prüfung der biokompatibilität von implantatwerkstoffen mit ... · danksagung iv danksagung an...
TRANSCRIPT

Prüfung der Biokompatibilität von
Implantatwerkstoffen mit Methoden
der digitalen Bildverarbeitung
Dissertation
zur Erlangung des akademischen Grades
DOKTOR–INGENIEUR
der Fakultät für
Mathematik und Informatik
der FernUniversität
in Hagen
von
Sven Buhl M.Sc.
Menden
Hagen 2015

Kurzfassung ii
Kurzfassung
Die vorliegende Dissertation beschreibt die Entwicklung einer vielseitig einsetz-
baren Zellanalysesoftware für die Biokompatibilitätsprüfung von Medizinproduk-
ten. Derzeit besteht ein hoher Bedarf an Zellanalysesystemen mit automati-
scher Auswertung im Rahmen der Biokompatibilitätsprüfung, da die Prüfungen
manuell und somit sehr kostenintensiv sowie fehleranfällig durchgeführt wer-
den. Im Rahmen der Arbeit werden die zwei Zelltypen L929 und MC3T3 unter-
sucht. Die Herausforderung liegt zum einen in der starken morphologischen
Variation der Zelltypen und die vergleichsweise kontrastschwache May-
Grünwald Zellfärbung. Die Linie L929 findet im Gegensatz zum Typ MC3T3 im
Bereich der Biokompatibilitätsprüfungen häufig Anwendung. Der Typ MC3T3
weist hingegen eine andere morphologische Erscheinung auf und dient zur
Verallgemeinerung der Analysealgorithmen. Die Aufnahme der May-Grünwald
angefärbten Zellen auf metallischen Oberflächen erfolgt mit einem Olympus
Bx51M bei 100-facher Vergrößerung. Für Zellen auf dem Material Polystyrol
kommt das Durchlichtmikroskop CKX41 ebenfalls von Olympus zum Einsatz.
Die Bilder für die automatische Zellauswertung werden mit der Farbkamera
Olympus XC10 mit einer Auflösung von 1376x1032 Pixel aufgenommen. Die
vorliegenden Bilder können aufgrund des Prozesses der Probenaufbereitung
Kratzer und Verunreinigungen enthalten, die im Rahmen der Bildvorverarbei-
tung zunächst reduziert werden. Die anschließende Segmentierung der Zellbe-
reiche erfolgt abhängig vom gewählten Mikroskopkontrast mit einem Schwell-
wertverfahren oder einer histogrammbasierten Methode. Zur Bestimmung des
Biokompatibilitätsgrades eines untersuchten Materials ist die Proliferationsrate
ein wichtiges Kriterium. Daher ist eine Erfassung agglomerierter Zellen uner-
lässlich. Zur Trennung zusammengewachsener Zellen kommen abhängig vom
Zelltyp verschiedene Verfahren zum Einsatz. Liegen Zellen vor, die meistens
Einschnürungen an den Zellkontaktstellen aufweisen, z.B. der Typ L929, kommt
ein Separationsverfahren auf Basis der dominanten Konturpunkte zum Einsatz.
Große und meist flächige Agglomerate, z.B. beim Typ MC3T3, werden hinge-
gen mit einer Wachstumssimulation ausgehend von den mit dem Histogram
Backprojection Algorithmus detektierten Kernbereichen aufgetrennt. Eine kon-
textbasierte Trennfunktion mit automatischer Parameterfestlegung durch einen
genetischen Algorithmus kommt zum Einsatz um die benötigte Rechenzeit zu
reduzieren und die Genauigkeit der Zellzählung zu erhöhen. Die Software ist so
konzipiert, dass die Auswahl des passenden Zellsegmentierungs- und Separa-
tionsverfahrens automatisch erfolgen kann. Die Abweichung der automatischen
Zellzahlbestimmung zur Referenzzählung liegt mit durchschnittlich 1,27% für
den Zelltyp L929 und 3,5% für den Typ MC3T3 im akzeptablen Bereich für die
Biokompatibilitätsprüfung.

Abstract iii
Abstract
This thesis describes the development of a versatile applicable cell analysis
software regarding the biocompatibility testing of medical products. At present
there is a high demand for cell analysis systems with automatic evaluation in
the field of biocompatibility testing due to the expansive and error-prone manual
process. In the context of this thesis the two cell types L929 and MC3T3 are
investigated. The challenge is the strong morphological variation of the cell
types and the comparative low contrast May-Grünwald cell staining. In contrast
to the type MC3T3 the type L929 is often used for biocompatibility tests. The
type MC3T3 is characterized by another morphological appearance and is used
to generalize the algorithms. The image acquisition of the May-Grünwald
stained cells on top of the metallic surface is done by a Olympus Bx51M at
magnification factor 100. Cells on top of polystyrol are analyzed by the Olympus
transmitted-light microscope CKX41. The images used for the automatic cell
analysis are acquired by the color camera Olympus XC10 with a resolution of
1376x1032 pixels. The images are containing scratches caused by the process
of the sample preparation which are in a first step reduced by an image prepro-
cessing step. The following cell segmentation is done depending on the chosen
microscope contrast with a threshold method or a histogram based algorithm.
The proliferation rate is an important criterion to determine the grade of biocom-
patibility. Therefor a detection of clustered cells is essential. Connected cells
are divided depending of the cell type with different methods. If cells have to be
analyzed which have joints at their contact areas the separation method based
on dominant contour points is used. Big Clusters without joints at their contact
areas are divided by a growth simulation starting from the cell nuclei detected
by the histogram backprojection algorithm. To reduce the calculation time and
to improve the cell count accuracy a context based separation method with au-
tomatic parameter adjustment done by a genetic algorithm is used. The choice
of the suitable cell segmentation and separation method can be done automati-
cally. The difference of the automatic cell count to the reference cell count is on
average 1.27% for the type L929 and 3.5% for the type MC3T3 which is ac-
ceptable in the field of biocompatibility testing.

Danksagung iv
Danksagung
An dieser Stelle möchte ich mich bei Allen bedanken, die zum Gelingen dieser
Arbeit in Form von fachlichen Ratschlägen, Diskussionen und Motivation beige-
tragen haben.
Mein Dank gilt auch Prof. Dr. Jürgen Jahns, der es mir ermöglichte, dieses Vor-
haben an der FernUniversität Hagen umzusetzen und mit Rat zur Seite stand
sowie Prof. Dr. Wolfram Schiffmann für seine konstruktiven Ratschläge. Insbe-
sondere möchte ich Prof. Dr. Burkhard Neumann danken, der mir in zahlreichen
Diskussionen stets mit seinem äußerst umfangreichen Fachwissen zur Seite
stand und mir darüber hinaus neue Sichtweisen über Problemstellungen im
Rahmen des Projektes eröffnete. Des Weiteren möchte ich allen im Rahmen
des Forschungsprojektes wirkenden Personen dafür danken, dass ich ein sehr
interessantes Thema bearbeiten durfte. Durch die interdisziplinäre Aufstellung
des Projektteams und den sich daraus ergebenen fachübergreifenden Diskus-
sionen hat sich mein zu Beginn sehr eingeschränktes Vorwissen auf dem Ge-
biet der Biotechnologie deutlich erweitert. Ebenso möchte ich Anna Severing
meinen Dank aussprechen, dafür dass sie mit starkem Engagement zahlreiche
biotechnologische Versuche durchgeführt und mir in vielen Projektbesprechun-
gen die Welt der Biotechnologie näher gebracht hat. Darüber hinaus bedanke
ich mich bei Martin Föller und Sebastian Christopher Schäfer, die im Rahmen
des Forschungsprojektes bei der Softwareentwicklung mitgewirkt haben.
Beim BMBF bedanke ich mich für die Finanzierung des interdisziplinären For-
schungsprojektes „Computer Vision zur Biokompatibilitätsprüfung von Implan-
tatwerkstoffen“ (Förderkennzeichen: 17033X10).
Des Weiteren möchte ich mich bei meinen ehemaligen Kollegen an der FH
Südwestfalen, insbesondere Herrn Friedhelm Krause für die sehr angenehmen
Jahre der Zusammenarbeit bedanken.
Ebenso bedanke ich mich bei den Herren Bernd Dippel und Christian Schäfer
für das Korrekturlesen der Arbeit.
Von Herzen möchte ich mich bei meinen Eltern bedanken, ohne Euch hätte ich
das nie geschafft! Auch meiner Freundin Senada Mulalic gilt mein Dank für ihre
Unterstützung, ihr Verständnis und ihre Geduld.

Inhaltsverzeichnis v
Inhaltsverzeichnis
Kurzfassung ........................................................................................................ ii
Abstract .............................................................................................................. iii
Danksagung ....................................................................................................... iv
Inhaltsverzeichnis ................................................................................................ v
Symbolverzeichnis ............................................................................................. ix
Tabellenverzeichnis ............................................................................................. x
Abbildungsverzeichnis .................................................................................... xviii
1. Einleitung ........................................................................................................ 1
1.1 Motivation und Zielsetzung .................................................................. 3
1.2 Gliederung der Arbeit ........................................................................... 6
2. Stand der Forschung ...................................................................................... 8
2.1 Segmentierungsverfahren für biomedizinische Anwendungen ............ 8
2.2 Verfahren zur Separation agglomerierter Zellen ................................ 10
2.3 Bestehende Zellanalysesysteme ....................................................... 12
2.4 Schlussfolgerung ............................................................................... 13
3. Vorbereitungen für die Biokompatibilitätsanalyse ......................................... 16
3.1 Probenaufarbeitung ........................................................................... 16
3.2 Zellkultivierung ................................................................................... 17
3.2.1 Vergleich der Färbemethoden ............................................ 19
4. Verwendete Hard- und Software .................................................................. 21
4.1 Visual Studio 2010 ............................................................................. 21
4.2 Halcon Bildverarbeitungsbibliothek .................................................... 21
4.3 Softwarearchitektur ............................................................................ 22
4.3.1 Dynamisches Einbinden von Programmmodulen ............... 22
4.3.2 Grundlegender Programmablauf ........................................ 24
4.4 Mikroskop und Kamera ...................................................................... 26
5. Segmentierung zytochemisch gefärbter Zellbereiche ................................... 27
5.1 Bildvorverarbeitung ........................................................................... 27
5.1.1 Reduktion von Bildartefakten durch Ausnutzung besonderer
Farbwertbeziehungen im RGB-Farbraum ........................... 27
5.1.2 Ergebnisse ......................................................................... 29
5.2 Untersuchung verschiedener Verfahren zur Segmentierung der
Zellbereiche ....................................................................................... 35
5.2.1 Schwellwertverfahren ............................................................... 35
5.2.1.1 Ergebnisse ............................................................................. 39

Inhaltsverzeichnis vi
5.2.2 Histogrammbasierte Methode ............................................ 41
5.2.2.1 Ergebnisse ............................................................................. 43
5.2.3 Histogram-Backprojection-Algorithmus .............................. 44
5.2.3.1 Ergebnisse ............................................................................. 46
5.3 Diskussion der Ergebnisse zur Segmentierung zytochemisch gefärbter
Zellbereiche ....................................................................................... 46
6. Separation agglomerierter Zellen ................................................................. 52
6.1 Kontextbasiertes Trennverfahren ....................................................... 54
6.1.1 Ergebnisse ......................................................................... 64
6.2 Klassifizierung segmentierter Zellbereiche ........................................ 66
6.2.1 Ergebnisse ......................................................................... 67
6.3 Zellkerndetektion ................................................................................ 68
6.3.1 Zellkerndetektion mit Hilfe eines Künstlich Neuronalen
Netzes (KNN) ...................................................................... 68
6.3.1.1 Datenerzeugung für das KNN ................................................ 69
6.3.1.2 Verbesserung der Klassifikationsleistung .............................. 70
6.3.1.3 Architektur und Lernparameter des KNN ............................... 71
6.3.1.4 Ergebnisse ............................................................................. 72
6.3.2 Zellkerndetektion auf Basis des Histogram-Backprojection-
Algorithmus ......................................................................... 73
6.3.2.1 Separation der Zellkernbereiche zur Optimierung der
Kerndetektion.................................................................... 77
6.3.2.2 Ergebnisse ............................................................................. 79
6.3.3 Diskussion der Ergebnisse ................................................. 80
6.4 Zellseparation in Clustern mit Verengungen an Zellkontaktstellen ..... 83
6.4.1 Lokalisierung der Verengungen an den Zellkontaktstellen . 83
6.4.2. Berechnung der kürzesten Pfade zwischen benachbarten
Zellkernbereichen ............................................................... 85
6.4.3 Generierung der Zellseparationslinien innerhalb der Cluster87
6.4.4 Ergebnisse ......................................................................... 89
6.5 Zellseparation in Clustern ohne Verengungen an Zellkontaktstellen . 91
6.5.1 Wachstumssimulation ........................................................ 91
6.5.2 Ergebnisse ......................................................................... 98
6.6 Zellseparation in Clustern mit Verengungen an Zellkontaktstellen ohne
Berücksichtigung der Zellkerne ......................................................... 99
6.6.1 Ergebnisse ....................................................................... 100
6.7 Randzellenbehandlung .................................................................... 101
6.7.1 Ergebnisse ....................................................................... 104
6.8 Diskussion der Ergebnisse zu den Separationsverfahren ................ 105
7. Automatisierung der Algorithmenauswahl .................................................. 112
7.1 Automatische Auswahl der geeigneten Zellsegmentierungsmethode113
7.2 Automatische Auswahl der geeigneten Zellseparationsmethode ..... 115

Inhaltsverzeichnis vii
8. Automatische Parametereinstellung ........................................................... 118
8.1 Automatische Parametereinstellung für die Zellsegmentierung ....... 118
8.2 Automatische Parametereinstellung für die CBS Methode .............. 123
8.2.1 Ergebnisse zur automatischen Parametereinstellung für die
CBS Methode ................................................................... 125
8.3 Diskussion der Ergebnisse zu den automatischen
Parametereinstellungen ................................................................... 126
9. Bewertung des Biokompatibilitätsgrades .................................................... 129
9.1 Quantitative Bewertung der Biokompatibilität................................... 129
9.2 Untersuchung verschiedener Stichprobenumfänge zur Berechnung der
durchschnittlichen Zellanzahl einer Probe ....................................... 132
9.3 Simulation unterschiedlich bioverträglicher Materialien ................... 136
9.4 Qualitative Bewertung der Biokompatibilität ..................................... 138
9.5 Diskussion der Ergebnisse zur Bewertung des
Biokompatibilitätsgrades .................................................................. 140
10. Ergebnisse und Diskussion ...................................................................... 142
10.1 Generalisierungsleistung der Software .......................................... 149
11. Zusammenfassung und Ausblick .............................................................. 155
12. Literaturverzeichnis .................................................................................. 158
13. Anhang ..................................................................................................... 165
13.1 Untersuchung zur Farbverteilung von Korrosionserscheinungen auf
dem Material Titan* ......................................................................... 165
13.2 Untersuchung zur Farbzusammensetzung der verwendeten
Materialien ....................................................................................... 168
13.3 Bewertung der in Kap. 5.2 vorgestellten Verfahren zur
Segmentierung von Zellen ............................................................... 171
13.4 Untersuchung zur Klassifizierung der Zellbereiche mit Hilfe
geometrischer Merkmale ................................................................. 176
13.5 Untersuchung zur Farbzusammensetzung der Kernbereiche und der
restlichen Zelle ................................................................................ 177
13.6 Untersuchung zur Zellzählung vor Anwendung der
Separationsverfahren ...................................................................... 181
13.7. Zellzählung mit dem Algorithmus aus Kap. 6.4 bei vorgeschalteter
CBS und ohne CBS ......................................................................... 183
13.8. Vergleich der automatischen Kernzählung auf Basis eines KNN zur
Referenz .......................................................................................... 184
13.9 Vergleich der automatischen Kernzählung auf Basis des HB-
Algorithmus mit der Referenzzählung .............................................. 185
13.10 Zellzählung mit der Wachstumssimulation im Vergleich zur
Referenzzählung ............................................................................. 189

Inhaltsverzeichnis viii
13.11 Zellzählung mit dem Algorithmus aus Kap. 6.6 und Vergleich mit der
Referenzzählung ............................................................................. 191
13.12 Bewertung der in Kap. 6 vorgestellten Verfahren zur Separation
zusammengewachsener Zellen ....................................................... 192
13.13 Bewertung der in Kap. 8 vorgestellten Methoden zur automatischen
Parametereinstellung ....................................................................... 194
13.14 Untersuchung verschiedener Stichproben der Probenoberfläche 197
13.15 Untersuchung zur manuellen Zellzählung in Suspension und auf der
Probenoberfläche ............................................................................ 198
13.16 Prüfzeiten zur Bestimmung der Proliferationsrate ........................ 200
13.17. Elimination von Ausreißern mit Hilfe des Grubbs-Tests .............. 201
Publikationsliste .............................................................................................. 204
Lebenslauf ...................................................................................................... 206
Eidesstattliche Erklärung ................................................................................ 208

Symbolverzeichnis ix
Symbolverzeichnis
L929: mouse connective tissue fibroblast established from the normal subcuta neous areolar and adipose tissue of a male C3H/An mouse MC3T3: MC3T3- E1 mouse embryo/fetus calvaria fibroblasts Titan*: Cp- Titan grade2, commercially pure Stahl*: rostfreier Stahl 14404 εB(Z): Erosion des Bildbereiches Z mit einem Strukturelement B Epithel: Bei diesem Begriff handelt es sich um eine Sammelbezeichnung für Deckgewebe und Drüsengewebe. Das Epithel ist neben Muskel-, Nerven- und Bindegewebe eine der vier Grundgewebearten. BHK-21: Baby Hamster Kidney – Nierenzellen vom Hamster L132: humane Lungenendothelzellen

Tabellenverzeichnis x
Tabellenverzeichnis
Tabelle 1: Kriterien für die in vitro-Biokompatibilität in Zell- und
Gewebekulkulturen (3) ............................................................................... 2
Tabelle 2: Durchzuführende Schleif- und Polierschritte .................................... 17
Tabelle 3: Materialspezifische Einwirkzeiten der Kristallviolett und May-
Grünwald Färbung ................................................................................... 19
Tabelle 4: Berechnung des Jaccard-Koeffizient für 25 ausgewählte Zellen mit
naheliegenden Artefakten, deren Segmentierungsergebnis durch die
Bildvorverarbeitung manipuliert wird. Als Segmentierungsmethode dient
das Schwellwertverfahren, beschrieben in Kap. 5.2.1 ............................. 31
Tabelle 5: Vergleich der Zellzählung mit der vorgestellten Bildvorverarbeitung
(1) und ohne Bildvorverarbeitung (2) zur Referenzzählung bei
Verwendung des Materials Stahl* und Titan*. Eingesetzt wird hier das
Segmentierungsverfahren, beschrieben in Kap. 5.2.1 und das
Separationsverfahren, beschrieben in Kap. 6.4 mit vorgeschalteter CBS
(Kap. 6.1); AZ: Automatische Zellzählung; FP: Falsch-positiv (zuviel)
Detektion; FN: Falsch-negativ (vergessen) Detektion; KD: Korrekte
Detektion (AZ-FP); REF: Referenzzählung; RFP: Relative Falsch-positiv
Detektion (FP / REF * 100); RFN: Relative falsch-negativ Detektion (FN /
REF * 100); ERR: Relativer Fehler (((REF – AZ) / REF) * 100) ............... 32
Tabelle 6: Vergleich der Zellzählung mit (1) der vorgestellten
Bildvorverarbeitung und ohne (2) Bildvorverarbeitung zur Referenzzählung
bei Verwendung des Materials Aluminium. Verwendung findet hier das
Segmentierungsverfahren, beschrieben in Kap. 5.2.1 und das
Separationsverfahren, beschrieben in Kap. 6.4; Zeichenerklärung siehe
Tabelle 5 .................................................................................................. 34
Tabelle 7: Schwellwerte zur Segmentierung der L929- und MC3T3-Zellbereiche
bei unterschiedlichen Materialien; SO,R: Oberer Schwellwert des
Rotkanals; SU,R: Unterer Schwellwert des Rotkanals; SO,G: Oberer
Schwellwert des Grünkanals; SU,G: Unterer Schwellwert des Grünkanals;
SO,B: Oberer Schwellwert des Blaukanals; SU,B: Unterer Schwellwert des
Blaukanals ............................................................................................... 36
Tabelle 8: Schwellwerte zur Segmentierung der Materialbereiche innerhalb der
bereits erfassten Zellbereiche; SO,R: Oberer Schwellwert des Rotkanals;
SU,R: Unterer Schwellwert des Rotkanals; SO,G: Oberer Schwellwert des
Grünkanals; SU,G: Unterer Schwellwert des Grünkanals; SO,B: Oberer
Schwellwert des Blaukanals; SU,B: Unterer Schwellwert des Blaukanals 38
Tabelle 9: Bewertung der Qualität des Schwellwertverfahrens für
unterschiedliche Zelllinien, Materialien und Mikroskopkontraste mit Hilfe
des Jaccard-Koeffizienten; A-HF: Auflicht- Hellfeld; A-DF: Auflicht-

Tabellenverzeichnis xi
Dunkelfeld; D-HF: Durchlicht- Hellfeld; D-PK: Durchlicht- Phasenkontrast
................................................................................................................. 40
Tabelle 10: Bewertung der Qualität des histogrammbasierten
Schwellwertverfahrens für unterschiedliche Zelllinien und
Mikroskopkontraste mit Hilfe des Jaccard-Koeffizienten; A-HF: Auflicht-
Hellfeld; A-DF: Auflicht-Dunkelfeld; D-HF: Durchlicht-Hellfeld; D-PK:
Durchlicht-Phasenkontrast ....................................................................... 43
Tabelle 11: Bewertung der Qualität des Histogram-Backprojection-Verfahrens
für unterschiedliche Zelllinien und Mikroskopkontraste mit Hilfe des
Jaccard-Koeffizienten; A-HF: Auflicht-Hellfeld; A-DF: Auflicht-Dunkelfeld;
D-HF: Durchlicht-Hellfeld; D-PK: Durchlicht-Phasenkontrast ................. 46
Tabelle 12: Benötigte Rechenzeit der in Kap. 5.2.1 bis Kap. 5.2.3 vorgestellten
Segmentierungsverfahren auf Basis eines Intel Core i7, 3.2GHz ............ 47
Tabelle 13: Vergleich der Jaccard-Koeffizienten der in Kap. 5.2.1 – 5.2.3
vorgestellten Zellsegmentierungsverfahren für die Zelltypen L929 und
MC3T3 bei unterschiedlichen Materialien und Mikroskopkontrasten; Die
grün hinterlegten Tabellenzellen zeigen das für den aktuell betrachteten
Mikroskopkontrast am besten geeignete Segmentierungsverfahren ....... 48
Tabelle 14: Statistische Daten zur automatischen Zellzählung mit dem
Segmentierungsverfahren aus Kap. 5.2.1 vor Anwendung der
Separationsmethoden .............................................................................. 51
Tabelle 15: Schematische Darstellung möglicher Objektgeometrien an einer
Kontaktstelle ............................................................................................ 61
Tabelle 16: Durchschnittliche relative Abweichung der automatischen Zählung
von 2376 L929-Zellen mit der Referenzzählung für unterschiedliche Werte
des Parameters r1 .................................................................................... 62
Tabelle 17: Durchschnittliche relative Abweichung der automatischen Zählung
von 2376 L929-Zellen mit der Referenzzählung für unterschiedliche Werte
des Parameters a ..................................................................................... 63
Tabelle 18: Durchschnittliche relative Abweichung der automatischen
Zellzählung von 2376 Zellen mit der Referenzzählung für unterschiedliche
Werte des Parameters x .......................................................................... 63
Tabelle 19: Durchschnittliche relative Abweichung der automatischen
Zellzählung von 2376 Zellen mit der Referenzzählung für unterschiedliche
Werte des Parameters y .......................................................................... 63
Tabelle 20: Vergleich der Rechenzeit für den Separationsalgorithmus (Kap. 6.4)
ohne CBS und mit CBS auf einer i7 3,2GHz CPU mit Parallelverarbeitung.
................................................................................................................. 64
Tabelle 21: Statistische Daten zur automatischen Zellzählung mit dem
Separationsverfahren aus Kap. 6.4 ohne und mit vorgeschalteter CBS .. 65
Tabelle 22: Vergleich der automatischen Klassifizierung der Zellbereiche in
Einzelzellen mit der Klassifizierung durch einen Experten; REF: Anzahl
klassifizierter Zellen durch einen Experten; FK: Anzahl falsch klassifizierter

Tabellenverzeichnis xii
Zellbereiche durch die Software; Relativer Fehler: Relativer Fehler der
Klassifikation durch die Software ............................................................. 67
Tabelle 23: Statistische Daten zur automatischen Kernzählung auf Basis eines
KNN ......................................................................................................... 73
Tabelle 24: Parametereinstellungen für die verwendeten Farbtemplates zur
Zellkerndetektion mittels HB-Algorithmus; ACMin: Minimal zulässige
Kernfläche; c: Anzahl der verwendeten Farben bei der Unterabtastung der
verwendeten 24bit-RGB Bilder (2563 Farben) zur Reduktion der
Rechenzeit, vgl. (55) ................................................................................ 75
Tabelle 25: Statistische Daten zur automatischen Kernzählung auf Basis des
HB-Algorithmus ........................................................................................ 79
Tabelle 26: Statistische Daten zur automatischen Kernzählung auf Basis des
HB-Algorithmus mit anschließender Kernseparation ............................... 79
Tabelle 27: Statistische Daten zur automatischen Zellzählung unter
Verwendung des in Kap. 6.4 beschriebenen Verfahrens ......................... 90
Tabelle 28: Statistische Daten zur automatischen Zellzählung unter
Verwendung des in Kap. 6.4 beschriebenen Verfahrens mit
vorgeschalteter CBS ................................................................................ 91
Tabelle 29: Vergleich der Flächenapproximation von 100 MC3T3 Zellen innerhalb
verschiedener Cluster bei unterschiedlichen maximalen
Wachstumsgeschwindigkeiten p .................................................................. 97
Tabelle 30: Durchschnittlicher Jaccard-Koeffizient für die Separation von 100
L929- und 100 MC3T3 Zellen mit Hilfe des in Kap. 6.5 vorgestellten
Separationsverfahrens ............................................................................. 98
Tabelle 31: Statistische Daten zur automatischen Zellzählung mit der
Wachstumssimulation .............................................................................. 99
Tabelle 32: Statistische Daten zur automatischen Zellzählung unter
Verwendung des in Kap. 6.6 beschriebenen Verfahrens ....................... 101
Tabelle 33: Vergleich der automatischen Zellzählung ohne
Randzellenbehandlung mit der Referenzzählung; AZ: Computergestützte
Zellzählung; REF: Referenzzellzählung durch einen Experten; ERR:
Relative Abweichung der automatischen Zellzählung von der
Referenzzählung .................................................................................... 104
Tabelle 34: Vergleich der Rechenzeiten zwischen den drei
Separationsverfahren für einen Probenausschnitt der Linie L929 auf einer
Intel Core i7 3,2GHz CPU ...................................................................... 106
Tabelle 35: Vergleich des Jaccard-Koeffizienten für 3 verschiedene
Separationsverfahren auf Basis von 200 Zellen (100 L929- und 100
MC3T3 Zellen) ....................................................................................... 107
Tabelle 36: Vergleich der durchschnittlichen Abweichungen der automatischen
Zellzählung zur Referenzzählung für die Separationsverfahren aus Kap.
6.4 – 6.6 für die Linien L929 und MC3T3 unter Berücksichtigung eines
Konfidenzintervalls von 99%. Untersucht wurden pro Zelllinie 100

Tabellenverzeichnis xiii
Probenausschnitte mit insgesamt ca. 20.000 L929 Zellen und ca. 10.000
MC3T3 Zellen ........................................................................................ 110
Tabelle 37: Schwellwerte zur Segmentierung der L929- und MC3T3-
Zellbereiche bei unterschiedlichen Materialien; S1O,R: Oberer Schwellwert
des Rotkanals; S1U,R: Unterer Schwellwert des Rotkanals; S1O,G: Oberer
Schwellwert des Grünkanals; S1U,G: Unterer Schwellwert des Grünkanals;
S1O,B: Oberer Schwellwert des Blaukanals; S1U,B: Unterer Schwellwert des
Blaukanals ............................................................................................. 118
Tabelle 38: Schwellwerte zur Segmentierung der Materialbereiche innerhalb der
bereits erfassten Zellbereiche; S2O,R: Oberer Schwellwert des Rotkanals;
S2U,R: Unterer Schwellwert des Rotkanals; S2O,G: Oberer Schwellwert des
Grünkanals; S2U,G: Unterer Schwellwert des Grünkanals; S2O,B: Oberer
Schwellwert des Blaukanals; S2U,B: Unterer Schwellwert des Blaukanals
............................................................................................................... 120
Tabelle 39: Schwellwerte zur Segmentierung der L929- und MC3T3-
Zellbereiche bei unterschiedlichen Materialien; S1O,R: Oberer Schwellwert
des Rotkanals; S1U,R: Unterer Schwellwert des Rotkanals; S1O,G: Oberer
Schwellwert des Grünkanals; S1U,G: Unterer Schwellwert des Grünkanals;
S1O,B: Oberer Schwellwert des Blaukanals; S1U,B: Unterer Schwellwert des
Blaukanals ............................................................................................. 121
Tabelle 40: Automatische Parameterfestlegung des genetischen Algorithmus
10-mal hintereinander ausgeführt ............................................... 126
Tabelle 41: Bewertung der Qualität des Schwellwertverfahrens mit manueller
und automatischer Parameterfestlegung für unterschiedliche Zelllinien,
Materialien und Mikroskopkontraste mit Hilfe des Jaccard-Koeffizienten; A-
HF: Auflicht-Hellfeld; A-DF: Auflicht-Dunkelfeld; D-HF: Durchlicht-Hellfeld;
D-PK: Durchlicht-Phasenkontrast .......................................................... 126
Tabelle 42: Zytotoxische Einteilung der Prüfmaterialien laut DIN ISO Norm
10993-5 (5) ............................................................................................ 129
Tabelle 43: Zuordnung der Zellproliferation zu den vier Stufen der Zytotoxizität
............................................................................................................... 132
Tabelle 44: Verschiedene Stichprobenumfänge mit der jeweiligen Abweichung
zum Referenzwert. Für jeden Stichprobenumfang wurden 10 Versuche
durchgeführt. Zeichenerklärung: Abw.: Die maximale positive und negative
Abweichung der durchschnittlichen Zellanzahl vom Referenzwert [%] .. 133
Tabelle 45: Anpassung der Zellproliferationsgrenzen für die Einordnung in die
Zytotoxizitätsstufe „nicht zytotoxisch“ unter Berücksichtigung der
Gesamtabweichung bei Verwendung einer Stichprobe mit 15 Bildern und
bei Verwendung der vollständigen Probe für die Zellzählung ................ 136
Tabelle 46: Formaldehyd-Verdünnungsreihe mit einer Konzentration von 0,0
µg/ml bis 0,5 µg/ml in 0,1 µg/ml Schritten für die Linie L929 und bis 1,0
µg/ml für MC3T3 Zellen auf dem Material Polystyrol. Abhängig von der

Tabellenverzeichnis xiv
Proliferationsrate erfolgt eine Interpretation des zytotoxischen Zustandes
der Zellen ............................................................................................... 137
Tabelle 47: Auswertung der morphologischen Merkmale für eine Formaldehyd-
Verdünnungsreihe mit einer Konzentration von 0,0 µg/ml bis 0,5 µg/ml in
0,1 µg/ml Schritten für die Linie L929 und bis 1,0 µg/ml für MC3T3 Zellen
auf dem Material Polystyrol. Bei den in der Tabelle angegebenen Daten
handelt sich jeweils um Durchschnittswerte aus 15 verschiedenen
Probenausschnitten; ∆K: Relative Abweichung zur Kompaktheit bei einer
Formaldehydkonzentration von 0,0 µg/ml .............................................. 139
Tabelle 48: Übersicht der statistischen Daten zur Genauigkeit der Zellzählung
der verschiedenen Separationsverfahren .............................................. 146
Tabelle 49: Abweichung des Rot- und Blaukanals zum Grünkanal für korrodierte
Artefakte ................................................................................................ 166
Tabelle 50: Abweichung des Rot- und Blaukanals zum Grünkanal für den
Hintergrundbereich ohne korrodierte Artefakte ...................................... 167
Tabelle 51: Farbzusammensetzung des Materials Titan* bei May-Grünwald-
Färbung unter Verwendung eines Auflichtmikroskopes im Hellfeldkontrast
............................................................................................................... 168
Tabelle 52: Farbzusammensetzung des Materials Stahl* bei May-Grünwald-
Färbung unter Verwendung eines Auflichtmikroskopes im Hellfeldkontrast
............................................................................................................... 169
Tabelle 53: Farbzusammensetzung des Materials Polystyrol bei May-Grünwald-
Färbung unter Verwendung eines Durchlichtmikroskopes im
Hellfeldkontrast ...................................................................................... 170
Tabelle 54: Bewertung der Segmentierungsleistung der Verfahren TF:
Schwellwertverfahren, Kap. 5.2.2; TB: Histogrammbasierte Methode, Kap.
5.2.3; HB: Histogram-Backprojection-Algorithmus Kap. 5.2.4 für die
verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-DF:
Auflicht-Dunkelfeld. Die L929-Zellen befinden sich auf dem Substrat Stahl*
und Titan*, daher kommt das Durchlichtmikroskop nicht zum Einsatz ... 172
Tabelle 55: Zeichenerklärung siehe Tabelle 46. Die L929-Zellen befinden sich
auf dem transparenten Substrat Polystyrol, daher kommt das
Auflichtmikroskop nicht zum Einsatz ...................................................... 173
Tabelle 56: Zeichenerklärung siehe Tabelle 46. Die MC3T3-Zellen befinden sich
auf dem Substrat Stahl* und Titan*, daher kommt das Durchlichtmikroskop
nicht zum Einsatz ................................................................................... 174
Tabelle 57: Zeichenerklärung siehe Tabelle 46. Die MC3T3-Zellen befinden sich
auf dem transparenten Substrat Polystyrol, daher kommt das
Auflichtmikroskop nicht zum Einsatz ...................................................... 175
Tabelle 58: Histogrammschwerpunkte Rs, Gs und Bs des Rot- Grün- und
Blaukanals für 50 verschiedene MC3T3 Zellen...................................... 179
Tabelle 59: Histogrammschwerpunkte Rs, Gs und Bs des Rot- Grün- und
Blaukanals für 50 verschiedene L929 Zellen ......................................... 180

Tabellenverzeichnis xv
Tabelle 60: Vergleich der automatischen L929-Zellzählung mit dem
Segmentierungsverfahren aus Kap. 5.2.1 vor Anwendung der
Separationsmethoden im Vergleich zur Referenzzählung; AZ:
Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte
Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR:
Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100) ............. 181
Tabelle 61: Vergleich der automatischen MC3T3-Zellzählung mit dem
Segmentierungsverfahren aus Kap. 5.2.1 vor Anwendung der
Separationsmethoden im Vergleich zur Referenzzählung; AZ:
Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte
Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR:
Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100) ............. 182
Tabelle 62: Vergleich der automatischen L929-Zellzählung mit dem Algorithmus
aus Kap. 6.4 bei vorgeschalter CBS (1) und ohne CBS (2); AZ:
Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte
Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR:
Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100) ............. 183
Tabelle 63: Vergleich der automatischen Kernzählung auf Basis eines KNN mit
der Referenzzählung; AZ: Automatische Zellzählung; REF:
Referenzzählung; REF Norm: Normierte Referenzzählung; AZ Norm:
Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm
– AZ Norm) / REF Norm) * 100) ............................................................. 184
Tabelle 64: Vergleich der automatischen L929-Kernzählung auf Basis des HB-
Algorithmus mit der Referenzzählung; AZ: Automatische Zellzählung;
REF: Referenzzählung; REF Norm: Normierte Referenzzählung; AZ Norm:
Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm
– AZ Norm) / REF Norm) * 100) ............................................................. 185
Tabelle 65: Vergleich der automatischen MC3T3-Kernzählung auf Basis des
HB-Algorithmus mit der Referenzzählung; AZ: Automatische Zellzählung;
REF: Referenzzählung; REF Norm: Normierte Referenzzählung; AZ Norm:
Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm
– AZ Norm) / REF Norm) * 100). ............................................................ 186
Tabelle 66: Automatische L929-Kernzählung auf Basis des HB-Algorithmus mit
anschließender Kernseparation im Vergleich zur Referenzzählung; AZ:
Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte
Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR:
Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100) ............. 187
Tabelle 67: Automatische MC3T3-Kernzählung auf Basis des HB-Algorithmus
mit anschließender Kernseparation im Vergleich zur Referenzzählung; AZ:
Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte
Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR:
Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100) ............. 188

Tabellenverzeichnis xvi
Tabelle 68: Vergleich der L929-Zellzählung (1) sowie der MC3T3-Zellzählung
(2) mit der Wachstumssimulation zur Referenzzählung; AZ 1:
Automatische L929-Zellzählung; AZ 2: Automatische MC3T3-Zellzählung;
REF: Referenzzählung; REF Norm: Normierte Referenzzählung; AZ 1
Norm: Normierte automatische L929-Zellzählung; AZ 2 Norm: Normierte
automatische MC3T3-Zellzählung; ERR 1: Relativer Fehler für den Typ
L929 (((REF Norm – AZ Norm) / REF Norm) * 100); ERR 2: Relativer
Fehler für den Typ MC3T3 ..................................................................... 189
Tabelle 69: L929-Zellzählung auf Basis des Algorithmus in Kap. 6.6 im
Vergleich zur Referenzzählung; AZ: Automatische Zellzählung; REF:
Referenzzählung; REF Norm: Normierte Referenzzählung; AZ Norm:
Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm
– AZ Norm) / REF Norm) * 100) ............................................................. 191
Tabelle 70: Berechnung des Jaccard-Koeffizienten für 100 separierte L929-
Zellen aus 10 verschiedenen Probenausschnitten. Verfahren A: Kap. 6.4;
Verfahren B: Kap. 6.5; Verfahren C: Kap. 6.6 ........................................ 192
Tabelle 71: Berechnung des Jaccard-Koeffizienten für 100 separierte MC3T3-
Zellen aus 10 verschiedenen Probenausschnitten. Verfahren A: Kap. 6.4;
Verfahren B: Kap. 6.5; Verfahren C: Kap. 6.6 ........................................ 193
Tabelle 72: Bewertung der Segmentierungsleistung des in Kap. 8.1
beschriebenen Verfahrens für die verschiedenen Mikroskopkontraste A-
HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld. Die L929-Zellen
befinden sich auf dem Substrat Stahl* und Titan*, daher kommt das
Durchlichtmikroskop nicht zum Einsatz .................................................. 194
Tabelle 73: Bewertung der Segmentierungsleistung des in Kap. 8.1
beschriebenen Verfahrens für die verschiedenen Mikroskopkontraste A-
HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld. Die L929-Zellen
befinden sich auf dem Substrat Polystyrol, daher kommt das
Auflichtmikroskop nicht zum Einsatz ..................................................... 195
Tabelle 74: Bewertung der Segmentierungsleistung des in Kap. 8.1
beschriebenen Verfahrens für die verschiedenen Mikroskopkontraste A-
HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld. Die MC3T3-Zellen
befinden sich auf dem Substrat Stahl* und Titan*, daher kommt das
Durchlichtmikroskop nicht zum Einsatz .................................................. 195
Tabelle 75: Bewertung der Segmentierungsleistung des in Kap. 8.1
beschriebenen Verfahrens für die verschiedenen Mikroskopkontraste A-
HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld. Die MC3T3-Zellen
befinden sich auf dem Substrat Polystyrol, daher kommt das
Auflichtmikroskop nicht zum Einsatz ...................................................... 196
Tabelle 76: Verschiedene Stichprobenumfänge mit der jeweiligen Abweichung
zum Referenzwert. Für jeden Stichprobenumfang wurden 10 Versuche
durchgeführt. Die zufälligen Stichproben umfassen 50, 30 und 10 Bilder,
die gezielt ausgewählten Stichproben hingegen 15, 10 und 5 Bilder.

Tabellenverzeichnis xvii
Zeichenerklärung: Ø: Durchschnittliche Zellanzahl der Stichprobe; σ:
Standardabweichung der Stichprobe; ∆REF: relative Abweichung der
durchschnittlichen Zellanzahl der Stichprobe zum Referenzwert; Abw.: Die
maximale positive und negative Abweichung der durchschnittlichen
Zellzahl vom Referenzwert [%] .............................................................. 197
Tabelle 77: Manuell ermittelte Zellanzahl für 6 Proben in Suspension. Pro Probe
wurden 10 Zählungen durchgeführt und der Mittelwert gebildet ............ 198
Tabelle 78: Manuell ermittelte Zellanzahl auf der Oberfläche von 3 Proben .. 199
Tabelle 79: Manuell ermittelte Zellanzahlen für 6 Proben in Suspension
(Versuchsreihe A) und 3 Proben unter dem Mikroskop, zunächst manuell
(Versuchsreihe B) und dann computergestützt gezählt (Versuchsreihe C).
Für jede Probe in Suspension wurden 10 Zählungen eines bestimmten
Volumens durchgeführt und auf das Gesamtvolumen hochgerechnet. Aus
den 10 Messungen wird schließlich der Mittelwert gebildet. .................. 200
Tabelle 80: Prüfzeiten zur Bestimmung der Proliferationsrate. Es werden 4
verschiedene Methoden der Prüfung vorgestellt. A: Manuelle Zellzählung
mit Hilfe einer Fuchs-Rosenthal-Zählkammer; B: Computergestützte
Zellzählung anhand einer Stichprobe mit einem Umfang von 15
Probenausschnitten; C: Computergestützte Zellzählung der vollständigen
Probe; D: Computergestützte Zellzählung ebenfalls von der vollständigen
Probe, wobei hier die Probe im Gegensatz zur Methode C automatisch
durch ein an das Mikroskop installiertes Achssystem abgescannt wird. Die
Rechenzeiten in der Spalte „Automatische Zellzählung“ sind in Klammern
gesetzt, da es sich hier ausschließlich um automatisierte Arbeitsschritte
handelt ................................................................................................... 200
Tabelle 81: Testwerte des Grubbs-Tests für die Versuchsreihen A, B und C zur
Identifikation von Ausreißern. Als Signifikanzniveau wurde α = 5%
gewählt. Aus der Grubbs-Tabelle (84) erhält man für n = 6 Messwerte und
α=5% einen Schwellwert SA = 1,82. Für n = 3 Messwerte und ebenfalls
α=5% resultiert ein Schwellwert SB = SC = 1,15. .................................... 202

Abbildungsverzeichnis xviii
Abbildungsverzeichnis
Abbildung 1: Zellmorphologische Änderung bei Abnahme der Biokompatibilität
(3) .............................................................................................................. 2
Abbildung 2: Links: Agglomerierte L929 Zellen auf dem Material Stahl*. Sie
bilden oft Verengungen an den Zellkontaktstellen. Rechts: Kompakt
zusammenhängende Zellen vom Typ MC3T3 auf dem Material Titan*. Die
Bilder wurden im Biotechnologielabor der Fachhochschule Südwestfalen
aufgenommen ............................................................................................ 4
Abbildung 3: Zellkernfärbung mit DRAQ5
Abbildung 4: Zellfärbung mit May- Grünwald...................................................... 6
Abbildung 5: Ablauf bei der Aufarbeitung der Proben ...................................... 16
Abbildung 6: Kristallviolett gefärbte MC3T3-Zellen ........................................... 19
Abbildung 7: May-Grünwald gefärbte MC3T3-Zellen ....................................... 19
Abbildung 8: Architektur der Halcon Bibliothek (Quelle: MVTec GmbH) .......... 22
Abbildung 9: Programmablauf zur dynamischen Einbindung von
Programmmodulen .................................................................................. 23
Abbildung 10: Aktivitätsdiagramm zum allgemeinen Programmablauf bei der
Zellanalyse ............................................................................................... 25
Abbildung 11: L929 Zellen auf dem Substrat Titan mit einigen Bildartefakten.
Der vergrößerte Ausschnitt zeigt eine Zelle im Teilungsprozess mit
benachbarter Störquelle q. Diese Störquelle wird als Teil der Zelle erfasst
(grüne Kontur). ......................................................................................... 28
Abbildung 12: Links: Segmentierter Zellbereich ohne Bildvorverarbeitung;
Rechts: Segmentierter Zellbereich mit Bildvorverarbeitung .................... 29
Abbildung 13: a: Zwei Objekte liegen übereinander. Der Jaccard-Koeffizient
beträgt 1; b: Zwei Objekte überlappen sich teilweise. Der Jaccard-
Koeffizient beträgt 0,33; c: Zwei Objekte besitzen keine gemeinsame
Schnittmenge. Der Jaccard-Koeffizient ist 0 ............................................ 30
Abbildung 14: MC3T3-Zellen auf dem Substrat Aluminium. Es liegen starke
Korrosionserscheinungen im Bild vor ....................................................... 33
Abbildung 15: Ergebnis der Bildvorverarbeitungsmethode nach Anwendung auf
den in Abb. 14 dargestellten MC3T3-Probenausschnitt ........................... 34
Abbildung 16: Verschiedene Zellbereiche ........................................................ 36
Abbildung 17: Links: Zellcluster ohne Detektion der eingeschlossenen Titan*-
Bereiche. Rechts: Die im Zellbereich erfassten Titan*-Flächen (orange
markiert) ................................................................................................... 37
Abbildung 18: Schematischer Ablauf der Zellsegmentierung mit Hilfe der
Schwellwertmethode für alle Farbkanäle eines RGB-Bildes .................... 39
Abbildung 19: MC3T3 Zellen auf dem Material Stahl unter Verwendung des
Hellfeldkontrastes beim Durchlichtmikroskop. Die große Zelle mittig im Bild

Abbildungsverzeichnis xix
weist einen teilweise sehr schwachen Kontrastunterschied zum Substrat
auf. Dies beeinflusst die Segmentierungsgenauigkeit. ............................. 40
Abbildung 20: Ablaufdiagramm zur automatischen Bestimmung der
Schwellwertgrenzen SO und SU ................................................................ 42
Abbildung 21: Ablauf der Zellsegmentierung nach der histogrammbasierten
Methode ................................................................................................... 43
Abbildung 22: L929-Zellen auf dem Substrat Stahl* bei 100-facher
Vergrößerung ........................................................................................... 44
Abbildung 23: Farbtemplate für den HB Algorithmus ....................................... 45
Abbildung 24: Ergebnis des HB mit Hilfe des Farbtemplates aus Abb. 22 ....... 45
Abbildung 25: Links: Hellfeldbild von ungefärbten Hefezellen (57); Rechts:
Gradientenbild von Abb. 24 Links ............................................................ 49
Abbildung 26: Zellen des Typs L929 auf dem Material Stahl*. Vereinzelnd
können Agglomerate (grüne Kontur) auftreten ......................................... 52
Abbildung 27: Zellen des Typs MC3T3 auf dem Material Stahl*. Diese Zelllinie
neigt zur Clusterbildung (grüne Kontur) ................................................... 52
Abbildung 28: Ablaufdiagramm zur Zellvereinzelung. Nähere Erläuterungen
siehe Text ................................................................................................ 53
Abbildung 29: Links: Die Kontur des Objektes RC wird mit einem kreisförmigen
Strukturelement C1 abgefahren. Mitte: Der in C1 liegende Hintergrund B
wird durch RA,n in zwei Teilmengen B1 und B2 aufgeteilt. Rechts: Mit Hilfe
von C1 und CR wird geprüft, ob eine Verengung vorliegt .......................... 55
Abbildung 30: Ergebnis der Objekttrennung nach 3 Separationsvorgängen. Es
resultieren 3 neue Objektbereiche RS,1, RS,2, RS,3. Die Objektbereiche RA,2
und RA,3 sind zufällig dem separierten Objekt RS,3 und der Objektbereich
RA,1 des getrennten Objektes RS,1 zugeordnet worden ............................ 56
Abbildung 31: Unregelmäßigkeiten bei einer Objektkontur die dazu führen dass
1BR und 2BR deutlich unterschiedliche Flächenanteile besitzen. ........... 57
Abbildung 32: Programmablaufplan zur Auftrennung eines Objektes RC an
seinen n Engpässen. Erläuterung im Text. .............................................. 58
Abbildung 33: Programmablaufplan zur CBS ................................................... 59
Abbildung 34: Links: 2 Objekte; Mitte: Zwei verbundene Objekte; Rechts: Zwei
verbundene Objekte, wobei ein Objektbereich in C1 hineinragt, der zum
linken Objekt gehört. ................................................................................ 61
Abbildung 35: Links: Die Abtrennung eines Zellausläufers von seiner Zelle ist
nicht erwünscht; Rechts: Die Aufspaltung eines Zellausläufers ist ebenfalls
unerwünscht............................................................................................. 64
Abbildung 36: Die cyan gefärbte Kontur repräsentiert den ursprünglichen
Zellbereich Zi. Die grün gefärbten Konturen repräsentieren Zi,13 (das
Ergebnis nach 13 Iterationen der Erosion). .............................................. 67
Abbildung 37: Durchlauf des Zellbereiches mit einem Fenster der Größe 20s20
Pixel zur Ermittlung der vier Inputinformationen für das KNN .................. 70

Abbildungsverzeichnis xx
Abbildung 38: Links: Prüfbereich zur Klassifizierung des eingeschlossenen
Zellareals; Rechts: Aufteilung des Prüfbereiches bei nicht eindeutiger
Klassifizierung .......................................................................................... 71
Abbildung 39: Ausgewählte Netzarchitektur für die Klassifizierung der
verschiedenen Zellbestandteile................................................................ 72
Abbildung 40: Trainings- und Validierungsfehler in Abhängigkeit der Lernzyklen
................................................................................................................. 72
Abbildung 41: MC3T3-Zellen auf dem Substrat Titan* bei 100-facher
Vergrößerung mit vergrößerten Bildausschnitten verschiedenfarbiger
Kernbereiche............................................................................................ 74
Abbildung 42: L929-Zellen auf dem Substrat Stahl* bei 100-facher
Vergrößerung mit vergrößerten Bildausschnitten verschiedenfarbiger
Kernbereiche............................................................................................ 74
Abbildung 43: Vergrößerte Darstellung der erstellten Farbtemplates für die
Detektion der Zellkernbereiche mittels HB-Algorithmus. Links: Template 1
zur Detektion dunkler Kernbereiche; Rechts: Template 2 zur Detektion
vergleichsweise heller Kernbereiche ........................................................ 74
Abbildung 44: a: Segmentierter Zellbereich (grüne Kontur); b: Ergebnisbild E1
des HB-Algorithmus mit Template 1; c: Ergebnisbild E2 des HB-
Algorithmus mit Template 2; d: Ergebnis der Zellkerndetektion nach der
Weiterverarbeitung von c. Bei den schwarz eingekreisten Kernbereichen
handelt es sich jeweils um zwei Kerne, die jedoch nur als ein Kernbereich
erkannt wurden ........................................................................................ 76
Abbildung 45: Ergebnis der Markergenerierung für MC3T3-Kernbereiche, die
Kontur der ursprünglichen Kernbereiche ist gelb, die generierten Marker
grün dargestellt ........................................................................................ 77
Abbildung 46: Flussdiagramm zum schematischen Ablauf der
Kernbereichdetektion mit Hilfe des HB Algorithmus ................................. 78
Abbildung 47: a: Zellcluster vom Typ MC3T3; b: Kerndetektion mit Hilfe des
Histogram Backprojection Algorithmus und anschließendem Closing. Nicht
korrekt detektierte Kernbereiche sind durch rote Pfeile gekennzeichnet; c:
Inverse Distanztransformation von b; d: Wasserscheidentransformation
angewendet auf c (Schwellwert: 10). Übersegmentierte Bereiche sind
durch Pfeile gekennzeichnet; e: Inverse Distanztransformation von b; d:
Wasserscheidentransformation angewendet auf c (Schwellwert: 15). Nicht
alle Kernbereiche wurden korrekt separiert, gekennzeichnet durch die
Pfeile; f: Segmentierte Kernbereiche mit der Methode beschrieben in Kap.
6.3.2 ......................................................................................................... 82
Abbildung 48: Links: Segmentierter L929-Cluster (blaue Kontur); Rechts:
Ermittelte DCPs (blaue Kreisstrukturen) für den im Bild segmentierten
Cluster (orange Kontur). Der rote Pfeil kennzeichnet einen für die
Separation ungeeigneten DCP ................................................................ 84

Abbildungsverzeichnis xxi
Abbildung 49: Definition eines Konturpixels als lokaler maximaler Abstand zum
Rand. Zunächst liegen n Konturpunkte vor dem Maximum, deren
Abstände sich zur linken Seite des umschließenden Rechtecks
vergrößern. Nach dem Maximum folgen n Konturpunkte, deren Abstände
sich zur linken Rechteckseite verkleinern ................................................ 84
Abbildung 50: Mit dem A*-Algorithmus berechneter Pfad (weiß gefärbt) durch
die Zellkernschwerpunkte (schwarze Kreuze) im Cluster ........................ 85
Abbildung 51: Ausgangssituation für die Berechnung des kürzesten Pfades von
der Startposition (grünes Quadrat) zur Zielposition (rotes Quadrat) unter
Berücksichtigung von nicht passierbaren Hindernissen (blaue Quadrate).
Für jedes Nachbarfeld werden die Kosten F, G und H angegeben. ......... 86
Abbildung 52: Berechneter Pfad vom Ausgangspunkt (grünes Quadrat) zum
Zielpunkt (rotes Quadrat) unter Berücksichtigung nicht passierbarer
Hindernisse (blaue Quadrate). Die zu dem kürzesten Pfad gehörenden
Felder sind durch einen roten Kreis gekennzeichnet. .............................. 87
Abbildung 53: Separation zweier zusammengewachsener Zellen.
Zeichenerklärung siehe Text .................................................................... 87
Abbildung 54: Flussdiagramm zum schematischen Ablauf der Zellseparation mit
der in Kap. 6.4 beschriebenen Methode .................................................. 89
Abbildung 55: L929 Zellen auf dem Material Stahl*. Bei den gelb markierten
Bereichen handelt es sich um Zellen, die sich gerade in einem
Teilungsprozess befinden ........................................................................ 90
Abbildung 56: Zwei Objektcluster mit jeweils unterschiedlicher Anzahl an
Objektmerkmalen im Bild ......................................................................... 92
Abbildung 57: Links: Die Objektmerkmale C1,1 und C1,2 befinden sich im
Wachstumsprozess. Der Parameter p ist auf p = 2 gesetzt. Bei
Verwendung von Gl. 21 resultiert p1,1 = 2 und p1,2 = 1. Rechts: Zweite
Iteration des Wachstums. Die weiße Linie repräsentiert die Grenze der
beiden Objekte, nachdem kein Wachstum mehr möglich ist. ................... 93
Abbildung 58: Schematischer Ablauf des Wachstumsprozesses. Die in Li
enthaltenen Objektmerkmale werden durch numerische Werte für jeden
Iterationsschritt repräsentiert. Die Pfeile zeigen auf das nach dem
Wachstumsprozess neu entstandene Objektmerkmal. Weiterführende
Erklärungen sind im nachfolgenden Text enthalten ................................. 95
Abbildung 59: a: Zusammengewachsene MC3T3-Zellen auf dem Material Titan;
b: Ergebnis des HB-Algorithmus zur Kerndetektion; c: Der segmentierte
Zellbereich ist grün umrandet, die detektierten Kernbereiche schwarz
umrandet. Teilweise wurden zwei Kerne als ein Bereich segmentiert; d:
Detektierte Kernbereiche mit starken Verengungen wurden aufgetrennt,
da es sich wahrscheinlich um zwei unterschiedliche Kernbereiche handelt.
................................................................................................................. 96
Abbildung 60: Die Kernbereichflächen (V), aufgetragen über die
Gesamtzellflächen (U) ............................................................................. 97

Abbildungsverzeichnis xxii
Abbildung 61: Ergebnis der Wachstumssimulation. Die Zellgrenzen sind grün
und die Kernbereiche schwarz dargestellt. .............................................. 98
Abbildung 62: MC3T3 Zellen auf dem Material Stahl*; Zellen, die sich nur
teilweise im Bild befinden (rot umrandet), müssen eliminiert werden ..... 101
Abbildung 63: Schematischer Probenausschnitt; Generierung einer
Rechteckkontur (schwarze Kontur) als Hilfsmittel zur Bestimmung der
Randzellen ............................................................................................. 102
Abbildung 64: Ablaufdiagramm zur Randzellenbehandlung nach der
Zellsegmentierung ................................................................................. 103
Abbildung 65: Allgemeine Vorgehensweise bei der Bildanalyse .................... 112
Abbildung 66: Punktdiagramm zur Standardabweichung der Grauwerte für 100
Bilder, aufgenommen am Auflichtmikroskop (blau) und 100 Bilder,
aufgenommen am Durchlichtmikroskop( rot) ......................................... 113
Abbildung 67: Dichtefunktionen der Standardabweichungen von Graustufen in
100 Auflichtbildern (blau) und 100 Durchlichtbildern (rot) ...................... 114
Abbildung 68: Anzahl der Zellkerne pro Cluster im Vergleich für die Zelltypen
L929 und MC3T3 ................................................................................... 115
Abbildung 69: Dichtefunktion des Merkmals KA für den Zelltyp L929 ............ 116
Abbildung 70: Dichtefunktion des Merkmals KA für den Zelltyp MC3T3 ........ 116
Abbildung 71: Histogramm des Rotkanals eines Zellbildes. Die Zellen sind vom
Typ L929, May-Grünwald gefärbt und auf dem Material Stahl aufgebracht.
Das globale Maximum liegt auf dem Index 229 mit 161.299 Pixeln ....... 119
Abbildung 72: Flussdiagramm zum Ablauf der automatischen
Parametereinstellung ............................................................................. 121
Abbildung 73: Zwei Histogramme von Bildern mit Zellen jeweils vom Typ L929,
May-Grünwald gefärbt, aufgenommen am Auflichtmikroskop. Das zweite
Bild ist etwas heller, es besitzt ein leicht nach rechts verschobenes
Histogramm (unten) ............................................................................... 122
Abbildung 74: Beispielhafte fitnessproportionale Aufteilung von 10 Individuen
auf einer Scheibe. Ein Zufallswert im Bereich 1 bis 360 (schwarzer Pfeil)
wählt ein Individuum für die Reproduktion aus....................................... 124
Abbildung 75: Vergleich der Arbeitsschritte der herkömmlichen
Zellzahlbestimmung mit Hilfe von Zählkammern (Verfahren A) mit der
computergestützten Auswertung der Zellbilder (Verfahren B). Erklärung
siehe Text. ............................................................................................. 130
Abbildung 76: Suspensionsmethode zum Aufbringen der Zellen auf die
Probenoberfläche ................................................................................... 131
Abbildung 77: Links: L929-Zellen auf dem Material Stahl mit vergleichsweise
schwacher Zelldichte. Rechts: Deutlich höhere Zelldichte auf derselben
Probe an einer anderen Position ............................................................ 133
Abbildung 78: Zellanzahl für 96 verschiedene Ausschnitte einer Probe (blaue
Raute). Die durchschnittliche Zellanzahl aller Probenausschnitte beträgt
477,23 Zellen (rotes Quadrat) ................................................................ 134

Abbildungsverzeichnis xxiii
Abbildung 79: Schematische Darstellung der Vorgehensweise bei den
Bildaufnahmen der vollständigen Probe. Die Aufnahmen beginnen am
rechten unteren und enden am linken oberen Probenbereich ............... 135
Abbildung 80: Morphologieänderung von L929-Zellen bei unterschiedlichen
Formaldehyd-Konzentrationen. Die unteren Strahlen besitzen einen
farblichen Verlauf von grün (bioverträglich) nach rot (biounverträglich). Der
obere Strahl zeigt die Bioverträglichkeit für den Zelltyp MC3T3 und der
untere Strahl für den Typ L929. Die gestrichelte Linie zeigt ab welcher
Formaldehydkonzentration eine stark zytotoxische Wirkung auf die Zellen
vorliegt ................................................................................................... 141
Abbildung 81: Prüfzeiten zur Bestimmung der Proliferationsrate. Es werden 4
verschiedene Methoden der Prüfung vorgestellt. Methode A: Manuelle
Zellzählung mit Hilfe einer Fuchs-Rosenthal-Zählkammer; Methode B:
Computergestützte Zellzählung anhand einer Stichprobe mit einem
Umfang von 15 Probenausschnitten; Methode C: Computergestützte
Zellzählung der vollständigen Probe; Methode D: Computergestützte
Zellzählung ebenfalls von der vollständigen Probe, wobei hier die Probe
im Gegensatz zu den Methoden B und C automatisch durch ein an das
Mikroskop installiertes Achssystem abgerastert wird. ............................ 142
Abbildung 82: Versuchsreihe A: Durch einen Experten ermittelte
durchschnittliche Zellanzahl von jeweils 6 Proben (A1 bis A6) des gleichen
Materials mit Hilfe einer Fuchs-Rosenthal-Zählkammer. ....................... 144
Abbildung 83: Vergleich der computergestützten vollständigen Zellzählung von
3 Proben (C1 bis C3) mit einer durch einen Experten durchgeführten
Referenzzählung (B1 bis B3). ................................................................ 144
Abbildung 84: Links: Durchschnittliche Zellanzahl der Versuchsreihen A-C vor
Ausschluß der Ausreißer; Rechts: Durchschnittliche Zellanzahl der
Versuchsreihen A-C nach Ausschluss der mit Hilfe des Grubbs-Tests
ermittelten Ausreißer .............................................................................. 145
Abbildung 85: Visualisierung der in Tabelle 48 hinterlegten statistischen Daten
zur Zellzählung der verschiedenen Separationsverfahren. Auf der x-Achse
sind jeweils für die 4 im Vergleich stehenden Verfahren die
Konfidenzintervalle 95%, 99%, 99,9% und 99,99% aufgetragen. Der y-
Wert repräsentiert die Abweichung zur Referenzzählung. Die farbige
Markierung innerhalb der jeweiligen Toleranzbereiche repräsentiert den
Mittelwert. .............................................................................................. 147
Abbildung 87: Links: Segmentierungsergebnis von kristallviolett gefärbten
MC3T3-Zellen; Rechts: Segmentierungsergebnis von May-Grünwald
gefärbten MC3T3-Zellen ........................................................................ 151
Abbildung 88: Links: Segmentierungsergebnis von zweifach
fluoreszenzgefärbten MC3T3 Zellen. Die Zellkerne sind mit dem
Fluoreszenzfarbstoff DAPI und der restliche Zellbereich mit dem
Fluoreszenzfarbstoff Phalloidin angefärbt. Rechts:

Abbildungsverzeichnis xxiv
Segmentierungsergebnis von DAPI-fluoreszenzgefärbten Kernbereichen
humaner Hautzellen ............................................................................... 151
Abbildung 89: Links: Segmentierung von BHK-21 Zellen; Rechts:
Segmentierung von L132 Zellen. Zellflächen weisen eine blaue und
Kernbereiche eine grüne Kontur auf ...................................................... 152
Abbildung 90: Segmentierung von in Suspension befindlichen L929 Zellen. Die
Suspension befindet sich in einer Zählkammer, daher ist das Raster im
Bild sichtbar. .......................................................................................... 153
Abbildung 91: Links: Kalibrierung der Software mit Hilfe eines
Referenzobjektes, hier der bei 10mm eingestellte Messschieber. Rechts:
Vermessung eines 30mm Endmaßes .................................................... 154
Abbildung 92: Zellen vom Typ L929 auf dem Substrat Titan*. Bildartefakte,
hervorgerufen durch Korrosionsprozesse, sind rot und
Hintergrundbereiche ohne Artefakte weiß markiert. ............................... 165
Abbildung 93: Abweichung des Rot- und Blaukanals zum Grünkanal für 10
verschiedene Bildartefakte, hervorgerufen durch Materialkorrosion ...... 166
Abbildung 94: Abweichung des Rot- und Blaukanals zum Grünkanal für 10
verschiedene Hintergrundbereiche ohne Artefakte, hervorgerufen durch
Materialkorrosion ................................................................................... 167
Abbildung 95: Summe der relativen Abweichungen des Rot- und Blaukanals
vom Grünkanal für 10 verschiedene Artefakt- sowie Hintergrundbereiche
............................................................................................................... 168
Abbildung 96: Berechnung der Farbzusammensetzung des Materials Titan* auf
Basis der 5 in grün markierten Bildbereiche .......................................... 169
Abbildung 97: Berechnung der Farbzusammensetzung des Materials Stahl* auf
Basis der 5 in grün markierten Bildbereiche .......................................... 170
Abbildung 98: Berechnung der Farbzusammensetzung des Materials Polystyrol
auf Basis der 5 in grün markierten Bildbereiche .................................... 171
Abbildung 99: Vergleich der Fläche von 100 Einzelzellen und 100 Zellcluster
aus fünf unterschiedlichen Proben der Linie L929 ................................. 176
Abbildung 100: Vergleich der Kompaktheit von 100 Einzelzellen und 100
Zellcluster aus fünf unterschiedlichen Proben der Linie L929 ................ 176
Abbildung 101: Die Kompaktheit aufgetragen über die Zellfläche für 100
Einzelzellen (blau) und 100 Zellcluster (rot) ........................................... 177

1. Einleitung 1
1. Einleitung
Der Einsatz von Implantaten im menschlichen Körper ist ein wichtiger Bestand-
teil der modernen Medizin. So führen beispielsweise Gelenkprothesen zur Stei-
gerung der Lebensqualität, wenn die Funktionalität eines oder mehrerer Gelen-
ke nicht mehr gegeben ist. Darüber hinaus können Stents oder Herzschrittma-
cher die Lebensdauer erkrankter Menschen verlängern.
Untersuchungen zur demographischen Entwicklung (1) zeigen, dass die durch-
schnittliche Lebenserwartung der Bevölkerung stetig anwächst. Dies führt zu
einer steigenden Nachfrage an Implantaten, da im fortgeschrittenen Alter ver-
mehrt Ermüdungserscheinungen verschiedener Körperteile, z.B. des Hüftge-
lenks auftreten, die teilweise durch geeignete Implantate kompensiert werden
können. Ein weiterer Grund für die zunehmende Nachfrage an Implantatwerk-
stoffen liegt in der ständigen Erweiterung des Anwendungsspektrums. So
kommen derzeit z.B. auch Materialien zur Behandlung von arteriellen Erkran-
kungen (2) zum Einsatz.
Die eingesetzten Biomaterialien müssen einerseits die gewünschte Funktion
(Biofunktionalität) erfüllen, z.B. die Übertragung von Lasten. Andererseits ist die
Sicherstellung der Körperverträglichkeit (Biokompatibilität) zu gewährleisten (3).
Der Grad der Biokompatibilität für Knochenimplantate wird nach Schenk (4) in
Inkompatibel, Biokompatibel, Bioinert, Bioaktiv, Induktiv und Konduktiv unter-
teilt. Inkompatible Materialien setzen bestimmte Substanzen in toxischen Men-
gen frei, die Abwehrreaktionen des Körpers hervorrufen und im schlimmsten
Fall zu einer Abstoßung des eingesetzten Implantates führen können. Um dies
zu verhindern, erfolgt vor dem Einsatz eines Implantates eine mehrstufige Bio-
kompatibilitätsprüfung des zugrundeliegenden Materials. Die erste Stufe um-
fasst in-vitro1-Untersuchungen mit isolierten Zellen, die auch Gegenstand der
vorliegenden Arbeit sind. In der zweiten Stufe werden Tierversuche (in-vivo-
Tests) und in der letzten Stufe klinische Untersuchungen am Menschen durch-
geführt (3).
Damit die von unterschiedlichen Laborfachkräften durchgeführte Biokompatibili-
tätsprüfung möglichst einheitlich umgesetzt wird, muss die Durchführung der in-
vitro-Versuche nach der Norm EN ISO 10993-5 (5) erfolgen. Bei der Bewertung
1 In-vitro: Hierbei handelt es sich um organische Vorgänge die außerhalb des lebenden Orga-
nismus stattfinden, z.B. Experimente mit Reagenzgläsern

1. Einleitung 2
der Biokompatibilität wird die Interaktion des Werkstoffs u.a. mit adhärent2
wachsenden Zellen analysiert. Dabei haben sich verschiedene Zellmerkmale
als geeignete Indikatoren für die Bewertung des zugrundeliegenden Materials
herausgestellt (Tabelle 1) (3).
Tabelle 1: Kriterien für die in vitro-Biokompatibilität in Zell- und Gewebekulkulturen (3)
Kriterium (Zellverhalten)
schlecht biokompatibel -> gut biokompatibel
Wachstum / Zelldichte Zellen sterben ab Zellen vermehren sich
Morphologie Abgekugelt Ausgebreitet
Adhäsion Schwach Stark
Benetzung Schlecht Gut
Stoffwechselprodukte Verändert Unverändert
Im Allgemeinen breiten sich Zellen auf einem biokompatiblen Material wesent-
lich stärker aus als auf einem weniger biokompatiblen Werkstoff. Ebenso ver-
ändert sich die Zellmorphologie mit abnehmender Biokompatibilität von flächi-
gen über spindelförmigen bis hin zu kleinen kugelförmigen Zellbereichen (Abb.
1).
Abbildung 1: Zellmorphologische Änderung bei Abnahme der Biokompatibilität (3)
Bei der Durchführung von in-vitro-Tests sollten möglichst die gleichen Zelllinien3
verwendet werden, die auch bei den in-vivo-Tests am Tier oder Mensch im di-
rekten Kontakt mit dem eingesetzten Implantat stehen (3). So ist die Wechsel-
wirkung zwischen dem später eingesetzten Werkstoff und dem umgebenen
Zellgewebe absehbarer. Ein gängiger Zelltyp bei der Biokompatibilitätsprüfung
ist die Linie L929. Dabei handelt es sich um Fibroblasten, die den Bindege-
webszellen zugeordnet sind und ein Implantat im Muskel oder unter der Haut
umgeben können (2).
2 Adhärent wachsende Zellen: Die Zellen haften auf der Probenoberfläche und beginnen zu
wachsen 3 Zelllinie: Ein bestimmter Typ einer Zelle, z.B. L929

1. Einleitung 3
1.1 Motivation und Zielsetzung
In-vitro-Prüfungen werden derzeit meist durch geschulte Laboranten durchge-
führt. Die in Tabelle 1 dargestellten Kriterien zur Beurteilung der Biokompatibili-
tät werden teilweise visuell bestimmt. Der Vorteil einer Sichtprüfung durch La-
borfachkräfte liegt in der Anwendung des großen Erfahrungsschatzes, der
durch ein computergestütztes Verfahren kaum zu ersetzen ist. Nachteile sind
zum einen der größere Zeitaufwand sowie die Fehleranfälligkeit bei den manu-
ell durchgeführten Prüfungen (6; 7; 8). Der Einsatz einer Analysesoftware als
Assistenzsystem für die Laborfachkraft bringt einige Vorteile mit sich. Zu nen-
nen sind die Reduktion der Prüfzeit, u.a. bei der Zellzählung, und die damit ein-
hergehende Kostenreduktion sowie eine objektive Zellanalyse mit reproduzier-
baren Ergebnissen. Mit Hilfe dieser Voruntersuchungen wäre es möglich, die
Kosten für den Prüfaufwand durch ein zertifiziertes Labor zu reduzieren, indem
einige Werkstoffe bereits im Vorfeld ausgeschlossen werden können. Darüber
hinaus können verschiedene Zellmerkmale, z.B. die durchschnittliche Einzel-
zellfläche oder die Kompaktheit bestimmt werden, die von Laborfachkräften oh-
ne zusätzliche Hilfsmittel nicht quantitativ erfassbar sind. Unter der Kompaktheit
ist wie üblich in der Bildverarbeitung
2
4
LK
A (9) zu verstehen. (Gl. 1)
L entspricht der Konturlänge des Objektes und A der Objektfläche.
Das Ziel dieser Arbeit umfasst die Entwicklung einer flexibel einsetzbaren Ana-
lysesoftware, um die genannten Vorteile in den Prozess der Biokompatibilitäts-
prüfung einfließen zu lassen. Unter flexibel ist hier zu verstehen, dass die Soft-
ware nicht nur bei einer bestimmten Zelllinie Anwendung findet, sondern bei
morphologisch unterschiedlichen Zelltypen eine genaue Analyse gewährleisten
soll. Aus diesem Grund wird neben der Zelllinie L929 auch der Typ MC3T3 bei
der Entwicklung der Analysemethoden berücksichtigt. Somit kann das compu-
tergestützte Verfahren neben der Biokompatibilitätsprüfung auch für andere
Applikationen eingesetzt werden, z.B. die Detektion von Korrosionserscheinun-
gen oder das Auswerten von Suspensionskulturen.
Aufgrund der genannten Ziele ergibt sich ein Anforderungsprofil an die zu ent-
wickelnde Software. Sie sollte eine Analyse von Zellen des Typs L929 und
MC3T3 auf unterschiedlichen Materialien wie Glas, Kunststoff oder Metall er-
möglichen. Eine Analyse weiterer Zelllinien ist wünschenswert. Innerhalb der
Zellen ist eine Segmentierung der Zellkerne sowie die genaue Bestimmung der

1. Einleitung 4
Zellanzahl innerhalb des Probenausschnittes zu gewährleisten und auf die ge-
samte Probe hochzurechnen. Des Weiteren ist es erforderlich die Zelldichte und
geometrische Charakteristika wie die Zellfläche und Rundheit im Probenaus-
schnitt zu bestimmen. Für eine individuelle Zellauswertung müssen zu einem
Zellcluster zusammengewachsene Zellen getrennt werden. Neben den automa-
tischen Analysemethoden soll der Anwender die Möglichkeit besitzen im Zwei-
felsfall manuell die Analyse zu optimieren, z.. durch entfernen, editieren oder
trennen von segmentierten Objektbereichen. Eine Stapelverarbeitung soll die
automatisierte Auswertung von mehreren Bildern ermöglichen und Ergebnisda-
ten sowie Bilder zur Laufzeit des Programms abspeichern.Zur späteren Analy-
se von bereits gespeicherten Daten ist eine Importfunktion vorzuse-
henWünschenswert ist eine einfach zu bedienende Benutzeroberfläche auf der
notwendige Parametereinstellungen für die Algorithmen auf separater Ebene
von dafür autorisierten Personen vorgenommen werden.
Die vorliegende Aufgabe beinhaltet einige besondere Herausforderungen. Wäh-
rend des Zellwachstums auf dem zu untersuchenden Material bilden sich oft
agglomerierte Zellbereiche. Zur exakten Zellzahlbestimmung und der Auswer-
tung bestimmter Merkmale wie z.B. der durchschnittlichen Einzelzellfläche oder
Kompaktheit müssen die Zellen separiert vorliegen. Zusammenhängende Zell-
bereiche weisen jedoch unterschiedliche morphologische Charakteristika zwi-
schen verschiedenen Zelllinien auf (Abb. 2).
Abbildung 2: Links: Agglomerierte L929 Zellen auf dem Material Stahl*. Sie bilden oft Veren-
gungen an den Zellkontaktstellen. Rechts: Kompakt zusammenhängende Zellen vom Typ
MC3T3 auf dem Material Titan*. Die Bilder wurden im Biotechnologielabor der Fachhochschule
Südwestfalen aufgenommen
Abb. 2 zeigt links agglomerierte Zellen der Linie L929 mittig im Bild. Sie sind oft
durch Verengungen an den Zellkontaktstellen gekennzeichnet. Hingegen
wachsen Zellen des Typs MC3T3 oft flächig zusammen und bilden kompakte

1. Einleitung 5
Formen. Diese Eigenschaften müssen bei der Analyse berücksichtigt werden,
denn eine hinreichend genaue Zellseparation ist Voraussetzung für eine
korrekte Beurteilung der Biokompatibilität des Substrates.
Darüber hinaus variiert die Morphologie der Zellen relativ stark, so dass die
Klassifikation in vitale und abgestorbene Zellen erschwert wird. Eine weitere
Herausforderung stellt die verwendete Färbemethode dar. Allgemein wird eine
Zellfärbung zur optischen Kennzeichnung verschiedener Bereiche wie z.B. dem
Zellkern durchgeführt, damit eine visuelle Überprüfung der gefärbten Zellareale
möglich ist. Dazu stehen verschiedene Färbemethoden zur Verfügung. Bei der
Fluoreszenzfärbung wird ein Protein der Zelle mit einem fluoreszierenden
Farbstoff (Fluorchrom) markiert. Nach Anregung des Fluorchroms mit Licht
eines definierten Wellenlängenintervalls emittiert es ebenfalls Licht. Das Protein
ist vermehrt in einem bestimmten Zellareal vorzufinden, z.B. dem
Zellkernbereich. Nach diesem Prinzip ist auch eine Mehrfachmarkierung
verschiedener Proteine möglich. Die Visualisierung verschiedener
Zellbestandteile geschieht dann mit Hilfe eines Fluoreszenzmikroskops (10).
Vorteil dieser Färbemethode ist der starke Kontrast im Bild, der die
Zellsegmentierung mittels Bildverarbeitungsmethoden einfach gestaltet.
Nachteil ist der im Vergleich zu konventionellen Auflichtmikroskopen hohe
Anschaffungspreis eines Fluoreszenzmikroskopes und ebenfalls höhere Kosten
für die Farbstoffe. Darüber hinaus ist eine dauerhafte Präparation der Zellen
nicht möglich. Im Rahmen dieser Arbeit werden die Bilder mit Hilfe einer
zytochemischen4 Färbung ohne Fluoreszenz analysiert. Zum Einsatz kommt die
so genannte May-Grünwald Färbung (10). Vorteile gegenüber der
Fluoreszenzfärbung sind zum einen die optische Hervorhebung verschiedener
Zellbereiche mit nur einem Färbevorgang. Zum anderen ist die Durchführung
des Färbeprozesses weniger aufwendig und für die Visualisierung kommt ein
konventionelles Durchlicht/- Auflichtmikroskop zum Einsatz. Allerdings fällt der
Bildkontrast bei dieser Färbemethode im Vergleich zum fluoreszenzbasierten
Verfahren geringer aus.
4 Zytochemische Färbung: Die Zellen werden durch eine chemische Reaktion angefärbt

1. Einleitung 6
Abbildung 3: Zellkernfärbung mit DRAQ5 Abbildung 4: Zellfärbung mit May- Grünwald
an lebenden HEK- Zellen (10) an L929 Zellen
In Abb. 3 erscheinen die Zellkerne mit Hilfe einer Fluoreszenzfärbung rot. Auf-
fällig ist der starke Kontrast zum Bildhintergrund. In Abb. 4 sind die Zellen mit
der May-Grünwald Färbung präpariert worden. Bei den größeren Zellen treten
die verschiedenen Zellbereiche durch unterschiedliche Farben auf Kosten eines
geringeren Kontrastes hervor. Folglich wird die Zellsegmentierung bei Verwen-
dung der May-Grünwald Färbung erschwert.
1.2 Gliederung der Arbeit
Nach der Einleitung in das Thema der Biokompatibilitätsprüfung und den Erläu-
terungen zur Problemstellung (Kap. 1.1) leiten sich daraus die Ziele für diese
Arbeit ab. Im folgenden Kapitel (Kap. 2) wird der Stand der Forschung im Be-
reich der Zellsegmentierung sowie Zellseparation beschrieben und daraus ge-
folgert, welche bekannten Verfahren zur Lösung der vorliegenden Aufgabe bei-
tragen könnten und demnach umfassender untersucht werden und für welche
Problemstellungen die Entwicklung neuer Methoden sinnvoll erscheint. Es folgt
die Erläuterung der für die Zellanalyse notwendigen Vorbereitungen. Diese um-
fassen die Probenaufbereitung und die Zellkultivierung (Kap. 3). Anschließend
werden die Softwareanforderungen und das Konzept der Programmarchitektur
vorgestellt (Kap. 4). Im Anschluss daran werden geeignete Verfahren für die
automatische Analyse der untersuchten Zelltypen vorgestellt und diskutiert. Ge-
genstand sind zum einen verschiedene Segmentierungsalgorithmen (Kap. 5)
zur Erfassung der Zellen ebenso wie unterschiedliche Methoden zur Separation
agglomerierter Zellen (Kap. 6). Es schließt sich ein Konzept zur Automatisie-
rung der Algorithmenauswahl (Kap. 7) und zur automatischen Festlegung der
Algorithmenparameter (Kap. 8) an. Die Beurteilung des Biokompatibilitätsgra-
des geschieht unter Zuhilfenahme verschiedener Merkmale der segmentierten
Zellen, wobei geeignete Merkmale in Kap. 9 ausgearbeitet werden. Im Ergeb-
nis- und Diskussionsteil (Kap. 10) wird das computergestützte Verfahren zur

1. Einleitung 7
Beurteilung der Biokompatibilität mit dem konventionellen Verfahren verglichen
und die Vor- und Nachteile diskutiert. Darüber hinaus werden weitere Anwen-
dungsmöglichkeiten mit Hilfe der Zellanalysemethode vorgestellt. Den Schluss
der Arbeit bildet die Zusammenfassung mit einer kurzen Erläuterung von der
Motivation über die Umsetzung der geforderten Ziele bis zu den Ergebnissen.
Die in Kap. 3.1 und Kap. 3.2 beschriebenen Vorgänge zur Probenaufbereitung
sowie Allgemein die Arbeitsschritte vom Aufbringen der verwendeten Zellen auf
die Probenoberfläche bis hin zum Färbeprozess werden im Rahmen des inter-
disziplinären Projektes „BioCV“ von Mitarbeitern des Labors für Biotechnologie
an der FH Südwestfalen durchgeführt.

