScaphopoda
C St.24 2763m
050
100150200250300350400450
Indi
vidu
en (n
/m²)
Nematoda
Bivalvia
Anisopoda
Scaphopoda
Isopoda
Abb.4: Individuenzahlen pro m² der Makrofauna, geordnet nach Stämmen, Klassen oder Familien
(ohne Polychaeta) an den beprobten Stationen des Vigo-Profils (A-C) und des Nazaré-
Canyon (D und E). An Station 27 des Nazaré-Canyon wurde keine Fauna gefunden.
22

Ergebnisse
An der Station 26 (2894m) des Nazaré-Canyons bietet sich ein völlig anderes Bild als an den
Stationen des Vigo-Profils. Es konnten, abgesehen von den Polychaeta, nur zwei Gruppen
nachgewiesen werden. Zum einen die Klasse der Nematoda, die hier mit 25666 Ind. m-2
extrem stark vertreten ist, und zum anderen die Bivalvia. Von dieser Klasse wurden 828 Ind.
m-2 ermittelt. Die Besonderheiten dieser Station sind die enorm hohen Individuenzahlen (im
Vergleich zu den anderen Stationen beider Transekte) in großer Tiefe.
An Station 25 (3514m) wurden 6 Gruppen nachgewiesen, von denen auch hier wieder die
Nematoda die größte Gruppe sind (1494 Ind. m-2). Die Scaphopoda sind mit 844 Ind. m-2
vertreten. Ebenso ist mit 844 Ind. m-2 die Familie der Prochaetodermatidae (Stamm: Mollusca
⇒ Klasse: Caudofoveata (Schildfüßer)) nachgewiesen. Die Bivalvia sind mit 130 Ind. m-2
gezählt. Die Crustacea sind mit einer Familie, den Anisopoda (260 Ind. m-2) vertreten. Eine
kleine Besonderheit stellen die 65 Ind. m-2 der Porifera dar.
An Station 25 sowie 26 des Nazaré-Canyons wurden Holothurioidae (Seegurken) gefunden.
Sie konnten jedoch nicht in die Datenerhebung aufgenommen werden. Sie wurden in
Kastengreifern gefunden, die als fehlgeschlagen eingestuft werden mussten. Die GKGs waren
zu tief in das Sediment eingedrungen. Das Sediment quoll oben aus den Kastengreifern
heraus. Eine genaue Quantifizierung dieser Proben war somit nicht mehr möglich. Es sei
jedoch für die spätere Diskussion darauf hingewiesen, dass Holothurien gefunden wurden.
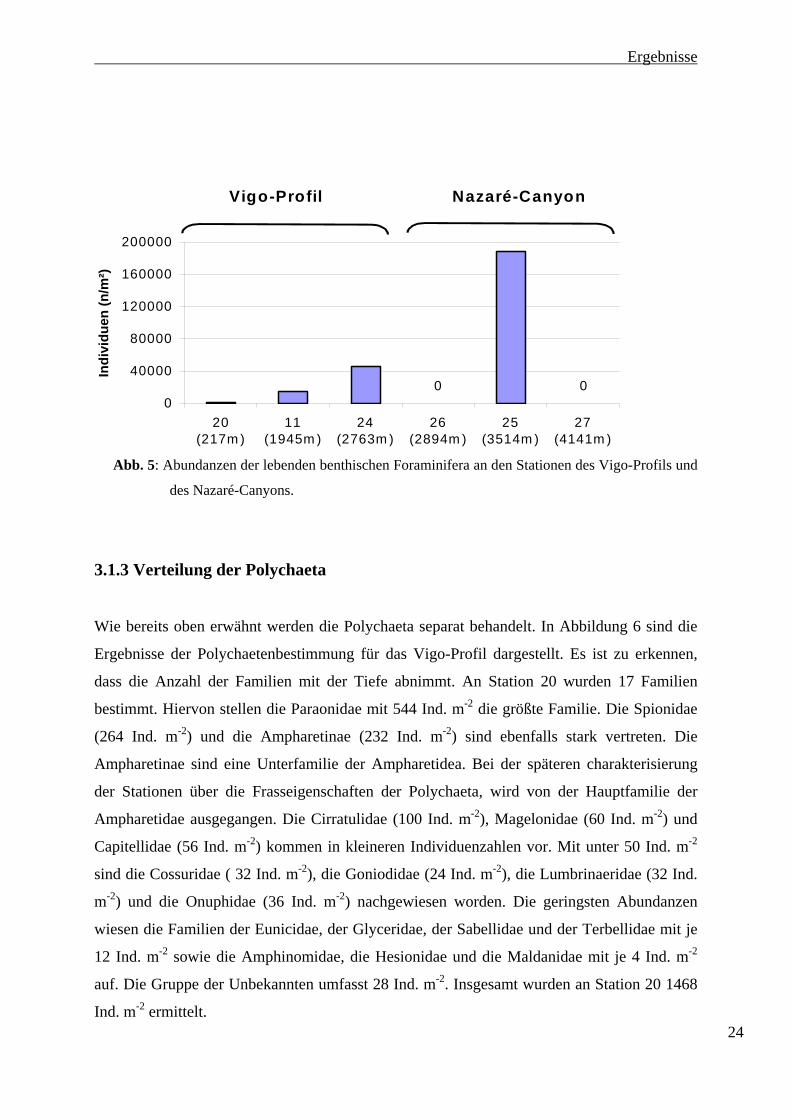
3.1.2 Verteilung der Benthischen Foraminiferen
Eine weitere nicht in der Datenerhebung der Makrofauna aufgeführte Klasse ist die der
Foraminifera. Auch hier wurde nur die Menge Individuen ermittelt, die auf Grund ihrer Größe
oder durch Verkleben mit Sedimentbestandteilen auf dem verwendeten 500µm-Sieb zurück
geblieben sind. Die Abundanzen der lebenden Foraminifera an den beprobten Stationen
beider Transekte sind in Abbildung 5 dargestellt. Es ist deutlich zu erkennen, wie bei den
Stationen 20, 11 und 24 des Vigo-Profils die Individuenzahl der Foraminiferen mit
zunehmender Tiefe ansteigt. Von Station 20 zu Station11 steigt die Individuenzahl um den
Faktor 9 von 1600 Ind. m-2 auf 14945 Ind. m-2. Der Unterschied zwischen Station11 (14945
Ind. m-2 ) und Station 24 (46296 Ind. m-2) liegt etwa bei Faktor 3.
An Station 25 wurden 188700 Ind. m-2 ermittelt. Im Nazaré-Canyon an Station 26 und 27
wurden jedoch keine lebenden Foraminiferen gefunden.
23
Ergebnisse
Vigo-Profil Nazaré-Canyon
0 00
40000
80000
120000
160000
200000
20(217m )
11(1945m )
24(2763m )
26(2894m )
25(3514m )
27(4141m )
Indi
vidu
en (n
/m²)
Abb. 5: Abundanzen der lebenden benthischen Foraminifera an den Stationen des Vigo-Profils und
des Nazaré-Canyons.
3.1.3 Verteilung der Polychaeta
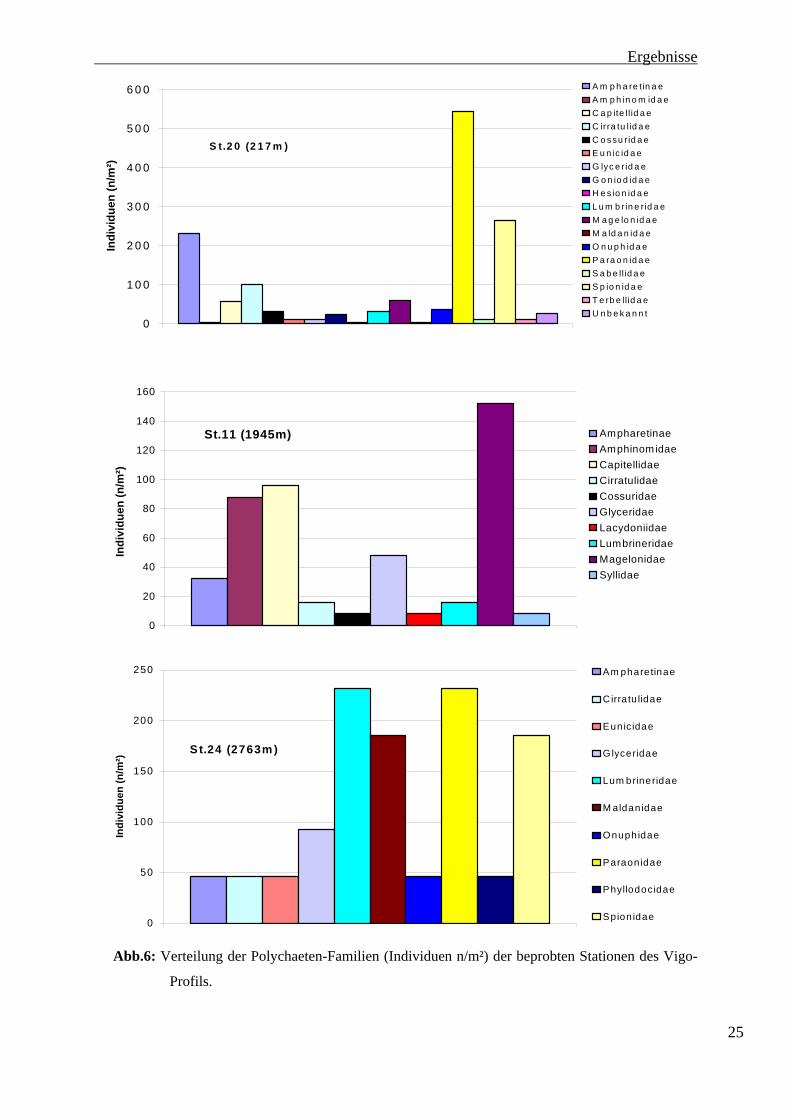
Wie bereits oben erwähnt werden die Polychaeta separat behandelt. In Abbildung 6 sind die
Ergebnisse der Polychaetenbestimmung für das Vigo-Profil dargestellt. Es ist zu erkennen,
dass die Anzahl der Familien mit der Tiefe abnimmt. An Station 20 wurden 17 Familien
bestimmt. Hiervon stellen die Paraonidae mit 544 Ind. m-2 die größte Familie. Die Spionidae
(264 Ind. m-2) und die Ampharetinae (232 Ind. m-2) sind ebenfalls stark vertreten. Die
Ampharetinae sind eine Unterfamilie der Ampharetidea. Bei der späteren charakterisierung
der Stationen über die Frasseigenschaften der Polychaeta, wird von der Hauptfamilie der
Ampharetidae ausgegangen. Die Cirratulidae (100 Ind. m-2), Magelonidae (60 Ind. m-2) und
Capitellidae (56 Ind. m-2) kommen in kleineren Individuenzahlen vor. Mit unter 50 Ind. m-2
sind die Cossuridae ( 32 Ind. m-2), die Goniodidae (24 Ind. m-2), die Lumbrinaeridae (32 Ind.
m-2) und die Onuphidae (36 Ind. m-2) nachgewiesen worden. Die geringsten Abundanzen
wiesen die Familien der Eunicidae, der Glyceridae, der Sabellidae und der Terbellidae mit je
12 Ind. m-2 sowie die Amphinomidae, die Hesionidae und die Maldanidae mit je 4 Ind. m-2
auf. Die Gruppe der Unbekannten umfasst 28 Ind. m-2. Insgesamt wurden an Station 20 1468
Ind. m-2 ermittelt. 24
Ergebnisse
S t.2 0 (2 1 7 m )
0
1 0 0
2 0 0
3 0 0
4 0 0
5 0 0
6 0 0
Indi
vidu
en (n
/m²)
A m p h a re t in a eA m p h in o m id a eC a p ite ll id a eC irra tu lid a eC o ssu rid a eE u n ic id a eG lyc e r id a eG o n io d id a eH e s io n id a eL u m b rin e rid a eM a g e lo n id a eM a ld a n id a eO n u p h id a eP a ra o n id a eS a b e llid a eS p io n id a eT e rb e llid a eU n b e k a n n t
St.11 (1945m)
0
20
40
60
80
100
120
140
160
Indi
vidu
en (n
/m²)
AmpharetinaeAmphinomidaeCapitellidaeCirratulidaeCossuridaeGlyceridaeLacydoniidaeLumbrineridaeMagelonidaeSyllidae
S t.24 (2763m )
0
50
100
150
200
250
Indi
vidu
en (n
/m²)
Am pharetinae
C irra tu lidae
Eunic idae
Glyceridae
Lum brineridae
M aldanidae
Onuphidae
Paraonidae
Phyllodocidae
Spion idae
Abb.6: Verteilung der Polychaeten-Familien (Individuen n/m²) der beprobten Stationen des Vigo-
Profils.
25

Ergebnisse
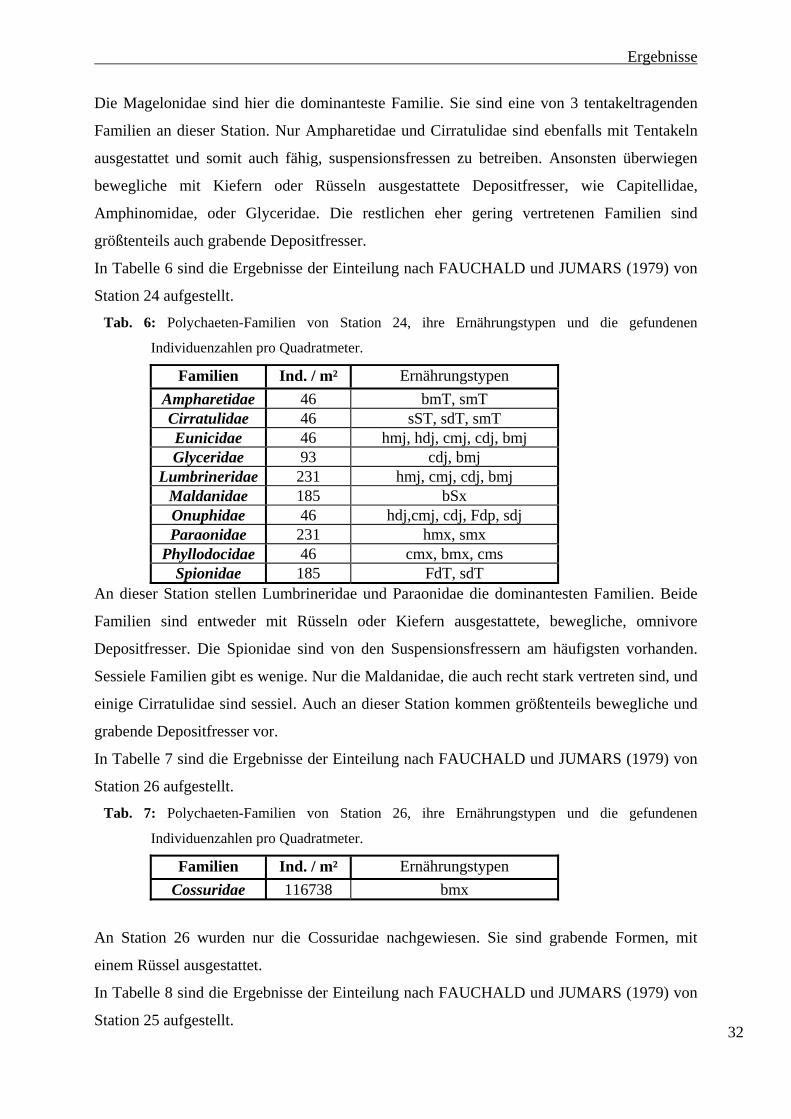
Die Station 11 weist 10 Familien auf und liegt damit deutlich unter der Familien-Anzahl von
Station 20. Die an Station 11 am stärksten vertretenen Familien sind die Magelonidae (152
Ind. m-2), die Capitellidae mit 96 Ind. m-2 und die Familie der Amphinomidae mit 88 Ind. m-2.
Die Familien der Glyceridae (48 Ind. m-2) und der Ampharetinae (32 Ind. m-2) sind mit
mittleren Individuenzahlen nachgewiesen worden. Die Cirratulidae (16 Ind. m-2), die
Lumbrineridae (16 Ind. m-2), die Cossuridae (8 Ind. m-2), die Lacydoniidae (8 Ind. m-2) und
die Syllidae (8 Ind. m-2) wurden nur in geringen Zahlen nachgewiesen. Die Individuenzahlen
sind an dieser Station deutlich niedriger als an Station 20. An Station 11 wurden insgesamt
472 Ind. m-2 gezählt.
An Station 24 wurden, wie an Station 11, 10 Familien nachgewiesen. Hier wurden jedoch die
Familien mit sehr viel höheren Individuenzahlen ermittelt. Die Lumbrineridae und die
Paraonidae wurden mit je 231 Ind. m-2 gezählt. Die Maldanidae und die Spionidae mit je 185
Ind. m-2.
1468
472
1157
0
200
400
600
800
1000
1200
1400
1600
20 11 24
Stationen
Indi
vidu
enza
hl (n
/m²)
Abb. 7: Gesamtindividuenzahlen (Ind. m-2) der Polychaeta an den beprobten Stationen des Vigo-
Profils.
Die weniger abundanten Familien (Ampharetinae, Cirratulidae, Eunicidae, Onuphidae,
Phyllodocidae) sind außer den Glyceridae (93 Ind. m-2) mit 46 Ind. m-2 vertreten. An Station
24 wurden insgesamt 1157 Ind. m-2 nachgewiesen. Die Unterschiede in den
Gesamtindividuenzahlen der Polychaeta der Stationen des Vigo-Profils sind in Abbildung 7
dargestellt.
An den Stationen des Nazaré-Canyons zeigt sich ein anderes Bild. Die Ergebnisse sind in
Abbildung 8 dargestellt. Zum einen wurde, wie oben erwähnt, an Station 27 keine Fauna 26
Ergebnisse
St.25 (3514m)
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Indi
vidu
en (n
/m²)
Ampharetinae
Cossuridae
Glyceridae
Goniodidae
Lumbrineridae
Paraonidae
Spionidae
B
St.26 (2894m)
0
20000
40000
60000
80000
100000
120000
140000
Indi
vidu
en (n
/m²)
Cossuridae
A
Vergleich der Gesamtindividuen der Polychaeta an den Stationen des Nazaré-Canyons
2857 0
116738
0
20000
40000
60000
80000
100000
120000
140000
26 25 27
Stationen
Indi
vidu
en (n
/m²)
C
Abb.8: Verteilung der Polychaeten-Familien (Individuen/m²) an den beprobten Stationen 25 (A)
und 26 (B) des Nazaré-Canyons und der Vergleich der Gesamtindividuenzahlen (C) im
Nazaré-Canyon. 27

Ergebnisse
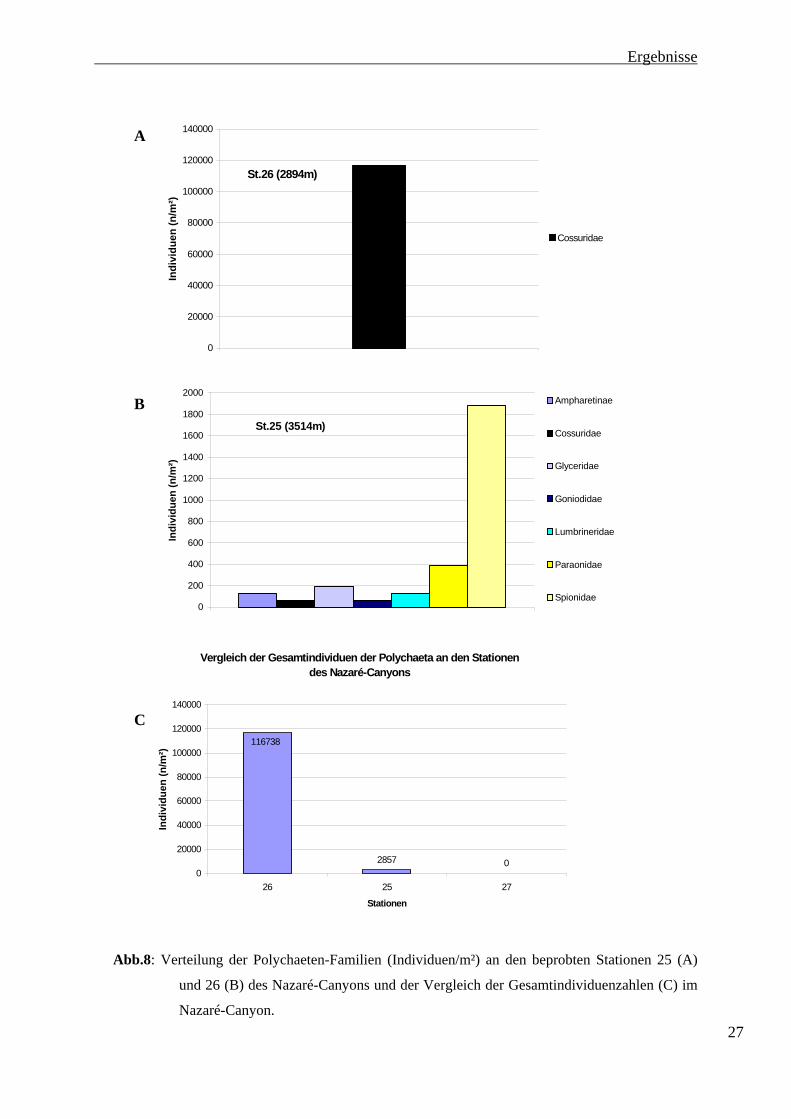
nachgewiesen. Zum anderen unterscheiden sich die Abundanzen der Familien deutlich von
denen des Vigo-Profils.An der flachsten Station des Nazaré-Canyons (Station 26) wurde nur
eine Polychaeten-Familie gefunden. Die Cossuridae treten hier in einer Dichte von 116738
Ind. m-2 auf und sind nach den Foraminifera an Station 25 die Gruppe mit den höchsten
ermittelten Individuenzahlen im gesamten Untersuchungsgebiet. An Station 25 wurden 7
Familien gezählt, von denen die Spionidae (1883 Ind. m-2) die höchsten Individuenzahlen
aufweisen. Sie setzen sich damit deutlich von den übrigen Familien an Station 25. ab. Die
Paraonidae kommen in einer Dichte von 390 Ind. m-2 vor. Die Glyceridae wurden mit 195
Ind. m-2 gezählt. Ampharetinae und Lumbrineridae sind mit je 130 Ind.-2 vertreten, Cossuridae
und Goniodidae mit je 65 Ind. m-2. Die Verteilung der Gesamtindividuenzahlen der
Polychaeta an den Stationen des Nazaré-Canyons sind ebenfalls in Abbildung 8 dargestellt.
3.1.4 Einteilung der Polychaeta nach Ernährungstypen
Die an den Stationen des Probennahmegebietes gefundenen Polychaeten-Familien, die
jeweiligen Ernährungstypen und die Anzahl der Arten der Familien sind in Tabelle 3
aufgelistet. Die Einteilung in Ernährungstypen erfolgt nach FAUCHALD und JUMARS
(1979).
Die Ampharetinae sind eine Unterfamilie der Ampharetidea. Sie sind sessiele Oberflächen-
Depositfresser (s) und mit Tentakeln ausgestattet. Oberflächen-Depositfresser werden auch
als Grenzschichtfresser bezeichnet (TAGHON und GREEN, 1992 aus THOMSEN, 1998). Sie
leben in Röhren und nehmen mit ihren Tentakeln Partikel aus nächster Umgebung auf, oder
wechseln je nach hydrodynamik zu passiven Suspensionfressen. Die Amphinomidae sind
carnivore, frei bewegliche (m) mit einem vorstülpbaren Rüssel (x) ausgestattete Polychaeta.
Die Capitellidae sind grabende (b) Depositfresser (s), die sich durch das Sediment bewegen
und auch Partikel von der Sedimentoberfläche fressen. Auch sie sind mit einem vorstülpbaren
Rüssel ausgestattet (x). Die Cirratulidae sind Oberflächen-Depositfresser (s), mit Tentakeln
ausgestattet. Je nach Art sind sie sessiel, bedingt beweglich (d) oder frei beweglich (m). Die
Cossuridae sind grabende (b), frei bewegliche (m) mit einem vorstülpbaren Rüssel
ausgestattete Polychaeta (x), die sich durch das Sediment graben. Die Eunicidae sind
omnivore Polychaeta, d.h. einige Arten sind herbivor andere carnivor. Ein Teil der Eunicidae
bewegt sich frei (m) grabend (b) durch das Sediment oder auf dem Sediment. Andere Arten
der Eunicidae sind weniger frei beweglich (d). Sie sind alle mit Kiefern ausgestattet (j). Die 28

Ergebnisse
Flabelligeridae sind eine ähnlich vielschichtige Familie wie die Eunicidae. Sie sind mit
Tentakeln ausgestattet und größtenteils frei beweglich (m), mit einigen weniger beweglichen
(d) Arten. Zum einen sind sie Filtrierer und zum anderen Depositfresser (s).
Tab.3: Auflistung der an den Stationen des Probennahmegebietes gefundenen Polychaeten-Familien,
ihre Ernährungstypen und die Anzahl der Arten die diese Familien umfassen. Die Bedeutung
der Abkürzungen der Ernährungstypen sind im Kasten unter der Tabelle 3 aufgelistet.
Familien Ernährungstypen Anzahl der Arten Ampharetidae sST 204 Amphinomidae cmx 110
Capitellidae smx, bmx 134 Cirratulidae sST, sdT, smT 134 Cossuridae bmx 15 Eunicidae hmj, hdj, cmj, cdj, bmj 241
Flabelligeridae FdT, smT, sdT 126 Glyceridae cdj, bmj 72 Goniodidae cdj 93 Hesionidae hmj, cmj, smj, bmj 130
Lacydoniidae bmx 8 Lumbrineridae hmj, cmj, cdj, bmj 188 Magelonidae sdT 35 Maldanidae bSx 218 Onuphidae hdj,cmj, cdj, Fdp, sdj 190 Paraonidae hmx, smx 50
Phyllodocidae cmx, bmx, cms 295 Sabellidae FST, sdT 292 Spionidae FdT, sdT 275 Syllidae hmj, cmj 602
Terbellidae sdT, sST 357
b ⇒ grabende Depositfresser c ⇒ carnivor F ⇒ Filtrierer (Suspensionsfresser) h ⇒ herbivor s ⇒ Oberflächen-Depositfresser o. Grenzschichtfresser d ⇒ bedingt beweglich m ⇒ frei beweglich S ⇒ sessiel j ⇒ mit Kiefern p ⇒ pumpend T ⇒ mit Tentakeln (Suspensionsfresser) x ⇒ andere Strukturen, meist feste vorstülpbare Rüssel
Buchstaben in Position 1
Buchstaben in Position 3
Buchstaben in Position 2
29

Ergebnisse
Die Glyceridae sind (c)arnivore, die sich je nach Art mal mehr, mal weniger beweglich (m &
d) durch das Sediment graben (b). Ebenso verhält es sich bei den Goniodidae. Beide Familien
haben Kiefer (j). Die Hesionidae sind frei bewegliche (m) und grabende (b) mit Kiefren (j)
ausgestattete omnivore (c & h) Depositfresser (s). Die Lacydoniidae graben (b) sich frei
beweglich (m) durch das Sediment und besitzen Kiefer (j). Die Lumbrineridae sind frei
bewegliche (m) und grabende (b) mit Kiefren (j) ausgestattete Omnivore (c & h).
Die Magelonidae sind mit Tentakeln ausgestattete bedingt bewegliche (d) Oberflächen-
Depositfresser (s). Die Maldanidae sind grabende (b) sessiele Depositfresser (s). Die
Onuphidae sind die facettenreichsten der gefundenen Familien. Sie sind in der Regel alle mit
Kiefern (j) ausgestattet und bedingt frei beweglich (d). Sie sind omnivor (c & h) und können
(F)iltrieren, pumpen und depositionsfressen (s). WILDISH und KRISTMANSON (1997)
bezeichnen solche „allround-Künstler“ als Grenzschichtfresser (aus THOMSEN 1998). Die
Paraonidae sind frei bewegliche (m) mit einem starken vorstülpbaren Rüssel (x) ausgestattete
(h)erbivore.
Die Autoren gehen hier jedoch von nur 1 Art aus obwohl die Paraonidae 50 Arten umfassen.
RÖDER (1971) beschrieb, dass Paraonidae durchaus Foraminfera oder kleine Crustacea
fressen. Die Phyllodocidae sind carnivore frei bewegliche (m) Depositfresser (s) und Jäger
(mit Rüssel x). Die Sabellidae sind meist sessiele Filtrierer, die mit ihren Tentakeln Partikel
fangen. Es gibt jedoch auch bedingt bewegliche (d) Depositfresser (s), die sich langsam durch
das Sediment bewegen. Die Spionidae sind bedingt beweglich (d). Sie besitzen Tentakeln mit
denen sie (F)iltrieren oder Deposit-Fressen (s) betreiben. Die Syllidae sind mit Kiefern
ausgestattete (j) gut bewegliche (m) Allesfresser (c & h). Die Terbellidea haben Tentakeln.
Sie sind sessiele oder bedingt bewegliche (d) Depositfresser (s).
3.1.5 Die Polychaeta der einzelnen Stationen und ihre Ernährungstypen
Für die Charakterisierung der einzelnen Stationen werden hier nochmal die Familien der
beprobten Stationen und ihre Ernährungstypen aufgestellt. In Tabelle 4 sind die Ergebnisse
der Einteilung nach FAUCHALD und JUMARS (1979) von Station 20 aufgestellt. Die
dominantesten Familien an Station 20 sind die Paraonidae, Spionidae und Ampharetidae. Sie
sind, außer den Ampharetidae, mehr oder minder frei beweglich und mit Tentakeln oder
einem starken vorstülpbaren Rüssel ausgfestattet. Die Ampharetidae sind sessiel und leben in
Röhren. Die Spionidae können Filtrieren (suspensionsfressen) und depositfressen
(Grenzschichtfresser). Die Cirratulidae, Capitellidae und Magelonidae sind mit über 50 Ind. 30

Ergebnisse
m-2 auch noch häufiger vertreten. Die Capitellidae sind eher grabende Depositfresser. Die
Cirratulidae und Magelonidae können neben depositfressen mit ihren Tenatakeln auch
suspensionsfressen.
Tab. 4: Polychaeten-Familien von Station 20 ihre Ernährungstypen und die gefundenen Individuenzahlen pro Quadratmeter.
Familien Ind.m-2 Ernährungstypen Ampharetidae 232 sST Amphinomidae 4 cmx
Capitellidae 56 smx, bmx Cirratulidae 100 sST, sdT, smT Cossuridae 32 bmx Eunicidae 12 hmj, hdj, cmj, cdj, bmj Glyceridae 12 cdj, bmj Goniodidae 24 cdj Hesionidae 4 hmj, cmj, smj, bmj
Lumbrineridae 32 hmj, cmj, cdj, bmj Magelonidae 60 sdT Maldanidae 4 bSx Onuphidae 36 hdj,cmj, cdj, Fdp, sdj Paraonidae 544 hmx, smx Sabellidae 12 FST, sdT Spionidae 264 FdT, sdT
Terbellidae 12 sdT, sST Die Ernährungstypen sind an Station 20 homogen verteilt. Es kommen Depositfresser sowie
Suspensionfresser vor. Die Tiere sind zum einen mit Tentakeln zum anderen mit festen
vorstülpbaren Rüsseln ausgesttattet. Das Verhältniss zwischen kiefer- rüssel- oder
tentakeltragenden Tieren ist relativ ausgeglichen.
In Tabelle 5 sind die Ergebnisse der Einteilung nach FAUCHALD und JUMARS (1979) von
Station 11 aufgestellt.
Tab. 5: Polychaeten-Familien von Station 11, ihre Ernährungstypen und die gefundenen Individuenzahlen pro Quadratmeter.
Familien Ind. / m² Ernährungstypen Ampharetidae 32 bmT, smT Amphinomidae 88 cmx
Capitellidae 96 smx, bmx Cirratulidae 16 sST, sdT, smT Cossuridae 8 bmx Glyceridae 48 cdj, bmj
Lacydoniidae 8 bmx Lumbrineridae 16 hmj, cmj, cdj, bmj Magelonidae 152 sdT
31
Ergebnisse
Die Magelonidae sind hier die dominanteste Familie. Sie sind eine von 3 tentakeltragenden
Familien an dieser Station. Nur Ampharetidae und Cirratulidae sind ebenfalls mit Tentakeln
ausgestattet und somit auch fähig, suspensionsfressen zu betreiben. Ansonsten überwiegen
bewegliche mit Kiefern oder Rüsseln ausgestattete Depositfresser, wie Capitellidae,
Amphinomidae, oder Glyceridae. Die restlichen eher gering vertretenen Familien sind
größtenteils auch grabende Depositfresser.
In Tabelle 6 sind die Ergebnisse der Einteilung nach FAUCHALD und JUMARS (1979) von
Station 24 aufgestellt.
Tab. 6: Polychaeten-Familien von Station 24, ihre Ernährungstypen und die gefundenen
Individuenzahlen pro Quadratmeter.
Familien Ind. / m² Ernährungstypen Ampharetidae 46 bmT, smT Cirratulidae 46 sST, sdT, smT Eunicidae 46 hmj, hdj, cmj, cdj, bmj Glyceridae 93 cdj, bmj
Lumbrineridae 231 hmj, cmj, cdj, bmj Maldanidae 185 bSx Onuphidae 46 hdj,cmj, cdj, Fdp, sdj Paraonidae 231 hmx, smx
Phyllodocidae 46 cmx, bmx, cms Spionidae 185 FdT, sdT
An dieser Station stellen Lumbrineridae und Paraonidae die dominantesten Familien. Beide
Familien sind entweder mit Rüsseln oder Kiefern ausgestattete, bewegliche, omnivore
Depositfresser. Die Spionidae sind von den Suspensionsfressern am häufigsten vorhanden.
Sessiele Familien gibt es wenige. Nur die Maldanidae, die auch recht stark vertreten sind, und
einige Cirratulidae sind sessiel. Auch an dieser Station kommen größtenteils bewegliche und
grabende Depositfresser vor.
In Tabelle 7 sind die Ergebnisse der Einteilung nach FAUCHALD und JUMARS (1979) von
Station 26 aufgestellt.
Tab. 7: Polychaeten-Familien von Station 26, ihre Ernährungstypen und die gefundenen
Individuenzahlen pro Quadratmeter.
Familien Ind. / m² Ernährungstypen Cossuridae 116738 bmx
An Station 26 wurden nur die Cossuridae nachgewiesen. Sie sind grabende Formen, mit
einem Rüssel ausgestattet.
In Tabelle 8 sind die Ergebnisse der Einteilung nach FAUCHALD und JUMARS (1979) von
Station 25 aufgestellt. 32

Ergebnisse
Mit 1883 Ind. m-2 ist die Spionidae an Station 25 die dominanteste Familie. Sie sind mit
Tentakeln ausgestattete Grenzschichtfresser. Sie können Suspensions- sowie Depositfressen.
Die Paraonidae ist die zweithäufigste Familie. Sie sind mit Rüsseln ausgestattete,
warscheinlich omnivore Depositfresser. Die Glyceridae sind carnivore, grabende
Depositfresser, ebenso wie die omnivoren Lumbrineridae. Die einzige Famile, die neben den
Spionidae noch als Suspensionfresser in Frage kommt, ist die mit Tentakeln ausgestattete
Ampharetidea. Cossuridae und Goniodidae kommen am wenigsten vor und zählen auch zu
den grabenden Depositfressern.
Tab. 8: Polychaeten-Familien von Station 25, ihre Ernährungstypen und die gefundenen
Individuenzahlen pro Quadratmeter.
Familien Ind. / m² Ernährungstypen Ampharetidae 130 bmT, smT
Cossuridae 65 bmx Glyceridae 195 cdj, bmj Goniodidae 65 cdj
Lumbrineridae 130 hmj, cmj, cdj, bmj Paraonidae 390 hmx, smx Spionidae 1883 FdT, sdT
Alles in allem wurden auch an Station 25 größtenteils Depositfresser vorgefunden. Jedoch
sind die Spionidae als Suspensionsfresser eindeutig häufiger als die übrigen depositfressenden
Familien vertreten.
Insgesamt wurden an den beprobten Stationen des Versuchsgebietes eher depositfressende
Familien nachgewiesen als Suspensionsfresser.
3.1.6 Betrachtung der Gesamtmakrofauna (Ind. m-2 und Prozentuale
Verteilung)
Die Gesamtindividuenzahlen der Makrofauna an den beprobten Stationen im Rahmen dieser
Arbeit sind in Abbildung 9 dargestellt. Hierbei wird auf die Nematoda und Foraminifera
verzichtet. Eine Bearbeitung der beiden Gruppen hätte den Rahmen dieser Arbeit gesprengt,
da sie speziell behandelt werden müssen. Die hier dargestellten Ergebnisse der Nematoda
sowie der Foraminifera basieren auf wenigen Individuenfunden. Diese vereinzelten Tiere
blieben im Sediment hängen oder auf dem verwendeten 500 µm Sieb (z.B. Nematoda die
länger als 500 µm sind). Jedoch werden hier bei der Gesamtbetrachtung die Polychaeta als
wichtiger Bestandteil der Benthosgemeinschaft mit einbezogen. Die Gesamtindividuenzahl 33
Ergebnisse
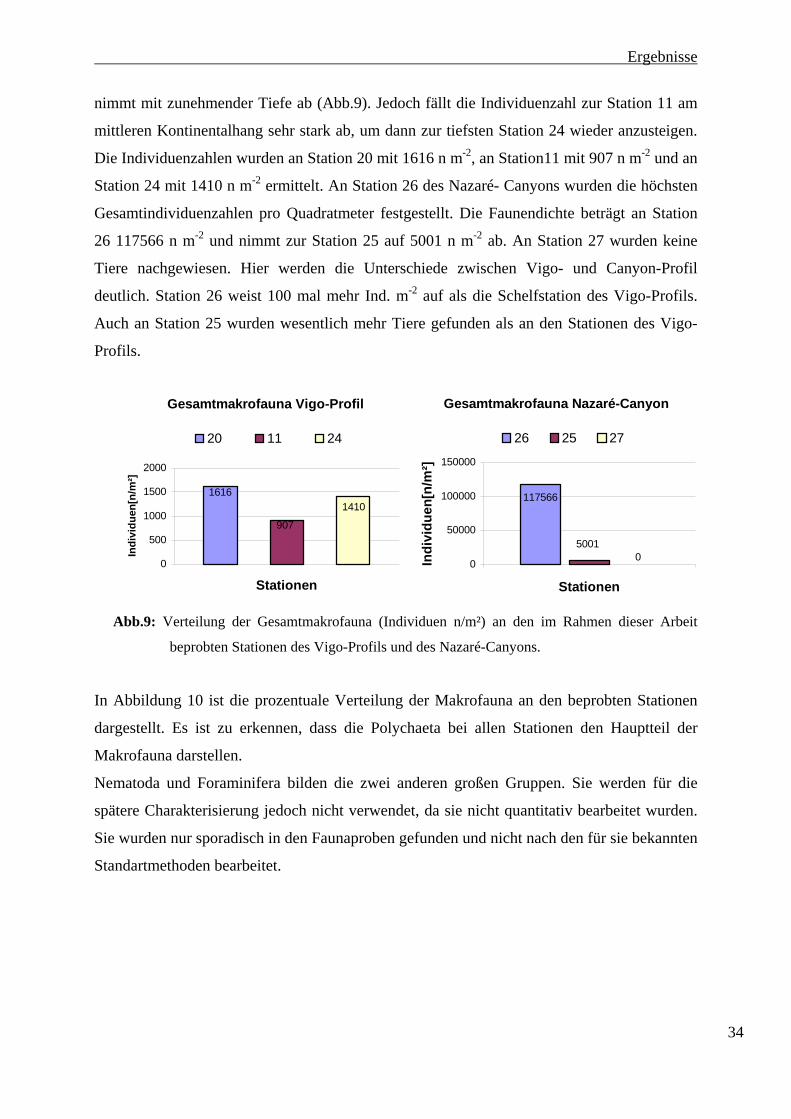
nimmt mit zunehmender Tiefe ab (Abb.9). Jedoch fällt die Individuenzahl zur Station 11 am
mittleren Kontinentalhang sehr stark ab, um dann zur tiefsten Station 24 wieder anzusteigen.
Die Individuenzahlen wurden an Station 20 mit 1616 n m-2, an Station11 mit 907 n m-2 und an
Station 24 mit 1410 n m-2 ermittelt. An Station 26 des Nazaré- Canyons wurden die höchsten
Gesamtindividuenzahlen pro Quadratmeter festgestellt. Die Faunendichte beträgt an Station
26 117566 n m-2 und nimmt zur Station 25 auf 5001 n m-2 ab. An Station 27 wurden keine
Tiere nachgewiesen. Hier werden die Unterschiede zwischen Vigo- und Canyon-Profil
deutlich. Station 26 weist 100 mal mehr Ind. m-2 auf als die Schelfstation des Vigo-Profils.
Auch an Station 25 wurden wesentlich mehr Tiere gefunden als an den Stationen des Vigo-
Profils.
Gesamtmakrofauna Vigo-Profil
1616
907
1410
0
500
1000
1500
2000
Stationen
Indi
vidu
en[n
/m²]
20 11 24
Gesamtmakrofauna Nazaré-Canyon
117566
50010
0
50000
100000
150000
Stationen
Indi
vidu
en[n
/m²]
26 25 27
Abb.9: Verteilung der Gesamtmakrofauna (Individuen n/m²) an den im Rahmen dieser Arbeit
beprobten Stationen des Vigo-Profils und des Nazaré-Canyons.
In Abbildung 10 ist die prozentuale Verteilung der Makrofauna an den beprobten Stationen
dargestellt. Es ist zu erkennen, dass die Polychaeta bei allen Stationen den Hauptteil der
Makrofauna darstellen.
Nematoda und Foraminifera bilden die zwei anderen großen Gruppen. Sie werden für die
spätere Charakterisierung jedoch nicht verwendet, da sie nicht quantitativ bearbeitet wurden.
Sie wurden nur sporadisch in den Faunaproben gefunden und nicht nach den für sie bekannten
Standartmethoden bearbeitet.
34