2. Stand der Forschung 8
2. Stand der Forschung
Die Bildverarbeitung nimmt in der modernen Medizintechnik einen immer höhe-
ren Stellenwert ein und ist für viele Anwendungsbereiche ein wichtiges Werk-
zeug geworden (11; 12). So hat sich eine programmtechnische Auswertung z.B.
zur Detektion von Tumorgewebe (13) oder in der Radiologie zur Unterstützung
des Arztes bei der Diagnosestellung bereits etabliert (3). Hingegen wird die in-
vitro Biokompatibilitätsprüfung von Implantatmaterialien derzeit nach einem
standardisierten Verfahren mit dem geschulten Auge von Laborfachkräften
durchgeführt (5). Zur Unterstützung der Laborfachkräfte bei der Biokompatibili-
tätsprüfung muss ein computergestütztes System zunächst analog zu anderen
biomedizinischen Anwendungsgebieten Zellen in mikroskopischen Bildern
segmentieren können. Dazu finden sich in der Literatur verschiedene Verfahren
(Kap. 2.1).
2.1 Segmentierungsverfahren für biomedizinische Anwendungen
Für die Segmentierung von Zellen in mikroskopischen Bildern steht eine Viel-
zahl an Methoden zur Verfügung. Diese werden typischerweise in die Katego-
rien pixel-, kanten-, regionen-, modell- und texturbasierte Verfahren eingeteilt.
Die in (14; 15; 16; 17) aufgeführten Verfahren stellen beispielsweise verschie-
dene pixelbasierte Schwellwertmethoden vor, die für die Zellsegmentierung
eingesetzt werden können. M. Sezgin et al. (18) haben ca. 40 verschiedene
schwellwertbasierte Verfahren, abhängig von der genutzten Information (z.B.
histogrammbasierte- oder lokalbasierte Schwellwertbildung), in sechs Gruppen
kategorisiert und für verschiedene Anwendungen auf ihre Leistungsfähigkeit hin
getestet. Das Ergebnis zeigt, dass, abhängig vom Anwendungsgebiet, unter-
schiedliche Schwellwertverfahren am besten für die Segmentierung geeignet
sind und keine Methode ausreichend genaue Ergebnisse für alle geprüften An-
wendungsfälle liefert. In (19) werden Zellen mit Hilfe eines Region-Growing-
Algorithmus segmentiert. Nach Auswahl der benötigten Startpunkte innerhalb
grob vorsegmentierter Zellbereiche werden die Nachbarpixel nacheinander auf
Homogenität geprüft. Homogenitätskriterien sind in diesem Fall die Abweichung
des Grauwertes und des Gradientenbetrags zu dem durchschnittlichen Grau-
wert und Gradientenbetrag aller Pixel aus der Vorsegmentierung. Sind die Ho-
mogenitätskriterien erfüllt, wird der benachbarte Pixel der Region hinzugefügt.
Modellbasierte Verfahren werden in (8; 20; 21; 22) vorgestellt. Sie verwenden
zur Objektdetektion im Vorfeld generierte formbasierte Modelle des zu suchen-

2. Stand der Forschung 9
den Objektes. Da dieses nicht immer die gleiche Form im Bild aufweist, können
unter Zuhilfenahme deformierbarer Templates (23; 24; 25) morphologisch leicht
variierende Objekte segmentiert werden. Swain und Ballard (26) entwickelten
den Histogram-Backprojection-Algorithmus. Dabei werden nicht formbasierte
Merkmale, sondern Farbwerte im Modell hinterlegt. Auf Basis der Farbhisto-
gramme des Modells und des Bildes, in dem das Objekt gesucht wird, kann auf
die Position des Objektes im Bild geschlossen werden.
In der medizinischen Bildauswertung haben hybride Segmentierungsverfahren
einen großen Stellenwert erlangt (27). Diese Methoden stellen eine Mischung
der Algorithmen aus den o.g. Segmentierungskategorien dar und versuchen die
jeweiligen Vorteile für eine bessere Segmentierung zu kombinieren. Die Wass-
erscheidentransformation ist eine Mischform von regionenbasierter- und kan-
tenbasierter Segmentierung, die in der medizinischen Bildverarbeitung zum
Einsatz kommt.
T. Pommerencke et al. (28) beschreiben ein Verfahren, das mit Hilfe der Wass-
erscheidentransformation Zellkerne und Zellen in mehrfach fluoreszenzgefärb-
ten Gewebeschnitten detektiert. Nach einer Kontrasterhöhung und anschlie-
ßender Gewebesegmentierung erfolgt die Detektion der Zellkerne mit einem
adaptiven Schwellwertverfahren. Überschreiten die Kerne eine bestimmte Flä-
che, wird angenommen, dass es sich um einen Cluster handelt. Die enthaltenen
Kerne werden mit der Wasserscheidentransformation auf Basis lokaler Intensi-
tätsmaxima segmentiert. Die anschließende Ermittlung der Zellgrenzen erfolgt
ebenfalls mit der Wasserscheidentransformation.
Ein weiteres hybrides Segmentierungsverfahren stellen die aktiven Konturen,
auch Snakes genannt, dar (29; 30; 31; 32; 33). Tscherepanow et al. (30) stellen
ein auf aktiven Konturen basierendes Verfahren zur Segmentierung ungefärbter
Zellen in Hellfeld-Mikroskopbildern vor. Im ersten Schritt erfolgt die Generierung
der für die Snakes notwendigen Startkonturen mit Hilfe morphologischer Opera-
tionen und der Distanz-Transformation. Im nächsten Schritt werden die Zellen
mit Hilfe aktiver Konturen segmentiert. Die Vorgehensweise ist dem Balloon-
Verfahren von Cohen (34) sehr ähnlich. Der Unterschied besteht jedoch darin,
dass nicht die Normalenvektoren, sondern die Entfernung der Snakes zur Start-
kontur berücksichtigt wird. Dadurch vermeidet man laut Tscherepanow eine
Konturüberlappung beim Wachstum konkaver Startkonturen.
Mit Methoden aus dem Bereich der Computational Intelligence (CI) stehen wei-
tere Verfahren zur Zellsegmentierung zur Verfügung (35; 36; 37; 38). Die CI ist
ein Teilgebiet der künstlichen Intelligenz und beinhaltet die Fuzzy-Logik, Evolu-
tionäre Algorithmen und Künstliche Neuronale Netze (KNN), die nach dem Vor-

2. Stand der Forschung 10
bild der Natur versuchen technische Probleme zu lösen (39). S. Colantonio et
al. (35) entwickelten ein fuzzy-neuronales Verfahren zur Segmentierung von
Kernen der Lymphzellen in mikroskopischen Bildern. Zunächst werden mit ei-
nem fuzzy-c-means basierenden Algorithmus grob die Zellen und der Hinter-
grund segmentiert. Anschließend kommt ein KNN zum Einsatz, dass die vor-
segmentierten Zellen in verschiedene Bereiche, z.B. die Zellkontur oder das
Zytoplasma, klassifiziert. Dazu ist das KNN im Vorfeld mit korrekt klassifizierten
Daten auf die Problemstellung zu trainieren. Zur Klassifizierung einer vorseg-
mentierten Zelle dienen dem KNN die Werte geeigneter Bildmerkmale, z.B. der
durchschnittliche Farbwert oder der Gradientenbetrag, als Inputdaten.
R. Q. Feitosa et al. (37) bieten mit Hilfe eines genetischen Algorithmus zur au-
tomatischen Bestimmung von Parametern eines regionenbasierten Segmentie-
rungsverfahrens eine wissensbasierte Lösung an. Die Wissensbasis besteht
aus im Vorfeld korrekt segmentierten Bildsegmenten. Nach einer zufälligen Ini-
tialisierung der Parameterwerte werden diese in einem evolutionären Prozess
optimiert. Der Prozess endet nach einer vorgegebenen Anzahl an Iterationen
mit der Segmentierung des Bildes unter Verwendung des zuletzt generierten,
entsprechend der vorgegebenen Fitnessfunktion, optimalen Parametersatzes.
Metzler et al. (40) verwenden mit Hilfe eines evolutionären Algorithmus eben-
falls ein wissensbasiertes Verfahren, hier nicht zur automatischen Parametrisie-
rung eines vorgegebenen Segmentierungsverfahrens, sondern zur direkten
Segmentierung verschiedener Gehirnareale in MR-Aufnahmen.
Reham Mohammed et al. (41) stellen ein Verfahren zur Segmentierung von ge-
färbten Leukemie Krebszellen vor. Dazu werden die Farbbilder zunächst in den
C-Y Farbraum transformiert und die Zellsegmentierung auf Basis des Y-Bild-
Histogramms durchgeführt. Zur Bildnachbearbeitung kommt ein Medianfilter
zum Einsatz, anschließend erfolgt die Rücktransformation in den RGB-
Farbraum.
2.2 Verfahren zur Separation agglomerierter Zellen
Die Auswertung von geeigneten Zellmerkmalen bei der Biokompatibilitätsprü-
fung ist ein elementarer Bestandteil, denn die Merkmale dienen zur Beurteilung
des Biokompatibilitätsgrades. Aus diesem Grund müssen möglichst alle im Bild
vorhandenen Zellen segmentiert werden. Allerdings können diese, abhängig
vom Toxizitätsgrad des verwendeten Substrates und vom Zelltyp selbst, im Ver-

2. Stand der Forschung 11
lauf ihres Wachstums agglomerieren. Daher ist es notwendig, diese Bereiche
durch geeignete Methoden zu vereinzeln.
B. Nilsson und A. Heyden (42) segmentieren dicht besiedelte Leukozyten aus
dem Knochenmark in mikroskopischen Bildern mit Hilfe der Wasserscheiden-
transformation. Nach einer Segmentierung der Kernbereiche über ein Schwell-
wertverfahren wird eine grauwertbasierte Distanztransformation durchgeführt.
Daran schließt sich eine Wasserscheidentransformation zur Separation sich
teilweise überschneidender Zellkerne an. Um einer durch die Wasserscheiden-
transformation verursachten Übersegmentierung entgegenzuwirken, folgt eine
Zusammenfassung benachbarter Segmente. Das betrifft die Segmente, deren
Durchmesser einen vorgegebenen Schwellwert unterschreiten.
Yung-Kuan Chang et al. (43) segmentieren HEp-2 Zellen in fluoreszenzgefärb-
ten Bildern und trennen zusammengewachsene Zellen voneinander. Zunächst
erfolgt eine Bildvorverarbeitung zur Transformation der Farbbilder in Graustufen
und zur Reduzierung des Rauschens. Die Zellkonturen werden über ein Gradi-
entenverfahren bestimmt und überlappende Zellbereiche auf Basis einer Wass-
erscheidentransformation mit anschließender Distanztransformation separiert.
U. Pal et al. (44) nutzen die bei manchen Zellagglomeraten auftretenden Ver-
jüngungen an den Kontaktstellen zur Separation der Einzelzellen aus. Diese
Verengungen spiegeln sich in den lokalen Minima richtungsabhängigen Ab-
standsfunktionen wider. Unter Einbeziehung eines zellspezifischen Regelwer-
kes werden die resultierenden dominanten Konturpunkte miteinander verbun-
den und separieren die zusammenhängenden Zellen.
M. Grobe et al. (45) stützen sich für die Separation neben den Verengungen an
den Zellkontaktstellen noch auf Farbunterschiede im Überlappungsbereich der
Zellen. Die Zellen sowie deren Kerne werden mit Hilfe eines histogrammbasier-
ten Schwellwertverfahrens aus zwei verschiedenen fluoreszenzgefärbten Bil-
dern segmentiert. Die Separation erfolgt analog mit Hilfe eines histogrammba-
sierten Schwellwertverfahrens unter den Prämissen, dass die überlappenden
Zellbereiche dunkler erscheinen als sich nicht überlappende Bereiche. Des
Weiteren muss jeder Überlappungsbereich durch zwei Einschnürungspunkte in
der Kontur gekennzeichnet sein.
In (46) und (47) werden weitere Separationsmethoden vorgestellt, die auf Basis
der Konturverengungen die zusammenhängenden Zellen aufspalten.
V. Metzler e t al. (38) entwickelten ein Verfahren zur Trennung von agglomerier-
ten L929 Zellen basierend auf morphologischen Operationen. Mit Hilfe eines auf
der Erosion beruhenden Algorithmus generieren sie zuerst Marker für die Identi-

2. Stand der Forschung 12
fizierung der einzelnen Zellen in den Zellverbänden. Die Rekonstruktion der
Zellen erfolgt daraufhin mit Hilfe eines erweiterten Dilatationsprozesses.
2.3 Bestehende Zellanalysesysteme
Die Cognition Network Technology (CNT) (48) und Gipsy (49) sind Software-
programme, die zur Lösung des vorgestellten Problems geeignet sein könnten.
Bei der CNT handelt es sich um ein objektbasiertes Bildanalysesystem für bio-
medizinische Fragestellungen, das von der Firma Definiens AG 1996 entwickelt
wurde. Das Konzept basiert darauf, die Analyse nicht ausschließlich pixelbasiert
zu betrachten, sondern Objekthierarchien (z.B. Zellkernbereiche) erzeugen und
bearbeiten zu können. Zur Zellerfassung stehen verschiedene Segmentie-
rungsverfahren zur Verfügung, die applikationsabhängig zum Einsatz kommen.
Ein interaktives Fuzzy- Klassifikationssystem ermöglicht dem Anwender, Bildbe-
reiche in unterschiedliche Objektklassen einzuordnen (3). Das CNT-System ist
in dem Softwarepaket „Definiens Developer XD“ integriert und kommerziell er-
hältlich.
Gipsy wurde für die optische Inspektion von Materialien in Echtzeit entwickelt
und verfolgt einen wissensbasierten Ansatz. Im Vorfeld erstellte Trainingsdaten
enthalten die für die Anwendung benötigten Objektklassen sowie deren Merk-
malswerte. Die Generierung dieser Daten erfolgt durch einen anwendungsspe-
zifischen Bildverarbeitungsalgorithmus. Lernverfahren erzeugen daraus ein se-
mantisches Netzwerk, wobei der Anwender bei Bedarf Modifikationen am Netz,
z.B. das Hinzufügen oder Entfernen von Merkmalen, vornehmen kann.
A. Rao und E.A. Vaisberg (50) haben sich 2007 ein Verfahren zur automati-
schen Analyse von gefärbten Leberzellen patentieren lassen. Zur Visualisierung
unterschiedlicher Zellareale dienen verschiedene Farbstoffe. Die Segmentie-
rung der durch die Farbstoffe markierten Zellbereiche wird mit Hilfe von
schwellwert- und kantenbasierten Verfahren sowie einem Algorithmus basie-
rend auf der Wasserscheidentransformation durchgeführt. Durch eine anschlie-
ßende Charakterisierung der erfassten Zellbereiche können Aussagen über
verschiedene medizinische Fragestellungen, z.B. das aktuelle Stadium einer
Erkrankung, getroffen werden.
Mit TissueQuest und HistQuest stehen zwei weitere Softwareanalyseprogram-
me der Firma TissueGnostics GmbH zur Analyse von fluoreszenz- und zyto-
chemisch gefärbten Zellen zur Verfügung. Da es sich um kommerziell vertrie-
bene Produkte handelt, sind die für die Zellanalyse verwendeten Algorithmen
nicht bekannt.

2. Stand der Forschung 13
2.4 Schlussfolgerung
Bis heute wurde kein universell einsetzbares computergestütztes Verfahren
entwickelt, das allen biomedizinischen Anforderungen gerecht wird (3) (27). Die
Gründe dafür sind vielseitig. Zum einen existieren verschiedene Bilderzeu-
gungssysteme (z.B. MRT, Ultraschall, REM, CCD Kameras,…), die Bildmaterial
mit stark unterschiedlichen Kontrasten liefern. Teilweise ist auch das Experten-
wissen einer erfahrenen Fachkraft zur Analyse notwendig. Die Integration des
Expertenwissens in ein computergestütztes Verfahren ist allerdings nur schwer
möglich und erfordert eine fachübergreifende Zusammenarbeit von Experten.
Nach Wintermantel (3) ist die Leistungsfähigkeit einer automatischen Bildaus-
wertung z.B. für eine automatische Diagnosestellung nach Auswertung eines
CT-Bildes „…noch weit entfernt von der kognitiv-diagnostischen Expertise eines
erfahrenen Radiologen“.
Die in Kap. 2.3 genannten Softwaretools CNT und Gipsy könnten zur Bearbei-
tung des vorliegenden Problems aufgrund ihres objekt- und wissensbasierten
Ansatzes sowie des hohen Funktionsumfanges geeignet sein. Die Untersu-
chungen in (7) zeigen jedoch, dass die Zellsegmentierung von anderen Verfah-
ren im Vergleich zur CNT bessere Ergebnisse liefern. Weitere Schwachpunkte
sind die unübersichtliche Programmoberfläche, die viel Zeit zur Einarbeitung
benötigt sowie des Umstandes, dass die Software nur kommerziell erhältlich ist.
Die Verwendung des automatischen Inspektionssystems Gipsy erfordert im
Vorfeld die Erzeugung vieler Trainingsdatensätze, um eine möglichst gute Wis-
sensbasis zu erhalten. Zudem werden keine Erweiterungsmöglichkeiten der
enthaltenen Algorithmen für komplexe Problemstellungen erläutert und es er-
scheint fraglich, ob eine Quantisierung von Merkmalswerten in vier Stufen für
verschiedene Analyseanforderungen hinreichend genau ist.
Das im Patent (50) beschriebene Zellanalyseverfahren ist für die Lösung der
vorliegenden Problemstellung ebenfalls nicht geeignet. Die Anwendung kon-
zentriert sich auf die Analyse von Leberzellen und benötigt verschiedene Fluo-
reszenzfarbstoffe zur Markierung unterschiedlicher Zellareale und ist somit nicht
für Anwendungen im Rahmen histologisch gefärbter Zellen einsetzbar. Eine in
der Patentschrift hinterlegte Skizze lässt außerdem darauf schließen, dass es
sich bei den untersuchten Zellen um eine morphologisch einfache Zellstruktur
handelt.
Die zu untersuchenden Zelltypen besitzen eine starke morphologische sowie
farbliche Variation, so dass der Einsatz von Segmentierungsmethoden, basie-
rend auf Wasserscheidentransformation, Kantenfiltern und geometrischen

2. Stand der Forschung 14
Merkmalen ungeeignet erscheint. Hingegen sollte eine Untersuchung verschie-
dener Schwellwertmethoden aufgrund ihrer Schnelligkeit sowie der aktiven Kon-
turen wegen ihrer robusten Segmentierung auch in kontrastarmen Bildern für
die vorliegende Problemstellung erfolgen. Ebenso sinnvoll erscheint der Einsatz
eines Künstlichen Neuronalen Netzes (KNN) zur Segmentierung unterschiedli-
cher Zellbereiche, da KNNs gut auf Klassifikationsprobleme ansprechen.
Viele Verfahren zur Zellseparation in mikroskopischen Bildern stützen sich auf
sehr spezielle Zellcharakteristika. So werden oft runde oder elliptische Zellgeo-
metrien (46; 47; 51) oder Farbunterschiede im Überlappungsbereich der Zellen
(45) für die Separation vorausgesetzt. Diese Verfahren lassen sich dadurch auf
keine oder wenige andere Zelllinien erweitern. Zum Beispiel die von Yung-Kuan
Chan et al. (43) vorgestellte Methode beschränkt sich ausschließlich auf die
Segmentierung und Separation von fluoreszenzgefärbten HEp-2 Zellen, wobei
die Zellmorphologie dieses Typs aufgrund fehlender Zellausläufer und der rela-
tiv runden Geometrie einfacher automatisch zu analysieren ist im Vergleich zum
Typ L929 und MC3T3. Im Ergebnisteil der vorgestellten Methode ist für 20
Testbilder mit insgesamt 1242 Zellen eine Abweichung von 5% zur Referenz-
zellzahl angegeben. Es ist davon auszugehen dass sich bei der hier vorliegen-
den komplexeren Zellmorphologie sowie der kontrastärmeren May-
Grünwaldfärbung schlechtere Ergebnisse einstellen die zu einer ungenauen
automatischen Bestimmung des Zytotoxizitätsgrades eines vorliegenden Mate-
rials führen.
Das Zellsegmentierungsverfahren von Reham Mohammed et al. (41) eignet
sich gut für die Segmentierung von Zellen die sich farblich deutlich vom Hinter-
grund unterscheiden. Die gezeigten Krebszellen weisen einen starken Kontrast
und eine andere Farbgebung zu den übrigen Bildbereichen auf. Durch die
Transformation in den C-Y Farbraum und die Verwendung des Y-Bildes liegen
die Krebszellen relativ isoliert im Bild vor. Der Einsatz dieser Methode eignet
sich jedoch nur für die Segmentierung fluoreszenzgefärbter Zellen da die ge-
färbten Zellbereiche im Vergleich zu der hier verwendeten May-
Grünwaldfärbung einen deutlich stärkeren Kontrast zur Umgebung aufweisen.
Das Separationsverfahren von U. Pal et al. (44) zur Detektion von Konturveren-
gungen liefert hingegen einen guten Ansatz für die Trennung agglomerierter
Zellen, die Verengungen an den Zellkontaktstellen aufweisen. Eine Erweiterung
des Verfahrens, so dass trotz teilweisem Ausbleiben von Konturverengungen
weiterhin aus biologischer Sicht korrekte Zelltrennungen möglich sind, erscheint
hier zweckmäßig, da die L929 Zellen nicht an allen Zellkontaktstellen solche

2. Stand der Forschung 15
Verengungen aufweisen. Für die Separation von agglomerierten MC3T3 Zellen
sowie ähnlichen Typen sind die vorgestellten Verfahren ungeeignet, denn die
Agglomerate weisen kaum Verengungen in der Kontur auf und sind oft durch
eine große kompakte geflechtartige Struktur gekennzeichnet. Hier erscheint die
Entwicklung eines Verfahrens zur Separation zusammengewachsener Zellen
ohne Verengungen an den Zellkontaktstellen zweckmäßig.
Des Weiteren ist die Entwicklung eines Verfahrens zur automatischen Auswahl
des passenden Zelltrennungsverfahrens für die Separation verschiedener Zell-
linien sinnvoll. So kann die Software bei noch unbekannten Zelltypen anhand
definierter Kriterien das am besten geeignetste Trennverfahren selbstständig
ausführen.

3. Vorbereitungen für die Biokompatibilitätsanalyse 16
3. Vorbereitungen für die Biokompatibilitätsanalyse
Die zu prüfenden Substrate erfordern vor Durchführung der Zellanalyse zuerst
eine Oberflächenbehandlung, damit sie über eine einheitliche Oberflächenbe-
schaffenheit verfügen und so gleiche Voraussetzungen für das Zellwachstum
erfüllen. Nach der Probenaufarbeitung sollten möglichst wenige Schleif- und
Polierkratzer vorliegen, da mit steigendem Anteil an Bildartefakten die automa-
tische Zellanalyse zu ungenauen Ergebnissen führen kann.
3.1 Probenaufarbeitung
Der Arbeitsablauf bei der Probenaufarbeitung ist in Abb. 5 dargestellt.
Abbildung 5: Ablauf bei der Aufarbeitung der Proben
Die Aufbereitung der Metallproben beginnt mit dem Einbetten des Materials.
Dieser Schritt ist für den anschließenden Schleifprozess notwendig. Dazu wird
das Kalteinbettmittel „VariKwick“ verwendet, das einen Kunststoffmantel um das
Substrat bildet. Der Schleifprozess erfolgt manuell mit einer Schleifmaschine
bei 300 Umdrehungen pro Minute unter ständiger Wasserzufuhr. Es kommen,
angefangen mit der gröbsten Körnung, fünf verschiedene Feinheitsgrade der
Siliciumcarbid (SiC) Schleifscheiben zum Einsatz (Tabelle 2). Das anschließen-
de Polieren mit Hilfe einer Diamantsuspension dient zur Herstellung einer spie-
gelpolierten Oberfläche.

3. Vorbereitungen für die Biokompatibilitätsanalyse 17
Tabelle 2: Durchzuführende Schleif- und Polierschritte
Schritt Aufarbeitungsmittel Ziel
1 240er SiC Planschleifen
2 500er SiC Grobes Schleifen
3 800er SiC Mittleres Schleifen
4 1000er SiC Feines Schleifen
5 2500er SiC Sehr feines Schleifen
6 3µm Polierscheibe Polieren
Im weiteren Verlauf werden die polierten Proben aus dem Kunststoff ausgebet-
tet und nacheinander jeweils 8 Minuten in Aceton, Ethanol und dest. Wasser
von Verunreinigungen wie z.B. Fettfilmen gesäubert. Der letzte Aufbereitungs-
schritt umfasst das Autoklavieren5. Dieser Prozess sterilisiert die Proben unter
genormten Bedingungen bei 121°C und 1,2bar in einem Autoklaven. Der Sterili-
sationsvorgang dauert 25 Minuten.
3.2 Zellkultivierung
Die in-vitro Kultivierung von bestimmten Zelllinien und Geweben ist ein wesent-
licher Bestandteil der modernen Zellbiologie. Mit dieser Methode ist es möglich,
ganze Biosysteme außerhalb des Körpers (in vitro) heranzuzüchten. Die Zellen
wachsen dabei in Kulturflaschen, die unter speziellen Bedingungen (5% CO2-
Begasung, 37°C) in einem Brutschrank lagern und nach Bedarf für die jeweili-
gen Versuche zur Verfügung stehen. Da bei der in-vitro Zellkultivierung keine
Blutgefäße zur Nährstoffversorgung und kein Immunsystem für die natürliche
Abwehr vorhanden sind, erfüllt hier das Nährmedium diese Aufgabe (52; 53).
In den Kulturflaschen wachsen die Zellen je nach Typ 2-5 Tage, bis ca. 85%
des Flaschenbodens mit einem Zellrasen bedeckt sind. In diesem Fall besitzen
die Zellen eine optimale Ausgangsvoraussetzung für das Wachstum auf der
Probenoberfläche.
Um die Vitalität der Kultur aufrecht zu erhalten, müssen die Zellen in regelmä-
ßigen Abständen passagiert werden. Das Passagieren umfasst das Ablösen
der Zellen von der Kulturflasche und das verdünnte Aussäen in neue Zellkultur-
gefäße (53). Das enzymatische Ablösen der Zellen vom Flaschenboden ge-
schieht unter Zugabe einer Trypsin-Lösung und einer kurzen Einwirkzeit im
Brutschrank. Der Ablösevorgang wird durch ein Kulturmedium gestoppt und das
5 Autoklav: Ein Autoklav ist ein verschließbarer Druckbehälter. Die darin befindlichen Materialien
werden zur Sterilisation einer thermischen Behandlung unterzogen

3. Vorbereitungen für die Biokompatibilitätsanalyse 18
Gemisch in ein Zentrifugenröhrchen gegeben und zentrifugiert, so dass die Zel-
len getrennt von den flüssigen Substanzen vorliegen. Anschließend erfolgt das
Resuspendieren des entstandenen Zellpellets unter Verwendung von frischem
Nährmedium. Zur Bestimmung der Zellanzahl wird ein bestimmtes Volumen der
Lösung entnommen und mit Hilfe einer Fuchs-Rosenthal-Zählkammer6 manuell
unter dem Mikroskop ausgezählt. Der Passagiervorgang endet mit der verdünn-
ten Zugabe des resuspendierten Zell-Nährmedium-Gemisches in eine neue
Kulturflasche. Auf diese Weise bleibt die Zellkultur erhalten und kann für neue
Versuche verwendet werden. Nach einer zelltypabhängigen Anzahl an Passa-
giervorgängen ist die maximale Teilungsrate der Zellkultur erreicht, und es
müssen neue Zellen herangezogen werden.
Für den eigentlichen Versuchsansatz wird anschließend die erforderliche Zell-
zahl auf die Proben ausgesät. Experimentelle Untersuchungen haben gezeigt,
dass mit einer Startpopulation von ca. 25.000 Zellen auf biokompatiblem Mate-
rial eine für die automatische Zellanalyse gut auswertbare Zelldichte entsteht.
Substrate mit geringerer Biokompatibilität stellen aufgrund der dünneren Be-
siedlungsdichte kein Problem bei der automatischen Zellanalyse dar. Nach Zu-
gabe von jeweils 25.000 Zellen auf die Probe werden diese für drei Tage in den
Brutschrank inkubiert7. Innerhalb dieser Zeit haften die Zellen an dem Proben-
material an und beginnen, abhängig vom Toxizitätsgrad des verwendeten Sub-
strates, unterschiedlich schnell zu wachsen und sich auszubreiten.
Das Zellwachstum dauert drei Tage. Nach Absaugen des Nährmediums werden
die Proben anschließend mit einer speziellen Pufferlösung (PBS, phosphat-
gepufferte Salzlösung) gewaschen. Abschließend erfolgt die Anfärbung der Zel-
len, wobei hier die histologische May-Grünwald Färbemethode aus im nachfol-
genden Absatz näher erläuterten Gründen zum Einsatz kommt. Zu den Zellen
wird eine probenabhängige Menge an Farblösung gegeben. Die Einwirkdauer
hängt vom verwendeten Material ab. Zuletzt gibt man den Proben die gleiche
Menge an destilliertem Wasser hinzu und lässt dieses ebenso eine materialab-
hängige Zeitspanne einwirken (Tabelle 3: Materialspezifische Einwirkzeiten der
Kristallviolett und May-Grünwald Färbung).
6 Fuchs-Rosenthal-Zählkammer: Bei der Fuchs-Rosenthal-Zählkammer handelt es sich um ein
Messgerät bestehend aus einem optischen Spezialglas. Damit kann unter dem Mikroskop die
Zellanzahl in Suspension ermittelt werden. Hilfreich für den Zählvorgang sind die 16 großen
Zählkammern, die jeweils aus 16 kleinen Zählkammern bestehen. Das Volumen einer kleinen
Zählkammer beträgt 3,2µl. 7 Inkubieren: Unter inkubieren ist das heranzüchten der Zellen bei bestimmten Aussenbedin-
gungen (5% CO2-Begasung, 37°C) im Inkubator (Brutschrank) zu verstehen.

3. Vorbereitungen für die Biokompatibilitätsanalyse 19
Tabelle 3: Materialspezifische Einwirkzeiten der Kristallviolett und May-Grünwald Färbung
Probenmaterial Färbemethode Einwirkzeit Farbstoff
Einwirkzeit dest. H2O
Polystyrol Kristallviolett 9:30min 0:00min
Polystyrol May-Grünwald 4:30min 9:00min
Metall Kristallviolett 8:00min 0:00min
Metall May-Grünwald 3:30min 7:00min
3.2.1 Vergleich der Färbemethoden
Es wurden zwei verschiedene Färbeverfahren untersucht, die Kristallviolett-
(Abb. 6) und die May-Grünwald (Abb. 7) Färbemethode.
Abbildung 6: Kristallviolett gefärbte MC3T3-Zellen
Abbildung 7: May-Grünwald gefärbte MC3T3-Zellen

3. Vorbereitungen für die Biokompatibilitätsanalyse 20
Bei der Kristallviolett Färbemethode werden die Zellen im Vergleich zur May-
Grünwald Färbetechnik dunkler angefärbt. Abb. 6 zeigt, dass in den meisten
Fällen der Farbunterschied zwischen verschiedenen Zellbereichen sehr gering
ist, teilweise erscheinen kleine Zellen vollständig schwarz und können so mit
Korrosionspartikeln oder abgestorbenen Zellen verwechselt werden. Da bei den
untersuchten Fibroblasten-Zellen eine starke Variation der Morphologie vorliegt
und eine Separation der Zellen, nur auf Basis geometrischer Eigenschaften, zu
ungenau ist, muss eine farbliche Unterscheidung verschiedener Zellbereiche,
wie beispielsweise Kernbereiche und Zytoplasma, ermöglicht werden. Diese
Anforderungen erfüllt die May-Grünwald Färbung. Der Farbunterschied zwi-
schen den Kernbereichen und den übrigen Zellen ist zwar relativ gering, aber
für eine optische Differenzierung verschiedener Zellbereiche ausreichend.
Da sich ein schwacher Farbunterschied verschiedener Zellareale maßgeblich
auf die Komplexität der zu entwickelnden Analyseverfahren auswirkt, ist der
Einsatz einer multiplen Fluoreszenzfärbung für die Anfärbung verschiedener
Zellbereiche denkbar. Auf diese Weise entstehen kontrastreiche Bilder mit star-
ken Farbunterschieden verschiedener Zellbereiche, die eine automatische Ana-
lyse im Vergleich zur May-Grünwald Färbung vereinfachen. Dafür wird jedoch,
wie in Kap. 1.1 bereits erwähnt, ein teures Fluoreszenzmikroskop benötigt, das
nicht zwingend zur Ausrüstung eines Standard-Labors gehört. Ebenfalls ist die
Färbetechnik im Vergleich zur May-Grünwald Färbung aufwändiger und die
Zellpräparation nicht dauerhaft, so dass im Rahmen dieses Projektes die May-
Grünwald Färbung gewählt wurde.

4. Verwendete Hard- und Software 21
4. Verwendete Hard- und Software
Dieses Kapitel stellt neben den verwendeten Hard- und Softwarekomponenten
das Konzept der Programmarchitektur sowie den geplanten Programmablauf
vor.
4.1 Visual Studio 2010
Das Zellanalyseprogramm wurde mit der Entwicklungsumgebung Visual Studio
2010 von der Firma Microsoft entwickelt und basiert auf dem .NET-Framework8
4. Bei der Programmierung mit Visual Studio 2010 kann zwischen verschiede-
nen Sprachen gewählt werden (C++, C#, VisualBasic, F#), wobei hier die Spra-
che C# zum Einsatz kam.
4.2 Halcon Bildverarbeitungsbibliothek
Bildverarbeitungsbibliotheken finden oft bei der Entwicklung bildverarbeitender
Applikationen Verwendung. Diese bieten eine Zusammenstellung verschiede-
ner Standardfunktionen, z.B. Schwellwertmethoden, Kantenfilter, morphologi-
sche Filter, etc., so dass die Entwicklungszeit der Software im Vergleich zur
Programmierung aller benötigten Operatoren signifikant reduziert wird. Für die
Entwicklung der Zellanalysesoftware kam die Bibliothek Halcon der Firma
MVTec zum Einsatz.
8 .NET Framework: Bei dem .NET Framework handelt es sich um eine von der Fa. Microsoft
entwickelte Softwareplattform. Sie umfasst bereits eine Sammlung an Funktionen, die für die
Entwicklung eigener Programme verwendet werden können

4. Verwendete Hard- und Software 22
Abbildung 8: Architektur der Halcon Bibliothek (Quelle: MVTec GmbH)
Abb. 8 zeigt die Architektur der Halcon Software. Sie baut auf einer Bibliothek
mit ca. 1600 Bildverarbeitungsoperatoren auf, die durch so genannte Extension
Packages mit selbst erstellten Funktionen erweiterbar ist. Über verschiedene
Interfaces können Bilder von Kameras mit unterschiedlichen Kommunikations-
schnittstellen (USB, IEEE 1394, etc.) aufgenommen werden. Halcon lässt sich
in Hochsprachen wie C, C++, C#, Visual Basic usw. integrieren. Da die Zellana-
lysesoftware auf Basis des .NET-Frameworks in C# entwickelt wird, handelt es
sich somit um eine kompatible Bildverarbeitungsbibliothek. Halcon liefert mit
dem Programm HDevelop ein Tool zur Entwicklung von Bildverarbeitungsalgo-
rithmen. Durch die Visualisierung von Zwischenergebnissen eines Algorithmus
ist schnell die Eignung des Algorithmus für die Problemstellung bestimmbar. Bei
Bedarf lässt sich der entwickelte Algorithmus exportieren und in die gewünschte
Entwicklungsumgebung einbinden.
4.3 Softwarearchitektur
4.3.1 Dynamisches Einbinden von Programmmodulen
Für die Zellanalysesoftware wurde eine modulare Architektur gewählt. Auf diese
Weise bleibt die Hauptanwendung übersichtlich, denn komplexe Algorithmen
werden in ein Programmmodul, auch Assembly genannt, gekapselt. Das
Hauptprogramm bindet beim Programmstart alle in einem bestimmten Ver-
zeichnis liegenden Assemblies dynamisch ein. Unter dynamisch ist hier zu ver-
stehen, dass die Anzahl der zu ladenden Assemblies nicht festgelegt ist, son-
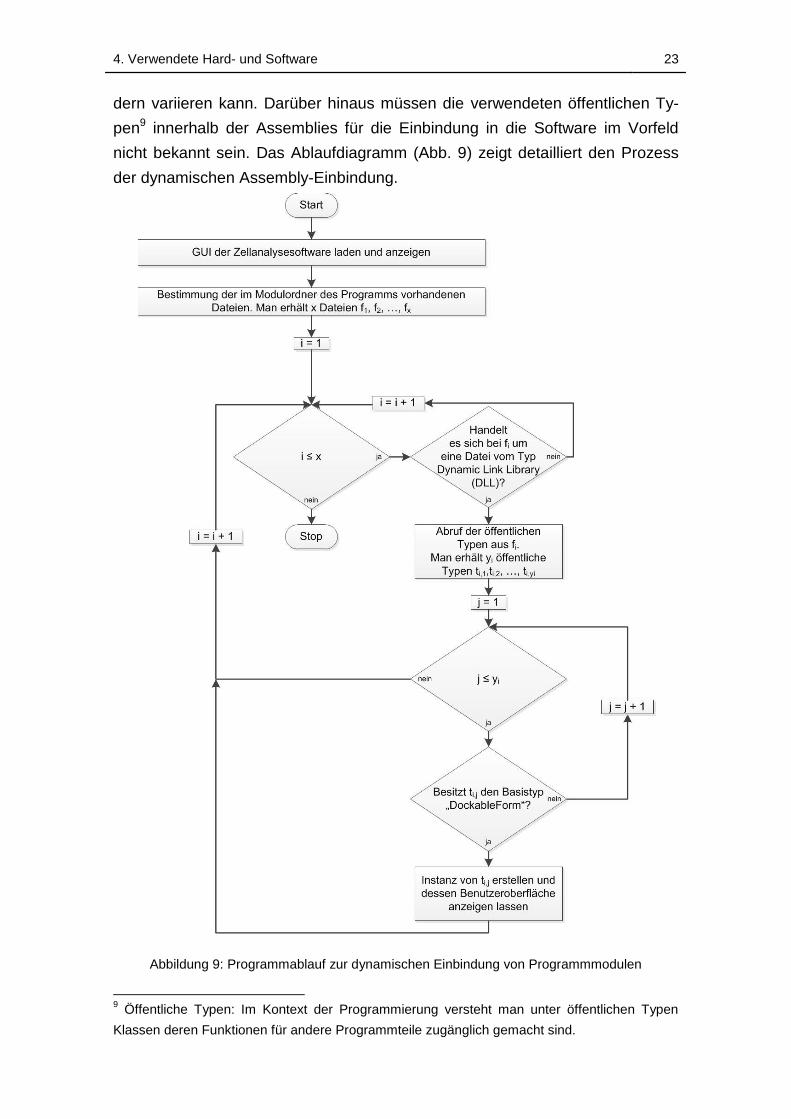
4. Verwendete Hard- und Software 23
dern variieren kann. Darüber hinaus müssen die verwendeten öffentlichen Ty-
pen9 innerhalb der Assemblies für die Einbindung in die Software im Vorfeld
nicht bekannt sein. Das Ablaufdiagramm (Abb. 9) zeigt detailliert den Prozess
der dynamischen Assembly-Einbindung.
Abbildung 9: Programmablauf zur dynamischen Einbindung von Programmmodulen
9 Öffentliche Typen: Im Kontext der Programmierung versteht man unter öffentlichen Typen
Klassen deren Funktionen für andere Programmteile zugänglich gemacht sind.

4. Verwendete Hard- und Software 24
Nach Anzeige der Benutzeroberfläche (GUI) werden alle in einem vorgegebe-
nen Verzeichnis vorhandenen Dateien ermittelt. Man erhält x Dateien f1, f2, …,
fx. Für jede Datei wird geprüft, ob ihr Datentyp einer Dynamic Link Library (DLL)
entspricht. In diesem Fall werden die in der DLL enthaltenen öffentlichen Typen
abgerufen. Es resultieren für jede Datei fi yi öffentliche Typen ti,1, ti,2, …, ti,yi mit i
= 1, 2, …, x. Entspricht der Typ ti,j mit j = 1, 2, …, yi einer DockableForm10, han-
delt es sich um eine zusätzliche Benutzeroberfläche, die nach Erstellung einer
Instanz11 von ti,j angezeigt werden soll. Auf die beschriebene Weise können nun
verschiedene Programmmodule mit oder ohne Benutzeroberflächen geladen
werden und stehen mit ihrer Funktionalität der Software zur Verfügung.
4.3.2 Grundlegender Programmablauf
Entsprechend den Anforderungen an die Software, vorgestellt in Kap 1.1, ergibt
sich der in Abb. 10 dargestellte Programmablauf.
10
DockableForm: Hierbei handelt es sich um einen Datentyp der es ermöglicht, das Programm-
fenster an das übergeordnete Steuerelement anzudocken 11
Instanz: Bei der Instanz handelt es sich um ein Objekt, dass aus einer Klasse zur Laufzeit
des Programms erstellt wird
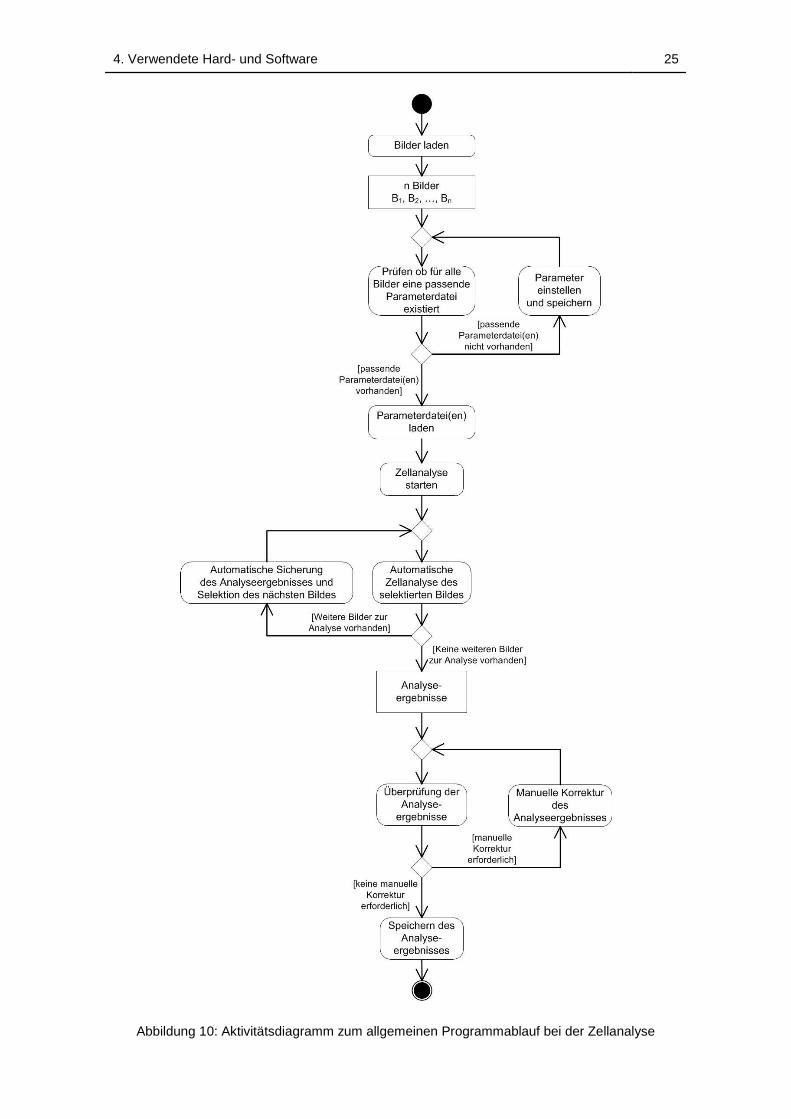
4. Verwendete Hard- und Software 25
Abbildung 10: Aktivitätsdiagramm zum allgemeinen Programmablauf bei der Zellanalyse

4. Verwendete Hard- und Software 26
4.4 Mikroskop und Kamera
Das Auflichtmikroskop Olympus Bx51M nimmt die Zellen auf metallischen
Substraten bei 100-facher Vergrößerung auf. Für Zellen auf dem Material Poly-
styrol kommt das Durchlichtmikroskop CKX41 von Olympus zum Einsatz. Die
Bilder für die automatische Zellauswertung werden mit der Farbkamera Olym-
pus XC10 mit einer Auflösung von 1376x1032 Pixel aufgenommen.

5. Segmentierung zytochemisch gefärbter Zellbereiche 27
5. Segmentierung zytochemisch gefärbter
Zellbereiche
5.1 Bildvorverarbeitung
Durch den Prozess der Probenaufbereitung, beschrieben in Kap. 3.1, können
Polierkratzer auf der Probenoberfläche entstehen. Diese können die Segmen-
tierung von Zellen negativ beeinflussen, so dass eine Eliminierung der Kratzer
sinnvoll erscheint. Da es sich bei Polierkratzern um stark lokalisierte Artefakte
handelt, ist der Einsatz der Fourier-Transformation zur Elimination der Kratzer
denkbar. Untersuchungen haben jedoch ergeben, dass neben den Kratzern
auch viele Zellausläufer eliminiert werden, da sie sich im Frequenzbereich nicht
voneinander unterscheiden. Aus diesem Grund ist die Fourier-Transformation
keine geeignete Lösung zum Entfernen der Polierkratzer.
Während des Projektverlaufs wurde der Probenaufbereitungsprozess durch die
beteiligten Biotechnologen optimiert, so dass die Anzahl der Polierkratzer stark
reduziert wurde und somit keinen weiteren negativen Einfluss auf die Zellanaly-
se ausüben. Daher wird auf ein Verfahren zur Elimination von Polierkratzern
verzichtet.
Des Weiteren können Korrosionserscheinungen des verwendeten Materials die
Segmentierung der übrigen Zellen negativ beeinflussen. Kap. 5.1.1 stellt ein
Verfahren zur Reduktion dieser Artefakte vor.
5.1.1 Reduktion von Bildartefakten durch Ausnutzung besonderer
Farbwertbeziehungen im RGB-Farbraum
Bildartefakte in Form von Korrosionserscheinungen des zugrunde liegenden
Probenmaterials können die Segmentierung der Zellbereiche stören (Abb.11).
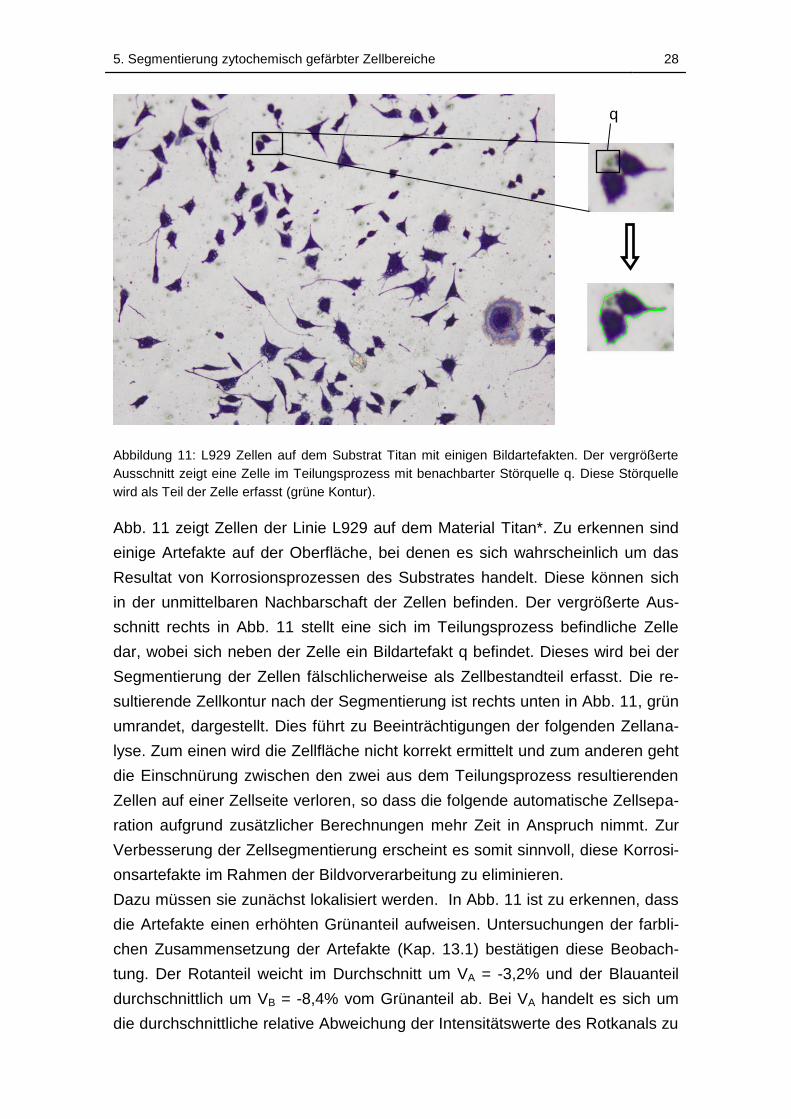
5. Segmentierung zytochemisch gefärbter Zellbereiche 28
Abbildung 11: L929 Zellen auf dem Substrat Titan mit einigen Bildartefakten. Der vergrößerte
Ausschnitt zeigt eine Zelle im Teilungsprozess mit benachbarter Störquelle q. Diese Störquelle
wird als Teil der Zelle erfasst (grüne Kontur).
Abb. 11 zeigt Zellen der Linie L929 auf dem Material Titan*. Zu erkennen sind
einige Artefakte auf der Oberfläche, bei denen es sich wahrscheinlich um das
Resultat von Korrosionsprozessen des Substrates handelt. Diese können sich
in der unmittelbaren Nachbarschaft der Zellen befinden. Der vergrößerte Aus-
schnitt rechts in Abb. 11 stellt eine sich im Teilungsprozess befindliche Zelle
dar, wobei sich neben der Zelle ein Bildartefakt q befindet. Dieses wird bei der
Segmentierung der Zellen fälschlicherweise als Zellbestandteil erfasst. Die re-
sultierende Zellkontur nach der Segmentierung ist rechts unten in Abb. 11, grün
umrandet, dargestellt. Dies führt zu Beeinträchtigungen der folgenden Zellana-
lyse. Zum einen wird die Zellfläche nicht korrekt ermittelt und zum anderen geht
die Einschnürung zwischen den zwei aus dem Teilungsprozess resultierenden
Zellen auf einer Zellseite verloren, so dass die folgende automatische Zellsepa-
ration aufgrund zusätzlicher Berechnungen mehr Zeit in Anspruch nimmt. Zur
Verbesserung der Zellsegmentierung erscheint es somit sinnvoll, diese Korrosi-
onsartefakte im Rahmen der Bildvorverarbeitung zu eliminieren.
Dazu müssen sie zunächst lokalisiert werden. In Abb. 11 ist zu erkennen, dass
die Artefakte einen erhöhten Grünanteil aufweisen. Untersuchungen der farbli-
chen Zusammensetzung der Artefakte (Kap. 13.1) bestätigen diese Beobach-
tung. Der Rotanteil weicht im Durchschnitt um VA = -3,2% und der Blauanteil
durchschnittlich um VB = -8,4% vom Grünanteil ab. Bei VA handelt es sich um
die durchschnittliche relative Abweichung der Intensitätswerte des Rotkanals zu
q

5. Segmentierung zytochemisch gefärbter Zellbereiche 29
den Intensitätswerten des Grünkanals. VB gibt die durchschnittliche relative Ab-
weichung der Intensitätswerte des Blaukanals zu den Intensitätswerten des
Grünkanals an. Hingegen ist bei der farblichen Zusammensetzung des Hinter-
grundbereiches festzustellen, dass der Rotanteil im Vergleich zum Grünanteil
mit VA = 1,4% leicht höher ausfällt. Der Blaukanal weist mit VB = 0,2% Abwei-
chung vom Grünkanal ähnliche Werte auf. Mit dieser Beobachtung kann über
eine einfache Regel die Zugehörigkeit eines Pixelfarbwertes zum Hintergrund
oder Artefakt festgelegt werden. Es wird SA = VA + VB berechnet und geprüft ob
SA ≤ -1.0% ist. In diesem Fall wird der Pixel aufgrund des dominierenden Grün-
anteils als Artefaktpixel, ansonsten als Hintergrundpixel definiert. Zur Eliminie-
rung eines Artefaktes muss der Farbwert der zugehörigen Pixel neu gesetzt
werden. Dazu wird der häufigste im Bild vorkommende Farbwert ermittelt und
die Artefaktpixel auf diesen Farbwert gesetzt. Dieser Vorgang beruht auf der
Annahme, dass der häufigste Farbwert des Bildes dem Hintergrund (Substrat)
zugeordnet werden kann.
Abb 12. (links) zeigt die Zellsegmentierung vor, Abb. 12 (rechts) die Zellseg-
mentierung nach der Bildvorverarbeitung. Die Zellfläche wird nach der Bildvor-
verarbeitung genauer bestimmt und vereinfacht die spätere Zellseparation
durch die resultierende „Hantelform“.
5.1.2 Ergebnisse
Zur Bewertung der Effizienz des vorgestellten Verfahrens wurden aus 10 Pro-
ben der Linie L929 auf dem Material Stahl* und Titan* die Anzahl der Zellen mit
und im Vergleich dazu ohne Bildvorverarbeitung bestimmt und mit der von ei-
nem Experten ermittelten Zellzahl verglichen. Im Rahmen der automatischen
Zellzählung kommen für die Zellsegmentierung das Schwellwertverfahren, be-
schrieben in Kap. 5.2.1, und für die Separation agglomerierter Zellen das Ver-
fahren, beschrieben in Kap. 6.4, zum Einsatz. Mit Hilfe des Jaccard-
Abbildung 12: Links: Segmentierter Zellbereich ohne Bildvorverarbeitung; Rechts: Segmen-
tierter Zellbereich mit Bildvorverarbeitung

5. Segmentierung zytochemisch gefärbter Zellbereiche 30
Koeffizienten (54) wird eine geeignete Methode zur Bestimmung der Segmen-
tierungsgenauigkeit vorgestellt. Dieser beschreibt den Anteil der gemeinsamen
Schnittmenge zweier Punktmengen A und B an der Gesamtmenge:
,A B
J A BA B
(Gl. 2)
Zur Veranschaulichung dient Abb. 13.
Abbildung 13: a: Zwei Objekte liegen übereinander. Der Jaccard-Koeffizient beträgt 1; b: Zwei
Objekte überlappen sich teilweise. Der Jaccard-Koeffizient beträgt 0,33; c: Zwei Objekte
besitzen keine gemeinsame Schnittmenge. Der Jaccard-Koeffizient ist 0
Abb. 13a zeigt zwei Objekte, die genau übereinander liegen. Der Jaccard-
Koeffizient beträgt 1. Dieser kann Werte zwischen 0 und 1 annehmen, wobei
ein Koeffizient von 1 bedeutet, dass die gemeinsame Schnittmenge der beiden
zu vergleichenden Objekte 100% der Gesamtfläche entspricht. Abb. 13b zeigt
zwei Objekte deren gemeinsame Schnittmenge einen Anteil von 33% zur Ge-
samtfläche annimmt (Jaccard-Koeffizient: 0,33). In Abb. 13c werden zwei Ob-
jekte gezeigt, die keine gemeinsame Schnittmenge aufweisen, der Jaccard-
Koeffizient beträgt 0.
Der Jaccard-Koeffizient wurde von 25 Zellen aus verschiedenen Probenaus-
schnitten, bei denen sich Artefakte in unmittelbarer Nachbarschaft befinden, zur
Bewertung der Segmentierungsgenauigkeit mit und ohne erfolgter Bildvorverar-

5. Segmentierung zytochemisch gefärbter Zellbereiche 31
beitung berechnet und einer Referenzsegmentierung durch einen Experten ge-
genübergestellt. Die Anzahl der untersuchten Zellen zur Berechnung des Jac-
card-Koeffizienten ist hier mit 25 Zellen sehr gering. Der Grund liegt darin, dass
nur vereinzelt Zellflächen auf dem Material Stahl* oder Titan* durch Korrosions-
erscheinungen gestört sind.
Die Referenzsegmentierung wird durch eine Laborfachkraft erstellt, indem sie
ein Zellbild in das Programm lädt und die Zellkonturen nachzeichnet. Die erstell-
ten Zellkonturen werden als Referenz abgespeichert. Es folgt die Ausführung
der automatischen Zellsegmentierung, worauf die computergenerierten Zellkon-
turen resultieren. Der Anwender lädt nun die Referenzkonturen in das Pro-
gramm und lässt den Jaccard-Koeffizienten zwischen den automatisch gene-
rierten Zellkonturen und den dazugehörigen Referenzkonturen berechnen. Aus
dieser Vorgehensweise resultieren die in Tabelle 4 gezeigten Daten.
Tabelle 4: Berechnung des Jaccard-Koeffizient für 25 ausgewählte Zellen mit naheliegenden
Artefakten, deren Segmentierungsergebnis durch die Bildvorverarbeitung manipuliert wird. Als
Segmentierungsmethode dient das Schwellwertverfahren, beschrieben in Kap. 5.2.1
Zellnummer Jaccard-Koeffizient mit
Bildvorverarbeitung Jaccard-Koeffizient ohne
Bildvorverarbeitung
1 0,81 0,75
2 0,80 0,73
3 0,87 0,80
4 0,86 0,85
5 0,83 0,80
6 0,88 0,86
7 0,85 0,80
8 0,86 0,78
9 0,88 0,82
10 0,89 0,79
11 0,86 0,81
12 0,85 0,80
13 0,81 0,78
14 0,86 0,79
15 0,80 0,74
16 0,88 0,83
17 0,85 0,76
18 0,88 0,78
19 0,87 0,83
20 0,82 0,86
21 0,88 0,79
22 0,84 0,79
23 0,81 0,82
24 0,86 0,79
25 0,89 0,83
Durchschnitt 0,85 0,80

5. Segmentierung zytochemisch gefärbter Zellbereiche 32
Ein Vergleich der Zellzählung mit und ohne Bildvorverarbeitung geht aus der nachfolgenden
Tabelle 5 hervor.
Tabelle 5: Vergleich der Zellzählung mit der vorgestellten Bildvorverarbeitung (1) und ohne
Bildvorverarbeitung (2) zur Referenzzählung bei Verwendung des Materials Stahl* und Titan*.
Eingesetzt wird hier das Segmentierungsverfahren, beschrieben in Kap. 5.2.1 und das Separa-
tionsverfahren, beschrieben in Kap. 6.4 mit vorgeschalteter CBS (Kap. 6.1); AZ: Automatische
Zellzählung; FP: Falsch-positiv (zuviel) Detektion; FN: Falsch-negativ (vergessen) Detektion;
KD: Korrekte Detektion (AZ-FP); REF: Referenzzählung; RFP: Relative Falsch-positiv Detekti-
on (FP / REF * 100); RFN: Relative falsch-negativ Detektion (FN / REF * 100); ERR: Relativer
Fehler (((REF – AZ) / REF) * 100)
Probe AZ 1
FP 1
FN 1
KD 1
AZ 2
FP 2
FN 2
KD 2
REF RFP
1 (%)
RFN 1
(%)
RFP 2
(%)
RFN 2
(%)
ERR1
(%)
ERR 2
(%)
Probe 1 104 0 1 104 104 0 1 104 105 0,0 1,0 0,0 1,0 1,0 1,0
Probe 2 82 1 2 81 82 1 2 81 83 1,2 2,4 1,2 2,4 1,2 1,2
Probe 3 93 1 1 92 92 0 1 92 93 1,1 1,1 0,0 1,1 0,0 1,1
Probe 4 91 3 3 88 92 4 3 88 91 3,3 3,3 4,4 3,3 0,0 1,1
Probe 5 99 1 1 98 101 3 1 98 99 1,0 1,0 3,0 1,0 0,0 2,0
Probe 6 91 1 0 90 91 1 0 90 90 1,1 0,0 1,1 0,0 1,1 1,1
Probe 7 92 3 0 89 91 2 0 89 89 3,4 0,0 2,2 0,0 3,4 2,2
Probe 8 73 2 4 71 74 1 2 73 75 2,7 5,3 1,3 2,7 2,7 1,3
Probe 9 61 1 2 60 61 1 2 60 62 1,6 3,2 1,6 3,2 1,6 1,6
Probe 10 94 0 2 94 93 0 3 93 96 0,0 2,1 0,0 3,1 2,1 3,1
Probe 11 81 1 3 80 84 1 0 83 83 1,2 3,6 1,2 0,0 2,4 1,2
Probe 12 100 0 1 100 99 0 2 99 101 0,0 1,0 0,0 2,0 1,0 2,0
Probe 13 77 1 0 76 77 2 1 75 76 1,3 0,0 2,6 1,3 1,3 1,3
Probe 14 89 2 2 87 91 3 1 88 89 2,2 2,2 3,4 1,1 0,0 2,2
Probe 15 91 3 6 88 91 0 3 91 94 3,2 6,4 0,0 3,2 3,2 3,2
Probe 16 92 0 1 92 92 1 2 91 93 0,0 1,1 1,1 2,2 1,1 1,1
Probe 17 102 2 3 100 105 2 0 103 103 1,9 2,9 1,9 0,0 1,0 1,9
Probe 18 75 0 2 75 75 2 4 73 77 0,0 2,6 2,6 5,2 2,6 2,6
Probe 19 81 1 0 80 82 3 1 79 80 1,3 0,0 3,8 1,3 1,3 2,5
Probe 20 88 0 0 88 88 0 0 88 88 0,0 0,0 0,0 0,0 0,0 0,0
Probe 21 72 0 0 72 72 1 1 71 72 0,0 0,0 1,4 1,4 0,0 0,0
Probe 22 66 2 5 64 67 1 3 66 69 2,9 7,2 1,4 4,3 4,3 2,9
Probe 23 95 1 2 94 96 0 0 96 96 1,0 2,1 0,0 0,0 1,0 0,0
Probe 24 92 0 0 92 93 2 1 91 92 0,0 0,0 2,2 1,1 0,0 1,1
Probe 25 86 1 0 85 87 2 0 85 85 1,2 0,0 2,4 0,0 1,2 2,4
Ø Fehler 1,3 1,6
Tabelle 5 zeigt eine minimale Verbesserung der automatischen Zellzahlbe-
stimmung durch Anwendung der beschriebenen Bildvorverarbeitung. Mit akti-
vierter Bildvorverarbeitung sinkt die durchschnittliche Abweichung der automati-
schen Zellzählung von 1,6% auf 1,3% im Vergleich zur Referenzzählung. Der
Grund liegt darin, dass die in Abb. 11 gezeigten Korrosionserscheinungen in
unmittelbarer Nachbarschaft einer Zelle zum einen nicht häufig auftreten und
zum anderen wenig Einfluss auf die Separation zusammenhängender Zellen
besitzen. Die Qualität der Segmentierung von Zellen mit benachbarten Korrosi-
onsartefakten wird unter Verwendung der Bildvorverarbeitung verbessert, der
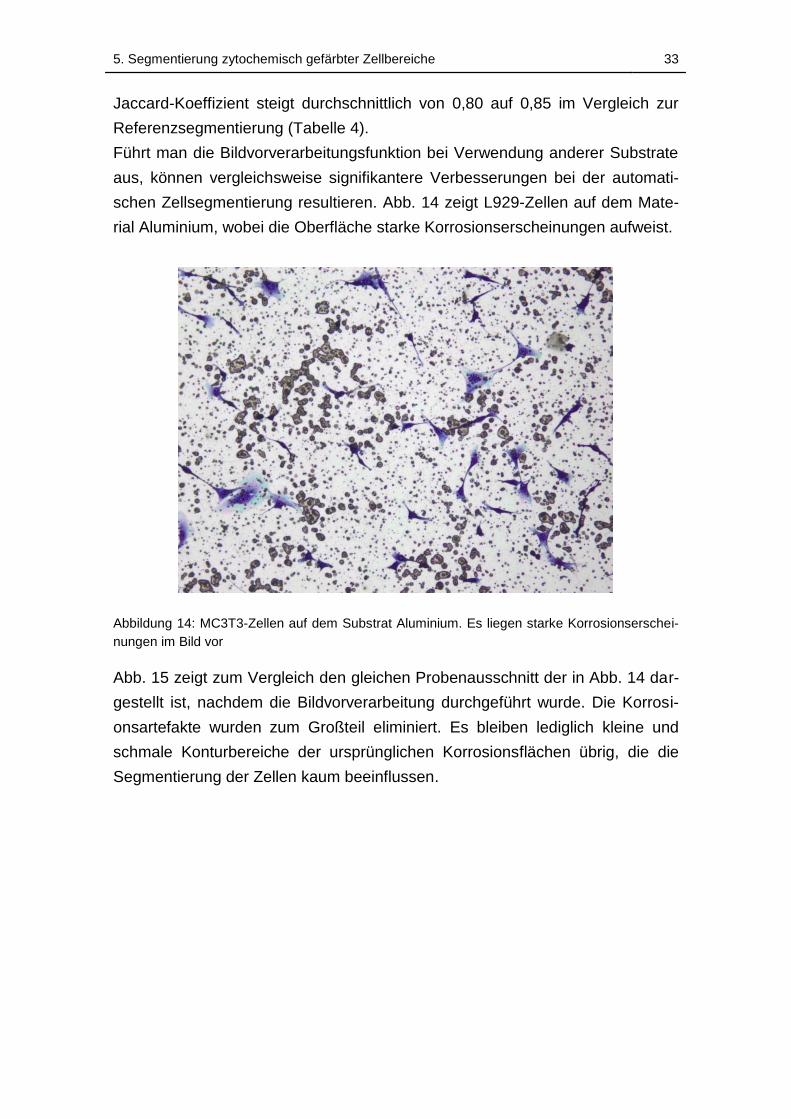
5. Segmentierung zytochemisch gefärbter Zellbereiche 33
Jaccard-Koeffizient steigt durchschnittlich von 0,80 auf 0,85 im Vergleich zur
Referenzsegmentierung (Tabelle 4).
Führt man die Bildvorverarbeitungsfunktion bei Verwendung anderer Substrate
aus, können vergleichsweise signifikantere Verbesserungen bei der automati-
schen Zellsegmentierung resultieren. Abb. 14 zeigt L929-Zellen auf dem Mate-
rial Aluminium, wobei die Oberfläche starke Korrosionserscheinungen aufweist.
Abbildung 14: MC3T3-Zellen auf dem Substrat Aluminium. Es liegen starke Korrosionserschei-
nungen im Bild vor
Abb. 15 zeigt zum Vergleich den gleichen Probenausschnitt der in Abb. 14 dar-
gestellt ist, nachdem die Bildvorverarbeitung durchgeführt wurde. Die Korrosi-
onsartefakte wurden zum Großteil eliminiert. Es bleiben lediglich kleine und
schmale Konturbereiche der ursprünglichen Korrosionsflächen übrig, die die
Segmentierung der Zellen kaum beeinflussen.

5. Segmentierung zytochemisch gefärbter Zellbereiche 34
Abbildung 15: Ergebnis der Bildvorverarbeitungsmethode nach Anwendung auf den in Abb. 14
dargestellten MC3T3-Probenausschnitt
Der Vergleich der automatisch ermittelten Zellzahl mit und ohne Bildvorverar-
beitung mit der Referenzzählung zeigt, dass die Bildvorverarbeitung in diesem
Fall zu einer deutlichen Verbesserung der automatischen Zellzählgenauigkeit
beiträgt (Tabelle 6).
Tabelle 6: Vergleich der Zellzählung mit (1) der vorgestellten Bildvorverarbeitung und ohne (2)
Bildvorverarbeitung zur Referenzzählung bei Verwendung des Materials Aluminium. Verwen-
dung findet hier das Segmentierungsverfahren, beschrieben in Kap. 5.2.1 und das Separations-
verfahren, beschrieben in Kap. 6.4; Zeichenerklärung siehe Tabelle 5
Probe AZ 1
FP 1
FN 1
KD 1
ACC 2
FP 2
FN 2
KD 2
REF RFP
1 (%)
RFN 1
(%)
RFP 2
(%)
RFN 2
(%)
ERR 1
(%)
ERR 2
(%)
Probe 1 61 9 2 52 203 151 2 52 54 16,7 3,7 279,6 3,7 13,0 275,9
Probe 2 72 10 5 62 161 99 5 62 67 14,9 7,5 147,8 7,5 7,5 140,3
Probe 3 46 10 4 36 117 81 4 36 40 25,0 10,0 202,5 10,0 15,0 192,5
Probe 4 63 7 5 56 146 90 5 56 61 11,5 8,2 147,5 8,2 3,3 139,3
Probe 5 64 26 2 38 177 139 2 38 40 65,0 5,0 347,5 5,0 60,0 342,5
Probe 6 94 46 6 48 176 128 6 48 54 85,2 11,1 237,0 11,1 74,1 225,9
Probe 7 34 7 3 27 172 145 3 27 30 23,3 10,0 483,3 10,0 13,3 473,3
Probe 8 85 32 8 53 133 80 8 53 61 52,5 13,1 131,1 13,1 39,3 118,0
Probe 9 86 35 4 51 180 129 4 51 55 63,6 7,3 234,5 7,3 56,4 227,3
Probe 10 70 7 4 63 156 93 4 63 67 10,4 6,0 138,8 6,0 4,5 132,8
Probe 11 68 7 2 61 163 105 5 58 63 11,1 3,2 166,7 7,9 7,9 158,7
Probe 12 77 10 3 67 148 80 2 68 70 14,3 4,3 114,3 2,9 10,0 111,4
Probe 13 52 5 1 47 173 129 4 44 48 10,4 2,1 268,8 8,3 8,3 260,4
Probe 14 65 13 3 52 121 72 6 49 55 23,6 5,5 130,9 10,9 18,2 120,0
Probe 15 58 10 4 48 139 90 3 49 52 19,2 7,7 173,1 5,8 11,5 167,3
Probe 16 80 8 5 72 173 101 5 72 77 10,4 6,5 131,2 6,5 3,9 124,7
Probe 17 68 20 2 48 162 114 2 48 50 40,0 4,0 228,0 4,0 36,0 224,0
Probe 18 44 17 4 27 183 160 8 23 31 54,8 12,9 516,1 25,8 41,9 490,3
Probe 19 40 9 6 31 158 125 4 33 37 24,3 16,2 337,8 10,8 8,1 327,0

5. Segmentierung zytochemisch gefärbter Zellbereiche 35
Probe 20 51 11 4 40 126 85 3 41 44 25,0 9,1 193,2 6,8 15,9 186,4
Probe 21 55 6 2 49 184 139 6 45 51 11,8 3,9 272,5 11,8 7,8 260,8
Probe 22 49 8 5 41 162 119 3 43 46 17,4 10,9 258,7 6,5 6,5 252,2
Probe 23 62 10 4 52 190 137 3 53 56 17,9 7,1 244,6 5,4 10,7 239,3
Probe 24 76 15 2 61 183 124 4 59 63 23,8 3,2 196,8 6,3 20,6 190,5
Probe 25 47 7 3 40 181 140 2 41 43 16,3 7,0 325,6 4,7 9,3 320,9
Ø Fehler 20,1 228,1
Tabelle 6 zeigt, dass sich bei Verwendung der Bildvorverarbeitung die automa-
tische Zellzahlbestimmung auf dem Substrat Aluminium von 228,1% auf 20,1%
Abweichung zur Referenzzählung reduziert. Die sehr hohe Abweichung ohne
Verwendung der Bildvorverarbeitung kommt dadurch zustande, dass viele Kor-
rosionsbereiche irrtümlich als Zellflächen segmentiert werden.
5.2 Untersuchung verschiedener Verfahren zur Segmentierung der
Zellbereiche
Dieses Kapitel stellt drei verschiedene Segmentierungsverfahren (Kap. 5.2.1 bis
Kap. 5.2.3) vor, mit denen die untersuchten Zelllinien auf den Materialien Stahl*,
Titan* und Polystyrol bei verschiedenen Mikroskopkontrasten detektierbar sind.
Im Anschluss werden in Kap. 5.3 die Vor- und Nachteile der vorgestellten Ver-
fahren diskutiert und begründet, warum andere in der Literatur bekannte Ver-
fahren hier nicht zum Einsatz kommen.
5.2.1 Schwellwertverfahren
Die Schwellwertmethode ist ein Verfahren zur Segmentierung von Objektberei-
chen im Bild, die sich aufgrund ihres Intensitätswertes vom Hintergrund unter-
scheiden. Allgemein ist für die Schwellwertmethode eine obere und untere
Schwellwertgrenze SO und SU im Intervall [0, 1, ..., 255] festzulegen. Pixel die
einen Grauwert ≥ SU und ≤ SO aufweisen werden für die weitere Verarbeitung
übernommen, die restlichen Pixel ignoriert.
Durch Verwendung einer zytochemischen Färbung der Zellen resultiert in mik-
roskopischen Zellaufnahmen ein ausreichender Kontrast zwischen den Zellen
und dem verwendeten Substrat zur Zellsegmentierung mit Hilfe eines Schwell-
wertverfahrens.