Ergebnisse
Prozentuale Verteilung (/m²) der Gesamtmacrofauna
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
20 11 24 26 25 27
Stationen
Proz
entu
aler
Ant
eil e
iner
Gru
ppe
ForaminiferaMolluscaPolychaetaPoriferaAnthozoaSipunculidaEchinodermataCrustaceaNematoda
Abb.10: Prozentuale Verteilung pro m² der Hauptgruppen der gefundenen Fauna an den beprobten
Stationen im Rahmen dieser Arbeit.
Die Foraminifera sind nur an den Stationen 11, 24 und 25 in größerer Zahl gefunden worden.
Die Crustacea und die Mollusca wurden an den Stationen 11-25 in mittleren Individuenzahlen
gefunden. Porifera, Anthozoa, Echinodermata und Sipunculida wurden jeweils nur an
einzelnen Stationen gefunden.
3.2 Biomasse
Die Ergebnisse der Biomassebestimmung ( in g C m-2) des Vigo-Profils sind in Abbildung 11
dargestellt, die des Nazaré-Canyons in Abbildung 12. An der Schelf-Station 20 (217m) wurde
die höchste Biomasse am Vigo-Profil nachgewiesen (0,244 g C m-2). Der Gehalt an Biomasse
nimmt mit der Tiefe kontinuierlich ab. An Station 11 (1945m) wurden 0,223 g C m-2 und an
Station 24 (2763m) 0,212 g C m-2 ermittelt. An den Stationen des Nazaré-Canyons ist die
Verteilung der Makrofauna-Biomasse anders (Abbildung12).
An der flachsten Station des Canyon-Profils (Station26) in 2894 m Tiefe wurde mit 0,217 g C
m-2 ähnlich viel Biomasse nachgewiesen wie an der tiefsten Station (Station 24) des Vigo-
Profils. Zur Station 25 in 3514m Tiefe nimmt die Biomasse jedoch um den Faktor 6 zu und
steigt auf 1,271 g C m-2. 35

Ergebnisse
0
0,05
0,1
0,15
0,2
0,25
0,3
Vigo-Profil
g C
/ m
²
20 (217m) 11 (1945m) 24 (2763m)
Abb.11: Biomasse in g C m-2 der beprobten Stationen des Vigo-Profils.
Wie oben bereits erwähnt, wurde an Station 27 keine Fauna gefunden. Es konnte somit im
Rahmen dieser Arbeit keine Makrofauna-Biomasse an Station 27 nachgewiesen werden. Wie
schon bei den Individuenzahlen wird hier der Unterschied zwischen den beiden Transekten
deutlich. Der Canyon setzt sich durch höhere Biomassen vom Hang-Profil vor Vigo ab.
Wie oben bereits erwähnt, wurde an Station 27 keine Fauna gefunden. Es konnte somit im
Rahmen dieser Arbeit keine Makrofauna-Biomasse an Station 27 nachgewiesen werden. Wie
schon bei den Individuenzahlen wird hier der Unterschied zwischen den beiden Transekten
deutlich. Der Canyon setzt sich durch höhere Biomassen vom Hang-Profil vor Vigo ab.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
Nazaré-Canyon
C.o
rg (g
/m²)
26 (2894m) 25 (3514m) 27 (4121m)
Abb.12: Biomasse in g C m-2 der beprobten Stationen des Nazaré-Canyons. Abb.12: Biomasse in g C m-2 der beprobten Stationen des Nazaré-Canyons.
3.3 Chlorophyll a 3.3 Chlorophyll a
In Abbildung 13 sind die Ergebnisse der Chlorophyll a Bestimmung, sowie die ermittelten
Zerfallskonstanten und Halbwertzeiten dargestellt.
In Abbildung 13 sind die Ergebnisse der Chlorophyll a Bestimmung, sowie die ermittelten
Zerfallskonstanten und Halbwertzeiten dargestellt.
An den Stationen des Vigo-Profils nimmt die Konzentration von Chlorophyll a mit
zunehmender Wassertiefe ab. Die höchsten Chl.a Konzentrationen wurden an Station 20
(217m) gefunden. In der Sedimentschicht 0-0,5 cm beträgt die Chl.a Konzentration 0,319 µg
cm-3. Die Konzentration steigt dann in der Schicht 0,5-1 cm auf 0,352 µg cm-3 an.
An den Stationen des Vigo-Profils nimmt die Konzentration von Chlorophyll a mit
zunehmender Wassertiefe ab. Die höchsten Chl.a Konzentrationen wurden an Station 20
(217m) gefunden. In der Sedimentschicht 0-0,5 cm beträgt die Chl.a Konzentration 0,319 µg
cm-3. Die Konzentration steigt dann in der Schicht 0,5-1 cm auf 0,352 µg cm-3 an. 36

Ergebnisse
Vigo-Profil Nazaré-Canyon
a d
b e
Abb.13: Chlorophyll a Konzentrationen (µg cm-3) in den ersten 9 cm der Sedimente an den
beprobten Stationen des Vigo-Profils (a-c) und des Nazaré-Canyons (d-e). Die
Datenpunkte sind Mittelwerte aus drei Parallelen. Die Fehlerbalken zeigen die
Standartabweichung. In die Diagramme sind die Halbwertzeiten T 1/2 (d) und
Zerfallskonstanten Kd (d-1) für Chlorophyll a an der betreffenden Station eingefügt. 37

Ergebnisse
Nach leichtem Abfallen der Konzentration wird in 4 cm Tiefe nochmals ein Peak von 0,317
µg cm-3 erreicht. Die Chl.a Konzentration fällt in 8 cm Sedimenttiefe auf 0,292 µg cm-3 ab.
Die Zerfallskonstante von Chl.a an Station 20 beträgt 0,031 d-1 und die Halbwertzeit beläuft
sich auf 22,0 Tage. Der Inventar, µg Chl.a cm-2 von 0-9cm Sedimenttiefe, beträgt 2,16 ±
0,319 µg cm-2.
An Station 11 (1952m) ist die Chl.a Konzentration in den Oberflächenschichten des
Sediments mit 0,14 µg cm-3 (0-0,5 cm) bzw. 0,091 µg cm-3 (0,5-1 cm) geringer als auf dem
Schelf. Der Minimalwert ist bei einer Sedimenttiefe von 2,5 cm erreicht (0,06 µg cm-3). Von
hier nimmt die Chl.a Konzentration mit steigender Tiefe wieder zu. In der Sedimentschicht
bei 6 cm erreicht die Chl.a Konzentration einen Peak von 0,119 µg cm-3. Sie nimmt bis in 8
cm Tiefe nur noch auf 0,11 µg cm-3 ab. Die Zerfallskonstante beträgt 0,035 d-1 und die
Halbwertzeit 19,5 Tage. Der Inventar, µg Chl.a cm-2 von 0-9cm Sedimenttiefe, beträgt 0,653
± 0,225 µg cm-2.
An Station 24 (2765 m) wurden die geringsten Konzentrationen von Chlorophyll a im
gesamten Probennahmegebiet ermittelt. Die Konzentration nimmt von der
Sedimentoberfläche 0-0,5 cm (0,056 µg cm-3) mit steigender Sedimenttiefe kontinuierlich ab.
In einer Tiefe von 8 cm beträgt die Chl.a Konzentration nur noch 0,005 µg cm-3. Die
Zerfallskonstante beträgt 0,031 d-1. Die Halbwertzeit liegt bei 22,9 Tagen. Der Inventar, µg
Chl.a cm-2 von 0-9cm Sedimenttiefe, beträgt 0,232 ± 0,043 µg cm-2.
Die Stationen im Nazaré-Canyon weisen höhere Chlorophyll a Konzentrationen auf. Sie
liegen in den Oberflächenschichten des Sediments zwischen 0,42-0,46 µg cm-3 Chl.a und
somit um 0,1 µg cm-3 oder um den Faktor 1,5-6,5 höher als am Vigo-Profil obwohl sie in
größeren Wassertiefen liegen.
An Station 26 (2878m) des Nazaré-Canyons nimmt die Chl.a Konzentration innerhalb des
ersten Zentimeters von 0,463 auf 0,413 µg cm-3 ab. Sie steigt dann jedoch mit zunehmender
Sedimenttiefe von 0,541 µg cm-3 (1-2 cm) auf 0,595 µg cm-3 in 8 cm Tiefe. Die
Zerfallskonstante nimmt auf 0,026 d-1 ab. Die Halbwertzeit liegt bei 26,5 Tagen. Der Inventar, µg
Chl.a cm-2 von 0-9cm Sedimenttiefe, beträgt 3,208 ± 0,804 µg cm-2.
An Station 25 (3554 m) zeigt sich ein ähnliches Bild wie an Station 26. Die Chl.a
Konzentration in der Oberflächenschicht ist 0,463 µg cm-3. Es folgt ein Abfallen der
Konzentration auf 0,453 µg cm-3. In der Schicht 2-3 cm wurde ein Peak von 0,536 µg cm-3
gefunden. Die Chl.a Konzentration nimmt dann mit zunehmender Sedimenttiefe bis auf 0,389
µg cm-3 ab. Die Zerfallskonstante Kd ist an dieser Station sehr klein (0,011 d-1). Dies hat eine
38

Ergebnisse
sehr hohe Halbwertzeit von 65,3 Tagen zur Folge. Der Inventar, µg Chl.a cm-2 von 0-9cm
Sedimenttiefe, beträgt 3,119 ± 0,802 µg cm-2.
Die tiefste Station im Nazaré-Canyon (Station 27, 4121 m) zeichnet sich ebenfalls durch
Oberflächenkonzentrationen von über 0,4 µg cm-3 aus. In der Schicht 0-0,5 cm wurden 0,425
µg cm-3 nachgewiesen, gefolgt von einem Subsurface-Maximum bei 0,5-1 cm Sedimenttiefe
(0,454 µg cm-3). Die Chlorophyll a Konzentration wird mit zunehmender Sedimenttiefe
kontinuierlich kleiner. Bei 4 cm Sedimenttiefe tritt ein Peak von 0,345 µg cm-3 auf. In der
Schicht 7-9 cm liegt die Konzentration nur noch bei 0,009 µg cm-3. Da Station 27 die letzte zu
beprobende Benthosstation auf dieser Ausfahrt war und das Schiff bereits zwei Tage später in
Cádiz einlaufen musste, herrschte erheblicher Zeitmangel. Die Probenausbeute war somit sehr
gering, und es war nicht möglich, für diese Station eine eigene Zerfallfskonstante sowie
Halbwertzeit zu bestimmen. Es wurden daher die Werte von Station 25 übernommen (Kd=
0,011 d-1 und T1/2= 65,3 d). Der Inventar, µg Chl.a cm-2 von 0-9cm Sedimenttiefe, beträgt
2,241 ± 0,419 µg cm-2.
3.4 Bioturbation
Die Ergebnisse der Bioturbationsmodellierung sind in den folgenden Abbildungen dargestellt.
Es wird bei jeder Station das diffusionsanaloge Modell (Modell 2) einem nonlocal-Mixing-
Modell (3-5) gegenüber gestellt. Aus Platzgründen wird hier immer nur das Modell mit dem
besten Fit dargestellt. Die übrigen Ergebnisse der Bioturbationsmodellierung sind im Anhang
zu finden. In Abbildung 14 sind die Ergebnisse von Station 20 (217m) des Vigo-Profils
dargestellt. Dem diffusionsanalogen Modell 2 ist das Modell 5 gegenübergestellt.
Mit Modell 2 konnte kein guter Fit errechnet werden. Der gewonnene Fit gleicht einer
Geraden, die durch das Chlorophyll-Profil gelegt wurde. Der von Modell 2 berechnete Db-
Wert liegt bei 174 cm² d-1, und ist zu hoch. Der Oberflächenfluss beträgt 0,791 µg Chl.a cm-2
d-1. Dieser Oberflächenfluss wird benötigt um stady-state-Bedingungen zu haben. Db-Wert
und Oberflächenfluss sind unwarscheinlich hoch und nicht realistisch. Aus diesem Grund
muß hier ein anderes Modell verwendet werden.
Mit dem Modell 5 wurde ein besserer Fit errechnet. Das berechnete Profil passt zu dem
natürlichen Profilverlauf. Der Db-Wert liegt hier bei 2,1 cm² d-1. An Station 20 wurde mit
dem Modell 5 der passendste Fit aller Modelle berechnet. Hier konnte nonlocal-mixing
nachgewiesen werden. Der Oberflächenfluss beträgt 0,069 µg Chl.a cm-2 d-1. Der über das 39
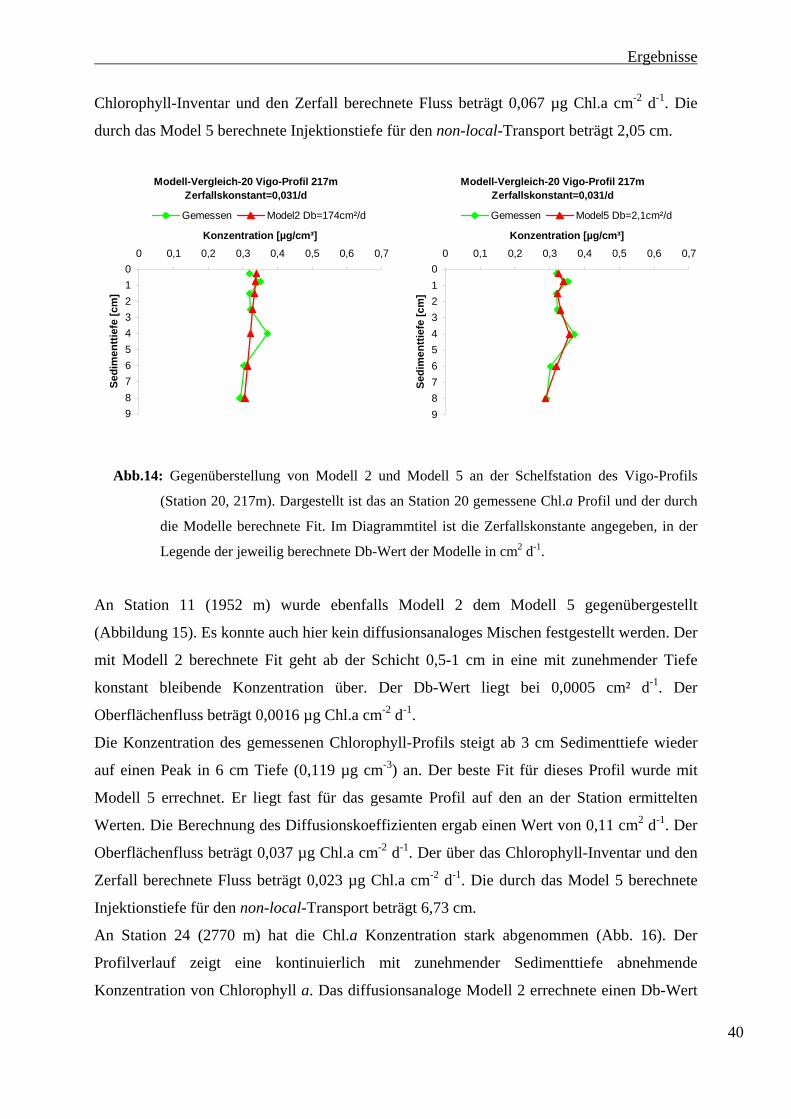
Ergebnisse
Chlorophyll-Inventar und den Zerfall berechnete Fluss beträgt 0,067 µg Chl.a cm-2 d-1. Die
durch das Model 5 berechnete Injektionstiefe für den non-local-Transport beträgt 2,05 cm.
Modell-Vergleich-20 Vigo-Profil 217mZerfallskonstant=0,031/d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model2 Db=174cm²/d
Modell-Vergleich-20 Vigo-Profil 217mZerfallskonstant=0,031/d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model5 Db=2,1cm²/d
Abb.14: Gegenüberstellung von Modell 2 und Modell 5 an der Schelfstation des Vigo-Profils
(Station 20, 217m). Dargestellt ist das an Station 20 gemessene Chl.a Profil und der durch
die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante angegeben, in der
Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
An Station 11 (1952 m) wurde ebenfalls Modell 2 dem Modell 5 gegenübergestellt
(Abbildung 15). Es konnte auch hier kein diffusionsanaloges Mischen festgestellt werden. Der
mit Modell 2 berechnete Fit geht ab der Schicht 0,5-1 cm in eine mit zunehmender Tiefe
konstant bleibende Konzentration über. Der Db-Wert liegt bei 0,0005 cm² d-1. Der
Oberflächenfluss beträgt 0,0016 µg Chl.a cm-2 d-1.
Die Konzentration des gemessenen Chlorophyll-Profils steigt ab 3 cm Sedimenttiefe wieder
auf einen Peak in 6 cm Tiefe (0,119 µg cm-3) an. Der beste Fit für dieses Profil wurde mit
Modell 5 errechnet. Er liegt fast für das gesamte Profil auf den an der Station ermittelten
Werten. Die Berechnung des Diffusionskoeffizienten ergab einen Wert von 0,11 cm2 d-1. Der
Oberflächenfluss beträgt 0,037 µg Chl.a cm-2 d-1. Der über das Chlorophyll-Inventar und den
Zerfall berechnete Fluss beträgt 0,023 µg Chl.a cm-2 d-1. Die durch das Model 5 berechnete
Injektionstiefe für den non-local-Transport beträgt 6,73 cm.
An Station 24 (2770 m) hat die Chl.a Konzentration stark abgenommen (Abb. 16). Der
Profilverlauf zeigt eine kontinuierlich mit zunehmender Sedimenttiefe abnehmende
Konzentration von Chlorophyll a. Das diffusionsanaloge Modell 2 errechnete einen Db-Wert
40
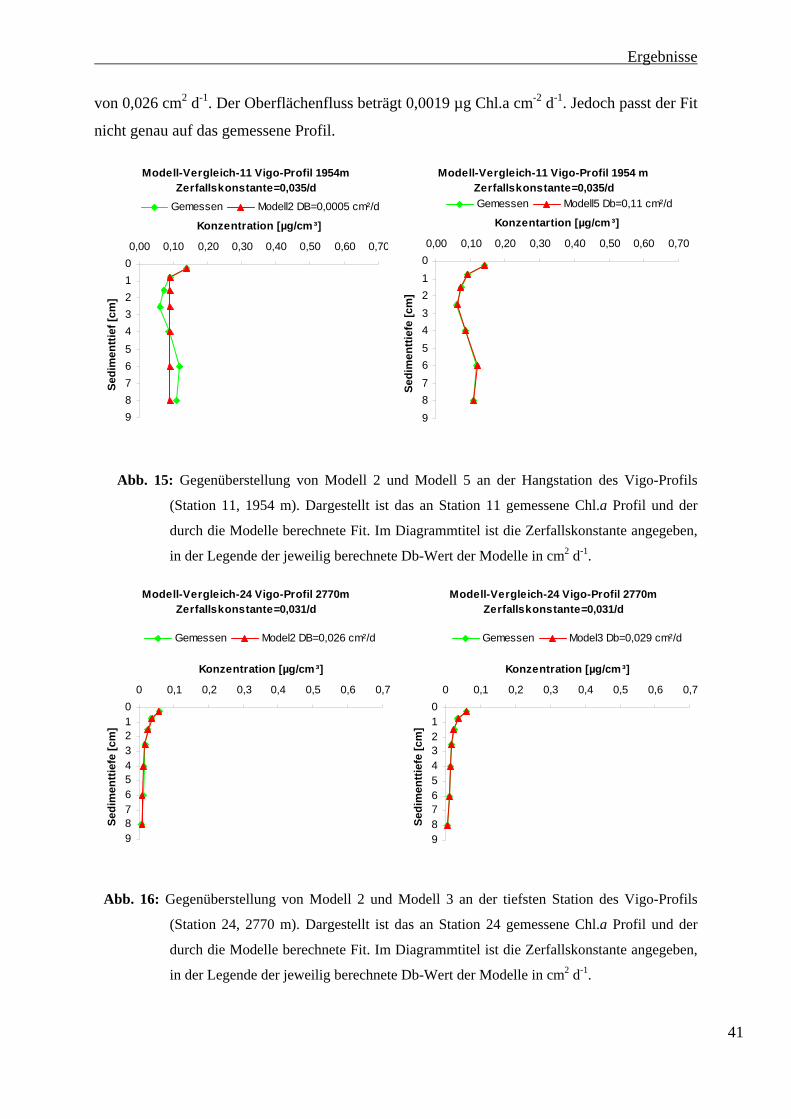
Ergebnisse
von 0,026 cm2 d-1. Der Oberflächenfluss beträgt 0,0019 µg Chl.a cm-2 d-1. Jedoch passt der Fit
nicht genau auf das gemessene Profil.
Modell-Vergleich-11 Vigo-Profil 1954mZerfallskonstante=0,035/d
0123456789
0,00 0,10 0,20 0,30 0,40 0,50 0,60 0,70
Konzentration [µg/cm³]
Sedi
men
ttief
[cm
]
Gemessen Modell2 DB=0,0005 cm²/d
Modell-Vergleich-11 Vigo-Profil 1954 mZerfallskonstante=0,035/d
0123456789
0,00 0,10 0,20 0,30 0,40 0,50 0,60 0,70
Konzentartion [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Modell5 Db=0,11 cm²/d
Abb. 15: Gegenüberstellung von Modell 2 und Modell 5 an der Hangstation des Vigo-Profils
(Station 11, 1954 m). Dargestellt ist das an Station 11 gemessene Chl.a Profil und der
durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante angegeben,
in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
Modell-Vergleich-24 Vigo-Profil 2770mZerfallskonstante=0,031/d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model2 DB=0,026 cm²/d
Modell-Vergleich-24 Vigo-Profil 2770mZerfallskonstante=0,031/d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model3 Db=0,029 cm²/d
Abb. 16: Gegenüberstellung von Modell 2 und Modell 3 an der tiefsten Station des Vigo-Profils
(Station 24, 2770 m). Dargestellt ist das an Station 24 gemessene Chl.a Profil und der
durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante angegeben,
in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
41