5. Segmentierung zytochemisch gefärbter Zellbereiche 36
Abbildung 16: Verschiedene Zellbereiche
Da die Zellen je nach Vitalitätszustand unterschiedlich stark ausgeprägte Endo-
und Ektoplasmabereiche besitzen, die jeweils durch unterschiedliche Farben
repräsentiert werden (Abb. 16), ist eine getrennte Schwellwertoperation im Rot-,
Grün- und Blaukanal des Bildes sinnvoll. Für die Linien L929 und MC3T3 haben
sich für die Materialien Stahl* und Titan* die in Tabelle 7 enthaltenen Schwell-
werte als geeignet herausgestellt. Zur Bestimmung geeigneter Schwellwerte
zeichnet ein Experte zunächst 50 Konturen verschiedener Zellen in das Bild.
Dieselben Zellen werden daraufhin automatisch mit den Start-Schwellwerten TR
= TG = TB = 128 segmentiert. Mit Hilfe des Jaccard-Koeffizienten kann die Ähn-
lichkeit der automatisch generierten Zellfläche mit der Referenzzellfläche ge-
messen werden. Die automatische Zellsegmentierung erfolgt nun iterativ für
verschiedene Schwellwerte wobei schließlich die Schwellwerte mit dem größten
Jaccard-Koeffizienten verwendet werden.
Tabelle 7: Schwellwerte zur Segmentierung der L929- und MC3T3-Zellbereiche bei unter-
schiedlichen Materialien; SO,R: Oberer Schwellwert des Rotkanals; SU,R: Unterer Schwellwert
des Rotkanals; SO,G: Oberer Schwellwert des Grünkanals; SU,G: Unterer Schwellwert des
Grünkanals; SO,B: Oberer Schwellwert des Blaukanals; SU,B: Unterer Schwellwert des Blauka-
nals
Proben- material
SO,R SU,R SO,G SU,G SO,B SU,B
L929 Stahl* 210 0 210 0 215 0
Titan* 190 0 190 0 190 0
Polystyrol 175 0 175 0 180 0
MC3T3
Stahl* 210 0 210 0 215 0
Titan* 185 0 185 0 190 0
Polystyrol 170 0 170 0 180 0
Äußeres Endoplasma
Zellkernbereich (Endoplasma)
Ektoplasma

5. Segmentierung zytochemisch gefärbter Zellbereiche 37
Die drei resultierenden Binärbilder BR, BG und BB werden dann zu einem Bild
BGes = R G BB B B vereinigt und einem Labeling12 unterzogen, damit nicht
zusammenhängende Zellflächen individuell auswertbar sind. Die segmentierten
Zellbereiche müssen eine Mindestfläche AMin aufweisen, andernfalls werden sie
von der weiteren Verarbeitung ausgeschlossen. Zur Bestimmung der minimalen
Zellfläche AMin des Typs L929 wurden die Zellen in 50 Bildern automatisch
segmentiert und ihre Zellfläche berechnet. Anschließend erfolgt eine manuelle
Prüfung des Ergebnisses daraufhin ob es sich bei den segmentierten Bereichen
um Zellen oder Artefakte von der Substratoberfläche handelt. Der kleinste seg-
mentierte Zellbereich weist eine Fläche von CMin = 276 Pixel auf und das größte
segmentierte Bildartefakt besitzt eine Fläche von EMax = 102 pixel. AMin ent-
spricht dem Mittelwert des kleinsten Zellbereiches und des größten Artefaktbe-
reiches:
189
2
Min MaxMin
C EA pixel
Eine Zelle kann mehrere eingeschlossene Bereiche des zugrundeliegenden
Materials besitzen (Abb. 17).
Abbildung 17: Links: Zellcluster ohne Detektion der eingeschlossenen Titan*-Bereiche. Rechts:
Die im Zellbereich erfassten Titan*-Flächen (orange markiert)
Abb. 17 zeigt einen segmentierten Zellcluster, der mehrere nicht zusammen-
hängende Bereiche des Materials Titan* enthält, hier durch die orangene Kontur
gekennzeichnet. Diese Bereiche müssen ebenfalls segmentiert werden, da sie
die anschließende Zellseparation vereinfachen und die Genauigkeit der Zellflä-
chenbestimmung erhöhen. Dazu wird innerhalb des bereits segmentierten Zell-
12
Labeling: Nicht zusammenhängende segmentierte Bildbereiche werden gelabelt, damit sie für
die weitere Verarbeitung individuell erfasst werden können. Das Label kann z.B. ein Grauwert
oder ein nummerischer Wert sein.
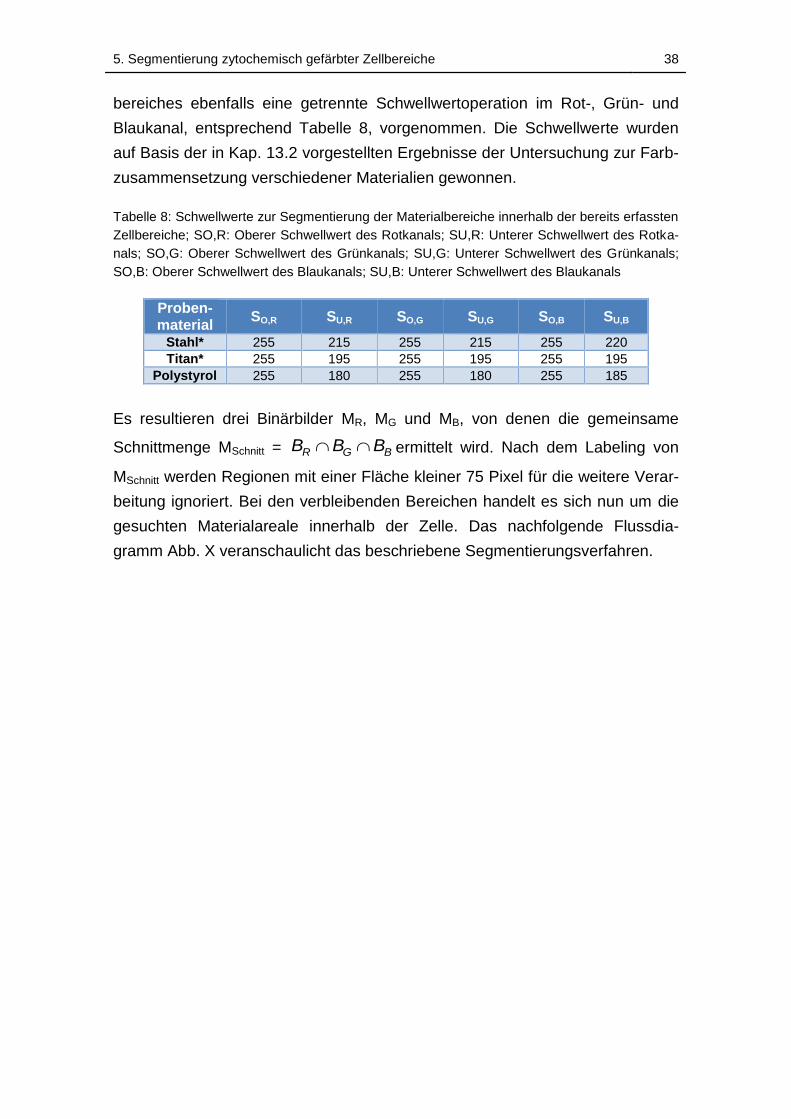
5. Segmentierung zytochemisch gefärbter Zellbereiche 38
bereiches ebenfalls eine getrennte Schwellwertoperation im Rot-, Grün- und
Blaukanal, entsprechend Tabelle 8, vorgenommen. Die Schwellwerte wurden
auf Basis der in Kap. 13.2 vorgestellten Ergebnisse der Untersuchung zur Farb-
zusammensetzung verschiedener Materialien gewonnen.
Tabelle 8: Schwellwerte zur Segmentierung der Materialbereiche innerhalb der bereits erfassten
Zellbereiche; SO,R: Oberer Schwellwert des Rotkanals; SU,R: Unterer Schwellwert des Rotka-
nals; SO,G: Oberer Schwellwert des Grünkanals; SU,G: Unterer Schwellwert des Grünkanals;
SO,B: Oberer Schwellwert des Blaukanals; SU,B: Unterer Schwellwert des Blaukanals
Proben- material
SO,R SU,R SO,G SU,G SO,B SU,B
Stahl* 255 215 255 215 255 220
Titan* 255 195 255 195 255 195
Polystyrol 255 180 255 180 255 185
Es resultieren drei Binärbilder MR, MG und MB, von denen die gemeinsame
Schnittmenge MSchnitt = R G BB B B ermittelt wird. Nach dem Labeling von
MSchnitt werden Regionen mit einer Fläche kleiner 75 Pixel für die weitere Verar-
beitung ignoriert. Bei den verbleibenden Bereichen handelt es sich nun um die
gesuchten Materialareale innerhalb der Zelle. Das nachfolgende Flussdia-
gramm Abb. X veranschaulicht das beschriebene Segmentierungsverfahren.

5. Segmentierung zytochemisch gefärbter Zellbereiche 39
Abbildung 18: Schematischer Ablauf der Zellsegmentierung mit Hilfe der Schwellwertmethode
für alle Farbkanäle eines RGB-Bildes
5.2.1.1 Ergebnisse
Zur Beurteilung der Qualität des in Kap. 5.2.1 vorgestellten Segmentierungsver-
fahrens ist eine Referenzsegmentierung notwendig. Dazu sind durch einen Ex-
perten pro Zelllinie und Mikroskopkontrastart jeweils 50 Zellen manuell segmen-
tiert worden. Mit Hilfe des Jaccard-Koeffizienten kann nun eine Aussage über
die durchschnittliche Abweichung der automatisch generierten Zellkonturen zu
den Referenzkonturen erfolgen. Die kompletten Daten sind in Kap. 13.3 aufge-

5. Segmentierung zytochemisch gefärbter Zellbereiche 40
listet, Tabelle 9 beinhaltet den Durchschnittswert des Jaccard-Koeffizienten auf
Basis von 50 Zellen jeweils für einen Mikroskopkontrast.
Tabelle 9: Bewertung der Qualität des Schwellwertverfahrens für unterschiedliche Zelllinien,
Materialien und Mikroskopkontraste mit Hilfe des Jaccard-Koeffizienten; A-HF: Auflicht- Hellfeld;
A-DF: Auflicht- Dunkelfeld; D-HF: Durchlicht- Hellfeld; D-PK: Durchlicht- Phasenkontrast
Materi-
al L929 MC3T3
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
Stahl*
/Titan* 0,89 0,85 --- --- 0,87 0,90 --- ---
Poly-
styrol --- --- 0,87 0,60 --- --- 0,76 0,53
Grundsätzlich wird für Bildaufnahmen von metallischen Proben (Stahl*, Titan*)
ein Auflichtmikroskop und für transparente Materialien (Polystyrol) ein Durch-
lichtmikroskop eingesetzt.
Bei Anwendung des schwellwertbasierten Verfahrens resultieren für Zellen des
Typs L929 und MC3T3 auf dem Material Stahl*/Titan* ähnliche Ergebnisse
(0,85 – 0,9) für die beiden Auflichtkontrastarten.
Für die Linien L929 und MC3T3 ist beim Einsatz von Polystyrolproben die
schwellwertbasierte Methode lediglich für den Durchlicht-Hellfeldkontrast geeig-
net. Hier unterscheiden sich die Ergebnisse bzgl. des Jaccard-Koeffizienten für
die beiden Zelllinien. Der Grund liegt darin, dass die im Vergleich zur Linie L929
flächiger gewachsenen MC3T3 Zellen im Randbereich kontrastärmer erschei-
nen (Abb. 18) und somit die Differenzierung des Hintergrundes schwieriger ist.
Abbildung 19: MC3T3 Zellen auf dem Material Stahl unter Verwendung des Hellfeldkontrastes
beim Durchlichtmikroskop. Die große Zelle mittig im Bild weist einen teilweise sehr schwachen
Kontrastunterschied zum Substrat auf. Dies beeinflusst die Segmentierungsgenauigkeit.
Unter Verwendung des Durchlicht-Phasenkontrastes resultieren keine verwert-
baren Ergebnisse da sowohl Zellen als auch das Substrat teilweise die gleiche
Farbgebung aufweisen und durch global einstellbare Schwellwerte somit nicht

5. Segmentierung zytochemisch gefärbter Zellbereiche 41
mehr hinreichend genau zwischen Vorder- und Hintergrund unterschieden wer-
den kann.
5.2.2 Histogrammbasierte Methode
Die histogrammbasierte Methode ist ein Verfahren, dass Objekte auf Basis des
Bildhistogramms segmentiert. Zunächst erfolgt die Bestimmung der oberen und
unteren Schwellwertgrenze SO und SU zur Segmentierung des Substrates. Da-
für ist die Berechnung des relativen Histogramms HR des Bildes notwendig. Da-
raufhin wird iterativ mit einem Fenster der Breite m über HR gefahren und in
jedem Iterationsschritt die Summe Si des relativen Bildanteils innerhalb des
Fensters berechnet,
( )i m
i R
j i
S H j
mit i = 0, 1, …, 255-m. (Gl. 3)
Die Untersuchung zur Farbverteilung unterschiedlicher Materialien (Kap. 13.2)
zeigt, dass bei Einsatz eines Durchlichtmikroskopes im Hellfeldkontrast das zu
untersuchende Substrat (Polystyrol) eine homogene Farbverteilung aufweist.
Da hingegen die verschiedenen Zellbereiche, z.B. Ekto- oder Endoplasma,
durch unterschiedliche Farben repräsentiert werden, erscheint in diesem Fall
die Segmentierung des Substrates sinnvoll, denn mit dem Segmentierungser-
gebnis kann anschließend durch Differenzbildung vom ursprünglichen Bild auf
die Zellbereiche geschlossen werden.
Zu Beginn erfolgt die Bestimmung der oberen und unteren Schwellwertgrenze
SO und SU zur Segmentierung des Substrates. Dafür ist die Berechnung des
relativen Histogramms HR des Bildes notwendig. Dann wird iterativ mit einem
Fenster der Breite m über HR gefahren und in jedem Iterationsschritt die Sum-
me Si des relativen Bildanteils innerhalb des Fensters berechnet,
( )i m
i R
j i
S H j
mit i = 0, 1, …, 255-m. (Gl. 3)
Zu Beginn wird eine Hilfsvariable SMax = 0 initialisiert. Nach jeder Iteration wird
geprüft, ob Si > SMax. In diesem Fall erhält SMax den Wert von Si und der obere
Schwellwert SO den Wert SO = j. Die untere Segmentierungsschwelle SU wird
mit SU = SO – m festgelegt. So wird genau der Histogrammausschnitt mit der
Breite m ermittelt, in dem sich die meisten Pixel befinden. Das nachfolgende
Ablaufdiagramm beschreibt die automatische Berechnung der Schwellwert-
grenzen SO und SU (Abb. 19).

5. Segmentierung zytochemisch gefärbter Zellbereiche 42
Abbildung 20: Ablaufdiagramm zur automatischen Bestimmung der Schwellwertgrenzen SO und
SU
Mit Hilfe der ermittelten Schwellwerte SO und SU erfolgt nun die Segmentierung
des Hintergrundbereiches, man erhält das Binärbild T1. Eine weitere Schwell-
wertoperation mit der unteren Schwelle 0 und der oberen Schwelle 255, ange-
wendet auf das Ausgangsbild, liefert ein Binärbild T2 des gesamten Bildberei-
ches. Daraufhin wird der Hintergrundbereich T1 vom gesamten Bildbereich T2
subtrahiert, man erhält TD = T2 \ T1. TD wird nun einem Closing (59) und einem
Labeling unterzogen, man erhält x segmentierte Objekte O1, O2,..., Ox.
Histogrammbasierte Methode zur Detektion von Zellbereichen:
Die Untersuchung zur Farbverteilung unterschiedlicher Materialien (Kap. 13.2)
zeigt, dass bei Einsatz eines Durchlichtmikroskopes im Hellfeldkontrast das zu
untersuchende Substrat (Polystyrol) eine homogene Farbverteilung aufweist.
Da hingegen die verschiedenen Zellbereiche, z.B. Ekto- oder Endoplasma,
durch unterschiedliche Farben repräsentiert werden, erscheint in diesem Fall
die Segmentierung des Substrates sinnvoll, denn mit dem Segmentierungser-
gebnis kann anschließend durch Differenzbildung vom ursprünglichen Bild auf
die Zellbereiche geschlossen werden. Die segmentierten Zellkandidaten müs-
sen eine Mindestfläche AMin aufweisen, andernfalls werden sie verworfen, wobei
für die im Rahmen des Projektes untersuchten Zelllinien, wie bereits in Kap.
5.2.1 erläutert, AMin = 189 Pixel eine gute Wahl ist. Nach Ausschluss der Objek-

5. Segmentierung zytochemisch gefärbter Zellbereiche 43
te mit einer Fläche < AMin resultieren y Zellbereiche Z1, Z2, …, Zy. Das Ablaufdi-
agramm (Abb. 20) fasst die beschriebene Segmentierung der Zellen zusam-
men.
Abbildung 21: Ablauf der Zellsegmentierung nach der histogrammbasierten Methode
5.2.2.1 Ergebnisse
Die Beurteilung der Qualität des in Kap. 5.2.2 vorgestellten Segmentierungsver-
fahrens folgt der Vorgehensweise in Kap. 5.2.1.1. Die vollständigen Daten sind
in Kap. 13.3 aufgelistet. Tabelle 10 zeigt den Durchschnittswert des Jaccard-
Koeffizienten.
Tabelle 10: Bewertung der Qualität des histogrammbasierten Schwellwertverfahrens für unter-
schiedliche Zelllinien und Mikroskopkontraste mit Hilfe des Jaccard-Koeffizienten; A-HF: Auf-
licht-Hellfeld; A-DF: Auflicht-Dunkelfeld; D-HF: Durchlicht-Hellfeld; D-PK: Durchlicht-
Phasenkontrast
Materi-
al L929 MC3T3
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
Stahl*
/Titan* 0,86 0,80 --- --- 0,86 0,85 --- ---
Poly-
styrol --- --- 0,87 0,77 --- --- 0,82 0,82

5. Segmentierung zytochemisch gefärbter Zellbereiche 44
Analog zum Kap. 5.2.1.1 gilt auch hier, dass grundsätzlich für Bildaufnahmen
von metallischen Proben (Stahl*, Titan*) ein Auflichtmikroskop und für transpa-
rente Materialien (Polystyrol) ein Durchlichtmikroskop eingesetzt wird.
Unter Einsatz der histogrammbasierten Schwellwertmethode resultieren für alle
Kontrastarten bei beiden Zelllinien verwertbare Ergebnisse.
5.2.3 Histogram-Backprojection-Algorithmus
Der Histogram-Backprojection-Algorithmus dient zur Lokalisierung von Objekten
in einem Bild auf Basis ihrer Farbwerte (26). Dazu muss im Vorfeld ein Farb-
template erzeugt werden, dass die Farben des zu suchenden Objektes beinhal-
tet. Das Verfahren kann mit Hilfe der Farbtemplates auf beliebige Objekte an-
gewandt werden. Zur Beschleunigung der Methode wird über einen Parameter
der Grad der Unterabtastung der Farbkanäle vorgenommen.
Abb. 21 zeigt, dass die Zellen einen farblichen Kontrast zum Hintergrundbereich
aufweisen. Hier bietet sich mit dem Histogram-Backprojection-Verfahren eine
geeignete Segmentierungsmethode an.
Abbildung 22: L929-Zellen auf dem Substrat Stahl* bei 100-facher Vergrößerung
Das Ergebnis des Histogram-Backprojection-Algorithmus (HB) ist mit Hilfe des
im Vorfeld erzeugten Templates (Abb. 22) in Abb. 23 dargestellt.

5. Segmentierung zytochemisch gefärbter Zellbereiche 45
Abbildung 23: Farbtemplate für den HB Algorithmus
Abbildung 24: Ergebnis des HB mit Hilfe des Farbtemplates aus Abb. 22
Für die Durchführung des HB-Algorithmus ist die Einstellung definierter
Parameter notwendig. Um eine Übersegmentierung der Zellen zu verhindern,
wird eine Mindestfläche ACMin für die segmentierten Zellbereiche festgelegt.
Dabei ist für die Zelltypen L929 und MC3T3, wie bereits in Kap. 5.2.1 erläutert,
ACMin = 189 Pixel eine gute Wahl. Wie in (55) beschrieben, wird aus
Geschwindigkeitsgründen meist eine Unterabtastung der Farbkanäle
vorgenommen. In diesem Fall ist mit c = 16 Farben eine optimale
Unterabtastung für die untersuchten Zelllinien eingestellt.
Im weiteren Verlauf wird das in Abb. 23 gezeigte Ergebnis mit der standard
Bildverarbeitungsoperation Closing (56) bearbeitet, um kleine
Segmentierungslücken zu schließen. Daraufhin erfolgt ein Labling, damit
zusammenhängende Flächen individuell auswertbar sind.

5. Segmentierung zytochemisch gefärbter Zellbereiche 46
5.2.3.1 Ergebnisse
Die Vorgehensweise zur Beurteilung der Qualität des in Kap. 5.2.3 vorgestellten
Segmentierungsverfahrens erfolgt analog zu Kap. 5.2.1.1 und Kap. 5.2.2.1. Die
vollständigen Daten sind in Kap. 13.3 aufgelistet, Tabelle 11 beinhaltet auch
hier den Durchschnittswert des Jaccard-Koeffizienten.
Tabelle 11: Bewertung der Qualität des Histogram-Backprojection-Verfahrens für unterschiedli-
che Zelllinien und Mikroskopkontraste mit Hilfe des Jaccard-Koeffizienten; A-HF: Auflicht-
Hellfeld; A-DF: Auflicht-Dunkelfeld; D-HF: Durchlicht-Hellfeld; D-PK: Durchlicht-Phasenkontrast
Materi-al
L929 MC3T3
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
Stahl* /Titan*
0,78 0,80 --- --- 0,85 0,86 --- ---
Polystyrol --- --- 0,81 0,63 --- --- 0,78 0,58
Analog zum Kap. 5.2.2.1 gilt auch hier dass grundsätzlich für Bildaufnahmen
von metallischen Proben (Stahl*, Titan*) ein Auflichtmikroskop und für transpa-
rente Materialien (Polystyrol ) ein Durchlichtmikroskop eingesetzt wird.
Im Auflicht-Hellfeld (A-HF) und Auflicht-Dunkelfeld (A-DF) resultieren für beide
Zelltypen ähnliche Ergebnisse.
Bei Verwendung von Polystyrolproben resultieren auch für die Durchlicht-
Hellfeldaufnahmen (D-HF) ähnliche Ergebnisse für beide Zelllinien. Im Durch-
licht-Phasenkontrast (D-PK) ergibt sich für beide Zelllinien eine geringere Über-
einstimmung der automatisch generierten Zellflächen zu den Referenzzellen
(Jaccard-Koeffizient: 0,63 und 0,58). Der Grund liegt darin, dass das erstellte
Template nicht alle Farbvariationen der Zellen abdeckt und so teilweise Zellbe-
standteile nicht segmentiert werden.
5.3 Diskussion der Ergebnisse zur Segmentierung zytochemisch
gefärbter Zellbereiche
Es wurden 3 Verfahren zur Segmentierung von May-Grünwald gefärbten L929-
und MC3T3 Zellen in mikroskopischen Bildaufnahmen (Kap. 5.2) vorgestellt.
Vorteile der schwellwertbasierten Methode, beschrieben in Kap. 5.2.1, sind zum
einen die geringe Rechenzeit. Für die Segmentierung eines Bildes der Größe
1376 x 1032 Pixel werden mit einem handelsüblichen PC (Intel Core i7 CPU)
50ms benötigt. Zum anderen ist die Parametrisierung der Methode einfach, es
muss zur Zellsegmentierung lediglich pro Zelllinie und Materialtyp ein Schwell-

5. Segmentierung zytochemisch gefärbter Zellbereiche 47
wert für den R-, G-, und B- Kanal gewählt werden. Durch die separate Einstel-
lung der Schwellwerte für jeden Farbkanal und der anschließenden Vereinigung
der Binärbilder ist die Segmentierung von Objekten, die sich aus unterschiedli-
chen Farben zusammensetzen, möglich.
Ein Nachteil dieser Methode ist die Anfälligkeit gegenüber Helligkeits- und Far-
bänderungen im Bild. Daher ist ein reproduzierbarer Färbevorgang der Zellen
Voraussetzung für eine gute Segmentierung. Da es sich bei der Zellfärbung im
Rahmen des Projektes um einen standardisierten Prozess handelt, beschrieben
in Kap. 3.2, ist die Voraussetzung für dieses Verfahren hier erfüllt.
Das histogrammbasierte Segmentierungsverfahren, beschrieben in Kap. 5.2.2,
benötigt eine moderate Rechenzeit. Sie liegt bei 1,26s für die Zellsegmentie-
rung innerhalb eines Bildes der Größe 1376 x 1032 Pixel. Eine Parametrisie-
rung durch den Anwender ist hierbei nicht erforderlich, da der für die Segmen-
tierung benötigte Schwellwert automatisch aus den Histogrammdaten des Bil-
des berechnet wird. So ist die Methode in der Lage, Helligkeits- und Farbände-
rungen von einem Bild zum nächsten Bild zu kompensieren. Voraussetzung für
eine gute Segmentierung ist jedoch, dass das Substrat eine homogene Färbung
in einem Bild aufweist. Dies ist idealerweise beim Durchlichtkontrast gegeben,
z.B. bei der Analyse von Polystyrolproben. Im Auflichtkontrast liefert dieses Ver-
fahren aufgrund farblich inhomogener Hintergrundbereiche schlechtere Ergeb-
nisse im Vergleich zur Methode aus Kap. 5.2.1.
Mit dem Histogram-Backprojection-Algorithmus (Kap. 5.2.3), ist durch iterative
Ausführung mit mehreren Farbtemplates die Segmentierung verschiedenfarbi-
ger Objekte in einem Bild möglich. So können z.B. unterschiedlich angefärbte
Zellen ohne Anpassung der Parameter segmentiert werden. Ein Nachteil ist die
im Vergleich zu den Verfahren aus Kap. 5.2.1 und Kap. 5.2.2 benötigte relativ
hohe Rechenzeit von über 2s (Tabelle 12).
Tabelle 12: Benötigte Rechenzeit der in Kap. 5.2.1 bis Kap. 5.2.3 vorgestellten Segmentie-
rungsverfahren auf Basis eines Intel Core i7, 3.2GHz
Segmentierungsverfahren
Durchschnittliche Rechenzeit für die Zellsegmentierung eines Pro-
benausschnittes der Größe 1376x1032 Pixel
Schwellwertbasierte Methode (Kap. 5.2.1)
50ms
Histogrammbasiertes Verfahren (Kap. 5.2.2)
1,26s
Histogram-Backprojection-Algorithmus (Kap. 5.2.3)
2,25s

5. Segmentierung zytochemisch gefärbter Zellbereiche 48
Damit ist diese Methode ca. doppelt so langsam im Vergleich zum histogramm-
basierten Schwellwertverfahren (Kap. 5.2.2) und ca. 40-mal langsamer als das
Schwellwertverfahren, beschrieben in Kap. 5.2.1.
Des Weiteren ist im Vorfeld eine Templateherstellung für jede Zelllinie und Ma-
terialtyp notwendig. Dieser Vorgang ist mit einer Bearbeitungszeit im Minuten-
bereich deutlich zeitaufwändiger als die Ausführung des Algorithmus. Allerdings
ist dieser Vorgang nur einmalig durchzuführen, da die erzeugten Templates zur
weiteren Verwendung gespeichert werden.
Die beschriebenen Segmentierungsverfahren werden entsprechend ihrer Vor-
und Nachteile jeweils für unterschiedliche Kontrastarten ausgewählt (Tabelle
13).
Tabelle 13: Vergleich der Jaccard-Koeffizienten der in Kap. 5.2.1 – 5.2.3 vorgestellten Zellseg-
mentierungsverfahren für die Zelltypen L929 und MC3T3 bei unterschiedlichen Materialien und
Mikroskopkontrasten; Die grün hinterlegten Tabellenzellen zeigen das für den aktuell betrachte-
ten Mikroskopkontrast am besten geeignete Segmentierungsverfahren
L929 MC3T3
Stahl* / Titan* Polystyrol Stahl* / Titan* Polystyrol
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
Algo Kap. 5.2.1
0,89 0,85 0,87 0,60 0,87 0,90 0,76 0,53
Algo Kap. 5.2.2
0,86 0,80 0,87 0,77 0,86 0,85 0,82 0,82
Algo Kap. 5.2.3
0,78 0,80 0,81 0,63 0,85 0,86 0,78 0,58
Tabelle 13 veranschaulicht, dass bei Verwendung eines Auflichtmikroskopes im
Hellfeld- und im Dunkelfeldkontrast das Schwellwertverfahren (Kap. 5.2.1) am
besten geeignet ist. Dies trifft sowohl bei den L929- als auch MC3T3-Zellen zu.
Die Abweichungen zu den Jaccard-Koeffizienten des histogrammbasierten Ver-
fahrens (Kap. 5.2.2) sind jedoch gering, so dass diese Methode ebenfalls für die
Zellsegmentierung einsetzbar ist.
Für die Analyse von Polystyrolproben im Durchlicht-Hellfeldkontrast (D-HF) sind
die schwellwertbasierte Methode (Kap. 5.2.1) sowie das histogrammbasierte
Schwellwertverfahren für Zellen der Linie L929 gleichermaßen geeignet, die
automatisch rekonstruierten Zellflächen weichen in beiden Fällen um 13% von
den Referenzzellflächen ab. Bei der Untersuchung von MC3T3-Zellen können
ebenfalls die Schwellwertmethode sowie das histogrammbasierte Verfahren

5. Segmentierung zytochemisch gefärbter Zellbereiche 49
eingesetzt werden, wobei dass histogrammbasierte Verfahren (Kap. 5.2.2)
leicht besser abschneidet.
Bei Verwendung eines Durchlichtmikroskops im Phasenkontrast (D-PK) ist so-
wohl für L929- als auch MC3T3 Zellen nur das histogrammbasierte Verfahren
(Kap. 5.2.2) geeignet. Der Grund liegt darin, dass dieses Verfahren zunächst
die Substratfläche segmentiert, und da das verwendete Material Polystyrol eine
homogene Färbung aufweist, erfolgt eine vergleichsweise sichere Segmentie-
rung. Durch die anschließende Subtraktion des detektierten Hintergrundes vom
Gesamtbild werden die Zellbereiche indirekt ermittelt. Die Zellfärbung variiert im
Phasenkontrast so stark, dass die übrigen Segmentierungsverfahren keine ak-
zeptablen Segmentierungsergebnisse liefern.
Das Histogram-Backprojection-Verfahren liefert für keinen Mikroskopkontrast
bessere Ergebnisse als die anderen Methoden, so dass dieser Algorithmus für
die Segmentierung von L929- und MC3T3 Zellen unter Verwendung der ge-
nannten Mikroskopkontraste nicht zum Einsatz kommt.
Neben den bereits vorgestellten Verfahren wurden auch andere Algorithmen
aus der Literatur auf ihre Eignung als Zellsegmentierungsverfahren untersucht.
Eine Methode basierend auf Kantenfilter ist in (57) zu finden. Dort werden Zel-
len zwar mit Hilfe eines standard Schwellwertverfahrens segmentiert, allerdings
auf Basis des Gradientenbildes, das zuvor mit Hilfe geeigneter Kantenfilter be-
rechnet wird. Diese Vorgehensweise ist für das in (57) verwendete Bildmaterial
geeignet (Abb. 24), da sich die Zellgrenzen mit gutem Kontrast von dem Hinter-
grund abheben und so ein verwertbares Gradientenbild entsteht.
Abbildung 25: Links: Hellfeldbild von ungefärbten Hefezellen (57); Rechts: Gradientenbild von
Abb. 24 Links
Das Bildmaterial im Rahmen dieses Projektes weist hingegen keine vergleich-
bar kontrastreichen Übergänge von Zelle zu Hintergrund auf und außerdem
erscheinen die Zellen aufgrund der unterschiedlich gefärbten Zellbereiche we-

5. Segmentierung zytochemisch gefärbter Zellbereiche 50
sentlich inhomogener als die in (57) bearbeiteten Zellen. Aus diesen Gründen
ist eine Segmentierung auf Basis von Kantenalgorithmen für die vorliegende
Aufgabenstellung nicht geeignet.
In (28) wird die Wasserscheidentransformation zur Zellsegmentierung einge-
setzt. Nach einer Fluoreszenzanfärbung von Zellmembran und Zellkernen wird
das Epithel nach einer Kontrasterhöhung mit Hilfe eines Schwellwertverfahrens
und morphologischen Operationen segmentiert. Die Detektion der Zellkerne
erfolgt ebenfalls über eine adaptive Schwellwertmethode mit anschließender
Wasserscheidentransformation. Die Erkennung der Zellgrenzen erfolgt auch mit
Hilfe einer Wasserscheidentransformation. Im Ergebnis resultiert laut (28) eine
Übersegmentierung der Zellen von ca. 7% im Vergleich zu einer Referenzzäh-
lung. Durch die 3-fache Fluoreszenzanfärbung verschiedener Zellbereiche liegt
ein optimaler farblicher Kontrast zur Segmentierung der gewünschten Zellberei-
che vor. In diesem Fall kann die Wasserscheidentransformation eingesetzt
werden, da sich die Zellkonturen kontrastreich im Bild hervorheben. Dennoch
resultiert bereits in diesem Fall eine Übersegmentierung von ca. 7%. Im Rah-
men dieses Projektes wird statt einer Fluoreszenzfärbung eine zytochemische
May-Grünwald Zellfärbung eingesetzt. Im Vergleich zur Fluoreszenzfärbung
resultiert ein schlechterer Kontrast zur Unterscheidung zwischen verschiedenen
Zellbestandteilen oder agglomerierten Zellen im Bildmaterial. Infolgedessen ist
von einem schlechten Segmentierungsergebnis auf Basis einer Wasserschei-
dentransformation auszugehen. Somit ist die Anwendung eines Verfahrens ba-
sierend auf diesem Algorithmus hier nicht sinnvoll.
Darüber hinaus werden aktive Konturen, auch Snakes genannt, bei der
Zellsegmentierung eingesetzt (29; 30; 31; 32; 33).
Aktive Konturen bieten den Vorteil, dass sie bei kontrastarmen und teilweise
verrauschten Bildern im Vergleich zu Schwellwertverfahren akzeptable Seg-
mentierungsergebnisse liefern.
Nachteil dieses Verfahrens ist die schlechte Anpassung an topologische
Veränderungen und die sehr hohe Rechenzeit. Für die Segmentierung einer
Zelle werden bereits 1-5s benötigt. Die Zellzahl in einem Probenausschnitt der
Linie L929 beträgt bei mittlerer Zelldichte ca. 200-300 Zellen, so dass für die
Zellsegmentierung eines Bildes mehrere Minuten benötigt werden. Ein anderer
Nachteil ist die notwendige Initialisierung einer Startkontur nahe der zu
suchenden realen Objektkontur. Die Startkontur muss entweder interaktiv durch
den Softwarebediener erstellt werden oder es werden weitere Algorithmen für
eine automatische Bestimmung der Startkonturen benötigt. Aus diesen
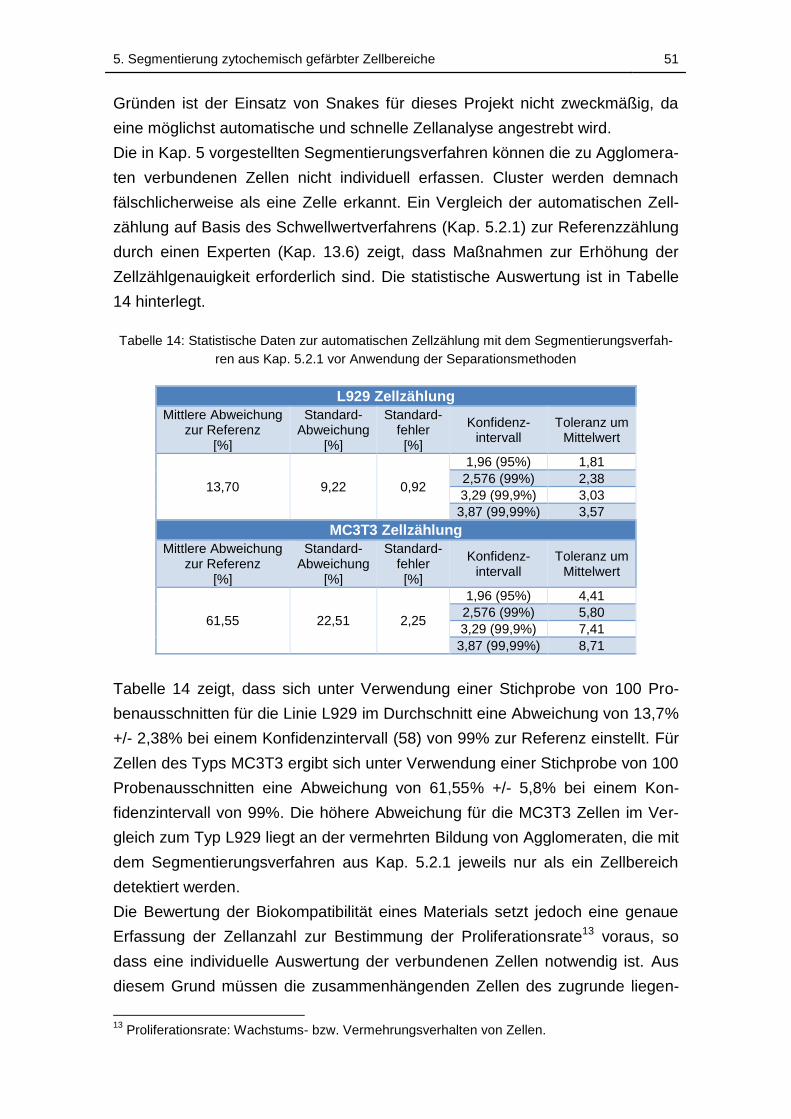
5. Segmentierung zytochemisch gefärbter Zellbereiche 51
Gründen ist der Einsatz von Snakes für dieses Projekt nicht zweckmäßig, da
eine möglichst automatische und schnelle Zellanalyse angestrebt wird.
Die in Kap. 5 vorgestellten Segmentierungsverfahren können die zu Agglomera-
ten verbundenen Zellen nicht individuell erfassen. Cluster werden demnach
fälschlicherweise als eine Zelle erkannt. Ein Vergleich der automatischen Zell-
zählung auf Basis des Schwellwertverfahrens (Kap. 5.2.1) zur Referenzzählung
durch einen Experten (Kap. 13.6) zeigt, dass Maßnahmen zur Erhöhung der
Zellzählgenauigkeit erforderlich sind. Die statistische Auswertung ist in Tabelle
14 hinterlegt.
Tabelle 14: Statistische Daten zur automatischen Zellzählung mit dem Segmentierungsverfah-
ren aus Kap. 5.2.1 vor Anwendung der Separationsmethoden
L929 Zellzählung Mittlere Abweichung
zur Referenz [%]
Standard- Abweichung
[%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Mittelwert
13,70 9,22 0,92
1,96 (95%) 1,81
2,576 (99%) 2,38
3,29 (99,9%) 3,03
3,87 (99,99%) 3,57
MC3T3 Zellzählung
Mittlere Abweichung zur Referenz
[%]
Standard- Abweichung
[%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Mittelwert
61,55 22,51 2,25
1,96 (95%) 4,41
2,576 (99%) 5,80
3,29 (99,9%) 7,41
3,87 (99,99%) 8,71
Tabelle 14 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten für die Linie L929 im Durchschnitt eine Abweichung von 13,7%
+/- 2,38% bei einem Konfidenzintervall (58) von 99% zur Referenz einstellt. Für
Zellen des Typs MC3T3 ergibt sich unter Verwendung einer Stichprobe von 100
Probenausschnitten eine Abweichung von 61,55% +/- 5,8% bei einem Kon-
fidenzintervall von 99%. Die höhere Abweichung für die MC3T3 Zellen im Ver-
gleich zum Typ L929 liegt an der vermehrten Bildung von Agglomeraten, die mit
dem Segmentierungsverfahren aus Kap. 5.2.1 jeweils nur als ein Zellbereich
detektiert werden.
Die Bewertung der Biokompatibilität eines Materials setzt jedoch eine genaue
Erfassung der Zellanzahl zur Bestimmung der Proliferationsrate13 voraus, so
dass eine individuelle Auswertung der verbundenen Zellen notwendig ist. Aus
diesem Grund müssen die zusammenhängenden Zellen des zugrunde liegen-
13
Proliferationsrate: Wachstums- bzw. Vermehrungsverhalten von Zellen.
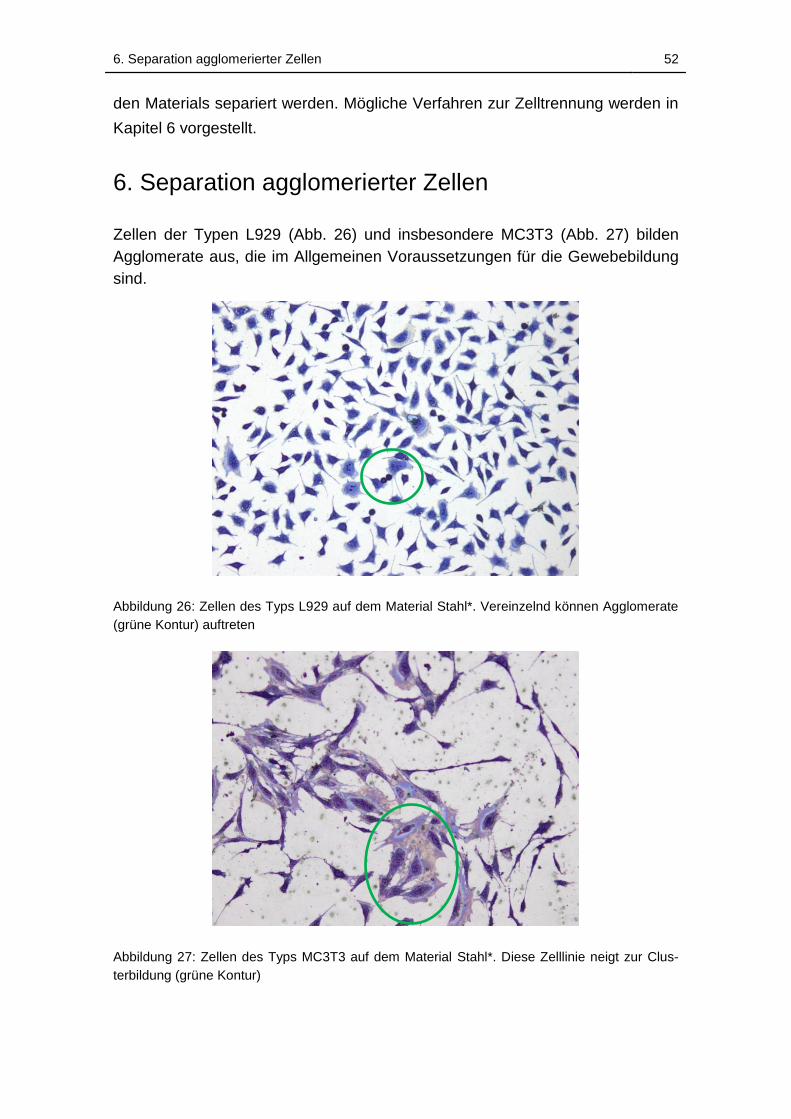
6. Separation agglomerierter Zellen 52
den Materials separiert werden. Mögliche Verfahren zur Zelltrennung werden in
Kapitel 6 vorgestellt.
6. Separation agglomerierter Zellen
Zellen der Typen L929 (Abb. 26) und insbesondere MC3T3 (Abb. 27) bilden
Agglomerate aus, die im Allgemeinen Voraussetzungen für die Gewebebildung
sind.
Abbildung 26: Zellen des Typs L929 auf dem Material Stahl*. Vereinzelnd können Agglomerate
(grüne Kontur) auftreten
Abbildung 27: Zellen des Typs MC3T3 auf dem Material Stahl*. Diese Zelllinie neigt zur Clus-
terbildung (grüne Kontur)

6. Separation agglomerierter Zellen 53
Für die Separation von L929 und MC3T3 Cluster sowie Zelltypen mit ähnlicher
Morphologie werden in diesem Kapitel geeignete Separationsverfahren vorge-
stellt. Das Konzept des Separationsvorgangs ist in Abb. 28 dargestellt.
Abbildung 28: Ablaufdiagramm zur Zellvereinzelung. Nähere Erläuterungen siehe Text
In Kap. 6.1 wird ein kontextbasiertes Trennverfahren (CBS) vorgestellt, das alle
Zellen an Verengungen, wie sie bei der Zelllinie L929 überwiegend an Zellgren-
zen innerhalb der Agglomerate vorzufinden sind, aufspaltet (59). Diese Metho-
de wird unabhängig vom verwendeten Zelltyp immer durchgeführt, da auch bei
Typen mit kompakter Clusterbildung, z.B. MC3T3, vereinzelnd Verengungen an
Zellkontaktstellen auftreten können. Zelltrennalgorithmen kommen zur Einspa-
rung von Rechenzeit nur für Zellbereiche, die als Agglomerate klassifiziert wur-

6. Separation agglomerierter Zellen 54
den, zum Einsatz. Eine geeignete Methode zur Klassifizierung der Zellbereiche
in Einzelzellen und Agglomerate wird in Kap. 6.2 erläutert. Im nächsten Schritt
erfolgt die Detektion der Kerne innerhalb der als Cluster definierten Zellbereiche
(Kap. 6.3). Werden Kernbereiche ermittelt, kommen, abhängig von der Zellmor-
phologie, zwei unterschiedliche Separationsverfahren zum Einsatz. Kap. 6.4
beschreibt ein Trennverfahren spezialisiert auf Zelltypen, die an den Kontakt-
stellen innerhalb der Agglomerate Verengungen an der Kontur aufweisen. Kap.
6.5 hingegen beschreibt ein Separationsverfahren, dass für Zelllinien mit kom-
pakter Clusterbildung ohne Verengungen an den Zellkontaktstellen zum Einsatz
kommt. Können keine Kernbereiche ermittelt werden, erfolgt die Separation der
Cluster mit Hilfe des in Kap. 6.6 beschriebenen Verfahrens.
Durch den Bildrand abgeschnittene Zellbereiche verfälschen die Analyse, z.B.
bei der Bestimmung der mittleren Zellfläche und müssen eliminiert werden. Ein
Verfahren zur Verarbeitung der Randzellen wird in Kap. 6.7 vorgestellt. Damit
ist die Erfassung der individuellen Zellbereiche abgeschlossen.
Die Auswahl der verschiedenen Verfahren wird von der Laborfachkraft im Vor-
feld einer Untersuchung festgelegt und in einer Parameterdatei gespeichert.
Liegen für die verschiedenen Kombinationen von Zelltyp und Substrat jeweils
Parameterdateien vor, so muss zu Beginn einer Analyse die passende Parame-
terdatei geladen werden. Weitere Parametereinstellungen sind nicht notwendig.
6.1 Kontextbasiertes Trennverfahren
Die CBS-Methode ist ein Verfahren zur Separation beliebiger zusammenhän-
gender Objekte (59). Die zusammenhängenden Objekte müssen zunächst mit
Hilfe eines geeigneten Verfahrens segmentiert werden, z.B. mit einer der Me-
thoden aus Kap. 5.2. Die Separation der segmentierten Objektbereiche erfolgt
im ersten Schritt mit Hilfe eines kreisförmigen Strukturelementes C1 mit Radius
r1, das die Kontur des Objektes RC abfährt (Abb. 29 Links). Da RC im weiteren
Verlauf des Algorithmus manipuliert wird, ist die Erstellung einer Kopie RC_Copy
von RC notwendig. Für jeden i-ten Konturpunkt wird geprüft, ob die von C1 ein-
geschlossene Hintergrundmenge B aus zwei voneinander getrennten Teilmen-
gen B1 und B2 besteht. B1 und B2 müssen durch die von C1 eingeschlossene
Objektregion getrennt sein (Abb. 29 Mitte). Hierbei handelt es sich um eine
notwendige Bedingung für eine lokale Verengung des Objektes. Sie ist durch
die Aufweitungen an den Enden gekennzeichnet. Um dies festzustellen, wird
zuerst eine zu C1 konzentrisch angeordnete Kreismaske C2 mit dem Radius r2 =
a • r1 generiert. Bei der Einstellung des Parameters a ist zu beachten dass die-

6. Separation agglomerierter Zellen 55
ser Abhängig von der Objektgröße ist. Liegen große Objekte im Bild vor, ist a
größer zu wählen als wenn kleinere Objekte vorliegen. Sollen in einem Bild un-
terschiedlich große Objekte getrennt werden kann die CBS iterativ mit ver-
schiedenen Einstellungen für den Parameter a ausgeführt werden. Mit Hilfe von
C1 und C2 wird dann die Kreisringmaske CR = C2 \ C1 berechnet und im weite-
ren Verlauf des Algorithmus bestimmt (Abb. 29 rechts).
Abbildung 29: Links: Die Kontur des Objektes RC wird mit einem kreisförmigen Strukturelement
C1 abgefahren. Mitte: Der in C1 liegende Hintergrund B wird durch RA,n in zwei Teilmengen B1
und B2 aufgeteilt. Rechts: Mit Hilfe von C1 und CR wird geprüft, ob eine Verengung vorliegt
Abb. 29 rechts zeigt, dass die Punktmenge RB aus 2 nicht überlappenden oder
angrenzenden Punktmengen 1BR und
2BR besteht. Die Entscheidungsfunkti-
on f1(x) wird durch Gl. 4 definiert. Wenn f1(x) den Wert true annimmt, liegt eine
Verengung vor.
1 21 , ,B A n B A nf x R R x R R x . (Gl. 4)
Das Symbol |..| steht für die Anzahl Bildpunkte einer Menge. Der Parameter x
gibt vor, um welchen Anteil 1BR und
2BR größer sein müssen als der von C1
eingeschlossene Objektbereich. Die Menge RB kann sich auch aus l nicht über-
lappenden oder angrenzenden Punktmengen zusammensetzen. Die Entschei-
dungsfunktion f1(x) erweitert sich und wird entsprechend Gl. 5 definiert. Nimmt
f1(x) den Wert true an, liegt eine Verengung vor.
1 21 , , ,...lB A n B A n B A nf x R R x R R x R R x . (Gl. 5)
Liefert f1(x) den Wert true, erfolgt die Objektseparation mit der Zuweisung
',\C C A nR R R
':C CR R .
Der Objektbereich RC wird überschrieben, um Mehrfachtrennungen an benach-
barten Konturpunkten zu vermeiden. RA,n wird als Element der Menge RA_Ges
hinzugefügt. Der beschriebene Separationsvorgang wird an den n Engstellen
des Objektclusters durchgeführt.
1BR 2BR
1 2B B BR R R

6. Separation agglomerierter Zellen 56
Abbildung 30: Ergebnis der Objekttrennung nach 3 Separationsvorgängen. Es resultieren 3
neue Objektbereiche RS,1, RS,2, RS,3. Die Objektbereiche RA,2 und RA,3 sind zufällig dem sepa-
rierten Objekt RS,3 und der Objektbereich RA,1 des getrennten Objektes RS,1 zugeordnet worden
Im dargestellten Objektcluster (Abb. 30) erhält man RS,1, RS,2 und RS,3 als Er-
gebnis des Labelings von RC, nachdem im Rahmen der 3 Separationsprozesse
RA,1, RA,2 und RA,3 von RC subtrahiert und das Ergebnis wieder RC zugewiesen
wurde. Da die in RA_Ges gespeicherten Objektbereiche Teilmengen des ur-
sprünglichen Objektbereiches RC_Copy sind, werden sie zufällig dem jeweils be-
nachbarten Bereich zugeordnet. In Abb. 30 wurden zwei der in RA_Ges gespei-
cherten Objektbereiche RS,3 und ein in RA_Ges gespeicherter Objektbereich RS,1
zugeordnet.
',3 ,3 ,1 ,3S S A AR R R R
',3 ,3:S SR R
',1 ,1 ,2S S AR R R
',1 ,1:S SR R
Es können Situationen vorliegen, bei denen Objekttrennungen nicht sinnvoll
sind, z.B. bei Unregelmäßigkeiten in der Objektkontur, aber f1(x) true als Ergeb-
nis hat. Aus diesem Grund wird die Entscheidungsfunktion f1(x) für eine Objektt-
rennung erweitert.
Eine Teilung unterbleibt für den Fall, wenn eine der beiden Regionen 1BR oder
2BR um das y-fache größer ist als die andere (Abb. 31).
RS,1 RS,3
RA,3
RA,1 RA,2
RS,2

6. Separation agglomerierter Zellen 57
Abbildung 31: Unregelmäßigkeiten bei einer Objektkontur die dazu führen dass 1BR und
2BR
deutlich unterschiedliche Flächenanteile besitzen.
Die erweiterte Entscheidungsfunktion lautet 1 2( , ) ( ) ( )f x y f x f y mit
1 2 2 12 B B B Bf y R y R R y R . (Gl. 6)
Liefert f1(x) den Wert true, liegt eine Verengung vor. Nimmt f2(y) den Wert true
an, ist sichergestellt, dass die Flächen von 1BR und
2BR weniger als das y-
fache voneinander abweichen, das Vorliegen einer Objektunregelmäßigkeit ist
unwahrscheinlich. Werden f1(x) und f2(x) logisch UND Verknüpft, liefert die er-
weiterte Entscheidungsfunktion f(x,y) ebenso den Wert true und eine Separati-
on wird durchgeführt. Der gesamte Programmablauf inklusive CBS ist in Abb.32
dargestellt.
2BR
1BR
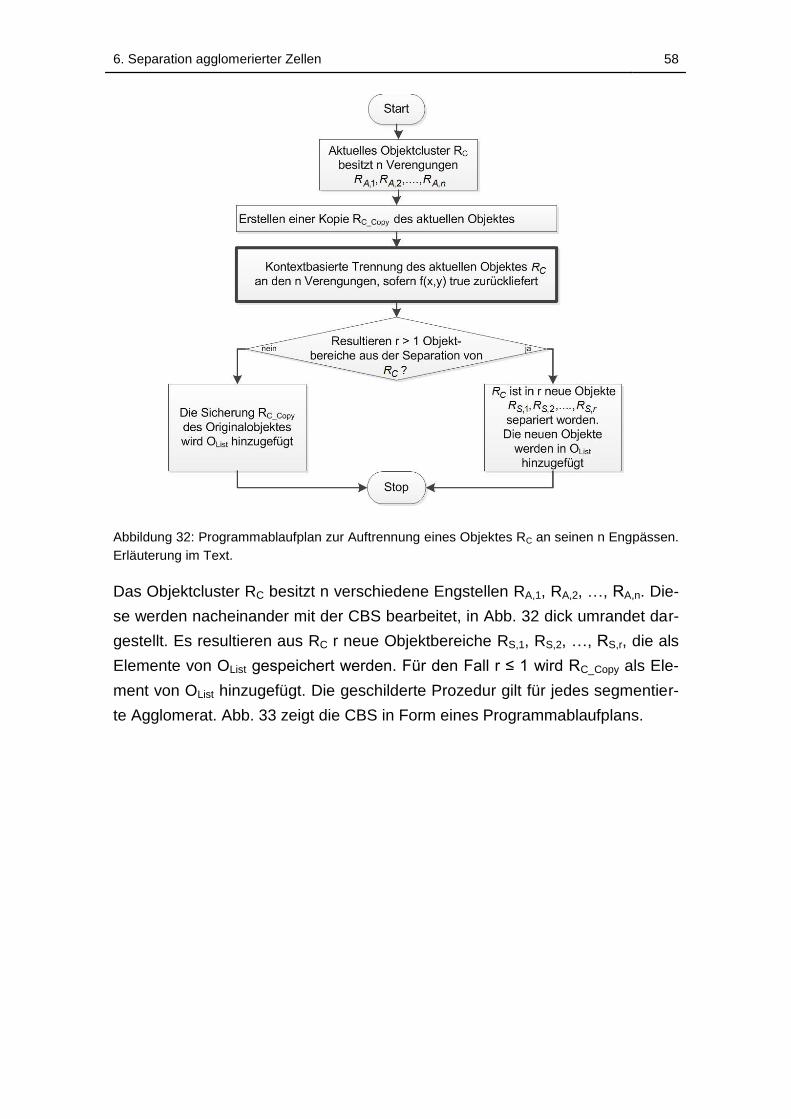
6. Separation agglomerierter Zellen 58
Abbildung 32: Programmablaufplan zur Auftrennung eines Objektes RC an seinen n Engpässen.
Erläuterung im Text.
Das Objektcluster RC besitzt n verschiedene Engstellen RA,1, RA,2, …, RA,n. Die-
se werden nacheinander mit der CBS bearbeitet, in Abb. 32 dick umrandet dar-
gestellt. Es resultieren aus RC r neue Objektbereiche RS,1, RS,2, …, RS,r, die als
Elemente von OList gespeichert werden. Für den Fall r ≤ 1 wird RC_Copy als Ele-
ment von OList hinzugefügt. Die geschilderte Prozedur gilt für jedes segmentier-
te Agglomerat. Abb. 33 zeigt die CBS in Form eines Programmablaufplans.

6. Separation agglomerierter Zellen 59
Abbildung 33: Programmablaufplan zur CBS
Abb. 33 zeigt eine entscheidungsbasierte Struktur des Algorithmus, mit der die
möglichen geometrischen Variationen der Objekte bearbeitet werden. Nach Be-

6. Separation agglomerierter Zellen 60
rechnung von , 1A n CR C R mit anschließendem Labeling erhält man k separa-
te Punktmengen. Für den Fall k > 1, wird nur die Punktmenge behandelt, die
den Mittelpunkt von C1 beinhaltet. Wenn k = 1, folgt die Berechnung von
B R CR C R . RB wird einem Labeling unterzogen, wodurch l Teilmengen
1 2, ,...,
lB B BR R R entstehen, die, vereinigt mit RA,n, eine zusammenhängende Men-
ge bilden. Wenn l = 2, prüft die Entscheidungsfunktion f(x,y), ob die beiden
Teilmengen 1BR und
2BR eine Verengung repräsentieren. Im Fall l > 2 wird ge-
prüft, ob alle Teilmengen
max,
p
p p
B
B A n B
R
R x R Ry
mit p = 1, 2,..., l; x = 1.5; y = 3 (Gl. 7)
erfüllen, wobei die größte Teilmenge von 1 2, ,...,
lB B BR R R aus maxpBR Elemen-
ten besteht. Enthält 1 2, ,...,
lB B BR R R mindestens zwei Mengen, die problemspezifi-
sche Bedingungen (anschließender Abschnitt „Anwendung der CBS für die Se-
paration zusammengewachsener L929-Zellen“) erfüllen, wird der Trennungs-
prozess eingeleitet und es entstehen i separate Objektbereiche RS,1, RS,2,…,
RS,i. Existieren q Mengen in 1 2, ,...,
lB B BR R R , die die Bedingungen nicht erfüllen,
werden sie als Elemente in RBD hinzugefügt, RBD = 1 2, ,...,
qBD BD BDR R R . Nach
der Objekttrennung erfolgt mit Hilfe der in RBD vorhandenen Elemente die Er-
mittlung der unerwünscht abgetrennten Objektbereiche. Ein abgetrennter Ob-
jektbereich RS,j mit j = 1, 2,…, i gilt als unerwünscht, wenn für ,jt tU S j BDR R R
(mit t = 1, 2,…,q) |RUjt| > 0 erfüllt ist. Ist ein unerwünschter Objektbereich RS,j
ermittelt, wird dieser einem der übrigen Objektbereiche zugewiesen, wobei die
Auswahl zufällig erfolgt. Auf diese Weise kann eine Objektseparation auch dort
erfolgen, wo mehrere Objektbereiche zusammenlaufen, aber diese nicht alle
getrennt werden dürfen (Tabelle 15, Fall 5). Die für eine Objektseparation auf-
tretenden Fälle sind in Tabelle 15 aufgeführt. Abb. 34 dient als Zeichenerklä-
rung für Tabelle 15.
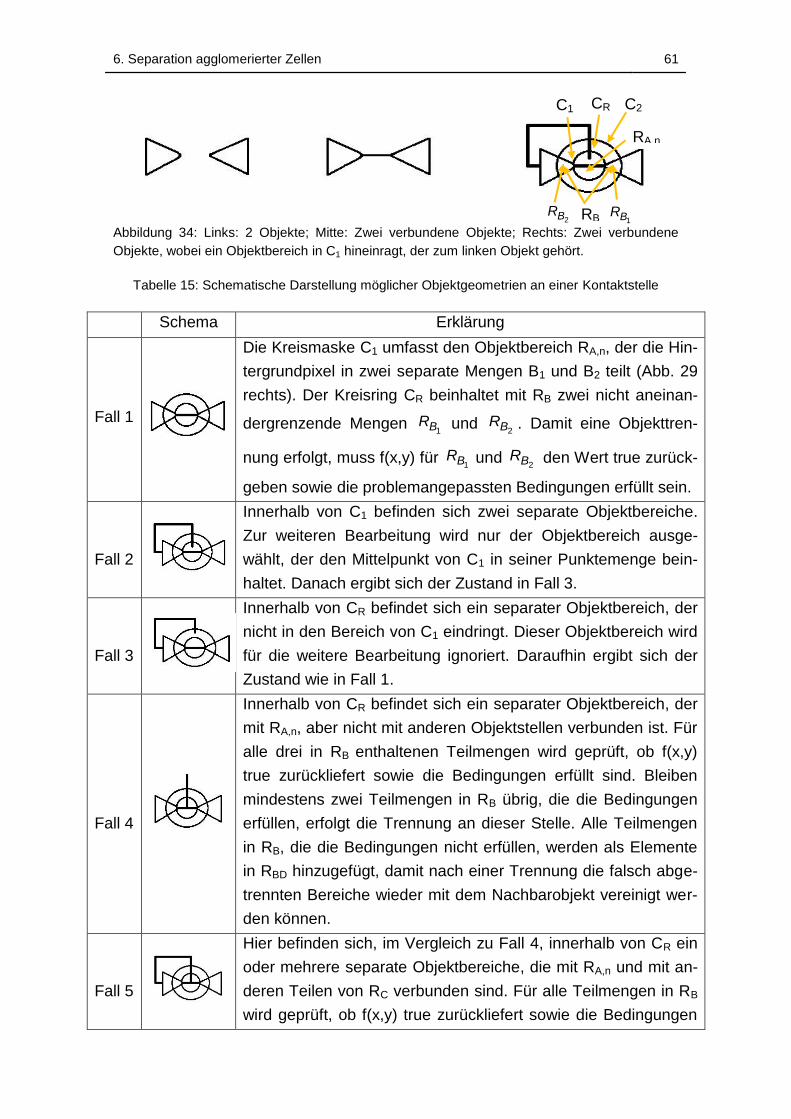
6. Separation agglomerierter Zellen 61
Abbildung 34: Links: 2 Objekte; Mitte: Zwei verbundene Objekte; Rechts: Zwei verbundene
Objekte, wobei ein Objektbereich in C1 hineinragt, der zum linken Objekt gehört.
Tabelle 15: Schematische Darstellung möglicher Objektgeometrien an einer Kontaktstelle
Schema Erklärung
Fall 1
Die Kreismaske C1 umfasst den Objektbereich RA,n, der die Hin-
tergrundpixel in zwei separate Mengen B1 und B2 teilt (Abb. 29
rechts). Der Kreisring CR beinhaltet mit RB zwei nicht aneinan-
dergrenzende Mengen 1BR und
2BR . Damit eine Objekttren-
nung erfolgt, muss f(x,y) für 1BR und
2BR den Wert true zurück-
geben sowie die problemangepassten Bedingungen erfüllt sein.
Fall 2
Innerhalb von C1 befinden sich zwei separate Objektbereiche.
Zur weiteren Bearbeitung wird nur der Objektbereich ausge-
wählt, der den Mittelpunkt von C1 in seiner Punktemenge bein-
haltet. Danach ergibt sich der Zustand in Fall 3.
Fall 3
Innerhalb von CR befindet sich ein separater Objektbereich, der
nicht in den Bereich von C1 eindringt. Dieser Objektbereich wird
für die weitere Bearbeitung ignoriert. Daraufhin ergibt sich der
Zustand wie in Fall 1.
Fall 4
Innerhalb von CR befindet sich ein separater Objektbereich, der
mit RA,n, aber nicht mit anderen Objektstellen verbunden ist. Für
alle drei in RB enthaltenen Teilmengen wird geprüft, ob f(x,y)
true zurückliefert sowie die Bedingungen erfüllt sind. Bleiben
mindestens zwei Teilmengen in RB übrig, die die Bedingungen
erfüllen, erfolgt die Trennung an dieser Stelle. Alle Teilmengen
in RB, die die Bedingungen nicht erfüllen, werden als Elemente
in RBD hinzugefügt, damit nach einer Trennung die falsch abge-
trennten Bereiche wieder mit dem Nachbarobjekt vereinigt wer-
den können.
Fall 5
Hier befinden sich, im Vergleich zu Fall 4, innerhalb von CR ein
oder mehrere separate Objektbereiche, die mit RA,n und mit an-
deren Teilen von RC verbunden sind. Für alle Teilmengen in RB
wird geprüft, ob f(x,y) true zurückliefert sowie die Bedingungen
RB
C1 CR C2
RA,n
2BR1BR
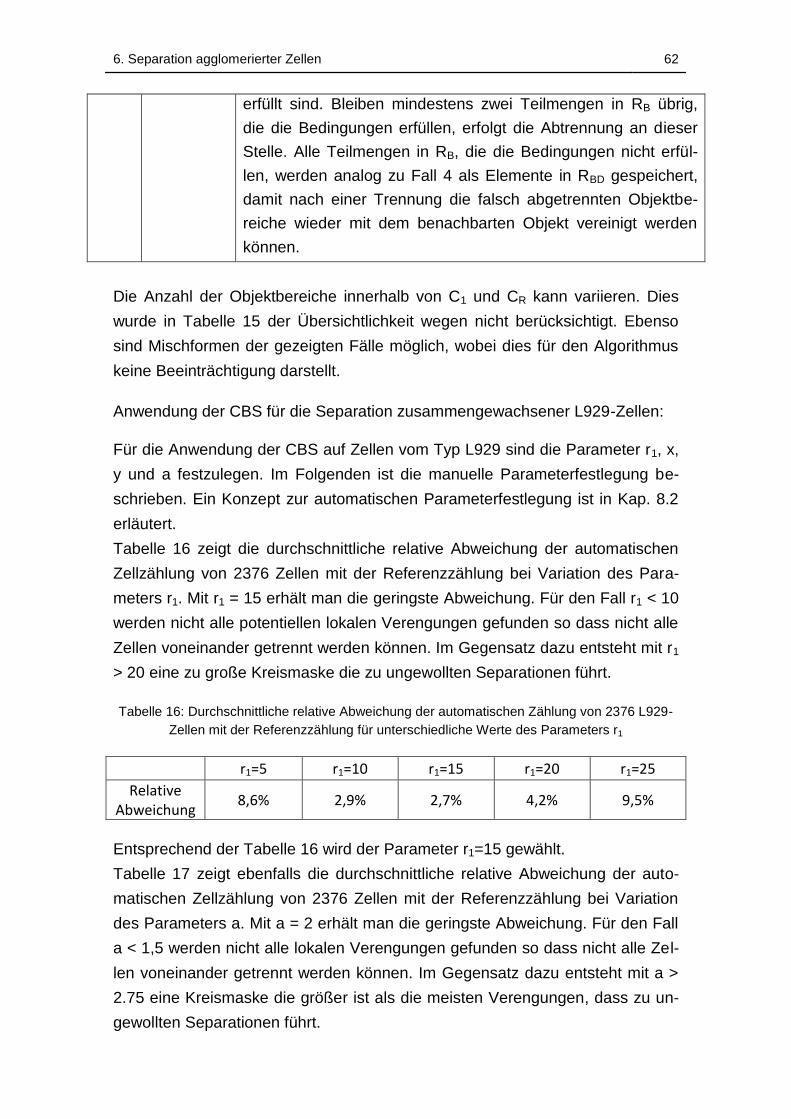
6. Separation agglomerierter Zellen 62
erfüllt sind. Bleiben mindestens zwei Teilmengen in RB übrig,
die die Bedingungen erfüllen, erfolgt die Abtrennung an dieser
Stelle. Alle Teilmengen in RB, die die Bedingungen nicht erfül-
len, werden analog zu Fall 4 als Elemente in RBD gespeichert,
damit nach einer Trennung die falsch abgetrennten Objektbe-
reiche wieder mit dem benachbarten Objekt vereinigt werden
können.
Die Anzahl der Objektbereiche innerhalb von C1 und CR kann variieren. Dies
wurde in Tabelle 15 der Übersichtlichkeit wegen nicht berücksichtigt. Ebenso
sind Mischformen der gezeigten Fälle möglich, wobei dies für den Algorithmus
keine Beeinträchtigung darstellt.
Anwendung der CBS für die Separation zusammengewachsener L929-Zellen:
Für die Anwendung der CBS auf Zellen vom Typ L929 sind die Parameter r1, x,
y und a festzulegen. Im Folgenden ist die manuelle Parameterfestlegung be-
schrieben. Ein Konzept zur automatischen Parameterfestlegung ist in Kap. 8.2
erläutert.
Tabelle 16 zeigt die durchschnittliche relative Abweichung der automatischen
Zellzählung von 2376 Zellen mit der Referenzzählung bei Variation des Para-
meters r1. Mit r1 = 15 erhält man die geringste Abweichung. Für den Fall r1 < 10
werden nicht alle potentiellen lokalen Verengungen gefunden so dass nicht alle
Zellen voneinander getrennt werden können. Im Gegensatz dazu entsteht mit r1
> 20 eine zu große Kreismaske die zu ungewollten Separationen führt.
Tabelle 16: Durchschnittliche relative Abweichung der automatischen Zählung von 2376 L929-
Zellen mit der Referenzzählung für unterschiedliche Werte des Parameters r1
r1=5 r1=10 r1=15 r1=20 r1=25
Relative Abweichung
8,6% 2,9% 2,7% 4,2% 9,5%
Entsprechend der Tabelle 16 wird der Parameter r1=15 gewählt.
Tabelle 17 zeigt ebenfalls die durchschnittliche relative Abweichung der auto-
matischen Zellzählung von 2376 Zellen mit der Referenzzählung bei Variation
des Parameters a. Mit a = 2 erhält man die geringste Abweichung. Für den Fall
a < 1,5 werden nicht alle lokalen Verengungen gefunden so dass nicht alle Zel-
len voneinander getrennt werden können. Im Gegensatz dazu entsteht mit a >
2.75 eine Kreismaske die größer ist als die meisten Verengungen, dass zu un-
gewollten Separationen führt.
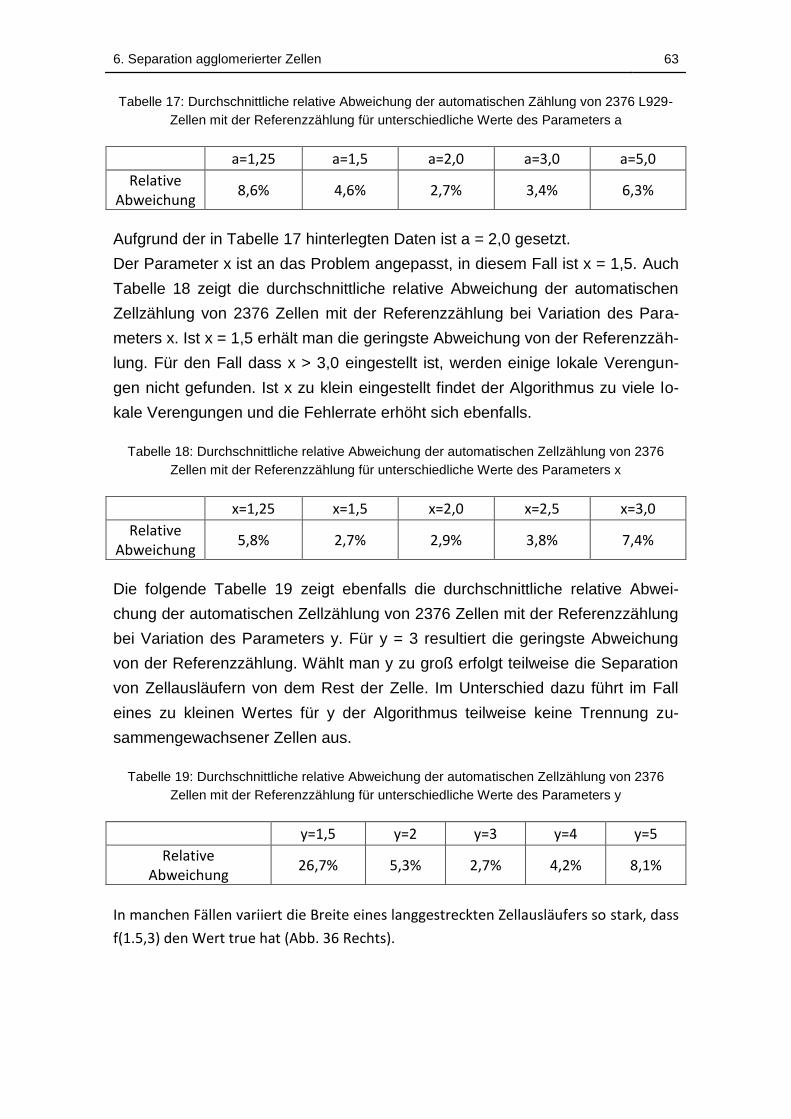
6. Separation agglomerierter Zellen 63
Tabelle 17: Durchschnittliche relative Abweichung der automatischen Zählung von 2376 L929-
Zellen mit der Referenzzählung für unterschiedliche Werte des Parameters a
a=1,25 a=1,5 a=2,0 a=3,0 a=5,0
Relative Abweichung
8,6% 4,6% 2,7% 3,4% 6,3%
Aufgrund der in Tabelle 17 hinterlegten Daten ist a = 2,0 gesetzt.
Der Parameter x ist an das Problem angepasst, in diesem Fall ist x = 1,5. Auch
Tabelle 18 zeigt die durchschnittliche relative Abweichung der automatischen
Zellzählung von 2376 Zellen mit der Referenzzählung bei Variation des Para-
meters x. Ist x = 1,5 erhält man die geringste Abweichung von der Referenzzäh-
lung. Für den Fall dass x > 3,0 eingestellt ist, werden einige lokale Verengun-
gen nicht gefunden. Ist x zu klein eingestellt findet der Algorithmus zu viele lo-
kale Verengungen und die Fehlerrate erhöht sich ebenfalls.
Tabelle 18: Durchschnittliche relative Abweichung der automatischen Zellzählung von 2376
Zellen mit der Referenzzählung für unterschiedliche Werte des Parameters x
x=1,25 x=1,5 x=2,0 x=2,5 x=3,0
Relative Abweichung
5,8% 2,7% 2,9% 3,8% 7,4%
Die folgende Tabelle 19 zeigt ebenfalls die durchschnittliche relative Abwei-
chung der automatischen Zellzählung von 2376 Zellen mit der Referenzzählung
bei Variation des Parameters y. Für y = 3 resultiert die geringste Abweichung
von der Referenzzählung. Wählt man y zu groß erfolgt teilweise die Separation
von Zellausläufern von dem Rest der Zelle. Im Unterschied dazu führt im Fall
eines zu kleinen Wertes für y der Algorithmus teilweise keine Trennung zu-
sammengewachsener Zellen aus.
Tabelle 19: Durchschnittliche relative Abweichung der automatischen Zellzählung von 2376
Zellen mit der Referenzzählung für unterschiedliche Werte des Parameters y
y=1,5 y=2 y=3 y=4 y=5
Relative Abweichung
26,7% 5,3% 2,7% 4,2% 8,1%
In manchen Fällen variiert die Breite eines langgestreckten Zellausläufers so stark, dass
f(1.5,3) den Wert true hat (Abb. 36 Rechts).

6. Separation agglomerierter Zellen 64
Abbildung 35: Links: Die Abtrennung eines Zellausläufers von seiner Zelle ist nicht erwünscht;
Rechts: Die Aufspaltung eines Zellausläufers ist ebenfalls unerwünscht
In Abb. 35 (Links) ist der Fall2 1
3B BR R skizziert. Damit ist f2(3) = false und
somit f(1.5,3) = false. Eine Trennung darf nicht erfolgen, denn es wurde ein zur
Zelle gehörender Zellausläufer erkannt. In manchen Fällen variiert die Breite
eines langgestreckten Zellausläufers so stark, dass f(1.5,3) den Wert true hat
(Abb. 35 Rechts). Auch dies ist eine offensichtliche Fehlentscheidung und muss
mit weiteren Bedingungen unterbunden werden.
Bedingung 1: Alle durch eine Trennung entstehenden Objektbereiche müssen
jeweils eine Mindestgröße Amin aufweisen, andernfalls erfolgt keine Trennung.
Bedingung 2: Bis zu einer maximalen Objektgröße Amax muss die Kompaktheit
aller Objektbereiche den Wert K a annehmen, mit a = 10 in diesem Fall.
Besitzt ein Element eine Fläche > Amax, handelt es sich um eine schmale lang-
gezogene Einzelzelle und eine Trennung ist korrekt.
6.1.1 Ergebnisse
Um die Effizienz des hier vorgestellten Verfahrens zu beurteilen, wurden, neben
einer Zeitmessung für 10 verschiedene L929-Probenausschnitte (Tabelle 20),
aus 100 Proben des Typs L929 auf dem Substrat Stahl* sowie Titan* die Anzahl
der Zellen mit dem Algorithmus aus Kap. 6.4 in Kombination mit der hier vorge-
stellten CBS bestimmt und mit der von einem Experten ermittelten Zellzahl ver-
glichen (Tabelle 21).
Tabelle 20: Vergleich der Rechenzeit für den Separationsalgorithmus (Kap. 6.4) ohne CBS und
mit CBS auf einer i7 3,2GHz CPU mit Parallelverarbeitung.
Probe Berechnungs-zeit ohne CBS
(s)
Berechnungs-zeit mit CBS (s)
Speed- Up
Probe 1 12,4 1,9 6,53
Probe 2 5,8 2,1 2,76
Probe 3 4,4 0,9 4,89
1BR
2BR 1BR
2BR

6. Separation agglomerierter Zellen 65
Probe 4 11,8 1,9 6,21
Probe 5 2,1 0,6 3,50
Probe 6 4,1 1,1 3,73
Probe 7 1,1 0,4 2,75
Probe 8 2,0 0,5 4,00
Probe 9 6,2 0,5 12,40
Probe 10 15,1 0,4 37,75
Ø Geschwindigkeitszuwachs 8,45
Tabelle 21: Statistische Daten zur automatischen Zellzählung mit dem Separationsverfahren
aus Kap. 6.4 ohne und mit vorgeschalteter CBS
Zellzählung ohne vorgeschalteter CBS Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
2,41 1,90 0,19
1,96 (95%) 0,37
2,576 (99%) 0,49
3,29 (99,9%) 0,63
3,87 (99,99%) 0,74
Zellzählung mit vorgeschalteter CBS
Mittlere Abweichung zur Referenz [%]
Standard- Abweichung [%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Abweichung
1,27 0,87 0,09
1,96 (95%) 0,17
2,576 (99%) 0,22
3,29 (99,9%) 0,29
3,87 (99,99%) 0,34
Aus Tabelle 20 geht hervor, dass durch Vorschalten der CBS im Mittel ein
Speed-Up um den Faktor 8 im Vergleich zur Zellzählung ohne Einsatz der CBS
erzielt wird. Tabelle 21 zeigt, dass unter Verwendung einer Stichprobe von 100
Probenausschnitten für den Einsatz der CBS im Durchschnitt die Abweichung
von 2,41% +/-0,49% auf 1,27% +/-0,22% bei einem Konfidenzintervall von 99%
sinkt (Kap. 13.7).
Die für die Entscheidungsfunktion f(x,y) notwendigen Parameter wurden für die
Zelllinie L929 experimentell angepasst. Mit f(1.5,3) erhält man die geringste
Abweichung der automatisierten Zellzahlzählung im Vergleich zum Referenz-
wert. Die zellspezifischen Regeln zu Objektgröße und Kompaktheit (Kap. 6.1
Bedingung 1 und Bedingung 2) wurden speziell für die vorliegende Anwendung
aufgestellt und können bei Bedarf durch andere Regeln ersetzt werden, ohne
die Methode zu verändern. Hier dient das Verfahren als Ergänzung zu dem in
Kap. 6.4 vorgestellten Separationsverfahren, um die Rechenzeit zu verkürzen
sowie die Genauigkeit der Zellzählung zu steigern. Es können jedoch auch
Operationen an Objektverengungen unabhängig von anderen Algorithmen vor-
genommen werden, wobei in dem Fall die Rechenzeit im Millisekunden Bereich
liegt. Die CBS ist übertragbar auf andere Objekte und kann somit auch Ver-
wendung für andere Problemstellungen finden.

6. Separation agglomerierter Zellen 66
6.2 Klassifizierung segmentierter Zellbereiche
Nachdem die Zellen mit einer der in Kap. 5 beschriebenen Methoden segmen-
tiert und die CBS-Methode aus Kap. 6.1 ausgeführt wurde, erfolgt die Klassifi-
zierung der resultierenden Bereiche entweder als „Einzelzelle“ oder „Zellclus-
ter“. Die als „Zellcluster“ klassifizierten Zellbereiche werden dann abhängig vom
Zelltyp durch die in Kap. 6.4, 6.5 und 6.6 beschriebenen Trennverfahren sepa-
riert, damit sie individuell auswertbar sind. Untersuchungen zeigen, dass die
Klassifizierung mit Hilfe geometrischer Merkmale, z.B. Zellfläche oder Kom-
paktheit, nicht geeignet ist (Kap. 13.4). Ebenso ist eine farbliche Unterschei-
dung von Einzelzellen und Agglomeraten nicht möglich. Da Zellen der Linie
L929 oft Verengungen an den Zellkontaktstellen aufweisen, ist mit Hilfe einer
iterativen Ausführung der Erosion mit anschließendem Labeling eine Klassifizie-
rung der Zellbereiche durchführbar.
Die Klassifizierung erfolgt nacheinander für alle n segmentierten Zellbereiche
Z1, Z2, …, Zn, wobei jeder Zellbereich als endliche Pixelmenge aufgefasst wird.
Dazu wird ein Zellbereich Zi iterativ mit einem kreisförmigen Strukturelement B,
dessen Radius 1,5 Pixel beträgt, erodiert. Der erodierte Zellbereich
Zi,j+1 = ,( )B i jZ mit j = 0, 1,…, JMax (Gl. 8)
wird einem Labeling unterzogen. Resultieren mindestens zwei nicht zusam-
menhängende Zellbereiche mit einer Fläche ≥ AMin, bricht das Verfahren ab und
der Zellbereich wird als „Zellcluster“ eingestuft. Liefert das Labeling nur einen
zusammenhängenden Zellbereich, wird die Erosion mit anschließendem La-
beling wiederholt, bis zwei nicht zusammenhängende Zellbereiche entstehen
oder die maximale Anzahl Iterationen JMax erreicht ist. Bei JMax handelt es sich
um einen einstellbaren Parameter, wobei sich JMax = 25 als geeignet erweist.
Wenn die maximale Anzahl Iterationen erreicht wurde und nur ein zusammen-
hängender Zellbereich nach dem Labeling vorliegt, erfolgt die Klassifizierung
des Zellbereiches als „Einzelzelle“.