Ergebnisse
Ab ca. 4 cm Sedimenttiefe weicht das berechnete Profil leicht von dem an der Station
gemessenen Profil ab. Modell 3 hingegen lieferte einen fast identischen Profilverlauf. Die
Unterschiede zu Modell 2 sind nicht gravierend. Dies spiegelt sich auch im durch Modell 3
berechneten Db-Wert wider (0,029 cm2 d-1). Er liegt nur 0,003 cm2 d-1 höher als der Db von
Modell 2. Der Oberflächenfluss beträgt 0,0021 µg Chl.a cm-2 d-1. Der von Model 3 berechnete
Injektionsfluss beträgt 0,0007 µg cm-2 d-1. Addiert man Oberflächenfluss und Injektionsfluss
bekommt man einen Gesamtfluss von 0,0028 µg cm-2 d-1. Der über das Chlorophyll-Inventar
und den Zerfall berechnete Fluss beträgt 0,007 µg Chl.a cm-2 d-1. Die durch das Model 3
berechnete Injektionstiefe für den non-local-Transport beträgt 3,09 cm. Hier kann
diffusionsanaloges mit einem geringen Beitrag durch nonlocal Mischen angenommen werden.
An der flachsten Station (Station 26) des Nazaré-Canyons steigt die Chl.a Konzentration mit
zunehmender Sedimenttiefe. Mit Modell 2 (Abbildung 17) wurde ein Db-Wert von 2,55 1028
cm-2 d-1 berechnet. Der Oberflächenfluss beträgt 1,89 1010 µg Chl.a cm-2 d-1.
Das ermittelte Profil bleibt mit der Sedimenttiefe konstant. Es konnte hier kein adäquater Fit
berechnet werden. Auf Grund der problematischen Tracerverteilung im Sediment (Zunahme
der Konzentration mit zunehmender Sedimenttiefe) war es schwierig, mit den verwendeten
Modellen einen guten Fit zu erlangen. Den besten Fit errechnete das Modell 4b. Jedoch liegt
der Fit nur in den ersten 1,5 cm auf dem gemessenen Profil der Station.
Modell-Vergleich-26 Nazaré-Canyon 2800mZerfallskonstante=0,026 /d
01234
56789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model4b Db=0,015 cm²/d
Modell-Vergleich-26 Nazaré-Canyon 2800mZerfallskonstante=0,026 /d
01
23
45
67
89
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model2 Db=2,55 E28 cm²/d
Abb. 17: Gegenüberstellung von Modell 2 und Modell 4b an der flachsten Station des Nazaré-
Canyons (Station 26, 2800 m). Dargestellt ist das an Station 26 gemessene Chl.a Profil
und der durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante
angegeben, in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
42

Ergebnisse
In den unteren Sedimentschichten bleibt er von 2,5 cm bis 6 cm mit der Tiefe konstant. Erst
bei 8 cm liegen die beiden Profile wieder übereinander. Der Db-Wert bträgt 0,007 cm2 d-1.
Der Oberflächenfluss beträgt 0,094 µg Chl.a cm-2 d-1. Der über das Chlorophyll-Inventar und
den Zerfall berechnete Fluss beträgt 0,083 µg Chl.a cm-2 d-1. Die durch das Model 4b
berechnete Injektionstiefe für den non-local-Transport beträgt 9,82 cm. Allein durch
Betrachtung des Chlorophyll a Profils müsste hier von nonlocal-mixing ausgegangen werden.
An Station 25 (3500 m) wurde mit Modell 2 ebenfalls kein adäquater Fit errechnet (Abb.18).
Der ermittelte Db-Wert beträgt 19,85 cm2 d-1. Der Oberflächenfluss beträgt 0,23 µg Chl.a cm-
2 d-1. Die berechnete Kurve ist direkt durch das gemessene Profil gelegt. Das gemessene Profil
zeigt ein Subsurface-Maximum bei 2,5 cm (0,536 µg cm-3). Dieser Profilverlauf deutet auf
nonlocal-mixing hin. Das hierfür ausgewählte Modell 4b zeigt den optisch besten Fit. Es
liegen etwa 5 Punkte in direkter Nähe des gemessenen Profils (bei 1,5 cm, 2,5 cm, 4 cm und 8
cm). Der berechnete Db-Wert beträgt 0,007 cm2 d-1. Der Oberflächenfluss beträgt 0,0034 µg
Chl.a cm-2 d-1. Der über das Chlorophyll-Inventar und den Zerfall berechnete Fluss beträgt
0,034 µg Chl.a cm-2 d-1. Die durch das Model 4b berechnete Injektionstiefe für den non-local-
Transport beträgt 2,5cm.
Modell-Vergleich-25 Nazaré-Canyon 3500mZerfallskonstante=0,011 /d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model4b Db=0,007 cm²/d
Modell-Vergleich-25 Nazaré-Canyon 3500mZerfallskonstante=0,011 /d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model2 Db=19,85 cm²/d
Abb. 18: Gegenüberstellung von Modell 2 und Modell 4b an der mittleren Station des Nazaré-
Canyons (Station 25, 3500 m). Dargestellt ist das an Station 25 gemessene Chl.a Profil
und der durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante
angegeben ,in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
An Station 27 findet sich ein ähnliches Profil wie an Station 25 (Abb.19). Ein kleines
Subsurface- Maximum bei 0,75 cm Sedimenttiefe (0,454 µg cm-3) sowie ein weiteres 43

Ergebnisse
Maximum bei 4 cm (0,345 µg cm-3) wurde beobachtet. Dass hier kein diffusionsanaloges
Mischen angenommen werden kann zeigt auch der durch Modell 2 berechnete Fit. Wie bei
Station 25 geht hier eine Gerade durch das gemessene Chl.a Profil. Der Db-Wert liegt bei
0,17 cm2 d-1. Der Oberflächenfluss beträgt 0,021 µg Chl.a cm-2 d-1. Für die Berechnung des
nonlocal-mixing wurde hier Modell 3 ausgewählt, da es den besten Fit ermittelte.
Es liegen nur 3 Punkte in direkter Nähe zu den gemessenen Datenpunkten (bei 1,5 cm, 4 cm
und 8 cm). Jedoch liegt der Profilverlauf des Fit’s hier der gemessenen Kurve am nächsten.
Der Db-Wert 0,024 cm2 d-1 wurde ermittelt. Der Oberflächenfluss beträgt 0,0084 µg Chl.a cm-
2 d-1. Der von Model 3 berechnete Injektionsfluss beträgt 0,0117 µg cm-2 d-1. Addiert man
Oberflächenfluss und Injektionsfluss bekommt man einen Gesamtfluss von 0,02 µg cm-2 d-1.
Der über das Chlorophyll-Inventar und den Zerfall berechnete Fluss beträgt 0,025 µg Chl.a
cm-2 d-1. Die durch das Model 3 berechnete Injektionstiefe für den non-local-Transport beträgt
3,89 cm.
Modell-Vergleich-27 Nazaré-Canyon 4121mZerfallskonstante=0,011 /d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model2 Db=0,17cm²/d
Modell-Vergleich-27 Nazaré-Canyon 4121mZerfallskonstante=0,011 /d
0123456789
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
Konzentration [µg/cm³]
Sedi
men
ttief
e [c
m]
Gemessen Model3 Db=0,024cm²/d
Abb. 19: Gegenüberstellung von Modell 2 und Modell 3 an der tiefsten Station des Nazaré-
Canyons (Station 27, 4212 m). Dargestellt ist das an Station 27 gemessene Chl.a Profil
und der durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante
angegeben, in der Legende ist der jeweilig berechnete Db-Wert der Modelle angeführt in
cm2 d-1.
44

Diskussion
4 Diskussion 4.1 Der Kontinentalhang der Iberischen Westküste Schelfgebiete und Kontinentalhänge machen zusammen 15 % der gesamten Ozeanoberfläche
aus (WOLLAST, 1989). Sie sind trotz ihres geringen Flächenanteils mit 26-28 % an der
Produktion des globalen marinen partikulären Kohlenstoffes beteiligt (WOLLAST, 1991;
THOMSEN, 1998). Die hochproduktiven Schelfgebiete exportieren ihr Material über die
Kontinentalhänge in tieferes Wasser des offenen Ozeans (THOMSEN, 1998). Die
Topographie des Kontinentalhanges ist, neben der Sedimentation von organischem Material
aus dem Pelagial, ein weiterer Faktor der für die Versorgung der Tiefsee mit organischem
Material verantwortlich ist. THOMSEN (1998) unterscheidet 3 Typen von
Kontinentalhängen.
37
Abb.19: Darstellung der drei von THOMSEN (1998) untersuchten Kontinentalhangtypen. (aus
THOMSEN, 1998) Unter den jeweiligen abbildungen sind die durchschnittlichen Breiten
der Kontinentalhangtypen eingefügt.

Diskussion
Der Typ I entspricht dem SEEP-Arbeitsgebiet an der Ostküste der USA (siehe BISCAYE et
al. 1988; BISCAYE et al. 1994; ANDERSON et al. 1994). Typ II wäre das Goban-Spur-
Gebiet westlich Irlands am Keltischen Kontinentalhang (siehe THOMSEN, 1998).
Der Kontinentalhang vor Vigo ist nach ein Typ III Hang. Der schmale Hang (20-40 km) ist
hochenergetisch und es findet eine direkte Kopplung zwischen Schelf und Tiefsee statt. Es
können Ereignissgesteuert große Mengen akkumulierten Schelfamterials in die Tiefe
transportiert werden. Dieser Typ wird vor allem in Auftriebsgebieten vorgefunden. Ferner ist
der gesamte Kontinentalhang mit Canyon durchzogen, die Teilweise bis zur Küste reichen
und Verbindung mit Schluchten an Land zeigen. Diese Canyon oder schmalen Trogsysteme
verursachen oder unterstützen den aktiven Transport von Material in die Tiefe. Solche
Canyonsysteme gelten nach HUTHNANCE (1995) und MONACO et al. (1990) als sehr
bedeutende morphologiche Strukturen für den Transport von organischem Material in die
Tiefsee.
4.2 Charakterisierung des Vigo-Profils Der gängige Trend der exponentiellen Abnahme der Makrofaunadichte mit zunehmender
Wassertiefe und Entfernung von Land (SIBUET et al. 1989; FLACH & HEIP, 1996;
SOLTWEDEL, 1997; FLACH, et al. 1998; PFANNKUCHE, et al. 1998) konnte am Vigo-
Profil beobachtet werden. Die Faunenzusammensetzung (Abb. 4) entspricht den Ergebnissen
anderer Versuchsgebiete wie Goban Spur, dem Kontinentalhang Galiziens, (FLACH und
HEIP, 1996; FLACH et al., 1998; HEIP et al.,, 1999; SOLTWEDEL, 1997) oder des Cap-
Ferret Canyons (Biscaya) (SORBE, 1999). Auch die mit zunehmender Tiefe steigenden
Individuenzahlen der gefundenen Foraminiferen folgen den bisher bekannten Befunden aus
anderen Kontinentalhanguntersuchungen (Abb. 5) (HANSEN, 1965; MURRAY, 1970 a;
MURRAY, 1979; SAIDOVA, 1970; LANKFORD und PHLEGER, 1973 alle aus
NUGLISCH, 1985). Ihre Individuenzahlen steigen mit zunehmender Tiefe.
38
Es sollt hier jedoch vor weg gesagt werden, dass die ermittelten Makrofauna-Ergebnisse
alleine nicht ausreichen, die Stationen zu charakterisieren. Da für die Auswertung der
Faunaproben pro Station immer nur ein Kastengreifer zur Verfügung stand, besteht die
Möglichkeit, dass hier Bereiche im Sediment getroffen wurden, in denen die
Makrofaunadichte sehr gering ist. Aussagen zur Patchiness können hier nicht gemacht
werden.

Diskussion
Die Gesamtindividuenzahlen (Abb.9) an der Schelfstation (St.20) sind mit 1616 Individuen
pro Quadratmeter (Ind. m-2) für dieses Gebiet sehr gering. FLACH und HEIP (1996) wiesen
im Goban Spur-Gebiet zwischen 4000 und 8000 Ind. m-2 nach. Unseren Ergebnissen
ähnlichere Resultate erlangten FLACH et al., (1998) im OMEX II-II Gebiet (zwischen Küste
und Galcia-Bank) etwa 60 km nördlich unseres Versuchsgebietes. Dort wiesen sie in einer
Tiefe von 774m 1200 Ind. m-2 nach. Die Autoren begründen die geringe Makrofaunadichte
mit geringen Corg-Gehalten. FRANKE (2000) wies an den Stationen des Vigo-Profils
ebenfalls geringe Corg-Werte nach. An Station 20 ermittelte FRANKE (2000) Corg-Werte von
0,2-0,4 %, und relativ sandiges Sediment mit Korngrößen zwischen 125 µm und <63µm.
LOHSE et al.,, (1998) wiesen an den Schelfstationen im Goban-Spur-Gebiet auf
Porenwasserströme hin, die einen erhöhten Export von organischem Material zur Folge haben
können.
Nach BISCAYE et al., (1994) und LOHSE et al., (1998) weisen Schelf-Sedimente generell
weniger Kohlenstoffgehalte auf als Kontinetalhänge. Dies könnte auch der Grund für die
geringe Biomasse auf dem Schelf von 0,244 g C m-2 sein.
HUTHNANCE et al., (pers. Mitteilung) wiesen im Winter 97/98 Strömungs-
geschwindigkeiten von bis zu 45 cm s-1 am Schelf vor Kap Finnisteré nach. Diese
tidenabhängigen Strömungsgeschwindigkeiten können zu erheblicher Resuspension von
Material führen, das dann über die Schelfkante und den oberen Kontinentalhang in tieferes
Wasser transportiert werden kann. THOMSEN und VAN WEERING (1998) beschreiben am
Beispiel von Goban Spur, dass unter normalen Strömungsbedingungen
(Freistromgeschwindigkeit ≈ 20 cm s-1) die oberste lockere Sedimentschicht während nahezu
jedem Gezeitenzyklus resuspendiert wird. In Abb.20 veranschaulichen WILDISH und
KRISTMANSON (1997) den Einfluss von Hydrodynamik und Ernährungstyp. Die
Bodenschubspannungsgeschwindigkeit U* ist ein Mass für die Scherkräfte, die auf den
Meeresboden und somit auf alle aktiven Organissmen an der Sediment-Wasser-Grenzschicht
einwirken. U* ist abhängig von der Strömungsgeschwindigkeit und dem Abstand zum
Meeresboden. Je höher die Strömungsgeschwindigkeit, desto höher sind die Scheerkräfte (U*)
an der Sedimentoberfläche, und um so mehr wird resuspendiert (THOMSEN, 1998).
Die gemischten Ernährungstypen werden demnach in Regionen mit erhöhter Suspension
(großes U*) vorgefunden. Diese Tatsachen spiegeln sich auch in der Zusammensetzung der
Polychaeta an Station 20 wieder (siehe 3.1.5, Tab.4).
39

Diskussion
Abb. 20: Diagramm zur benthischen Gemeinschaftsstruktur an Kontinentalhängen entlang eines
Gradienten veränderter Strömungsbedingungen. U* = Bodenschubspannungs-
geschwindigkeit (aus THOMSEN, 1998 nach WILDISH und KRISTMANSON, 1997)
Die homogene Verteilung zwischen Oberflächen-Depositfressern (Grenzschichtfresser) und
Suspensionsfressern könnte somit ein Indiez für erhöhte Suspension sein Die dominantesten
Familien an Station 20 sind die Paraonidae, Spionidae und die Ampharetidae. Die Paraonidea
sind eher grabende Depositfresser, die selektiv Partikel fressen. Die Spionidae sind auf Grund
ihrer eingeschränkten Beweglichkeit und ihren Tentakeln mehr zu den Filtrieren, also den
Suspensionsfressern zu zählen. Für die Ampharetidae gilt das Gleiche, den auch sie sind mit
Tentakeln ausgestattet, zudem sind sie sessiel. Zu den Suspensionsfressern sind ebenfalls die
Cirratulidae und die Magelonidae zu zählen. Auch sie kommen an Station 20 häufiger vor.
Die restlichen Familien sind eher zu den Grenzschichtfressern zu zählen, wie vor allem die
Onuphidae, d.h. sie sind auch in der Lage Depositfressen zu betreiben.
40
In den meisten Fällen ist die umfangreiche Aufnahme und Wiederabgabe von Partikeln, im
Sediment oder an der Sedimentoberfläche (subsurface- oder surface-depositfeeding), die
vorherrschende Art des Partikelmischens (ALLER, 1982; THAYER, 1983, aus
WHEATCROFT, et al., 1990). Vor allem Depositfresser (ca. 80 % der Makrofauna der
Tiefsee, JUMARS und GALLAGHER, 1982) besiedeln Lebensräume, die sich durch
Nahrungsmangel kennzeichen. Das metabolisierbare Material macht zum Teil nur 5% der
aufgenommenen Sedimentmenge aus, und Depositfresser müssen daher sehr viel Sediment
pro Zeit aufnehmen (LOPEZ und LEVINTON, 1987). Weiterhin ist es für die Organissmen

Diskussion
vorteilhaft das partikuläre Material zu selektieren, und vornehmlich die feine
Sedimentfraktion aufzunehmen, da diese den größten Anteil an Corg aufweisen (MAHAUT
und GRAF, 1987; WHEATCROFT und JUMARS, 1987; SELF und JUMARS, 1988 aus
WHEATCROFT et al., 1990). So sorgen sie für einen starke Umverteilung von
Sedimentmaterial, vor allem von feinen Partikeln, die von der Sedimentoberfläche in tiefere
Sedimentschichten transportiert werden. Die Depositfresser stellen also ein wichtiges Glied in
der Kette des Kohlenstoffkreislaufs dar, sie jegliche Art von organischem Materail in das
Sediment verfrachten
Die Bioturbation wurde hier mit Chl. a Profilen modeliert. Die Menge der gefundenen Chl. a
Equivalente ist für dieses Gebiet gering (1,342 µg Chl. a cm-3 innerhalb des ersten
Sedimentzentimeters). PFANNKUCHE und SOLTWEDEL, (1998) wiesen im Goban Spur
Gebiet zwischen 1,0 und 8,0 µg Chl. a cm-3 innerhalb des ersten Sedimentzentimeters nach.
Auf Grund der höheren Primärproduktionsraten (250-350 mg C m-2 d-1) am Kontinentalen
Schelf vor Portugal im Gegensatz zu Goban Spur (100-200 mg C m-2 d-1) oder dem offenen
Ozean (100-150 mg C m-2 d-1)(ANTOINE et al. 1996), müssten höhere Chlorophyll a
Mengen im Sediment gefunden werden. Eine Ursache für die gegenteiligen Befunde könnten
die im Winter vorkommenden hohen Strömungsgeschwindigkeiten und die damit verbundene
Resuspension sein (siehe Oben). Eine andere Möglichkeit ist ein stärkerer Abbau des
Materials durch Zooplankton in den oberen Wasserschichten oder der schnelle Abbau durch
die Benthosgemeinschaft. Die für die Modellierung benötigte Halbwertzeit und
Zerfallskonstante von Chlorophyll a wurde experimentell ermittelt. Die Halbwertzeit von 22
Tagen und der Zerfall 0,031 d-1 decken sich mit Ergebnissen von GRAF (1989),
PFANNKUCHE und SOLTWEDEL (1998) sowie SUN et al. (1993).
41
Es sollte herausgefunden werden, ob an den Stationen diffusionsanaloges oder non-local
Mischen vorherrscht. Die hierfür verwendetetn Modelle 2 und 5 ergaben, dass non-local
Mischen vorherrscht. Das diffusionsanaloge Modell errechnette einen Fit der keinen der
gemessenen Punkte trifft. Nach BOUDREAU (1986b) kann bei non-local Mischen erst nach
langer Zeit ein diffusionsanloges Profil erreicht werden, d.h., dass der Prozess des Mischens
hier noch nicht abgeschlossen ist (GRAF, 1995). Der errechnete Db-Wert von 174 cm2 d-1
und der Oberflächenfluss von 0,791 µg cm-2 d-1 sind unrealistisch. Der Oberflächenfluss
übertrifft den Chlorophyll a Gehalt der ersten 0,5 cm des Sediments um über das Doppelte.
Die Werte sind um den Faktor 1000 größer als die gängigen Annahmen für flachere Gebiete
wie die Schelfmeere (Db für 234Th, zwischen 0,14 und 0,38 cm2 d-1 BOUDREAU, 1994).
GERINO et al., (1997) zeigten, dass mit dem Tracer Chl. a 2-3 mal höhere