6. Separation agglomerierter Zellen 67
Abbildung 36: Die cyan gefärbte Kontur repräsentiert den ursprünglichen Zellbereich Z i. Die
grün gefärbten Konturen repräsentieren Zi,13 (das Ergebnis nach 13 Iterationen der Erosion).
Abb. 36 zeigt einen Zellbereich Zi,j, der nach 13 Iterationen der Erosion mit an-
schließendem Labeling zwei neue Zellbereiche liefert. Der Zellbereich wird folg-
lich als „Zellcluster“ klassifiziert.
Da sich agglomerierte Zellen vom Typ MC3T3 oft durch eine kompakte Geo-
metrie auszeichnen, ist eine Klassifizierung der Zellbereiche mit Hilfe der iterati-
ven Erosion nicht ideal. Durch Ergänzung der Methode mit nachfolgender Regel
lassen sich auch Zellbereiche des Typs MC3T3 in Cluster und Einzelzellen
klassifizieren.
Liegen nach der Kerndetektion, beschrieben in Kap. 6.3, mindestens 2 unter-
schiedliche Kernbereiche in einem als Einzelzelle klassifizierten Zellbereich vor,
wird nachträglich die Klassifikation in „Zellcluster“ geändert, da mit hoher Wahr-
scheinlichkeit eine Fehlklassifizierung vorliegt.
6.2.1 Ergebnisse
Zur Bewertung der Leistungsfähigkeit der vorgestellten Klassifikationsmethode
wurden in 40 Probenausschnitten (20 L929 und 20 MC3T3 Bilder) die Zellberei-
che durch einen Experten in Zellcluster und Einzelzellen eingeteilt und der au-
tomatischen Klassifizierung gegenübergestellt (Tabelle 22).
Tabelle 22: Vergleich der automatischen Klassifizierung der Zellbereiche in Einzelzellen mit der
Klassifizierung durch einen Experten; REF: Anzahl klassifizierter Zellen durch einen Experten;
FK: Anzahl falsch klassifizierter Zellbereiche durch die Software; Relativer Fehler: Relativer
Fehler der Klassifikation durch die Software
Proben Nr.
REF FK Relativer
Fehler (%) Proben
Nr. REF FK
Relativer Fehler (%)
1 L929 198 1 0,5 11 L929 234 1 0,4
2 L929 197 5 2,5 12 L929 262 5 1,9
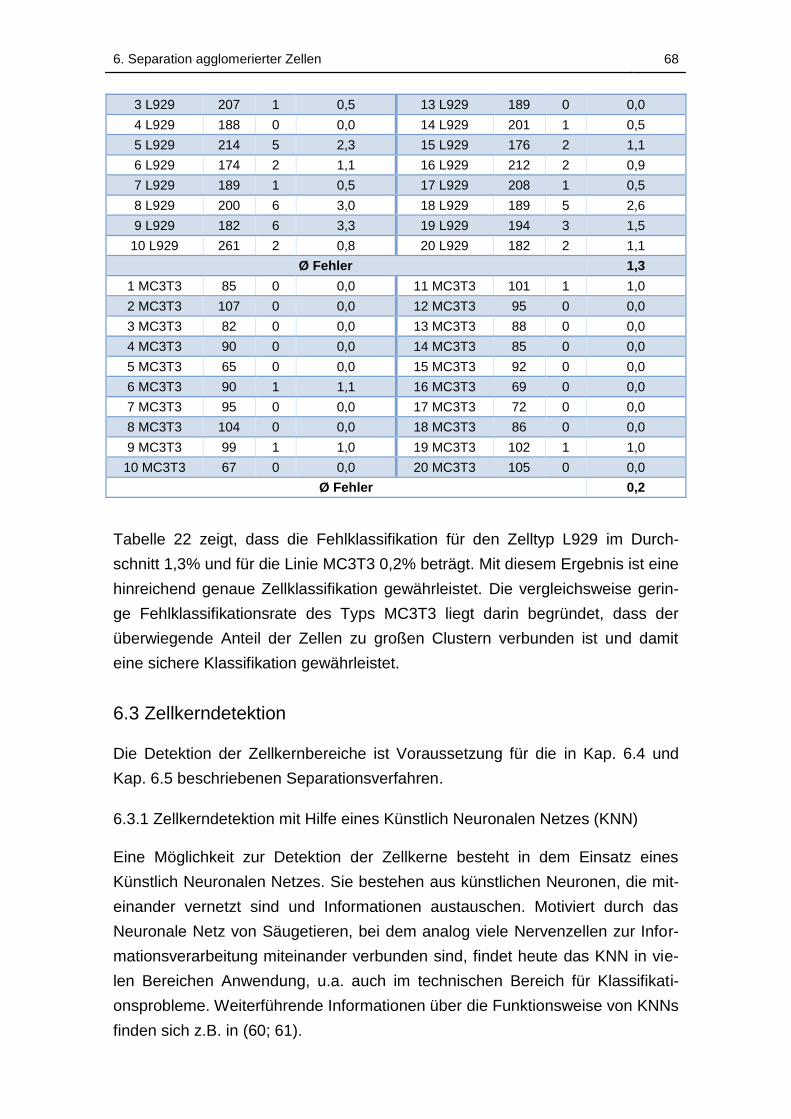
6. Separation agglomerierter Zellen 68
3 L929 207 1 0,5 13 L929 189 0 0,0
4 L929 188 0 0,0 14 L929 201 1 0,5
5 L929 214 5 2,3 15 L929 176 2 1,1
6 L929 174 2 1,1 16 L929 212 2 0,9
7 L929 189 1 0,5 17 L929 208 1 0,5
8 L929 200 6 3,0 18 L929 189 5 2,6
9 L929 182 6 3,3 19 L929 194 3 1,5
10 L929 261 2 0,8 20 L929 182 2 1,1
Ø Fehler 1,3
1 MC3T3 85 0 0,0 11 MC3T3 101 1 1,0
2 MC3T3 107 0 0,0 12 MC3T3 95 0 0,0
3 MC3T3 82 0 0,0 13 MC3T3 88 0 0,0
4 MC3T3 90 0 0,0 14 MC3T3 85 0 0,0
5 MC3T3 65 0 0,0 15 MC3T3 92 0 0,0
6 MC3T3 90 1 1,1 16 MC3T3 69 0 0,0
7 MC3T3 95 0 0,0 17 MC3T3 72 0 0,0
8 MC3T3 104 0 0,0 18 MC3T3 86 0 0,0
9 MC3T3 99 1 1,0 19 MC3T3 102 1 1,0
10 MC3T3 67 0 0,0 20 MC3T3 105 0 0,0
Ø Fehler 0,2
Tabelle 22 zeigt, dass die Fehlklassifikation für den Zelltyp L929 im Durch-
schnitt 1,3% und für die Linie MC3T3 0,2% beträgt. Mit diesem Ergebnis ist eine
hinreichend genaue Zellklassifikation gewährleistet. Die vergleichsweise gerin-
ge Fehlklassifikationsrate des Typs MC3T3 liegt darin begründet, dass der
überwiegende Anteil der Zellen zu großen Clustern verbunden ist und damit
eine sichere Klassifikation gewährleistet.
6.3 Zellkerndetektion
Die Detektion der Zellkernbereiche ist Voraussetzung für die in Kap. 6.4 und
Kap. 6.5 beschriebenen Separationsverfahren.
6.3.1 Zellkerndetektion mit Hilfe eines Künstlich Neuronalen Netzes (KNN)
Eine Möglichkeit zur Detektion der Zellkerne besteht in dem Einsatz eines
Künstlich Neuronalen Netzes. Sie bestehen aus künstlichen Neuronen, die mit-
einander vernetzt sind und Informationen austauschen. Motiviert durch das
Neuronale Netz von Säugetieren, bei dem analog viele Nervenzellen zur Infor-
mationsverarbeitung miteinander verbunden sind, findet heute das KNN in vie-
len Bereichen Anwendung, u.a. auch im technischen Bereich für Klassifikati-
onsprobleme. Weiterführende Informationen über die Funktionsweise von KNNs
finden sich z.B. in (60; 61).

6. Separation agglomerierter Zellen 69
Im Rahmen dieser Arbeit wurde die Eignung eines KNN zur Klassifizierung vor-
segmentierter Zellbereiche (Kap. 5.2) in drei Zellbestandteile, gemäß Kap.
5.2.2, untersucht (62). Dabei handelt es sich um das Ektoplasma (brauner
Randbereich der Zelle), äußere Endoplasma (hellblauer Zellbereich) und den
Kernbereich bzw. das Endoplasma, wobei sich im weiteren Projektverlauf her-
ausgestellt hat, dass nur die Information über die Position des Kernbereiches
benötigt wird.
6.3.1.1 Datenerzeugung für das KNN
Damit das KNN zwischen dem Kernbereich und der übrigen Zellfläche unter-
scheiden kann, müssen geeignete Merkmale gefunden werden. Untersuchun-
gen zeigen, dass die Farbzusammensetzung der Kernbereiche innerhalb eines
Bildes variiert, aber meistens einen Unterschied zur Färbung der restlichen Zel-
le aufweist (Kap. 13.5). Dem KNN werden vier Informationen zur Klassifizierung
bereitgestellt. Bei den ersten drei Merkmalen RGB
RV , RGB
GV , RGB
BV handelt es
sich um normierte Farbverhältnisse. Dazu werden die Schwerpunkte Rs, Gs und
Bs aus den lokalen Histogrammen nach folgender Vorschrift berechnet:
255
2550
0
1S k
kk
k
R m k
m k = 0 bis 255 (Gl. 9)
In (Gl. 9) entspricht mk der Anzahl Pixel an der Position k im Histogramm des
Rotkanals.
Die Schwerpunkte des Grün- und Blaukanals werden analog ermittelt. Die Nor-
mierung dient zur Kompensation von Helligkeitsschwankungen und erfolgt nach
der Vorschrift
RGB SR
S S S
RV
R G B , (Gl. 10)
RGB SG
S S S
GV
R G B , (Gl. 11)
RGB SB
S S S
BV
R G B. (Gl. 12)
Bei der vierten Inputinformation für das KNN handelt es sich um die Farbsätti-
gung S, da die Untersuchungen weiterhin zeigen, dass die Kernbereiche meis-
tens eine höhere Farbsättigung als die restliche Zellfläche aufweisen (Kap.

6. Separation agglomerierter Zellen 70
13.5). Eine Transformation in den HSV-Farbraum (63) ist hier vorteilhaft, da so
schnell auf die Farbsättigung eines Pixels zugegriffen werden kann.
Die vier Inputinformationen für das KNN werden in lokalen Fenstern mit der
Größe 20x20 Pixel berechnet (Abb. 37).
Abbildung 37: Durchlauf des Zellbereiches mit einem Fenster der Größe 20s20 Pixel zur Ermitt-
lung der vier Inputinformationen für das KNN
Dazu wird das kleinste umschließende Rechteck des aktuellen Zellbereiches
ermittelt und die Fenstermaske beginnend links oben iterativ durch diesen ver-
schoben. Bei jeder Iteration wird geprüft, ob mindestens 70% der Fensterfläche
durch Zellpixel gefüllt sind. Ist diese Bedingung erfüllt, werden die vier Inputin-
formationen für das KNN ermittelt, andernfalls erfolgt für diesen Fensterbereich
keine Klassifizierung. Die Fenstergröße ist variabel, jedoch für die vorliegende
Anwendung optimal mit 20x20 Pixel eingestellt. Die Verwendung eines kleine-
ren Fensters liefert zu wenige Pixel für eine sichere Klassifizierung, ein größerer
Bereich hingegen würde oftmals mehrere Zellbereiche beinhalten und so zu
keiner eindeutigen Klassifikation führen. Es lässt sich nicht vermeiden, dass
teilweise auch verschiedene Zellbereiche in einem Fenster der Größe 20x20
Pixel enthalten sind. Kap. 6.3.1.2 stellt eine Methode vor, so dass auch in die-
sem Fall eine eindeutige Klassifikation möglich ist.
6.3.1.2 Verbesserung der Klassifikationsleistung
Der Übergang verschiedener Zellbereiche, z.B. des Zellkerns zum umliegenden
äußeren Endoplasma, kann sich innerhalb eines Prüffensters befinden, so dass
ohne weitere Maßnahmen keine eindeutige Klassifizierung möglich ist, Abb. 38
(links).

6. Separation agglomerierter Zellen 71
Abbildung 38: Links: Prüfbereich zur Klassifizierung des eingeschlossenen Zellareals; Rechts:
Aufteilung des Prüfbereiches bei nicht eindeutiger Klassifizierung
In Abb. 38 (links) ist ein Prüfbereich dargestellt, in dem die für die Klassifizie-
rung benötigten Daten ermittelt werden. Da sowohl ein Teil dem Kernbereich
als auch dem äußeren Endoplasma zugeordnet ist, liefert das Netz keine ein-
deutige Klassifikation. Der Prüfbereich wird in diesem Fall automatisch in zwei
Bereiche aufgeteilt und erneut klassifiziert, Abb. 38 (rechts). Dieser Vorgang
wird maximal 2-mal wiederholt, da weitere Iterationen aufgrund zu kleiner Prüf-
bereiche zu unsicheren Klassifikationen führen. Auf diese Weise ist eine ge-
nauere Klassifizierung verschiedener Zellbestandteile, insbesondere an den
Übergängen verschiedener Zellbereiche, möglich.
6.3.1.3 Architektur und Lernparameter des KNN
Die Architektur eines KNN setzt sich standardmäßig aus einer Eingabe-, Aus-
gabe- und versteckten Schicht zusammen (64). Die Anzahl der Eingabeneuro-
nen ist durch die Anzahl der benötigten Merkmale zur Klassifizierung vorgege-
ben, in diesem Fall vier Neuronen, vgl. 6.3.1.1. Die Anzahl der Neuronen in der
Ausgabeschicht entspricht der Anzahl der vorliegenden unterschiedlichen Klas-
sen. Im Rahmen des Projektes sind es drei unterschiedliche Ausgabeneuronen,
vgl. 6.3.1. Zwischen Eingabe- und Ausgabeneuronen liegen zwei versteckte
Schichten mit jeweils vier Neuronen. Hierbei handelt es sich nach eingehenden
Untersuchungen um die optimale Topologie für diese Problemstellung (Abb.
39).
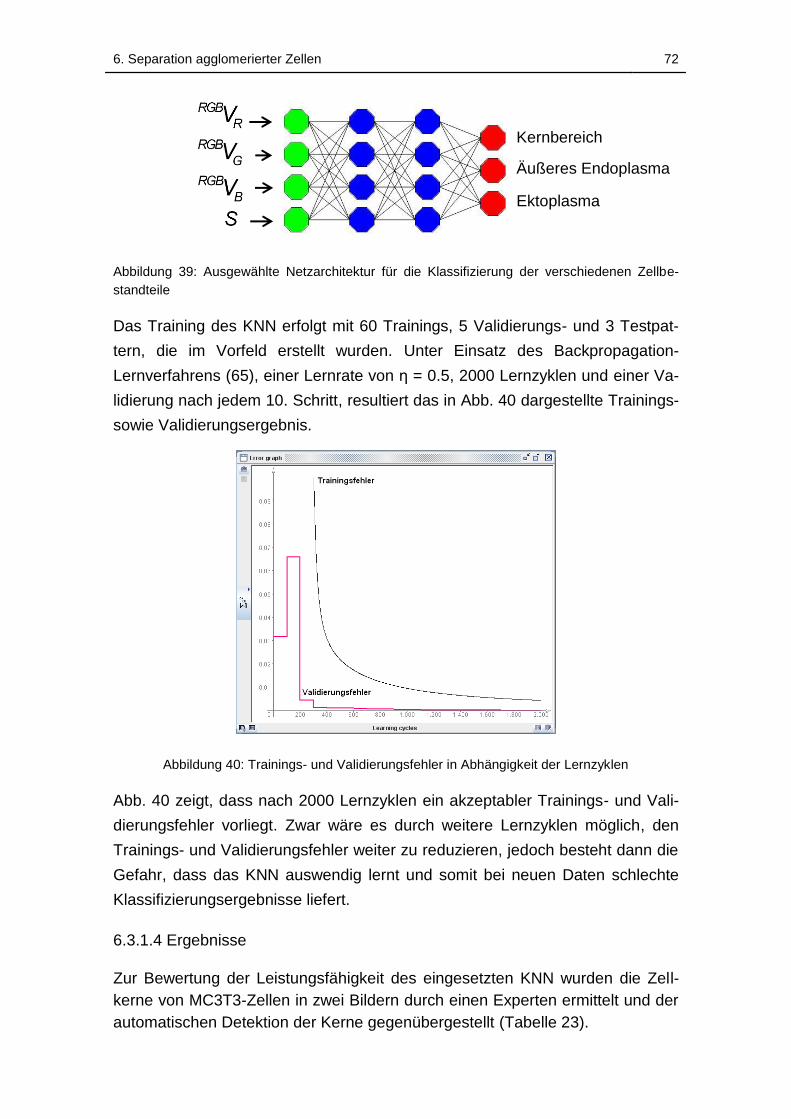
6. Separation agglomerierter Zellen 72
Abbildung 39: Ausgewählte Netzarchitektur für die Klassifizierung der verschiedenen Zellbe-
standteile
Das Training des KNN erfolgt mit 60 Trainings, 5 Validierungs- und 3 Testpat-
tern, die im Vorfeld erstellt wurden. Unter Einsatz des Backpropagation-
Lernverfahrens (65), einer Lernrate von η = 0.5, 2000 Lernzyklen und einer Va-
lidierung nach jedem 10. Schritt, resultiert das in Abb. 40 dargestellte Trainings-
sowie Validierungsergebnis.
Abbildung 40: Trainings- und Validierungsfehler in Abhängigkeit der Lernzyklen
Abb. 40 zeigt, dass nach 2000 Lernzyklen ein akzeptabler Trainings- und Vali-
dierungsfehler vorliegt. Zwar wäre es durch weitere Lernzyklen möglich, den
Trainings- und Validierungsfehler weiter zu reduzieren, jedoch besteht dann die
Gefahr, dass das KNN auswendig lernt und somit bei neuen Daten schlechte
Klassifizierungsergebnisse liefert.
6.3.1.4 Ergebnisse
Zur Bewertung der Leistungsfähigkeit des eingesetzten KNN wurden die Zell-
kerne von MC3T3-Zellen in zwei Bildern durch einen Experten ermittelt und der
automatischen Detektion der Kerne gegenübergestellt (Tabelle 23).
Kernbereich
Äußeres Endoplasma
Ektoplasma

6. Separation agglomerierter Zellen 73
Tabelle 23: Statistische Daten zur automatischen Kernzählung auf Basis eines KNN
Klassifizierung von L929 Zellen
Mittlere Abweichung
zur Referenz [%]
Standard- Abweichung [%]
Standard- fehler [%]
Mittlere Rechenzeit
Konfidenz- Intervall
Toleranz um Abweichung
14,1 4,40 0,44 418s
1,96 (95%) 0,89
2,576 (99%) 1,13
3,29 (99,9%) 1,45
3,87 (99,99%) 1,70
Tabelle 23 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten für die Linie L929 (Kap. 13.8) im Durchschnitt eine Abwei-
chung von 14,1% +/- 1,13% bei einem Konfidenzintervall von 99% zur Referenz
einstellt. Die Rechenzeit hängt von der Anzahl der im Bild vorhandenen Zellen
bzw. deren Zellfläche ab, beträgt hier im Durchschnitt für 273 Zellen 418 Se-
kunden.
6.3.2 Zellkerndetektion auf Basis des Histogram-Backprojection-Algorithmus
Die Zellkerne der Linien MC3T3 und L929 können farblich variieren, weisen
aber meistens einen schwachen Kontrast zu ihrer lokalen Umgebung auf (Abb.
41 und 42). Daher erscheint eine Detektion der Kerne mit Hilfe des HB-
Algorithmus für diese Aufgabe sinnvoll, da der Algorithmus zur Lokalisierung
von Objekten auf Basis ihrer Farbwerte dient.

6. Separation agglomerierter Zellen 74
Abbildung 41: MC3T3-Zellen auf dem Substrat Titan* bei 100-facher Vergrößerung mit vergrö-
ßerten Bildausschnitten verschiedenfarbiger Kernbereiche
Abbildung 42: L929-Zellen auf dem Substrat Stahl* bei 100-facher Vergrößerung mit vergrößer-
ten Bildausschnitten verschiedenfarbiger Kernbereiche
Die vergrößerten Ausschnitte in Abb. 41 und Abb. 42 zeigen die Farbvariation
für die Zellkernbereiche innerhalb eines Probenausschnittes. Damit eine Detek-
tion verschiedenfarbiger Kerne möglich ist, muss der HB-Algorithmus iterativ mit
verschiedenen im Vorfeld erstellten Farbtemplates ausgeführt werden. Für die
May-Grünwald angefärbten Zellen des Typs MC3T3 und L929 kommen zwei
erstellte Farbtemplates für die Detektion der Kernbereiche zum Einsatz, in Abb.
43 vergrößert dargestellt.
Abbildung 43: Vergrößerte Darstellung der erstellten Farbtemplates für die Detektion der Zell-
kernbereiche mittels HB-Algorithmus. Links: Template 1 zur Detektion dunkler Kernbereiche;
Rechts: Template 2 zur Detektion vergleichsweise heller Kernbereiche
Für jedes Template werden analog zur Zellsegmentierung, beschrieben in Kap.
5.2.4, templatespezifische Parameter eingestellt. Tabelle 24 zeigt die für diese
Anwendung optimalen Parameter für jedes Template.

6. Separation agglomerierter Zellen 75
Tabelle 24: Parametereinstellungen für die verwendeten Farbtemplates zur Zellkerndetektion
mittels HB-Algorithmus; ACMin: Minimal zulässige Kernfläche; c: Anzahl der verwendeten Farben
bei der Unterabtastung der verwendeten 24bit-RGB Bilder (2563 Farben) zur Reduktion der
Rechenzeit, vgl. (55)
Template L929 MC3T3
ACMin c ACMin c
1 150 7 150 6
2 250 8 250 8
Nach Ausführung des HB-Algorithmus resultieren entsprechend der Anzahl der
verwendeten Templates (Abb. 43) zwei Ergebnisbilder E1 und E2 (Abb. 44b,
Abb. 44c). Diese werden jeweils zur Erfassung der einzelnen Kernbereiche mit
weiteren Bildverarbeitungsverfahren bearbeitet. Die Bildverarbeitungsoperation
Closing (56) dient zum Schließen kleiner Segmentierungslücken und das an-
schließende Labeling zur individuellen Auswertung zusammenhängender Kern-
bereiche. Es resultieren für E1 x Kernbereiche C11, C12, …, C1x und für E2 y
Kernbereiche C21, C22, …, C2y, wobei ein Kernbereich als endliche Pixelmenge
und die Größe eines Kernbereiches, z.B. C11, als die Anzahl der enthaltenen
Pixel 11C aufgefasst wird.

6. Separation agglomerierter Zellen 76
Abbildung 44: a: Segmentierter Zellbereich (grüne Kontur); b: Ergebnisbild E1 des HB-
Algorithmus mit Template 1; c: Ergebnisbild E2 des HB-Algorithmus mit Template 2; d: Ergebnis
der Zellkerndetektion nach der Weiterverarbeitung von c. Bei den schwarz eingekreisten Kern-
bereichen handelt es sich jeweils um zwei Kerne, die jedoch nur als ein Kernbereich erkannt
wurden
Im Vergleich zur Zellsegmentierung mit Hilfe des HB-Algorithmus (Kap. 5.2.3)
folgen hier nun weitere Schritte. Zunächst wird geprüft, ob Schnittmengen der
Kernbereiche aus E1 und E2 nach folgender Vorschrift existieren:
1 1
1 2 0yx
i j
i j
C C . (Gl. 13)
Ist dies der Fall, wird nur der größere der beiden Kernbereiche, entweder C1i
oder C2j, nach der Vorschrift
max 1 , 2i jC C (Gl. 14)
übernommen.
a
b c
d

6. Separation agglomerierter Zellen 77
Die schwarzen Markierungen in Abb. 44d zeigen, dass teilweise zwei benach-
barte Kernbereiche als ein großer Kernbereich (gelbe Kontur) detektiert wurden,
da sie keine farbliche Unterscheidung aufweisen und zusammenhängend sind.
Oft ist jedoch die Kontur zwischen zwei Kernen durch eine Einschnürung ge-
kennzeichnet, ähnlich wie bei zwei aneinandergrenzenden Zellen, z.B. des
Typs L929. Daher ist eine Separation der detektierten Kernbereiche mit ausge-
prägten Einschnürungen hilfreich, um die Kerndetektion zu optimieren.
6.3.2.1 Separation der Zellkernbereiche zur Optimierung der Kerndetektion
Die Separation der Kernbereiche erfolgt in 2 Schritten. Zuerst werden Marker
der separierten Kernbereiche generiert. Dies erfolgt analog zur Klassifikation
der segmentierten Zellbereiche (Kap. 6.2) mit Hilfe einer iterativen Erosion. Der
Unterschied zum Verfahren in Kap. 6.2 liegt darin, dass die iterative Erosion
nicht beendet wird, sobald nach dem Labeling mindestens zwei neue Bereiche
entstanden sind, da weitere Separationen eines bereits getrennten Kernberei-
ches in kleinere Kernbereiche möglich sind.
Abbildung 45: Ergebnis der Markergenerierung für MC3T3-Kernbereiche, die Kontur der ur-
sprünglichen Kernbereiche ist gelb, die generierten Marker grün dargestellt
Abb. 45 verdeutlicht die Markergenerierung für MC3T3-Zellkernbereiche. Am
Beispiel des Kernbereiches CA9 (gelbe Kontur) wird der Separationsprozess
erläutert. Nachdem CA9 n-mal einer Erosion mit anschließendem Labeling un-
terzogen wurde, entstehen zwei separate Kernbereiche CA9,1 und CA9,2 (grüne
Konturen). Eine iterative Erosion der Kernbereiche CA9,1 und CA9,2 führt zu kei-
nen weiteren Separationen, so dass es sich hier um zwei Marker handelt, die im
CA1
CA2
CA3
CA6
CA5
CA4
CA7
CA8
CA9
CA10
CA11 CA12
CA16
CA15
CA13
CA14
CA9,1
CA9,2

6. Separation agglomerierter Zellen 78
weiteren Verlauf für die Rekonstruktion der Kernfläche dienen. Die Rekonstruk-
tion der Kernbereiche ist notwendig, da durch die Erosion ein Teil der Kernflä-
che eliminiert wurde, die im nächsten Schritt wieder hergestellt werden muss
(weiße Schraffur in Abb. 45). Für die Rekonstruktion der Kernbereiche kommt
die in Kap. 6.6 beschriebene Wachstumssimulation zum Einsatz. Der Unter-
schied zu Kap. 6.6 liegt darin, dass keine Zellbereiche aus dem Wachstums-
prozess resultieren sondern die optimierten Kernbereiche für den anschließen-
den Separationsvorgang innerhalb der Agglomerate. Der Algorithmus muss je-
doch nicht geändert werden, so dass hier für eine detaillierte Beschreibung des
Rekonstruktionsprozesses auf Kap. 6.6 verwiesen wird. Das beschriebene Ver-
fahren ist abschließend mit Hilfe eines Flussdiagramms schematisch darge-
stellt.
Abbildung 46: Flussdiagramm zum schematischen Ablauf der Kernbereichdetektion mit Hilfe
des HB Algorithmus

6. Separation agglomerierter Zellen 79
6.3.2.2 Ergebnisse
Zur Bewertung der Leistungsfähigkeit des hier vorgestellten Verfahrens wurden
in jeweils 100 Proben von L929- und MC3T3-Zellen die Kernbereiche durch
einen Experten ermittelt und der automatischen Kerndetektion gegenüberge-
stellt (Kap. 13.9). Tabelle 25 zeigt die statistische Auswertung für die Kernde-
tektion ohne anschließende Kernseparation an engen Stellen, Tabelle 26 hin-
gegen mit anschließender Kernseparation.
Tabelle 25: Statistische Daten zur automatischen Kernzählung auf Basis des HB-Algorithmus
L929-Kernzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
3,09 2,19 0,22
1,96 (95%) 0,43
2,576 (99%) 0,56
3,29 (99,9%) 0,72
3,87 (99,99%) 0,85
MC3T3-Kernzählung
Mittlere Abweichung zur Referenz [%]
Standard- Abweichung [%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Abweichung
7,58 4,12 0,41
1,96 (95%) 0,81
2,576 (99%) 1,06
3,29 (99,9%) 1,35
3,87 (99,99%) 1,59
Tabelle 26: Statistische Daten zur automatischen Kernzählung auf Basis des HB-Algorithmus
mit anschließender Kernseparation
L929-Kernzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
2,06 1,72 0,17
1,96 (95%) 0,34
2,576 (99%) 0,44
3,29 (99,9%) 0,56
3,87 (99,99%) 0,66
MC3T3-Kernzählung
Mittlere Abweichung zur Referenz [%]
Standard- Abweichung [%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Abweichung
3,52 2,36 0,24
1,96 (95%) 0,46
2,576 (99%) 0,61
3,29 (99,9%) 0,78
3,87 (99,99%) 0,92
Tabelle 25 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten für die Linie L929 ohne anschließende Kernseparation im
Durchschnitt eine Abweichung von 3,09% +/- 0,56% bei einem Konfidenzinter-
vall von 99% zur Referenz einstellt. Für Zellen des Typs MC3T3 ergibt sich oh-
ne anschließende Kernseparation unter Verwendung einer Stichprobe von 100

6. Separation agglomerierter Zellen 80
Probenausschnitten eine Abweichung von 7,58% +/- 1,06% bei einem Kon-
fidenzintervall von 99%.
Anhand Tabelle 26 ist zu erkennen, dass sich unter Verwendung einer Stich-
probe von 100 Probenausschnitten für die Linie L929 mit anschließender Kern-
separation im Durchschnitt eine Abweichung von 2,06% +/- 0,44% bei einem
Konfidenzintervall von 99% zur Referenz einstellt. Für Zellen des Typs MC3T3
ergibt sich mit anschließender Kernseparation unter Verwendung einer Stich-
probe von 100 Probenausschnitten eine Abweichung von 3,52% +/- 0,61% bei
einem Konfidenzintervall von 99%.
6.3.3 Diskussion der Ergebnisse
Die Detektion der Zellkerne vom Typ L929 mit Hilfe eines KNN (Kap. 6.3.1)
weist mit 14,1% eine hohe Abweichung zum Referenzwert auf. Zwar wurden die
meisten Kernbereiche vom KNN ermittelt, jedoch werden benachbarte Kerne oft
in Form eines zusammenhängenden Bereiches zurückgegeben und somit nicht
einzeln detektiert. Die benötigte Rechenzeit ist ebenfalls hoch. Abhängig von
der Anzahl der Zellen in einem Bild sind ca. vier bis acht Minuten für die Zell-
kerndetektion notwendig. Die sehr hohe Rechenzeit liegt darin begründet, dass
in einem Bild sehr viele Klassifikationen mit dem KNN durchgeführt werden. Ein
zu klassifizierender Bereich beträgt 20x20 Pixel, wobei ein Bild insgesamt 1376
x 1032 Pixel beträgt, so dass theoretisch bis zu 3550 Klassifizierungen pro Bild
durchgeführt werden (66).
Ein weiterer Nachteil ist der Umstand, dass das KNN mit den in Kap. 6.3.1 vor-
gestellten Inputinformationen auf die hier verwendete Zellfärbung trainiert wur-
de. Kommt nun eine andere Färbemethode zum Einsatz ist ein erneutes Trai-
ning des KNN sehr wahrscheinlich notwendig.
Der Vorteil bei Verwendung eines KNN liegt darin, dass neben der Klassifizie-
rung von Kernbereichen auch andere Zellbereiche, z.B. das Ektoplasma, bei
Bedarf detektiert werden können, sofern dem Netz Merkmale zur Differenzie-
rung der unterschiedlichen Zellbereiche vorliegen. Die Ausführungsgeschwin-
digkeit würde sich dadurch nicht signifikant ändern.
Die Detektion der Zellkerne auf Basis des Histogram-Backprojection-
Algorithmus, beschrieben in Kap. 6.3.2, liefert für die Linie MC3T3 eine Abwei-
chung von 3,52% und für die Linie L929 eine Abweichung von 2,06% zum Refe-
renzwert. Bei den MC3T3-Zellen entsteht eine höhere Abweichung im Vergleich
zum Typ L929, da die Kernbereiche in großen, kompakten MC3T3-Clustern
größer ausfallen und einer stärkeren farblichen Variation unterliegen.

6. Separation agglomerierter Zellen 81
Im Vergleich zum Einsatz eines KNN liefert das Kerndetektionsverfahren auf
Basis des Histogram-Backprojection-Algorithmus jedoch deutlich bessere Er-
gebnisse, und darüber hinaus beträgt die Rechenzeit für einen Probenauss-
schnitt im Durchschnitt mit 10,7s nur 2,6% der Rechenzeit bei Verwendung ei-
nes KNN.
Eine weitere geeignete Methode zur Separation von Zellkernen ist die Wasser-
scheidentransformation (67) basierend auf der inversen Distanztransformation
der segmentierten Zellbereiche (68).
Das Verfahren ist gut auf Kernbereiche, die durch eine runde Geometrie ge-
kennzeichnet sind, anwendbar. Für Kernbereiche des Typs MC3T3 hingegen
resultieren aufgrund der starken Variation der Morphologie schlechtere Ergeb-
nisse im Vergleich zum Verfahren, beschrieben in Kap. 6.3.2 (Abb. 47).

6. Separation agglomerierter Zellen 82
Abbildung 47: a: Zellcluster vom Typ MC3T3; b: Kerndetektion mit Hilfe des Histogram Backpro-
jection Algorithmus und anschließendem Closing. Nicht korrekt detektierte Kernbereiche sind
durch rote Pfeile gekennzeichnet; c: Inverse Distanztransformation von b; d: Wasserscheiden-
transformation angewendet auf c (Schwellwert: 10). Übersegmentierte Bereiche sind durch
Pfeile gekennzeichnet; e: Inverse Distanztransformation von b; d: Wasserscheidentransformati-
on angewendet auf c (Schwellwert: 15). Nicht alle Kernbereiche wurden korrekt separiert, ge-
kennzeichnet durch die Pfeile; f: Segmentierte Kernbereiche mit der Methode beschrieben in
Kap. 6.3.2
Abb. 47a zeigt ein Cluster vom Typ MC3T3. Die Anwendung des Histogram
Backprojection Algorithmus gefolgt von der Closing-Methode führt zum Ergeb-
nis in Abb. 47b. Allerdings werden nicht alle Kerne korrekt detektiert, gekenn-
zeichnet durch die roten Pfeile. Das Ergebnis der Wasserscheidentransformati-
on (Abb. 47d) auf Basis der inversen Distanztransformation (Abb. 47c) zeigt
eine Übersegmentierung, ebenfalls durch rote Pfeile gekennzeichnet. Eine An-
passung des Wasserscheiden-Schwellwertes reduziert zwar die Übersegmen-
a b c
d e f

6. Separation agglomerierter Zellen 83
tierung, allerdings werden dann nicht mehr alle Kernbereiche voneinander ge-
trennt segmentiert (Abb. 47e, rote Pfeile).
Im Vergleich führt die Kerndetektion mit Hilfe des in Kap. 6.3.2 vorgestellten
Verfahrens zu keinen ungewollten Kernseparationen.
Aufgrund der Farbvariation und des geringen Kontrastes der Zellkerne zu um-
gebenden Zellbereichen sind Bildverarbeitungsverfahren wie z.B. Schwellwert-
verfahren, Kantenfilter und modellbasierte Verfahren für diese Aufgabenstellung
ungeeignet, da sie aufgrund ihrer Funktionsweise einen guten Kontrast des zu
detektierenden Objektes zur Umgebung voraussetzen. Im Gegensatz dazu ist
eine sichere Kerndetektion trotz variierender Farbgebung mit dem Histogram-
Backprojection-Algorithmus möglich, wenn mehrere Farbtemplates erstellt wur-
den, die die Farbvariation der Kerne abdecken.
6.4 Zellseparation in Clustern mit Verengungen an Zellkontaktstellen
Nach der Klassifikation der Zellbereiche in Einzelzellen sowie Cluster (Kap. 6.2)
und der Detektion der Kernbereiche, wird nun ein Verfahren vorgestellt, dass
mit Hilfe der ermittelten Kernbereiche die in den Clustern enthaltenen Zellen
separiert (69; 70). Voraussetzung für die erfolgreiche Separierung der zusam-
mengewachsenen Zellen ist, dass die zu untersuchende Zelllinie meistens Ver-
engungen an den Zellkontaktstellen bildet. Diese Bedingung ist z.B. von Zellen
der Linie L929 erfüllt, hingegen weisen die Agglomerate der Linie MC3T3 oft
keine Verengungen an den Zellkontaktstellen auf, so dass für diese ein anderes
Separationsverfahren erforderlich ist (Kap. 6.5).
6.4.1 Lokalisierung der Verengungen an den Zellkontaktstellen
Die Lokalisierung der Verengungen an den Zellkontaktstellen innerhalb der
Cluster geschieht über die Berechnung der so genannten dominanten Kontur-
punkte. Das in (44) beschriebene Verfahren wurde jedoch erweitert. Der erste
Schritt besteht in der Berechnung des Abstandes für jeden Konturpunkt eines
Clusters in eine vorgegebene Richtung zum umschließenden Rechteck. Des
Weiteren wird von jedem Konturpunkt der nächste Konturpixel daraufhin ge-
prüft, ob dieser in Richtung zur aktuell betrachteten Seite des umschließenden
Rechtecks Teil der Zelle ist. Ist dies der Fall, liefert die Prüffunktion den Wert
true, andernfalls false zurück. Somit werden nur die Punkte als dominante Kon-
turpunkte (DCP) definiert, die einerseits ein lokales Minimum der Abstandsfunk-
tion widerspiegeln und andererseits den Wert true von der Prüffunktion erhal-

6. Separation agglomerierter Zellen 84
ten. Die geschilderte Vorgehensweise wird für die übrigen drei Richtungen
(oben, unten, links) wiederholt.
Abbildung 48: Links: Segmentierter L929-Cluster (blaue Kontur); Rechts: Ermittelte DCPs
(blaue Kreisstrukturen) für den im Bild segmentierten Cluster (orange Kontur). Der rote Pfeil
kennzeichnet einen für die Separation ungeeigneten DCP
In Abb. 48 (Rechts) sind in der Farbe blau für alle vier Richtungen die ermittel-
ten DCPs eingezeichnet. Die Bestimmung der Maxima erfolgt hier im Vergleich
zu (44) jedoch anders. Es wird nur der Konturpixel als lokales Maximum defi-
niert, bei dem sich der Abstand von n vorherigen Konturpixeln zu einer Seite
des umschließenden Rechtecks monoton vergrößert und für die nachfolgenden
n Konturpunkte monoton verringert hat (Abb. 49).
Abbildung 49: Definition eines Konturpixels als lokaler maximaler Abstand zum Rand. Zunächst
liegen n Konturpunkte vor dem Maximum, deren Abstände sich zur linken Seite des umschlie-
ßenden Rechtecks vergrößern. Nach dem Maximum folgen n Konturpunkte, deren Abstände
sich zur linken Rechteckseite verkleinern
Die Vorgehensweise liegt darin begründet, dass die Clusterkontur aufgrund der
Segmentierung oft stufig aufgebaut ist und so eine Bestimmung der lokalen
Maxima durch digitale Ableitung unmöglich macht. Die Anzahl n der notwendi-
gen Stufen, bei denen sich monoton der Abstand zum umschließenden Recht-
eck vergrößern muss, ist parametrisiert und abhängig vom Zelltyp. Für den hier
vorliegenden Zelltyp L929 ist n=3 eine gute Wahl. So kann die Sensibilität des
Maximum
Abstand verkleinert sich n-mal
Abstand vergrößert sich n-mal
Ungeeigneter
DCP

6. Separation agglomerierter Zellen 85
Algorithmus festgelegt und je nach Bedarf mehr oder weniger DCPs erzeugt
werden. Wie in Abb. 48 zu sehen ist, kann der Fall eintreten, dass DCPs an
ungeeigneten Konturabschnitten erzeugt werden. Demnach wird eine Methode
zur Verbindung der korrekten DCPs benötigt.
6.4.2. Berechnung der kürzesten Pfade zwischen benachbarten
Zellkernbereichen
Die Herausforderung liegt nun darin, die DCPs an den aus biologischer Sicht
optimalen Stellen miteinander zu verbinden, so dass eine sinnvolle Zelltrennung
entsteht. Dazu werden zunächst von den gefundenen Zellkernen im Cluster die
Schwerpunkte berechnet. Ausgehend von dem links liegenden Schwerpunkt
wird der kürzeste Weg innerhalb des Zellbereiches zum nächstgelegenen Zell-
kernschwerpunkt bestimmt und so fortfahrend. Dazu dient der A*-Algorithmus
(71; 72), der häufig auch in der Spielebranche Anwendung findet (Abb. 50).
Abbildung 50: Mit dem A*-Algorithmus berechneter Pfad (weiß gefärbt) durch die Zellkern-
schwerpunkte (schwarze Kreuze) im Cluster
Dabei müssen mögliche Hindernisse wie z.B. innenliegende Löcher in der Zelle
berücksichtigt werden, denn der kürzeste Pfad soll Hindernisse nicht durch-
kreuzen. Der Algorithmus stellt ausgehend vom Startpunkt für alle Nachbarpixel
eine Kostenfunktion auf:
( , ) ( , ) ( , ) ( . 15)F x y G x y H x y Gl
G(x,y) beschreibt die Kosten, vom Startpunkt ausgehend zum aktuellen Punkt,
und H(x,y) die geschätzten Kosten vom aktuellen Punkt zum Zielpunkt. Abb. 51
und Abb. 52 zeigen anhand eines Beispiels die Vorgehensweise bei der Be-
rechnung des kürzesten Pfades zwischen zwei Punkten unter Berücksichtigung
von Hindernissen.

6. Separation agglomerierter Zellen 86
Abbildung 51: Ausgangssituation für die Berechnung des kürzesten Pfades von der Startpositi-
on (grünes Quadrat) zur Zielposition (rotes Quadrat) unter Berücksichtigung von nicht passier-
baren Hindernissen (blaue Quadrate). Für jedes Nachbarfeld werden die Kosten F, G und H
angegeben.
Abb. 51 zeigt anhand des Nachbarfeldes rechts vom Startpunkt die Zuordnung
der nummerischen Werte zu den Kosten F, G und H. Das Nachbarfeld erhält für
G den Wert 10, da es sich unmittelbar neben dem Startpunkt befindet. Diagonal
angrenzende Nachbarfelder erhalten hingegen den Wert G=14. H erhält den
Wert 30, da die geschätzte Distanz zum Zielpunkt 3 Felder beträgt. Die Kosten
für H(x,y) werden in diesem Fall über den Manhatten-Block Abstand berechnet,
da die Rechenzeit erheblich kürzer ist als im Vergleich zum euklidischen Ab-
stand. Somit ergeben sich für das Nachbarfeld rechts vom Startpunkt Gesamt-
kosten von F = 10 + 30 = 40. Nach Berechnung der Kosten für jeden Nachbar-
punkt wird der Nachbar mit den geringsten Gesamtkosten ausgewählt und die
Prozedur wiederholt. Ein wichtiger Aspekt ist, dass jeder untersuchte Punkt ei-
nen Zeiger auf seinen Vorgängerpunkt erhält. So kann der Weg vom Zielpunkt
zum Startpunkt zurückverfolgt werden. Der ermittelte Pfad vom Start- zum Ziel-
punkt ist in Abb. 52 durch die roten Kreise in den Feldern dargestellt.

6. Separation agglomerierter Zellen 87
Abbildung 52: Berechneter Pfad vom Ausgangspunkt (grünes Quadrat) zum Zielpunkt (rotes
Quadrat) unter Berücksichtigung nicht passierbarer Hindernisse (blaue Quadrate). Die zu dem
kürzesten Pfad gehörenden Felder sind durch einen roten Kreis gekennzeichnet.
Ist der kürzeste Pfad zwischen zwei Zellkernen berechnet, werden von den
Pfadpunkten nur die übernommen, die sich außerhalb der Zellkernbereiche be-
finden, da eine Zelltrennung durch einen vorhandenen Zellkern keinen Sinn
macht.
6.4.3 Generierung der Zellseparationslinien innerhalb der Cluster
Zuerst werden für die Pfadpunkte zwischen zwei Zellkernen die Abstände zu
allen DCPs des aktuellen Clusters berechnet und der erste DCP übernommen,
der den kürzesten Abstand zu einem Pfadpunkt außerhalb der Kernregion auf-
weist (Abb. 53, DCP 1). Der Pfadpunkt P und der Konturpunkt DCP1 bilden den
Anfangs- und Endpunkt des ersten Abschnittes der Trennlinie.
Abbildung 53: Separation zweier zusammengewachsener Zellen. Zeichenerklärung siehe Text
DCP1
Kern-Schwerpunkt 1
Erster Abschnitt der Trennlinie
Zweiter Abschnitt der Trennlinie
DCP2
Kern-Schwerpunkt 2 Pfadpunkt mit kleinstem
Abstand zu DCP 1
P

6. Separation agglomerierter Zellen 88
Der erste Abschnitt der Trennlinie muss allerdings die Bedingung erfüllen, kei-
nen Zellkernbereich zu schneiden. Schneidet der erste Linienabschnitt den
Kernbereich wird er wieder verworfen und das nächste Punktepaar (DCP und
Pfadpunkt) mit dem niedrigsten Abstand gesucht. Dieser Vorgang wiederholt
sich, bis ein gültiger erster Trennlinienabschnitt erzeugt wurde.
Nach Erzeugen des ersten Trennlinienabschnittes muss der zweite Trennlinien-
abschnitt, bestehend aus dem nächstgelegenen DCP auf der anderen Seite des
Pfades, gefunden werden, der mit demselben Pfadpunkt verbunden wird und
die Zelltrennung vervollständigt. Um zu prüfen, ob ein DCP auf der anderen
Seite des Pfades liegt, wird das Skalarprodukt der beiden potentiellen Tren-
nungsvektoren gebildet, mit der Bedingung, dass das Ergebnis < 0 sein muss.
Nicht in jedem Fall muss es einen passenden zweiten DCP geben. Es kann der
Fall eintreten, dass alle Trennlinien durch einen Zellkern führen und somit nicht
verwendet werden können. Alternativ wird dann der Pfadpunkt auf der Hälfte
des Weges zwischen den beiden Kernen selektiert und von dort aus der nächst
liegende Konturpunkt auf der einen Seite des Pfades sowie derjenige auf der
anderen Seite des Pfades bestimmt. Wenn die beiden Abschnitte der Trennlinie
keinen Zellkernbereich schneiden, handelt es sich um den neuen Grenzbereich
der Zellen, ansonsten werden die Trennlinienabschnitte zum nächstgelegenen
Konturpunkt geprüft, solange, bis eine gültige Trennlinie erzeugt wurde.
Für eine schematische Übersicht der beschriebenen Methode dient folgendes
Flussdiagramm.

6. Separation agglomerierter Zellen 89
Abbildung 54: Flussdiagramm zum schematischen Ablauf der Zellseparation mit der in Kap. 6.4
beschriebenen Methode
6.4.4 Ergebnisse
Zur Beurteilung der Genauigkeit bezüglich der generierten Einzelzellflächen
innerhalb von Agglomeraten des in Kap. 6.4 vorgestellten Separationsverfah-
rens ist eine Referenzsegmentierung notwendig. Dazu sind durch einen Exper-
ten aus 10 verschiedenen Proben 100 in verschiedene Cluster verbundene
L929 Zellen manuell segmentiert worden. Mit Hilfe des Jaccard-Koeffizienten
kann nun eine Aussage über die durchschnittliche Abweichung der automatisch
generierten Zellflächen zu den Referenzzellflächen erfolgen. Der Durch-
schnittswert des Jaccard-Koeffizienten auf Basis von 100 aus Clustern separier-
ten L929 Zellen für den Auflicht-Hellfeld Mikroskopkontrast beträgt 0,8. Vor der
Zellseparation in den Clustern müssen alle Zellbereiche zuvor mit einem der in
Kap. 5 vorgestellten Methoden segmentiert werden. Dazu kommt hier das
schwellwertbasierte Verfahren, beschrieben in Kap. 5.2.1, zum Einsatz.
Zur Bewertung der Genauigkeit bezüglich der Zellzählung des vorgestellten Se-
parationsverfahrens wurden 100 verschiedene Probenausschnitte der Linie
L929 aufgenommen und die automatische Zellzählung der Referenzzählung

6. Separation agglomerierter Zellen 90
durch eine Laborfachkraft gegenübergestellt (Kap. 13.7). Die statistische Aus-
wertung ist in Tabelle 27 hinterlegt.
Tabelle 27: Statistische Daten zur automatischen Zellzählung unter Verwendung des in Kap.
6.4 beschriebenen Verfahrens
L929-Zellzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
2,41 1,90 0,19
1,96 (95%) 0,37
2,576 (99%) 0,49
3,29 (99,9%) 0,63
3,87 (99,99%) 0,74
Aus Tabelle 27 geht hervor, dass sich unter Verwendung einer Stichprobe von
100 Probenausschnitten für die Linie L929 im Durchschnitt eine Abweichung
von 2,41% +/- 0,49% zur Referenz bei einem Konfidenzintervall von 99% ein-
stellt. Die hier vorgestellte Separationsmethode weist Probleme beim Trennen
von Zellen auf, die sich gerade im Teilungsprozess befinden (Abb. 55, gelb um-
randete Zellen).
Abbildung 55: L929 Zellen auf dem Material Stahl*. Bei den gelb markierten Bereichen handelt
es sich um Zellen, die sich gerade in einem Teilungsprozess befinden
Diese Zellen weisen keinen oder einen sehr schwachen farblichen Kontrast des
Kernbereiches zu den umgebenen Zellarealen auf. Da die hier beschriebene
Methode für eine korrekte Separation der Cluster u.a. auf die darin vorhande-
nen Kernbereiche zugreifen muss, ist ein erkennbarer farblicher Kontrast der
Kernbereiche jedoch Voraussetzung für eine korrekte Separation. In Kombina-
tion mit der CBS-Methode (Kap. 6.1) können somit die sich im Teilungsprozess
befindlichen Zellen ebenso separiert werden, so dass dadurch eine Reduktion
der Abweichung zur Referenzzählung von 2,41% auf 1,27% erfolgt (Tabelle 28).

6. Separation agglomerierter Zellen 91
Tabelle 28: Statistische Daten zur automatischen Zellzählung unter Verwendung des in Kap.
6.4 beschriebenen Verfahrens mit vorgeschalteter CBS
L929-Zellzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
1,27 0,87 0,09
1,96 (95%) 0,17
2,576 (99%) 0,22
3,29 (99,9%) 0,29
3,87 (99,99%) 0,34
Tabelle 28 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten für die Linie L929 mit vorgeschalteter CBS im Durchschnitt
eine Abweichung von 1,27% +/- 0,22% zur Referenz bei einem Konfidenzinter-
vall von 99% einstellt.
6.5 Zellseparation in Clustern ohne Verengungen an
Zellkontaktstellen
6.5.1 Wachstumssimulation
Befinden sich mehrere Objekte mit einem visuell erfassbaren Merkmal in einem
Objektcluster und sind die einzelnen Objektkonturen von Interesse kann die hier
beschriebene Wachstumssimulation auf Basis der Objektmerkmale zum Einsatz
kommen.
Die Simulation des Zellwachstums erfolgt nacheinander für alle n im Bild klassi-
fizierten Objektcluster A1, A2, …, An, wobei jeder Cluster Ai als endliche Pixel-
menge aufgefasst wird. Die Klassifikation eines Objektbereiches als Cluster
erfolgt, wenn in diesem mehr als ein Objektmerkmal, z.B. Kerne, enthalten sind.
Für jedes Cluster wird ein Zielbild Ti zur Dokumentation des Wachstumsprozes-
ses erzeugt. Die Größe von Ti entspricht der Größe des kleinsten umschließen-
den Rechtecks von Ai. Allen Pixeln in Ti wird zu Beginn der Farbwert (R, G, B) =
(255, 255, 255) zugeordnet. Die Pixelmenge i i iS T A erhält den Farbwert (0,
0, 0). Innerhalb von Ai befinden sich mi nicht zusammenhängende Objektmerk-
malsbereiche ,1 ,2 ,, ,...,ii i i mK K K . Alle Mengen , ,i j i i jC T K mit j = 1, 2, …, mi sollen
Elemente von Li sein (Abb. 56).

6. Separation agglomerierter Zellen 92
Abbildung 56: Zwei Objektcluster mit jeweils unterschiedlicher Anzahl an Objektmerkmalen im
Bild
Allen Elementen in Li wird jeweils eine Marke zugeordnet, die aus dem Index j
wie folgt resultiert:
2( . 17)
256j
jR Gl
2mod 256( .18)
256j
jG Gl
2mod 256 mod 256 ( .19)jB j Gl
So ist eine Unterscheidung von maximal 3256 2 (16.777.214) Objekten in ei-
nem Cluster möglich.
Unter der Größe eines Objektmerkmalbereiches ,i jC ist die Anzahl der Bildpunk-
te ,i jC zu verstehen. Die in Li enthaltenen Elemente werden ihrer Größe nach
absteigend sortiert, so dass der Wachstumsprozess mit dem größten Objekt-
merkmal startet. Mit Hilfe eines Parameters p {1, 2, …, 10} lässt sich die
Wachstumsgeschwindigkeit pi,j für jedes Objektmerkmal abhängig von seiner
Größe mit
pi,j = , , , ,min max
, ,max min
i j i j i j i j
i j i j
p C C C C
C C
berechnen. (Gl. 20)
pi,j kann folgende Werte annehmen:
S1
S2 T1 T2
C1,1
C1,2
C1,3
C2,1
C2,2
C2,3
C2,4
C2,5
A1
A1

6. Separation agglomerierter Zellen 93
(Gl. 21)
Für den unwahrscheinlichen Fall, dass alle in einem Cluster enthaltenen Ob-
jektmerkmale die gleiche Größe aufweisen, wird die Wachstumsgeschwindig-
keit auf pi,j = 1 gesetzt.
Das Verfahren arbeitet iterativ und beginnt die Wachstumssimulation im Cluster
Ai mit dem größten Element ,1iC aus Li. Dazu werden in Ti die Nachbarpixel
aller Elemente von ,1iC in Form einer 4er-Nachbarschaftsbeziehung daraufhin
geprüft, ob sie innerhalb des Clusters Ai liegen und keinem Objektmerkmal zu-
geordnet sind. Erfüllen alle Nachbarpixel die Bedingungen bilden sie die
Punktmenge Ni,1,k mit k = 1, 2, …, pi,1. Die Bedeutung der Indizes von Ni,j,k ist
wie folgt:
i: Index des ausgewählten Objektclusters
j: Index des ausgewählten Objektmerkmals aus dem i.ten Cluster
k: Aktuelle Wachstumsiteration mit k = 1, 2, …, pi,j
Im Allgemeinen kann Ni,j,k als Flächenzuwachs von Ci,j gesehen werden, sche-
matisch in Abb. 57 dargestellt.
Abbildung 57: Links: Die Objektmerkmale C1,1 und C1,2 befinden sich im Wachstumsprozess.
Der Parameter p ist auf p = 2 gesetzt. Bei Verwendung von Gl. 21 resultiert p1,1 = 2 und p1,2 = 1.
Rechts: Zweite Iteration des Wachstums. Die weiße Linie repräsentiert die Grenze der beiden
Objekte, nachdem kein Wachstum mehr möglich ist.
Abb. 57 zeigt, dass die Nachbarpixel N1,1,1 des Objektmerkmals C1,1 die ge-
nannten Bedingungen erfüllen und Teil des neuen Objektmerkmals
C1,1 (4 pixel)
Agglomerat A1
N1,1,1
N1,1,2
C1,2 (2 pixel)
N1,2,1
C1,3 (25 pixel)
N1,3,1
N1,3,2
C1,4 (12 pixel)
N1,4,1
, , max
1, , , , ,min max
1 , , min
p für c ci j i j
p p p für C C Ci j i j i j i j i j
für C Ci j i j

6. Separation agglomerierter Zellen 94
1,3 1,1,1 1,1C N C werden. Das neue Objektmerkmal wächst ein weiteres Mal (p1,1
= 2), wobei das Ergebnis in C1,3 überschrieben wird.
1,3 1,1,2 1,3:C N C
Ist das Wachstum von 1,1C beendet, wird C1,3 als Element in Li hinzugefügt.
Danach beginnt das Wachstum von C1,2. Die Nachbarpixel N1,2,1 von C1,2 erfül-
len die genannten Bedingungen und werden Teil des neuen Objektmerkmals
1,4 1,2,1 1,2C N C . Die Wachstumsprozedur wird in diesem Fall nicht wiederholt
da p1,2 auf 1 gesetzt ist. Das Ergebnis C1,4 kommt als Element in Li hinzu. Li
beinhaltet nun 4 Elemente C1,2, C1,2, C1,3 und C1,4. Abb. 57 (Rechts) zeigt das
Wachstum von C1,3 und C1,4. Zunächst wächst C1,3 wieder in zwei Iterations-
schritten:
1,5 1,3,1 1,3C N C
1,5 1,3,2 1,5:C N C
wobei das Ergebnis C1,5 als Element in Li hinzugefügt wird. Daraufhin erfolgt
das Wachstum von C1,4. Das Ergebnis 1,6 1,4,1 1,4C N C wird ebenso als Ele-
ment in Li hinzugefügt. Nun ist kein weiteres Wachstum der beiden ursprüngli-
chen Objektmerkmale möglich, die weiße Linie in Abb. 57 repräsentiert die
Grenze der beiden gewachsenen Objekte.
Im Allgemeinen erfolgt die Wachstumssimulation nacheinander für die nächsten
Elemente aus Li, bis eine näher beschriebene Abbruchbedingung des Algorith-
mus erfüllt ist. Für den Abbruch des Verfahrens stehen zwei Variablen u und v
bereit. u wird mit mi (Anzahl nicht zusammenhängender Objektmerkmale von Ai)
und v mit 0 initialisiert. Erhält Li nach Prüfung aller Nachbarpixel ein neues Ele-
ment , 1ii mC , wird die Variable u um eins inkrementiert. Die Variable v erhöht
sich nach der Nachbarschaftsprüfung, unabhängig davon, ob Ci,j gewachsen ist,
immer um eins. Der Algorithmus bricht ab, wenn v den Wert von u angenom-
men hat. Abb. 58 zeigt schematisch den Ablauf des Wachstumsprozesses.

6. Separation agglomerierter Zellen 95
Abbildung 58: Schematischer Ablauf des Wachstumsprozesses. Die in Li enthaltenen
Objektmerkmale werden durch numerische Werte für jeden Iterationsschritt repräsentiert. Die
Pfeile zeigen auf das nach dem Wachstumsprozess neu entstandene Objektmerkmal.
Weiterführende Erklärungen sind im nachfolgenden Text enthalten
Abb. 58 verdeutlicht schematisch den Ablauf des Wachstumsprozesses. Zu
Beginn liegen beispielsweise zwei Elemente Ci,1 und Ci,2 in Li vor (Iterations-
schritt v=0). Bei jedem weiteren Iterationsschritt (v = 1, 2, …, x) entsteht ein
neues Objektmerkmal Ci,u+1, das in Li hinzugefügt wird. Bei v = x handelt es sich
um den letzten Iterationsschritt, bei dem beide Objektmerkmale wachsen. Li
enthält u = x + 2 Elemente und v = x. Vom Iterationsschritt v = x zum Schritt v =
x + 1 findet kein Wachstum statt, Li behält u = x + 2 Elemente und v erhält den
Wert v = x + 1. Ab dem Iterationsschritt v = x + 2 wächst das übrige wachstums-
fähige Objektmerkmal weiter an, bis nach v = y Iterationen kein Wachstum mehr
möglich ist. Li enthält nun u = y + 1 Elemente und v = y. Im Iterationsschritt v = y
+ 1 findet kein weiteres Wachstum mehr statt, Li erhält somit kein neues Ele-
ment und u behält seinen aktuellen Wert. Die Variable v wird jedoch bei jedem
Iterationsschritt um eins inkrementiert, so dass v letztlich den Wert von u an-
nimmt und die Abbruchbedingung des Algorithmus erfüllt ist.
Anwendung der beschriebenen Methode für die Wachstumssimulation von
MC3T3-Zellen:
Zellen der Linie MC3T3 bilden oft große kompakte Agglomerate, die an den
Zellkontaktstellen meist keine Verengungen aufweisen. Deshalb ist eine Zellse-
paration in Agglomeraten auf Basis geometrischer Überlegungen nicht zweck-

6. Separation agglomerierter Zellen 96
mäßig. Betrachtet man den Zellverbund in Abb. 59a, scheint die Fläche des
Kernbereiches mit der Fläche der Zelle in Zusammenhang zu stehen.
Abbildung 59: a: Zusammengewachsene MC3T3-Zellen auf dem Material Titan; b: Ergebnis des
HB-Algorithmus zur Kerndetektion; c: Der segmentierte Zellbereich ist grün umrandet, die de-
tektierten Kernbereiche schwarz umrandet. Teilweise wurden zwei Kerne als ein Bereich seg-
mentiert; d: Detektierte Kernbereiche mit starken Verengungen wurden aufgetrennt, da es sich
wahrscheinlich um zwei unterschiedliche Kernbereiche handelt.
Daraus leitet sich die Idee ab, die in den Agglomeraten enthaltenen individuel-
len Zellflächen auf Basis der detektierten Kernbereiche (Abb. 59d) durch eine
Wachstumssimulation zu approximieren (74). Betrachtet man die Gesamtfläche
einer Zelle als Merkmal U und die Fläche eines Kernbereiches als Merkmal V
mit jeweils q Merkmalswerten iu sowie iv mit i = 1, 2, …, q, gibt der Korrelati-
onskoeffizient (75)
1 2
2 21,
1 1
1
1 1; ; ( .16)
1 1
q
i i q qi
m m u i v i
i iu v
u u v vq
r s u u s v v Gls s q q
aufschluss über den linearen Zusammenhang zwischen der Zellfläche und der
Zellkernfläche.
2 Kernbereiche als eine große Region erfasst
Separierte Zellkernbereiche

6. Separation agglomerierter Zellen 97
Abbildung 60: Die Kernbereichflächen (V), aufgetragen über die Gesamtzellflächen (U)
Abb. 60 zeigt die Zellkernfläche, aufgetragen über die gesamte Zellfläche. Für
eine Stichprobe von 100 Zellen des Typs MC3T3 ergibt sich ein Korrelations-
wert , 0,86u vr . Für die Zelltypen L929 und L132 wurde ebenfalls der Korrelati-
onswert für eine Stichprobe von jeweils 100 Zellen berechnet. Es resultiert für
die Linie L929 ein Korrelationswert von 0,92 und für die Linie L132 ein Korrela-
tionswert von 0,95. Da die drei Zelltypen deutliche Unterschiede in ihrer Mor-
phologie aufweisen kann für weitere Zelltypen eine ähnliche Korrelation zwi-
schen Kernbereich und Zellfläche angenommen werden. Ein Test mit einem
weiteren Zelltyp bestätigt dies. Für Zellen des Typs BHK-21 resultiert ein Korre-
lationswert von 0,89. Auf Basis dieser Erkenntnisse erscheint eine Zellflächen-
approximation durch eine Wachstumssimulation, ausgehend von den Kernbe-
reichen, sinnvoll.
Das beschriebene Verfahren benötigt die Festlegung des Parameters p für die
maximale Wachstumsgeschwindigkeit der Objektmerkmale, in diesem Fall der
Zellkernbereiche. Tabelle 29 zeigt, dass für die Flächenapproximation von 100
in verschiedenen Clustern enthaltenen MC3T3 Zellen p = 5 eine gute Wahl ist.
Tabelle 29: Vergleich der Flächenapproximation von 100 MC3T3 Zellen innerhalb verschiedener Cluster
bei unterschiedlichen maximalen Wachstumsgeschwindigkeiten p
p=1 p=5 p=10
Durchschnittlicher Jaccard-Koeffizient
0,67 0,79 0,69
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
0 10000 20000 30000 40000
Zell
kern
flä
ch
e (
Pix
el)
Gesamtzellfläche (Pixel)
Kernbereichflächen aufgetragen über die Gesamtzellflächen

6. Separation agglomerierter Zellen 98
6.5.2 Ergebnisse
Das Ergebnis der Zellseparation mit Hilfe der Wachstumssimulation ist in Abb.
61 zu sehen.
Abbildung 61: Ergebnis der Wachstumssimulation. Die Zellgrenzen sind grün und die Kernbe-
reiche schwarz dargestellt.
Um die Genauigkeit der hier angewendeten Zellflächenrekonstruktion innerhalb
von Clustern zu beurteilen, wurden 200 von einem Experten manuell festgeleg-
te Zellbereiche (100 L929- und 100 MC3T3 Zellen) mit den automatisch seg-
mentierten Zellbereichen verglichen (Kap. 13.12, Tabelle 70 und Tabelle 71).
Mit Hilfe des Jaccard-Koeffizienten kann eine Aussage über die durchschnittli-
che Abweichung der automatisch generierten Zellkonturen zu den Referenzkon-
turen erfolgen (Tabelle 30).
Tabelle 30: Durchschnittlicher Jaccard-Koeffizient für die Separation von 100 L929- und 100
MC3T3 Zellen mit Hilfe des in Kap. 6.5 vorgestellten Separationsverfahrens
Separation von L929 Cluster
Separation von MC3T3 Cluster
Jaccard-Koeffizient
0,79 0,79

6. Separation agglomerierter Zellen 99
Im Durchschnitt besitzen die untersuchten Zellen vom Typ L929 sowie vom Typ
MC3T3 einen Jaccard-Koeffizienten von 0,79.
Die Zellzahlbestimmung basiert hier auf der Anzahl der detektierten Kerne nach
der Methode, beschrieben in Kap. 6.3.2. Dazu wurden aus jeweils 100 Proben
der Linien L929 und MC3T3 auf den Substraten Stahl* sowie Titan* die automa-
tisch bestimmte Anzahl der Zellen mit der von einem Experten ermittelten kor-
rekten Zellanzahl verglichen (Kap. 13.10).
Tabelle 31: Statistische Daten zur automatischen Zellzählung mit der Wachstumssimulation
L929-Zellzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
1,31 0,96 0,10
1,96 (95%) 0,19
2,576 (99%) 0,25
3,29 (99,9%) 0,31
3,87 (99,99%) 0,37
MC3T3-Zellzählung
Mittlere Abweichung zur Referenz [%]
Standard- Abweichung [%]
Standard- fehler [%]
Konfidenz- intervall
Toleranz um Abweichung
3,50 2,36 0,24
1,96 (95%) 0,46
2,576 (99%) 0,61
3,29 (99,9%) 0,78
3,87 (99,99%) 0,92
Tabelle 31 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten für die Linie L929 im Durchschnitt eine Abweichung von 1,31%
+/- 0,25% bei einem Konfidenzintervall von 99% zur Referenz einstellt. Für Zel-
len des Typs MC3T3 ergibt sich unter Verwendung einer Stichprobe von 100
Probenausschnitten hingegen eine Abweichung von 3,50% +/- 0,61% bei einem
Konfidenzintervall von 99%. Der Grund für die höhere Abweichung bei dem
Zelltyp MC3T3 liegt in der stärkeren Ausbildung von Agglomeraten im Vergleich
zum Typ L929 und einer stärkeren Variation der Zellmorphologie, die den Pro-
zess der Separation erschweren.
6.6 Zellseparation in Clustern mit Verengungen an Zellkontaktstellen
ohne Berücksichtigung der Zellkerne
Im Rahmen dieses Projektes wird nach der Zellkultivierung eine zytochemische
Färbung zur farblichen Unterscheidung zwischen Zellkernbereich und dem üb-
rigen Zellarealen durchgeführt. Bei diesen Bedingungen sind die in Kap. 6.4
und Kap. 6.5 vorgestellten Separationsmethoden für eine Trennung agglome-

6. Separation agglomerierter Zellen 100
rierter Zellen mit und ohne Verengungen an den Zellkontaktstellen geeignet. Es
finden sich auch andere Anwendungen, bei denen ungefärbte Zellen detektiert
werden sollen, z.B. bei der Verfolgung von sich bewegenden Zellen (76; 77).
Für die Separation ungefärbter Zellagglomerate sind die in Kap. 6.4 und Kap.
6.5 vorgestellten Verfahren nicht geeignet.
Das Ziel dieser Arbeit ist die Entwicklung eines computergestützten Auswer-
teprogrammes, das für unterschiedliche Aufgaben eingesetzt werden kann. Da-
her wird eine weitere Zell-Separationsmethode vorgestellt, mit der, im Unter-
schied zu den zuvor genannten, ohne Berücksichtigung der Kernbereiche eine
Zelltrennung in Clustern erfolgt. (38) stellt ein dafür geeignetes Separationsver-
fahren vor, das auf Basis einer iterativen Erosion in Kombination mit einer Ope-
ning-Methode (78) zunächst Marker für die anschließende Rekonstruktion der
Zellbereiche generiert. Diese Marker werden anschließend mit Hilfe einer erwei-
terten Dilatationsmethode zu Zellbereichen rekonstruiert. Dieses Verfahren ist
für die Trennung zusammengewachsener Zellen nicht auf die Detektion der in
den Clustern enthaltenen Kernbereiche angewiesen. Das in (38) beschriebene
Verfahren wird als alternative Separationsmethode in dieser Arbeit übernom-
men. Da aus (38) keine Informationen zur Genauigkeit der Zellflächenrekon-
struktion sowie zur Genauigkeit bei der automatischen Zellzahlbestimmung her-
vorgehen, liefert Kap. 6.6.1 die fehlenden Untersuchungsergebnisse. Mit Hilfe
der Daten ist es anschließend möglich die in (38) beschriebenen Methode mit
den in Kap. 6.4 und 6.5 beschriebenen Separationsverfahren objektiv zu ver-
gleichen.
6.6.1 Ergebnisse
Zur Beurteilung der Genauigkeit bezüglich der generierten Einzelzellflächen
innerhalb von Agglomeraten des in Kap. 6.6 vorgestellten Separationsverfah-
rens ist eine Referenzsegmentierung notwendig. Dazu sind durch einen Exper-
ten aus 5 verschiedenen Proben 100 in verschiedene Cluster verbundene L929
Zellen manuell segmentiert worden. Mit Hilfe des Jaccard-Koeffizienten kann
nun eine Aussage über die durchschnittliche Abweichung der automatisch ge-
nerierten Zellflächen zu den Referenzzellflächen erfolgen. Der Durchschnitts-
wert des Jaccard-Koeffizienten auf Basis von 100 aus Clustern separierten
L929 Zellen für den Auflicht-Hellfeld Mikroskopkontrast beträgt 0,77. Vor der
Zellseparation in den Clustern müssen alle Zellbereiche zuvor mit einem der in
Kap. 5 vorgestellten Methoden segmentiert werden. Das geschieht mit Hilfe des
schwellwertbasierten Verfahrens, beschrieben in Kap. 5.2.1.