Diskussion
Bioturbationskoeffizienten errechnet werden als mit dem radioaktiven Tracer 234Th. Das non-
local-Mixing-Modell 5 lieferte dgegen einen Db-Wert von 2,1 cm-2 d-1 und einen
Oberflächenfluss von 0,069 µg Chl.a cm-2 d-1. Dieser Fluss deckt sich fast mit dem über das
Chlorophyll Inventar und den Zerfall berechnetetn Chl.a-Fluss von 0,067 µg cm-2 d-1 und
wird benötigt um stady-state- Bedingungen zu erfüllen. Der berechnete Fit liegt fast genau auf
dem gemessenen Profil. Der Db-wert zeigt, daß hier eine hohe Mischungaktivität vorherrscht.
Dies deckt sich mit den gefundenen Polychaeten-Familien, von denen viele
Grenzschichtfresser sind und somit auch Depositfressen betreiben, was einen entscheidenden
Einfluss auf die Bioturbation hat (GRAF et al., 1995; SOETAERT et al.,, 1996). Die durch
die oben erwähnte Strömung resuspendierten Partikel werden von den Suspensionsfressern
aufgenommen, und können in tiefere Sedimentschichten transportiert werden. Hier werden
die abgegebenen Fecal-Pellets von den Deposit-Fressern weiterverarbeiet, und entweder noch
tiefer in das Sediment oder an die Sedimentoberfläche transportiert. Das Modell 5 ist das
klompizierteste Modell von SOETAERT et al., (1996). Es modelliert mit vier non-local-
Teilen in verschiedenen Sedimentschichten. Hier besteht nun die Möglichkeit den Fluss aus
der Schicht X0-0 in die Schicht L-δx – L+δx. (siehe 2.6.2, Model 5)zu modellieren. Bei dem
von uns ermittelten Chl.a Profil könnte der Fluss aus der Schicht um 0,5 cm Sedimenttiefe
stammen und in die Schicht um 3-5 cm gelangen. Die mit dem Model errechnete
Injektionstiefe von 2,05cm passt nicht ganz zu dem Profil.
42
Hier wird also Material aktiv von einer höher gelegenen Schicht in eine tiefere Transportiert,
was durch Depositfresser oder aber auch durch die an Station 20 gefundenen Thalassinidae
(Maulwurfskrebse) durchgeführt werden kann (ROWDEN und JONES ,1994 aus GRAF und
ROSENBERG, 1997). Durch die Menge der gefundenen Fauna, die geringen Mengen an
Chlorophyll a und den relativ hohen Db-Wert kann davon ausgegangen werden, dass das
Sediment sehr gut durchmischt ist. Da die Strömungen Tidenabhängig sind, kann es sein, dass
Grenzschichtfresser von Depositfressen zu Suspensionsfressen wechseln und umgekehrt. Dies
hat auch zur Folge, dass die Sedimentoberfläche sehr stark aufgelockert ist, wodurch
Resuspension begünstigt wird. Durch die Suspensionsfresser steigt der Fluss von organischem
Material in das Sediment erheblich an (GRAF, 1992) und kann dann durch Depositfressen
noch tiefer in das Sediment transportiert werden. Die gute Durchmischung des Sediments
zeigt sich auch in der gleichmäßigen Verteilung der Korngrößen bis in ein Tiefe von 9cm
(FRANKE, 2000), was nach FLACH und HEIP (1996b) und FLACH et al., (1998) auf
erhöhte Bioturbation oder auf gleichmäßigen Eintrag von sedimentiertem Material hinweist.
Hier sind beide Möglichkeiten vorstellbar. Auf Grund dieser Ergebnisse kann hier über ein

Diskussion
Exportgebiet nachgedacht werden, wo überschüssiges organisches Material über die
Schelfkante auf den Kontinentalhang transportiert wird (siehe 4.4).
An Station 11 nimmt die Makrofaunadichte auf 907 Ind. m-2 ab. Dies erscheint
ungenwöhnlich, da FRANKE (2000) an dieser Station die höchsten Corg-Werte für das Vigo-
Profil ermittelte (ca. 1,1 % Corg). Ebenfalls ungewöhnlich ist die Zunahme der Fauna-Gruppen
an Station 11. So wurden an Station 20 neun Gruppen nachgewiesen, an Station 11 jedoch elf.
Der Varianz der Faunadichte und Zusammensetzung kann mit mesoskaligen
hydrodynamischen Effekten zusammenhängen. So können Gezeitenströme oder interne
Wellen für stake Resuspension und die Umverteilungen von organischem Material sowie der
Fauna sorgen (SOLTWEDEL, 1997). Gezeitenströme die bodennah und hangparallel der
Iberischen Halbinsel in einer Tiefe von 1500m verlaufen, können nach VAN WEERING und
DE STIGTER (1999) 15-25 cm s-1 erreichen. Durch diese Gezeitenströme können erhebliche
Mengen Material resuspendiert worden sein, und dann in einer topographischen Senke im
Strömungsberuhigten mittleren Hangteil abgelagert werden. Dies wurde im SEEP-I und –II
Untersuchungsgebiet nachgewiesen (ANDERSON et al.,, 1988; BISCAY et al.,, 1994;
ROWE et al.,, 1994). Nachdem sich das Sediment abgelagert hat wird es neu besiedelt.
Faunendichte und zusammensetzung variieren hierbei (SMITH und HESSLER, 1987).
FRANKE (2000) fand an Station 11 sehr feines Material, was für einen Strömungsberuhigten
Bereich sprechen würde. Es kann hier jedoch die Annahme von FRANKE (2000) untermauert
werden, dass es sich an Station 11 um kein Akkumulationsgebiet handelt da die Faunendichte
zu gering ist.
43
Die gefundenen Chlorophyll a Mengen sind um die Hälfte geringer als an Station 20 und
folgen so dem gängigen Trend der Abnahme der Chlorophyll-Menge im Sediment mit
zunehmender Wassertiefe (PFANNKUCHE und SOLTWEDEL, 1998). Das Verhältniss von
Chlorophyll a zu Phaeophytin ist an dieser Station in der obersten Sedimentschicht bis 0,5 cm
sehr hoch(Abb.21). Da diese Station einen strömungsberuhigten Teil des Hangs darstellen
könnte ist die Warscheinlichkeit groß, dass sich hier resuspendiertes Sediment aus den oberen
Hangregionen ablagert. Es handelt sich meistens um refraktäres Material. FRANCKE (2000)
wies an den Stationen des Vigo-Profils homogene Verteilung von Corg bis in 9cm
Sedimenttiefe nach, was auf refraktäres Material hindeutet (ANDERSON et al., 1994). Da das
Verhältniss zwischen Chlorophyll a und Phaeophytin (siehe Anhang) sehr hoch ist, besteht
nach STEPHENS et al.,(1997) die Möglichkeit, dass das sedimentierte Material relativ frisch
ist. Dies beruht darauf, dass der Abbau der Phaeopigmente langsamer Abläuft, als der des
labilen Chlorophylls, bei den bis zu 10% des sehr reaktiven Teils geschützt sein können.

Diskussion
Durch diesen Schutz vor Abbau kann Chlorophyll bis in große Sedimenttiefen nachgewiesen
werden. Wie oben bereits erwähnt selektieren die Tiere der Benthosgemeinschaft ihre
Nahrung. Nach GRAF et al.,, (1995) geht der Abbau von Chlorophyll innerhalb von wenigen
Tagen von statten, und reicht in kürzester Zeit bis in Sedimenttiefen von 10 cm.
Die an Station 11 gefundenen Polychaeten-Familien könnten diese Annahme untermauern. Es
wurden hauptsächlich Depositfresser gefunden. Nur die Magelonidae, Ampahartidae und
Cirratulidae wären in der Lage Suspensionsfressen zu betreiben. Nach FLACH und HEIP
(1996b) sorgen vor allem die Depositfresser durch hohe Vergrabungsraten für eine homogene
Verteilung der C/N-Verhältnisse im Sediment bis 9 cm Tiefe. Die geringe Faunendichte
könnte bei, im Gegensatz zu Station 20, nur geringfüg kleinerer Biomasse (0,223 g C m-2)
tatsächlich ein Indiez für eine Neubesiedlung nach einem heftigen Sedimentationsereigniss
sein. Wenn frisches Material ankommt könnten die großen Tiere schneller als kleinere sein
und so die Biomasseverteilung beeinflussen. Hinzukommt, dass die suspensionsfresenden
Familien oft eingeschränkt beweglich sind, im gegensatz zu den meist gut beweglichen
Depositfressern (FAUCHALD und JUMARS, 1979).
Der Db-Wert an dieser Station beträgt 0,11 cm2 d-1 und würde die oben gemachten Annahmen
unterstützen. Für die Modellierung wurde Model 5 verwendet, da es den besten Fit
berechnete. Das Model 2 kann nach den ersten 0,5 cm keinen brauchbaren Fit mehr
berechnen. Hier kann kein diffusionsanaloges Mischen angenommen werden.
Der über das Chlorophyll-Inventar berechnete Chl. a Fluss beträgt 0,023 µg cm-2 d-1 und ist
etwas kleiner als der Fluss den das Model 5 berechnete (0,037 µg cm-2 d-1). Die von Model 5
berechnete Injektionstiefe beträgt 6,73 cm. Bei 6cm Sedimenttief liegt ein Peak von 0,119 µg
Chl.a cm-2 was gut zu der ermittelten Injektiontiefe passen würde, und zu der Annahme, dass
Depositfresser hier das labile Chlorophyll sehr schnell in tieferes Sediment transportieren.
Eine Gruppe der Makrofauna die dies noch forcieren kann sind die Sipunculida (72 Ind. m-2).
Am Vøring Plateau wiesen GRAF et al. (1995) bis zu 90 Ind. m-2 nach, und trauten ihnen zu
alle erhältlichen organischen Partikel von der Sedimentoberfläche aufzunehmen und sie in
ihre Röhren zu verfrachten, um sie später als Fecal-Pellets wieder an die Sedimentoberfläche
abzugeben. Unsere hier gefundenen 72 Ind. m-2 sind warscheinlich nicht in der Lage das an
dieser Station alleine zu bewerkstelligen, haben aber gewiss einen großen Anteil am Transport
von Partikel in tiefere Sedimentschichten. Es könnte sein, dass hier nach einem heftigen
Sedimentationsereigniss ein lokales Akkumulationsgebiet entsteht. Um dies zu bestätigen
wären mehrere Probennahmen nötig, zu verschiedenen Zeiten in kurzem Abstand. Erst dann
44
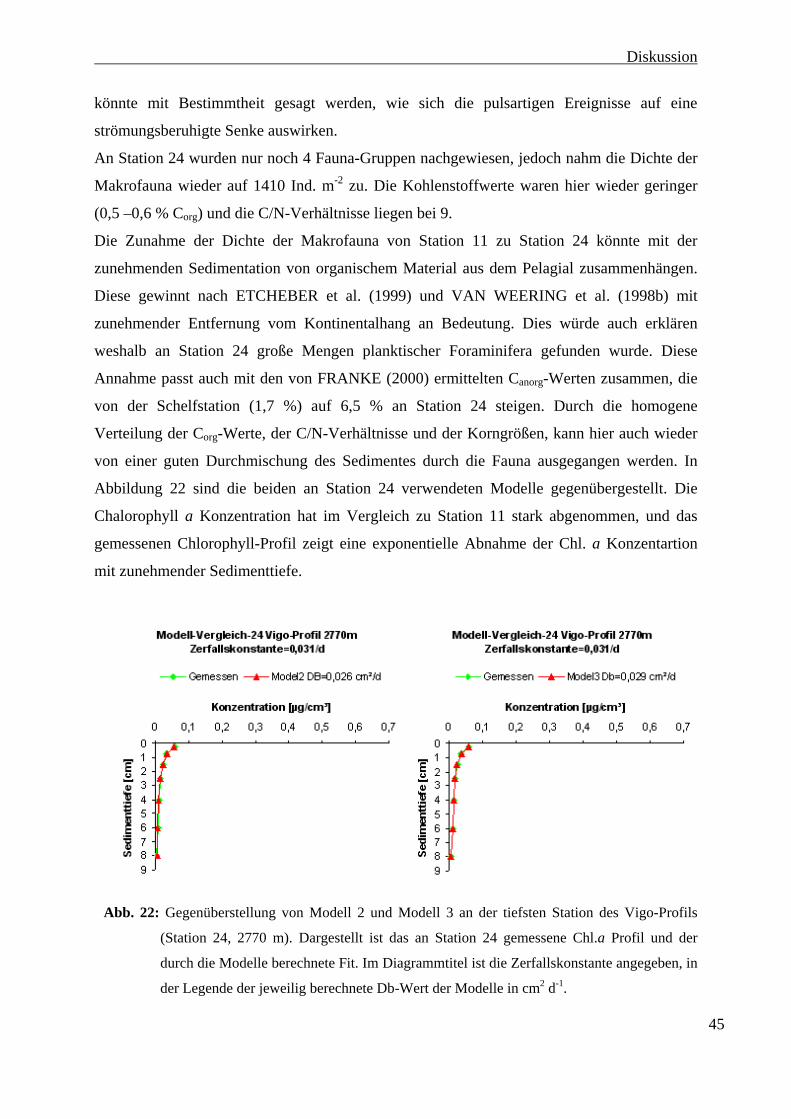
Diskussion
könnte mit Bestimmtheit gesagt werden, wie sich die pulsartigen Ereignisse auf eine
strömungsberuhigte Senke auswirken.
An Station 24 wurden nur noch 4 Fauna-Gruppen nachgewiesen, jedoch nahm die Dichte der
Makrofauna wieder auf 1410 Ind. m-2 zu. Die Kohlenstoffwerte waren hier wieder geringer
(0,5 –0,6 % Corg) und die C/N-Verhältnisse liegen bei 9.
Die Zunahme der Dichte der Makrofauna von Station 11 zu Station 24 könnte mit der
zunehmenden Sedimentation von organischem Material aus dem Pelagial zusammenhängen.
Diese gewinnt nach ETCHEBER et al. (1999) und VAN WEERING et al. (1998b) mit
zunehmender Entfernung vom Kontinentalhang an Bedeutung. Dies würde auch erklären
weshalb an Station 24 große Mengen planktischer Foraminifera gefunden wurde. Diese
Annahme passt auch mit den von FRANKE (2000) ermittelten Canorg-Werten zusammen, die
von der Schelfstation (1,7 %) auf 6,5 % an Station 24 steigen. Durch die homogene
Verteilung der Corg-Werte, der C/N-Verhältnisse und der Korngrößen, kann hier auch wieder
von einer guten Durchmischung des Sedimentes durch die Fauna ausgegangen werden. In
Abbildung 22 sind die beiden an Station 24 verwendeten Modelle gegenübergestellt. Die
Chalorophyll a Konzentration hat im Vergleich zu Station 11 stark abgenommen, und das
gemessenen Chlorophyll-Profil zeigt eine exponentielle Abnahme der Chl. a Konzentartion
mit zunehmender Sedimenttiefe.
Abb. 22: Gegenüberstellung von Modell 2 und Modell 3 an der tiefsten Station des Vigo-Profils
(Station 24, 2770 m). Dargestellt ist das an Station 24 gemessene Chl.a Profil und der
durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante angegeben, in
der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
45

Diskussion
Das diffusionsanaloge Model 2 errechnete einen Db von 0,026 cm-2 d-1 und einen
Oberflächenfluss von 0,0019 µg Chl.a cm-2 d-1. Das einfachste der non-local Modelle, Model
3, errechnete einen Db von 0,029 cm-2 d-1 und einen Oberflächenfluss von 0,0028 µg Chl.a
cm-2 d-1. Der über das Chlorophyll-Inventar berechnete Fluss beträgt 0,007 µg Chl.a cm-2 d-1.
Beide Modelle berechneten einen niedrigeren Oberflächenfluss, aber einen fast identischen
Db-Wert. Dies zeigt, dass advectives (non-local) Mischen nach längerem Anhalten zu
diffusionsanalogen Partikelverteilungen führt (BOUDREAU, 1986b). Die Makrofaunadichte
wird hier sehr stark von den Polychaeten beeinflusst. Die 4 anderen Makrofaunagruppen der
Bivalvia, Anisopoda, Isopoda und Scaphopoda machen nur 255 Ind. m-2 an dieser Station aus.
Die Polychaeta dominieren mit 1157 Ind. m-2. Auch an dieser Station werden die
Ernährungstypen der Polychaeta von den Deposit und Grenschichtfressern dominiert, was
sich mit den Ergebnissen von FLACH und HEIP (1996b) deckt. Die C/N-Verhältnisse lassen
auch wieder den Schluss zu, dass es sich um refraktäres Material handelt (FRANKE, 2000),
wobei LOHSE et al., (1998) darauf hinweisen, dass an Kontinentalhängen C/N-Werte
lediglich ein Indiez für das Vorhandensein von terrigenem Material sind. Das würde auch die
gute Durchmischung erklären, da die Depositfresser hier das Sediment immer wieder
aufnehmen um den geringen Teil an labilem Kohlenstoff zu verarbeiten. Durch ihre
Grabtätigkeit können die Depositfresser die Sedimentoberfläche und somit das
hydrodynamische Rgime beeinflussen und damit zur indirekten Biodeposition führen. Die
Grenzschichtfresser könnten je nach vorherrschender Strömung von Deposit zu
Suspensionfressen wechseln und so Partikel aktiv zur Biodeposition bringen(THOMSEN,
1998).
46
Die an Station 24 gefundenen Polychaeten-Familien zeigen wie oben schon erwähnt die
klassischenVerteilungsmuster. Die Paraonidae und Lumbrineridae sind am häufigsten
vertreten, gefolgt von den Maldanidae und den Spionidae. Die Spiopnidae stellen einen
Großteil der hier gefundenen Suspensionsfresser. Ansonsten kommen dafür nur noch einige
Gattungen der Onuphidae und Cirratulidae in Frage. Die restlichen Familien sind mehr den
Grenzschichtfressern zuzuordnen, oder aber von vorne herein Depositfresser. Die
vorgefundenen 185 Ind. m-2 der Spionidae könnten ein Indiez für erhöhte
Strömungsgeschwindigkeiten sein. FRANKE (2000) fand an Station 24 gröberes Material als
an Station 11. THOMSEN und VAN WEERING (1998) fanden am Konbtinentalhang bei
Goban Spur in 1450 m Strömunggeschwindigkeiten von bis zu 40 cm s-1 und VAN
WEERING et al., (1998b) fanden dort auch gröberes Sediment. Bei den vorherrschenden
nördlichen Strömungrichtungen (DICKSON et al., 1985) könnte es sein, dass das Plateau auf

Diskussion
dem Station 24 liegt, zwischen Galicia-Bank und Kontinentalhang eine Art Kanal darstellt, in
dem erhöhte Bodenströmungen herrschen. Durch die oben erwähnten Aktivitäten der Fauna
werden feine Partikel vermehrt resuspendiert und können so von der Strömung abtransportiert
werden.
4.2 Charakterisierung des Nazaré-Canyons Etwa 50 % der Fläche der europäischen Kontinentalhänge ist von Tiefseecanyon bedeckt.
Diese Canyonsysteme gelten nach HUTHNANCE (1995) und MONACO et al. (1990) als
sehr bedeutende morphologiche Strukturen für den Transport von organischem Material in die
Tiefsee. MONACO et al., (1990) wiesen im Lacaze-Duthiers-Canyon und anderern Canyon
im Golf von Lion nach, dass nur ca. 10 % des in ihnen transportierten Materials
sedimentieren, und die restlichen 90 % in größere Tiefen transportiert werden.
SORBE, (1999) unterscheidet „aktive“ und „inaktive“ Canyonsysteme. In einem aktiven
Canyon wird Material kontinuierlich in größere Tiefen transportiert. Die
Makrofaunagemeinschaft dieses Canyontypus unterscheidet sich in Dichte und
Zusammensetzung meist nicht von den Makrofaunagemeinschaften der benachbarten
Kontinentalhänge. An der Canyonmündung in die Tiefsee-Ebene akkumuliert das durch den
Canyon transportierte Material, wodurch auch die Makrofaunagemeinschft hier größer sein
kann als in den benachbarten Gebieten.
Inaktive Canyonsysteme fungieren als Depotcenter. In ihnen sammelt sich, meist in den
flacheren Teilen des Canyons, sehr feines, organisch angereichertes Sediment. In diesen
Bereichen finden sich dann große Abundanzen bestimmter Faunengruppen wie z.B.
Holothurien. Diese werden als „Canyon-Indikatoren“ bezeichnet. Die Makrofaunadichte in
einem inaktiven Canyon ist generell höher als die Dichte der Makrofauna der angrenzenden
Gebiete (SORBE, 1999).
47
Station 26 ist die flachste Station der drei Stationen im Nazaré-Canyon und weist Merkmale
für einen inaktiven Canyon auf. Die Makrofaunadichte liegt bei 117566 Ind. m-2. Hierzu muß
jedoch gesagt werden, daß die vorgefundenen 116738 Cossuridae m-2 durchgehend ohne
Abdomen waren, d.h. nur Kopf und Torax vorhanden waren (Abb. 23). Ähnliche Ergebnisse
erlangte CÚRDIA (pers. Mitteilung). In der Zoologie werden Polychaeta die ohne Kopf
gefunden werden nicht gezählt. In wie Fern es sich im ungekehrten Fall verhält ist unklar, und
sie werden deshalb mitgezählt. Es soll hier versucht werden, zu erklären wie die große Menge
an Cossuridae an diese Station gekommen ist.

Diskussion
Wie oben bereits erwähnt fungieren inaktive Canyon als Depotcenter, indem organisch
reiches Material in den flacheren Canyonteilen abgelagert wird. Die Topographie des Nazaré-
Canyons lässt solche Depotcenter in den flacheren Bereichen des Canyons warscheinlich
nicht zu (siehe Material und Methoden 2.1). Der obere (flache) Teil des Canyons hat sehr
steile Flanken und es könnte sein, dass dort größere Mengen Material nicht abgelagert werden
können.
Organisch reiches Material kommt in der Regel vom Schelf, wo es durch tidenabhängige
Strömungen suspendiert wird. Ein Teil des Materials wird über die Schelfkante hangabwärts
transportiert, der Rest wird in hangparallelen Strömungen bis zu einem Canyon transportiert.
Das Material sedimentiert im Canyon-Kopf und akkumuliert bevor es erneut resuspendiert
werden kann (BISCAYE und ANDERSON, 1994; HEUSSNER et al.,, 1999). Durch
intensive Bioturbation, die durch den Eintrag des relativ frischen organischen Materials
forciert wird, kommt es zu Sedimentinstabilitätet(DILLON und ZIMMERMAN, 1970;
HECKER, 1982; MALAHOFF et al., 1982 aus GAGE und TYLER, 1991). Durch interne
Wellen, Erdbeben oder Tidenabhängige Strömungen kann das Sediment ins Rutschen
kommen (LUCKENBACH, 1986) und gelangt so in tiefere Bereiche des Canyon, wo es in
einer topographischen Senke liegen bleibt.
Nach HARTMANN-SCHRÖDER (1996) wurden Cossuridae bisher erst für Wassertiefen bis
etwa 900 m beschrieben, und sie gelten als sehr bewegliche das Sediment durchrabende
Polychaeta (FAUCHALD und JUMARS, 1979). Es wäre daher vorstellbar, dass die
Cossuridae im oberen Hangbereich frisches organisches Material besiedelt und es dabei stark
durchwühlt haben.
Abb. 23: Darstellung eines Cossura longocirrata WEBSTER&BENEDICT, 1887 (Cossuridae,
DAY, 1963). Die gestrichelte Linie zeigt den Bereich wo die Tiere von ihrem Abdomen
getrennt wurden.
48

Diskussion
Hierdurch könnte es zu einer Sedimentinstabilität gekommen sein und eine große Menge
Material ist abgerutscht. Dies könnte erklären, weshalb die Cossuridae alle nur halb
vorgefunden wurden (Abb. 23), da sie auf dem weg in die Tiefe durch die Turbulenzen
zerstört wurden.
Es wäre aber auch möglich, dass die Cossuridae beim Aussieben zerstört wurden. Da aber
keine vollständigen Cossuridae gefunden wurden ist das eher unwarscheinlich.
Außer den Cossuridae wurden an Station 26 noch 828 Bivalvia m-2 in der
Makrofaunagrößenklasse nachgewiesen. Die Bivalvia wurden im Rahmen dieser Arbeit nicht
weiter bearbeitet, und daher ist nicht bekannt um welche Familien oder Gattungen es sich
handelt, und es kann daher auch nichts zu ihrem vorkommen an Station 26 gesagt werden.
Für eine große Menge frisch abgerutschten Sediments sprechen auch die homogenen
Verteilungen der Chlorophyll a Equivalente sowie die Verteilung des ATP und der hohe
Sedimentwassergehalt (89-75 Vol. %) an dieser Station.(FRANCKE, 2000). Diese Verteilung
könnte auch durch intensive Bioturbation zustande kommen.
Mit dem diffusionsanalogen Model 2 wurde ein astronomischer Diffusionkoeffizient (Db) von
2,55·1028 cm2 d-1 und ein gigantischer Oberflächenfluss von 1,89·1010 µg cm-2 d-1 errechnet.
Diese Werte sind völlig unrealistisch.
Abb. 24: Gegenüberstellung von Modell 2 und Modell 4b an der flachsten Station des Nazaré-
Canyons (Station 26, 2800 m). Dargestellt ist das an Station 26 gemessene Chl.a Profil
und der durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante
angegeben, in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.
49

Diskussion
In Abbildung 24 sind nochmals die beiden verwendeten Modelle gezeigt. Gut zu erkennen ist
der zunehmende Profilverlauf des Chlorophyll-Profils. Das Model 4b ermittelte einen Db-
Wert von 0,015 cm2 d-1 was ca 5,5 cm2 a-1 bedeuten würde. Das wäre in anbetracht dessen,
dass das Sediment warscheinlich relativ frisch ist ein plausiebler Wert. Frischer Phytodetritus
kann nach GRAF (1989) mit einer Geschwindigkeit von 1 cm d-1 in das Sediment
transportiert werden. Für diesen Transport kommen die gefundenen Cossuridae nicht in Frage,
da nur die Thoraca gefunden wurden. Also muß diese Bioturbationsleistung zum einen von
den an Station 26 gefundenen Bivalvia, und/oder aber von Holothurioidae (Seegurken)
vollzogen werden. An Station 26 wurden einige ca. 10 cm große Seegurken gefunden, jedoch
nicht mit in die Datenerhebung aufgenomme (siehe 3.1.1). Es wäre vorstellbar, dass diese
Holothurioidae für die massive Bioturbation verantwortlich sind, kann aber nicht mit
bestimmtheit gesagt werden, da nicht bekannt ist wie viele Seegurken wirklich im Bereich
dieser Station zu finden sind. Für die Bioturbation durch die Seeegurken sprechen auch die
durch Model 4b (0,094 µg Chl.a cm-2 d-1) und den Chlorophyll-Inventar (0,083 µg Chl.a cm-2
d-1 ) ermittelten Oberflächenflüsse. Beide Flüsse liegen deutlich höher als die
Oberflächenflüsse an z.B. der Schelfstation 20 (0,069/0,067 µg Chl.a cm-2 d-1). Die von
Model 4b berechnete Injektionstiefe beträgt 9,82 cm. Nach BOUDREAU (1994) ist 9,8 ± 4,5
cm die Sedimenttiefe, die im Mittel Weltweit als Sedimenttiefe für den Aufenthalt von
Depositfressern angesehen wird. Dieser Sedimentbereich ist somit auch die am meisten durch
Depositfresser beeinflusste Schicht. Anhand eines einfachen Models verbesserte
BOUDREAU (1998) diese Sedimenttiefe auf 9,7 cm, was aber auch noch sehr dicht bei
unseren Ergebnissen liegt.
An Station 25 wurden weniger Tiere gefunden, jedoch mehrere Familien. Es wurden
insgesamt 5001 Ind. m-2 nachgewiesen, die sich aus 7 Polychaeten-Familien und 5
Makrofauna-Gruppen zusammmensetzten. Die Makrofaunadichte ist um den Faktor 3-5,5
größer als am Vigo-Profil. Dies würde nach SORBE (1999) für einen inaktiven Canyon
sprechen (siehe Oben). Nach FRANCKE (2000) weisen die von ihm gefundenen Corg-Werte
von bis zu 2% an den Stationen 26 und 25 auf lokale Akkumulationsgebiete oder aber auf
erhöhten Eintrag organischen Materials hin. Es könnte sich bei Station 25 also ebenfalls um
ein Depotcenter handeln. Außerdem wurden an Station 25 ebenfalls Holothurien gefunden,
die wie oben bereits erwähnt als Canyon-indikatoern in inaktiven Canyon zu seghen sind.
50
Die Zusammensetzung der gefundenen Fauna an Station 25 spricht ebenfalls für eine
Anreicherung organischen Materials. Es wurden Scaphopoda und Prochaetodermatidae mit je
844 Ind. m-2 nachgewiesen, die nach GAGE und TYLER (1991) sowie RIEDL (1983)

Diskussion
Bewohner organisch angereicherter feinschlammiger Böden sind. Von den 7 gefundenen
Polychaeten-Familien sind 6 Depositfresser. Nur die Spionidae, die mit 1883 Ind. m-2
gefunden wurden sind fähig Suspensionsfressen zu betreiben, können jedoch als
Grenzschichtfresser eingestuft werden (FAUCHALD und JUMARS, 1979). Da die Spionidae
in so großer Zahl angetroffen werden, kann davon ausgegangen werden, dass an Station 25
vermutlich regelmäßig Strömungen auftreten, die Material resuspendieren, oder aber
suspendiertes Material vorbeitransportieren. Die Spionidae könnten dann von Depositfressen
zu Suspensionsfressen wechseln.
An Station 25 wurden außerdem die meisten Foraminifera im Untersuchungsgebiet
nachgewiesen (188700 Ind. m-2), was für ein gutes Nahrungsangebot spricht
(NUGLISCH,1985) und somit für erhöhten Eintag organischen Materials stehen könnte.
Durch die Anwesenheit der Foraminifera wurde an Station 25 vmit 1, 271 g C m-2 die höchste
Biomasse im Untersuchungsgebiet ermittelt.
Die gefundenen Chlorophyll a Mengen sind auch an Station 25 wieder um den Faktor 2-5
höher als am Vigo-Profil und deuten somit auf einen erhöhten Materialeintrag.
Der Db-Wert nimmt von Station 26 zu Station 25 um die Hälfte ab. Mit Model 4b wurde ein
Db von 0,007 cm2 d-1 errechnet. Der Oberflächenfluss beträgt 0,0034 µg cm-2 d-1 und ist um
den Faktor 10 kleiner als der aus dem Inventar berechnete Fluss von 0,034 µg cm-2 d-1. Die
Injektionstiefe liegt bei 2,5 cm, was genau zu dem Peak in 2,5 cm Tiefe passen würde. (Abb.
25).
51
Abb. 25: Gegenüberstellung von Modell 2 und Modell 4b an der mittleren Station des Nazaré-
Canyons (Station 25, 3500 m). Dargestellt ist das an Station 25 gemessene Chl.a Profil
und der durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante
angegeben ,in der Legende der jeweilig berechnete Db-Wert der Modelle in cm2 d-1.

Diskussion
Wie in Abbildung 25 zu erkennen konnte hier mit Model 2 kein guter Fit errechnet, und somit
auch kein diffusionsanaloges Mischen festgestellt werden.
Der niedrigere Db-Wert an Station 25 im gegensatz zur Station 26 kann nach STEPHENS et
al., (1997) mit kleineren Zerfallskonstatnten und längeren Halbwertzeiten zusammenhängen.
Die längeren Halbwertzeiten resultieren aus refraktärem Material welches nicht so schnell
abgebaut werden kann und meist in tiefren Sedimentschichten zu finden ist. FRANKE (2000)
wies für den Nazaré-Canyon steigende C/N-Verhältnisse nach, was für refrakteres Material
spricht und verwies darauf, dass eventuell durch eine Turbidit frisches Material an die Station
kam. Tuirbidite sind hangabwärts gerichtete Trübeströme die oft große Materialfrachten in die
Tiefe transportieren und dabei sehr hohe Strömungsgeschwindigkeiten erreichen können
(HUTHNANCE, 1995; MONACO et al., 1990). Das würde zumindest erklären weshalb so
hohe Chlorophyll a Werte dem hohen C/N-Verhältniss von 12,5 gegenüber stehen. An Station
25 wurden zudem erhebliche Mengen von Pflanzenresten gefunden, die z.B. von Seegrass
stammen könnten. SORBE (1999) fand im Cap-Ferret-Canyon ebenfalls größere Mengen von
Pflanzenfragmenten und deutetete sie als Indikator für den Transport von Material durch den
Canyon. Sie werden vor allem in den tieferen Canyonabschnitten gefunden, da dort
warscheinlich mehr Material akkumuliert. In diesem Zusammenhang erwähnt er GRIGGS et
al., (1969), der herausfand, dass solche Pflanzereste weniger von Depositfressern sondern von
Mikroorganissmen abgebaut werden. Dies würde auch zu den hohen Sauerstoffzehrungwerten
von FRANKE (2000) passen (St.25 = 6,4 mmol O2 m-2 d-1).
52
Station 27 stellt im Fall dieser Arbeit eine Besonderheit dar, da an dieser Station keine Fauna
nachgewiesen werden konnte. Das kann zum einen am an dieser Station von FRANKE (2000)
nachgewiesenen C/N-Verhältnissen (14) liegen, zum anderen besteht aber auch die
Möglichkeit, dass kurz vor der Probennahme ein Turbidit den Großteil der oberen
Sedimentschichten weggespühlt hat. Hierfür würden auch die von FRANKE (2000)
ermittelten Korngrößen mit einem Meridian von 282,4µm sprechen. Bei der Probennahme
viel an Station 27 eine regelmäßige Schichtung des Sediments auf. Auf eine ca. 0,5-1 cm
dicke Sandschicht folgte eine etwa 5-8cm dicke feinschlammige Schicht, die dann wieder von
einer Sandschicht überdeckt war. Dies konnte bis zu einer Sedimenttiefe von 50 cm
beobachtet werden. Dies könnte ein Indiez für regelmäßige Turbidite sein mit
zwischenzeitlichen weniger umfangreichen Sedimentationsereignissen. So könnte immer
wieder frisches Material akkumulieren, wobei die gröberen Sedimentanteile in untere
Sedimentlagen sacken. Das feine Fluffartige Material wird entweder von einem erneuten
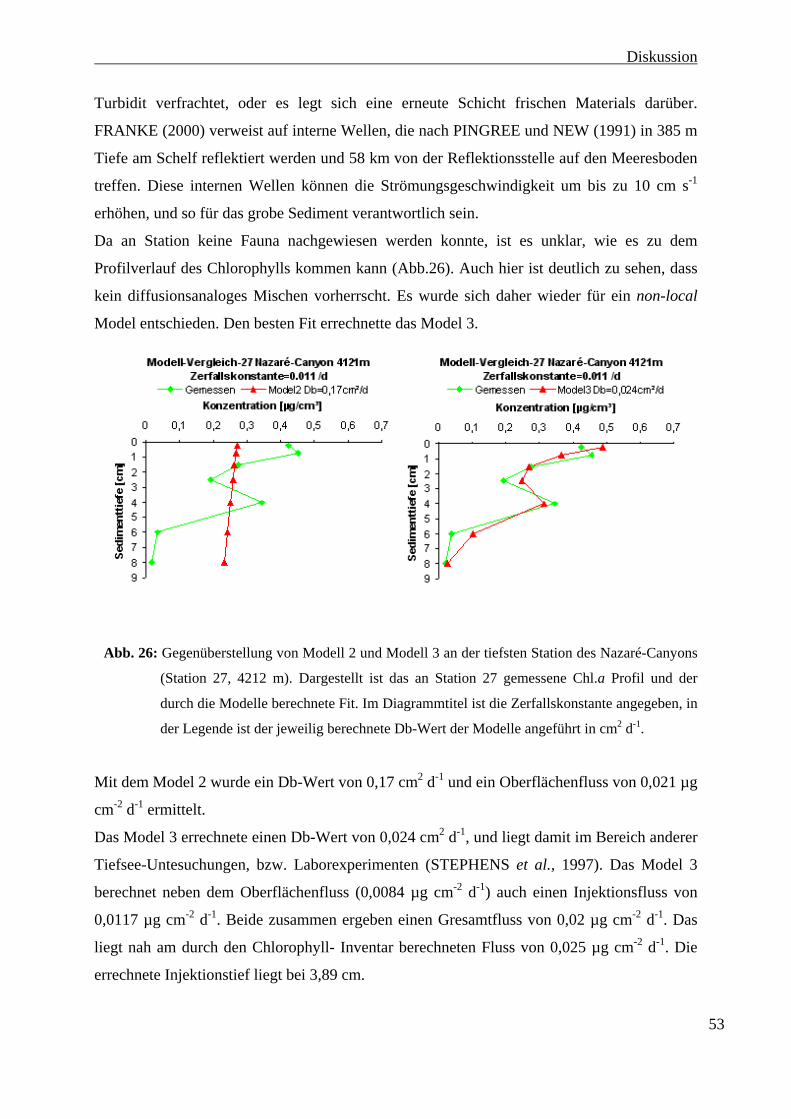
Diskussion
Turbidit verfrachtet, oder es legt sich eine erneute Schicht frischen Materials darüber.
FRANKE (2000) verweist auf interne Wellen, die nach PINGREE und NEW (1991) in 385 m
Tiefe am Schelf reflektiert werden und 58 km von der Reflektionsstelle auf den Meeresboden
treffen. Diese internen Wellen können die Strömungsgeschwindigkeit um bis zu 10 cm s-1
erhöhen, und so für das grobe Sediment verantwortlich sein.
Da an Station keine Fauna nachgewiesen werden konnte, ist es unklar, wie es zu dem
Profilverlauf des Chlorophylls kommen kann (Abb.26). Auch hier ist deutlich zu sehen, dass
kein diffusionsanaloges Mischen vorherrscht. Es wurde sich daher wieder für ein non-local
Model entschieden. Den besten Fit errechnette das Model 3.
Abb. 26: Gegenüberstellung von Modell 2 und Modell 3 an der tiefsten Station des Nazaré-Canyons
(Station 27, 4212 m). Dargestellt ist das an Station 27 gemessene Chl.a Profil und der
durch die Modelle berechnete Fit. Im Diagrammtitel ist die Zerfallskonstante angegeben, in
der Legende ist der jeweilig berechnete Db-Wert der Modelle angeführt in cm2 d-1.
Mit dem Model 2 wurde ein Db-Wert von 0,17 cm2 d-1 und ein Oberflächenfluss von 0,021 µg
cm-2 d-1 ermittelt.
Das Model 3 errechnete einen Db-Wert von 0,024 cm2 d-1, und liegt damit im Bereich anderer
Tiefsee-Untesuchungen, bzw. Laborexperimenten (STEPHENS et al., 1997). Das Model 3
berechnet neben dem Oberflächenfluss (0,0084 µg cm-2 d-1) auch einen Injektionsfluss von
0,0117 µg cm-2 d-1. Beide zusammen ergeben einen Gresamtfluss von 0,02 µg cm-2 d-1. Das
liegt nah am durch den Chlorophyll- Inventar berechneten Fluss von 0,025 µg cm-2 d-1. Die
errechnete Injektionstief liegt bei 3,89 cm.
53

Diskussion
Wie kommt nun dieser Profilverlauf zustande? Da keine Fauna angetroffen wurde müssen
hier Annahmen gemacht werden. Wie oben bereits erwähnt gelten Holothurien als Canyon-
Indikatoren und sind dort anzutreffen wo es viel organisches Material zu verarbeiten gibt
(SORBE 1999). Nach DUINEVELD et al., (1997) gelten Holothurien als Indikatoren für
pulsartigen, sporadischen Phytoplanktoneintrag. Es muss daher davon ausgegangen werden,
dass an dieser Station Holothurien vorkommen, wir aber auf Grund der Patchiness keine in
unserem Kastengreifer hatten. Weshalb von der restliche Fauna ebenfalls nichts gefunden
wurde kann hier nicht erklärt werden. Eine Möglichkeit wäre Anoxie. Jedoch ist das Sediment
an dieser Station erst ab ca. 3 cm anoxisch (CÚRDIA, pers. Mitteilung). Eine andere
Möglichkeit für die unregelmäßige Verteiluing des Chlorophylls könnten die sporadischen
pulsartigen Einträge von Material sein. Hierüber ist jedoch nichts bekannt.
4.4 Vergleich der bearbeiteten Gebiete im Rahmen dieser Arbeit
Der von den meisten Kontinentalhängen bekannte Trend in der Abnahme der
Makrofaunadichte mit zunehmender Tiefe und entfernung von Land (SIBUET et al. 1989;
FLACH & HEIP, 1996; SOLTWEDEL, 1997; FLACH, et al. 1998; PFANNKUCHE, et al.
1998) konnte am Vigo-Profil nachgewiesen werden, jedoch nicht im Nazaré-Canyon. Zum
einen liegt es daran, dass die falchste beprobte Station im Canyon ca. 100 m tiefer lag als am
Vigo-Profil, und somit keine Makrofaunawerte für mit dem Vigo-Profil vergleichbare Tiefen
zur verfügung standen.
54
Zum anderen liegt es daran, dass der Nazaré-Canyon aller warscheinlichkeit nach ein
inaktiver Canyon ist. Inaktive Canyonsysteme zeichnen sich durch wesentlich höhere
Makrofaunazahlen als an den ihn umgebenen Kontinetalhanggebieten aus. Ausserdem gelten
sie als Depotcenter für organisch reiches Material, was zur Folge hat, dass in bestimmten
Bereichen sehr hohe Makrofaunazahlen anzuterffen sind und in anderen Bereich wieder sehr
viel weniger (SORBE, 1999). So könnten zumindest die hohehn Individeunzahlen an Station
26 erklärt werden, obwohl unklar ist, ob die dort gefunden Tiere nicht durch einen Turbidit
dort hingelangten. Die immer noch sehr hohen Makrofaunazahlen an Station 25 könnten
hingegen eine Beweis für ein lokales Akkumulationsgebiet sein. Dies würde auch mit den für
diese Wassertiefe hohen Corg-Werten (2%) von FRANKE (2000) zusammenpassen. Die
völlige Abwesenheit von Fauna an Station 27 kann hier nur durch Patchiness erklärt werden,
da wir nur einen Großkastengreifer für die Bilanzierung der Fauna zur Verfügung hatten. Ob