6. Separation agglomerierter Zellen 101
Die Bewertung der Leistungsfähigkeit bezüglich der Zellzählung des vorgestell-
ten Separationsverfahrens erfolgt durch Gegenüberstellung der automatischen
Zellzählung von 100 Probenausschnitten der Linie L929 mit einer Referenzzäh-
lung durch eine Laborfachkraft (Kap. 13.11). Die statistische Auswertung der
Daten ist in Tabelle 32 aufgeführt.
Tabelle 32: Statistische Daten zur automatischen Zellzählung unter Verwendung des in Kap.
6.6 beschriebenen Verfahrens
Zellzählung Mittlere Abweichung
zur Referenz [%] Standard-
Abweichung [%] Standard- fehler [%]
Konfidenz- Intervall
Toleranz um Abweichung
3,13 1,37 0,14
1,96 (95%) 0,27
2,576 (99%) 0,35
3,29 (99,9%) 0,45
3,87 (99,99%) 0,53
Tabelle 32 zeigt, dass sich unter Verwendung einer Stichprobe von 100 Pro-
benausschnitten im Durchschnitt eine Abweichung von 3,13% +/- 0,35% bei
einem Konfidenzintervall von 99% zur Referenz einstellt. Da Zellen vom Typ
MC3T3 oft kompakte flächige Agglomerate ausbilden und demnach nicht durch
Verengungen an den Zellkontaktstellen gekennzeichnet sind, liegen hier keine
Ergebnisse vor.
6.7 Randzellenbehandlung
Bei der Aufnahme eines Probenausschnittes befinden sich oft nur Teilbereiche
von Zellen am Bildrand (Abb. 62 rot markiert), die übrigen zu den Zellen gehö-
renden Areale verlaufen über den Bildrand hinaus. Diese Zellbestandteile in-
nerhalb des Probenausschnittes werden im Folgenden Randzellen genannt.
Abbildung 62: MC3T3 Zellen auf dem Material Stahl*; Zellen, die sich nur teilweise im Bild be-
finden (rot umrandet), müssen eliminiert werden

6. Separation agglomerierter Zellen 102
Die geometrischen Merkmale der Randzellen, z.B. die Fläche oder Kompakt-
heit, können aufgrund des Umstandes, dass sich nur ein Teil der Zelle im Bild
befindet, die morphologische Analyse der Zellen innerhalb des Probenaus-
schnittes verfälschen. Des Weiteren ist eine standardisierte Zählweise notwen-
dig, damit die Ergebnisse der automatischen und manuellen Zellzählung ver-
gleichbar sind, z.B. zur Bewertung der Genauigkeit bei der automatischen Zell-
zählung. Aus diesen Gründen ist eine Elimination der Randzellen zweckmäßig.
Zunächst müssen von den n vorhandenen Zellbereichen Z = {Z1, Z2, …, Zn} im
Probenausschnitt die Randzellen klassifiziert werden. Dazu wird eine Recht-
eckkontur R mit der Größe des zugrundeliegenden Bildes und einer Dicke von
einem Pixel generiert, Abb. 63.
Abbildung 63: Schematischer Probenausschnitt; Generierung einer Rechteckkontur (schwarze
Kontur) als Hilfsmittel zur Bestimmung der Randzellen
Von R und jeder im Bild segmentierten Zelle Zi wird jeweils die gemeinsame
Schnittmenge i iS Z R ermittelt. Ist |Si| > 0, wird das Verhältnis
ii
i
ZLV
S (Gl. 22)
bestimmt, wobei ZLi hier als die Zellkonturlänge der Randzelle Zi aufzufassen
ist. Überschreitet Vi den Wert eines einstellbaren Parameters F, so wird die
Randzelle nicht direkt verworfen, da nur ein verhältnismäßig geringer Teil der
Zellkontur den Rand schneidet und somit von einem Experten bei einer Zellzäh-
lung wahrscheinlich gezählt wird. Im Rahmen dieser Arbeit wird F = 30 gesetzt.
Nun erfolgt die Bestimmung der Kompaktheit Ki, Kreisförmigkeit KFi und die
Fläche Ai der Randzelle Zi. Untersuchungen haben gezeigt, wenn Ki > KMax ˄
KFi < KFMax ˄ Ai < AMax, dann handelt es sich bei Zi wahrscheinlich um einen
Zellausläufer. Sind die genannten Bedingungen erfüllt, wird Zi demnach ge-
löscht. Andernfalls ist Zi eine gültige Zelle für die Analyse. Für die Zelllinien
L929 und MC3T3 haben sich die Parameter KMax = 10, KFMax = 0.1 und AMax =
1500 Pixel bewährt, für andere Zelllinien können die Parameter entsprechend

6. Separation agglomerierter Zellen 103
angepasst werden. Die Randzellenverarbeitung ist zusammenfassend in Form
eines Ablaufdiagrammes erläutert (Abb. 64).
Abbildung 64: Ablaufdiagramm zur Randzellenbehandlung nach der Zellsegmentierung
Abb. 64 zeigt das Ablaufdiagramm der bereits beschriebenen Randzellenverar-
beitung. Die Dekrementierung der Laufvariablen i, für den Fall der Löschung
von Zi aus Z ist notwendig, da nach dem Löschen des Elementes an der Positi-

6. Separation agglomerierter Zellen 104
on i der Index aller nachfolgenden Elemente automatisch um eins dekrementiert
wird.
6.7.1 Ergebnisse
Zur Bewertung der Qualität der Randzellenbehandlung wurde die automatische
Zellzahlerkennung mit und ohne Randzellenbehandlung einer Referenzzählung
durch einen Experten gegenübergestellt (Tabelle 33).
Tabelle 33: Vergleich der automatischen Zellzählung ohne Randzellenbehandlung mit der Refe-
renzzählung; AZ: Computergestützte Zellzählung; REF: Referenzzellzählung durch einen Ex-
perten; ERR: Relative Abweichung der automatischen Zellzählung von der Referenzzählung
Probe AZ REF ERR (%)
L929 1 266 232 14,7
L929 2 418 369 13,3
L929 3 280 250 12,0
L929 4 235 199 18,1
L929 5 279 240 16,3
L929 6 225 199 13,1
L929 7 244 215 13,5
L929 8 240 210 14,3
L929 9 250 215 16,3
L929 10 316 280 12,9
Ø Fehler 14,9
MC3T3 1 108 85 27,1
MC3T3 2 122 107 14,0
MC3T3 3 98 82 19,5
MC3T3 4 117 90 30,0
MC3T3 5 87 65 33,8
MC3T3 6 110 90 22,2
MC3T3 7 112 95 17,9
MC3T3 8 123 104 18,3
MC3T3 9 110 99 11,1
MC3T3 10 88 67 31,3
Ø Fehler 22,5
Für den Typ L929 kommt das Verfahren, beschrieben in Kap. 6.4, für die Linie
MC3T3 hingegen die Methode, beschrieben in Kap. 6.5 zum Einsatz. Tabelle
32 belegt, dass die automatische Zellzahlbestimmung für den Typ L929 ohne
Randzellenbehandlung durchschnittlich 14,9% Abweichung zur Referenzzäh-
lung beträgt. Bei aktivierter Randzellenbehandlung sinkt die Abweichung von
14,9% auf 1,27% (Tabelle 28). Für Zellen der Linie MC3T3 resultiert im Durch-
schnitt eine Abweichung von 22,5% ohne Randzellenbehandlung und 3,50%
Abweichung mit der Randzellenbehandlung (Tabelle 31). Aus Tabelle 33 geht

6. Separation agglomerierter Zellen 105
weiterhin hervor, dass die Abweichung ohne Randzellenbehandlung beim Typ
MC3T3 im Vergleich zum Typ L929 um 7,6% höher ausfällt. Der Grund liegt
darin, dass die MC3T3 Cluster im Vergleich zu L929-Agglomeraten mehr Zel-
len beinhalten und die am Bildrand gelegenen Cluster teilweise vom Bildrand
durchschnitten werden. In Relation zur Linie L929 werden demnach häufiger
MC3T3 Randzellen verworfen, da sie sonst die Analyse verfälschen würden.
Wenn die computergestützte Randzellenbehandlung nicht ausgeführt wird, blei-
ben jedoch alle Randzellen erhalten. Da der Anteil an MC3T3-Randzellen im
Vergleich zum Anteil an L929-Randzellen größer ausfällt, führt dies beim Typ
MC3T3 im Durchschnitt zu einer größeren Abweichung zur Referenzzählung.
6.8 Diskussion der Ergebnisse zu den Separationsverfahren
Die in Kap. 6.1 vorgestellte Methode CBS findet Anwendung für die Separation
agglomerierter Zellen mit Verengungen an deren Kontaktstellen. Der Vorteil
dieser Methode wird deutlich, wenn sie dem Separationsverfahren, beschrieben
in Kap. 6.4, vorangestellt wird. Hierdurch kann die Abweichung der automati-
schen Zellzählung zur Referenzzählung von 2,3% auf 1,27% reduziert werden.
Außerdem wird eine Reduzierung der Rechenzeit um ca. Faktor 8 erzielt. Ein
weiterer Vorteil liegt in der Anwendbarkeit auf allgemeine Objekte, die an Ver-
engungen aufgetrennt werden sollen. Für Zelltypen deren Agglomerate keine
Verengungen aufweisen, z.B. Zellen der Linie MC3T3, ist dieses Verfahren un-
geeignet.
Die in Kap. 6.2 beschriebene Methode zur Klassifizierung der segmentierten
Zellbereiche auf Basis einer iterativen Erosion führt zu einer mittleren Fehlklas-
sifizierung von 1,3% für den Zelltyp L929 und von 0,2% für den Zelltyp MC3T3.
Eine Klassifizierung der L929-Zellbereiche in Einzelzellen oder Agglomerate
ausschließlich mit der Auswertung der Anzahl der Kerne, würde zu einer 0,9%
schlechteren Klassifikation im Vergleich zur eingesetzten Methode führen. Dies
ist darauf zurückzuführen, dass kleine Zellen, die gerade einen Teilungsprozess
abschließen, oft über keine optisch erkennbaren Zellkernbereiche verfügen und
so fälschlicherweise als Einzelzelle klassifiziert werden.
Kap. 6.4 stellt ein Separationsverfahren vor, das Zellen an Engstellen teilt. Es
handelt sich um eine Kombination aus verschiedenen in der Literatur bekannten
Algorithmen, wobei teilweise Anpassungen der Methoden an diese Aufgaben-
stellung notwendig waren, beschrieben in Kap. 6.4.1. Der Vorteil dieses Verfah-
rens liegt darin, dass es auf alle Zelltypen anwendbar ist, die Verengungen an
den Zellkontaktstellen aufweisen. Die Methode ist in der Lage, trotz teilweise

6. Separation agglomerierter Zellen 106
nicht vorhandener Verengungen an den Zellkontaktstellen eine Separation mit
Hilfe bestimmter Konturpunkte durchzuführen. Jedoch sollte der Anteil an Eng-
stellen an den Zellkontaktstellen für ein genaues Separationsergebnis überwie-
gen.
Der Algorithmus benötigt trotz vorgeschalteter CBS (Kap. 6.1) im Vergleich zu
den Separationsverfahren, beschrieben in den Kapiteln 6.5 und 6.6, ca. die
dreifache Rechenzeit (Tabelle 34). Für ein Bild werden, abhängig von der An-
zahl und Größe vorhandener Agglomerate, ca. 1s Rechenzeit in Anspruch ge-
nommen. Das Verfahren setzt die Detektion der Kernbereiche innerhalb der
Agglomerate voraus. Im Rahmen des Projektes geschieht dies durch die in
Kap. 6.3.2 beschriebene Methode auf Basis des Histogram-Backprojection-
Algorithmus.
Tabelle 34 bietet einen Überblick der Rechenzeiten der drei vorgestellten Sepa-
rationsverfahren.
Tabelle 34: Vergleich der Rechenzeiten zwischen den drei Separationsverfahren für einen Pro-
benausschnitt der Linie L929 auf einer Intel Core i7 3,2GHz CPU
Separationsverfahren Durchschnittliche Rechenzeit des
Separationsvorgangs für einen Probenausschnitt der Linie L929
Separationsverfahren (Kap. 6.4) 1,03s
Separationsverfahren (Kap. 6.5) 0,27s
Separationsverfahren auf Basis von (38) (Kap. 6.6)
0,31s
Das Trennverfahren (Kap. 6.4) ist mit einer Rechenzeit von ca. 1s im Vergleich
zu den anderen beiden Verfahren am langsamsten. Die Methoden (Kap. 6.5
und Kap. 6.6) benötigen jeweils Rechenzeiten von ca. 300ms.
Das Separationsverfahren (Kap. 6.5) führt eine Zellflächenrekonstruktion mit
Hilfe einer Wachstumssimulation auf Basis der im Vorfeld detektierten Kernbe-
reiche (Kap. 6.3.2) durch. Der Vorteil dieser Methode ist die Anwendbarkeit auf
alle Zelllinien, bei denen eine Kerndetektion möglich ist. Die Segmentierung der
Kerne ist z.B. mit Hilfe einer zytochemischen Färbung der Zellen durchführbar.
Somit kann der Algorithmus, im Gegensatz zu den Verfahren, beschrieben in
Kap. 6.1 und 6.4, völlig unabhängig von der Zellgeometrie die Separation von
Agglomeraten durchführen. Ein weiterer Vorteil dieser Methode ist die ver-
gleichsweise schnelle Verarbeitungszeit. Für ein Bild werden im Durchschnitt
ca. 0,3s benötigt. Die Qualität dieses Verfahrens hängt stark von der Detektier-
barkeit der Kernbereiche ab. So führt eine ungenaue Segmentierung der Kerne
mit hoher Wahrscheinlichkeit auch zu einer nicht zufriedenstellenden Separati-

6. Separation agglomerierter Zellen 107
on in den Zellclustern. Mit dem Verfahren zur Detektion der Kernbereiche, be-
schrieben in Kap. 6.3.2, ist jedoch eine hinreichende Segmentierungsgenauig-
keit für Zellen vom Typ MC3T3 und L929 gegeben. Da auf einfache Weise mit
Hilfe eines im Rahmen dieses Projektes entwickelten Softwaretools neue Farb-
templates erstellt werden können, stellt eine Anpassung der Kerndetektion an
andere Zelllinien kein Problem dar.
Kap. 6.6. beschreibt ein weiteres Separationsverfahren, dass im Vergleich zu
den zwei zuvor genannten Trennmethoden (Kap. 6.4 und Kap. 6.5) ohne vorhe-
rige Detektion der Kernbereiche arbeitet. Der Separationsvorgang dieser Me-
thode ist in (38) beschrieben. Der Vorteil liegt darin, dass in diesem Fall keine
Anfärbung und Segmentierung der Zellkerne vorgenommen werden müssen.
Zudem benötigt diese Methode mit ca. 0,3s pro Bild eine geringe Rechenzeit.
Ein Nachteil dieser Methode ist, dass aus biologischer Sicht teilweise unsinnige
Trennvorgänge durchgeführt werden, z.B. durch einen Zellkernbereich. Dies
liegt an der variablen Morphologie der verwendeten Zelltypen. So sind auch an
engen Zellbereichen teilweise Kernflächen vorzufinden, die bei einer iterativen
Erosion fälschlicherweise aufgetrennt werden. Darüber hinaus ist diese Metho-
de nicht für die Separation kompakt gewachsener Cluster, z.B. vom Typ
MC3T3, geeignet, da diese Zelllinie kaum Verengungen an den Zellkontaktstel-
len ausbildet. Tabelle 35 zeigt die durchschnittlichen Jaccard-Koeffizienten für
die in Kap. 6.4 bis 6.6 beschriebenen Separationsverfahren, um die Qualität der
Zellflächenrekonstruktion vergleichen zu können. Die Jaccard-Koeffizienten al-
ler untersuchten Zellen sind in Kap. 13.12 hinterlegt.
Tabelle 35: Vergleich des Jaccard-Koeffizienten für 3 verschiedene Separationsverfahren auf
Basis von 200 Zellen (100 L929- und 100 MC3T3 Zellen)
Separation von L929 Cluster
Wachstumssimulation (Kap. 6.5)
Separationsverfahren auf Basis von (38) (Kap. 6.6)
Separationsverfahren (Kap. 6.4)
Jaccard-Koeffizienten Jaccard-Koeffizienten Jaccard-Koeffizienten
0,79 0,77 0,80
Separation von MC3T3 Cluster
Wachstumssimulation (Kap. 6.5)
Separationsverfahren (Kap. 6.6)
Separationsverfahren (Kap. 6.4)
Jaccard-Koeffizienten Jaccard-Koeffizienten Jaccard-Koeffizienten
0,79 0,38 0,42
Das Separationsverfahren auf Basis von (38) liefert für 100 segmentierte L929
Referenzzellen einen durchschnittlichen Jaccard-Koeffizient von 0,77. Mit die-
sem Ergebnis schneidet das Verfahren auf Basis von (38) etwas schlechter ab
als der Algorithmus aus Kap. 6.4. Der Grund liegt darin, dass die Methode im

6. Separation agglomerierter Zellen 108
Gegensatz zu dem Verfahren aus Kap. 6.4 nicht in der Lage ist zu erkennen, ob
Zellkernbereiche durch den Separationsprozess aufgetrennt werden. Der Algo-
rithmus, beschrieben in Kap. 6.5, liefert ebenfalls etwas schlechtere Ergebnisse
als das Verfahren aus Kap. 6.4, da es die Separation nicht anhand der für die
Linie L929 charakteristischen Verengungen vornimmt, sondern die Zellflächen-
approximation, ausgehend von den detektierten Zellkernbereichen, durchführt.
Mit der Trennmethode aus Kap. 6.4 ist die, im Vergleich zu den Verfahren aus
Kap. 6.5 und Kap. 6.6, präziseste Zellflächenrekonstruktion von L929 Zellen
möglich (Jaccard-Koeffizient: 0,80). Allerdings ist diese Methode sehr rechenin-
tensiv (Tabelle 34) und analog zum Trennverfahren auf Basis von (38) (Kap.
6.6) nur für Zelltypen geeignet, die Verengungen an den Kontaktstellen der Zel-
len aufweisen. Zur Trennung von kompakten Zell-Agglomeraten, z.B. der Linie
MC3T3, ist von den vorgestellten Verfahren ausschließlich die Wachstumssimu-
lation, beschrieben in Kap. 6.5, geeignet. Für eine Separation werden keine
Engstellen der Zellkontur benötigt.
Andere Separationsverfahren basieren oft auf der Auswertung geometrischer
Merkmale (46; 47).
In (46) wird von einem zuvor segmentierten Zellcluster dessen Skelett berech-
net. Die Positionen der Konturverengungen des Clusters werden mit Hilfe eines
Abstandshistogramms ermittelt, in dem für jeden Konturpunkt der kürzeste Ab-
stand zum Skelett enthalten ist. Ein definiertes Regelwerk steuert die Auswahl
der Konturpunkte, die zur Trennung des Clusters verbunden werden.
Vorteil dieses Verfahrens ist die gute Anwendbarkeit auf elliptisch geformte Zel-
len, die bei Agglomeratbildung Verengungen an den Zellkontaktstellen aufwei-
sen. Ein Nachteil liegt darin, dass die Methode nicht auf Zellen mit variabler
Morphologie anwendbar ist. Es wird keine Methode vorgestellt, mit der die Se-
paration von Clustern mit teilweise fehlenden Verengungen vorgenommen wird.
In (46) wird darüber hinaus keine Beurteilung der Separationsleistung geliefert,
z.B. durch Vergleich der automatischen Zellzählung nach der Zellseparation mit
einer Referenzzählung. Ein Bild aus (46) zeigt Zellen, die trotz vorhandener
Verengungen der Zellclusterkonturen nicht getrennt wurden, so dass eine gute
Separationsleistung angezweifelt werden muss. Aufgrund der genannten Nach-
teile wird dieses Verfahren nicht für die Auftrennung von L929 und MC3T3 Ag-
glomeraten eingesetzt.
Eine weitere aus der Literatur bekannte Separationsmethode wird in (47) vor-
gestellt. Die mit Hilfe einer Kantenerkennung segmentierten Zellkonturen wer-
den jeweils in Abschnitte mit einer spezifizierten Länge unterteilt und für jeden
Abschnitt die Normale berechnet. Durch bestimmte Winkelbeziehungen der

6. Separation agglomerierter Zellen 109
Normalen zu der x-Achse wird auf eine Verengung der Kontur geschlossen.
Punkte, die jeweils eine Engstelle in der Kontur repräsentieren, werden unter
Verwendung spezieller Regeln verbunden und so die Cluster aufgetrennt. Es
folgt eine Ellipsenapproximation der separierten Bereiche zur Rekonstruktion
der individuellen Zellflächen.
Diese Methode ist durch die elliptische Approximation der separierten Zellberei-
che ausschließlich auf ellipsenförmige Zellen begrenzt. Für Zellen mit variieren-
der Morphologie, z.B. vom Typ L929, ist trotz häufiger Verengungen an den
Zellkontaktstellen keine ausreichende Zellflächenrekonstruktion dieser Art nach
der Separation gewährleistet, da die Zellgeometrie stark von der Form einer
Ellipse abweichen kann. Darüber hinaus zeigen die Ergebnisse in (47), dass für
lediglich vier Bilder mit jeweils ca. 300 – 400 Zellen ein Vergleich der automati-
schen Zellzählung mit einer Referenzzählung vorgenommen wurde. Die Abwei-
chung der automatischen Zellzählung zur Referenzzählung variiert von Bild zu
Bild zwischen -7,09% und 8,59%. Daraus lässt sich schlussfolgern, dass die
vorgestellte Methode keine reproduzierbaren Ergebnisse, selbst bei Verwen-
dung derselben Zelllinie, liefert. Aus diesen Gründen wird das Verfahren aus
(47) für die Separation der in diesem Projekt vorliegenden Zelltypen nicht ein-
gesetzt.
Ein weiteres Verfahren stützt sich auf die farbliche Unterscheidung von zusam-
mengewachsenen Zellen (45). Hier wird eine zweifache Fluoreszenzfärbung
einer Probe zur farblichen Darstellung des Zytoplasmas und der Zellkerne vor-
genommen. Die Segmentierung der Zellplasmen wird mit einem histogrammba-
sierten Schwellwertverfahren im PAP-gefärbten Durchlichtbild und die Detektion
der Kerne ebenfalls mit einem histogrammbasierten Schwellwertverfahren im
DAPI-gefärbten Fluoreszenzbild ausgeführt. Die Trennung von Zellclustern er-
folgt mit Hilfe der Segmentierung von dunkler erscheinenden Überlappungsbe-
reichen zusammenhängender Zellen durch das Otsu Schwellwertverfahren. Zur
Bewertung der Separationsleistung wurden als Referenzwert 1665 Zellen ma-
nuell gezählt und der automatischen Zellzählung gegenübergestellt. Daraus
resultiert für die automatische Zellzählung eine Abweichung von 24% zum Re-
ferenzwert. Diese Abweichung ist im Vergleich zu den anderen Separationsver-
fahren sehr hoch (Tabelle 35).
Vorteil dieses Verfahrens ist eine geringe Rechenzeit. Es liegen zwar keine An-
gaben über die Ausführungsgeschwindigkeit vor, aber die vorgestellten Bildver-
arbeitungsfunktionen besitzen erfahrungsgemäß Rechenzeiten jeweils im hun-
dertstel bis zehntel Sekundenbereich. Nachteil dieser Methode ist, dass die Se-
paration eine farbliche Unterscheidung des Überlappungsbereiches zusam-

6. Separation agglomerierter Zellen 110
mengewachsener Zellen voraussetzt. Diese Eigenschaft ist im Rahmen dieses
Projektes nicht gegeben, so dass auch dieses Verfahren hier keine Verwen-
dung findet.
Das in (43) beschriebene Verfahren segmentiert fluoreszenzgefärbte Zellen mit
Hilfe des Gradientenverfahrens und trennt zusammenhängende Zellbereiche
auf Basis einer Wasserscheidentransformation mit anschließender Distanz-
transformation. Dieser Algorithmus beschränkt sich jedoch ausschließlich auf
die Segmentierung und Separation von fluoreszenzgefärbten HEp-2 Zellen,
wobei die Zellmorphologie dieses Typs aufgrund fehlender Zellausläufer und
der relativ runden Geometrie einfacher automatisch zu analysieren ist im Ver-
gleich zum Typ L929 und MC3T3. Im Ergebnisteil des papers ist für 20 Testbil-
der mit insgesamt 1242 Zellen eine Abweichung von 5% zur Referenzzählung
angegeben. Es ist davon auszugehen dass sich bei der hier vorliegenden kom-
plexeren Zellmorphologie sowie der kontrastärmeren May-Grünwaldfärbung
schlechtere Ergebnisse einstellen die zu einer ungenauen automatische Be-
stimmung des Zytotoxizitätsgrades eines vorliegenden Materials führen.
In (38) werden Zellagglomerate vom Typ L929 einer Separation, basierend auf
morphologischen Operationen, unterzogen. Die automatische Zellzählung mit
Hilfe dieses Verfahrens liefert im Vergleich zu den Ergebnissen der in Kap. 6.4
und 6.5 vorgestellten Trennverfahren größere Abweichungen zum Referenzwert
(Tabelle 36).
Tabelle 36: Vergleich der durchschnittlichen Abweichungen der automatischen Zellzählung zur
Referenzzählung für die Separationsverfahren aus Kap. 6.4 – 6.6 für die Linien L929 und
MC3T3 unter Berücksichtigung eines Konfidenzintervalls von 99%. Untersucht wurden pro Zell-
linie 100 Probenausschnitte mit insgesamt ca. 20.000 L929 Zellen und ca. 10.000 MC3T3 Zel-
len
Separationsverfahren MC3T3 L929
Verfahren A (Kap. 6.4) --- 1,27% +/- 0,22%
Verfahren B (Kap. 6.5) 3,5% +/- 0,61% 1,31% +/- 0,25%
Verfahren aus (38) (Kap. 6.6) --- 3,13% +/- 0,35%
Tabelle 36 zeigt, dass das Separationsverfahren aus (38) im Durchschnitt eine
1,86% höhere Abweichung der automatischen L929-Zellzählung zur Referenz-
zählung aufweist als das in Kap. 6.4 beschriebene Trennverfahren. Im Ver-
gleich zum Separationsverfahren (Kap. 6.5) liegt ebenfalls eine höhere Abwei-
chung (1,82%) vor. Für die Vereinzelung von Zellclustern der Linie MC3T3 sind
die Verfahren aus (38) und Kap. 6.4 im Gegensatz zu dem Trennverfahren aus

6. Separation agglomerierter Zellen 111
Kap. 6.5, das hier eine Abweichung von 3.5% zum Referenzwert aufweist, nicht
geeignet.
Auf Basis der diskutierten Methoden lässt sich schlussfolgern, dass für die
Trennung von Zellagglomeraten des Typs L929 sowie ähnlich gearteter Zellen
das Separationsverfahren, beschrieben in Kap. 6.4, mit vorgeschalteter CBS
(Kap. 6.1) die zu bevorzugende Methode ist. Alternativ kann mit ähnlichen Er-
gebnissen der Wachstumsalgorithmus aus Kap. 6.5 zur Separation und Rekon-
struktion der Zellflächen herangezogen werden, insbesondere wenn eine ver-
gleichsweise kurze Prüfzeit erforderlich ist. Bei der Analyse von MC3T3 Zellen
oder morphologisch ähnlichen Typen eignet sich das Wachstumsverfahren
(Kap. 6.5). Für den Fall, dass Zellen ohne Färbung der Kernbereiche analysiert
werden sollen, lassen sich Agglomerate mit dem Verfahren auf Basis von (38)
auftrennen.

7. Automatisierung der Algorithmenauswahl 112
7. Automatisierung der Algorithmenauswahl
In den Kapiteln 5 und 6 wurden die Zellsegmentierungs- und separationsverfah-
ren vorgestellt. Das vorliegende Kapitel geht nun auf das Konzept der automati-
schen Zellanalyse ein. Es wird ein Verfahren vorgestellt, dass das geladene
Bildmaterial hinsichtlich der Zellmorphologie und des Mikroskopkontrastes ana-
lysiert und die für die Analyse am besten geeignete Kombination an Algorith-
men ausgewählt. Abbildung 65 gibt einen Überblick des allgemeinen Analyse-
konzeptes.
Abbildung 65: Allgemeine Vorgehensweise bei der Bildanalyse
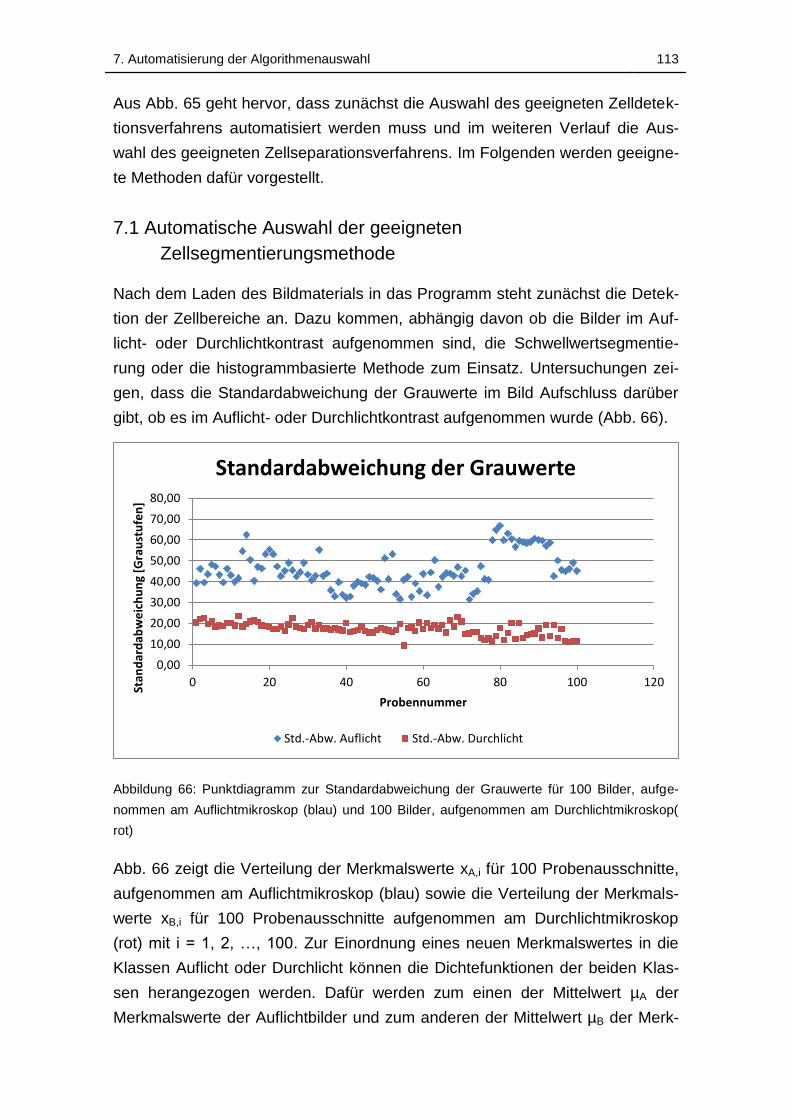
7. Automatisierung der Algorithmenauswahl 113
Aus Abb. 65 geht hervor, dass zunächst die Auswahl des geeigneten Zelldetek-
tionsverfahrens automatisiert werden muss und im weiteren Verlauf die Aus-
wahl des geeigneten Zellseparationsverfahrens. Im Folgenden werden geeigne-
te Methoden dafür vorgestellt.
7.1 Automatische Auswahl der geeigneten
Zellsegmentierungsmethode
Nach dem Laden des Bildmaterials in das Programm steht zunächst die Detek-
tion der Zellbereiche an. Dazu kommen, abhängig davon ob die Bilder im Auf-
licht- oder Durchlichtkontrast aufgenommen sind, die Schwellwertsegmentie-
rung oder die histogrammbasierte Methode zum Einsatz. Untersuchungen zei-
gen, dass die Standardabweichung der Grauwerte im Bild Aufschluss darüber
gibt, ob es im Auflicht- oder Durchlichtkontrast aufgenommen wurde (Abb. 66).
Abbildung 66: Punktdiagramm zur Standardabweichung der Grauwerte für 100 Bilder, aufge-
nommen am Auflichtmikroskop (blau) und 100 Bilder, aufgenommen am Durchlichtmikroskop(
rot)
Abb. 66 zeigt die Verteilung der Merkmalswerte xA,i für 100 Probenausschnitte,
aufgenommen am Auflichtmikroskop (blau) sowie die Verteilung der Merkmals-
werte xB,i für 100 Probenausschnitte aufgenommen am Durchlichtmikroskop
(rot) mit i = 1, 2, …, 100. Zur Einordnung eines neuen Merkmalswertes in die
Klassen Auflicht oder Durchlicht können die Dichtefunktionen der beiden Klas-
sen herangezogen werden. Dafür werden zum einen der Mittelwert µA der
Merkmalswerte der Auflichtbilder und zum anderen der Mittelwert µB der Merk-
0,00
10,00
20,00
30,00
40,00
50,00
60,00
70,00
80,00
0 20 40 60 80 100 120
Stan
dar
dab
we
ich
un
g [G
rau
stu
fen
]
Probennummer
Standardabweichung der Grauwerte
Std.-Abw. Auflicht Std.-Abw. Durchlicht

7. Automatisierung der Algorithmenauswahl 114
malswerte für die Durchlichtbilder benötigt. Des Weiteren sind die Standardab-
weichungen σA ebenfalls für die Merkmalswerte der Auflichtbilder und σB für die
Merkmalswerte der Durchlichtbilder zu ermitteln. Mit dem Mittelwert und der
Standardabweichung kann die Dichtefunktion der Normalverteilung (79) be-
stimmt werden (Gl. 23).
21
22 1, ,
2
x
x e
(Gl. 23)
Die Dichtefunktionen φA der Auflichtverteilung und φB der Durchlichtverteilung
sind in Abb. 67 dargestellt.
Abbildung 67: Dichtefunktionen der Standardabweichungen von Graustufen in 100 Auflichtbil-
dern (blau) und 100 Durchlichtbildern (rot)
Für die Klassifikation eines Bildes in die Klasse Auflicht oder Durchlicht wird
nun der Merkmalswert x für das Bild bestimmt und in die beiden Dichtefunktio-
nen φA und φB eingesetzt und der jeweilige Funktionswert bestimmt. Wenn der
Funktionswert φA(x, µA, σA) ≥ φB(x, µB, σB) resultiert, erfolgt die Einordnung des
Eingabebildes in die Klasse Auflicht, andernfalls in die Klasse Durchlicht. Liegt
eine Klassifizierung als Auflichtbild vor, kommt das Segmentierungsverfahren,
beschrieben in Kap. 5.2.1, zum Einsatz, ansonsten die Methode, beschrieben in
Kap. 5.2.2. Aus Abb. 67 geht hervor, dass sich die beiden Verteilungen nur ge-
0,00000
0,02000
0,04000
0,06000
0,08000
0,10000
0,12000
0,14000
0,16000
1 5 9
13
17
21
25
29
33
37
41
45
49
53
57
61
65
69
73
77
81
85
89
93
97
φ(x
,µ,σ
)
Standardabweichung x
Dichtefunktionen der Standardabweichungen von Graustufen in Auflicht- und Durchlichtbildern
Dichtefkt. Auflicht Dichtefkt. Durchlicht
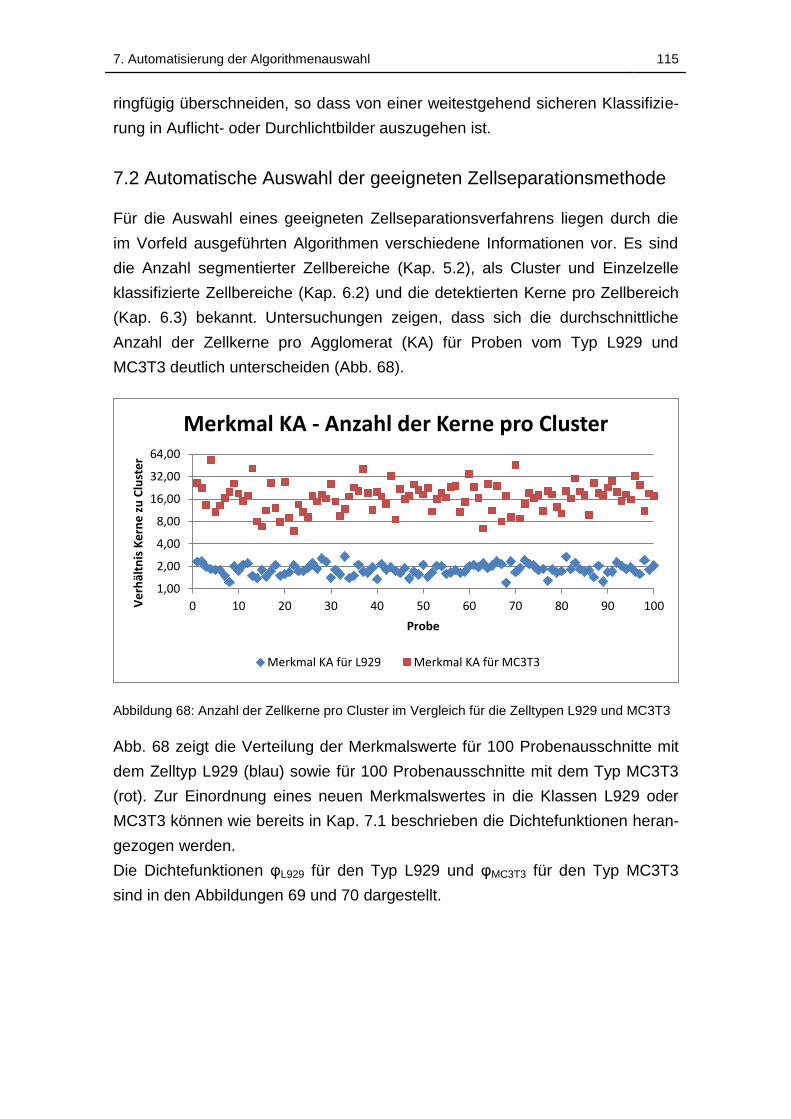
7. Automatisierung der Algorithmenauswahl 115
ringfügig überschneiden, so dass von einer weitestgehend sicheren Klassifizie-
rung in Auflicht- oder Durchlichtbilder auszugehen ist.
7.2 Automatische Auswahl der geeigneten Zellseparationsmethode
Für die Auswahl eines geeigneten Zellseparationsverfahrens liegen durch die
im Vorfeld ausgeführten Algorithmen verschiedene Informationen vor. Es sind
die Anzahl segmentierter Zellbereiche (Kap. 5.2), als Cluster und Einzelzelle
klassifizierte Zellbereiche (Kap. 6.2) und die detektierten Kerne pro Zellbereich
(Kap. 6.3) bekannt. Untersuchungen zeigen, dass sich die durchschnittliche
Anzahl der Zellkerne pro Agglomerat (KA) für Proben vom Typ L929 und
MC3T3 deutlich unterscheiden (Abb. 68).
Abbildung 68: Anzahl der Zellkerne pro Cluster im Vergleich für die Zelltypen L929 und MC3T3
Abb. 68 zeigt die Verteilung der Merkmalswerte für 100 Probenausschnitte mit
dem Zelltyp L929 (blau) sowie für 100 Probenausschnitte mit dem Typ MC3T3
(rot). Zur Einordnung eines neuen Merkmalswertes in die Klassen L929 oder
MC3T3 können wie bereits in Kap. 7.1 beschrieben die Dichtefunktionen heran-
gezogen werden.
Die Dichtefunktionen φL929 für den Typ L929 und φMC3T3 für den Typ MC3T3
sind in den Abbildungen 69 und 70 dargestellt.
1,00
2,00
4,00
8,00
16,00
32,00
64,00
0 10 20 30 40 50 60 70 80 90 100Ve
rhäl
tnis
Ke
rne
zu
Clu
ste
r
Probe
Merkmal KA - Anzahl der Kerne pro Cluster
Merkmal KA für L929 Merkmal KA für MC3T3

7. Automatisierung der Algorithmenauswahl 116
Abbildung 69: Dichtefunktion des Merkmals KA für den Zelltyp L929
Abbildung 70: Dichtefunktion des Merkmals KA für den Zelltyp MC3T3
Für die Klassifikation eines Bildes in die Klasse L929 oder MC3T3 wird nun der
Merkmalswert x für das Bild bestimmt, dieser in die beiden Dichtefunktionen
φL929 und φMC3T3 eingesetzt und der jeweilige Funktionswert bestimmt. Wenn der
Funktionswert φA(x, µL929, σL929) ≥ φB(x, µMC3T3, σMC3T3) resultiert, erfolgt die Ein-
ordnung des Eingabebildes in die Klasse L929, andernfalls in die Klasse
MC3T3. Liegt eine Klassifizierung als L929 Zellbild vor, kommt das Separati-
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
0,8
0
0,9
2
1,0
4
1,1
6
1,2
8
1,4
0
1,5
2
1,6
4
1,7
6
1,8
8
2,0
0
2,1
2
2,2
4
2,3
6
2,4
8
2,6
0
2,7
2
φ(x
,µ,σ
)
Anz.Kerne/Cluster x
Dichtefunktionen des Merkmals KA für den Zelltyp L929
Dichtefkt. Merkmal KA L929
0,00
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0,08
0,0
0
2,0
0
4,0
0
6,0
0
8,0
0
10
,00
12
,00
14
,00
16
,00
18
,00
20
,00
22
,00
24
,00
26
,00
28
,00
30
,00
32
,00
34
,00
36
,00
38
,00
φ(x
,µ,σ
)
Anz.Kerne/Cluster x
Dichtefunktion des Merkmals KA für den Zelltyp MC3T3
Dichtefkt. Merkmal KA MC3T3

7. Automatisierung der Algorithmenauswahl 117
onsverfahren, beschrieben in Kap. 6.4, zum Einsatz, ansonsten die Methode,
beschrieben in Kap. 6.5.
Zur Bewertung der Qualität des Entscheidungsmerkmals KA wurde für 100
Proben durch die Software automatisch das Separationsverfahren ausgewählt
und geprüft ob die resultierenden Ergebnisse im Vergleich zu den Ergebnissen
mit dem nicht ausgewählten Separationsverfahren besser oder gleich gut sind.
Die Untersuchung zeigt, dass die Software zu 99% das am besten geeignete
Separationsverfahren für die Zelltrennung in Clustern auswählt.
Die Separationsverfahren, beschrieben in den Kapiteln 6.4 und 6.5, setzen die
Detektion der Kernbereiche voraus. Sollte der Fall eintreten, dass keine Zell-
kerne detektiert wurden kommt das Separationsverfahren, beschrieben in Kap.
6.6 zum Einsatz das unter Verwendung morphologischer Operationen die Tren-
nung zusammengewachsener Zellbereiche ohne Berücksichtigung der Kernbe-
reiche vornimmt.

8. Automatische Parametereinstellung 118
8. Automatische Parametereinstellung
Die Parametereinstellungen der für die Zellanalyse benötigten Algorithmen
werden durch die Fachkraft an die zu untersuchende Zelllinie und das Material
angepasst. Die Einstellungen sind in einer Datei speicherbar. Auf diese Weise
ist nur ein einmaliges Einstellen aller Parameter für eine bestimmte Zelllinie und
Material notwendig.
Ist für alle zu untersuchenden Zelltypen und Materialien eine Parameterdatei
generiert worden, ist zu Beginn der Analyse die passende Parameterdatei vom
Anwender auszuwählen.
Zur Einstellung der Algorithmenparameter ist ein bestimmtes Maß an Fachwis-
sen im Bereich Computer Vision notwendig, die Anwender der Software müs-
sen entsprechend geschult werden. Aus diesem Grund ist eine automatische
Lösung zur Bestimmung der Algorithmenparameter für eine Applikation hilf-
reich.
8.1 Automatische Parametereinstellung für die Zellsegmentierung
Das Segmentierungsverfahren, beschrieben in Kap. 5.2.1 kommt z.B. zum Ein-
satz, wenn Proben am Auflichtmikroskop aufgenommen und die darauf befindli-
chen Zellen zuvor angefärbt wurden. Der Algorithmus benötigt 6 Parameter
(S1O,R, S1U,R, S1O,G, S1U,G, S1O,B, S1U,B) zur Einstellung pro Zelltyp und ver-
wendetem Material (Tabelle 37).
Tabelle 37: Schwellwerte zur Segmentierung der L929- und MC3T3-Zellbereiche bei unter-
schiedlichen Materialien; S1O,R: Oberer Schwellwert des Rotkanals; S1U,R: Unterer Schwellwert
des Rotkanals; S1O,G: Oberer Schwellwert des Grünkanals; S1U,G: Unterer Schwellwert des
Grünkanals; S1O,B: Oberer Schwellwert des Blaukanals; S1U,B: Unterer Schwellwert des Blauka-
nals
Proben- material
S1O,R S1U,R S1O,G S1U,G S1O,B S1U,B
L929 Stahl* 210 0 210 0 215 0
Titan* 190 0 190 0 190 0
Polystyrol 175 0 175 0 180 0
MC3T3
Stahl* 210 0 210 0 215 0
Titan* 185 0 185 0 190 0
Polystyrol 170 0 170 0 180 0
Die drei Parameter SU,R, SU,G und SU,B werden auf den Wert 0 gesetzt, da die
Zellbereiche stets dunkler erscheinen als das umgebene Material. Die automa-

8. Automatische Parametereinstellung 119
tische Einstellung der Parameter SO,R, SO,G und SO,B erfolgt nach folgender Vor-
gehensweise.
Zunächst wird das Farbbild in den Rot- Grün- und Blaukanal aufgetrennt. Im
Histogramm des Rotkanals (Abb. 71) folgt die Bestimmung des globalen Maxi-
mums.
Abbildung 71: Histogramm des Rotkanals eines Zellbildes. Die Zellen sind vom Typ L929, May-
Grünwald gefärbt und auf dem Material Stahl aufgebracht. Das globale Maximum liegt auf dem
Index 229 mit 161.299 Pixeln
Der Index des globalen Maximums entspricht dem häufigsten im Bild vorkom-
menden Grauwert. Dieser kann dem verwendeten Material zugeordnet werden,
da es die meiste Fläche im Bild belegt. Ausgehend vom Index des globalen
Maximums wird schrittweise der Index um eins verkleinert bis die Anzahl der
Pixel an der aktuell betrachteten Histogrammposition weniger als p = 3% der
Anzahl an Pixeln am absoluten Maximum entspricht. Der aktuelle Histogram-
mindex wird dem Parameter S1O,R zugewiesen. Aufgrund der Farbvielfalt der
Zellen ist die Wahrscheinlichkeit hoch dass Grauwerte bis zum aktuellen Histo-
grammindex den Zellbereichen zugeordnet werden können. Für die anderen
beiden Kanäle wird analog verfahren und die jeweiligen resultierenden Indizes
den Parametern SO,G und SO,B zugewiesen. Zur Bestimmung des optimalen
Prozentwertes p wurden von 100 zufällig ausgewählten Zellen die Referenzkon-
turen ermittelt und mit den automatisch generierten Zellkonturen verglichen. Als
Vergleichsmaß kam der Jaccard-Koeffizient zum Einsatz. Unter der Verwen-
dung verschiedener Prozentsätze hat sich p = 3% als optimal herausgestellt, da
für 100 untersuchte Zellen hierfür der größte Jaccard-Koeffizient resultierte.
Für die Detektion der Hintergrundbereiche innerhalb der Zellen (Abb. 17) sind
ebenfalls 3 der 6 Parameter automatisch einzustellen (Tabelle 38).

8. Automatische Parametereinstellung 120
Tabelle 38: Schwellwerte zur Segmentierung der Materialbereiche innerhalb der bereits erfass-
ten Zellbereiche; S2O,R: Oberer Schwellwert des Rotkanals; S2U,R: Unterer Schwellwert des
Rotkanals; S2O,G: Oberer Schwellwert des Grünkanals; S2U,G: Unterer Schwellwert des Grünka-
nals; S2O,B: Oberer Schwellwert des Blaukanals; S2U,B: Unterer Schwellwert des Blaukanals
Proben- material
S2O,R S2U,R S2O,G S2U,G S2O,B S2U,B
Stahl* 255 215 255 215 255 220
Titan* 255 195 255 195 255 195
Polystyrol 255 180 255 180 255 185
Da der Hintergrundbereich heller als die Zellen erscheint und die oberen
Schwellwerte für die Detektion der Zellbereiche bereits ermittelt sind, werden
die unteren Segmentierungsschwellen S2U,R, S2U,G und S2U,B auf folgende Wer-
te gesetzt:
S2U,R = S1O,R + 1
S2U,G = S1O,G + 1
S2U,B = S1O,B + 1
Der Algorithmenablauf ist in Form eines Flussdiagramms in Abb. 72 dargestellt.

8. Automatische Parametereinstellung 121
Abbildung 72: Flussdiagramm zum Ablauf der automatischen Parametereinstellung
Nach Ausführung der automatischen Parameterfestlegung ergeben sich für die
verschiedenen Materialen und Zelltypen die in Tabelle 39 enthaltenen Schwell-
werte.
Tabelle 39: Schwellwerte zur Segmentierung der L929- und MC3T3-Zellbereiche bei unter-
schiedlichen Materialien; S1O,R: Oberer Schwellwert des Rotkanals; S1U,R: Unterer Schwellwert
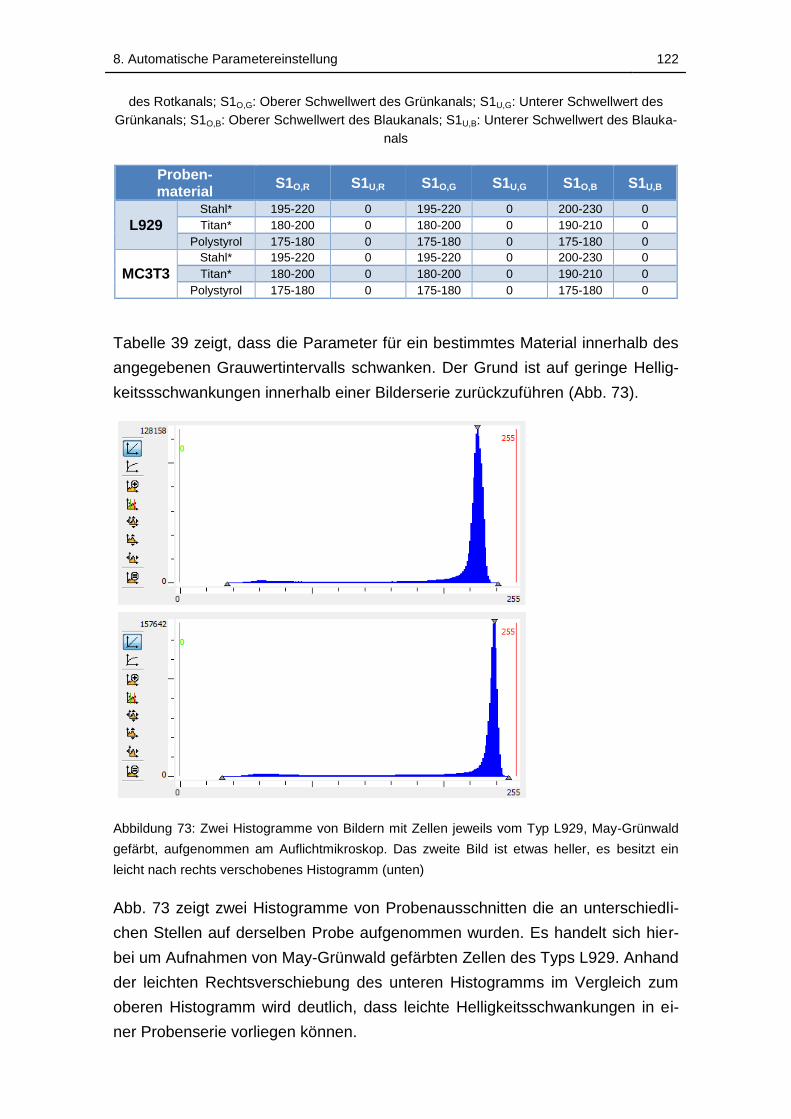
8. Automatische Parametereinstellung 122
des Rotkanals; S1O,G: Oberer Schwellwert des Grünkanals; S1U,G: Unterer Schwellwert des
Grünkanals; S1O,B: Oberer Schwellwert des Blaukanals; S1U,B: Unterer Schwellwert des Blauka-
nals
Proben- material
S1O,R S1U,R S1O,G S1U,G S1O,B S1U,B
L929 Stahl* 195-220 0 195-220 0 200-230 0
Titan* 180-200 0 180-200 0 190-210 0
Polystyrol 175-180 0 175-180 0 175-180 0
MC3T3 Stahl* 195-220 0 195-220 0 200-230 0
Titan* 180-200 0 180-200 0 190-210 0
Polystyrol 175-180 0 175-180 0 175-180 0
Tabelle 39 zeigt, dass die Parameter für ein bestimmtes Material innerhalb des
angegebenen Grauwertintervalls schwanken. Der Grund ist auf geringe Hellig-
keitssschwankungen innerhalb einer Bilderserie zurückzuführen (Abb. 73).
Abbildung 73: Zwei Histogramme von Bildern mit Zellen jeweils vom Typ L929, May-Grünwald
gefärbt, aufgenommen am Auflichtmikroskop. Das zweite Bild ist etwas heller, es besitzt ein
leicht nach rechts verschobenes Histogramm (unten)
Abb. 73 zeigt zwei Histogramme von Probenausschnitten die an unterschiedli-
chen Stellen auf derselben Probe aufgenommen wurden. Es handelt sich hier-
bei um Aufnahmen von May-Grünwald gefärbten Zellen des Typs L929. Anhand
der leichten Rechtsverschiebung des unteren Histogramms im Vergleich zum
oberen Histogramm wird deutlich, dass leichte Helligkeitsschwankungen in ei-
ner Probenserie vorliegen können.

8. Automatische Parametereinstellung 123
8.2 Automatische Parametereinstellung für die CBS Methode
Die kontextbasierte Separationsmethode (CBS), beschrieben in Kap. 6.1, wird
nach der Zellbereichsegementierung zur ersten Stufe der Zellseparation einge-
setzt. Die Einstellung des Algorithmus erfolgt über die folgenden vier Parame-
ter:
r: Radius der Kreismaske C1
a: Festlegung des Radius der Kreismaske C2 mit r2 = a · r
x, y: Parameter der Entscheidungsfunktion f2(x,y) für eine Engstelle in der Zelle
Zur automatischen Einstellung der Parameter ist der Einsatz von genetischen
Algorithmen möglich. Mit ihrer Hilfe können komplexe Optimierungsaufgaben in
vertretbarer Zeit gelöst werden. Die Funktionsweise lehnt sich, jedoch verein-
facht, an die natürliche Evolution von Lebewesen an und läuft prinzipiell nach
folgendem Schema ab (80):
1. An die Problemstellung angepasste genetische Codierung der Lösungskan-
didaten vornehmen
2. Initialisierung: Anfangspopulation erzeugen mit meist zufälliger Zuordnung
von Fitnesswerten.
3. Durchlaufe die folgenden Schritte, bis ein Abbruchkriterium erfüllt ist:
- Güte der Lösungskandidaten mit Hilfe der Fitnessfunktion berechnen
- Selektion der Eltern für die nachfolgende Rekombination
- Rekombination der ausgewählten Lösungskandidaten
- Zufällige Veränderung der Nachfahren (Mutation)
- Reproduktion der neuen Generation und Prüfen der Abbruchbedingung
Für die vorliegende Anwendung wird zunächst der Wertebereich der o.g. vier
Parameter wie folgt festgelegt.
r: 1, 2, 3, …, 50
a: 1.5, 2.0, 2.5, …, 6.0
x: 1.00, 1.05, 1.10, …, 3.00
y: 1.2, 1.4, 1.6, …, 10.0
Der Wertebereich und die Quantisierung der Parameter sind für die vorliegende
Anwendung angepasst. Mit den vorliegenden 4 Wertebereichen ergeben sich
50x10x41x45 = 922.500 Parameterkonfigurationen, aus denen eine für den

8. Automatische Parametereinstellung 124
Zelltyp L929 geeignete Parametereinstellung gefunden werden soll. Der Typ
L929 wird hier vorrangig untersucht, da dieser typischerweise für Biokompatibili-
tätstests herangezogen wird.
Als Codierungsvariante kommt in diesem Fall die Realzahlcodierung zum Ein-
satz, da die Parameterwerte nicht nur ganzzahlige Werte annehmen können.
Ein Individuum I bzw. ein Lösungskandidat besitzt die folgende Form:
I = {r, a, x, y}
Zur Bestimmung der Güte eines Individuums wird eine Fitnessfunktion benötigt.
Diese Funktion vergleicht die automatisch ermittelte Zellanzahl mit der Refe-
renzzählung unter Verwendung einer bestimmten Parameterkonfiguration. Für
die automatische Bestimmung der Zellanzahl kommt nach der Zellbereichseg-
mentierung die CBS Methode zur Separation zusammenhängender Zellen zum
Einsatz, wobei die verwendete Parameterkonfiguration von dem aktuell zu be-
wertendem Individuum repräsentiert wird. Das setzt eine vorherige manuelle
Zellzählung eines Referenzbildes mit Zellclustern voraus.
Im nächsten Schritt erfolgt die Selektion der Elternpaare für die anschließende
Rekombination. Dazu kommt das „Roulette-Wheel-Selection“ Verfahren zum
Einsatz, bei dem zunächst die Anteile der normierten Fitnesswerte
,
1
ii prop n
i
i
FF
F
(Gl. 24)
auf eine Drehscheibe aufgetragen werden. Abb. 74 verdeutlicht beispielhaft das
Vorgehen anhand von 10 Individuen.
Abbildung 74: Beispielhafte fitnessproportionale Aufteilung von 10 Individuen auf einer Scheibe.
Ein Zufallswert im Bereich 1 bis 360 (schwarzer Pfeil) wählt ein Individuum für die Reproduktion
aus.
Die Selektion erfolgt durch n-maliges Drehen der Scheibe, wobei nach jedem
Drehvorgang ein Elternteil ausgewählt wird. Dabei kann der Fall eintreten, dass
ein Individuum mehrmals ausgewählt wird. Der Drehvorgang wird hier durch die
Fitnessverteilung der Individuen Auswahl

8. Automatische Parametereinstellung 125
Bestimmung einer Zufallszahl zwischen 1 bis 360 simuliert. Ausgewählt ist da-
raufhin das Individuum, dessen Winkelbereich auf der Drehscheibe von der Zu-
fallszahl getroffen wurde.
Daraufhin steht die Rekombination der selektierten Elternpaare an. Dies ge-
schieht mit Hilfe der ganzarithmetischen Rekombination. Bei dieser Methode
werden für alle Gene neue Werte bestimmt. Die Werte ergeben sich aus der
gewichteten Summe der Elterngene, wobei die Gewichtung über den Parameter
α festgelegt ist:
1 1 1 11 , , ,I r x y
2 2 2 22 , , ,I r x y
1 2 1 2 1 2 1 21 1 , 1 , 1 , 1K r r a a x x y y (Gl. 25)
2 1 2 1, 2 1 2 12 1 , 1 1 , 1K r r a a x x y y (Gl. 26)
Zum Beispiel ergeben sich unter Verwendung des Parameters α = 0,7 für die
folgenden zwei Elternteile I1 und I2 die zwei Nachkommen K1 und K2:
I1 = {r1=8, a1=2.0, x1=1.1, y1=3.0} K1 = {8.9, 2.15, 1.19, 3.3}
I2 = {r2=11, a2=2.5, x2=1.40, y2=4.0} K2 = {10.1, 2.35, 1.31, 3.7}
Im letzten Schritt der Rekombination müssen die berechneten Genwerte auf
den nächstgelegenen gültigen Wert des Wertebereiches gerundet werden:
K1 = {9, 2.0, 1.20, 3.4}
K2 = {10, 2.5, 1.30, 3.8}
Nach der Rekombination erfolgt eine Mutation der Gene, die mit einer Wahr-
scheinlichkeit von p = 1% pro Gen auftritt. Mit Hilfe der Mutationsoperation ist
es möglich eine gewisse Inhomogenität in der Population aufrecht zu erhalten.
Erfolgt eine Mutation, so wird der Wert des betrachteten Gens zufällig auf einen
gültigen Wert innerhalb des Wertebereiches gesetzt.
8.2.1 Ergebnisse zur automatischen Parametereinstellung für die CBS Methode
Für die Durchführung der Parameterfestlegung mit dem genetischen Algorith-
mus wird eine Anfangspopulation von 20 Individuen gewählt und im Rahmen
von 50 Generationen nach dem besten Parametersatz gesucht. Dabei beträgt
die Rechenzeit auf einem intel Core i7 mit 3,2Ghz ca. 60s. Wird der genetische
α = 0,7
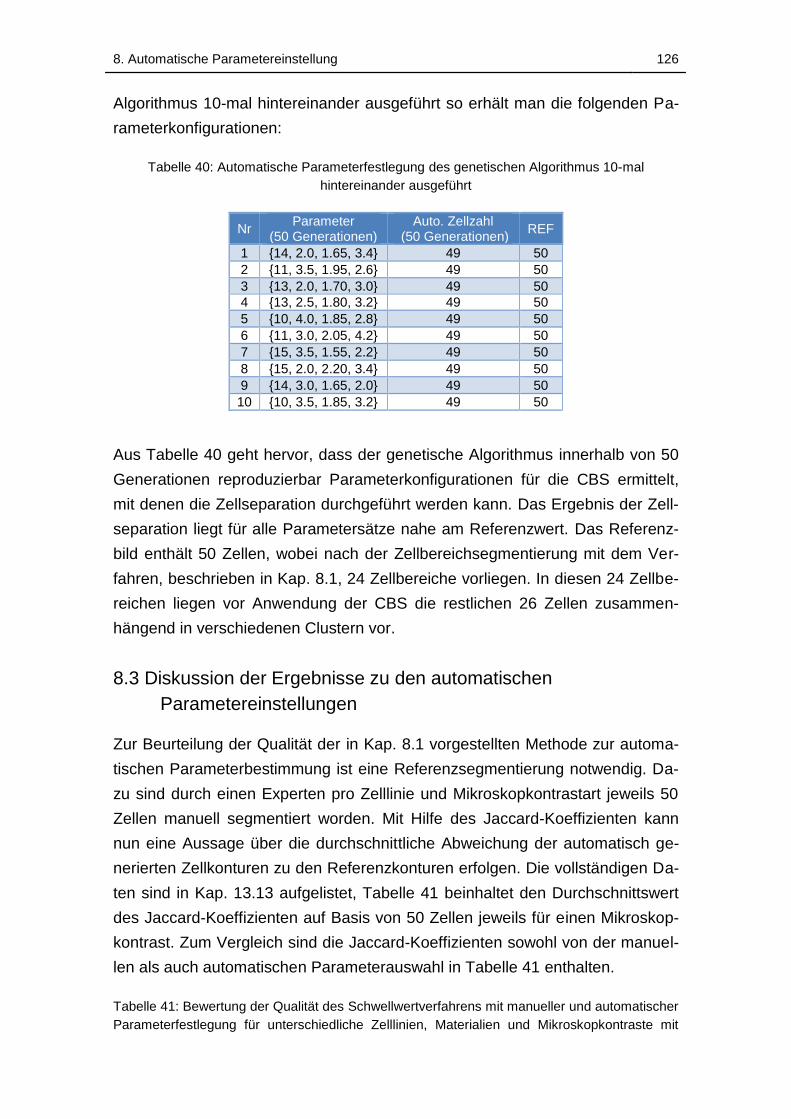
8. Automatische Parametereinstellung 126
Algorithmus 10-mal hintereinander ausgeführt so erhält man die folgenden Pa-
rameterkonfigurationen:
Tabelle 40: Automatische Parameterfestlegung des genetischen Algorithmus 10-mal
hintereinander ausgeführt
Nr Parameter
(50 Generationen) Auto. Zellzahl
(50 Generationen) REF
1 {14, 2.0, 1.65, 3.4} 49 50
2 {11, 3.5, 1.95, 2.6} 49 50
3 {13, 2.0, 1.70, 3.0} 49 50
4 {13, 2.5, 1.80, 3.2} 49 50
5 {10, 4.0, 1.85, 2.8} 49 50
6 {11, 3.0, 2.05, 4.2} 49 50
7 {15, 3.5, 1.55, 2.2} 49 50
8 {15, 2.0, 2.20, 3.4} 49 50
9 {14, 3.0, 1.65, 2.0} 49 50
10 {10, 3.5, 1.85, 3.2} 49 50
Aus Tabelle 40 geht hervor, dass der genetische Algorithmus innerhalb von 50
Generationen reproduzierbar Parameterkonfigurationen für die CBS ermittelt,
mit denen die Zellseparation durchgeführt werden kann. Das Ergebnis der Zell-
separation liegt für alle Parametersätze nahe am Referenzwert. Das Referenz-
bild enthält 50 Zellen, wobei nach der Zellbereichsegmentierung mit dem Ver-
fahren, beschrieben in Kap. 8.1, 24 Zellbereiche vorliegen. In diesen 24 Zellbe-
reichen liegen vor Anwendung der CBS die restlichen 26 Zellen zusammen-
hängend in verschiedenen Clustern vor.
8.3 Diskussion der Ergebnisse zu den automatischen
Parametereinstellungen
Zur Beurteilung der Qualität der in Kap. 8.1 vorgestellten Methode zur automa-
tischen Parameterbestimmung ist eine Referenzsegmentierung notwendig. Da-
zu sind durch einen Experten pro Zelllinie und Mikroskopkontrastart jeweils 50
Zellen manuell segmentiert worden. Mit Hilfe des Jaccard-Koeffizienten kann
nun eine Aussage über die durchschnittliche Abweichung der automatisch ge-
nerierten Zellkonturen zu den Referenzkonturen erfolgen. Die vollständigen Da-
ten sind in Kap. 13.13 aufgelistet, Tabelle 41 beinhaltet den Durchschnittswert
des Jaccard-Koeffizienten auf Basis von 50 Zellen jeweils für einen Mikroskop-
kontrast. Zum Vergleich sind die Jaccard-Koeffizienten sowohl von der manuel-
len als auch automatischen Parameterauswahl in Tabelle 41 enthalten.
Tabelle 41: Bewertung der Qualität des Schwellwertverfahrens mit manueller und automatischer
Parameterfestlegung für unterschiedliche Zelllinien, Materialien und Mikroskopkontraste mit

8. Automatische Parametereinstellung 127
Hilfe des Jaccard-Koeffizienten; A-HF: Auflicht-Hellfeld; A-DF: Auflicht-Dunkelfeld; D-HF: Durch-
licht-Hellfeld; D-PK: Durchlicht-Phasenkontrast
Material L929 MC3T3
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
Manuelle Parameterfestlegung
Stahl*/Titan* 0,89 0,85 --- --- 0,87 0,90 --- ---
Polystyrol --- --- 0,87 0,60 --- --- 0,76 0,53
Automatische Parameterfestlegung
Stahl* /Titan* 0,92 0,87 --- --- 0,90 0,92 --- ---
Polystyrol --- --- 0,90 0,65 --- --- 0,78 0,55
Aus Tabelle 41 geht hervor, dass bei der automatischen Parameterauswahl
geringfügig bessere Jaccard-Koeffizienten im Vergleich zur manuellen Parame-
terfestlegung resultieren. Der Grund liegt darin, dass bei der automatischen Pa-
rameterfestlegung die leichten Helligkeitsschwankungen zwischen einzelnen
Bildaufnahmen ausgeglichen werden.
Bei Verwendung des Durchlicht-Phasenkontrastes resultieren für beide Varian-
ten der Parameterauswahl mit Jaccard-Koeffizienten von 0,60 und 0,65 keine
verwertbaren Ergebnisse, da sowohl Zellen als auch das Substrat teilweise die
gleiche Farbgebung aufweisen und durch global einstellbare Schwellwerte nicht
mehr hinreichend genau zwischen Vorder- und Hintergrund unterschieden wer-
den kann.
Bei der Parameterauswahl für die CBS Methode mit Hilfe eines genetischen
Algorithmus resultieren bei mehrmaliger Ausführung nicht die gleichen Parame-
tersätze (Tabelle 40). Der Grund liegt in der zufälligen Festlegung der Startpa-
rameter sowie der zufälligen Mutation von Individuen, die schließlich zu unter-
schiedlichen Parameterwerten führen können. Allerdings weichen die Parame-
terkonfigurationen nicht deutlich voneinander ab und die CBS Methode liefert
für die in Tabelle 40 aufgeführten Parametersätze die gleiche Qualität. Diese
Eigenschaft der CBS Methode, für unterschiedliche Parameterkonfigurationen
gleiche Ergebnisse zu erzielen, befürwortet den Einsatz eines genetischen Al-
gorithmus zur automatischen Parameterfestlegung da die Wahrscheinlichkeit
hoch ist mit einer mäßigen Anzahl an Generationen eine gute Lösung zu erzie-
len. Die Verwendung der unterschiedlichen Parameterkonfigurationen aus Ta-
belle 40 führen bei der automatischen Zellzählung lediglich zu geringen Abwei-

128
chungen. Setzt man die folgenden drei Parametersätze für die CBS-Methode
ein
I1 = {14, 2.0, 1.65, 3.4}
I2 = {11, 3.0, 2.05, 4.2}
I3 = {10, 3.5, 1.85, 3.2}
und führt eine Zellzählung in Kombination mit dem Zellbereichsegmentierungs-
verfahren, beschrieben in Kap. 8.1, und dem Zellseparationsverfahren, be-
schrieben in Kap. 6.5 durch, so ergeben sich für jeweils 20 Probenausschnitte
durchschnittliche Abweichungen von 1,38%, 1,47% und 1,44% zur Referenz-
zählung. Eine Stichprobe von 20 Probenausschnitten reicht hier aus um zu zei-
gen, dass die Abweichungen der automatischen Zellzählungen bei Verwendung
unterschiedlicher Parametersätze, die alle als die beste Lösung vom geneti-
schen Algorithmus vorgeschlagen wurden, mit 0,09% und 0,06% sehr gering
sind.

9. Bewertung des Biokompatibilitätsgrades 129
9. Bewertung des Biokompatibilitätsgrades
Die in Kap. 5 und 6 vorgestellten Algorithmen ermöglichen die Segmentierung
und Separation von L929-, MC3T3- oder ähnlichen Zelltypen in mikroskopi-
schen Bildern, so dass nachfolgende Zellanalysen mittels der Software möglich
sind. Die Bewertung des Biokompatibilitätsgrades erfolgt laut DIN ISO Norm
10993-5 (5) entweder durch quantitative oder qualitative in-vitro-Zellversuche,
wobei das Prüfmaterial einer der in Tabelle 42 aufgeführten Zytotoxizitätsstu-
fen14 zugeordnet wird.
Tabelle 42: Zytotoxische Einteilung der Prüfmaterialien laut DIN ISO Norm 10993-5 (5)
Zytotoxizitäts-skala
Interpretation
0 Nicht zytotoxisch
1 Schwach zytotoxisch
2 Mäßig zytotoxisch
3 Stark zytotoxisch
9.1 Quantitative Bewertung der Biokompatibilität
Die quantitative Bewertung der Biokompatibilität eines Substrates (Probenwerk-
stoff) erfolgt mit Hilfe der Zellproliferation, d.h. es wird das Wachstums- und
Vermehrungsverhalten der Zellen nach einem festgelegten Testzeitraum in
Kontakt mit dem Probenwerkstoff untersucht. In Standardlaboren wird zur Be-
stimmung der Zellproliferation (nach Ablauf der Versuchsdauer) die Anzahl vita-
ler Zellen, die in Gegenwart des Probenwerkstoffes überlebt haben, mit speziel-
len Zählkammern ermittelt. Mit Hilfe der entwickelten Software ist hingegen eine
automatische Zellzählung und Morphologieanalyse für mikroskopische Aufnah-
men möglich. Die Arbeitsschritte im Rahmen der Zellzahlbestimmung mit Hilfe
der Zählkammern (Verfahren A) und unter Verwendung einer computergestütz-
ten Analyse (Verfahren B) sind in Abb. 75 dargestellt.
14
Zytotoxizität: Unter Zytotoxizität versteht man die schädigende Wirkung chemischer Substan-
zen (hier der Probenwerkstoff) auf damit in Verbindung stehende Zellen.

9. Bewertung des Biokompatibilitätsgrades 130
Abbildung 75: Vergleich der Arbeitsschritte der herkömmlichen Zellzahlbestimmung mit Hilfe
von Zählkammern (Verfahren A) mit der computergestützten Auswertung der Zellbilder (Verfah-
ren B). Erklärung siehe Text.
Zu Beginn müssen für beide Verfahren A und B die vorbereitenden Maßnah-
men für die zu testenden Probenwerkstoffe getroffen werden (Schritt 1). Dieser
Arbeitsschritt zur Probenaufbereitung ist bereits in Kap. 3 beschrieben. Es folgt,
ebenfalls für beide Verfahren analog, das Aufbringen der Zellen auf die Pro-
benoberfläche (Schritt 2). Dies geschieht mit Hilfe der Suspensionsmethode,
Abb. 76.

9. Bewertung des Biokompatibilitätsgrades 131
Abbildung 76: Suspensionsmethode zum Aufbringen der Zellen auf die Probenoberfläche
Das zu prüfende Material wird in ein Well (gefäßartige Struktur) gegeben und
vollständig mit der Zellsuspension übergossen. Aufgrund der im Vergleich zur
umgebenden Flüssigkeit höheren Dichte der Zellen, sinken diese mit fortschrei-
tender Zeit auf die Probenoberfläche.
Metallische Materialien neigen in wässriger Umgebung (z.B. Nährmedium oder
Blut) zur Korrosion, wobei Ionen in das Medium abgegeben werden (81; 82).
Diese Ionen können das Wachstum und die Morphologie der Zellen beeinflus-
sen. Aufgrund dieser Tatsache ist ein separater Ansatz der nichttoxischen Refe-
renzproben in einem eigenen Well erforderlich, da sonst der Referenzwert
durch die herausgelösten Metallionen verfälscht werden könnte.
Die mit Zellen behafteten Proben werden anschließend für 72 Stunden im Brut-
schrank inkubiert15 (Schritt 3). Während dieser Zeit durchlaufen die Zellen einen
Wachstums- und Teilungsprozess, der, abhängig von der Toxizität des zugrun-
deliegenden Materials, zu unterschiedlichen Populationen führt.
Die Vorgehensweise zur Bestimmung der Zellanzahl auf den Proben unter-
scheidet sich in den nachfolgenden Arbeitsschritten zwischen den beiden Me-
thoden.
Nach Verfahren A steht nun die Überführung der Metallproben in ein neues
Well an. Dadurch ist sichergestellt, dass nur die Zellen auf der Probenoberflä-
che abgelöst werden und nicht diejenigen, welche sich neben der Probe auf
dem Kulturgefäß angeheftet haben (Schritt 4.1A).
Beim Ablösen der Zellen von der Probenoberfläche entstehen 1000µl Zellsus-
pension (Schritt 4.2A). Von dieser Lösung werden 20µl in eine Fuchs-
Rosenthal-Zählkammer pipettiert. Es folgt die Zählung aller Zellen, die sich in
einem definierten Raster der Zählkammer mit einem Volumen von 3,2µl befin-
den. Die resultierende Zellanzahl wird auf das Volumen von 1000µl hochge-
rechnet. Methode A sieht eine 10-malige Wiederholung des Pipettier- und Zähl-
15
Inkubieren: Unter inkubieren ist das heranzüchten der Zellen unter bestimmten Aussenbedin-
gungen (5% CO2-Begasung, 37°C) im Inkubator (Brutschrank) zu verstehen.

9. Bewertung des Biokompatibilitätsgrades 132
vorgangs vor. Anschließend erfolgt die Mittelwertbildung der 10 resultierenden
Zellzahlen xi mit i = 1, 2, …, 10 gemäß Gl. 27 (Schritt 4.3.A).
10
1
11000
10 3,2
i
i
xx
(Gl. 27)
Verfahren B sieht nach dem Zellwachstum im Brutschrank im Gegensatz zu
Verfahren A eine Färbung der Zellen auf der Probenoberfläche vor (Schritt
4.1B). Zum Einsatz kommt hier die May-Grünwald-Färbung, aus den in Kap.
3.2.1 genannten Gründen. Damit ist die Voraussetzung für eine automatische
optische Detektion der Zellen und Kernbereiche auf der Probe gegeben. Nach
der Färbung folgt die Aufnahme der Probenoberfläche unter dem Mikroskop
(Schritt 4.2B). Untersuchungen zur Eignung einer Stichprobe bei der Aufnahme
der Materialoberfläche sind in Kap. 9.2 erläutert. Eine Analysesoftware ermittelt
daraufhin die Zellanzahl auf der Probenoberfläche (Schritt 4.3B).
Mit beiden Verfahren ist sowohl die Zellanzahl PR der Referenzprobe als auch
die Zellanzahl PM des zu testenden Materials zu bestimmen. Mit 100M
R
PP
P
wird die Proliferationsrate der Testprobe berechnet. Abhängig von der Prolifera-
tionsrate erfolgt, analog zu (82), die Einstufung des Testmaterials in eine Zyto-
toxizitätsstufe, gemäß Tabelle 43.
Tabelle 43: Zuordnung der Zellproliferation zu den vier Stufen der Zytotoxizität
Zytotoxizitäts-skala
Proliferationrate P [%]
Interpretation
0 100 – 81 Nicht zytotoxisch
1 80 – 71 Schwach zytotoxisch
2 70 – 61 Mäßig zytotoxisch
3 60 - 0 Stark zytotoxisch
9.2 Untersuchung verschiedener Stichprobenumfänge zur
Berechnung der durchschnittlichen Zellanzahl einer Probe
Zur Bestimmung der Proliferationsrate mit Hilfe der Software ist, wie bereits in
Kap. 9.1 beschrieben, die Ermittlung der durchschnittlichen Zellanzahl auf einer
Probe Voraussetzung. Die Probenoberfläche weist eine Fläche von 3,14cm2
auf. Für eine optimale automatische Zellanalyse werden die Bilder, abhängig
vom Zelltyp, mit einer Vergrößerung von Faktor 50 oder 100 aufgenommen.
Erfolgt die Aufnahme mit einer Vergrößerung von Faktor 50, sind bereits 128
Bildaufnahmen zur Abbildung der gesamten Probe notwendig. Zur Reduzierung

9. Bewertung des Biokompatibilitätsgrades 133
des damit verbundenen Arbeitsaufwandes ist die Verwendung einer Stichprobe
denkbar, die die durchschnittliche Zellanzahl der Probe repräsentieren soll.
Es werden verschiedene Stichprobenumfänge festgelegt und jeweils die Abwei-
chung vom Referenzwert bestimmt. Unter dem Referenzwert ist hier die durch-
schnittliche Zellanzahl aller Probenausschnitte zu verstehen, die keine Proben-
randbereiche aufweisen. Nach Abzug der Bilder, auf denen der Probenrand
sichtbar ist, verbleiben von den 128 noch 96 Probenausschnitte.
Problematisch bei der Auswahl der Ausschnitte ist jedoch, dass die Zelldichte
auf der Probenoberfläche variiert (Abb. 77).
Abbildung 77: Links: L929-Zellen auf dem Material Stahl mit vergleichsweise schwacher Zell-
dichte. Rechts: Deutlich höhere Zelldichte auf derselben Probe an einer anderen Position
Daher ist es sinnvoll, die Selektion der Ausschnitte nicht zufällig, sondern mit
Hilfe eines Experten durchzuführen, wobei Probenbereiche mit einer mittleren
Zelldichte ausgewählt werden. Die vollständigen Daten zur Untersuchung der
Eignung verschiedener Stichprobenumfänge sind in Kap. 13.14 aufgeführt. Ta-
belle 44 zeigt die aus der Untersuchung resultierende maximale positive und
negative Abweichung der durchschnittlichen Zellanzahl vom Referenzwert für
unterschiedliche Stichprobenumfänge.
Tabelle 44: Verschiedene Stichprobenumfänge mit der jeweiligen Abweichung zum Referenz-
wert. Für jeden Stichprobenumfang wurden 10 Versuche durchgeführt. Zeichenerklärung: Abw.:
Die maximale positive und negative Abweichung der durchschnittlichen Zellanzahl vom Refe-
renzwert [%]
Stichprobenumfänge
zufällige Auswahl der Stichprobe
Nr. 80 Bilder 50 Bilder 30 Bilder 15 Bilder
Abw. -4,2% bis 6,0% -7,5% bis +13,5% -17,4% bis +36,4% -20,6% bis +23,5%
gezielte Auswahl der Stichprobe
Nr. 20 Bilder 15 Bilder 10 Bilder 5 Bilder
Abw. -4,8% bis +4,6% -4,7% bis +2,2% -12,6% bis +6,4% -20,0% bis 9,6%

9. Bewertung des Biokompatibilitätsgrades 134
Tabelle 44 zeigt zunächst die Abweichungen der ermittelten durchschnittlichen
Zellanzahl zum Referenzwert bei zufälliger Auswahl des Bildmaterials. Für ei-
nen Stichprobenumfang von 50, der ca. die Hälfte der Gesamtmenge ohne
Probenrand entspricht, streut für 10 Versuche die Abweichung der durchschnitt-
lichen Zellanzahl der Stichprobe im Vergleich zum Referenzwert von -7,5% bis
+13,5%.
In Tabelle 43 ist für die Einstufung der Toxizität eines Materials in den Grad
„nicht zytotoxisch“ ein Intervall von 20% der Proliferationsratenänderung ange-
geben und für die Einstufung in die Klassen „schwach zytotoxisch“ sowie „mä-
ßig zytotoxisch“ noch 10%. Mit der genannten Streuung von -7,5% bis 13,5%
um den Referenzwert ist jedoch eine sichere Zuordnung in eine bestimmte Zy-
totoxizitätsstufe nicht gewährleistet. Daher ist der Einsatz einer zufälligen Stich-
probe hier ungeeignet. Der Grund liegt in der ungleichmäßigen Verteilung der
Zellen auf der Probenoberfläche (Abb. 78).
Abbildung 78: Zellanzahl für 96 verschiedene Ausschnitte einer Probe (blaue Raute). Die
durchschnittliche Zellanzahl aller Probenausschnitte beträgt 477,23 Zellen (rotes Quadrat)
Abb. 78 zeigt, dass die Zelldichte auf der Probenoberfläche variiert. Die meisten
Probenausschnitte liefern eine Population zwischen 300 – 500 Zellen. Einige
Bereiche weisen jedoch deutlich höhere (> 1000 Zellen) Populationen auf. Dies
führt zu einer hohen Standardabweichung von 352,65 Zellen. Die vergleichs-
weise hohe Zellanzahl von Probenausschnitten mit höherem Index ist darauf
zurückzuführen, dass die Zelldichte im oberen Probenbereich zunimmt (Abb.
78). Die unterschiedliche Zelldichteverteilung ist wahrscheinlich auf die Präpa-
rationstechnik zurückzuführen und nicht reproduzierbar.
0
500
1000
1500
2000
2500
0 20 40 60 80 100 120
Zella
nza
hl p
ro P
rob
en
auss
chn
itt
Bildnummer (Index) des Probenausschnittes
Zellanzahl proProbenausschnitt
Mittelwert

9. Bewertung des Biokompatibilitätsgrades 135
Abbildung 79: Schematische Darstellung der Vorgehensweise bei den Bildaufnahmen der voll-
ständigen Probe. Die Aufnahmen beginnen am rechten unteren und enden am linken oberen
Probenbereich
Die Aufnahmen der Probenausschnitte erfolgen mäanderförmig, beginnend am
unteren rechten Probenbereich. Die Bilder werden nummeriert, der Index läuft
von 1 bis 128.
Tabelle 44 zeigt weiterhin die Streuung der Abweichung der durchschnittlichen
Zellanzahl zum Referenzwert für verschiedene, gezielt ausgewählte Stichpro-
benumfänge. Die geringste Abweichung resultiert unter Verwendung einer
Stichprobe mit 15 Bildern. Für 10 Durchläufe ergibt sich eine Abweichung von
-4,7% bis +2,2% vom Referenzwert. Für einen Stichprobenumfang von 20 Bil-
dern findet keine Reduzierung der Abweichung mehr statt. Im Gegenteil, die
Abweichung zum Referenzwert erhöht sich mit -4,8% bis +4,6% leicht. Dies
lässt darauf schließen, dass sich die Abweichung zum Referenzwert bei geziel-
ter Auswahl der Bildausschnitte, trotz Einsatz eines größeren Stichprobenum-
fanges, im Durchschnitt nicht weiter verringern lässt.
Für die Biokompatibilitätsanalyse von Implantatmaterialien ist es notwendig, ein
zu prüfendes Material nicht bioverträglicher einzustufen als es tatsächlich der
Fall ist. Tritt dieser Fehler dennoch auf, könnte im schlimmsten Fall die Absto-
ßung des Implantates vom umliegenden Körpergewebe die Folge sein. Daher
ist es empfehlenswert, die, abhängig von der Vorgehensweise bei der Prüfung,
ermittelten Abweichungen zum Referenzwert bei der Einordnung in die Zytoto-
xizitätsstufen zu berücksichtigen. Tabelle 45 zeigt die Anpassung der unteren
Zellproliferationsgrenze für die Zytotoxizitätsstufe „nicht zytotoxisch“ unter Be-
rücksichtigung des Gesamtfehlers bei Verwendung eines Stichprobenumfanges
mit 15 Bildern sowie der Zellzählung der vollständigen Probe.
Start (Index:1)
Ende (Index: 128)

9. Bewertung des Biokompatibilitätsgrades 136
Tabelle 45: Anpassung der Zellproliferationsgrenzen für die Einordnung in die Zytotoxizitätsstu-
fe „nicht zytotoxisch“ unter Berücksichtigung der Gesamtabweichung bei Verwendung einer
Stichprobe mit 15 Bildern und bei Verwendung der vollständigen Probe für die Zellzählung
Zytotoxizitätsprüfung mit einer Stichprobe
von 15 Bildern
(99% Konfidenzintervall)
Zytotoxizitätsprüfung unter Berücksichtigung der voll-
ständigen Probe
(99% Konfidenzintervall)
Fehler bei Verwendung einer Stichprobe (Tabelle 43)
+/- 5% 0%
Fehler der computergestützten Zellzählung von L929-Zellen
1,43% +/- 0,69% 1,27% +/- 0,22%
Max. Gesamtfehler 7,12% ≈ 1,49% Erlaubte Proliferationsrate für die
Einstufung des untersuchten Materials in die Toxizitätsstufe
„nicht zytotoxisch“ gemäß Tabelle 43 unter Berücksichtigung des hier ermittelten Gesamtfehlers
100 – 88,12% 100 – 82,51%
Tabelle 45 (links) zeigt, dass unter Berücksichtigung des maximalen Gesamt-
fehlers von 7,12% für den Einsatz einer Stichprobe mit 15 Bildern bis zu einer
Proliferationsrate von 88,12% im Vergleich zur Referenzprobe eine zu 99% si-
chere Zuordnung in die Zytotoxizitätsstufe „nicht zytotoxisch“ gewährleistet ist.
Unter Verwendung der vollständigen Probe für die Zellzählung ist hingegen eine
zu 99% sichere Zuordnung in die Zytotoxizitätsstufe „nicht zytotoxisch“ mit einer
Proliferationsrate von bereits 82,51% zum Referenzwert möglich, da der max.
Gesamtfehler mit 1,49% erheblich kleiner ist als beim Einsatz der Stichprobe.
Die Bestimmung der Zellanzahl der vollständigen Probe erfordert jedoch 128
Probenausschnitte im Vergleich zu den 15 Ausschnitten der Stichprobe. Daraus
resultiert ein ca. 8,5-facher Arbeitsaufwand für die Bereitstellung des Bildmate-
rials.
9.3 Simulation unterschiedlich bioverträglicher Materialien
Die Vorgehensweise bei der Zuordnung der Zytotoxizitätsstufen mit Hilfe der
Zellanalysesoftware soll anhand der folgenden Untersuchung exemplarisch ge-
zeigt werden. Um eine kontinuierliche Abnahme der Zytotoxizität und somit un-
terschiedliche Vitalitätsstufen der L929- und MC3T3- Zellen hervorzurufen, setzt
man die Zellen einer zuvor hergestellten Verdünnungsreihe einer toxischen
Substanz (hier Formaldehyd) aus. Das Konzentrationsintervall wird für Zellen
vom Typ L929 auf 0,0 bis 0,5 µg/ml und für Zellen des Typs MC3T3 auf 0,0 bis
1,0 µg/ml in 0,1 µg/ml Schritten festgelegt. Nach 72 Stunden Versuchsdauer
färbt eine May-Grünwald-Lösung die Zellen an. Von jeder Konzentration werden

9. Bewertung des Biokompatibilitätsgrades 137
3 Ansätze hergestellt. Die Software wertet pro Formaldehyd-Konzentration 15
mit dem Mikroskop erstellte Bilder aus und ermittelt automatisch die durch-
schnittliche Anzahl der vitalen Zellen aus der Stichprobe. Als Referenz dient die
durchschnittliche Anzahl vitaler Zellen ohne zytotoxischen Einfluss. Das Ver-
hältnis der durchschnittlichen vitalen Zellen der Stichprobe im Vergleich zu den
durchschnittlichen vitalen Zellen der Referenzprobe wird in Form der Proliferati-
onsrate in Tabelle 36 dargestellt. Da hier lediglich die Tendenz der Proliferati-
onsratenänderung bei zunehmender Toxizität überprüft wird, ist eine exakte
Bestimmung der Proliferationsrate nicht entscheidend. Eine Abweichung von -
4,7% bis 2,2% (Tabelle 44) von der realen durchschnittlichen Zellanzahl einer
Probe ist hier vertretbar, da sich dadurch die Entwicklung der Proliferationsrate
bei steigender Toxizität nicht ändert.
Tabelle 46: Formaldehyd-Verdünnungsreihe mit einer Konzentration von 0,0 µg/ml bis 0,5 µg/ml
in 0,1 µg/ml Schritten für die Linie L929 und bis 1,0 µg/ml für MC3T3 Zellen auf dem Material
Polystyrol. Abhängig von der Proliferationsrate erfolgt eine Interpretation des zytotoxischen
Zustandes der Zellen
Zelllinie Formaldehyd-konzentration
[µg/ml]
Proliferation [%]
Interpretation
L929 0,0 100 Nicht zytotoxisch
L929 0,1 87,2 Nicht zytotoxisch
L929 0,2 73,9 Schwach zytotoxisch
L929 0,3 63,8 Mäßig zytotoxisch
L929 0,4 54,5 Stark zytotoxisch
L929 0,5 41,7 Stark zytotoxisch
MC3T3 0,0 100 Nicht zytotoxisch
MC3T3 0,1 84,2 Nicht zytotoxisch
MC3T3 0,2 71,8 Schwach zytotoxisch
MC3T3 0,3 68,1 Mäßig zytotoxisch
MC3T3 0,4 65,6 Mäßig zytotoxisch
MC3T3 0,5 61,0 Mäßig zytotoxisch
MC3T3 0,6 56,5 Stark zytotoxisch
MC3T3 0,7 52,5 Stark zytotoxisch
MC3T3 0,8 47,7 Stark zytotoxisch
MC3T3 0,9 41,3 Stark zytotoxisch
MC3T3 1,0 32,7 Stark zytotoxisch
Tabelle 46 zeigt, dass mit zunehmender Formaldehyd-Konzentration die Prolife-
rationsrate für beide Zelltypen (L929 und MC3T3) sinkt, da sich die toxische
Wirkung negativ auf das Wachstumsverhalten auswirkt. Die Proliferationsrate
sinkt bis zur Formaldehyd-Konzentration von 0,2 µg/ml für beide Zelllinien ähn-
lich stark. Bei höheren Konzentrationen verringert sich die Proliferationsrate für

9. Bewertung des Biokompatibilitätsgrades 138
den Typ L929 stärker im Vergleich zur Linie MC3T3. Während bei der Linie
L929 bereits bei einer Formaldehyd-Konzentration von 0,4 µg/ml eine starke
Zytotoxizität vorliegt, ist dies für den Typ MC3T3 erst bei einer Konzentration
von 0,6 µg/ml der Fall.
Für dieses Verhalten können verschiedene Zusammenhänge verantwortlich
sein. Eine mögliche Ursache kann ein im Vergleich zum Typ L929 weiter fortge-
schrittener Adhäsionsprozess der MC3T3-Zellen sein. Des Weiteren bilden Zel-
len der Linie MC3T3 im Gegensatz zu L929-Zellen im Durchschnitt größere und
kompaktere Agglomerate aus, die mit ihrer geflechtartigen komplexen Struktur
eventuell eine höhere Resistenz gegenüber toxischen Einflüssen besitzen. Ab-
hängig von der Stoffwechselaktivität einer Zelllinie ist es ebenso möglich, dass
der Typ L929 schneller Reaktionen auf einen toxischen Einfluss zeigt als Zellen
vom Typ MC3T3.
Im nächsten Schritt wird nun geprüft, ob neben der Proliferationsrate auch die
morphologischen Zellmerkmale der vorliegenden Zelllinien eine Aussage über
den Biokompatibilitätsgrad zulassen. Tritt der Fall ein, dass sich die Morpholo-
gie der Zellen für die o.g. Konzentrationen sicher messbar unterscheiden lässt,
so ist eine Berücksichtigung der Zellmorphologie als Ergänzung zur Proliferati-
onsrate als Bewertungskriterium für die Biokompatibilität sinnvoll.
9.4 Qualitative Bewertung der Biokompatibilität
Eine Methode zur qualitativen Bewertung der Biokompatibilität ist die Morpholo-
gieanalyse der Zellen mit Hilfe eines Mikroskopes. Die Schwierigkeit bei dieser
Untersuchungsmethode liegt im Gegensatz zur quantitativen Betrachtung darin,
dass morphologische Merkmale, z.B. die Kompaktheit oder die mittlere Zellflä-
che, nicht ohne Hilfsmittel durch eine Laborfachkraft exakt bestimmt werden
können. Die qualitative Analyse der Zellmorphologie entspricht daher einer sub-
jektiven Einschätzung auf Basis von Erfahrungswerten der jeweiligen Labor-
fachkraft. Mit Hilfe der entwickelten Analysesoftware lassen sich verschiedene
morphologische Zelleigenschaften, z.B. die Fläche und Kompaktheit, bestim-
men und daraus eine genauere qualitative Bewertung der Biokompatibilität ab-
leiten.
Zunächst werden die morphologischen Zellmerkmale „Kompaktheit“ und
„durchschnittliche Einzelzellfläche“ auf ihre Eignung zur Bestimmung der Bio-
kompatibilität untersucht, da sich die Zellmorphologie mit abnehmender Biover-
träglichkeit nach (3) von flächigen über spindelförmigen bis hin zu kleinen ku-

9. Bewertung des Biokompatibilitätsgrades 139
gelförmigen Zellbereichen verändert. Hierfür finden die bereits in Kap. 9.3 auf-
geführten Zellaufnahmen der Formaldehyd-Verdünnungsreihe Verwendung.
Die Ergebnisse der morphologischen Zellanalyse für unterschiedliche Formal-
dehyd-Konzentrationen sind in Tabelle 47 aufgeführt.
Tabelle 47: Auswertung der morphologischen Merkmale für eine Formaldehyd-
Verdünnungsreihe mit einer Konzentration von 0,0 µg/ml bis 0,5 µg/ml in 0,1 µg/ml Schritten für
die Linie L929 und bis 1,0 µg/ml für MC3T3 Zellen auf dem Material Polystyrol. Bei den in der
Tabelle angegebenen Daten handelt sich jeweils um Durchschnittswerte aus 15 verschiedenen
Probenausschnitten; ∆K: Relative Abweichung zur Kompaktheit bei einer Formaldehydkonzent-
ration von 0,0 µg/ml
Zell-linie
Formaldehyd-konzentration
[µg/ml]
Mittlere Kompakt-
heit K
∆K [%]
Standard-abweichung
für K
Durch-schnittliche
Einzelzellfläche [µm
2]
L929 0,0 1,75 0,0 0,11 480
L929 0,1 1,72 -1,7 0,13 440
L929 0,2 1,78 1,7 0,14 470
L929 0,3 2,01 14,9 0,20 549
L929 0,4 1,72 -1,7 0,19 478
L929 0,5 1,83 4,6 0,32 561
MC3T3 0,0 2,71 0,0 0,17 713
MC3T3 0,1 2,81 3,7 0,23 736
MC3T3 0,2 2,94 8,5 0,22 699
MC3T3 0,3 2,98 10,0 0,21 683
MC3T3 0,4 3,01 11,1 0,25 667
MC3T3 0,5 3,01 11,1 0,26 681
MC3T3 0,6 2,98 10,0 0,27 695
MC3T3 0,7 3,09 14,0 0,26 696
MC3T3 0,8 3,19 17,7 0,28 698
MC3T3 0,9 3,26 20,3 0,26 689
MC3T3 1,0 3,33 22,9 0,25 681
Die in Tabelle 47 enthaltenen Daten für die Linie L929 weisen einen Korrelati-
onskoeffizient KC1 = 0,31 zwischen der Formaldehyd-Konzentration und der
durchschnittlichen Kompaktheit auf. Des Weiteren resultiert ein Korrelationsko-
effizient KA1 = 0,67 mit der durchschnittlichen Einzelzellfläche. Für den Zelltyp
MC3T3 hingegen resultiert ein Korrelationskoeffizient KC2 = 0,96 zwischen der
Formaldehyd-Konzentration und der durchschnittlichen Kompaktheit. Für die
durchschnittliche Einzelzellfläche und der Konzentration ergibt sich ein Korrela-
tionskoeffizient KA2 = -0,49.

9. Bewertung des Biokompatibilitätsgrades 140
9.5 Diskussion der Ergebnisse zur Bewertung des
Biokompatibilitätsgrades
Im Gegensatz zur personellen Bewertung der Zellmorphologie ist es mit Hilfe
eines computergestützten Systems möglich, qualitative Merkmale quantitativ
auszuwerten. Die Daten in Tabelle 47 zeigen, dass mit den Merkmalen „durch-
schnittliche Einzelzellfläche“ und „Kompaktheit“ für die Linie L929 keine sichere
Bewertung der Biokompatibilität durchführbar ist. Es besteht keine ausreichen-
de Korrelation zw. der Formaldehyd-Konzentration und den Merkmalswerten im
Rahmen des vorgegebenen Konzentrationsintervalls (0 bis 0,5µg/ml). Die
durchschnittliche Einzelzellfläche des Zelltyps MC3T3 ist, analog zur Linie
L929, aufgrund einer unzureichenden Korrelation zur Formaldehyd-
Konzentration nicht als morphologisches Merkmal zur Bewertung der Biokom-
patibilität geeignet. Im Gegensatz dazu liegt mit KC2 = 0,96 eine gute Korrelation
zwischen der Konzentration und der durchschnittlichen Kompaktheit der MC3T3
Zellen vor. Jedoch ist die Änderung der durchschnittlichen Kompaktheit von
einer Konzentrationsstufe zur nächsten Stufe gering und im Vergleich dazu die
vorliegende Standardabweichung der durchschnittlichen Merkmalswerte für ei-
ne bestimmte Konzentrationsstufe hoch (Tabelle 47). So ist eine sichere Be-
stimmung der Zytotoxizitätsstufe auf Basis des Merkmals „Kompaktheit“ für die
Linie MC3T3 nicht gewährleistet.
Aus Tabelle 47 geht weiterhin hervor, dass für die Zelltypen L929 und MC3T3
keine wesentliche Änderung der Zellfläche bei der vorliegenden Formaldehyd-
Konzentrationsreihe vorliegt. Diese Beobachtung entspricht nicht den in (3) be-
schriebenen morphologischen Änderungen der Zellen von flächig über spindel-
förmig bis hin zu kugelig bei Zunahme der Biounverträglichkeit. Der Grund liegt
darin, dass keine Korrelation zwischen der in der Literatur angegebenen Mor-
phologieänderung bei toxischem Einfluss und der in der Norm hinterlegten Ein-
stufung der Proliferationsrate in die verschiedenen Zytotoxizitätsstufen gegeben
ist. Abb. 80 stellt den Zusammenhang zwischen der Morphologieänderung und
der Änderung der Proliferationsrate bei verschiedenen Formaldehyd-
Konzentrationsstufen dar.
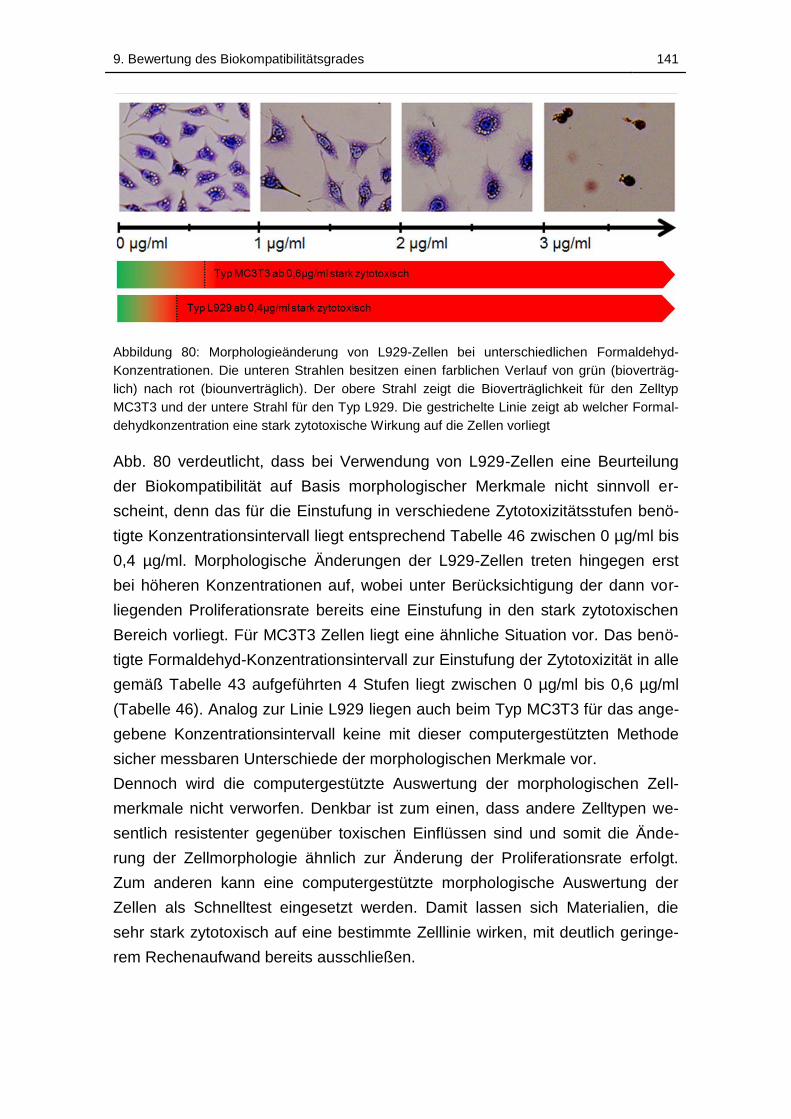
9. Bewertung des Biokompatibilitätsgrades 141
Abbildung 80: Morphologieänderung von L929-Zellen bei unterschiedlichen Formaldehyd-
Konzentrationen. Die unteren Strahlen besitzen einen farblichen Verlauf von grün (bioverträg-
lich) nach rot (biounverträglich). Der obere Strahl zeigt die Bioverträglichkeit für den Zelltyp
MC3T3 und der untere Strahl für den Typ L929. Die gestrichelte Linie zeigt ab welcher Formal-
dehydkonzentration eine stark zytotoxische Wirkung auf die Zellen vorliegt
Abb. 80 verdeutlicht, dass bei Verwendung von L929-Zellen eine Beurteilung
der Biokompatibilität auf Basis morphologischer Merkmale nicht sinnvoll er-
scheint, denn das für die Einstufung in verschiedene Zytotoxizitätsstufen benö-
tigte Konzentrationsintervall liegt entsprechend Tabelle 46 zwischen 0 µg/ml bis
0,4 µg/ml. Morphologische Änderungen der L929-Zellen treten hingegen erst
bei höheren Konzentrationen auf, wobei unter Berücksichtigung der dann vor-
liegenden Proliferationsrate bereits eine Einstufung in den stark zytotoxischen
Bereich vorliegt. Für MC3T3 Zellen liegt eine ähnliche Situation vor. Das benö-
tigte Formaldehyd-Konzentrationsintervall zur Einstufung der Zytotoxizität in alle
gemäß Tabelle 43 aufgeführten 4 Stufen liegt zwischen 0 µg/ml bis 0,6 µg/ml
(Tabelle 46). Analog zur Linie L929 liegen auch beim Typ MC3T3 für das ange-
gebene Konzentrationsintervall keine mit dieser computergestützten Methode
sicher messbaren Unterschiede der morphologischen Merkmale vor.
Dennoch wird die computergestützte Auswertung der morphologischen Zell-
merkmale nicht verworfen. Denkbar ist zum einen, dass andere Zelltypen we-
sentlich resistenter gegenüber toxischen Einflüssen sind und somit die Ände-
rung der Zellmorphologie ähnlich zur Änderung der Proliferationsrate erfolgt.
Zum anderen kann eine computergestützte morphologische Auswertung der
Zellen als Schnelltest eingesetzt werden. Damit lassen sich Materialien, die
sehr stark zytotoxisch auf eine bestimmte Zelllinie wirken, mit deutlich geringe-
rem Rechenaufwand bereits ausschließen.
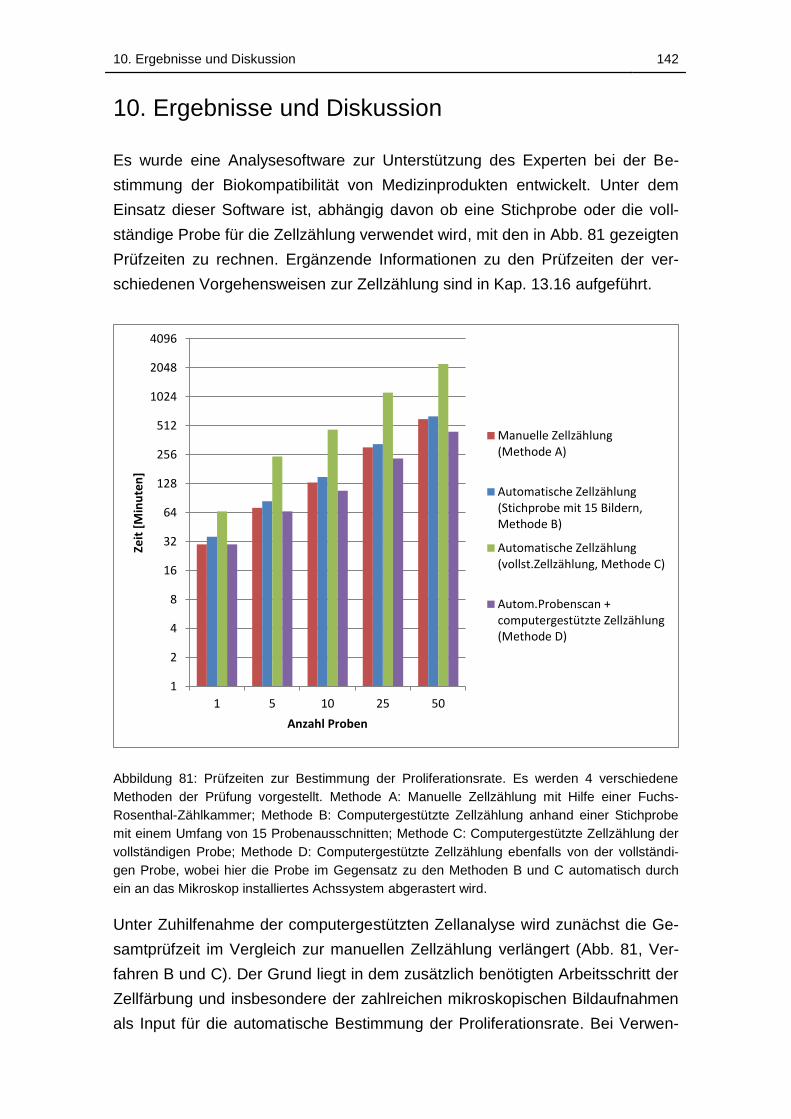
10. Ergebnisse und Diskussion 142
10. Ergebnisse und Diskussion
Es wurde eine Analysesoftware zur Unterstützung des Experten bei der Be-
stimmung der Biokompatibilität von Medizinprodukten entwickelt. Unter dem
Einsatz dieser Software ist, abhängig davon ob eine Stichprobe oder die voll-
ständige Probe für die Zellzählung verwendet wird, mit den in Abb. 81 gezeigten
Prüfzeiten zu rechnen. Ergänzende Informationen zu den Prüfzeiten der ver-
schiedenen Vorgehensweisen zur Zellzählung sind in Kap. 13.16 aufgeführt.
Abbildung 81: Prüfzeiten zur Bestimmung der Proliferationsrate. Es werden 4 verschiedene
Methoden der Prüfung vorgestellt. Methode A: Manuelle Zellzählung mit Hilfe einer Fuchs-
Rosenthal-Zählkammer; Methode B: Computergestützte Zellzählung anhand einer Stichprobe
mit einem Umfang von 15 Probenausschnitten; Methode C: Computergestützte Zellzählung der
vollständigen Probe; Methode D: Computergestützte Zellzählung ebenfalls von der vollständi-
gen Probe, wobei hier die Probe im Gegensatz zu den Methoden B und C automatisch durch
ein an das Mikroskop installiertes Achssystem abgerastert wird.
Unter Zuhilfenahme der computergestützten Zellanalyse wird zunächst die Ge-
samtprüfzeit im Vergleich zur manuellen Zellzählung verlängert (Abb. 81, Ver-
fahren B und C). Der Grund liegt in dem zusätzlich benötigten Arbeitsschritt der
Zellfärbung und insbesondere der zahlreichen mikroskopischen Bildaufnahmen
als Input für die automatische Bestimmung der Proliferationsrate. Bei Verwen-
1
2
4
8
16
32
64
128
256
512
1024
2048
4096
1 5 10 25 50
Zeit
[M
inu
ten
]
Anzahl Proben
Manuelle Zellzählung(Methode A)
Automatische Zellzählung(Stichprobe mit 15 Bildern,Methode B)
Automatische Zellzählung(vollst.Zellzählung, Methode C)
Autom.Probenscan +computergestützte Zellzählung(Methode D)

10. Ergebnisse und Diskussion 143
dung einer Stichprobe von 15 Probenausschnitten (Abb. 81, Verfahren B) ist mit
einer Gesamtprüfzeit von 1,4h für 5 Proben zu rechnen, für die vollständige
Zellzählung von 5 Proben sogar mit 4,1h. Im Gegensatz dazu erfolgt die manu-
elle Zellzählung mittels Zählkammer (Abb. 81, Verfahren A) in 1,2h. Dabei wird
je Probe 10-mal manuell ausgezählt und schließlich der Mittelwert gebildet.
Im Rahmen dieser Arbeit stand kein ansteuerbares Achssystem für das Mikro-
skop zur Verfügung. Jedoch könnte mit Einsatz eines Achssystems zur automa-
tischen Aufnahme der Probenoberfläche die Prüfzeit deutlich reduziert werden
(Abb. 81, Verfahren D). Lässt man die für die Zellanalyse benötigte Rechenzeit
des Computers unberücksichtigt, so verringert sich die Arbeitszeit für z.B. 5
Proben von 4,1h auf ca. 1,1h und liegt damit im Bereich der benötigten Zeit im
Rahmen der manuellen Zellzählung. Bei steigender Anzahl der zu analysieren-
den Proben erhöht sich die Zeitersparnis unter Verwendung der automatischen
Zellzählung, vorausgesetzt die mikroskopischen Bildaufnahmen werden auto-
matisch aufgenommen. Für die Analyse von z.B. 50 Proben würde demnach die
benötigte Arbeitszeit von 10h bei manueller Zellzählung auf 7,4h für die compu-
tergestützte Zählung sinken.
Die Zeit zur Bestimmung der Proliferationsrate hängt stark von der Erfahrung
der Laborfachkraft ab und kann daher um mehrere Minuten variieren. Hingegen
erfolgt die computergestützte Analyse bei jedem Auswertevorgang in nahezu
gleicher Zeit. Rechenzeitunterschiede im Sekundenbereich ergeben sich ledig-
lich durch die sich ändernden Laufzeiten der Separationsalgorithmen aufgrund
der variablen Zellmorphologie.
Unabhängig davon, ob eine manuelle Zählung von Zellen in Suspension oder
auf der Probenoberfläche vorgenommen wird, muss zu Beginn der Biokompati-
bilitätsprüfung eine bestimmte Ausgangspopulation auf das zu untersuchende
Material gebracht werden. Ideal wäre, wenn für verschiedene Proben tatsäch-
lich die gleiche Ausgangspopulation bereitgestellt werden kann. Untersuchun-
gen zeigen jedoch (Kap 13.15), dass die gleiche Ausgangspopulation für ver-
schiedene Proben nicht gewährleistet werden kann. Abbildungen 82 und 83
stellen die ermittelten Daten grafisch dar.
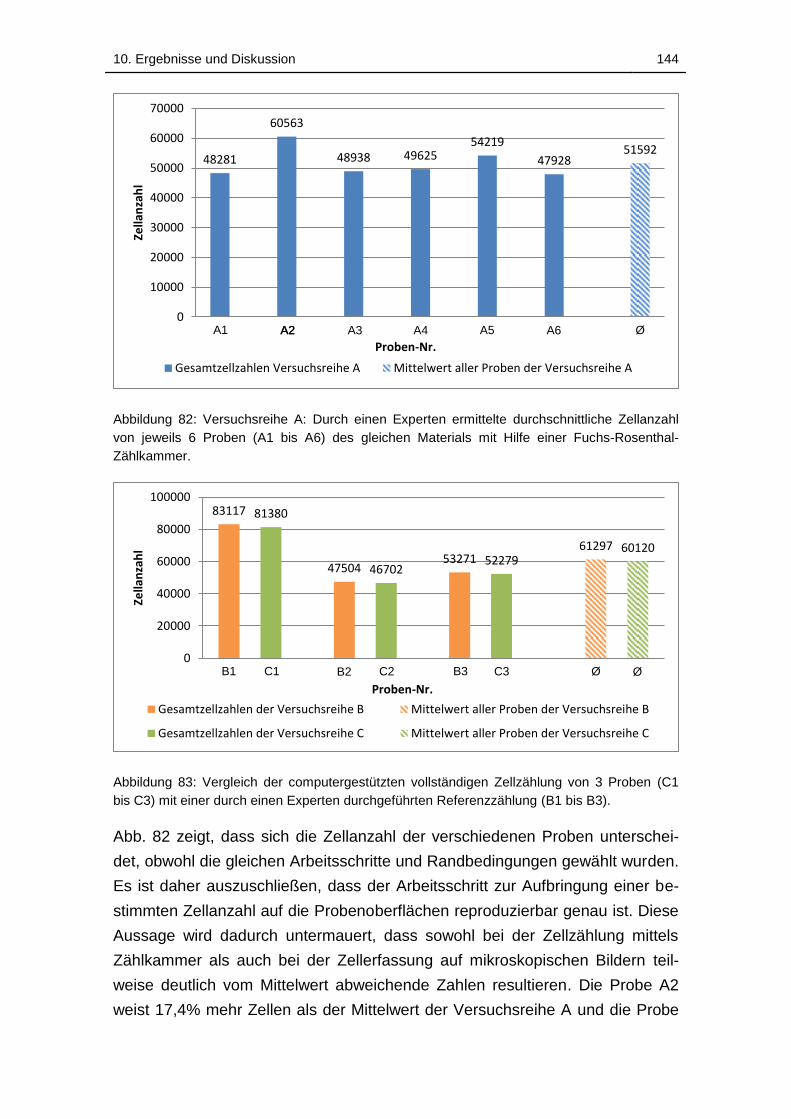
10. Ergebnisse und Diskussion 144
Abbildung 82: Versuchsreihe A: Durch einen Experten ermittelte durchschnittliche Zellanzahl
von jeweils 6 Proben (A1 bis A6) des gleichen Materials mit Hilfe einer Fuchs-Rosenthal-
Zählkammer.
Abbildung 83: Vergleich der computergestützten vollständigen Zellzählung von 3 Proben (C1
bis C3) mit einer durch einen Experten durchgeführten Referenzzählung (B1 bis B3).
Abb. 82 zeigt, dass sich die Zellanzahl der verschiedenen Proben unterschei-
det, obwohl die gleichen Arbeitsschritte und Randbedingungen gewählt wurden.
Es ist daher auszuschließen, dass der Arbeitsschritt zur Aufbringung einer be-
stimmten Zellanzahl auf die Probenoberflächen reproduzierbar genau ist. Diese
Aussage wird dadurch untermauert, dass sowohl bei der Zellzählung mittels
Zählkammer als auch bei der Zellerfassung auf mikroskopischen Bildern teil-
weise deutlich vom Mittelwert abweichende Zahlen resultieren. Die Probe A2
weist 17,4% mehr Zellen als der Mittelwert der Versuchsreihe A und die Probe
48281
60563
48938 49625 54219
47928 51592
0
10000
20000
30000
40000
50000
60000
70000
Zella
nza
hl
Proben-Nr.
Gesamtzellzahlen Versuchsreihe A Mittelwert aller Proben der Versuchsreihe A
83117
47504 53271
61297
81380
46702 52279
60120
0
20000
40000
60000
80000
100000
Zella
nza
hl
Proben-Nr.
Gesamtzellzahlen der Versuchsreihe B Mittelwert aller Proben der Versuchsreihe B
Gesamtzellzahlen der Versuchsreihe C Mittelwert aller Proben der Versuchsreihe C
A1 A2 A5 A4 A3 Ø A6
B1 B3 B2 C1 C2 C3 Ø Ø
A2

10. Ergebnisse und Diskussion 145
B1 35,6% mehr Zellen als der Mittelwert der Versuchsreihe B auf. Des Weiteren
liegen die Standardabweichungen zu den Zellzahlen der Proben A1 bis A6 in
der gleichen Größenordnung (Kap. 13.15, Tabelle 79), so dass man hier auf
einen reproduzierbaren Arbeitsablauf bei der Bestimmung der Zellanzahl
schließen kann.
Aufgrund der variierenden Zellanzahl ist die Untersuchung von nur einer Probe
zur Bestimmung des Biokompatibilitätsgrades nicht empfehlenswert. Sinnvoll ist
hingegen die Untersuchung von mehreren Proben des gleichen Materials, um
die resultierenden Ergebnisse einer statistischen Auswertung zu unterziehen
und ein sicheres Gesamtergebnis zu gewährleisten. Denkbar ist hier der Aus-
schluss von Ausreißern, z.B. mit Hilfe des Grubbs-Tests (84), und die nachfol-
gende Neuberechnung des Mittelwertes der Zellanzahl. Führt man den Grubbs-
Test für die Probenserien A, B und C durch, so stellen sich die Proben A2, B1
und C1 als Ausreißer heraus (Kap. 13.17).
Abbildung 84: Links: Durchschnittliche Zellanzahl der Versuchsreihen A-C vor Ausschluß der
Ausreißer; Rechts: Durchschnittliche Zellanzahl der Versuchsreihen A-C nach Ausschluss der
mit Hilfe des Grubbs-Tests ermittelten Ausreißer
Ohne Berücksichtigung der ermittelten Ausreißer resultiert ein neuer Mittelwert
für die Versuchsreihe A von 49798, für die Versuchsreihe B von 50388 und für
die Versuchsreihe C von 49491. Nach dem Ausschluss der Ausreißer liegen
nun für die unterschiedlichen Methoden zur Gewinnung der Zellanzahl ähnliche
Werte vor (Abb. 84).
51592
61297 60120
49798 50388 49491
0
10000
20000
30000
40000
50000
60000
70000
Zella
nza
hl
Durchschnittliche Zellanzahl der Versuchsreihen A B C vor Ausschluss der Ausreißer
Durchschnittliche Zellanzahl der Versuchsreihen A B C nach Ausschluss der Ausreißer

10. Ergebnisse und Diskussion 146
Abb. 83 zeigt weiterhin die Ergebnisse der automatischen Zellzahlbestimmung
(Versuchsreihe C) für dieselben Proben, die bereits in der Versuchsreihe B oh-
ne computergestütztes System gezählt wurden. Für die Probe C1 resultiert eine
Abweichung von 2,1% zur Probe B1, für C2 eine Abweichung von 1,7% zu B2
und für C3 eine Abweichung von 1,9% zu B3. Es fällt auf, dass mit steigender
Zellanzahl auf der Probenoberfläche die Abweichung zu den jeweiligen Mess-
werten aus Versuchsreihe B zunimmt. Der Grund liegt darin, dass bei zuneh-
mender Zelldichte im Bild die Separation der zusammenhängenden Zellen er-
schwert wird und folglich nicht alle Zellen erfasst werden.
Die hier genannten Abweichungen von 1,7% bis 2,1% sind leicht höher als die
in Kap. 6.5.2 genannten 1,31% für die Linie L929. Der Grund liegt darin, dass,
im Gegensatz zu den untersuchten Bildaufnahmen mit 100-facher Vergröße-
rung in Kap. 6.5.2, hier mit einer Vergrößerung von Faktor 50 das Bildmaterial
aufgenommen wurde. Bei einer Vergrößerung von Faktor 50 sind bereits 128
Aufnahmen für die vollständige Erfassung der Probenoberfläche notwendig,
deshalb wurde hier die Vergrößerung reduziert. Eine kleinere Vergrößerung der
Zellen stellt für die automatische Zellanalyse jedoch eine größere Herausforde-
rung dar, da pro Zelle weniger Bildpixel zur Verfügung stehen. Tauscht man die
bestehende Kamera durch eine entsprechend höher auflösende Kamera aus,
würden sich auch bei einer Vergrößerung von Faktor 50 ähnliche Abweichun-
gen wie in Kap. 6.5.2 einstellen.
Für die Bestimmung der Genauigkeit der in Kap. 6 vorgestellten Separations-
verfahren sind zunächst jeweils für 100 Probenausschnitte die mittlere Abwei-
chung zur Referenz ermittelt worden. Um eine Aussage über die statistische
Sicherheit der angegebenen Abweichung zu erhalten, sind für vier verschiede-
ne Konfidenzintervalle die Toleranzen um die mittlere Abweichung berechnet
worden. Es ist zu erwarten, dass mit der jeweils angenommenen Wahrschein-
lichkeit die mittlere Abweichung für 100 Probenausschnitte im jeweils angege-
benen Toleranzbereich liegt (Tabelle 48).
Tabelle 48: Übersicht der statistischen Daten zur Genauigkeit der Zellzählung der verschiede-
nen Separationsverfahren
Ver-fahren
Zell-typ
Mittlere Abweichung
zur Referenz
[%]
Standard- Abweichung
[%]
Standard- fehler
[%]
Konfidenz- Intervall
Toleranz um
Abweichung [%]
Methode aus Kap 6.4 mit CBS
L929 1,27 0,87 0,09
95% 1,27 +/- 0,17
99% 1,27 +/- 0,22
99,9% 1,27 +/- 0,29
99,99% 1,27+/- 0,34

10. Ergebnisse und Diskussion 147
Methode aus Kap.
6.5 L929 1,31 0,96 0,10
95% 1,31 +/- 0,19
99% 1,31 +/- 0,25
99,9% 1,31 +/- 0,31
99,99% 1,31 +/- 0,37
Methode aus Kap.
6.6 L929 3,13 1,37 0,14
95% 3,13 +/- 0,27
99% 3,13 +/- 0,35
99,9% 3,13 +/- 0,45
99,99% 3,13 +/- 0,53
Methode aus Kap.
6.5 MC3T3 3,50 2,36 0,24
95% 3,50 +/- 0,46
99% 3,50 +/- 0,61
99,9% 3,50 +/- 0,78
99,99% 3,50 +/- 0,92
Für einen schnellen visuellen Vergleich der in Tabelle 47 hinterlegten Daten
dient die folgende Abb. 85.
Abbildung 85: Visualisierung der in Tabelle 48 hinterlegten statistischen Daten zur Zellzählung
der verschiedenen Separationsverfahren. Auf der x-Achse sind jeweils für die 4 im Vergleich
stehenden Verfahren die Konfidenzintervalle 95%, 99%, 99,9% und 99,99% aufgetragen. Der y-
Wert repräsentiert die Abweichung zur Referenzzählung. Die farbige Markierung innerhalb der
jeweiligen Toleranzbereiche repräsentiert den Mittelwert.
Abb. 85 visualisiert die in Tabelle 48 hinterlegten Daten. Das Bild ist in 4 Ab-
schnitte unterteilt wobei jeder Abschnitt für ein zu vergleichendes Separations-
verfahren steht (Spalte 1 in Tabelle 48). Jeder Abschnitt enthält die Intervall-
grenzen, jeweils für das 95%-, 99%-, 99,9%- und 99,99%-Konfidenzniveau. Die
graue Markierung innerhalb der Intervalle repräsentiert den Mittelwert der Ab-
weichung zur Referenz. Anhand der Grafik wird deutlich, dass die Abweichun-
gen für die Verfahren, beschrieben in Kap. 6.4 mit vorgeschalteter CBS und
Kap. 6.5 für den Zelltyp L929 die geringsten Abweichungen zur Referenz sowie
die kleinsten Konfidenzintervalle aufweisen. Die Auswahl des passenden Kon-
fidenzintervalls hängt von der Problemstellung ab und muss im Einzelfall festge-
legt werden.

10. Ergebnisse und Diskussion 148
Welches Konfidenzintervall ist für die vorliegende Aufgabenstellung geeignet?
Bei der vorliegenden Anwendung ist es wichtig, dass ein Implantatmaterial nicht
besser biokompatibel eingestuft wird als es tatsächlich der Fall ist. Im schlimms-
ten Fall können Abstoßungsreaktionen des Körpers die Folge sein.
Aufgrund der variierenden Startpopulation der Zellen auf dem Proben- sowie
Referenzmaterial (Abbildung 82) wird ohnehin eine Mittelwertbildung der Refe-
renz-Zellzählung und Test-Zellzählung für 10 Proben durchgeführt so dass
durch diese Vorgehensweise die Verwendung des 99%-Konfidenzintervalls für
jede Probe ausreichend erscheint.
Es ist zu erwarten, dass sich mit fortschreitender Arbeitszeit die Fehlerwahr-
scheinlichkeit einer Laborfachkraft, hervorgerufen durch Konzentrationsverlust,
erhöht. Dies führt zu Fehlern bei der Bestimmung der Zellanzahl und Proliferati-
onsrate. Darüber hinaus kann nicht garantiert werden, dass mehrere La-
borfachkräfte für denselben Probenausschnitt die gleiche morphologische Be-
wertung vornehmen. Aus diesen Fehlerquellen können unterschiedliche Bewer-
tungsergebnisse für die Biokompatibilität eines Materials resultieren. Auch mit
den vorliegenden computergestützten Verfahren ist aufgrund der Komplexität
der Zellmorphologie keine fehlerfreie automatische Zellanalyse möglich. Durch
die automatisierte und damit reproduzierbare Anwendung der Algorithmen ist
jedoch im Gegensatz zur personellen Analyse ein objektives Ergebnis gewähr-
leistet.
Ist die Laborfachkraft mit dem automatisch generierten Segmentierungs- oder
Separationsergebnis der Zellen unzufrieden, kann sie bei Bedarf interaktiv das
Ergebnis modifizieren und die Berechnung des Biokompatibilitätsgrades erneut
durchführen lassen.
Aufgrund der Ergebnisse der Untersuchungen zu quantitativen und qualitativen
Zellmerkmalen (Kap. 9) wird für die Bewertung der Biokompatibilität mit Hilfe
der Zelllinien L929 und MC3T3 ausschließlich die Änderung der Proliferations-
rate im Vergleich zu einer Referenzprobe herangezogen und auf eine automati-
sche Beurteilung der morphologischen Zellmerkmale aus den in Kap. 9.5 ge-
nannten Gründen verzichtet. Mit Hilfe der Software ist jedoch eine automatische
Morphologieanalyse von Zellen möglich. Diese Funktion steht daher z.B. für
andere Zelllinien zur Verfügung und beansprucht nur unwesentlich mehr Re-
chenzeit.

10. Ergebnisse und Diskussion 149
10.1 Generalisierungsleistung der Software
Generalisierungsleistung der vorgestellten Segmentierungsmethoden
Die Form, Größe und Art der Objekte ist für die Detektion mit Hilfe des in Kap.
5.2.1 vorgestellten Segmentierungsverfahrens unerheblich, lediglich der Intensi-
tätswert der Pixel ist für die Erfassung von Bedeutung. Daher handelt es sich
hier um ein Segmentierungsverfahren dass für alle Objekte einsetzbar ist, die
einen Kontrastunterschied zum Hintergrund aufweisen. Voraussetzung für eine
ordentliche Segmentierung ist eine reproduzierbare Helligkeit von Bild zu Bild,
andernfalls ist eine Anpassung der Segmentierungsschwellen erforderlich.
Die histogrammbasierte Segmentierungsmethode, beschrieben in Kap. 5.2.2,
kann zur Detektion beliebiger Objekte im Bild angewandt werden. Auf Basis des
Bildhistogramms erfolgt die Bestimmung der Schwellwerte automatisch, so
dass eine manuelle Parametereingabe entfällt. Voraussetzung für eine erfolg-
reiche Segmentierung ist ein ausreichender Kontrastunterschied zwischen Vor-
der- und Hintergrund des Bildes. Vorranging findet dieses Verfahren dann An-
wendung wenn der Hintergrundbereich eines Bildes relativ homogen erscheint.
Dies ist z.B. meist bei Durchlichtaufnahmen der Fall.
Der Histogram Backprojection Algorithmus kann ebenfalls für die Detektion be-
liebiger Objekte im Bild angewendet werden sofern sie sich farblich von dem
Hintergrund unterscheiden. Treten Objekte mit unterschiedlicher Farbgebung
auf so können mit Hilfe mehrerer Farbtemplates alle Objekte in einem Bild
segmentiert werden. Das Verfahren kommt an seine Grenzen wenn eine deutli-
che Farbvariation eines Objektes von Bild zu Bild auftritt so dass Objektfarben
teilweise nicht im Template abgebildet sind. In diesem Fall ist eine Segmentie-
rung der entsprechenden Objektbereiche nicht möglich und das Farbtemplate
muss angepasst werden.
Generalisierungsleistung der vorgestellten Separationsmethoden:
Die CBS-Methode, beschrieben in Kap. 6.1, ist anwendbar auf alle Objekte die
sich zu Objektclustern zusammengeschlossen haben bzw. optisch der Erschei-
nung eines zusammengewachsenen Bereiches entsprechen und daher mit Hilfe
von Segmentierungsverfahren als ein Objektbereich detektiert werden. Diese
Methode benötigt für die Separation der Objekte Einschnürungen in der Kontur
des Objektclusters. Dies ist bei vielen Objekttypen der Fall, z.B. Partikel oder
verschiedene Zelllinien.
Mit Hilfe der einstellbaren Kreismaskendurchmesser C1 und C2 ist der Algorith-

10. Ergebnisse und Diskussion 150
mus in der Lage unterschiedlich große Objektcluster zu trennen. Eine iterative
Ausführung der CBS-Methode mit variierenden Kreismaskendurchmessern er-
möglicht die Trennung von unterschiedlich großen Objekten innerhalb eines
Bildes. Die CBS-Methode findet keine Anwendung für Objekttypen, dessen
Cluster keine Einschnürungen an den Objektkontaktstellen aufweisen, z.B. für
den Fall des Zelltyps MC3T3.
Das Separationsverfahren, beschrieben in Kap. 6.4, ist für alle Zelltypen an-
wendbar, die Einschnürungen an den Zellkontaktstellen aufweisen und deren
Kernbereiche so angefärbt sind, dass sie einen Kontrastunterschied zur Umge-
bung aufweisen. Dies ist z.B. für Zellen der Typen L929, L132 und HEp-2 der
Fall. Der Algorithmus ist hingegen nicht anwendbar auf Zellen die meistens
kompakte Cluster ohne Einschnürungen an den Zellkontaktstellen aufweisen,
z.B. beim Typ MC3T3.
Die Wachstumssimulation, beschrieben in Kap. 6.5, ist für die Separation aller
Objekte geeignet, die ein Objektmerkmal aufweisen das mit der Fläche des ge-
samten Objektes korreliert. Für diese Methode ist es, im Gegensatz zum CBS-
Verfahren oder dem Algorithmus aus Kap. 6.4, unerheblich ob die Objektcluster
Einschnürungen an den Kontaktstellen aufweisen oder nicht. Ein geeignetes
Objektmerkmal im Bereich der Zellanalyse ist die Kernfläche. Eine Korrelation
der Kernflächen zur gesamten Zellfläche wurde für die Zelltypen L929, MC3T3,
L132 und BHK-21 nachgewiesen. Es ist davon auszugehen, dass weitere Zell-
typen eine Korrelation zwischen Kern- und Zellfläche aufweisen, so dass das
Verfahren für viele weitere Zelltypen einsetzbar ist. Voraussetzung für eine er-
folgreiche Separation von zusammenhängenden Zellen ist die Detektion der
Kernbereiche, auf deren Basis die Konturen der Zellen innerhalb eines Clusters
berechnet werden. Dieses Verfahren ist ungeeignet für Objekte deren Objekt-
merkmal in keinem Zusammenhang mit der Objektgröße steht.
Die im Rahmen dieser Arbeit entwickelte Software lässt sich auch für andere
Färbemethoden und Anwendungen einsetzen. So kann neben der histologi-
schen May-Grünwald- oder der kristallviolett- ebenso eine Fluoreszenzfärbung
zur nachfolgenden Zellanalyse zum Einsatz kommen (Abb. 87 und Abb. 88).
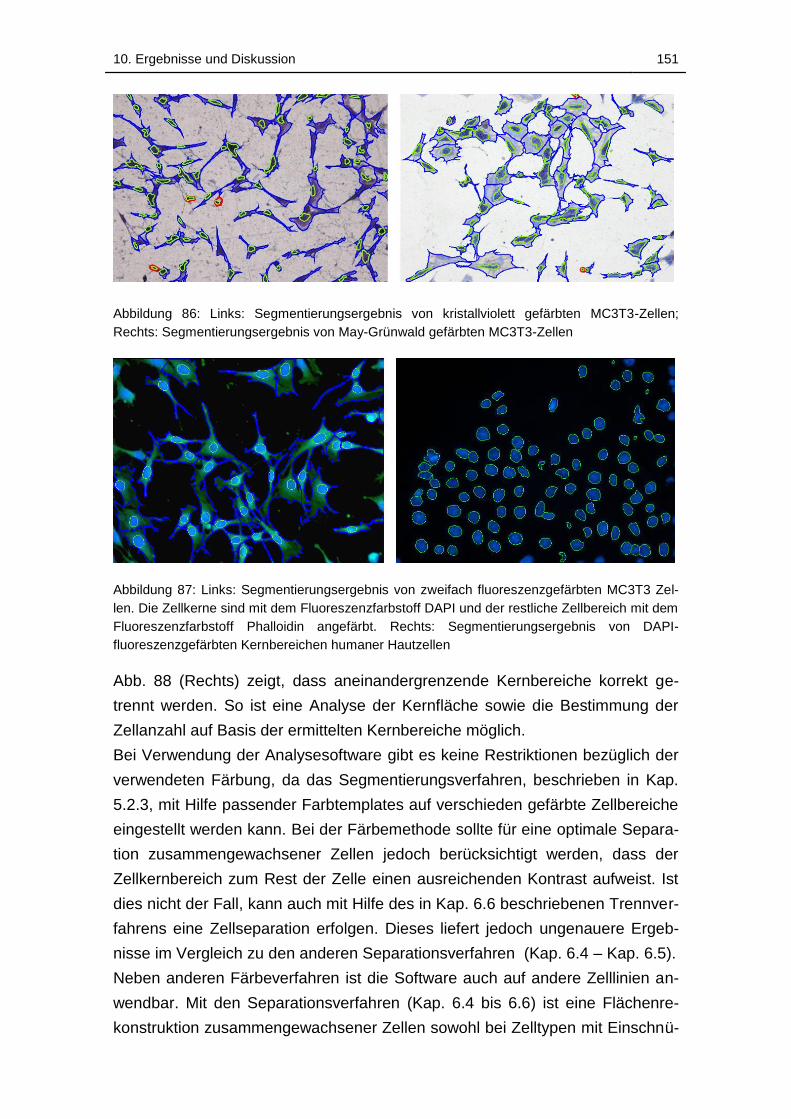
10. Ergebnisse und Diskussion 151
Abbildung 86: Links: Segmentierungsergebnis von kristallviolett gefärbten MC3T3-Zellen;
Rechts: Segmentierungsergebnis von May-Grünwald gefärbten MC3T3-Zellen
Abbildung 87: Links: Segmentierungsergebnis von zweifach fluoreszenzgefärbten MC3T3 Zel-
len. Die Zellkerne sind mit dem Fluoreszenzfarbstoff DAPI und der restliche Zellbereich mit dem
Fluoreszenzfarbstoff Phalloidin angefärbt. Rechts: Segmentierungsergebnis von DAPI-
fluoreszenzgefärbten Kernbereichen humaner Hautzellen
Abb. 88 (Rechts) zeigt, dass aneinandergrenzende Kernbereiche korrekt ge-
trennt werden. So ist eine Analyse der Kernfläche sowie die Bestimmung der
Zellanzahl auf Basis der ermittelten Kernbereiche möglich.
Bei Verwendung der Analysesoftware gibt es keine Restriktionen bezüglich der
verwendeten Färbung, da das Segmentierungsverfahren, beschrieben in Kap.
5.2.3, mit Hilfe passender Farbtemplates auf verschieden gefärbte Zellbereiche
eingestellt werden kann. Bei der Färbemethode sollte für eine optimale Separa-
tion zusammengewachsener Zellen jedoch berücksichtigt werden, dass der
Zellkernbereich zum Rest der Zelle einen ausreichenden Kontrast aufweist. Ist
dies nicht der Fall, kann auch mit Hilfe des in Kap. 6.6 beschriebenen Trennver-
fahrens eine Zellseparation erfolgen. Dieses liefert jedoch ungenauere Ergeb-
nisse im Vergleich zu den anderen Separationsverfahren (Kap. 6.4 – Kap. 6.5).
Neben anderen Färbeverfahren ist die Software auch auf andere Zelllinien an-
wendbar. Mit den Separationsverfahren (Kap. 6.4 bis 6.6) ist eine Flächenre-
konstruktion zusammengewachsener Zellen sowohl bei Zelltypen mit Einschnü-
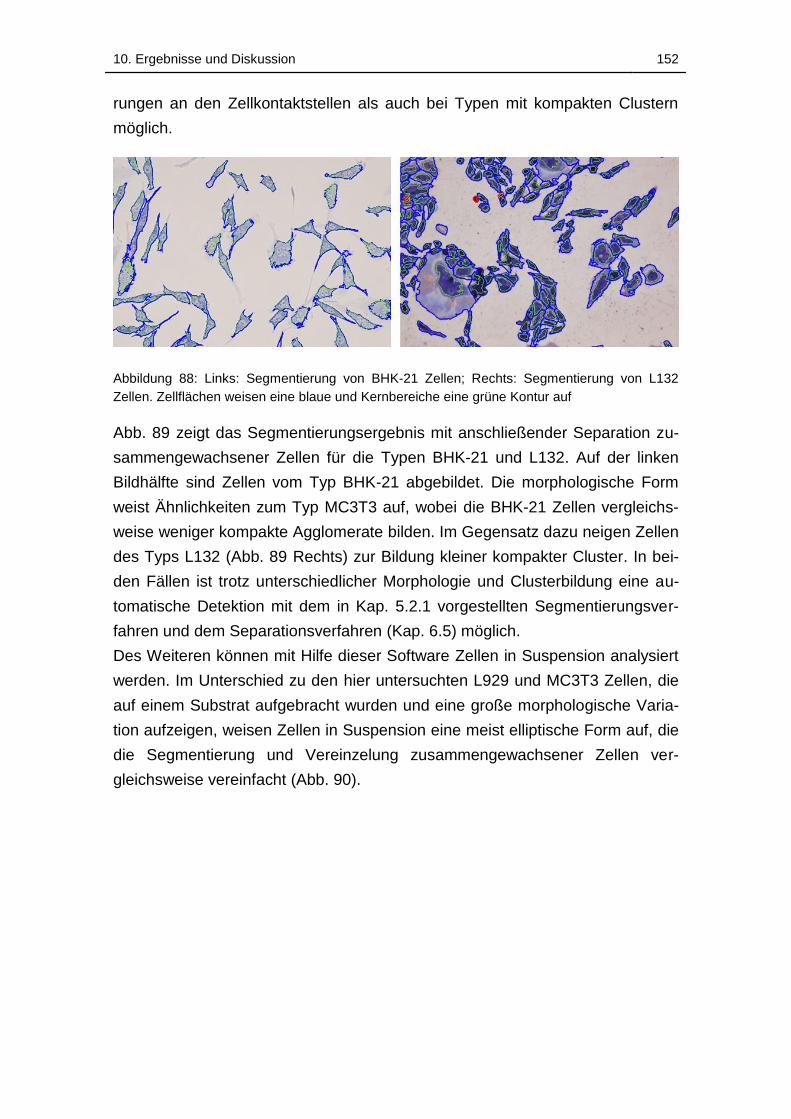
10. Ergebnisse und Diskussion 152
rungen an den Zellkontaktstellen als auch bei Typen mit kompakten Clustern
möglich.
Abbildung 88: Links: Segmentierung von BHK-21 Zellen; Rechts: Segmentierung von L132
Zellen. Zellflächen weisen eine blaue und Kernbereiche eine grüne Kontur auf
Abb. 89 zeigt das Segmentierungsergebnis mit anschließender Separation zu-
sammengewachsener Zellen für die Typen BHK-21 und L132. Auf der linken
Bildhälfte sind Zellen vom Typ BHK-21 abgebildet. Die morphologische Form
weist Ähnlichkeiten zum Typ MC3T3 auf, wobei die BHK-21 Zellen vergleichs-
weise weniger kompakte Agglomerate bilden. Im Gegensatz dazu neigen Zellen
des Typs L132 (Abb. 89 Rechts) zur Bildung kleiner kompakter Cluster. In bei-
den Fällen ist trotz unterschiedlicher Morphologie und Clusterbildung eine au-
tomatische Detektion mit dem in Kap. 5.2.1 vorgestellten Segmentierungsver-
fahren und dem Separationsverfahren (Kap. 6.5) möglich.
Des Weiteren können mit Hilfe dieser Software Zellen in Suspension analysiert
werden. Im Unterschied zu den hier untersuchten L929 und MC3T3 Zellen, die
auf einem Substrat aufgebracht wurden und eine große morphologische Varia-
tion aufzeigen, weisen Zellen in Suspension eine meist elliptische Form auf, die
die Segmentierung und Vereinzelung zusammengewachsener Zellen ver-
gleichsweise vereinfacht (Abb. 90).

10. Ergebnisse und Diskussion 153
Abbildung 89: Segmentierung von in Suspension befindlichen L929 Zellen. Die Suspension
befindet sich in einer Zählkammer, daher ist das Raster im Bild sichtbar.
Abb. 90 zeigt die Segmentierung von in Suspension befindlichen Zellen. Zu-
sammenhängende Zellen werden mit Hilfe des in Kap. 6.5 beschriebenen Se-
parationsverfahrens erfolgreich getrennt, so dass eine korrekte Zellzahlbestim-
mung auch in Suspension möglich ist.
Die Software lässt sich neben der Analyse von Zellen auch auf völlig andere
Aufgabenstellungen anwenden, z.B. zur Vermessung verschiedener Objekte.
Dies ist möglich, da die verwendeten Segmentierungsverfahren (Kap. 5.2) un-
abhängig von der Morphologie auf verschiedene Objekte angewendet und mit
Hilfe einer Kalibrierfunktion Pixel in das Längenmaß Meter umgerechnet wer-
den können. Für eine Kalibrierung wird ein Objekt benötigt, von dem das Maß in
der Basiseinheit Meter bekannt ist, z.B. ein Endmaß oder eine Schieblehre
(Abb. 91). Ist das System kalibriert, kann der Abstand zwischen zwei Punkten
durch interaktives Einzeichnen einer Geraden in das Bildfenster bestimmt wer-
den, unabhängig von der Art der vorliegenden Objekte.

10. Ergebnisse und Diskussion 154
Abbildung 90: Links: Kalibrierung der Software mit Hilfe eines Referenzobjektes, hier der bei
10mm eingestellte Messschieber. Rechts: Vermessung eines 30mm Endmaßes
Abb. 91 (Links) zeigt die Kalibrierung der Software mit Hilfe einer auf 10mm
eingestellten Schieblehre. Dazu wird interaktiv eine Gerade in das Bildfenster
gezeichnet, wobei der Startpunkt z.B. an der linken vorderen Kante des Mess-
schnabels gewählt und der Endpunkt an der rechten vorderen Kante des Mess-
schnabels gesetzt wird. Nach Eingabe des Abstandes in der Basiseinheit Meter
(hier 0,01m) ist das System nun in der Lage den Umrechnungsfaktor von Pixel
nach Meter zu bestimmen. Abb. 91 (Rechts) zeigt die interaktive Vermessung
eines 30mm Endmaßes nach erfolgter Kalibrierung.
Ein weiteres Anwendungsgebiet für den Einsatz dieser Software ist die optische
Qualitätskontrolle von Bauteilen auf Defekte wie z.B. Kratzer oder Luftein-
schlüsse bei lackierten Oberflächen. Da viele Oberflächen von beispielsweise
lackierten Kunststoffteilen eine relativ homogene Färbung aufweisen, fallen De-
fekte meist durch einen farblichen Unterschied zur lokalen Umgebung auf. Mit
Hilfe eines Templates, das die Farben der zu suchenden Defekte enthält, kön-
nen mit dem Segmentierungsverfahren (Kap. 5.2.3) gezielt Fehlstellen auf der
Materialoberfläche detektiert und ausgewertet werden (Fläche, Form, …).

11. Zusammenfassung und Ausblick 155
11. Zusammenfassung und Ausblick
Gegenstand der vorliegenden Arbeit ist die Entwicklung einer vielseitig einsetz-
baren Zellanalysesoftware für die Biokompatibilitätsprüfung von Medizinproduk-
ten. Zurzeit besteht großer Bedarf an automatisierten Zellanalysesystemen für
die Biokompatibilitätsprüfung, da die Prüfungen manuell und somit sehr kosten-
intensiv und fehleranfällig durchgeführt werden. Zwar lässt sich das Experten-
wissen einer erfahrenen Fachkraft nicht durch ein computergestütztes Verfah-
ren ersetzen, jedoch verringert der Einsatz einer Analysesoftware die Prüfzei-
ten, spart Kosten ein und führt zu einer reproduzierbaren, objektiven Bewertung
des Bildmaterials. Um das Expertenwissen einer erfahrenen Fachkraft einflie-
ßen zu lassen, ist die Software so konzipiert, dass bei Bedarf Änderungen an
der Analyse vorgenommen werden können. Derzeit existieren bereits verschie-
dene Zellanalyseverfahren, jedoch sind sie meistens für spezielle Anwendungs-
fälle konzipiert und somit nicht flexibel einsetzbar. Die in dieser Arbeit entwickel-
te Software verfügt über eine hohe Generalisierungsleistung. Sie kann für die
Analyse unterschiedlicher Zelllinien Verwendung finden und lässt sich neben
der Zellanalyse auch auf andere Problemstellungen erweitern, bei denen eine
Teilchenanalyse im Vordergrund steht. Eine besondere Herausforderung stellt
die starke morphologische Variation der untersuchten Zellen dar. Aus diesem
Grund wird die Separation agglomerierter Zellen erschwert. Eine weitere Prob-
lematik stellt die verwendete Färbemethode dar. Diese Anwendung wählt eine
zytochemische Färbung, da sich so mit nur einem Färbeprozess im Gegensatz
zu einer Fluoreszenzfärbung mehrere Zellmerkmale visualisieren lassen. Zu-
dem ist eine zytochemische Färbung kostengünstiger und einfacher umsetzbar.
Diesen Vorteilen stehen die Nachteile gegenüber, dass der Bildkontrast deutlich
geringer ausfällt und Farbvariationen der Zellbereiche auftreten.
Zu Beginn der Biokompatibilitätsprüfung muss das Probenmaterial für das an-
schließende Aufbringen der Zellen durch mehrere Prozessschritte vorbereitet
werden. Nachdem die Zellen einen festgelegten Zeitraum in Kontakt mit dem
Material verbracht haben, folgt die zytochemische Anfärbung und anschließend
die Analyse unter dem Mikroskop. Das vorgestellte computergestützte Verfah-
ren unterstützt diese Analyse. Die Software eliminiert zuerst Bildartefakte wie
z.B. Korrosionserscheinungen des Materials. Je nach verwendeter Zelllinie
kommen unterschiedliche Mikroskopkontraste zum Einsatz, für die jeweils taug-
liche Zellsegmentierungsverfahren ausgewählt werden. Unter den entstehen-
den getrennten Zellbereichen befinden sich oft auch agglomerierte Zellen, die
eine Separation erforderlich machen. Sind die Kerne der Zellen sichtbar, wird

11. Zusammenfassung und Ausblick 156
ihre Detektion mit dem Histogram-Backprojection-Algorithmus durchgeführt. Die
darauf folgende Separation der agglomerierten Zellen findet mit Hilfe von zwei
verschiedenen Verfahren statt. Das erste, im Rahmen dieser Arbeit entwickelte
Verfahren, eignet sich bei Zelllinien mit kompakten zusammenhängen Zellclus-
tern (z.B. MC3T3) und separiert die Zellen durch eine Wachstumssimulation,
ausgehend von den detektierten Kernbereichen. Das zweite Verfahren, dass
verschiedene aus der Literatur bekannte Verfahren kombiniert, nutzt die Tatsa-
che, dass manche Zelllinien (z.B. L929) bei agglomerierten Zellen Verengungen
an den Zellkontaktstellen bilden, die über die Berechnung der dominanten Kon-
turpunkte ermittelt werden. Der A*-Algorithmus berechnet anschließend die kür-
zesten Pfade zwischen den Schwerpunkten der Zellkerne. Unter Berücksichti-
gung verschiedener Regeln, die eine aus biologischer Sicht korrekte Trennung
gewährleisten, werden aus den Pfadpunkten außerhalb der Kernbereiche und
den dominanten Konturpunkten die Trennlinien der Zellen generiert. Eine Vor-
schaltung der, ebenfalls im Rahmen dieser Arbeit entwickelten, kontextbasier-
ten Trennfunktion (CBS) vor das zuletzt genannte Separationsverfahren führt zu
einer Verbesserung der Analysegenauigkeit und zur Reduktion der Rechenzeit.
Die Einstellung der CBS-Parameter kann mit Hilfe eines genetischen Algorith-
mus automatisch vorgenommen werden. Für die anderen beiden Separations-
verfahren ist jeweils nur ein Parameter einzustellen, daher wurde auf eine au-
tomatische Parameterfestlegung verzichtet.
Sind keine Zellkerne im Bild sichtbar, so steht für die Zellseparation in Clustern
ein weiteres aus der Literatur bekanntes Verfahren zur Verfügung. Es eignet
sich ausschließlich für Zelllinien mit Verengungen an den Zellkontaktstellen.
Nach der Zellseparation folgt die Prüfung der durch den Rand teilweise abge-
schnittenen Zellen dahingehend, ob sie für die Auswertung übernommen oder
verworfen werden. Zur Bewertung des Biokompatibilitätsgrades wurden ver-
schiedene Zellmerkmale auf ihre Tauglichkeit hin untersucht. Mit Hilfe der Soft-
ware lassen sich die durchschnittliche Kompaktheit, Zelldichte, Proliferationsra-
te sowie die durchschnittliche Einzelzellfläche berechnen.
Die Software erstellt zu Beginn von einem als 100% biokompatibel angenom-
menen Material Referenzdaten. Diese umfassen die Werte zu den bereits ge-
nannten Zellmerkmalen. Die Daten aus den Biokompatibilitätsprüfungen der zu
untersuchenden Materialien werden anschließend mit den zuvor erstellten Re-
ferenzdaten verglichen und daraus Kennzahlen für den Grad der Biokompatibili-
tät abgeleitet.
Für die Bestimmung der Analysegenauigkeit des computergestützten Verfah-
rens wurden die vorliegenden Zellzahlen von Experten ermittelt und mit den

11. Zusammenfassung und Ausblick 157
Ergebnissen aus der automatischen Zellzählung verglichen. Es stellte sich her-
aus, dass die automatische Zellzählung der Linie L929 bei Verwendung eines
Stichprobenumfanges von 100 Probenausschnitten zu 1,27% +/- 0,22% unter
Verwendung eines Konfidenzintervalls von 99% von der Referenzzählung ab-
weicht. Die automatische Zählung von MC3T3 Zellen weicht bei einem Umfang
von 100 Probenausschnitten zu 3,5% +/- 0,61% unter Verwendung eines Kon-
fidenzintervalls von 99% von der Referenzzählung ab. Die Zellmorphologie be-
treffenden Merkmale lassen sich auf diese Weise nicht verifizieren, da eine
Quantifizierung durch einen Experten ohne zusätzliche Hilfsmittel nicht möglich
ist. Die Untersuchungen verschiedener Zellmerkmale für die Bestimmung der
Biokompatibilität zeigen zwar, dass mit zunehmenden Toxizitätsgrad des Mate-
rials eine morphologische Änderung der Zellen eintritt, jedoch erst ab einem
stark zytotoxischen Zustand. Die Eignung der genannten Merkmale zur Beurtei-
lung der Biokompatibilität ist bei den Zelllinien L929 und MC3T3 somit nur be-
dingt gegeben, da sie ausschließlich zur Bestätigung eines stark zytotoxischen
Zustandes herangezogen werden können. Es ist jedoch nicht auszuschließen
dass andere Zelllinien stärkere morphologische Veränderungen unter toxi-
schem Einfluss zeigen und diese analysiert werden sollen. In dem Fall stellt die
Software ein geeignetes Hilfsmittel dar.
Die Arbeit ist im Rahmen des BMBF geförderten interdisziplinären Forschungs-
projektes „Computer Vision zur Biokompatibilitätsprüfung von Implantatwerk-
stoffen“ entstanden. Durch die Zusammenarbeit von Biotechnologen und Soft-
wareentwicklern konnte ein Zellanalysesystem entwickelt werden, das gezielt
auf die Bedürfnisse der Anwender bei der Arbeit mit einem Zellanalysepro-
gramm zugeschnitten ist.
Im weiteren Verlauf ist eine Auslagerung geeigneter Verfahren auf die Grafik-
karte zur Geschwindigkeitsoptimierung geplant, so dass sich ein weiterer Zeit-
gewinn für die automatische Zellzählung einstellt.
Eine Möglichkeit zur weiteren Automatisierung des Prüfverfahrens besteht da-
rin, die bisher durch einen Experten aufgenommenen Probenausschnitte auto-
matisch mit Hilfe einer Achssteuerung für das Mikroskop aufnehmen zu lassen.
Dadurch entfällt der hohe Zeitaufwand für die derzeit personengebundene Auf-
nahme der Probenausschnitte.

12. Literaturverzeichnis 158
12. Literaturverzeichnis
1. Gesundheitsausgaben und demografischer Wandel. Bundesgesundheitsblatt,
Berlin Heidelberg : Springer, 55 DOI 10.1007/s00103-012-1469-4. 2012, S.
614-623.
2. Silver, F., Doillon, C. Biocompatibility - Interactions of biological and
implantable materials. VCH Publishers Inc. 1989.
3. Wintermantel, E., Ha, S.-W. Medizintechnik Life Science Engineering. Berlin
Heidelberg : Springer, 2009.
4. Schenk, R. K. Bone response to grafts and implants. Perspectives on
biomaterials, Materials science monographs, Lin O.C.C., Chao E.Y.S. (eds.).
1986, S. 121-136.
5. ISO 10993-5, Biological testing of Medical Devices – Part 5: Tests for
Cytotoxicity, in vitro methods. 1991.
6. Comaniciu, D., Meer, P., Foran, D. Image Guided Decision Support System
for Pathology. Machine Vision and Applications, Vol 11(4). 2000, S. 213-224.
7. Mauch, D. Entwicklung eines benutzerorientierten Segmentiersystems für
biomedizinische Bilder. Hamburg : disserta Verlag, 2010.
8. Lee, K.-M., Street, W.N. Model-based Detection, Segmentation and
Classification for Image Analysis using On-line Shape Learning. Machine Vision
and Applications. 2003, S. 222-233.
9. Nischwitz, A., Fischer, M., Haberäcker, P., Socher, G. Computergrafik und
Bildverarbeitung. Wiesbaden : Vieweg+Teubner, 2011. S. 513.
10. Mulisch, M., Welsch, U. Romeis Mikroskopische Technik. Heidelberg :
Spektrum Akademischer Verlag, 2010. S. 274-275.
11. Handels, H., Horsch, A., Meinzer, H. P. Advances in medical image
computing. Methods Inf Med 46(3). 2007, S. 251-253.
12. Handels, H., Ehrhardt, J., Deserno, T.M., Meinzer, H. P., Tolxdorff, T.
Bildverarbeitung für die Medizin. Springer. 2011.
13. Athelogou, M., Schäpe, A., von Büren, E., Hummel, M., Stein, H., Binnig, G.
Vollautomatische, detaillierte Quantifizierung des Ki-67 Indizes bei Brustkrebs.
Biospektrum : Zeitschrift der Gesellschaft für Biologishe Chemie (GBCH) und
der Vereinigung für Allgemeine und Angewandte Mikrobiologie (VAAM),
Vol.10(6). 2004, S. 798-799.
14. Pham, T. D. Geo- Thresholding for Segmentation of Fluorescent
Microscopic Cell Images. Proc. Int. Conf. Mass-Data Analysis of Images and

12. Literaturverzeichnis 159
Signals in Medicine, Biotechnology, and Chemistry,Lecture Notes in Computer
Science Volume 4826/2007. 2007, S. 15-26.
15. Huang, Q., Gao, W., Cai, W. Thresholding technique with adaptive window
selection for uneven lighting image. Pattern Recognition Letters, Volume 26(6).
2005, S. 801-808.
16. Wu, H.S., Berba, J., Gil, J. Iterative thresholding for segmentation of cells
from noisy images. Journal of Microscopy, Volume 197(3). 2000, S. 296-404.
17. Qiao, Y., Hu, Q., Qian, G., Luo, S., Nowinski, W. L. Thresholding based on
variance and intensity contrast. Pattern Recognition, Volume 40(2). 2007, S.
596-608.
18. Sezgin, M., Sankur, B. Survey over image thresholding techniques and
quantitative performance evaluation. Journal of Electronic Imaging 13(1). 2004,
S. 146-165.
19. Umesh Adiga, P.S., Chaudhuri, B. Region based techniques for
segmentation of volumetric histo-pathological images. Computer Methods and
Programs in Biomedicine, 61(1). 2000, S. 23-47.
20. Kirschner, M., Wesarg, S. Automatische Initialisierung von Formmodellen
mittels modellbasierter Registrierung. Bildverarbeitung für die Medizin. 2011, S.
69-73.
21. Ruppertshofen, H., Lorenz, C., Beyerlein, P., Salah, Z., Rose, G., Schramm,
H. Fully Automatic Model Creation for Object Localization Utilizing the
Generalized Hough Transform. Bildverarbeitung für die Medizin, Springer. 2010,
S. 281-285.
22. Neumann, B. Bildverarbeitung für Einsteiger. Berlin, Heidelberg : Springer,
2005. S. 167-169.
23. Ghebreab, S., Smeulders, A.W.M. Strings: Variational Deformable Models
of Multivariate Continuous Boundary Features. IEEE Transactions on Pattern
Analysis And Machine Intelligence, Volume 25(11). 2003, S. 1399–1410.
24. Bronkorsta, P.J.H., Reinders, M.J.T., Hendriks, E.A., Grimbergen, J.,
Heethaar, R.M., Brakenhoff, G.J. On-line detection of red blood cell shape using
deformable templates. Pattern Recognition Letters, Volume 21(5). 2000, S. 413-
424.
25. Garrido, A., Pérez de la Blanca, N. Applying deformable templates for cell
image segmentation. Pattern Recognition, Volume 33(5). 2000, S. 821-832.
26. Swain, M. J., Ballard, D.H. Color indexing. International Journal Of
Computer Vision, Volume 7(1). 1991.

12. Literaturverzeichnis 160
27. Deserno, T. M. Medizinische Bildverarbeitung. Berlin Heidelberg : Springer,
2011.
28. Pommerencke, T., Dickhaus, H., Grabe, N. Vollautomatische
Einzelzellerkennung auf fluoreszenten Gewebeschnitten humaner Epidermis.
Bildverarbeitung für die Medizin. 2009, S. 311-315.
29. Velasco, F. A., Marroquín, J. L. Robust parametric active contours: the
Sandwich Snakes. Machine Vision and Applications Springer-Verlag 12(5).
2000, S. 238-242.
30. Tscherepanow, M., Zöllner, F., Hillebrand, M., Kunnert, F. Automatic
segmentation of unstained living cells in bright field microscope images.
International Conference on Mass-Data Analysis of Images and Signals in
Medicine, Biotechnology, Chemistry and Food Industry (MDA), Volume 5108.
2008, S. 158-172.
31. Murashov, D. Two-level method for segmentation of cytological images
using active contour model. Proc. of the 7th. Int. Conference on Pattern
Recognition and Image Analysis (PRIA-7), Volume 3. 2004, S. 814-817.
32. Sadeghian, F., Seman, Z., Ramli, A., Abdul Kahar, B., Saripan, M. A
Framework for White Blood Cell Segmentation in Microscopic Blood Images
Using Digital Image Processing. Biological Procedures Online, Volume 11.
2009, S. 196-206.
33. Aymeric Histace, Leila Meziou, Bogdan Matuszewski, Frederic Precioso,
Mark Murphy, Franck Carreiras. Statistical region based active contour using a
fractional entropy descriptor - Application to nuclei cell segmentation in confocal
microscopy images. Annals of British Machine Vision Association. 2013, 5, S. 1-
15.
34. Cohen, L. D. On active contour models and ballons. CVGIP: Image
Understanding, Volume 53(2). 1991, S. 211-218.
35. Colantonio, S., Gurevich, I., Salvetti, O. Automatic fuzzy-neural based
segmentation of microscopic cell images. International Journal of Signal and
Imaging Systems Engineering, 1(1). 2008, S. 18-24.
36. Meftah, B., Lezoray, O., Lecluse, M., Benyettou, A. Cell Microscopic
Segmentation with Spiking Neuron Networks. Artificial Neural Networks -
ICANN 2010, Lecture Notes in Computer Science, Volume 6352/2010. 2010, S.
117-126.
37. Feitosa, R. Q., Costa, G. A. O. P., Cazes, T. B. A genetic approach for the
automatic adaptation of segmentation parameters. 1st International Conference
on Object-based Image Analysis - ISPRS Proceedings. 2006.

12. Literaturverzeichnis 161
38. Metzler, V., Bienert, H., Lehmann, T., Mottaghy, K., Spitzer, K. A Novel
Method for Quantifying Shape Deformation Applied to Biocompatibility Testing.
ASAIO Journal, 45(4). 1999, S. 264-271.
39. Kramme, R. Medizintechnik. Berlin Heidelberg : Springer, 2011.
40. Metzler, V., Vandenhouten, R., Krone, J., Grebe, R. Wissensbasierte
Bildsegmentierung mittels stochastischer Optimierung. 5. Freiburger Workshop
Digitale Bildverarbeitung in der Medizin. 1997, S. 75-80.
41. Reham Mohammed, Omima Nomir, Iraky Khalifa. Segmentation of Acute
Lymphoblastic Leukemia Using C-Y Color Space. International Journal of
Advanced Computer Science and Applications (IJACSA). 2014, Bd. 5, 11.
42. Nilsson, B., Heyden, A. Segmentation of complex cell clusters in
microscopic images: Application to bone marrow samples. Cytometry Part A,
vol. 66A(1). 2005, S. 24-32.
43. Yung-Kuan Chan, Der-Chen Huang, Kuo-Ching Liu, Rong-Tai Chen, Xiaoyi
Jiang. An Automatic Indirect Immunofluorescence Cell Segmentation System.
Mathematical Problems in Engineering. 2014, Article ID 501206.
44. Pal, U., Rodenacker, K., Chaudhuri, B. Automatic cell segmentation in cyto-
and histometry using dominant contour feature points. Analytical cellular
pathology 17(4). 1998, S. 243-250.
45. Grobe, M., Volk, H., Münzenmayer, C., Wittenberg, T. Segmentierung von
überlappenden Zellen in Fluoreszenz- und Durchlichtaufnahmen.
Bildverarbeitung für die Medizin, Springer. 2003, S. 201-205.
46. Weixing, W., Hao, S. Cell Cluster Image Segmentation on Form Analysis. 3.
International Conference on Natural Computation, IEEE. 2007, S. 833-836.
47. Kothari, S., Chaudry, Q., Wang, M.D. Automated cell counting and cluster
segmentation using concavity detection and ellipse fitting techniques.
Biomedical Imaging IEEE International Symposium. 2009, S. 795-798.
48. Athelogou, M., Schmidt, G., Schönmeyer, R., Binnig, R., Binnig, G.
Kontextbasierte Bildanalyse mit Cognition Networks. Spektrum Akademischer
Verlag - Redaktion BIOspektrum. 2007, S. 657-659.
49. Dehning, O. Gipsy: Knowledge Based Surface Inspection. IAPR Workshop
on Machine Vision Applications. 1996, S. 237-240.
50. Rao, A., Vaisberg, E.A. Domain Segmentation and Analysis.
US2007/0202519 A1 USA, 30. August 2007.
51. Wurflinger, T., Stockhausen, J., Meyer Ebrecht, D., Bocking, A. Robust
automatic coregistration, segmentation, and classification of cell nuclei in

12. Literaturverzeichnis 162
multimodal cytopathological microscopic images. Comput. Med. Imaging Graph.
28. 2004, S. 87-98.
52. Boxberg, H.J. Leitfaden für die Zell- und Gewebekultur. Weinheim : WILEY-
VCH, 2007. S. 83, 128.
53. Lindl, T. Zell- und Gewebekultur. Heidelberg, Berlin : Spektrum, 2002. S. 73,
104.
54. Heimann, T., Thorn, M., Kunert, T., Meinzer, H. P. Empirische
Vergleichsmaße für die Evaluation von Segmentierungsergebnissen. Berlin
Heidelberg New York : Springer, 2004. S. 165.
55. Hafner, W. Segmentierung von Video-Bildfolgen durch Adaptive
Farbklassifikationen. München : Herbert Utz Verlag, 1999. S. 38.
56. Azad, P., Gockel, T., Dillmann, R. Computer Vision - Das Praxisbuch.
Aachen : Elektor-Verlag, 2007. S. 117.
57. Kvarnström, M., Logg, K., Diez, A., Bodvard, K., Käll, M. Image analysis
algorithms for cell contour recognition in budding yeast. Optics Express 16(17).
2008, S. 12943-12957.
58. Claus Brell, Juliana Brell, Siegfried Kirsch. Statistik von Null auf Hundert.
Berlin Heidelberg : Springer Spektrum, 2014, S. 125.
59. Buhl S., Neumann B., Schäfer S.C. Context-Based Separation of Cell
Clusters for the Automatic Biocompatibility Testing of Implant Materials. Journal
of Computational Medicine. 2014, Article ID 542521 doi:10.1155/2014/542521.
60. Zell, A. Simulation neuronaler Netze. Oldenbourg : Addison Wesley
Longman, 2003.
61. R. Kruse, C. Borgelt, F. Klawonn, C. Moewes, G. Ruß, M. Steinbrecher.
Computational Intelligence. Wiesbaden : Vieweg+Teubner, 2011.
62. Buhl, S., Neumann, B., Schneider, M., Lehmann, U., Eisenbarth, E.,
Weißbach, T. Automatische Segmentierung von mikroskopischen Zellbildern für
die Biokompatibilitätsprüfung mit Hilfe eines Künstlich Neuronalen Netzes. 19.
Workshop Computational Intelligence des VDI/GMA-FA 5.14 "Computational
Intelligence" und der GU-FG "Fuzzy-Systeme und Soft Computing". 2009.
63. Burger, W., Burge, M.-J. Digitale Bildverarbeitung. Berlin Heidelberg :
Springer, 2006. S. 254.
64. Crone, S. F. Neuronale Netze zur Prognose und Disposition im Handel.
Wiesbaden : Gabler Verlag, 2010. S. 179.
65. Rojas, Raul. Neural Networks - A Systematic Introduction. Berlin, New-
York : Springer, 1996. S. 151.