Diskussion
diese Phenomän mit Strömung oder Turbiditen zusammenhängt muss in Zukunft noch geklärt
werden
Die Makrofaunazahlen am Vigo-Profil sind für Kontinetalhänge sehr gering. FLACH et al.,
(1998) wiesen jedoch in unmittelbarer Nähe (Sommer) des Vigo-Profils ähnlich niedrige
Faunenzahlen nach, so das ausgeschlossen werden kann, dass die geringen Individuenzahlen
mit der Probennahme im Winter zu tun haben. Weshalb die Makrofaunadichte am
Kontinentalhang vor Vigo trotz der im Winter auch noch sehr hohehen
Primärproduktionsraten von 250-350 mg C m-2 d-1 (ANTOINE et al. 1996) so gering bleibt,
ist unklar. Ein Grund könnten die verstärkten Exportprozesse über die Schelfkante und den
Kontinentalhang in die Tiefsee sein, die für einen hochenergetischen Kontinentalhang
angenommen werden (HUTHNANCE, 1995; THOMSEN, 1998). Dies wäre auch eine
Erklärungsmöglichkeit für die geringe Fauna an Station 11 des mittelren Kontinentalhanges
sowie die Zunahme am Sockel (Station 24). Hierfür können hangabwärts gerichtete laterale
Transportprozesse in der Bodengrenzschicht (Benthic Boundary Layer) die Ursache sein, wie
sie THOMSEN (1998) für das Goban-Spur-Gebiet feststellte. Durch hohe
Strömungsgeschwindigeiten verstärkte Resuspension, bringt deponiertes Material immer
wieder in die BBL, wo es dann Hangabwärts transportiert werden kann. Die hohen
Strömungsgeschwindigkeiten an der Schelfkante von bis zu 45 cm s-1 (HUTHNANCE et al.,,
pers. Mitteilung) und Gezeitenströme mit Geschwindigkeiten von 15-25 cm s-1 die bodennah
und hangparallel der Iberischen Halbinsel in einer Tiefe von 1500m verlaufen (VAN
WEERING und DE STIGTER, 1999), könnten für wiederholte Resuspension sorgen. Es
könnte also sein, dass dem mittleren Kontinentalhang nicht ausreichend organisches Material
zur Verfügung steht, um höhere Individuenzahlen hervorzubringen. Dafür finden sich am
Kontinentalsockel wieder vermehrt Tiere, da dort eine große Menge des Hangabwärts
transportierten Materials akkumulieren kann.
Ca. 80 % der Makrofauna der Tiefsee sind Depositfresser (JUMARS und GALLAGHER,
1982). Dies konnte am Vigo-Profil beobachtet werden. Nur die Schelfstation wies
erwartungsgemäß eine homogen Verteilung zwischen Deposit- und Suspensionsfressern auf.
Die Dominanz der Deposit- bzw. Grenzschichtfresser steigt mit zunehmender Tiefe (vergl.
Tab. 4-6 unter 3.1.5).
55
Im Nazaré-Canyon überraschen die hohen Abundanzen der Spionidae an Station 25. Sie
machen mit 1883 Ind. m-2 zwei Drittel der gefundenen Polychaeta aus. Es sind
Grenzschichtfresser, die somit auch suspensionsfressen können, und eineige Arten sind reine
Filtrierer. Da hier nur bis auf Familienebene bestimmt wurde können keine genaueren

Diskussion
Angaben gemacht werden. Die Anzahl der gefundenen Familien im Nazaré-Canyon
verdeutlicht jedoch die Dominanz des Depositfressen im Tiefsee-Milieu.
Die Abnahme der gemessenen Chlorophyll-Werte mit zunehmender Tiefe am Vigo-Profil
entsprechen anderen Untersuchungen an Kontinentalhängen (DUINEVELD et al., 1997;
LAVALEYE et al., 1999). Die exponentielle Abnahme der Chlorophyll a Konzentration mit
zunehmnder Tiefe und entfernung von Land, verdeutlicht den zunehmenden marinen Einfluss
der weiter von Land entfernten Stationen 11 und 24. Dies spiegelt sich auch in den von
FRANKE (2000) ermittelten C/N-Verhältnissen wieder, die von 13,5 am Schelf auf 9 an den
tiefen Stationen zurückgehehn.
Die Chlorophyll a Konzentrationen im Nazaré-Canyon liegen um den Faktor 1,5 – 6 höher als
am Vigo-Profil und verdeutlichen einmal mehr den inaktiven Charakter des Canyons, und
seine Funktion als Depotcenter und Materialrutsche für organisch angereichertes Sediment.
Die über den Tracer Chlorophyll a modellierten Bioturbationskoeffizienten und Chlorophyll-
Flüsse korrelieren mit den ermittelten Makrofaunadaten (Individuenzahlen und Biomasse).
56
Der höchste Db-Wert von 2,1 cm2 d-1 wurde an der Schelfstation gemessen, wo auch die
meiste Fauna (1616 Ind. m-2) am Vigo-Profil entdeckt wurde und auch die höchste Biomasse
(0,244 g C m-2). Die beiden errechneten Oberflächenflüsse für Chlorophyll a liegen dicht
beieinander (Tab.9). Bei einem Chl.a/Corg-Verhältniss von 60, was etwa dem Verhältniss von
lebenden Algen entspricht (MURRAY et al., 1994), ergibt sich für Station 20 ein
Kohlenstofffluss von 4,02 mg C m-2d-1 (über den Inventar) und 4,13 mg C m-2d-1 (über Model
5). Nach FRANKE (2000) hat Station 20 einen Corg-Bedarf von 44,9 mg C m-2d-1. Das an
Station 20 Chlorophyll a würde also nicht ausreichen. Berechnet man den Fluss jedoch mit
Chl.a/Corg-Verhältnissen von 600 bzw. 1000, wie sie in der Tiefsee angetroffen werden (600-
7700, NEWTON und MURRAY, 1995, aus STEPHENS et al., 1997) so ergeben sich für die
Inventar-Methode 40,18 mg C m-2d-1 und für Model 5 41,32 mg C m-2d-1. Erst bei einem
Chl.a/Corg-Verhältnissen von 1000 wäre der Bedarf an organischem Kohlenstoff an Station 20
gedeckt (66,96 mg C m-2d-1 über Inventar 68,86 mg C m-2d-1 Model 5). An Station 11 wurde
ein höherer Db (0,11 cm2 d-1) ermittelt als an Station 24 (Db=0,029 cm2 d-1), obwohl an
Station 24 mehr Ind. m-2 (1410 Ind. m-2) gefunden wurden. Dies hängt mit der höheren
Biomasse an Station 11 zusammen (907 Ind. m-2 ergaben 0,223 g C m-2d-1). Hier scheinen
einige größere Tiere für höhere Bioturbationsraten verantwortlich zu sein, wie z.B. die 72
Sipunculiden pro Quadratmeter. Für die in Tabelle 9 aufgelisteten Chlorophyll-Flüsse ergeben
sich (mit den oben genannten Chl.a/Corg-Verhältnissen 60 / 600 / 1000) folgende Corg-Flüsse
an Station 11 und 24.

Diskussion
Tab.9: Aufgeführt sind der Chlorophyll a-Inventar der einzelnen Stationen sowie die jeweilig
ermittelte Zerfallskonstatnte für Chlorophyll a. Ausserdem sind die über das Inventar und den Zerfall
berechneten sowie die von den Modellen errechneten Chl.a-Flüsse aufgelistet. Stat. Tiefe
m
Chl.a-Inventar
µg cm-2
Kd
d-1
Chl.a-Fluss
über Inventar
µg cm-2 d-1
Chl.a-Fluss
über Model 3
µg cm-2 d-1
Chl.a-Fluss
über Model 4a
µg cm-2 d-1
Chl.a-Fluss
über Model 4b
µg cm-2 d-1
Chl.a-Fluss
über Model 5
µg cm-2 d-1
20
11
24
217
1945
2763
2,16
0,653
0,232
0,031
0,035
0,031
0,067
0,023
0,007
--
--
0,0021
--
--
--
--
--
--
0,069
0,037
--
26
25
27
2894
3514
4121
3,208
3,119
2,241
0,022
0,011
0,011
0,071
0,028
0,025
--
--
0,02
--
--
--
0,0094
0,0028
--
--
--
--
Station 11: Corg-Bedarf 19,2 mg C m-2 d-1
Inventar (60) = 1,37 g C m-2 d-1 (600) = 13,71 g C m-2 d-1 (1000) = 22,86 g C m-2 d-1
Model 5 (60) = 2,21 g C m-2 d-1 (600) = 22,1 g C m-2 d-1 (1000) = 36,84 g C m-2 d-1
Station 24: Corg-Bedarf 23 mg C m-2 d-1
Inventar (60) = 0,43 g C m-2 d-1 (600) = 4,32 g C m-2 d-1 (1000) = 7,19 g C m-2 d-1
Model 3 (60) = 0,17 g C m-2 d-1 (600) = 1,68 g C m-2 d-1 (1000) = 2,8 g C m-2 d-1
Der Kohlenstoffbedarf an Station 11 wurde von FRANKE (2000) mit 19,2 g C m-2 d-1
errechnet. Nach Model 5 wäre der Kohlenstoffbedarf an Station 11 bei einem Chl.a/Corg-
Verhältniss von 600 gedeckt. Station 24 hat einen Bedarf an Corg von 23 g C m-2 d-1. Durch die
Zufuhr von Chlorophyll a kann der Bedarf von Station 24 nicht gedeckt werden.
Im Nazaré-Canyon kann keine direkte Korrelation der Db-Werte mit den Makrofaunadaten
festgestellt werden.
Der höchste Db wurde mit 0,024 cm2 d-1 an Station 27 errechnet. Hier wurde jedoch keine
Makrofauna nachgewiesen. Die Station 25 hat die höchste Biomasse mit 1,271 g C m-2 aber
den kleinsten Db-Wert (0,005 cm2 d-1). An Station 26, wo größtenteils Cossuridae mit
unbekannter Herkunft gefunden wurden hat Model 4b einen Db-Wert von 0,015 cm2 d-1
errechnete, bei einer Biomasse von 0,217 g C m-2. Die Chlorophyll-Flüsse sind ebenfalss in
Tabelle 9 aufgelistet. Die Kohlenstoffflüsse für die drei Stationen des Nazaré-Canyon sind
nachstehend aufgeführt.
57

Diskussion
Station 26: Corg-Bedarf 48,4 mg C m-2 d-1
Inventar (60) = 5,0 g C m-2 d-1 (600) = 50,04 g C m-2 d-1 (1000) = 83,4 g C m-2 d-1
Model 4b (60) = 5,65 g C m-2 d-1 (600) = 56,49 g C m-2 d-1 (1000) = 94,15 g C m-2 d-1
Station 25: Corg-Bedarf 65,7 mg C m-2 d-1
Inventar (60) = 2,06 g C m-2 d-1 (600) = 20,59 g C m-2 d-1 (1000) = 34,31 g C m-2 d-1
Model 4b (60) = 0,17 g C m-2 d-1 (600) = 1,66 g C m-2 d-1 (1000) = 2,77 g C m-2 d-1
Station 27: Corg-Bedarf 8,1 mg C m-2 d-1
Inventar (60) = 1,48 g C m-2 d-1 (600) = 14,79 g C m-2 d-1 (1000) = 24,65 g C m-2 d-1
Model 3 (60) = 0,4 g C m-2 d-1 (600) = 3,96 g C m-2 d-1 (1000) = 6,6 g C m-2 d-1
Hier wird deutlich, dass der über die Modelle berechnete Kohlenstofffluss nur an Station 26
ausreicht, um den Bedarf an Corg zu decken. Bei einem Chl.a/Corg-Verhältniss von 600 wird an
2 Stationen des Vigo-Profils und einer Station des Nazaré-Canyons der von FRANKE (2000)
über die Sauerstoffzehrung berechnete Kohlenstoffbedarf gedeckt. Im Falle der Station 27
würde der Fluss, der über den Chlorophyll-Inventar berechnet wurde ausreichen um den
Kohlenstoffbedarf an dieser Station zu decken.
58
Das Defiziet an, aus dem Pelagial sedimentierendem, organischem Kohlenstoff für das
Benthos der Kontinentalhänge ist ein viel diskutiertes Thema (SMITH, 1987; SMITH, 1989;
JAHNKE et al., 1990; GRAF, 1992; GRAF et al., 1995; LAMPITT et al., 1995). Die Autoern
schlussfolgerten, dass ein erheblicher Teil organischen Materials über laterale Advektion dem
Benthos zugeführt werden muß. Diese Prozesse sollten in der Bodennepheloidschicht (BNL)
stattfinden (JAHNKE et al., 1990; BISCAYE et al., 1994; GRAF et al., 1995). Im Goban-
Spur-Gebiet haben THOMSEN und GRAF (1995) hauptsächlich refraktären Kohlenstoffs in
der BNL festgestellt, der somit auch nicht ausreicht um den benthischen Kohlenstoffbedarf zu
decken. THOMSEN (1998) stellte für das Goban-Spur-Gebiet eine Bilanzierung des
benthischen Kohlenstoffbedarfs über laterale hangabwärtsgerichtete Transportprozesse in der
Bodengrenzschicht dar. Er geht von pulsartig eingetragenen großen Mengen Phytodetritus in
den oberen Hangbereichen aus, die den dortigen Kohlenstoffbedarf übertreffen. Über mehrere
benthische Resuspensionschleifen gelangt das Material in strömungsberuhigte Bereiche und
steht dort dem Benthos zur verfügung. Das Material liegt größtenteils in Form größerer
Aggregate vor, in denen der Kohlenstoff gebunden ist. Die Aggregate sind von Bakterien

Diskussion
besiedelt, die gelöste Substanzen in partikuläre Biomasse umwandeln. Die
Bakterienbiomasse, die sich auf den Aggregaten befindet, stellt somit eine zusätzliche
Kohlenstoffquelle dar. Diese Möglichkeit wäre auf Grund der Beschaffenheit des
Kontinentalhanges vor Vigo für dieses Gebiet zutreffend.
Für den Nazaré-Canyon treffen eher die Annahmen von SORBE (1999) und MONACO
(1990) zu. Bis auf Station 26 sind die Stationen im Nazaré-Canyon auf erheblichen lateralen
Eintrag angewiesen (FRANKE, 2000). Der Nazaré-Canyon ist ein inaktiver Canyon, d.h. es
finden pulsartige Sedimentationsereignisse statt die große Mengen Material in die Tiefe
transportieren und in topographischen Senken ablagern. Solche Plume events wurden von
THOMSEN et al., (eingereicht, aus THOMSEN, 1998) am westlichen Barents See
Kontinentalhang beschrieben und werden für das spanisch/portugisische Auftriebsgebiet
erwartet.
Bei einem Chl.a/Corg-Verhältniss von 600 handelt es sich für Tiefsee-Bedingungen um relativ
frisches Material. Für Station 26 könnte ein kurz vor der Probennahme stattgefundenens
Sedimentationsereigniss für relativ frisches Material gesorgt haben. Oder aber es ist etwas
älteres Material, was im flacheren Teil des Canyons sedimentierte. Wie unter 4.3 beschrieben
könnte dieses Material durch erhöhte Bioturbation aufglockert worden sein (DILLON und
ZIMMERMAN, 1970; HECKER, 1982; MALAHOFF et al., 1982 aus GAGE und TYLER,
1991) und dann durch hydrodynamische Einflüsse abrutschen (LUCKENBACH, 1986).
Das der Kohlenstoffbedarf selbst bei einem Chl.a/Corg-Verhältniss von 1000 an den unteren
Stationen des Nazaré-Canyons noch nicht gedeckt wird, zeigt, dass das vorgefundene
Chlorophyll sehr alt sein muß. Da an diese Stationen erheblich Mengen Pflanzenfragmente
gefunden wurden und FRANKE (2000) C/N-Verhältnisse von 12,5 für Station 25 und 14 für
Station 27 ermittelt hat, kann davon ausgegangen werden, dass das vorgefundene Material
nicht aus einem „jungen“ Sedimentationsereigniss stammt. Viel mehr liegt die Vermutung
nahe, dass durch die oben erwähnten Plume events so große Mengen Material durch den
Canyon in die Tiefe transportiert werden, dass am Canyonende ein erhebliche Überschuss
vorhanden ist, der durch die dortige Fauna nicht sehr schnell abgebaut wird.
Sicher ist jedoch, dass der Nazaré-Canyon großen Anteil am Transport von auf dem Schelf
und dem Kontinentalhang produzierten Material in die Tiefsee hat.
59
Da der Bedarf bei 3 von 6 Sationen des Probennahmegebietes nicht über die Zufuhr von
Chlorophyll a gedeckt wird, müsste die oben erwähnte, auf Aggregaten lokalisierte
Bakterienbiomasse in Betracht gezogen werden. Vielleicht reicht diese aus, um das Defiziet
an den Stationen 24, 25 und 27 auszugleichen

Zusammenfassung
5 Zusammenfassung Ziel dieser Arbeit war es, das Benthal des nordwestlichen Kontinentalhanges der Iberischen
Halbinsel an Hand der Makrofauna und der Bioturbation zu charakterisieren. Die
Probennahme fand im Winter statt, und daher beziehen sich die Ergebnisse ausschließlich auf
diese Jahreszeit.
Das Probennahmegebiet an der Iberischen Westküste ist nach THOMSEN (1998) ein Typ III
Kontinentalhang. Auf Grund des schmalen Schelfs von 20-40 km und dem steilen
Kontinentalhang welcher mit zahlreichen Canyon durchzogen ist gilt dieser Typ als
hochreaktiv. Das Gebiet wird durch saisonal (Sommer/Winter) auftretende Auf- und
Abtriebsereignisse beeinflusst.
Die Makrofaunadichte am Kontinentalhang vor Vigo ist für ein so hochproduktives Gebiet
sehr gering (1616 Ind. m-2 an der Schelfstation 20) folgt aber dem gängigen Trend der
Abnahme der Faunendichte mit zunehmender Wassertiefe und größer werdendem Abstand
zum Land (Station 11 = 907 Ind. m-2 und Station 24 1410 Ind. m-2) ebenso wie die Biomasse.
Die Biomasse sinkt von 0,244 g C m-2 an Station 20 über 0,223 g C m-2 an Station 11 auf
0,212 g C m-2 an Station 24. Die Makrofaunazusammensetzung entspricht Untersuchungen an
anderen Kontinentalhängen wie Goban Spur (FLACH und HEIP, 1996) und Untersuchungen
am Kontinentalhang Galiziens (FLACH et al. 1999).
Die Makrofaunadichte des Nazaré-Canyons ist wesentlich höher als die des Vigo-Profils
(117566 Ind. m-2 an Station 26 und 5001 Ind. m-2 an Station 25). Dies wird als Merkmal für
den inaktiven Charakter des Canyons gedeutet (SORBE, 1999). Die höchste Biomasse wurde
an Station 25 mit 1,271 g C m-2 ermittelt. An Station 26 konnten nur 0,217 g C m-2
festgestellt werden, was mit der geringen Masse der Cossuriden-Köpfe zusammenhängt. Die
hohe Dichte der Cossuridae an Station 26 könnte ein Indiz für einen kurz vor der
Probennahme von statten gegangenen Turbidit sein.
Die Verteilung der Polychaeten-Familien am Vigo-Profil ist Vergleichbar mit anderen
Kontinentalhängen (Goban-Spur; Kontinentalhang Galiziens), ebenso wie deren
Ernährungstypen. Am Schelf besteht eine homogene Verteilung zwischen Deposit und
Suspensionsfressern. Mit zunehmender Wassertiefe steigt die Dominanz der Depositfresser.
Im Nazaré-Canyon können genaue Angaben zu der Verteilung der Polychaeten-Familien im
Vergleich zu anderen Canyons nicht gemacht werden, da nur an Station 25 eine Polychaeten-
Gemeinschaft nachgewiesen werden konnte. Diese entspricht aber Ergebnissen aus dem Cap-
68