12. Literaturverzeichnis 163
66. Buhl, S., Neumann, B., Grothe, A. Bewertung verschiedener Methoden zur
Untersuchung mikroskopischer Zellbilder mit CV und CI für die
Bioverträglichkeit von Implantatwerkstoffen. DGaO-Proceedings. 2010.
67. Vincent, L., Soille, P. Watersheds in Digital Spaces: An Efficient Algorithm
Based On Immersion Simulations. IEEE Transactions On Pattern Analysis And
Machine Intelligence, Vol 13, No. 6. 1991, S. 583 - 598.
68. Malpica, N., de Solórzano, C., Vaquero, J., Santos, A., Vallcorba, I., García-
Sagredo, J., del Pozo, F. Applying Watershed Algorithms to the Segmentation
of Clustered Nuclei. Cytometry, Vol 28(4). 1997, S. 289-297.
69. Buhl, S., Neumann, B, Eisenbarth, E. Segmentation Of Cytological Stained
Cell Areas And Generation Of Cell Boundaries In Complex Shaded Cell
Clusters. 55.IWK - International Scientific Colloquium. 2010, S. 511-514.
70. Buhl, S., Neumann, B., Schneider, M., Lehmann, U., Eisenbarth, E.
Automatische Segmentierung von Zellkernen und den dazugehörigen Zellen in
Zellclustern für die Biokompatibilitätsprüfung. 20. Workshop Computational
Intelligence des VDI/GMA-FA 5.14 "Computational Intelligence" und der GI-FG
"Fuzzy-Systeme und Soft-Computing". 2010.
71. Hart, P. E., Nilsson, N. J., Raphael, B. Correction to A Formal Basis for the
Heuristic Determination of Minimum Cost Paths. SIGART Newsletter 37. 1972,
S. 28-29.
72. Norvig, P., Russell, S. Künstliche Intelligenz. München : Pearson Studium,
2004.
73. Buhl, S., Neumann, B., Schäfer, S., Severing, A. Automatic cell
segmentation in strongly agglomerated cell networks for different cell types.
International Journal of Computational Biology and Drug Design. 2013, Bd. 6, 1,
S. 200 - 220.
74. Lehn, J., Wegmann, H. Einführung in die Statistik. Wiesbaden : Teubner
Verlag, 2006.
75. Wu, K., Gauthier, D., Levine, M. D. Live Cell Image Segmentation. IEEE
Transactions on Biomedical Engineering, 42(1). 1995, S. 1-12.
76. Peskin, A.P., Hoeppner, D.J., Stuelten, C.H. Segmentation and Cell
Tracking of Breast Cancer Cells. Advances in Visual Computing, LNCS Vol
6938, 7th International Symposium. 2011, S. 381-391.
77. Neumann, B. Bildverarbeitung für Einsteiger. Berlin, Heidelberg : Springer,
2005. S. 148-149.
78. Claus Brell, Juliana Brell, Siegfried Kirsch. Statistik von Null auf Hundert.
Berlin Heidelberg : Springer Spektrum, 2014. S. 88, 114.

12. Literaturverzeichnis 164
79. Koch, Sören. Genetische Algorithmen - Allgemeine Grundlagen. Genetische
Algorithmen für das Order Batching-Problem in manuellen
Komissioniersystemen. Wiesbaden : Springer Fachmedien, 2014, S. 59-93.
80. Shi, D. Biomaterials and Tissue Engineering. Berlin, Heidelberg,New York :
Springer, 2004. S. 1ff, 83-108.
81. Ratner, B., Hoffmann, A., Schoen, F., Lemons, J. Biomaterials Science: an
introduction to materials in medicine. Elsevier, Vol 2. 2004, S. 1-22, 355ff., 432-
437.
82. Fraunhofer-Institut für Grenzflächen- und Biverfahrenstechnik IGB.
Biologische Beurteilung von Medizinprodukten - Prüfungen auf in-vitro-
Zytotoxizität nach DIN ISO 10993-5. 2009.
83. Grubbs, F.E. Procedures for Detecting Outlying Observations in Samples.
Technometrics, Vol. 11, No. 1. 1969, S. 1-21.
84. Ens, K. Kombinierte Registrierung und Segmentierung. Wiesbaden : Vieweg
und Teubner, 2011. S. 29-32.
85. Kruse, R., Borgelt, C., Klawonn, F., Moewes, C., Ruß, G., Steinbrecher, M.
Computational Intelligence. Wiesbaden : Vieweg+Teubner, 2011.

13. Anhang 165
13. Anhang
13.1 Untersuchung zur Farbverteilung von Korrosionserscheinungen
auf dem Material Titan*
In Abb. 92 sind 10 verschiedene Bildartefakte (rot markiert) sowie 10 unter-
schiedliche Hintergrundbereiche ohne Bildartefakte (weiß markiert) dargestellt.
Die Artefakte scheinen eine leichte Grünfärbung aufzuweisen. Das Untersu-
chungsergebnis der Farbzusammensetzung aller markierten Artefakte bestätigt
die Annahme, aufgeführt in Tabelle 49.
Abbildung 91: Zellen vom Typ L929 auf dem Substrat Titan*. Bildartefakte, hervorgerufen durch
Korrosionsprozesse, sind rot und Hintergrundbereiche ohne Artefakte weiß markiert.

13. Anhang 166
Tabelle 49: Abweichung des Rot- und Blaukanals zum Grünkanal für korrodierte Artefakte
Proben-bereich
Mittlerer Farbwert Relative Abweichung in %
Rotkanal Grünkanal Blaukanal Rot- zum Grünkanal
Blau- zum Grünkanal
1 149,8 153,5 146,1 -2,4 -4,8
2 142,3 148,4 134,2 -4,1 -9,6
3 167,3 169,2 164,1 -1,1 -3,0
4 168,4 176,2 154,5 -4,4 -12,3
5 166,1 173,1 155,5 -4,0 -10,2
6 171,7 179,1 158,9 -4,1 -11,3
7 176,2 181,9 166,5 -3,1 -8,5
8 167,4 170,8 159,6 -2,0 -6,6
9 172,3 177,9 160 -3,1 -10,1
10 176,6 182,4 169,1 -3,2 -7,3
Durchschnitt -3,2 -8,4
Abbildung 92: Abweichung des Rot- und Blaukanals zum Grünkanal für 10 verschiedene Bildar-
tefakte, hervorgerufen durch Materialkorrosion
-15,0
-10,0
-5,0
0,0
5,0
Re
lati
ve A
bw
eic
hu
ng
(%)
Relative Abweichung des Rot- und Blaukanals zum Grünkanal für Bildartefakte hervorgerufen durch Materialkorrosion
Rotkanal zum Grünkanal Blaukanal zum Grünkanal

13. Anhang 167
Tabelle 50: Abweichung des Rot- und Blaukanals zum Grünkanal für den Hintergrundbereich
ohne korrodierte Artefakte
Proben-bereich
Mittlerer Farbwert Relative Abweichung in %
Rotkanal Grünkanal Blaukanal Rot- zum Grünkanal
Blau- zum Grünkanal
1 218,2 214,9 215,1 1,5 0,1
2 217,6 215,3 214,5 1,1 -0,4
3 215,8 212,8 212,9 1,4 0,0
4 216,3 213,5 213,2 1,3 -0,1
5 219,3 216,7 215,7 1,2 -0,5
6 214,9 211,8 211,5 1,5 -0,1
7 217,4 214,5 213,7 1,4 -0,4
8 212,1 208,7 208,2 1,6 -0,2
9 213,3 210,5 210,2 1,3 -0,1
10 210,8 207,3 207,6 1,7 0,1
Durchschnitt 1,4 -0,2
Abbildung 93: Abweichung des Rot- und Blaukanals zum Grünkanal für 10 verschiedene Hin-
tergrundbereiche ohne Artefakte, hervorgerufen durch Materialkorrosion
-5,0
-3,0
-1,0
1,0
3,0
5,0
1 2 3 4 5 6 7 8 9 10
Re
lati
ve A
bw
eic
hu
ng
(%)
Relative Abweichung des Rot- und Blaukanals zum Grünkanal für den Hintergrundbereich ohne Korrosionsartefakte
Rel. Abw. Rotkanal zum Grünkanal Rel. Abw. Blaukanal zum Grünkanal
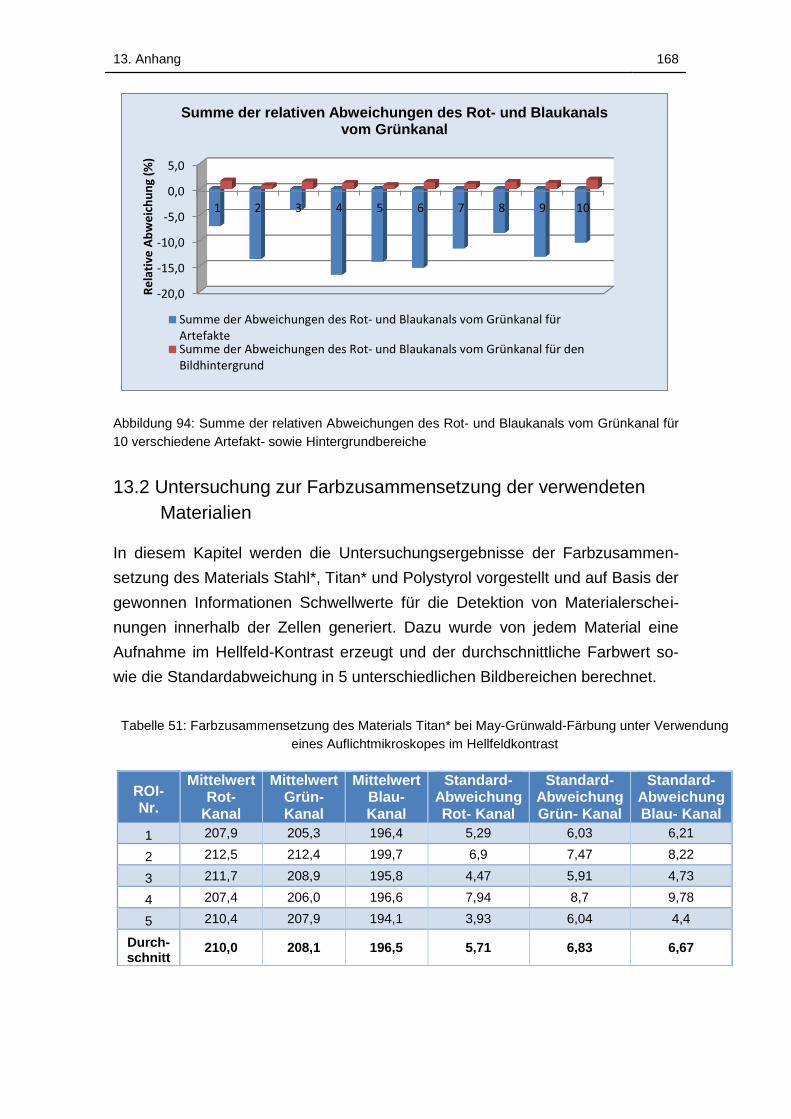
13. Anhang 168
Abbildung 94: Summe der relativen Abweichungen des Rot- und Blaukanals vom Grünkanal für
10 verschiedene Artefakt- sowie Hintergrundbereiche
13.2 Untersuchung zur Farbzusammensetzung der verwendeten
Materialien
In diesem Kapitel werden die Untersuchungsergebnisse der Farbzusammen-
setzung des Materials Stahl*, Titan* und Polystyrol vorgestellt und auf Basis der
gewonnen Informationen Schwellwerte für die Detektion von Materialerschei-
nungen innerhalb der Zellen generiert. Dazu wurde von jedem Material eine
Aufnahme im Hellfeld-Kontrast erzeugt und der durchschnittliche Farbwert so-
wie die Standardabweichung in 5 unterschiedlichen Bildbereichen berechnet.
Tabelle 51: Farbzusammensetzung des Materials Titan* bei May-Grünwald-Färbung unter Verwendung
eines Auflichtmikroskopes im Hellfeldkontrast
ROI-Nr.
Mittelwert Rot-
Kanal
Mittelwert Grün- Kanal
Mittelwert Blau- Kanal
Standard- Abweichung Rot- Kanal
Standard- Abweichung Grün- Kanal
Standard- Abweichung Blau- Kanal
1 207,9 205,3 196,4 5,29 6,03 6,21
2 212,5 212,4 199,7 6,9 7,47 8,22
3 211,7 208,9 195,8 4,47 5,91 4,73
4 207,4 206,0 196,6 7,94 8,7 9,78
5 210,4 207,9 194,1 3,93 6,04 4,4
Durch-schnitt
210,0 208,1 196,5 5,71 6,83 6,67
-20,0
-15,0
-10,0
-5,0
0,0
5,0
1 2 3 4 5 6 7 8 9 10
Re
lati
ve A
bw
eic
hu
ng
(%)
Summe der relativen Abweichungen des Rot- und Blaukanals vom Grünkanal
Summe der Abweichungen des Rot- und Blaukanals vom Grünkanal fürArtefakteSumme der Abweichungen des Rot- und Blaukanals vom Grünkanal für denBildhintergrund

13. Anhang 169
Abbildung 95: Berechnung der Farbzusammensetzung des Materials Titan* auf Basis der 5 in grün mar-
kierten Bildbereiche
Da das Material Titan* im Gegensatz zu den gefärbten Zellen deutlich heller erscheint,
werden die oberen Schwellwertgrenzen für den Rot- Grün- und Blaukanal auf SO,R =
SO,G = SO,B = 255 gesetzt. Der untere Schwellwert für den Rotkanal ergibt sich aus Ta-
belle 51, gerundet zu SU,R = , , 207,44 7,94 200.R i R iMIN MAX
x Die Schwell-
werte SU,G sowie SU,B werden analog berechnet, man erhält SU,G = 197 und SU,B = 184.
Tabelle 52: Farbzusammensetzung des Materials Stahl* bei May-Grünwald-Färbung unter Verwendung
eines Auflichtmikroskopes im Hellfeldkontrast
ROI-Nr.
Mittelwert Rot-
Kanal
Mittelwert Grün- Kanal
Mittelwert Blau- Kanal
Standard- Abweichung Rot- Kanal
Standard- Abweichung Grün- Kanal
Standard- Abweichung Blau- Kanal
1 207,2 209,3 209,6 2,52 2,94 1,84
2 209,5 213,5 212,9 2,66 3,89 2,01
3 208,3 211,6 211,8 2,25 2,56 1,73
4 205,1 207,7 208,2 2,98 4,06 2,35
5 205,3 207,7 208,5 2,83 3,82 2,25
Durch-schnitt
207,1 210,0 210,2 2,65 3,45 2,04
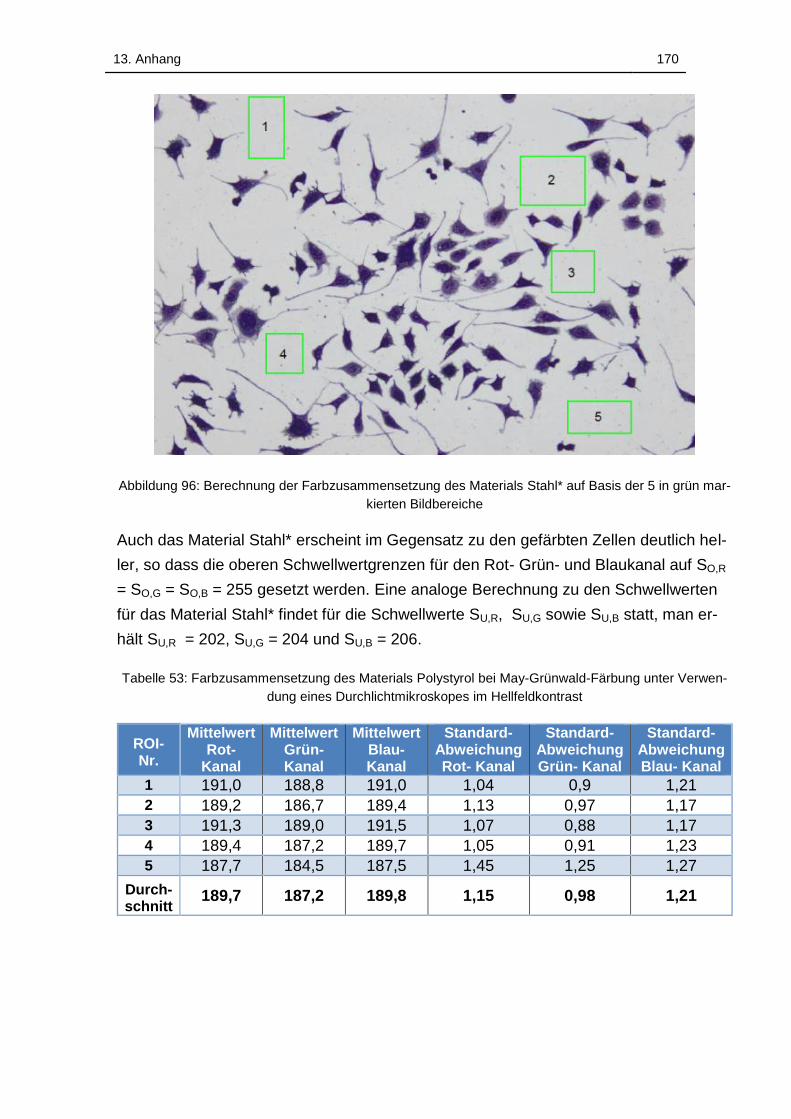
13. Anhang 170
Abbildung 96: Berechnung der Farbzusammensetzung des Materials Stahl* auf Basis der 5 in grün mar-
kierten Bildbereiche
Auch das Material Stahl* erscheint im Gegensatz zu den gefärbten Zellen deutlich hel-
ler, so dass die oberen Schwellwertgrenzen für den Rot- Grün- und Blaukanal auf SO,R
= SO,G = SO,B = 255 gesetzt werden. Eine analoge Berechnung zu den Schwellwerten
für das Material Stahl* findet für die Schwellwerte SU,R, SU,G sowie SU,B statt, man er-
hält SU,R = 202, SU,G = 204 und SU,B = 206.
Tabelle 53: Farbzusammensetzung des Materials Polystyrol bei May-Grünwald-Färbung unter Verwen-
dung eines Durchlichtmikroskopes im Hellfeldkontrast
ROI-Nr.
Mittelwert Rot-
Kanal
Mittelwert Grün- Kanal
Mittelwert Blau- Kanal
Standard- Abweichung Rot- Kanal
Standard- Abweichung Grün- Kanal
Standard- Abweichung Blau- Kanal
1 191,0 188,8 191,0 1,04 0,9 1,21
2 189,2 186,7 189,4 1,13 0,97 1,17
3 191,3 189,0 191,5 1,07 0,88 1,17
4 189,4 187,2 189,7 1,05 0,91 1,23
5 187,7 184,5 187,5 1,45 1,25 1,27
Durch-schnitt
189,7 187,2 189,8 1,15 0,98 1,21

13. Anhang 171
Abbildung 97: Berechnung der Farbzusammensetzung des Materials Polystyrol auf Basis der 5
in grün markierten Bildbereiche
Das Material Polystyrol erscheint ebenso wie Stahl* und Titan* im Gegensatz
zu den gefärbten Zellen heller, so dass die oberen Schwellwertgrenzen für den
Rot- Grün- und Blaukanal auf SO,R = SO,G = SO,B = 255 gesetzt werden. Im Ver-
gleich zu Stahl* und Titan* wirkt die Farbgebung des Polystyrols homogener
und es treten keine Bildartefakte wie Korrosionserscheinungen auf. Dies spie-
gelt sich in einer im Vergleich zu den anderen Substraten kleineren Stan-
dardabweichung wider. Die Schwellwerte SU,R, SU,G sowie SU,B werden analog
zu den Schwellwerten für die anderen Werkstoffe berechnet, man erhält SU,R =
186, SU,G = 183 und SU,B = 186.
13.3 Bewertung der in Kap. 5.2 vorgestellten Verfahren zur
Segmentierung von Zellen
Zur Bewertung der Segmentierungsleistung der in Kap. 5.2. vorgestellten Ver-
fahren wurden in den Mikroskopkontrasten Auflicht-Hellfeld, Auflicht-Dunkelfeld,
Durchlicht-Hellfeld und Durchlicht-Phasenkontrast von jeweils 50 Zellen des
Typs MC3T3 und L929 der Jaccard-Koeffizient berechnet. Als Referenzkontur
kommt die manuelle Segmentierung derselben 50 Zellen eines Experten zum
Einsatz. Tabelle 54 zeigt die ermittelten Daten der Linie L929 für das Material
Stahl*/Titan*, Tabelle 55 für Polystyrol. Tabelle 56 zeigt die Daten der Linie
MC3T3 ebenfalls für die Materialien Stahl*/Titan*, Tabelle 57 für Polystyrol.
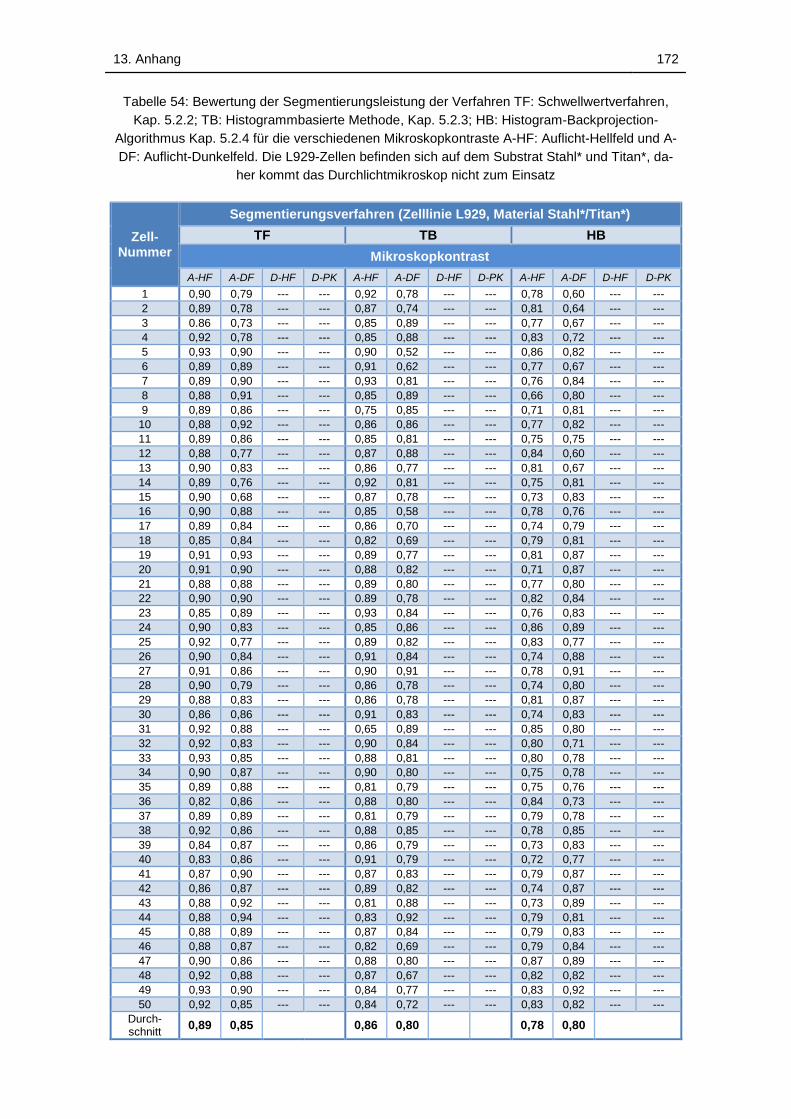
13. Anhang 172
Tabelle 54: Bewertung der Segmentierungsleistung der Verfahren TF: Schwellwertverfahren,
Kap. 5.2.2; TB: Histogrammbasierte Methode, Kap. 5.2.3; HB: Histogram-Backprojection-
Algorithmus Kap. 5.2.4 für die verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-
DF: Auflicht-Dunkelfeld. Die L929-Zellen befinden sich auf dem Substrat Stahl* und Titan*, da-
her kommt das Durchlichtmikroskop nicht zum Einsatz
Zell-Nummer
Segmentierungsverfahren (Zelllinie L929, Material Stahl*/Titan*)
TF TB HB
Mikroskopkontrast
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
1 0,90 0,79 --- --- 0,92 0,78 --- --- 0,78 0,60 --- ---
2 0,89 0,78 --- --- 0,87 0,74 --- --- 0,81 0,64 --- ---
3 0.86 0,73 --- --- 0,85 0,89 --- --- 0,77 0,67 --- ---
4 0,92 0,78 --- --- 0,85 0,88 --- --- 0,83 0,72 --- ---
5 0,93 0,90 --- --- 0,90 0,52 --- --- 0,86 0,82 --- ---
6 0,89 0,89 --- --- 0,91 0,62 --- --- 0,77 0,67 --- ---
7 0,89 0,90 --- --- 0,93 0,81 --- --- 0,76 0,84 --- ---
8 0,88 0,91 --- --- 0,85 0,89 --- --- 0,66 0,80 --- ---
9 0,89 0,86 --- --- 0,75 0,85 --- --- 0,71 0,81 --- ---
10 0,88 0,92 --- --- 0,86 0,86 --- --- 0,77 0,82 --- ---
11 0,89 0,86 --- --- 0,85 0,81 --- --- 0,75 0,75 --- ---
12 0,88 0,77 --- --- 0,87 0,88 --- --- 0,84 0,60 --- ---
13 0,90 0,83 --- --- 0,86 0,77 --- --- 0,81 0,67 --- ---
14 0,89 0,76 --- --- 0,92 0,81 --- --- 0,75 0,81 --- ---
15 0,90 0,68 --- --- 0,87 0,78 --- --- 0,73 0,83 --- ---
16 0,90 0,88 --- --- 0,85 0,58 --- --- 0,78 0,76 --- ---
17 0,89 0,84 --- --- 0,86 0,70 --- --- 0,74 0,79 --- ---
18 0,85 0,84 --- --- 0,82 0,69 --- --- 0,79 0,81 --- ---
19 0,91 0,93 --- --- 0,89 0,77 --- --- 0,81 0,87 --- ---
20 0,91 0,90 --- --- 0,88 0,82 --- --- 0,71 0,87 --- ---
21 0,88 0,88 --- --- 0,89 0,80 --- --- 0,77 0,80 --- ---
22 0,90 0,90 --- --- 0.89 0,78 --- --- 0,82 0,84 --- ---
23 0,85 0,89 --- --- 0,93 0,84 --- --- 0,76 0,83 --- ---
24 0,90 0,83 --- --- 0,85 0,86 --- --- 0,86 0,89 --- ---
25 0,92 0,77 --- --- 0,89 0,82 --- --- 0,83 0,77 --- ---
26 0,90 0,84 --- --- 0,91 0,84 --- --- 0,74 0,88 --- ---
27 0,91 0,86 --- --- 0,90 0,91 --- --- 0,78 0,91 --- ---
28 0,90 0,79 --- --- 0,86 0,78 --- --- 0,74 0,80 --- ---
29 0,88 0,83 --- --- 0,86 0,78 --- --- 0,81 0,87 --- ---
30 0,86 0,86 --- --- 0,91 0,83 --- --- 0,74 0,83 --- ---
31 0,92 0,88 --- --- 0,65 0,89 --- --- 0,85 0,80 --- ---
32 0,92 0,83 --- --- 0,90 0,84 --- --- 0,80 0,71 --- ---
33 0,93 0,85 --- --- 0,88 0,81 --- --- 0,80 0,78 --- ---
34 0,90 0,87 --- --- 0,90 0,80 --- --- 0,75 0,78 --- ---
35 0,89 0,88 --- --- 0,81 0,79 --- --- 0,75 0,76 --- ---
36 0,82 0,86 --- --- 0,88 0,80 --- --- 0,84 0,73 --- ---
37 0,89 0,89 --- --- 0,81 0,79 --- --- 0,79 0,78 --- ---
38 0,92 0,86 --- --- 0,88 0,85 --- --- 0,78 0,85 --- ---
39 0,84 0,87 --- --- 0,86 0,79 --- --- 0,73 0,83 --- ---
40 0,83 0,86 --- --- 0,91 0,79 --- --- 0,72 0,77 --- ---
41 0,87 0,90 --- --- 0,87 0,83 --- --- 0,79 0,87 --- ---
42 0,86 0,87 --- --- 0,89 0,82 --- --- 0,74 0,87 --- ---
43 0,88 0,92 --- --- 0,81 0,88 --- --- 0,73 0,89 --- ---
44 0,88 0,94 --- --- 0,83 0,92 --- --- 0,79 0,81 --- ---
45 0,88 0,89 --- --- 0,87 0,84 --- --- 0,79 0,83 --- ---
46 0,88 0,87 --- --- 0,82 0,69 --- --- 0,79 0,84 --- ---
47 0,90 0,86 --- --- 0,88 0,80 --- --- 0,87 0,89 --- ---
48 0,92 0,88 --- --- 0,87 0,67 --- --- 0,82 0,82 --- ---
49 0,93 0,90 --- --- 0,84 0,77 --- --- 0,83 0,92 --- ---
50 0,92 0,85 --- --- 0,84 0,72 --- --- 0,83 0,82 --- ---
Durch-schnitt
0,89 0,85 0,86 0,80 0,78 0,80

13. Anhang 173
Tabelle 55: Zeichenerklärung siehe Tabelle 46. Die L929-Zellen befinden sich auf dem transpa-
renten Substrat Polystyrol, daher kommt das Auflichtmikroskop nicht zum Einsatz
Zell-Nummer
Segmentierungsverfahren (Zelllinie L929, Material Polystyrol)
TF TB HB
Mikroskopkontrast
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
1 --- --- 0,88 0,64 --- --- 0,81 0,88 --- --- 0,77 0,71
2 --- --- 0,94 0,81 --- --- 0,91 0,71 --- --- 0,89 0,66
3 --- --- 0,86 0,13 --- --- 0,87 0,91 --- --- 0,82 0,19
4 --- --- 0,83 0,63 --- --- 0,82 0,70 --- --- 0,74 0,57
5 --- --- 0,89 0,40 --- --- 0,85 0,76 --- --- 0,85 0,70
6 --- --- 0,88 0,63 --- --- 0,94 0,71 --- --- 0,92 0,51
7 --- --- 0,87 0,56 --- --- 0,87 0,68 --- --- 0,86 0,54
8 --- --- 0,92 0,58 --- --- 0,87 0,77 --- --- 0,85 0,58
9 --- --- 0,85 0,45 --- --- 0,88 0,77 --- --- 0,75 0,57
10 --- --- 0,85 0,64 --- --- 0,87 0,82 --- --- 0,80 0,48
11 --- --- 0,86 0,55 --- --- 0,87 0,78 --- --- 0,83 0,59
12 --- --- 0,84 0,68 --- --- 0,86 0,83 --- --- 0,81 0,61
13 --- --- 0,88 0,65 --- --- 0,90 0,83 --- --- 0,87 0,69
14 --- --- 0,84 0,61 --- --- 0,86 0,72 --- --- 0,86 0,53
15 --- --- 0,84 0,67 --- --- 0,86 0,76 --- --- 0,83 0,62
16 --- --- 0,88 0,68 --- --- 0,89 0,65 --- --- 0,81 0,59
17 --- --- 0,88 0,66 --- --- 0,89 0,60 --- --- 0,84 0,67
18 --- --- 0,87 0,72 --- --- 0,88 0,66 --- --- 0,78 0,61
19 --- --- 0,84 0,62 --- --- 0,85 0,81 --- --- 0,73 0,65
20 --- --- 0,86 0,68 --- --- 0,86 0,74 --- --- 0,91 0,72
21 --- --- 0,93 0,45 --- --- 0,93 0,80 --- --- 0,88 0,66
22 --- --- 0,86 0,43 --- --- 0,89 0,72 --- --- 0,78 0,66
23 --- --- 0,89 0,42 --- --- 0,89 0,73 --- --- 0,84 0,72
24 --- --- 0,88 0,59 --- --- 0,88 0,77 --- --- 0,84 0,63
25 --- --- 0,85 0,59 --- --- 0,87 0,83 --- --- 0,76 0,70
26 --- --- 0,91 0,58 --- --- 0,90 0,79 --- --- 0,84 0,58
27 --- --- 0,89 0,61 --- --- 0,87 0,68 --- --- 0,82 0,62
28 --- --- 0,89 0,65 --- --- 0,87 0,69 --- --- 0,84 0,62
29 --- --- 0,90 0,63 --- --- 0,91 0,78 --- --- 0,84 0,70
30 --- --- 0,90 0,70 --- --- 0,88 0,79 --- --- 0,85 0,78
31 --- --- 0,87 0,57 --- --- 0,88 0,74 --- --- 0,82 0,52
32 --- --- 0,86 0,60 --- --- 0,88 0,77 --- --- 0,78 0,59
33 --- --- 0,80 0,72 --- --- 0,80 0,84 --- --- 0,66 0,73
34 --- --- 0,90 0,65 --- --- 0,91 0,81 --- --- 0,83 0,68
35 --- --- 0,89 0,70 --- --- 0,89 0,80 --- --- 0,85 0,61
36 --- --- 0,86 0,71 --- --- 0,88 0,76 --- --- 0,65 0,61
37 --- --- 0,81 0,63 --- --- 0,81 0,80 --- --- 0,73 0,64
38 --- --- 0,86 0,54 --- --- 0,86 0,86 --- --- 0,73 0,55
39 --- --- 0,86 0,66 --- --- 0,87 0,72 --- --- 0,73 0,63
40 --- --- 0,83 0,51 --- --- 0,85 0,86 --- --- 0,64 0,59
41 --- --- 0,93 0,65 --- --- 0,90 0,72 --- --- 0,80 0,68
42 --- --- 0,84 0,44 --- --- 0,84 0,75 --- --- 0,76 0,72
43 --- --- 0,95 0,76 --- --- 0,93 0,71 --- --- 0,89 0,72
44 --- --- 0,75 0,52 --- --- 0,78 0,78 --- --- 0,74 0,51
45 --- --- 0,89 0,67 --- --- 0,86 0,83 --- --- 0,81 0,74
46 --- --- 0,89 0,52 --- --- 0,86 0,81 --- --- 0,69 0,66
47 --- --- 0,86 0,59 --- --- 0,89 0,87 --- --- 0,88 0,73
48 --- --- 0,83 0,57 --- --- 0,84 0,76 --- --- 0,84 0,69
49 --- --- 0,88 0,73 --- --- 0,90 0,80 --- --- 0,87 0,79
50 --- --- 0,90 0,71 --- --- 0,90 0,86 --- --- 0,78 0,79
Durch-schnitt
0,87 0,60 0,87 0,77 0,81 0,63

13. Anhang 174
Tabelle 56: Zeichenerklärung siehe Tabelle 46. Die MC3T3-Zellen befinden sich auf dem Sub-
strat Stahl* und Titan*, daher kommt das Durchlichtmikroskop nicht zum Einsatz
Zell-Nummer
Segmentierungsverfahren (Zelllinie MC3T3, Material Stahl*/Titan*)
TF TB HB
Mikroskopkontrast
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
1 0,75 0,95 --- --- 0,81 0,94 --- --- 0,75 0,94 --- ---
2 0,85 0,95 --- --- 0,90 0,92 --- --- 0,87 0,90 --- ---
3 0,85 0,94 --- --- 0,85 0,93 --- --- 0,84 0,94 --- ---
4 0,87 0,94 --- --- 0,94 0,94 --- --- 0,94 0,93 --- ---
5 0,91 0,88 --- --- 0,91 0,78 --- --- 0,92 0,88 --- ---
6 0,85 0,86 --- --- 0,89 0,81 --- --- 0,92 0,91 --- ---
7 0,89 0,91 --- --- 0,92 0,93 --- --- 0,92 0,94 --- ---
8 0,91 0,91 --- --- 0,91 0,92 --- --- 0,87 0,93 --- ---
9 0,82 0,92 --- --- 0,91 0,93 --- --- 0,89 0,91 --- ---
10 0,87 0,90 --- --- 0,92 0,88 --- --- 0,90 0,90 --- ---
11 0,86 0,73 --- --- 0,87 0,73 --- --- 0,86 0,71 --- ---
12 0,92 0,90 --- --- 0,92 0,87 --- --- 0,79 0,91 --- ---
13 0,86 0,92 --- --- 0,88 0,90 --- --- 0,76 0,93 --- ---
14 0,87 0,93 --- --- 0,72 0,93 --- --- 0,83 0,91 --- ---
15 0,93 0,93 --- --- 0.94 0,93 --- --- 0,91 0,94 --- ---
16 0,92 0,93 --- --- 0,92 0,90 --- --- 0,89 0,86 --- ---
17 0,85 0,93 --- --- 0,85 0,90 --- --- 0,78 0,89 --- ---
18 0,80 0,90 --- --- 0,83 0,90 --- --- 0,79 0,84 --- ---
19 0,89 0,95 --- --- 0,85 0,94 --- --- 0,75 0,89 --- ---
20 0,92 0,90 --- --- 0,91 0,86 --- --- 0,89 0,86 --- ---
21 0,84 0,87 --- --- 0,84 0,86 --- --- 0,86 0,71 --- ---
22 0,91 0,87 --- --- 0,82 0,87 --- --- 0,91 0,78 --- ---
23 0,87 0,92 --- --- 0,85 0,84 --- --- 0,85 0,83 --- ---
24 0,86 0,93 --- --- 0,88 0,91 --- --- 0,81 0,87 --- ---
25 0,95 0,93 --- --- 0,94 0,94 --- --- 0,93 0,91 --- ---
26 0,88 0,87 --- --- 0,89 0,82 --- --- 0,81 0,83 --- ---
27 0,87 0,85 --- --- 0,69 0,82 --- --- 0,85 0,82 --- ---
28 0,84 0,88 --- --- 0,81 0,81 --- --- 0,85 0,88 --- ---
29 0,75 0,89 --- --- 0,81 0,80 --- --- 0,84 0,85 --- ---
30 0,86 0,84 --- --- 0,69 0,74 --- --- 0,50 0,81 --- ---
31 0,93 0,90 --- --- 0,92 0,87 --- --- 0,91 0,87 --- ---
32 0,93 0,86 --- --- 0,93 0,80 --- --- 0,93 0,82 --- ---
33 0,92 0,88 --- --- 0,93 0,73 --- --- 0,80 0,86 --- ---
34 0,90 0,83 --- --- 0,84 0,83 --- --- 0,88 0,83 --- ---
35 0,89 0,88 --- --- 0,86 0,78 --- --- 0,86 0,52 --- ---
36 0,91 0,86 --- --- 0,91 0,76 --- --- 0,91 0,84 --- ---
37 0,94 0,88 --- --- 0,92 0,74 --- --- 0,91 0,87 --- ---
38 0,89 0,91 --- --- 0,89 0,89 --- --- 0,86 0,91 --- ---
39 0,93 0,92 --- --- 0,86 0,91 --- --- 0,89 0,88 --- ---
40 0,91 0,90 --- --- 0,85 0,85 --- --- 0,85 0,89 --- ---
41 0,86 0,92 --- --- 0,88 0,83 --- --- 0,84 0,86 --- ---
42 0,92 0,89 --- --- 0,86 0,76 --- --- 0,90 0,80 --- ---
43 0,91 0,91 --- --- 0,91 0,79 --- --- 0,91 0,86 --- ---
44 0,90 0,88 --- --- 0,91 0,79 --- --- 0,89 0,85 --- ---
45 0,89 0,86 --- --- 0,83 0,57 --- --- 0,82 0,73 --- ---
46 0,75 0,92 --- --- 0,76 0,91 --- --- 0,80 0,90 --- ---
47 0,85 0,91 --- --- 0,89 0,86 --- --- 0,83 0,92 --- ---
48 0,88 0,92 --- --- 0,77 0,92 --- --- 0,74 0,93 --- ---
49 0,76 0,90 --- --- 0,83 0,89 --- --- 0,71 0,91 --- ---
50 0,77 0,82 --- --- 0,86 0,85 --- --- 0,80 0,79 --- ---
Durch-schnitt
0,87 0,90 0,86 0,85 0,85 0,86

13. Anhang 175
Tabelle 57: Zeichenerklärung siehe Tabelle 46. Die MC3T3-Zellen befinden sich auf dem trans-
parenten Substrat Polystyrol, daher kommt das Auflichtmikroskop nicht zum Einsatz
Zell-Nummer
Segmentierungsverfahren (Zelllinie MC3T3, Material Polystyrol)
TF TB HB
Mikroskopkontrast
A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK A-HF A-DF D-HF D-PK
1 --- --- 0,82 0,59 --- --- 0,86 0,85 --- --- 0,79 0,49
2 --- --- 0,74 0,45 --- --- 0,77 0,76 --- --- 0,76 0,57
3 --- --- 0,84 0,40 --- --- 0,77 0,83 --- --- 0,81 0,67
4 --- --- 0,78 0,71 --- --- 0,70 0,74 --- --- 0,83 0,56
5 --- --- 0,81 0,31 --- --- 0,86 0,77 --- --- 0,87 0,65
6 --- --- 0,80 0,82 --- --- 0,83 0,78 --- --- 0,75 0,68
7 --- --- 0,80 0,83 --- --- 0,84 0,83 --- --- 0,82 0,60
8 --- --- 0,72 0,46 --- --- 0,66 0,79 --- --- 0,71 0,44
9 --- --- 0,80 0,29 --- --- 0,68 0,92 --- --- 0,70 0,73
10 --- --- 0,70 0,41 --- --- 0,85 0,88 --- --- 0,86 0,79
11 --- --- 0,74 0,48 --- --- 0,69 0,85 --- --- 0,72 0,63
12 --- --- 0,73 0,28 --- --- 0,83 0,9 --- --- 0,77 0,78
13 --- --- 0,79 0,70 --- --- 0,86 0,85 --- --- 0,80 0,55
14 --- --- 0,73 0,23 --- --- 0,88 0,82 --- --- 0,87 0,48
15 --- --- 0,69 0,12 --- --- 0,80 0,70 --- --- 0,89 0,38
16 --- --- 0,83 0,58 --- --- 0,89 0,79 --- --- 0,92 0,40
17 --- --- 0,81 0,67 --- --- 0,88 0,81 --- --- 0,74 0,50
18 --- --- 0,69 0,73 --- --- 0,77 0,83 --- --- 0,75 0,64
19 --- --- 0,79 0,65 --- --- 0,80 0,81 --- --- 0,76 0,54
20 --- --- 0,75 0,61 --- --- 0,86 0,85 --- --- 0,85 0,58
21 --- --- 0,78 0,47 --- --- 0,71 0,88 --- --- 0,61 0,72
22 --- --- 0,67 0,65 --- --- 0,83 0,82 --- --- 0,80 0,59
23 --- --- 0,75 0,75 --- --- 0,85 0,82 --- --- 0,79 0,62
24 --- --- 0,72 0,69 --- --- 0,84 0,80 --- --- 0,83 0,61
25 --- --- 0,85 0,24 --- --- 0,86 0,80 --- --- 0,73 0,33
26 --- --- 0,84 0;65 --- --- 0,88 0,88 --- --- 0,88 0,55
27 --- --- 0,74 0,54 --- --- 0,87 0,85 --- --- 0,85 0,67
28 --- --- 0,77 0,66 --- --- 0,93 0,86 --- --- 0,84 0,42
29 --- --- 0,79 0,71 --- --- 0,87 0,85 --- --- 0,81 0,49
30 --- --- 0,69 0,64 --- --- 0,77 0,84 --- --- 0,87 0,58
31 --- --- 0,77 0,09 --- --- 0,86 0,90 --- --- 0,84 0,64
32 --- --- 0,77 0,57 --- --- 0,79 0,84 --- --- 0,85 0,69
33 --- --- 0,80 0,77 --- --- 0,81 0,89 --- --- 0,78 0,57
34 --- --- 0,78 0,64 --- --- 0,81 0,77 --- --- 0,73 0,5
35 --- --- 0,77 0,56 --- --- 0,84 0,86 --- --- 0,79 0,7
36 --- --- 0,77 0,77 --- --- 0,86 0,85 --- --- 0,81 0,6
37 --- --- 0,64 0,56 --- --- 0,79 0,83 --- --- 0,75 0,71
38 --- --- 0,81 0,66 --- --- 0,87 0,87 --- --- 0,82 0,56
39 --- --- 0,74 0,60 --- --- 0,84 0,79 --- --- 0,82 0,47
40 --- --- 0,78 0,76 --- --- 0,85 0.91 --- --- 0,80 0,56
41 --- --- 0,76 0,52 --- --- 0,83 0,80 --- --- 0,78 0,57
42 --- --- 0,61 0,48 --- --- 0,66 0,82 --- --- 0,64 0,58
43 --- --- 0,69 0,70 --- --- 0,84 0,76 --- --- 0,73 0,47
44 --- --- 0,81 0,60 --- --- 0,86 0,71 --- --- 0,81 0,47
45 --- --- 0,66 0,17 --- --- 0,73 0,67 --- --- 0,71 0,47
46 --- --- 0,70 0,28 --- --- 0,81 0,79 --- --- 0,77 0,68
47 --- --- 0,80 0,65 --- --- 0,8 0,86 --- --- 0,64 0,61
48 --- --- 0,81 0,50 --- --- 0,82 0,84 --- --- 0,66 0,59
49 --- --- 0,85 0,24 --- --- 0,86 0,88 --- --- 0,67 0,57
50 --- --- 0,79 0,15 --- --- 0,87 0,85 --- --- 0,82 0,50
Durch-schnitt
0,76 0,53 0,82 0,82 0,78 0,58

13. Anhang 176
13.4 Untersuchung zur Klassifizierung der Zellbereiche mit Hilfe
geometrischer Merkmale
Zur Überprüfung der Eignung geometrischer Merkmale zur Klassifizierung der
segmentierten Zellbereiche in Einzelzelle oder Zellcluster wurden aus fünf un-
terschiedlichen Probenbereichen von 100 als Cluster und 100 als Einzelzelle
eingeordneten Zellbereichen die Fläche und Kompaktheit ermittelt.
Abbildung 98: Vergleich der Fläche von 100 Einzelzellen und 100 Zellcluster aus fünf unter-
schiedlichen Proben der Linie L929
Abbildung 99: Vergleich der Kompaktheit von 100 Einzelzellen und 100 Zellcluster aus fünf
unterschiedlichen Proben der Linie L929
0
2000
4000
6000
8000
10000
12000
14000
16000
18000
20000
0 20 40 60 80 100 120
Fläc
he
(P
ixe
l)
Nummer der Einzelzelle bzw. des Clusters
Fläche von Einzelzellen im Vergleich zur Fläche der Zellcluster
Einzelzellen Cluster
0
2
4
6
8
10
12
0 20 40 60 80 100 120
Ko
mp
akth
eit
Nummer der Einzelzelle bzw. des Clusters
Kompaktheit von Einzelzellen im Vergleich zur Kompaktheit der Zellcluster
Einzelzellen Cluster
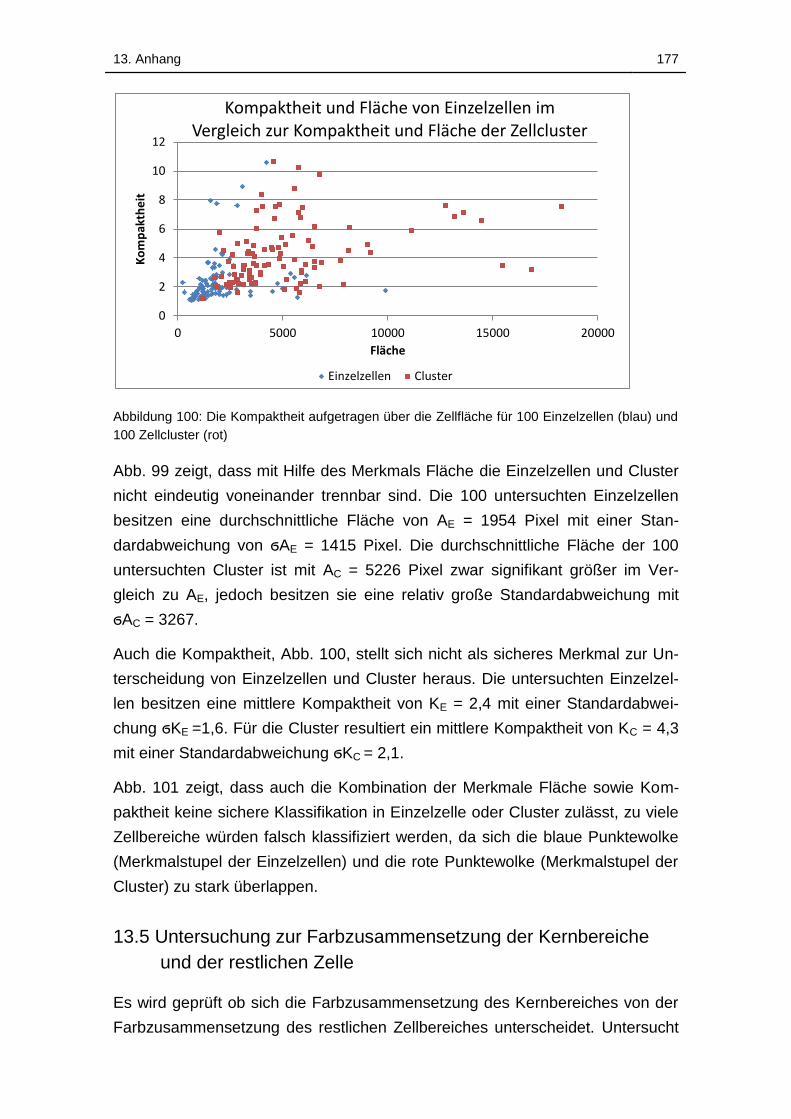
13. Anhang 177
Abbildung 100: Die Kompaktheit aufgetragen über die Zellfläche für 100 Einzelzellen (blau) und
100 Zellcluster (rot)
Abb. 99 zeigt, dass mit Hilfe des Merkmals Fläche die Einzelzellen und Cluster
nicht eindeutig voneinander trennbar sind. Die 100 untersuchten Einzelzellen
besitzen eine durchschnittliche Fläche von AE = 1954 Pixel mit einer Stan-
dardabweichung von ϭAE = 1415 Pixel. Die durchschnittliche Fläche der 100
untersuchten Cluster ist mit AC = 5226 Pixel zwar signifikant größer im Ver-
gleich zu AE, jedoch besitzen sie eine relativ große Standardabweichung mit
ϭAC = 3267.
Auch die Kompaktheit, Abb. 100, stellt sich nicht als sicheres Merkmal zur Un-
terscheidung von Einzelzellen und Cluster heraus. Die untersuchten Einzelzel-
len besitzen eine mittlere Kompaktheit von KE = 2,4 mit einer Standardabwei-
chung ϭKE =1,6. Für die Cluster resultiert ein mittlere Kompaktheit von KC = 4,3
mit einer Standardabweichung ϭKC = 2,1.
Abb. 101 zeigt, dass auch die Kombination der Merkmale Fläche sowie Kom-
paktheit keine sichere Klassifikation in Einzelzelle oder Cluster zulässt, zu viele
Zellbereiche würden falsch klassifiziert werden, da sich die blaue Punktewolke
(Merkmalstupel der Einzelzellen) und die rote Punktewolke (Merkmalstupel der
Cluster) zu stark überlappen.
13.5 Untersuchung zur Farbzusammensetzung der Kernbereiche
und der restlichen Zelle
Es wird geprüft ob sich die Farbzusammensetzung des Kernbereiches von der
Farbzusammensetzung des restlichen Zellbereiches unterscheidet. Untersucht
0
2
4
6
8
10
12
0 5000 10000 15000 20000
Ko
mp
akth
eit
Fläche
Kompaktheit und Fläche von Einzelzellen im Vergleich zur Kompaktheit und Fläche der Zellcluster
Einzelzellen Cluster

13. Anhang 178
werden jeweils 50 Zellen vom Typ L929 und MC3T3. Dabei wird jede Zellfläche
iZA und der darin enthaltene Kernbereich iKA segmentiert, mit i = 1, 2, …, 50.
Daraufhin werden die Histogrammschwerpunkte ,S RR , ,S RG und ,S RB der Histo-
gramme des Rot- Grün- und Blaukanals von den Zellflächen ohne Kernbereich
i i iR Z KA A A berechnet. Des Weiteren werden die Histogrammschwerpunkte
,S KR , ,S KG und ,S KB der Histogramme des Rot- Grün- und Blaukanals von den
Zellkernbereichen iKA berechnet.
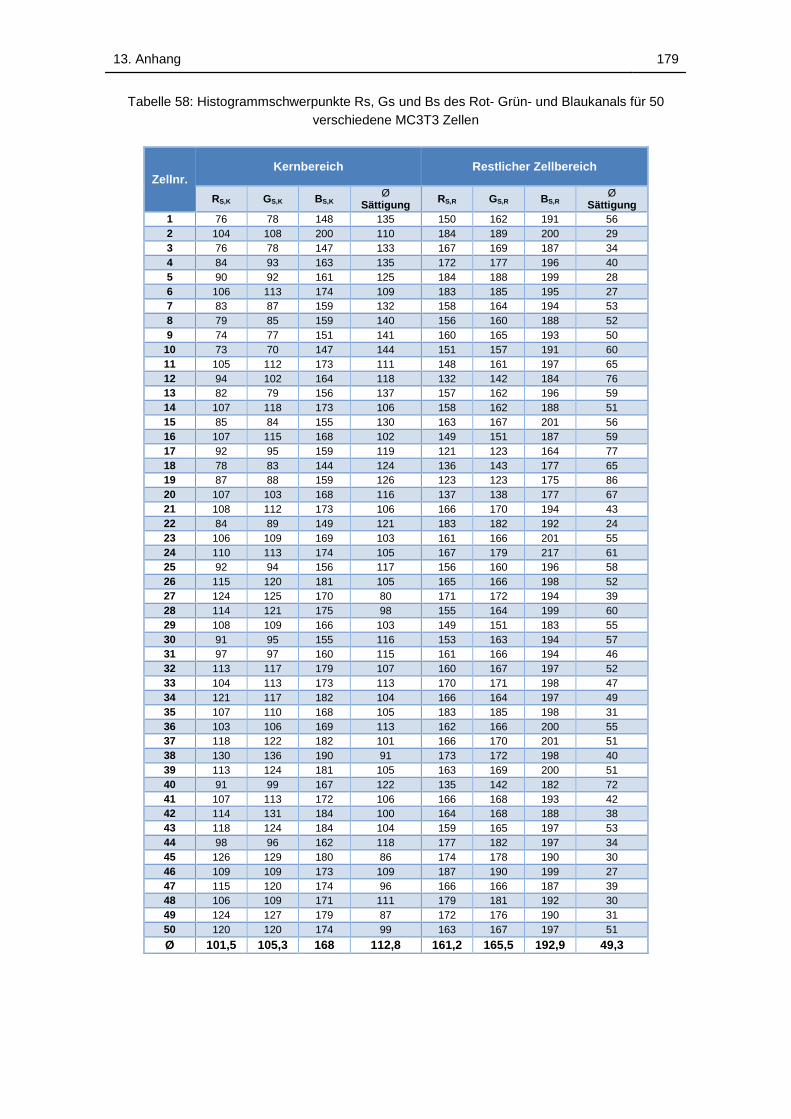
13. Anhang 179
Tabelle 58: Histogrammschwerpunkte Rs, Gs und Bs des Rot- Grün- und Blaukanals für 50
verschiedene MC3T3 Zellen
Zellnr. Kernbereich Restlicher Zellbereich
RS,K GS,K BS,K Ø
Sättigung RS,R GS,R BS,R
Ø Sättigung
1 76 78 148 135 150 162 191 56
2 104 108 200 110 184 189 200 29
3 76 78 147 133 167 169 187 34
4 84 93 163 135 172 177 196 40
5 90 92 161 125 184 188 199 28
6 106 113 174 109 183 185 195 27
7 83 87 159 132 158 164 194 53
8 79 85 159 140 156 160 188 52
9 74 77 151 141 160 165 193 50
10 73 70 147 144 151 157 191 60
11 105 112 173 111 148 161 197 65
12 94 102 164 118 132 142 184 76
13 82 79 156 137 157 162 196 59
14 107 118 173 106 158 162 188 51
15 85 84 155 130 163 167 201 56
16 107 115 168 102 149 151 187 59
17 92 95 159 119 121 123 164 77
18 78 83 144 124 136 143 177 65
19 87 88 159 126 123 123 175 86
20 107 103 168 116 137 138 177 67
21 108 112 173 106 166 170 194 43
22 84 89 149 121 183 182 192 24
23 106 109 169 103 161 166 201 55
24 110 113 174 105 167 179 217 61
25 92 94 156 117 156 160 196 58
26 115 120 181 105 165 166 198 52
27 124 125 170 80 171 172 194 39
28 114 121 175 98 155 164 199 60
29 108 109 166 103 149 151 183 55
30 91 95 155 116 153 163 194 57
31 97 97 160 115 161 166 194 46
32 113 117 179 107 160 167 197 52
33 104 113 173 113 170 171 198 47
34 121 117 182 104 166 164 197 49
35 107 110 168 105 183 185 198 31
36 103 106 169 113 162 166 200 55
37 118 122 182 101 166 170 201 51
38 130 136 190 91 173 172 198 40
39 113 124 181 105 163 169 200 51
40 91 99 167 122 135 142 182 72
41 107 113 172 106 166 168 193 42
42 114 131 184 100 164 168 188 38
43 118 124 184 104 159 165 197 53
44 98 96 162 118 177 182 197 34
45 126 129 180 86 174 178 190 30
46 109 109 173 109 187 190 199 27
47 115 120 174 96 166 166 187 39
48 106 109 171 111 179 181 192 30
49 124 127 179 87 172 176 190 31
50 120 120 174 99 163 167 197 51
Ø 101,5 105,3 168 112,8 161,2 165,5 192,9 49,3

13. Anhang 180
Tabelle 59: Histogrammschwerpunkte Rs, Gs und Bs des Rot- Grün- und Blaukanals für 50
verschiedene L929 Zellen
Zellnr. Kernbereich Restlicher Zellbereich
RS,K GS,K BS,K Ø
Sättigung RS,R GS,R BS,R
Ø Sättigung
1 71 66 153 154 133 137 187 85
2 71 69 146 142 111 106 170 107
3 63 56 134 150 117 113 177 105
4 83 91 168 134 160 169 205 59
5 94 102 178 128 146 156 204 76
6 80 97 172 140 136 145 199 86
7 84 85 157 127 165 170 206 55
8 82 81 160 135 168 170 212 60
9 69 62 132 138 155 160 193 59
10 78 75 151 136 126 120 177 92
11 66 78 168 162 108 117 181 114
12 62 72 151 155 145 158 206 79
13 95 117 187 128 163 177 209 58
14 102 116 183 119 149 156 183 55
15 59 59 143 159 120 125 179 95
16 87 97 177 133 147 156 197 73
17 72 72 148 140 160 165 202 62
18 63 61 140 149 124 133 176 86
19 61 60 141 155 132 144 187 83
20 63 60 144 155 137 137 195 87
21 57 52 130 158 110 115 171 103
22 78 79 151 135 176 181 200 39
23 73 71 147 141 132 138 183 81
24 95 97 170 123 174 178 205 46
25 82 83 162 136 162 167 201 57
26 73 67 143 141 172 177 208 50
27 74 79 164 150 135 143 194 85
28 74 69 153 149 168 170 210 60
29 60 50 113 145 128 135 173 78
30 94 96 166 121 170 171 200 47
31 110 116 187 113 176 183 205 40
32 75 85 167 150 151 158 205 73
33 98 103 178 123 186 191 209 33
34 84 88 162 130 167 171 206 55
35 78 78 160 142 180 185 211 43
36 69 76 155 153 141 143 200 85
37 57 53 114 139 141 142 193 83
38 91 94 172 131 176 179 212 50
39 77 77 148 131 162 167 200 53
40 81 81 164 138 161 163 211 69
41 99 108 184 127 170 182 217 58
42 91 97 173 127 173 177 205 45
43 66 61 131 142 157 162 195 59
44 80 90 163 133 158 167 199 57
45 62 59 134 151 120 130 174 85
46 80 91 168 138 154 159 202 68
47 75 72 144 135 167 171 199 47
48 73 65 144 149 145 145 194 77
49 70 74 148 141 150 151 187 59
50 64 68 139 140 154 157 191 56
Ø 76,9 79,1 155,3 139,4 150,4 155,4 196,1 68,3
Die Tabellen 58 und 59 zeigen, dass sich die Histogrammschwerpunkte für die
Kernbereiche von den Histogrammschwerpunkten der restlichen Zellbereiche
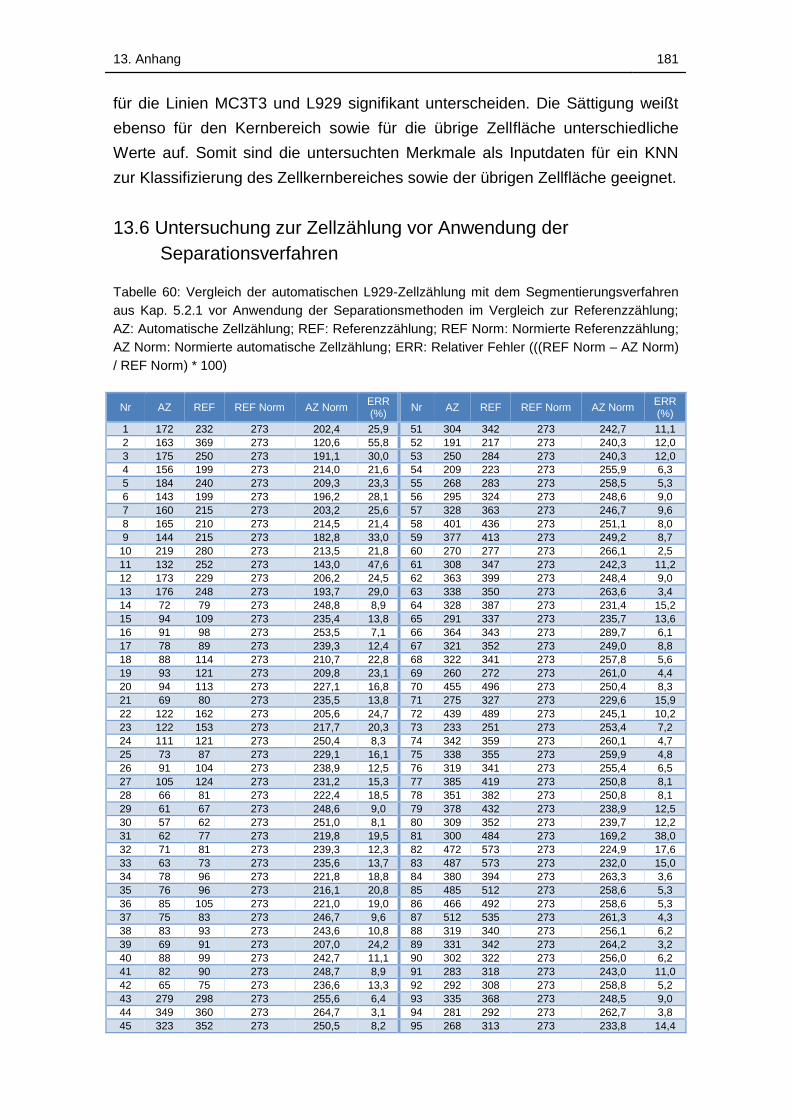
13. Anhang 181
für die Linien MC3T3 und L929 signifikant unterscheiden. Die Sättigung weißt
ebenso für den Kernbereich sowie für die übrige Zellfläche unterschiedliche
Werte auf. Somit sind die untersuchten Merkmale als Inputdaten für ein KNN
zur Klassifizierung des Zellkernbereiches sowie der übrigen Zellfläche geeignet.
13.6 Untersuchung zur Zellzählung vor Anwendung der
Separationsverfahren
Tabelle 60: Vergleich der automatischen L929-Zellzählung mit dem Segmentierungsverfahren
aus Kap. 5.2.1 vor Anwendung der Separationsmethoden im Vergleich zur Referenzzählung;
AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte Referenzzählung;
AZ Norm: Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm – AZ Norm)
/ REF Norm) * 100)
Nr AZ REF REF Norm AZ Norm ERR (%)
Nr AZ REF REF Norm AZ Norm ERR (%)
1 172 232 273 202,4 25,9 51 304 342 273 242,7 11,1
2 163 369 273 120,6 55,8 52 191 217 273 240,3 12,0
3 175 250 273 191,1 30,0 53 250 284 273 240,3 12,0
4 156 199 273 214,0 21,6 54 209 223 273 255,9 6,3
5 184 240 273 209,3 23,3 55 268 283 273 258,5 5,3
6 143 199 273 196,2 28,1 56 295 324 273 248,6 9,0
7 160 215 273 203,2 25,6 57 328 363 273 246,7 9,6
8 165 210 273 214,5 21,4 58 401 436 273 251,1 8,0
9 144 215 273 182,8 33,0 59 377 413 273 249,2 8,7
10 219 280 273 213,5 21,8 60 270 277 273 266,1 2,5
11 132 252 273 143,0 47,6 61 308 347 273 242,3 11,2
12 173 229 273 206,2 24,5 62 363 399 273 248,4 9,0
13 176 248 273 193,7 29,0 63 338 350 273 263,6 3,4
14 72 79 273 248,8 8,9 64 328 387 273 231,4 15,2
15 94 109 273 235,4 13,8 65 291 337 273 235,7 13,6
16 91 98 273 253,5 7,1 66 364 343 273 289,7 6,1
17 78 89 273 239,3 12,4 67 321 352 273 249,0 8,8
18 88 114 273 210,7 22,8 68 322 341 273 257,8 5,6
19 93 121 273 209,8 23,1 69 260 272 273 261,0 4,4
20 94 113 273 227,1 16,8 70 455 496 273 250,4 8,3
21 69 80 273 235,5 13,8 71 275 327 273 229,6 15,9
22 122 162 273 205,6 24,7 72 439 489 273 245,1 10,2
23 122 153 273 217,7 20,3 73 233 251 273 253,4 7,2
24 111 121 273 250,4 8,3 74 342 359 273 260,1 4,7
25 73 87 273 229,1 16,1 75 338 355 273 259,9 4,8
26 91 104 273 238,9 12,5 76 319 341 273 255,4 6,5
27 105 124 273 231,2 15,3 77 385 419 273 250,8 8,1
28 66 81 273 222,4 18,5 78 351 382 273 250,8 8,1
29 61 67 273 248,6 9,0 79 378 432 273 238,9 12,5
30 57 62 273 251,0 8,1 80 309 352 273 239,7 12,2
31 62 77 273 219,8 19,5 81 300 484 273 169,2 38,0
32 71 81 273 239,3 12,3 82 472 573 273 224,9 17,6
33 63 73 273 235,6 13,7 83 487 573 273 232,0 15,0
34 78 96 273 221,8 18,8 84 380 394 273 263,3 3,6
35 76 96 273 216,1 20,8 85 485 512 273 258,6 5,3
36 85 105 273 221,0 19,0 86 466 492 273 258,6 5,3
37 75 83 273 246,7 9,6 87 512 535 273 261,3 4,3
38 83 93 273 243,6 10,8 88 319 340 273 256,1 6,2
39 69 91 273 207,0 24,2 89 331 342 273 264,2 3,2
40 88 99 273 242,7 11,1 90 302 322 273 256,0 6,2
41 82 90 273 248,7 8,9 91 283 318 273 243,0 11,0
42 65 75 273 236,6 13,3 92 292 308 273 258,8 5,2
43 279 298 273 255,6 6,4 93 335 368 273 248,5 9,0
44 349 360 273 264,7 3,1 94 281 292 273 262,7 3,8
45 323 352 273 250,5 8,2 95 268 313 273 233,8 14,4
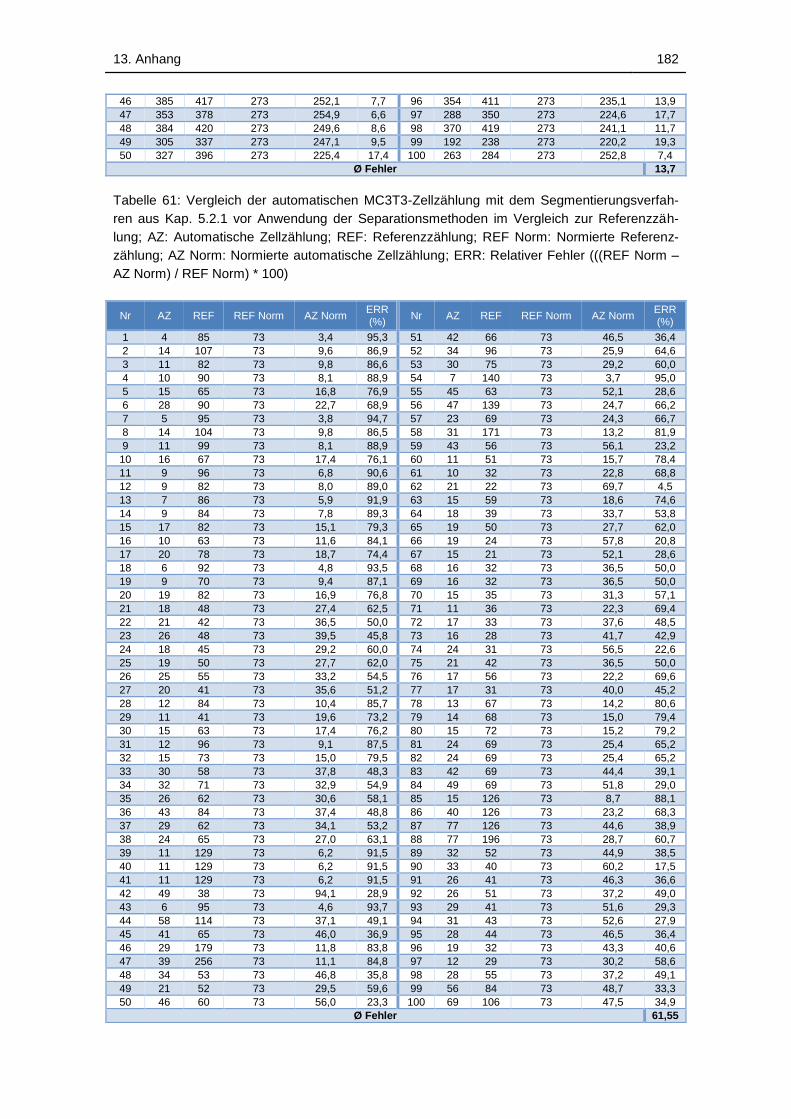
13. Anhang 182
46 385 417 273 252,1 7,7 96 354 411 273 235,1 13,9
47 353 378 273 254,9 6,6 97 288 350 273 224,6 17,7
48 384 420 273 249,6 8,6 98 370 419 273 241,1 11,7
49 305 337 273 247,1 9,5 99 192 238 273 220,2 19,3
50 327 396 273 225,4 17,4 100 263 284 273 252,8 7,4
Ø Fehler 13,7
Tabelle 61: Vergleich der automatischen MC3T3-Zellzählung mit dem Segmentierungsverfah-
ren aus Kap. 5.2.1 vor Anwendung der Separationsmethoden im Vergleich zur Referenzzäh-
lung; AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte Referenz-
zählung; AZ Norm: Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF Norm –
AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm AZ Norm ERR (%)
Nr AZ REF REF Norm AZ Norm ERR (%)
1 4 85 73 3,4 95,3 51 42 66 73 46,5 36,4
2 14 107 73 9,6 86,9 52 34 96 73 25,9 64,6
3 11 82 73 9,8 86,6 53 30 75 73 29,2 60,0
4 10 90 73 8,1 88,9 54 7 140 73 3,7 95,0
5 15 65 73 16,8 76,9 55 45 63 73 52,1 28,6
6 28 90 73 22,7 68,9 56 47 139 73 24,7 66,2
7 5 95 73 3,8 94,7 57 23 69 73 24,3 66,7
8 14 104 73 9,8 86,5 58 31 171 73 13,2 81,9
9 11 99 73 8,1 88,9 59 43 56 73 56,1 23,2
10 16 67 73 17,4 76,1 60 11 51 73 15,7 78,4
11 9 96 73 6,8 90,6 61 10 32 73 22,8 68,8
12 9 82 73 8,0 89,0 62 21 22 73 69,7 4,5
13 7 86 73 5,9 91,9 63 15 59 73 18,6 74,6
14 9 84 73 7,8 89,3 64 18 39 73 33,7 53,8
15 17 82 73 15,1 79,3 65 19 50 73 27,7 62,0
16 10 63 73 11,6 84,1 66 19 24 73 57,8 20,8
17 20 78 73 18,7 74,4 67 15 21 73 52,1 28,6
18 6 92 73 4,8 93,5 68 16 32 73 36,5 50,0
19 9 70 73 9,4 87,1 69 16 32 73 36,5 50,0
20 19 82 73 16,9 76,8 70 15 35 73 31,3 57,1
21 18 48 73 27,4 62,5 71 11 36 73 22,3 69,4
22 21 42 73 36,5 50,0 72 17 33 73 37,6 48,5
23 26 48 73 39,5 45,8 73 16 28 73 41,7 42,9
24 18 45 73 29,2 60,0 74 24 31 73 56,5 22,6
25 19 50 73 27,7 62,0 75 21 42 73 36,5 50,0
26 25 55 73 33,2 54,5 76 17 56 73 22,2 69,6
27 20 41 73 35,6 51,2 77 17 31 73 40,0 45,2
28 12 84 73 10,4 85,7 78 13 67 73 14,2 80,6
29 11 41 73 19,6 73,2 79 14 68 73 15,0 79,4
30 15 63 73 17,4 76,2 80 15 72 73 15,2 79,2
31 12 96 73 9,1 87,5 81 24 69 73 25,4 65,2
32 15 73 73 15,0 79,5 82 24 69 73 25,4 65,2
33 30 58 73 37,8 48,3 83 42 69 73 44,4 39,1
34 32 71 73 32,9 54,9 84 49 69 73 51,8 29,0
35 26 62 73 30,6 58,1 85 15 126 73 8,7 88,1
36 43 84 73 37,4 48,8 86 40 126 73 23,2 68,3
37 29 62 73 34,1 53,2 87 77 126 73 44,6 38,9
38 24 65 73 27,0 63,1 88 77 196 73 28,7 60,7
39 11 129 73 6,2 91,5 89 32 52 73 44,9 38,5
40 11 129 73 6,2 91,5 90 33 40 73 60,2 17,5
41 11 129 73 6,2 91,5 91 26 41 73 46,3 36,6
42 49 38 73 94,1 28,9 92 26 51 73 37,2 49,0
43 6 95 73 4,6 93,7 93 29 41 73 51,6 29,3
44 58 114 73 37,1 49,1 94 31 43 73 52,6 27,9
45 41 65 73 46,0 36,9 95 28 44 73 46,5 36,4
46 29 179 73 11,8 83,8 96 19 32 73 43,3 40,6
47 39 256 73 11,1 84,8 97 12 29 73 30,2 58,6
48 34 53 73 46,8 35,8 98 28 55 73 37,2 49,1
49 21 52 73 29,5 59,6 99 56 84 73 48,7 33,3
50 46 60 73 56,0 23,3 100 69 106 73 47,5 34,9
Ø Fehler 61,55
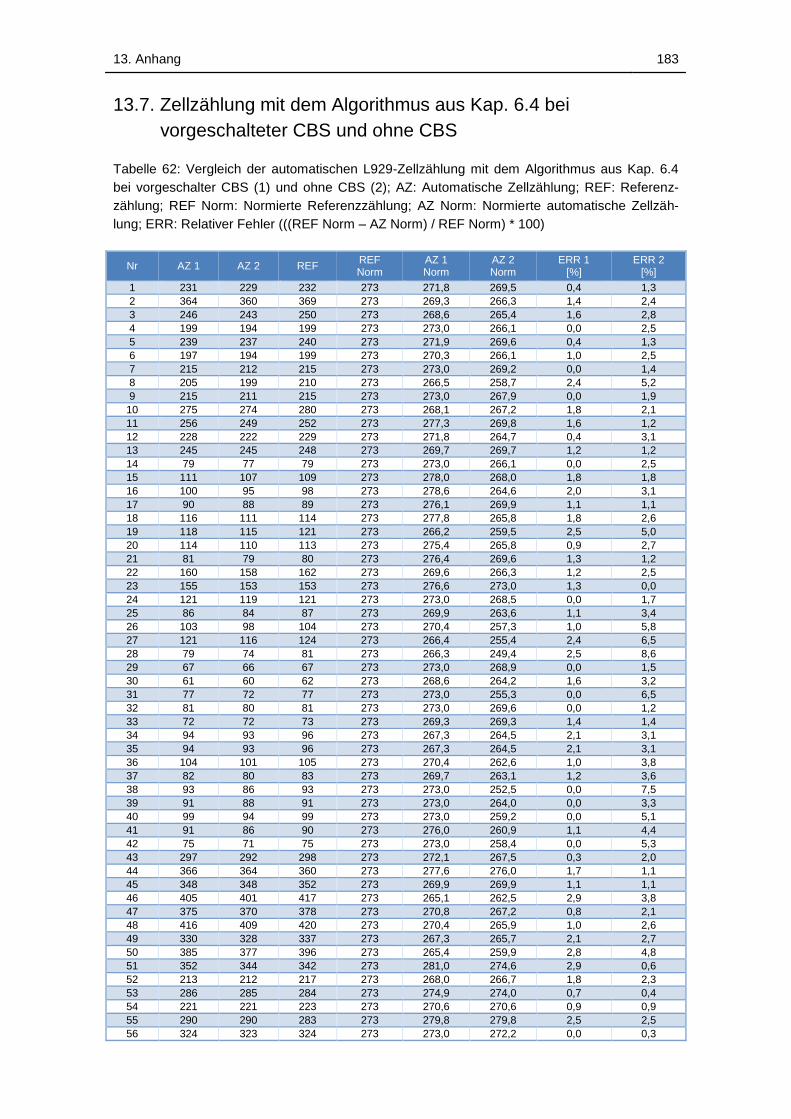
13. Anhang 183
13.7. Zellzählung mit dem Algorithmus aus Kap. 6.4 bei
vorgeschalteter CBS und ohne CBS
Tabelle 62: Vergleich der automatischen L929-Zellzählung mit dem Algorithmus aus Kap. 6.4
bei vorgeschalter CBS (1) und ohne CBS (2); AZ: Automatische Zellzählung; REF: Referenz-
zählung; REF Norm: Normierte Referenzzählung; AZ Norm: Normierte automatische Zellzäh-
lung; ERR: Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100)
Nr AZ 1 AZ 2 REF REF Norm
AZ 1 Norm
AZ 2 Norm
ERR 1 [%]
ERR 2 [%]
1 231 229 232 273 271,8 269,5 0,4 1,3
2 364 360 369 273 269,3 266,3 1,4 2,4
3 246 243 250 273 268,6 265,4 1,6 2,8
4 199 194 199 273 273,0 266,1 0,0 2,5
5 239 237 240 273 271,9 269,6 0,4 1,3
6 197 194 199 273 270,3 266,1 1,0 2,5
7 215 212 215 273 273,0 269,2 0,0 1,4
8 205 199 210 273 266,5 258,7 2,4 5,2
9 215 211 215 273 273,0 267,9 0,0 1,9
10 275 274 280 273 268,1 267,2 1,8 2,1
11 256 249 252 273 277,3 269,8 1,6 1,2
12 228 222 229 273 271,8 264,7 0,4 3,1
13 245 245 248 273 269,7 269,7 1,2 1,2
14 79 77 79 273 273,0 266,1 0,0 2,5
15 111 107 109 273 278,0 268,0 1,8 1,8
16 100 95 98 273 278,6 264,6 2,0 3,1
17 90 88 89 273 276,1 269,9 1,1 1,1
18 116 111 114 273 277,8 265,8 1,8 2,6
19 118 115 121 273 266,2 259,5 2,5 5,0
20 114 110 113 273 275,4 265,8 0,9 2,7
21 81 79 80 273 276,4 269,6 1,3 1,2
22 160 158 162 273 269,6 266,3 1,2 2,5
23 155 153 153 273 276,6 273,0 1,3 0,0
24 121 119 121 273 273,0 268,5 0,0 1,7
25 86 84 87 273 269,9 263,6 1,1 3,4
26 103 98 104 273 270,4 257,3 1,0 5,8
27 121 116 124 273 266,4 255,4 2,4 6,5
28 79 74 81 273 266,3 249,4 2,5 8,6
29 67 66 67 273 273,0 268,9 0,0 1,5
30 61 60 62 273 268,6 264,2 1,6 3,2
31 77 72 77 273 273,0 255,3 0,0 6,5
32 81 80 81 273 273,0 269,6 0,0 1,2
33 72 72 73 273 269,3 269,3 1,4 1,4
34 94 93 96 273 267,3 264,5 2,1 3,1
35 94 93 96 273 267,3 264,5 2,1 3,1
36 104 101 105 273 270,4 262,6 1,0 3,8
37 82 80 83 273 269,7 263,1 1,2 3,6
38 93 86 93 273 273,0 252,5 0,0 7,5
39 91 88 91 273 273,0 264,0 0,0 3,3
40 99 94 99 273 273,0 259,2 0,0 5,1
41 91 86 90 273 276,0 260,9 1,1 4,4
42 75 71 75 273 273,0 258,4 0,0 5,3
43 297 292 298 273 272,1 267,5 0,3 2,0
44 366 364 360 273 277,6 276,0 1,7 1,1
45 348 348 352 273 269,9 269,9 1,1 1,1
46 405 401 417 273 265,1 262,5 2,9 3,8
47 375 370 378 273 270,8 267,2 0,8 2,1
48 416 409 420 273 270,4 265,9 1,0 2,6
49 330 328 337 273 267,3 265,7 2,1 2,7
50 385 377 396 273 265,4 259,9 2,8 4,8
51 352 344 342 273 281,0 274,6 2,9 0,6
52 213 212 217 273 268,0 266,7 1,8 2,3
53 286 285 284 273 274,9 274,0 0,7 0,4
54 221 221 223 273 270,6 270,6 0,9 0,9
55 290 290 283 273 279,8 279,8 2,5 2,5
56 324 323 324 273 273,0 272,2 0,0 0,3