Zusammenfassung
Ferret-Canyon in der Biscaya (GERINO et al. 1999) und dem Carson-Canyon in der Region
der Grand Banks im nordwestlichen Atlantik (HOUSTON und HAEDRICH, 1984).
Die sinkenden Chlorophyll a Werte mit zunehmender Wassertiefe und wachsendem Abstand
vom Schelf (Station 20: 0,319 µg cm-3; Station 11: 0,14 µg cm-3; Station 24: 0,06 µg cm-3;
alle Werte aus dem 1 Sedimentzentimeter) entsprechen den typischen Beobachtungen an
Kontinentalhängen, und der sinkenden Primärproduktion in der euphotischen Zone mit größer
werdendem Abstand vom Schelf.
Die Chlorophyll a Werte im Canyon liegen um den Faktor 1,5-6,5 höher als am Vigo-Profil,
wobei keine deutliche Abnahme mit der Tiefe zu beobachten ist (Station 26: 0,463 µg cm-3;
Station 25: 0,464 µg cm-3; Station 27: 0,425 µg cm-3; alle Werte aus dem 1
Sedimentzentimeter).
Die mit den Bioturbationsmodellen nach SOETAERT et al., (1996) ermittelten
Bioturbationskoeffizienten und Oberflächenflüsse folgen am Vigo-Profil dem allgemeinen
Trend. An Station 20 wurde ein Db von 2,1 cm2 d-1 errechnet. Von Station 20 fällt der Db
über Station 11 mit 0,11 cm2 d-1 auf 0,029 cm2 d-1 an Station 24.
Im Nazaré-Canyon konnte solch ein Trend nicht festgestellt werden. Ebenso konnte keine
Korrelation zu den ermittelten Biomassedaten gefunden werden. Der höchste
Bioturbationskoeffizient wurde an der tiefsten Station im Canyon (St.27) errechnet. Hier
beträgt er 0,024 cm2 d-1. An Station 25, die die höchste Biomasse im gesamten
Untersuchungsgebiet aufweist wurde der geringste Db-Wert von 0,005 cm2 d-1 gefunden. An
Station 26 beträgt der Db 0,015 cm2 d-1.
Die von den Modellen berechneten Oberflächenflüsse für Chl.a an Station 24 des Vigo-Profils
sowie an den Stationen 25 und 27 des Nazaré-Canyons ergaben Corg-Flüsse, die nicht
ausreichen würden den Kohlenstoffbedarf (berechnet von FRANKE, 2000) an den
betreffenden Stationen zu decken.
69

Literatur
6 Literatur
Anderson, R.F., Rowe, G.T., Kemp, P.F., Trumbore S., Biscaye, P.E., 1994. Carbon
budget for the mid -slope depocenter of the Middle Atlantic Bight. Deep-Sea
Resaerch II 41(2/3), 669-703.
Antoine, D., André, J.M., Morel, A.,1996. Oceanic primary production 2. - Estimation at
global scale from satellite (coastal zone color scanner) chlorophyll. Global
Biogeochemical Cycles 10(1), 57-69.
Barnett, P.R.O., Watson, J., Conelly, J.W., 1984. A multiple corer for taking virtually
undisturbed samples from shelf, bathyal and abyssal sediments. Oceanologica Acta
Batten, M.L., Lopes-da-Costa,C.N., Nelson,-C.S., 1992. A numerical study of wind stress curl
effects on eddies and filaments off the northwest coast of the Iberian Peninsula.
Journal of Marine Systems 3(3), 249-266
Berner, R.A., 1980. EARLY DIAGENESIS, A Theoretical Approach. Princeton University
Press
Biscaye, P.E., Anderson, R.F., Deck, B.L., 1988. Fluxes of particles and constituents to the
eastern United States continental slope and rise: SEEP-I. Continental Shelf
Resaerch 8(2), 855-904.
Biscaye, P.E., Flagg,C.N., Falkowski,P.G., 1994. The shelf edge exchange processes
experiment, SEEP-II: An introduction to hypotheses, results and conclusions.
Deep-Sea Resaerch II 41(2/3), 231-252.
Blanton, J.O., Tenore, K.R., Castillejo, F., Atkins, L.P., Schwing, F.B., Lavín, A., 1987. The
relationship of upwelling to mussel production in the rías of the western coast of
Spain. Journal of Marine Research 45, 497-511.
Boudreau, B.P., 1986a. Mathmetics of Tracer Mixing in Sediments: 1. Spatially-Dependent,
Diffusifve Mixing. American Journal of Science 286, 161-198
Boudreau, B.P., 1986b. Mathmetics of Tracer Mixing in Sediments: 2. Nonlocal Mxing and
Biological Conveyor-Belt Phenomena. American Journal of Science 286, 199-238
Boudreau, B.P., Imbiden, D.M., 1987. Mathmetics of Tracer Mixing in Sediments: 3. The
Theory of Nonlocal Mixing within Sediments. American Journal of Science 287, 693-
719
70

Literatur
Boudreau, B.P., 1994. Is burial velocity a master parameter for bioturbation? Geochimica et
Cosmochimica Acta 58(4), 1243-1249
Boudreau, B.P., 1998. Mean mixed depht of sediments: The wherefore and the why. Limnol.
Oceanogr. 43(4), 524-526
Dickson, R.R., Gould, W.J., Müller, T.J., Maillard, C., 1985. Estimates of the mean
circulation in the deep (>2000 m) layer of the eastern North Atlantic. Progress in
Oceanography 14, 103-127.
Duineveld, G.C.A., Lavaleye, M.S.S., Berghuis, E.M., de Wilde, P.A.W.J., van der Weele,
J., Kok, A., Batten, S.D., De Leeuw, J.W., 1997. Patterns of benthic fauna and
benthic pespiration on the Celtic continental margin in relation to the distribution of
phytodetritus. Internationale Revue der gesamten Hydrobiologie 82(3), 395-424.
Etcheber, H., Relexans, J.C., Beliard, M., Weber, O., Buscail, R., Heussner, S., 1999.
Distribution and quality of sedimentary organic matter on the Aquitanian margin
(Bay of Biscay). Deep-sea Research II 46, 2249-2288.
Fauchald, K., Jumars, P.A., 1979. The Diet of Worms: A Study of Polychaete feeding
Guilds. Ocanog. Mar. Biol. Ann. Rev. 17, 193-284
Flach, E., Heip, C., 1996a. Seasonal variations in faunal distribution and activity across the
continental slope of the Goban Spur area (N.E. Atlantic). Journal of Sea Research
36(3/4), 203-215.
Flach, E., Heip, C., 1996b. Vertical distribution within the sediment of macrozoobenthos
along the continental slope in the Goban Spur area (N.E. Atlantic). Marine Ecology
Progress Series 141, 55-66.
Flach, E., Heip, C., Muthumbi, A., Sandee, A., 1999. Benthic community structure in relation
to sediment composition at the Iberian Margin. OMEX II-II sec. Annual Report, 229-
239
Fraga, F., 1981. Upwelling off the Galician coast, northwest Spain. In: Richards, F.A. (Ed.),
Coastal upwelling, American Geophysical Union, Washington, DC, 176-182.
Franke, U., 2000. Charakterisierung des Benthals des nordwestlichen Kontinentalhangs der
Iberischen Halbinsel: Biogeochemische Stoffflüsse. Dissertation, Univ. Rostock
Gage, J.D., Tyler, P.A., 1991. Deep-sea biologie. A natural history of organisms at the
deep-sea floor. Cambridge University Press Cambridge, 1-504.
Gardner, W.D., 1989. Baltimore Canyon as a modern conduit of sediment to the deep sea.
Deep-sea Research 36, 323-358.
71

Literatur
Gerino, M., Aller, R.C., Lee, C., Cochran, J.K., Aller, J.Y., Green, M.A., Hirschberg, D.,
1997. Comparison of Different Tracers and Methods Used to Quantify Bioturbation
During Spring Bloom: 234-Thorium, Luminophores and Chlorophyll a. Estuarien,
Coastal and Shelf Science 46, 531-547
Gerino, M., Stora, G., Weber, O., 1999. Evidence of bioturbation in the Cap-Ferret Canyon in
the deep northeastern Atlantic. Deep-Sea Resaerch II 46, 2289-2307
Graf, G., 1989. Benthic-pelagic coupling in a deep-sea benthic community. Nature 341, 437-
438
Graf, G., 1992. Benthic pelagic coupling: a benthic view. Oceanography and Marine
Biological Annual Reviews 30, 149-190.
Graf, G., Gerlach, S.A., Linke, P., Queisser, W., Ritzrau, W., Scheltz, A., Thomsen, L.,
Witte, U., 1995. Benthic-pelagic coupling in the Greenland-Norwegian Sea and its
effect on the geological record. Geologische Rundschau 84, 49-58.
Graf, G., Rosenberg, R., 1997. Bioresuspension and biodeposition: a review. Journal of
Marine Systems 11, 269-278
Graf, G., Ritzrau, W., 1997. Biologische Aepekte sedimentologischer Prozesse.
Geowissenschaften 15, 16-18
Hartmann-Schröder, G., 1996. Die Tierwelt Deutschlands, 58.Teil, Anelida, Borstenwürmer,
Polychaeta. Gustaf Fischer 2
Haynes, R., Barton, E.D., Pilling, I., 1993. Development, Persistance, and Variability of
Upwelling Filaments off the Atlantic Coast of the Iberian Peninsula. Journal of
Geophysical Research 98(C12) 22681-22692
Heussner, S., de Madron, X.D., Radakovitch, O., Beaufort, L., Biscaye, P.E., Carbonne, J.,
Delsaut, N., Etcheber, H., Monaco, A., 1999. Spatial and temporal patterns of
downward particle fluxes on the continental slope of the Bay of Biscay
(northeastern Atlantic). Deep-Sea Research II 46, 2101-2146.
Holm-Hansen, O., Lorenzen, C.J., Holmes, R.W., Strickland, D.H., 1965. Fluorometric
Determination of Chlorophyll. J. Cons. Int. Explor. Mer 30(1), 3-15
Houston, K. A., Haedrich, R.L., 1984. Abundances and biomass of macrobenthos in the
vicinity of Carson Submarine Canyon, northwest Atlantic Ocean. Marine Biology 82,
301-305
72

Literatur
Huthnance, J.M., 1995. Circulation, exchange and water masses at the ocean margin: the
role of physical processes at the shelf edge. Progress of Oceanography 35,
353-431.
Jahnke, R.A., Reimers, C.E., Craven, D.B., 1990. Intensification of recycling of organic
matter at the sea floor near ocean margins. Nature 348, 50-54.
Jeffrey, S.W., Humphrey, G.F, 1975, New Spectrophotometric Equations for Determination
Chlorophyll a,b,c1 and c2 in Higher Plants, Algae and Natural Phytoplankton. Biocem.
Physiol. Planzen 167, 191-194
Jumars, P.A., Gallagher, E.D., 1982. Deep-Sea Cmmunity Structure: Three Plays on the
Benthic Proscenium. 217-255
Lampitt, R.S., Raine, C.T., Billett, D.S.M., Rice, A.L., 1995. Material supply to the european
continental slope: A budget based on benthic oxygen demand and organic supply.
Deep-Sea Research I 42(11/12), 1865-1880.
Lavaleye, M.S.S., Duineveld, G.C.A., Berghuis, E.M., 1999. Megabenthos, benthic
respiration and phytopigments off N.W. Spain. OMEX II-II sec. Annual Report,
111-118.
Lohse, L., Helder, W., Epping, E.H.G., Balzer, W., 1998. Recycling of organic matter along
a shelf-slope transect across the N.W. european continental margin (Goban Spur).
Progress in Oceanorgaphy 42, 77-100.
López-Jamar, E., Cal, R.M., González, G., Hanson, R.B., Rey, J., Santiago, G., Tenore,
K.R.,1992. Upwelling aund outwelling on the benthic regime of the continental shelf
off Galicia, N.W. Spain. Journal of Marine Research 50, 465-488.
Luckenbach, M.W., 1986. Sediment stability around animal tubes: the role of
hydrodynamic processes and biotic activity. Limnology and Oceanorgaphy 31, 779-
Lutze, G.F., Altenbach, A., 1991.Technik und Signifikanz der Lebendfärbung benthischer
Foraminiferen mit Bengalrot. Geologisches Jahrbuch A128, 251-265
Miller, P., Groom, S., Smyth, T., Miguel-Lago, M., 1999. Remute sensing in OMEX II-II.
OMEX II-II sek. Annual Report, 73-82.
Monaco, A., Biscaye, P., Soyer, J., Pocklington, R., Heussner, S., 1990. Particle fluxes
and response on a continental margin: The 1985-1988 mediterranean ECOMARGE
experiment. Continental Shelf Research 10(9-10), 809-839.
Murray, J.W., Barber, R.T., Roman, M.R., Bacon, M.P., Feely, R.A., 1994. Physical and
biological controls on carbon cycling in the equatorial Pacific. Science 266, 58-65.
73

Literatur
Nuglisch, K., 1985. Foraminiferen- marine Mikroorganissmen. A. Ziemsen Verlag, DDR-
4600 Wittenberg Lutherstadt
Pfannkuche, O., Soltwedel, T., 1998. Small benthic size classes along the N.W. european
continental margin: spatial and temporal variability in activity and biomass. Progress
in Oceanorgaphy 42, 189-207.
Pingree, R.D., New, A.L., 1991. Abyssal penetration and reflexion of internal tidal energy
in the Bay of Biscay. Journal of Physical Oceanography 21, 28-39.
Prego, R., Bao, R., 1997. Upwelling influence on the Galician Coast: silicate in shelf water
and underlying surface sediments. Continental Shelf Research 17(3) 307-318
Richter, G., 1988. Stoffwechselphysiologie der Pflanzen. Georg Thieme 5
Riedl, R., 1983. Fauna und Flora des Mittelmeeres. Verlag Paul Parey 3
Sibuet, M., Lambert, C.E., Chesselet, R., Laubier, L., 1989.Density of the major size groups
of benthic fauna and trophic input in deep basins of the Atlantic Ocean. Journa of
Marine Reasearch 47, 851-867
Smith, C.R., Hessler, R.R., 1987. Colonization and Succession in Deep-sea Ecosystems. Tree
2(12), ?-?
Smith, K.L. Jr., 1987. Food energy supply and demand: A discrepancy between particulate
organic carbon flux and sediment community oxygen consumption in the deep ocean.
Limnology and Oceanography 32(1), 201-220.
Soetaert, K., Herman, P.M.J., Middelburg, J.J., Heip, C., deStigter, H.S., van Weering,
T.C.E., Epping, E., Helder, W., 1996. Modelling 210Pb-derived mixing activity in
ocean margin sediments: Diffusive versus nonlocal mixing. Journal of Marine
Research 54, 1207-1227
Soltwedel, T., 1997.Temporal variabilities in benthic activity and biomass on the western
European continental margin. Oceanologica Acta 20, 871-879
Sorbe, J.C., 1999. Deep-Sea macrofaunal assemblages within the Benthic Boundary Layer of
the Cap-Ferret Canyon (Bay of Biscay, NE Atlantic). Deep-Sea Research II 46, 2309-
2329
Sousa, F.M., Bricaud, A. 1992. Stellite-Derived Phytoplankton Pigment Structures in the
Portuguese Upwelling Area. Journal of Geophysical Research 97(C7), 11343-11356
Stephens, M.P., Kadko, D.C., Smith, C.R., Latasa, M., 1997. Chlorophyll-a and
pheopigments as tracers of labile organic carbon at the central equatorial Pacific
seafloor. Geochimica et Cosmochimica Acta 61(21), 4605-4619.
74

Literatur
Sun, M., Aller, R.C., Lee, C., 1993. Early diagenesis of Chlorophyll a in Long Island Sound
sediments: A measure of carbon flux and particle reworking. Journal of Marine
Research 49, 379-401
Sun, M., Aller, R.C., Lee, C., 1994. Spatial and temporal distributions of sedimentary
chloropigments as indicators of benthic processes in Long Island Sound. Journal of
Marine Research 52, 149-176
Tenore, K.R., Alonso-Noval, M., Alvarez-Ossorio, M., Atkinson, L.P., Cabanas, J.M.,
Cal, R.M., Campos, H.J., Castellejo, F., Chesney, E.J., Gonzalez, N., Hanson, R.B.,
McClain, C.R., Miranda, A., Roman, M.R., Sanchez, J., Santiago, G., Valdes, L.,
Varela, M., Yoder, J., 1995. Fisheries and oceanography off Galicia, N.W. Spain:
Mesoscale spatial and temporal changes in physical processes and resultant
patterns off biological productivity. Journal of Geophysical Research 100(6),
10943-10966.
Thomsen, L, Graf, G., 1995. Characteristics of suspended particulate matter in the benthic
boundery layer of the continental margin of the western Barent Sea. Oceanologica
Acta 17(6),597-607.
Thomsen, L., van Weering, Tj.C.E., 1998. Spatial and temporal variability of particulate
matter in the benthic boundary layer at the N.W. european margin (Goban Spur).
Progress in Oceanorgaphy 42, 61-71.
Thomsen, L., 1998. Prozesse in der Bodengrenzschicht an Kontinentalhängen und ihre
Bedeutung für den benthischen Kohlenstoffkreislauf. Habilitationsschrift,
Mathematisch-Naturwissenschaftliche Fakultät der Christian-Albrechts-Universität
zu Kiel.
Vanney, J.R., Mougenot, D., 1990. Un canyon sous marin du type „gouf“: Le canhao da
Nazaré (Portugal). Oceanologica Acta 13(1), 1-14.
Walsh, J.J., Rowe, G.T., Iverson, R.L., McRoy, C.P., 1981. Biological export of the shelf
carbon: a neglected sink of the global CO2 cycle. Nature 291, 196-201.
Walsh, J.J., 1991. Importance of continental margins in the marine biogeochemical cycling
of carbon and nitrogen. Nature 350, 53-55.
van Weering, T.C.E., McCave, I.N., Hall, I.R., 1998a. Ocean Margin Exchange (OMEX I)
benthic process study. Progress in Oceanography 42, 1-4.
van Weering, T.C.E., Hall, I.R., de Stigter, H.C., McCave, I.N., Thomsen, L., 1998b.
Recent sediments, sediment accumulation and carbon buriel at Goban Spur, N.W.
european continental margin (47-50° N). Progress in Oceanography 42, 5-35.
75

Literatur
van Weering, T.C.E., de Stigter, H.C., 1999. Recent sediment transport and accumulation
on the western Iberian margin. OMEX II-II sek. Annual Report, 83-92.
Wheatcroft, R.A., Jumars, P.A., Smith, C.R., Nowell, A.R.M., 1990. A mechanistic view of
the particle biodiffusion coefficient: Step lenght, rest periods and transport directions.
Journal of Marine Research 48, 177-207
Wheatcroft, R.A., 1992. Experimental test for particle size-dependent bioturbation in the deep
ocean. Limnol. Oceanogr. 37(1), 90-104
Wooster, W.S., Bakun, A., McLain, D.R., 1976. The seasonal upwelling cycle along the
eastern boundary of the North Atlantic. Journal of Marine Research 34, 131-141.
76

Danksagung
Herrn Prof. Graf danke Ich für die Bereitstellung dieses Diplomthemas und die konstruktive Kritik des
Manuskriptes, sowie die zum Ende hin immer angenehmere Betreuung.
Frau Prof. Lochte danke Ich für das kritische Lesen dieser Arbeit.
Ganz besonders möchte Ich mich bei Anja Kähler bedanken für die klasse Einführung in die
Chlorophyll-Bestimmung und das unermüdliche Beantworten meiner vielen und häufigen Fragen. Eva
Philipp danke ich für die Hilfe beim Wühlen im Schlamm.
Besonders bei Robert Turnewitsch und Martin Powilleit bedanke ich mich für viele gute, anregende
Gespräche und die hilfreichen Bemerkungen zum Manuskript.
Für all die Hilfe an Bord des FS Meteor und die gute Atmosphäre sei der Besatzung der Meteor
herzlich gedankt sowie den Mitreisenden, insbesondere Uli Franke, ohne deren Hilfe diese Arbeit nie
zustande gekommen wäre.
Ein besonderer Dank gilt Herrn Dr. Bick, der mir bei der Bestimmung der Polychaeta sehr geholfen
hat.
Holger Pielenz danke ich für seine Hilfsbereitschaft beim Lösen technischer Probleme.
Ausserdem möchte ich mich bei meinen Mitstreitern Kai, Markus und Uli bedanken, die immer mit
Rat und Tat und vor allem guter Laune zur Stelle waren.
Der größte Dank gilt meinen Eltern ohne die Ich nie so weit gekommen wäre. Sie haben mich in jeder
Hinsicht unterstützt und immer Verständnis und Interesse gezeigt. Genauso danke ich meiner lieben
Omi sowie Opa und Mummi, die an dieser Unterstützung maßgeblich beteiligt waren.

Erklärung
Hiermit erkläre Ich an Eides statt, daß Ich diese Arbeit selbständig und nur unter Verwendung der
angegebenen Hilfsmittel verfaßt habe.
Florian Peine
Rostock im Juli 2000