13. Anhang 184
57 367 366 363 273 276,0 275,3 1,1 0,8
58 435 432 436 273 272,4 270,5 0,2 0,9
59 414 409 413 273 273,7 270,4 0,2 1,0
60 275 279 277 273 271,0 275,0 0,7 0,7
61 350 345 347 273 275,4 271,4 0,9 0,6
62 405 400 399 273 277,1 273,7 1,5 0,3
63 359 362 350 273 280,0 282,4 2,6 3,4
64 390 388 387 273 275,1 273,7 0,8 0,3
65 341 339 337 273 276,2 274,6 1,2 0,6
66 353 349 343 273 281,0 277,8 2,9 1,7
67 359 350 352 273 278,4 271,4 2,0 0,6
68 351 343 341 273 281,0 274,6 2,9 0,6
69 277 280 272 273 278,0 281,0 1,8 2,9
70 506 505 496 273 278,5 278,0 2,0 1,8
71 323 317 327 273 269,7 264,7 1,2 3,1
72 493 492 489 273 275,2 274,7 0,8 0,6
73 258 253 251 273 280,6 275,2 2,8 0,8
74 362 362 359 273 275,3 275,3 0,8 0,8
75 357 353 355 273 274,5 271,5 0,6 0,6
76 349 342 341 273 279,4 273,8 2,3 0,3
77 425 411 419 273 276,9 267,8 1,4 1,9
78 391 381 382 273 279,4 272,3 2,4 0,3
79 436 421 432 273 275,5 266,0 0,9 2,5
80 360 357 352 273 279,2 276,9 2,3 1,4
81 481 446 484 273 271,3 251,6 0,6 7,9
82 581 561 573 273 276,8 267,3 1,4 2,1
83 577 565 573 273 274,9 269,2 0,7 1,4
84 397 392 394 273 275,1 271,6 0,8 0,5
85 507 503 512 273 270,3 268,2 1,0 1,8
86 490 490 492 273 271,9 271,9 0,4 0,4
87 543 540 535 273 277,1 275,6 1,5 0,9
88 339 332 340 273 272,2 266,6 0,3 2,4
89 340 342 342 273 271,4 273,0 0,6 0,0
90 322 321 322 273 273,0 272,2 0,0 0,3
91 317 313 318 273 272,1 268,7 0,3 1,6
92 302 300 308 273 267,7 265,9 1,9 2,6
93 361 348 368 273 267,8 258,2 1,9 5,4
94 295 288 292 273 275,8 269,3 1,0 1,4
95 307 302 313 273 267,8 263,4 1,9 3,5
96 401 391 411 273 266,4 259,7 2,4 4,9
97 343 339 350 273 267,5 264,4 2,0 3,1
98 407 402 419 273 265,2 261,9 2,9 4,1
99 233 219 238 273 267,3 251,2 2,1 8,0
100 279 277 284 273 268,2 266,3 1,8 2,5
Ø Fehler 1,27 2,41
13.8. Vergleich der automatischen Kernzählung auf Basis eines
KNN zur Referenz
Tabelle 63: Vergleich der automatischen Kernzählung auf Basis eines KNN mit der Referenz-
zählung; AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte Refe-
renzzählung; AZ Norm: Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF
Norm – AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm
AZ Norm
ERR (%)
Nr AZ REF REF Norm
AZ Norm
ERR (%)
1 198 232 273 233,0 14,7 51 287 342 273 229,1 16,1
2 330 369 273 244,1 10,6 52 178 217 273 223,9 18,0
3 211 250 273 230,4 15,6 53 256 284 273 246,1 9,9
4 176 199 273 241,4 11,6 54 193 223 273 236,3 13,5
5 214 240 273 243,4 10,8 55 251 283 273 242,1 11,3
6 179 199 273 245,6 10,1 56 293 324 273 246,9 9,6
7 198 215 273 251,4 7,9 57 310 363 273 233,1 14,6

13. Anhang 185
8 186 210 273 241,8 11,4 58 382 436 273 239,2 12,4
9 189 215 273 240,0 12,1 59 366 413 273 241,9 11,4
10 256 280 273 249,6 8,6 60 241 277 273 237,5 13,0
11 222 252 273 240,5 11,9 61 307 347 273 241,5 11,5
12 185 229 273 220,5 19,2 62 343 399 273 234,7 14,0
13 198 248 273 218,0 20,2 63 318 350 273 248,0 9,1
14 66 79 273 228,1 16,5 64 353 387 273 249,0 8,8
15 83 109 273 207,9 23,9 65 299 337 273 242,2 11,3
16 75 98 273 208,9 23,5 66 287 343 273 228,4 16,3
17 74 89 273 227,0 16,9 67 305 352 273 236,5 13,4
18 101 114 273 241,9 11,4 68 302 341 273 241,8 11,4
19 108 121 273 243,7 10,7 69 219 272 273 219,8 19,5
20 104 113 273 251,3 8,0 70 454 496 273 249,9 8,5
21 67 80 273 228,6 16,3 71 292 327 273 243,8 10,7
22 134 162 273 225,8 17,3 72 449 489 273 250,7 8,2
23 121 153 273 215,9 20,9 73 203 251 273 220,8 19,1
24 104 121 273 234,6 14,0 74 308 359 273 234,2 14,2
25 67 87 273 210,2 23,0 75 310 355 273 238,4 12,7
26 85 104 273 223,1 18,3 76 293 341 273 234,6 14,1
27 98 124 273 215,8 21,0 77 376 419 273 245,0 10,3
28 59 81 273 198,9 27,2 78 344 382 273 245,8 9,9
29 50 67 273 203,7 25,4 79 392 432 273 247,7 9,3
30 48 62 273 211,4 22,6 80 307 352 273 238,1 12,8
31 64 77 273 226,9 16,9 81 411 484 273 231,8 15,1
32 63 81 273 212,3 22,2 82 506 573 273 241,1 11,7
33 56 73 273 209,4 23,3 83 495 573 273 235,8 13,6
34 88 96 273 250,3 8,3 84 331 394 273 229,3 16,0
35 85 96 273 241,7 11,5 85 449 512 273 239,4 12,3
36 93 105 273 241,8 11,4 86 450 492 273 249,7 8,5
37 70 83 273 230,2 15,7 87 462 535 273 235,7 13,6
38 81 93 273 237,8 12,9 88 302 340 273 242,5 11,2
39 77 91 273 231,0 15,4 89 297 342 273 237,1 13,2
40 82 99 273 226,1 17,2 90 268 322 273 227,2 16,8
41 76 90 273 230,5 15,6 91 254 318 273 218,1 20,1
42 61 75 273 222,0 18,7 92 282 308 273 250,0 8,4
43 266 298 273 243,7 10,7 93 308 368 273 228,5 16,3
44 313 360 273 237,4 13,1 94 251 292 273 234,7 14,0
45 323 352 273 250,5 8,2 95 270 313 273 235,5 13,7
46 380 417 273 248,8 8,9 96 344 411 273 228,5 16,3
47 337 378 273 243,4 10,8 97 302 350 273 235,6 13,7
48 369 420 273 239,9 12,1 98 375 419 273 244,3 10,5
49 300 337 273 243,0 11,0 99 200 238 273 229,4 16,0
50 356 396 273 245,4 10,1 100 229 284 273 220,1 19,4
Ø Fehler 14,1
13.9 Vergleich der automatischen Kernzählung auf Basis des HB-
Algorithmus mit der Referenzzählung
Tabelle 64: Vergleich der automatischen L929-Kernzählung auf Basis des HB-Algorithmus mit
der Referenzzählung; AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm: Nor-
mierte Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR: Relativer Fehler
(((REF Norm – AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm
AZ Norm
ERR Nr AZ REF REF Norm
AZ Norm
ERR
1 227 232 273 267,1 2,2 51 340 342 273 271,4 0,6
2 366 369 273 270,8 0,8 52 209 217 273 262,9 3,7
3 241 250 273 263,2 3,6 53 281 284 273 270,1 1,1
4 194 199 273 266,1 2,5 54 218 223 273 266,9 2,2
5 234 240 273 266,2 2,5 55 285 283 273 274,9 0,7
6 197 199 273 270,3 1,0 56 319 324 273 268,8 1,5
7 212 215 273 269,2 1,4 57 365 363 273 274,5 0,6
8 198 210 273 257,4 5,7 58 417 436 273 261,1 4,4

13. Anhang 186
9 209 215 273 265,4 2,8 59 404 413 273 267,1 2,2
10 270 280 273 263,3 3,6 60 270 277 273 266,1 2,5
11 255 252 273 276,3 1,2 61 341 347 273 268,3 1,7
12 221 229 273 263,5 3,5 62 393 399 273 268,9 1,5
13 246 248 273 270,8 0,8 63 356 350 273 277,7 1,7
14 75 79 273 259,2 5,1 64 385 387 273 271,6 0,5
15 104 109 273 260,5 4,6 65 336 337 273 272,2 0,3
16 93 98 273 259,1 5,1 66 345 343 273 274,6 0,6
17 86 89 273 263,8 3,4 67 345 352 273 267,6 2,0
18 114 114 273 273,0 0,0 68 336 341 273 269,0 1,5
19 115 121 273 259,5 5,0 69 268 272 273 269,0 1,5
20 108 113 273 260,9 4,4 70 500 496 273 275,2 0,8
21 78 80 273 266,2 2,5 71 309 327 273 258,0 5,5
22 159 162 273 267,9 1,9 72 484 489 273 270,2 1,0
23 153 153 273 273,0 0,0 73 250 251 273 271,9 0,4
24 118 121 273 266,2 2,5 74 355 359 273 270,0 1,1
25 85 87 273 266,7 2,3 75 350 355 273 269,2 1,4
26 97 104 273 254,6 6,7 76 337 341 273 269,8 1,2
27 119 124 273 262,0 4,0 77 410 419 273 267,1 2,1
28 74 81 273 249,4 8,6 78 375 382 273 268,0 1,8
29 63 67 273 256,7 6,0 79 418 432 273 264,2 3,2
30 60 62 273 264,2 3,2 80 353 352 273 273,8 0,3
31 70 77 273 248,2 9,1 81 445 484 273 251,0 8,1
32 79 81 273 266,3 2,5 82 562 573 273 267,8 1,9
33 71 73 273 265,5 2,7 83 562 573 273 267,8 1,9
34 92 96 273 261,6 4,2 84 386 394 273 267,5 2,0
35 91 96 273 258,8 5,2 85 500 512 273 266,6 2,3
36 100 105 273 260,0 4,8 86 483 492 273 268,0 1,8
37 78 83 273 256,6 6,0 87 536 535 273 273,5 0,2
38 84 93 273 246,6 9,7 88 330 340 273 265,0 2,9
39 87 91 273 261,0 4,4 89 338 342 273 269,8 1,2
40 92 99 273 253,7 7,1 90 317 322 273 268,8 1,6
41 85 90 273 257,8 5,6 91 305 318 273 261,8 4,1
42 70 75 273 254,8 6,7 92 295 308 273 261,5 4,2
43 291 298 273 266,6 2,3 93 346 368 273 256,7 6,0
44 360 360 273 273,0 0,0 94 284 292 273 265,5 2,7
45 345 352 273 267,6 2,0 95 301 313 273 262,5 3,8
46 396 417 273 259,3 5,0 96 393 411 273 261,0 4,4
47 367 378 273 265,1 2,9 97 337 350 273 262,9 3,7
48 405 420 273 263,3 3,6 98 396 419 273 258,0 5,5
49 327 337 273 264,9 3,0 99 218 238 273 250,1 8,4
50 375 396 273 258,5 5,3 100 272 284 273 261,5 4,2
Ø Fehler 3,09
Tabelle 65: Vergleich der automatischen MC3T3-Kernzählung auf Basis des HB-Algorithmus
mit der Referenzzählung; AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm:
Normierte Referenzzählung; AZ Norm: Normierte automatische Zellzählung; ERR: Relativer
Fehler (((REF Norm – AZ Norm) / REF Norm) * 100).
Nr AZ REF REF Norm
AZ Norm
ERR Nr AZ REF REF Norm
AZ Norm
ERR
1 73 85 73 62,7 14,1 51 66 66 73 73,0 0,0
2 94 107 73 64,1 12,1 52 92 96 73 70,0 4,2
3 77 82 73 68,5 6,1 53 74 75 73 72,0 1,3
4 88 90 73 71,4 2,2 54 128 140 73 66,7 8,6
5 63 65 73 70,8 3,1 55 58 63 73 67,2 7,9
6 87 90 73 70,6 3,3 56 135 139 73 70,9 2,9
7 86 95 73 66,1 9,5 57 66 69 73 69,8 4,3
8 99 104 73 69,5 4,8 58 159 171 73 67,9 7,0
9 94 99 73 69,3 5,1 59 52 56 73 67,8 7,1
10 62 67 73 67,6 7,5 60 48 51 73 68,7 5,9
11 87 96 73 66,2 9,4 61 26 32 73 59,3 18,8
12 77 82 73 68,5 6,1 62 20 22 73 66,4 9,1
13 76 86 73 64,5 11,6 63 57 59 73 70,5 3,4
14 72 84 73 62,6 14,3 64 37 39 73 69,3 5,1
15 71 82 73 63,2 13,4 65 47 50 73 68,6 6,0
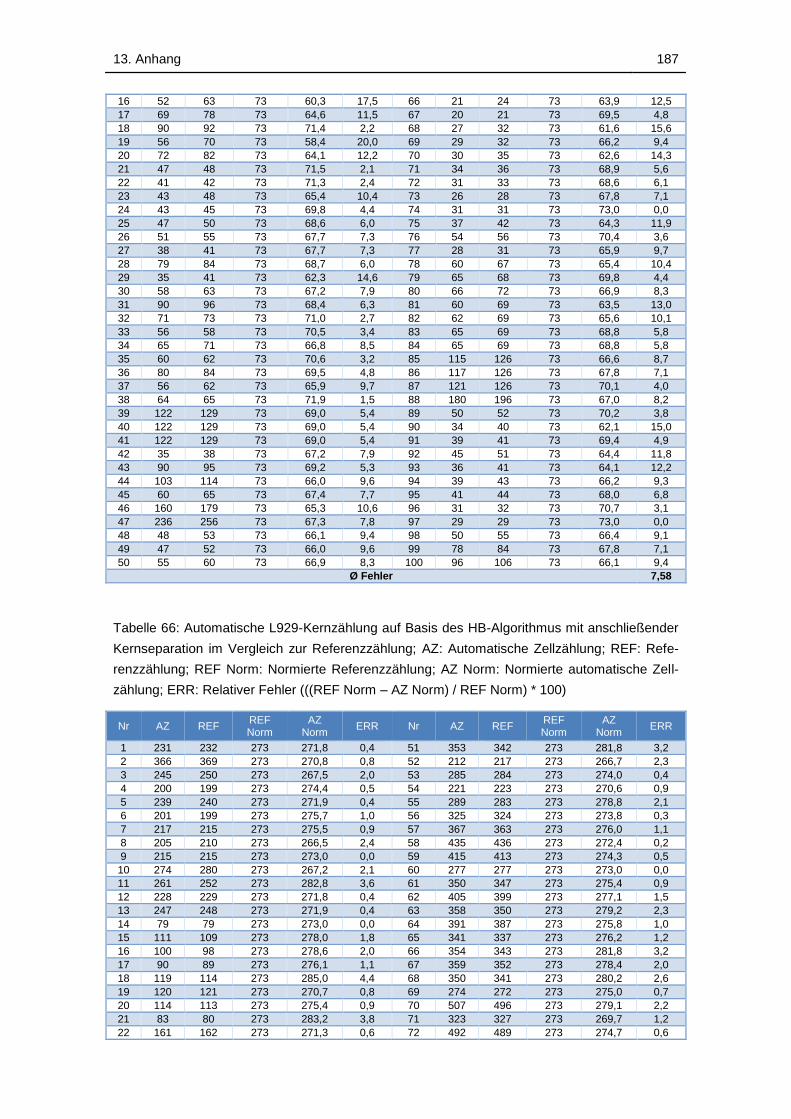
13. Anhang 187
16 52 63 73 60,3 17,5 66 21 24 73 63,9 12,5
17 69 78 73 64,6 11,5 67 20 21 73 69,5 4,8
18 90 92 73 71,4 2,2 68 27 32 73 61,6 15,6
19 56 70 73 58,4 20,0 69 29 32 73 66,2 9,4
20 72 82 73 64,1 12,2 70 30 35 73 62,6 14,3
21 47 48 73 71,5 2,1 71 34 36 73 68,9 5,6
22 41 42 73 71,3 2,4 72 31 33 73 68,6 6,1
23 43 48 73 65,4 10,4 73 26 28 73 67,8 7,1
24 43 45 73 69,8 4,4 74 31 31 73 73,0 0,0
25 47 50 73 68,6 6,0 75 37 42 73 64,3 11,9
26 51 55 73 67,7 7,3 76 54 56 73 70,4 3,6
27 38 41 73 67,7 7,3 77 28 31 73 65,9 9,7
28 79 84 73 68,7 6,0 78 60 67 73 65,4 10,4
29 35 41 73 62,3 14,6 79 65 68 73 69,8 4,4
30 58 63 73 67,2 7,9 80 66 72 73 66,9 8,3
31 90 96 73 68,4 6,3 81 60 69 73 63,5 13,0
32 71 73 73 71,0 2,7 82 62 69 73 65,6 10,1
33 56 58 73 70,5 3,4 83 65 69 73 68,8 5,8
34 65 71 73 66,8 8,5 84 65 69 73 68,8 5,8
35 60 62 73 70,6 3,2 85 115 126 73 66,6 8,7
36 80 84 73 69,5 4,8 86 117 126 73 67,8 7,1
37 56 62 73 65,9 9,7 87 121 126 73 70,1 4,0
38 64 65 73 71,9 1,5 88 180 196 73 67,0 8,2
39 122 129 73 69,0 5,4 89 50 52 73 70,2 3,8
40 122 129 73 69,0 5,4 90 34 40 73 62,1 15,0
41 122 129 73 69,0 5,4 91 39 41 73 69,4 4,9
42 35 38 73 67,2 7,9 92 45 51 73 64,4 11,8
43 90 95 73 69,2 5,3 93 36 41 73 64,1 12,2
44 103 114 73 66,0 9,6 94 39 43 73 66,2 9,3
45 60 65 73 67,4 7,7 95 41 44 73 68,0 6,8
46 160 179 73 65,3 10,6 96 31 32 73 70,7 3,1
47 236 256 73 67,3 7,8 97 29 29 73 73,0 0,0
48 48 53 73 66,1 9,4 98 50 55 73 66,4 9,1
49 47 52 73 66,0 9,6 99 78 84 73 67,8 7,1
50 55 60 73 66,9 8,3 100 96 106 73 66,1 9,4
Ø Fehler 7,58
Tabelle 66: Automatische L929-Kernzählung auf Basis des HB-Algorithmus mit anschließender
Kernseparation im Vergleich zur Referenzzählung; AZ: Automatische Zellzählung; REF: Refe-
renzzählung; REF Norm: Normierte Referenzzählung; AZ Norm: Normierte automatische Zell-
zählung; ERR: Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm
AZ Norm
ERR Nr AZ REF REF Norm
AZ Norm
ERR
1 231 232 273 271,8 0,4 51 353 342 273 281,8 3,2
2 366 369 273 270,8 0,8 52 212 217 273 266,7 2,3
3 245 250 273 267,5 2,0 53 285 284 273 274,0 0,4
4 200 199 273 274,4 0,5 54 221 223 273 270,6 0,9
5 239 240 273 271,9 0,4 55 289 283 273 278,8 2,1
6 201 199 273 275,7 1,0 56 325 324 273 273,8 0,3
7 217 215 273 275,5 0,9 57 367 363 273 276,0 1,1
8 205 210 273 266,5 2,4 58 435 436 273 272,4 0,2
9 215 215 273 273,0 0,0 59 415 413 273 274,3 0,5
10 274 280 273 267,2 2,1 60 277 277 273 273,0 0,0
11 261 252 273 282,8 3,6 61 350 347 273 275,4 0,9
12 228 229 273 271,8 0,4 62 405 399 273 277,1 1,5
13 247 248 273 271,9 0,4 63 358 350 273 279,2 2,3
14 79 79 273 273,0 0,0 64 391 387 273 275,8 1,0
15 111 109 273 278,0 1,8 65 341 337 273 276,2 1,2
16 100 98 273 278,6 2,0 66 354 343 273 281,8 3,2
17 90 89 273 276,1 1,1 67 359 352 273 278,4 2,0
18 119 114 273 285,0 4,4 68 350 341 273 280,2 2,6
19 120 121 273 270,7 0,8 69 274 272 273 275,0 0,7
20 114 113 273 275,4 0,9 70 507 496 273 279,1 2,2
21 83 80 273 283,2 3,8 71 323 327 273 269,7 1,2
22 161 162 273 271,3 0,6 72 492 489 273 274,7 0,6
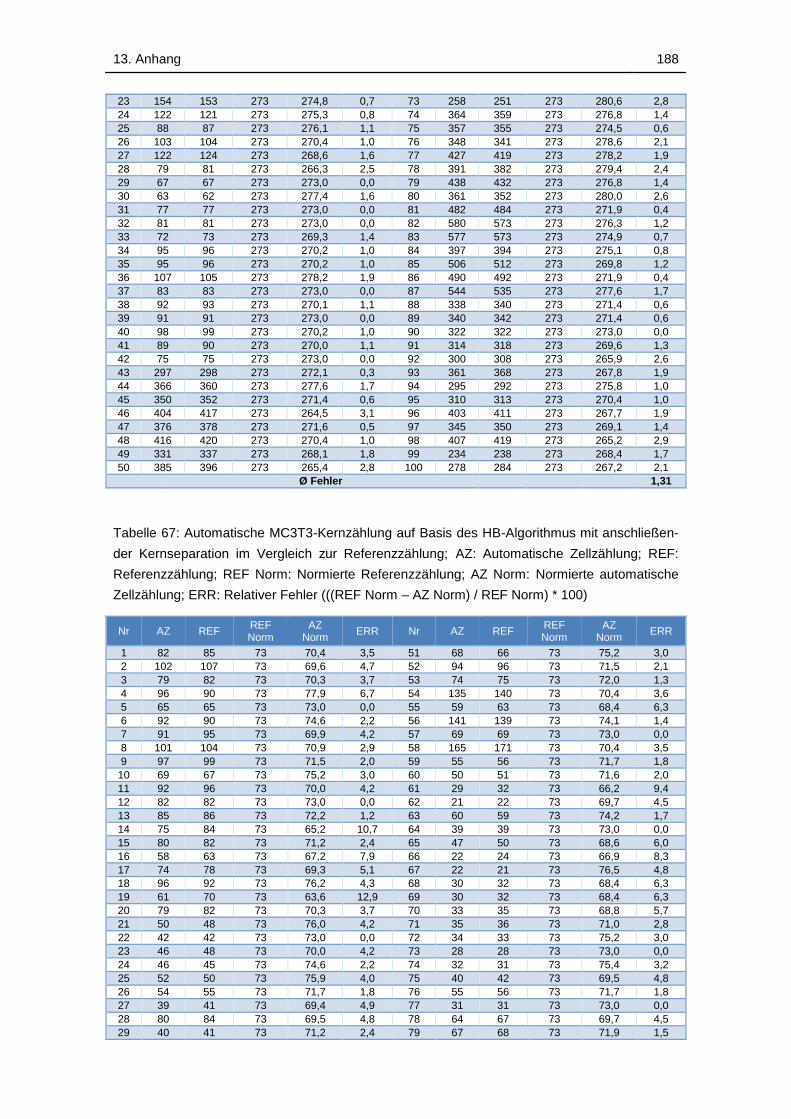
13. Anhang 188
23 154 153 273 274,8 0,7 73 258 251 273 280,6 2,8
24 122 121 273 275,3 0,8 74 364 359 273 276,8 1,4
25 88 87 273 276,1 1,1 75 357 355 273 274,5 0,6
26 103 104 273 270,4 1,0 76 348 341 273 278,6 2,1
27 122 124 273 268,6 1,6 77 427 419 273 278,2 1,9
28 79 81 273 266,3 2,5 78 391 382 273 279,4 2,4
29 67 67 273 273,0 0,0 79 438 432 273 276,8 1,4
30 63 62 273 277,4 1,6 80 361 352 273 280,0 2,6
31 77 77 273 273,0 0,0 81 482 484 273 271,9 0,4
32 81 81 273 273,0 0,0 82 580 573 273 276,3 1,2
33 72 73 273 269,3 1,4 83 577 573 273 274,9 0,7
34 95 96 273 270,2 1,0 84 397 394 273 275,1 0,8
35 95 96 273 270,2 1,0 85 506 512 273 269,8 1,2
36 107 105 273 278,2 1,9 86 490 492 273 271,9 0,4
37 83 83 273 273,0 0,0 87 544 535 273 277,6 1,7
38 92 93 273 270,1 1,1 88 338 340 273 271,4 0,6
39 91 91 273 273,0 0,0 89 340 342 273 271,4 0,6
40 98 99 273 270,2 1,0 90 322 322 273 273,0 0,0
41 89 90 273 270,0 1,1 91 314 318 273 269,6 1,3
42 75 75 273 273,0 0,0 92 300 308 273 265,9 2,6
43 297 298 273 272,1 0,3 93 361 368 273 267,8 1,9
44 366 360 273 277,6 1,7 94 295 292 273 275,8 1,0
45 350 352 273 271,4 0,6 95 310 313 273 270,4 1,0
46 404 417 273 264,5 3,1 96 403 411 273 267,7 1,9
47 376 378 273 271,6 0,5 97 345 350 273 269,1 1,4
48 416 420 273 270,4 1,0 98 407 419 273 265,2 2,9
49 331 337 273 268,1 1,8 99 234 238 273 268,4 1,7
50 385 396 273 265,4 2,8 100 278 284 273 267,2 2,1
Ø Fehler 1,31
Tabelle 67: Automatische MC3T3-Kernzählung auf Basis des HB-Algorithmus mit anschließen-
der Kernseparation im Vergleich zur Referenzzählung; AZ: Automatische Zellzählung; REF:
Referenzzählung; REF Norm: Normierte Referenzzählung; AZ Norm: Normierte automatische
Zellzählung; ERR: Relativer Fehler (((REF Norm – AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm
AZ Norm
ERR Nr AZ REF REF Norm
AZ Norm
ERR
1 82 85 73 70,4 3,5 51 68 66 73 75,2 3,0
2 102 107 73 69,6 4,7 52 94 96 73 71,5 2,1
3 79 82 73 70,3 3,7 53 74 75 73 72,0 1,3
4 96 90 73 77,9 6,7 54 135 140 73 70,4 3,6
5 65 65 73 73,0 0,0 55 59 63 73 68,4 6,3
6 92 90 73 74,6 2,2 56 141 139 73 74,1 1,4
7 91 95 73 69,9 4,2 57 69 69 73 73,0 0,0
8 101 104 73 70,9 2,9 58 165 171 73 70,4 3,5
9 97 99 73 71,5 2,0 59 55 56 73 71,7 1,8
10 69 67 73 75,2 3,0 60 50 51 73 71,6 2,0
11 92 96 73 70,0 4,2 61 29 32 73 66,2 9,4
12 82 82 73 73,0 0,0 62 21 22 73 69,7 4,5
13 85 86 73 72,2 1,2 63 60 59 73 74,2 1,7
14 75 84 73 65,2 10,7 64 39 39 73 73,0 0,0
15 80 82 73 71,2 2,4 65 47 50 73 68,6 6,0
16 58 63 73 67,2 7,9 66 22 24 73 66,9 8,3
17 74 78 73 69,3 5,1 67 22 21 73 76,5 4,8
18 96 92 73 76,2 4,3 68 30 32 73 68,4 6,3
19 61 70 73 63,6 12,9 69 30 32 73 68,4 6,3
20 79 82 73 70,3 3,7 70 33 35 73 68,8 5,7
21 50 48 73 76,0 4,2 71 35 36 73 71,0 2,8
22 42 42 73 73,0 0,0 72 34 33 73 75,2 3,0
23 46 48 73 70,0 4,2 73 28 28 73 73,0 0,0
24 46 45 73 74,6 2,2 74 32 31 73 75,4 3,2
25 52 50 73 75,9 4,0 75 40 42 73 69,5 4,8
26 54 55 73 71,7 1,8 76 55 56 73 71,7 1,8
27 39 41 73 69,4 4,9 77 31 31 73 73,0 0,0
28 80 84 73 69,5 4,8 78 64 67 73 69,7 4,5
29 40 41 73 71,2 2,4 79 67 68 73 71,9 1,5

13. Anhang 189
30 61 63 73 70,7 3,2 80 69 72 73 70,0 4,2
31 92 96 73 70,0 4,2 81 67 69 73 70,9 2,9
32 73 73 73 73,0 0,0 82 70 69 73 74,1 1,4
33 59 58 73 74,3 1,7 83 73 69 73 77,2 5,8
34 67 71 73 68,9 5,6 84 73 69 73 77,2 5,8
35 60 62 73 70,6 3,2 85 124 126 73 71,8 1,6
36 81 84 73 70,4 3,6 86 126 126 73 73,0 0,0
37 58 62 73 68,3 6,5 87 129 126 73 74,7 2,4
38 67 65 73 75,2 3,1 88 196 196 73 73,0 0,0
39 125 129 73 70,7 3,1 89 50 52 73 70,2 3,8
40 125 129 73 70,7 3,1 90 39 40 73 71,2 2,5
41 125 129 73 70,7 3,1 91 42 41 73 74,8 2,4
42 36 38 73 69,2 5,3 92 51 51 73 73,0 0,0
43 94 95 73 72,2 1,1 93 39 41 73 69,4 4,9
44 110 114 73 70,4 3,5 94 42 43 73 71,3 2,3
45 65 65 73 73,0 0,0 95 42 44 73 69,7 4,5
46 170 179 73 69,3 5,0 96 33 32 73 75,3 3,1
47 247 256 73 70,4 3,5 97 30 29 73 75,5 3,4
48 50 53 73 68,9 5,7 98 54 55 73 71,7 1,8
49 48 52 73 67,4 7,7 99 81 84 73 70,4 3,6
50 57 60 73 69,4 5,0 100 101 106 73 69,6 4,7
Ø Fehler 3,52
13.10 Zellzählung mit der Wachstumssimulation im Vergleich zur
Referenzzählung
Tabelle 68: Vergleich der L929-Zellzählung (1) sowie der MC3T3-Zellzählung (2) mit der
Wachstumssimulation zur Referenzzählung; AZ 1: Automatische L929-Zellzählung; AZ 2: Au-
tomatische MC3T3-Zellzählung; REF: Referenzzählung; REF Norm: Normierte Referenzzäh-
lung; AZ 1 Norm: Normierte automatische L929-Zellzählung; AZ 2 Norm: Normierte automati-
sche MC3T3-Zellzählung; ERR 1: Relativer Fehler für den Typ L929 (((REF Norm – AZ Norm) /
REF Norm) * 100); ERR 2: Relativer Fehler für den Typ MC3T3
Nr AZ 1 REF 1 AZ 2 REF 2 REF 1 Norm
REF 2 Norm
AZ 1 Norm
AZ 2 Norm
ERR 1 [%]
ERR 2 [%]
1 231 232 82 85 273 73 271,8 70,4 0,4 3,5
2 366 369 102 107 273 73 270,8 69,6 0,8 4,7
3 245 250 79 82 273 73 267,5 70,3 2,0 3,7
4 200 199 96 90 273 73 274,4 77,9 0,5 6,7
5 239 240 65 65 273 73 271,9 73,0 0,4 0,0
6 201 199 92 90 273 73 275,7 74,6 1,0 2,2
7 217 215 91 95 273 73 275,5 69,9 0,9 4,2
8 205 210 101 104 273 73 266,5 70,9 2,4 2,9
9 215 215 97 99 273 73 273,0 71,5 0,0 2,0
10 274 280 69 67 273 73 267,2 75,2 2,1 3,0
11 261 252 92 96 273 73 282,8 70,0 3,6 4,2
12 228 229 82 82 273 73 271,8 73,0 0,4 0,0
13 247 248 85 86 273 73 271,9 72,2 0,4 1,2
14 79 79 75 84 273 73 273,0 65,2 0,0 10,7
15 111 109 80 82 273 73 278,0 71,2 1,8 2,4
16 100 98 58 63 273 73 278,6 67,2 2,0 7,9
17 90 89 74 78 273 73 276,1 69,3 1,1 5,1
18 119 114 96 92 273 73 285,0 76,2 4,4 4,3
19 120 121 61 70 273 73 270,7 63,6 0,8 12,9
20 114 113 79 82 273 73 275,4 70,3 0,9 3,7
21 83 80 50 48 273 73 283,2 76,0 3,8 4,2
22 161 162 42 42 273 73 271,3 73,0 0,6 0,0
23 154 153 46 48 273 73 274,8 70,0 0,7 4,2
24 122 121 46 45 273 73 275,3 74,6 0,8 2,2
25 88 87 52 50 273 73 276,1 75,9 1,1 4,0
26 103 104 54 55 273 73 270,4 71,7 1,0 1,8
27 122 124 39 41 273 73 268,6 69,4 1,6 4,9

13. Anhang 190
28 79 81 80 84 273 73 266,3 69,5 2,5 4,8
29 67 67 40 41 273 73 273,0 71,2 0,0 2,4
30 63 62 61 63 273 73 277,4 70,7 1,6 3,2
31 77 77 92 96 273 73 273,0 70,0 0,0 4,2
32 81 81 73 73 273 73 273,0 73,0 0,0 0,0
33 72 73 59 58 273 73 269,3 74,3 1,4 1,7
34 95 96 67 71 273 73 270,2 68,9 1,0 5,6
35 95 96 60 62 273 73 270,2 70,6 1,0 3,2
36 107 105 81 84 273 73 278,2 70,4 1,9 3,6
37 83 83 58 62 273 73 273,0 68,3 0,0 6,5
38 92 93 67 65 273 73 270,1 75,2 1,1 3,1
39 91 91 125 129 273 73 273,0 70,7 0,0 3,1
40 98 99 125 129 273 73 270,2 70,7 1,0 3,1
41 89 90 125 129 273 73 270,0 70,7 1,1 3,1
42 75 75 36 38 273 73 273,0 69,2 0,0 5,3
43 297 298 94 95 273 73 272,1 72,2 0,3 1,1
44 366 360 110 114 273 73 277,6 70,4 1,7 3,5
45 350 352 65 65 273 73 271,4 73,0 0,6 0,0
46 404 417 170 179 273 73 264,5 69,3 3,1 5,0
47 376 378 247 256 273 73 271,6 70,4 0,5 3,5
48 416 420 50 53 273 73 270,4 68,9 1,0 5,7
49 331 337 48 52 273 73 268,1 67,4 1,8 7,7
50 385 396 57 60 273 73 265,4 69,4 2,8 5,0
51 353 342 68 66 273 73 281,8 75,2 3,2 3,0
52 212 217 94 96 273 73 266,7 71,5 2,3 2,1
53 285 284 74 75 273 73 274,0 72,0 0,4 1,3
54 221 223 135 140 273 73 270,6 70,4 0,9 3,6
55 289 283 59 63 273 73 278,8 68,4 2,1 6,3
56 325 324 141 139 273 73 273,8 74,1 0,3 1,4
57 367 363 69 69 273 73 276,0 73,0 1,1 0,0
58 435 436 165 171 273 73 272,4 70,4 0,2 3,5
59 415 413 55 56 273 73 274,3 71,7 0,5 1,8
60 277 277 50 51 273 73 273,0 71,6 0,0 2,0
61 350 347 29 32 273 73 275,4 66,2 0,9 9,4
62 405 399 21 22 273 73 277,1 69,7 1,5 4,5
63 358 350 60 59 273 73 279,2 74,2 2,3 1,7
64 391 387 39 39 273 73 275,8 73,0 1,0 0,0
65 341 337 47 50 273 73 276,2 68,6 1,2 6,0
66 354 343 22 24 273 73 281,8 66,9 3,2 8,3
67 359 352 22 21 273 73 278,4 76,5 2,0 4,8
68 350 341 30 32 273 73 280,2 68,4 2,6 6,3
69 274 272 30 32 273 73 275,0 68,4 0,7 6,3
70 507 496 33 35 273 73 279,1 68,8 2,2 5,7
71 323 327 35 36 273 73 269,7 71,0 1,2 2,8
72 492 489 34 33 273 73 274,7 75,2 0,6 3,0
73 258 251 28 28 273 73 280,6 73,0 2,8 0,0
74 364 359 32 31 273 73 276,8 75,4 1,4 3,2
75 357 355 40 42 273 73 274,5 69,5 0,6 4,8
76 348 341 55 56 273 73 278,6 71,7 2,1 1,8
77 427 419 31 31 273 73 278,2 73,0 1,9 0,0
78 391 382 64 67 273 73 279,4 69,7 2,4 4,5
79 438 432 67 68 273 73 276,8 71,9 1,4 1,5
80 361 352 69 72 273 73 280,0 70,0 2,6 4,2
81 482 484 67 69 273 73 271,9 70,9 0,4 2,9
82 580 573 70 69 273 73 276,3 74,1 1,2 1,4
83 577 573 73 69 273 73 274,9 77,2 0,7 5,8
84 397 394 73 69 273 73 275,1 77,2 0,8 5,8
85 506 512 124 126 273 73 269,8 71,8 1,2 1,6
86 490 492 126 126 273 73 271,9 73,0 0,4 0,0
87 544 535 129 126 273 73 277,6 74,7 1,7 2,4
88 338 340 196 196 273 73 271,4 73,0 0,6 0,0
89 340 342 50 52 273 73 271,4 70,2 0,6 3,8
90 322 322 39 40 273 73 273,0 71,2 0,0 2,5
91 314 318 42 41 273 73 269,6 74,8 1,3 2,4
92 300 308 51 51 273 73 265,9 73,0 2,6 0,0
93 361 368 39 41 273 73 267,8 69,4 1,9 4,9
94 295 292 42 43 273 73 275,8 71,3 1,0 2,3
95 310 313 42 44 273 73 270,4 69,7 1,0 4,5
96 403 411 33 32 273 73 267,7 75,3 1,9 3,1

13. Anhang 191
97 345 350 30 29 273 73 269,1 75,5 1,4 3,4
98 407 419 54 55 273 73 265,2 71,7 2,9 1,8
99 234 238 81 84 273 73 268,4 70,4 1,7 3,6
100 278 284 101 106 273 73 267,2 69,6 2,1 4,7
Ø Fehler 1,31 3,50
13.11 Zellzählung mit dem Algorithmus aus Kap. 6.6 und Vergleich
mit der Referenzzählung
Tabelle 69: L929-Zellzählung auf Basis des Algorithmus in Kap. 6.6 im Vergleich zur Referenz-
zählung; AZ: Automatische Zellzählung; REF: Referenzzählung; REF Norm: Normierte Refe-
renzzählung; AZ Norm: Normierte automatische Zellzählung; ERR: Relativer Fehler (((REF
Norm – AZ Norm) / REF Norm) * 100)
Nr AZ REF REF Norm
AZ Norm
ERR [%]
Nr AZ REF REF Norm
AZ Norm
ERR [%]
1 229 232 273 269,5 1,3 51 327 342 273 261,0 4,4
2 347 369 273 256,7 6,0 52 209 217 273 262,9 3,7
3 241 250 273 263,2 3,6 53 280 284 273 269,2 1,4
4 195 199 273 267,5 2,0 54 212 223 273 259,5 4,9
5 236 240 273 268,5 1,7 55 276 283 273 266,2 2,5
6 194 199 273 266,1 2,5 56 313 324 273 263,7 3,4
7 218 215 273 276,8 1,4 57 355 363 273 267,0 2,2
8 205 210 273 266,5 2,4 58 421 436 273 263,6 3,4
9 209 215 273 265,4 2,8 59 400 413 273 264,4 3,1
10 271 280 273 264,2 3,2 60 268 277 273 264,1 3,2
11 245 252 273 265,4 2,8 61 335 347 273 263,6 3,5
12 221 229 273 263,5 3,5 62 392 399 273 268,2 1,8
13 244 248 273 268,6 1,6 63 341 350 273 266,0 2,6
14 72 79 273 248,8 8,9 64 377 387 273 265,9 2,6
15 104 109 273 260,5 4,6 65 324 337 273 262,5 3,9
16 96 98 273 267,4 2,0 66 329 343 273 261,9 4,1
17 88 89 273 269,9 1,1 67 338 352 273 262,1 4,0
18 110 114 273 263,4 3,5 68 333 341 273 266,6 2,3
19 117 121 273 264,0 3,3 69 264 272 273 265,0 2,9
20 111 113 273 268,2 1,8 70 474 496 273 260,9 4,4
21 78 80 273 266,2 2,5 71 316 327 273 263,8 3,4
22 157 162 273 264,6 3,1 72 470 489 273 262,4 3,9
23 151 153 273 269,4 1,3 73 245 251 273 266,5 2,4
24 116 121 273 261,7 4,1 74 348 359 273 264,6 3,1
25 86 87 273 269,9 1,1 75 346 355 273 266,1 2,5
26 101 104 273 265,1 2,9 76 328 341 273 262,6 3,8
27 122 124 273 268,6 1,6 77 403 419 273 262,6 3,8
28 79 81 273 266,3 2,5 78 376 382 273 268,7 1,6
29 64 67 273 260,8 4,5 79 420 432 273 265,4 2,8
30 58 62 273 255,4 6,5 80 337 352 273 261,4 4,3
31 76 77 273 269,5 1,3 81 470 484 273 265,1 2,9
32 77 81 273 259,5 4,9 82 542 573 273 258,2 5,4
33 69 73 273 258,0 5,5 83 542 573 273 258,2 5,4
34 94 96 273 267,3 2,1 84 379 394 273 262,6 3,8
35 93 96 273 264,5 3,1 85 487 512 273 259,7 4,9
36 99 105 273 257,4 5,7 86 471 492 273 261,3 4,3
37 80 83 273 263,1 3,6 87 504 535 273 257,2 5,8
38 89 93 273 261,3 4,3 88 327 340 273 262,6 3,8
39 90 91 273 270,0 1,1 89 330 342 273 263,4 3,5
40 94 99 273 259,2 5,1 90 314 322 273 266,2 2,5
41 88 90 273 266,9 2,2 91 312 318 273 267,8 1,9
42 73 75 273 265,7 2,7 92 299 308 273 265,0 2,9
43 289 298 273 264,8 3,0 93 354 368 273 262,6 3,8
44 354 360 273 268,5 1,7 94 288 292 273 269,3 1,4
45 345 352 273 267,6 2,0 95 306 313 273 266,9 2,2
46 411 417 273 269,1 1,4 96 398 411 273 264,4 3,2
47 370 378 273 267,2 2,1 97 340 350 273 265,2 2,9
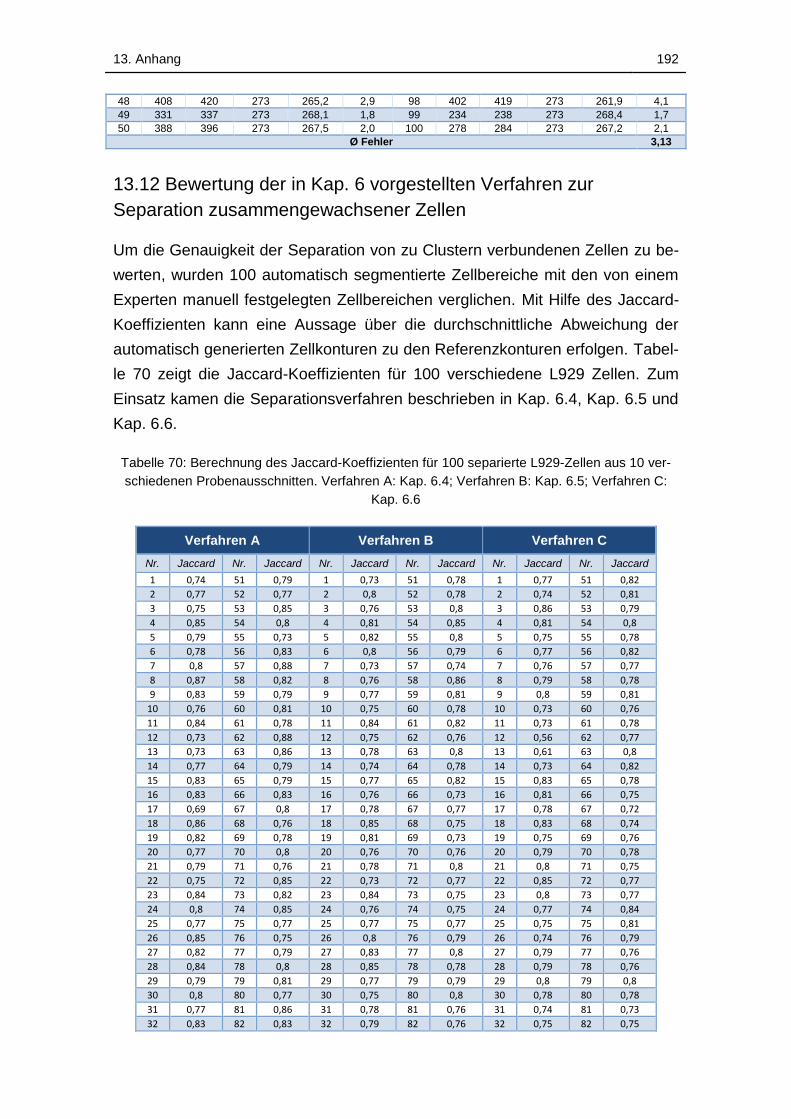
13. Anhang 192
48 408 420 273 265,2 2,9 98 402 419 273 261,9 4,1
49 331 337 273 268,1 1,8 99 234 238 273 268,4 1,7
50 388 396 273 267,5 2,0 100 278 284 273 267,2 2,1
Ø Fehler 3,13
13.12 Bewertung der in Kap. 6 vorgestellten Verfahren zur
Separation zusammengewachsener Zellen
Um die Genauigkeit der Separation von zu Clustern verbundenen Zellen zu be-
werten, wurden 100 automatisch segmentierte Zellbereiche mit den von einem
Experten manuell festgelegten Zellbereichen verglichen. Mit Hilfe des Jaccard-
Koeffizienten kann eine Aussage über die durchschnittliche Abweichung der
automatisch generierten Zellkonturen zu den Referenzkonturen erfolgen. Tabel-
le 70 zeigt die Jaccard-Koeffizienten für 100 verschiedene L929 Zellen. Zum
Einsatz kamen die Separationsverfahren beschrieben in Kap. 6.4, Kap. 6.5 und
Kap. 6.6.
Tabelle 70: Berechnung des Jaccard-Koeffizienten für 100 separierte L929-Zellen aus 10 ver-
schiedenen Probenausschnitten. Verfahren A: Kap. 6.4; Verfahren B: Kap. 6.5; Verfahren C:
Kap. 6.6
Verfahren A Verfahren B Verfahren C
Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard
1 0,74 51 0,79 1 0,73 51 0,78 1 0,77 51 0,82
2 0,77 52 0,77 2 0,8 52 0,78 2 0,74 52 0,81
3 0,75 53 0,85 3 0,76 53 0,8 3 0,86 53 0,79
4 0,85 54 0,8 4 0,81 54 0,85 4 0,81 54 0,8
5 0,79 55 0,73 5 0,82 55 0,8 5 0,75 55 0,78
6 0,78 56 0,83 6 0,8 56 0,79 6 0,77 56 0,82
7 0,8 57 0,88 7 0,73 57 0,74 7 0,76 57 0,77
8 0,87 58 0,82 8 0,76 58 0,86 8 0,79 58 0,78
9 0,83 59 0,79 9 0,77 59 0,81 9 0,8 59 0,81
10 0,76 60 0,81 10 0,75 60 0,78 10 0,73 60 0,76
11 0,84 61 0,78 11 0,84 61 0,82 11 0,73 61 0,78
12 0,73 62 0,88 12 0,75 62 0,76 12 0,56 62 0,77
13 0,73 63 0,86 13 0,78 63 0,8 13 0,61 63 0,8
14 0,77 64 0,79 14 0,74 64 0,78 14 0,73 64 0,82
15 0,83 65 0,79 15 0,77 65 0,82 15 0,83 65 0,78
16 0,83 66 0,83 16 0,76 66 0,73 16 0,81 66 0,75
17 0,69 67 0,8 17 0,78 67 0,77 17 0,78 67 0,72
18 0,86 68 0,76 18 0,85 68 0,75 18 0,83 68 0,74
19 0,82 69 0,78 19 0,81 69 0,73 19 0,75 69 0,76
20 0,77 70 0,8 20 0,76 70 0,76 20 0,79 70 0,78
21 0,79 71 0,76 21 0,78 71 0,8 21 0,8 71 0,75
22 0,75 72 0,85 22 0,73 72 0,77 22 0,85 72 0,77
23 0,84 73 0,82 23 0,84 73 0,75 23 0,8 73 0,77
24 0,8 74 0,85 24 0,76 74 0,75 24 0,77 74 0,84
25 0,77 75 0,77 25 0,77 75 0,77 25 0,75 75 0,81
26 0,85 76 0,75 26 0,8 76 0,79 26 0,74 76 0,79
27 0,82 77 0,79 27 0,83 77 0,8 27 0,79 77 0,76
28 0,84 78 0,8 28 0,85 78 0,78 28 0,79 78 0,76
29 0,79 79 0,81 29 0,77 79 0,79 29 0,8 79 0,8
30 0,8 80 0,77 30 0,75 80 0,8 30 0,78 80 0,78
31 0,77 81 0,86 31 0,78 81 0,76 31 0,74 81 0,73
32 0,83 82 0,83 32 0,79 82 0,76 32 0,75 82 0,75
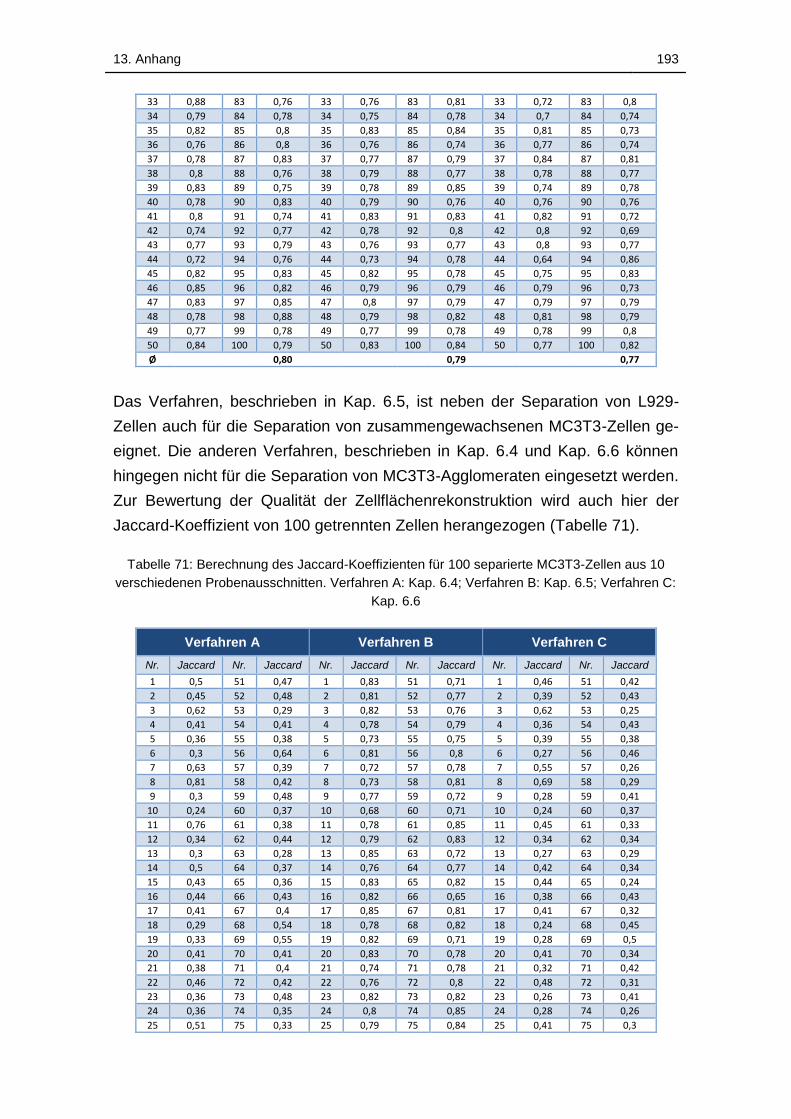
13. Anhang 193
33 0,88 83 0,76 33 0,76 83 0,81 33 0,72 83 0,8
34 0,79 84 0,78 34 0,75 84 0,78 34 0,7 84 0,74
35 0,82 85 0,8 35 0,83 85 0,84 35 0,81 85 0,73
36 0,76 86 0,8 36 0,76 86 0,74 36 0,77 86 0,74
37 0,78 87 0,83 37 0,77 87 0,79 37 0,84 87 0,81
38 0,8 88 0,76 38 0,79 88 0,77 38 0,78 88 0,77
39 0,83 89 0,75 39 0,78 89 0,85 39 0,74 89 0,78
40 0,78 90 0,83 40 0,79 90 0,76 40 0,76 90 0,76
41 0,8 91 0,74 41 0,83 91 0,83 41 0,82 91 0,72
42 0,74 92 0,77 42 0,78 92 0,8 42 0,8 92 0,69
43 0,77 93 0,79 43 0,76 93 0,77 43 0,8 93 0,77
44 0,72 94 0,76 44 0,73 94 0,78 44 0,64 94 0,86
45 0,82 95 0,83 45 0,82 95 0,78 45 0,75 95 0,83
46 0,85 96 0,82 46 0,79 96 0,79 46 0,79 96 0,73
47 0,83 97 0,85 47 0,8 97 0,79 47 0,79 97 0,79
48 0,78 98 0,88 48 0,79 98 0,82 48 0,81 98 0,79
49 0,77 99 0,78 49 0,77 99 0,78 49 0,78 99 0,8
50 0,84 100 0,79 50 0,83 100 0,84 50 0,77 100 0,82
Ø 0,80 0,79 0,77
Das Verfahren, beschrieben in Kap. 6.5, ist neben der Separation von L929-
Zellen auch für die Separation von zusammengewachsenen MC3T3-Zellen ge-
eignet. Die anderen Verfahren, beschrieben in Kap. 6.4 und Kap. 6.6 können
hingegen nicht für die Separation von MC3T3-Agglomeraten eingesetzt werden.
Zur Bewertung der Qualität der Zellflächenrekonstruktion wird auch hier der
Jaccard-Koeffizient von 100 getrennten Zellen herangezogen (Tabelle 71).
Tabelle 71: Berechnung des Jaccard-Koeffizienten für 100 separierte MC3T3-Zellen aus 10
verschiedenen Probenausschnitten. Verfahren A: Kap. 6.4; Verfahren B: Kap. 6.5; Verfahren C:
Kap. 6.6
Verfahren A Verfahren B Verfahren C
Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard Nr. Jaccard
1 0,5 51 0,47 1 0,83 51 0,71 1 0,46 51 0,42
2 0,45 52 0,48 2 0,81 52 0,77 2 0,39 52 0,43
3 0,62 53 0,29 3 0,82 53 0,76 3 0,62 53 0,25
4 0,41 54 0,41 4 0,78 54 0,79 4 0,36 54 0,43
5 0,36 55 0,38 5 0,73 55 0,75 5 0,39 55 0,38
6 0,3 56 0,64 6 0,81 56 0,8 6 0,27 56 0,46
7 0,63 57 0,39 7 0,72 57 0,78 7 0,55 57 0,26
8 0,81 58 0,42 8 0,73 58 0,81 8 0,69 58 0,29
9 0,3 59 0,48 9 0,77 59 0,72 9 0,28 59 0,41
10 0,24 60 0,37 10 0,68 60 0,71 10 0,24 60 0,37
11 0,76 61 0,38 11 0,78 61 0,85 11 0,45 61 0,33
12 0,34 62 0,44 12 0,79 62 0,83 12 0,34 62 0,34
13 0,3 63 0,28 13 0,85 63 0,72 13 0,27 63 0,29
14 0,5 64 0,37 14 0,76 64 0,77 14 0,42 64 0,34
15 0,43 65 0,36 15 0,83 65 0,82 15 0,44 65 0,24
16 0,44 66 0,43 16 0,82 66 0,65 16 0,38 66 0,43
17 0,41 67 0,4 17 0,85 67 0,81 17 0,41 67 0,32
18 0,29 68 0,54 18 0,78 68 0,82 18 0,24 68 0,45
19 0,33 69 0,55 19 0,82 69 0,71 19 0,28 69 0,5
20 0,41 70 0,41 20 0,83 70 0,78 20 0,41 70 0,34
21 0,38 71 0,4 21 0,74 71 0,78 21 0,32 71 0,42
22 0,46 72 0,42 22 0,76 72 0,8 22 0,48 72 0,31
23 0,36 73 0,48 23 0,82 73 0,82 23 0,26 73 0,41
24 0,36 74 0,35 24 0,8 74 0,85 24 0,28 74 0,26
25 0,51 75 0,33 25 0,79 75 0,84 25 0,41 75 0,3
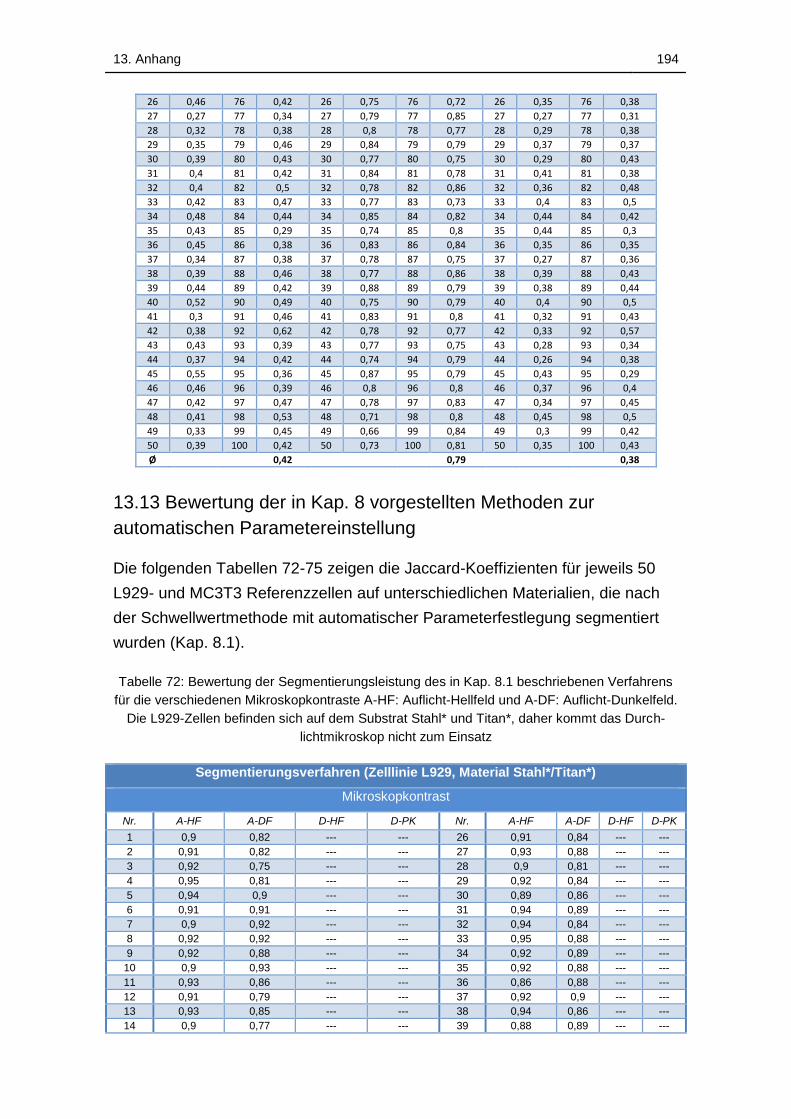
13. Anhang 194
26 0,46 76 0,42 26 0,75 76 0,72 26 0,35 76 0,38
27 0,27 77 0,34 27 0,79 77 0,85 27 0,27 77 0,31
28 0,32 78 0,38 28 0,8 78 0,77 28 0,29 78 0,38
29 0,35 79 0,46 29 0,84 79 0,79 29 0,37 79 0,37
30 0,39 80 0,43 30 0,77 80 0,75 30 0,29 80 0,43
31 0,4 81 0,42 31 0,84 81 0,78 31 0,41 81 0,38
32 0,4 82 0,5 32 0,78 82 0,86 32 0,36 82 0,48
33 0,42 83 0,47 33 0,77 83 0,73 33 0,4 83 0,5
34 0,48 84 0,44 34 0,85 84 0,82 34 0,44 84 0,42
35 0,43 85 0,29 35 0,74 85 0,8 35 0,44 85 0,3
36 0,45 86 0,38 36 0,83 86 0,84 36 0,35 86 0,35
37 0,34 87 0,38 37 0,78 87 0,75 37 0,27 87 0,36
38 0,39 88 0,46 38 0,77 88 0,86 38 0,39 88 0,43
39 0,44 89 0,42 39 0,88 89 0,79 39 0,38 89 0,44
40 0,52 90 0,49 40 0,75 90 0,79 40 0,4 90 0,5
41 0,3 91 0,46 41 0,83 91 0,8 41 0,32 91 0,43
42 0,38 92 0,62 42 0,78 92 0,77 42 0,33 92 0,57
43 0,43 93 0,39 43 0,77 93 0,75 43 0,28 93 0,34
44 0,37 94 0,42 44 0,74 94 0,79 44 0,26 94 0,38
45 0,55 95 0,36 45 0,87 95 0,79 45 0,43 95 0,29
46 0,46 96 0,39 46 0,8 96 0,8 46 0,37 96 0,4
47 0,42 97 0,47 47 0,78 97 0,83 47 0,34 97 0,45
48 0,41 98 0,53 48 0,71 98 0,8 48 0,45 98 0,5
49 0,33 99 0,45 49 0,66 99 0,84 49 0,3 99 0,42
50 0,39 100 0,42 50 0,73 100 0,81 50 0,35 100 0,43
Ø 0,42 0,79 0,38
13.13 Bewertung der in Kap. 8 vorgestellten Methoden zur
automatischen Parametereinstellung
Die folgenden Tabellen 72-75 zeigen die Jaccard-Koeffizienten für jeweils 50
L929- und MC3T3 Referenzzellen auf unterschiedlichen Materialien, die nach
der Schwellwertmethode mit automatischer Parameterfestlegung segmentiert
wurden (Kap. 8.1).
Tabelle 72: Bewertung der Segmentierungsleistung des in Kap. 8.1 beschriebenen Verfahrens
für die verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld.
Die L929-Zellen befinden sich auf dem Substrat Stahl* und Titan*, daher kommt das Durch-
lichtmikroskop nicht zum Einsatz
Segmentierungsverfahren (Zelllinie L929, Material Stahl*/Titan*)
Mikroskopkontrast
Nr. A-HF A-DF D-HF D-PK Nr. A-HF A-DF D-HF D-PK
1 0,9 0,82 --- --- 26 0,91 0,84 --- ---
2 0,91 0,82 --- --- 27 0,93 0,88 --- ---
3 0,92 0,75 --- --- 28 0,9 0,81 --- ---
4 0,95 0,81 --- --- 29 0,92 0,84 --- ---
5 0,94 0,9 --- --- 30 0,89 0,86 --- ---
6 0,91 0,91 --- --- 31 0,94 0,89 --- ---
7 0,9 0,92 --- --- 32 0,94 0,84 --- ---
8 0,92 0,92 --- --- 33 0,95 0,88 --- ---
9 0,92 0,88 --- --- 34 0,92 0,89 --- ---
10 0,9 0,93 --- --- 35 0,92 0,88 --- ---
11 0,93 0,86 --- --- 36 0,86 0,88 --- ---
12 0,91 0,79 --- --- 37 0,92 0,9 --- ---
13 0,93 0,85 --- --- 38 0,94 0,86 --- ---
14 0,9 0,77 --- --- 39 0,88 0,89 --- ---

13. Anhang 195
15 0,91 0,7 --- --- 40 0,9 0,87 --- ---
16 0,92 0,87 --- --- 41 0,9 0,91 --- ---
17 0,9 0,86 --- --- 42 0,89 0,87 --- ---
18 0,88 0,85 --- --- 43 0,9 0,93 --- ---
19 0,93 0,93 --- --- 44 0,92 0,94 --- ---
20 0,92 0,92 --- --- 45 0,91 0,9 --- ---
21 0,9 0,9 --- --- 46 0,93 0,88 --- ---
22 0,94 0,91 --- --- 47 0,92 0,89 --- ---
23 0,92 0,9 --- --- 48 0,94 0,88 --- ---
24 0,91 0,85 --- --- 49 0,95 0,9 --- ---
25 0,92 0,78 --- --- 50 0,94 0,86 --- ---
0,92 0,87
Tabelle 73: Bewertung der Segmentierungsleistung des in Kap. 8.1 beschriebenen Verfahrens
für die verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld.
Die L929-Zellen befinden sich auf dem Substrat Polystyrol, daher kommt das Auflichtmikroskop
nicht zum Einsatz
Segmentierungsverfahren (Zelllinie L929, Material Polystyrol)
Mikroskopkontrast
Nr. A-HF A-DF D-HF D-PK Nr. A-HF A-DF D-HF D-PK
1 0,91 0,66 --- --- 26 0,93 0,68 --- ---
2 0,95 0,83 --- --- 27 0,92 0,7 --- ---
3 0,88 0,14 --- --- 28 0,94 0,72 --- ---
4 0,87 0,66 --- --- 29 0,93 0,66 --- ---
5 0,92 0,38 --- --- 30 0,94 0,75 --- ---
6 0,91 0,66 --- --- 31 0,9 0,63 --- ---
7 0,9 0,6 --- --- 32 0,89 0,64 --- ---
8 0,93 0,62 --- --- 33 0,86 0,77 --- ---
9 0,88 0,58 --- --- 34 0,94 0,7 --- ---
10 0,89 0,69 --- --- 35 0,93 0,75 --- ---
11 0,89 0,58 --- --- 36 0,91 0,76 --- ---
12 0,86 0,72 --- --- 37 0,86 0,68 --- ---
13 0,9 0,7 --- --- 38 0,89 0,59 --- ---
14 0,87 0,65 --- --- 39 0,88 0,72 --- ---
15 0,85 0,67 --- --- 40 0,87 0,57 --- ---
16 0,88 0,7 --- --- 41 0,93 0,69 --- ---
17 0,9 0,71 --- --- 42 0,86 0,5 --- ---
18 0,92 0,73 --- --- 43 0,95 0,8 --- ---
19 0,9 0,65 --- --- 44 0,79 0,62 --- ---
20 0,91 0,67 --- --- 45 0,9 0,71 --- ---
21 0,93 0,5 --- --- 46 0,91 0,58 --- ---
22 0,89 0,47 --- --- 47 0,86 0,7 --- ---
23 0,92 0,42 --- --- 48 0,86 0,68 --- ---
24 0,93 0,64 --- --- 49 0,89 0,74 --- ---
25 0,9 0,66 --- --- 50 0,91 0,73 --- ---
0,90 0,65
Tabelle 74: Bewertung der Segmentierungsleistung des in Kap. 8.1 beschriebenen Verfahrens
für die verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld.
Die MC3T3-Zellen befinden sich auf dem Substrat Stahl* und Titan*, daher kommt das Durch-
lichtmikroskop nicht zum Einsatz
Segmentierungsverfahren (Zelllinie MC3T3, Material Stahl*/Titan*)
Mikroskopkontrast
Nr. A-HF A-DF D-HF D-PK Nr. A-HF A-DF D-HF D-PK
1 0,76 0,95 --- --- 26 0,91 0,9 --- ---
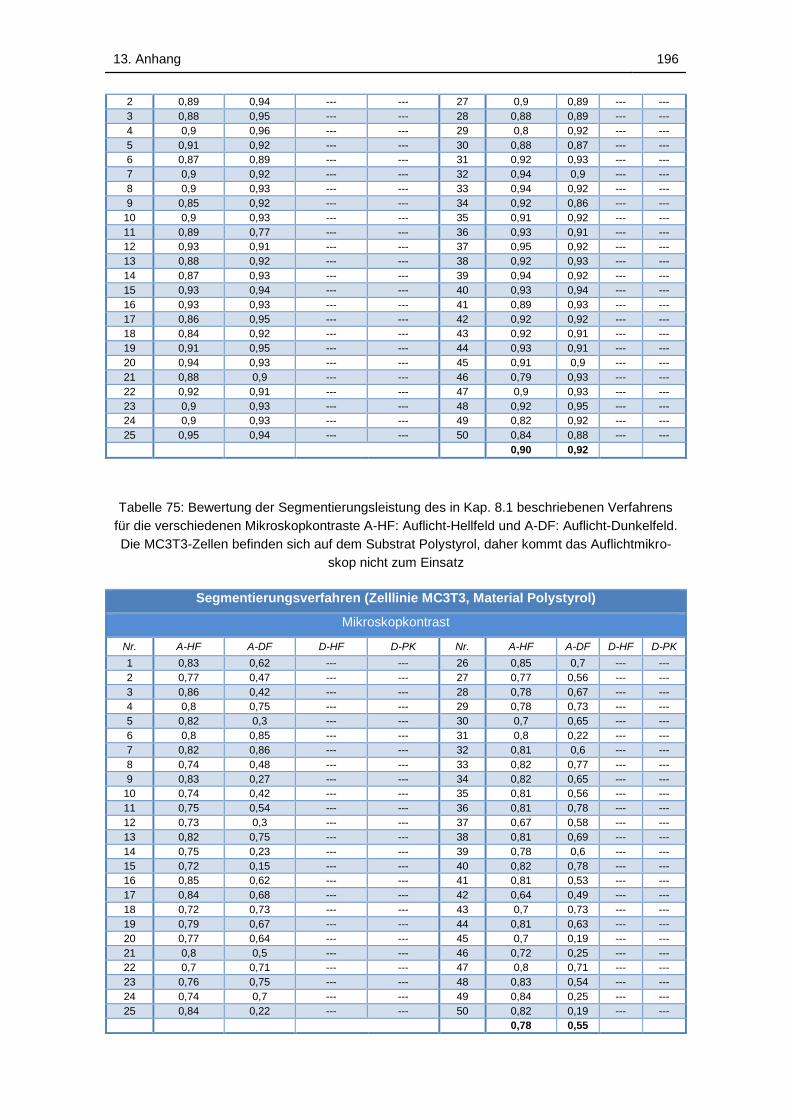
13. Anhang 196
2 0,89 0,94 --- --- 27 0,9 0,89 --- ---
3 0,88 0,95 --- --- 28 0,88 0,89 --- ---
4 0,9 0,96 --- --- 29 0,8 0,92 --- ---
5 0,91 0,92 --- --- 30 0,88 0,87 --- ---
6 0,87 0,89 --- --- 31 0,92 0,93 --- ---
7 0,9 0,92 --- --- 32 0,94 0,9 --- ---
8 0,9 0,93 --- --- 33 0,94 0,92 --- ---
9 0,85 0,92 --- --- 34 0,92 0,86 --- ---
10 0,9 0,93 --- --- 35 0,91 0,92 --- ---
11 0,89 0,77 --- --- 36 0,93 0,91 --- ---
12 0,93 0,91 --- --- 37 0,95 0,92 --- ---
13 0,88 0,92 --- --- 38 0,92 0,93 --- ---
14 0,87 0,93 --- --- 39 0,94 0,92 --- ---
15 0,93 0,94 --- --- 40 0,93 0,94 --- ---
16 0,93 0,93 --- --- 41 0,89 0,93 --- ---
17 0,86 0,95 --- --- 42 0,92 0,92 --- ---
18 0,84 0,92 --- --- 43 0,92 0,91 --- ---
19 0,91 0,95 --- --- 44 0,93 0,91 --- ---
20 0,94 0,93 --- --- 45 0,91 0,9 --- ---
21 0,88 0,9 --- --- 46 0,79 0,93 --- ---
22 0,92 0,91 --- --- 47 0,9 0,93 --- ---
23 0,9 0,93 --- --- 48 0,92 0,95 --- ---
24 0,9 0,93 --- --- 49 0,82 0,92 --- ---
25 0,95 0,94 --- --- 50 0,84 0,88 --- ---
0,90 0,92
Tabelle 75: Bewertung der Segmentierungsleistung des in Kap. 8.1 beschriebenen Verfahrens
für die verschiedenen Mikroskopkontraste A-HF: Auflicht-Hellfeld und A-DF: Auflicht-Dunkelfeld.
Die MC3T3-Zellen befinden sich auf dem Substrat Polystyrol, daher kommt das Auflichtmikro-
skop nicht zum Einsatz
Segmentierungsverfahren (Zelllinie MC3T3, Material Polystyrol)
Mikroskopkontrast
Nr. A-HF A-DF D-HF D-PK Nr. A-HF A-DF D-HF D-PK
1 0,83 0,62 --- --- 26 0,85 0,7 --- ---
2 0,77 0,47 --- --- 27 0,77 0,56 --- ---
3 0,86 0,42 --- --- 28 0,78 0,67 --- ---
4 0,8 0,75 --- --- 29 0,78 0,73 --- ---
5 0,82 0,3 --- --- 30 0,7 0,65 --- ---
6 0,8 0,85 --- --- 31 0,8 0,22 --- ---
7 0,82 0,86 --- --- 32 0,81 0,6 --- ---
8 0,74 0,48 --- --- 33 0,82 0,77 --- ---
9 0,83 0,27 --- --- 34 0,82 0,65 --- ---
10 0,74 0,42 --- --- 35 0,81 0,56 --- ---
11 0,75 0,54 --- --- 36 0,81 0,78 --- ---
12 0,73 0,3 --- --- 37 0,67 0,58 --- ---
13 0,82 0,75 --- --- 38 0,81 0,69 --- ---
14 0,75 0,23 --- --- 39 0,78 0,6 --- ---
15 0,72 0,15 --- --- 40 0,82 0,78 --- ---
16 0,85 0,62 --- --- 41 0,81 0,53 --- ---
17 0,84 0,68 --- --- 42 0,64 0,49 --- ---
18 0,72 0,73 --- --- 43 0,7 0,73 --- ---
19 0,79 0,67 --- --- 44 0,81 0,63 --- ---
20 0,77 0,64 --- --- 45 0,7 0,19 --- ---
21 0,8 0,5 --- --- 46 0,72 0,25 --- ---
22 0,7 0,71 --- --- 47 0,8 0,71 --- ---
23 0,76 0,75 --- --- 48 0,83 0,54 --- ---
24 0,74 0,7 --- --- 49 0,84 0,25 --- ---
25 0,84 0,22 --- --- 50 0,82 0,19 --- ---
0,78 0,55

13. Anhang 197
13.14 Untersuchung verschiedener Stichproben der
Probenoberfläche
Für eine computergestützte Analyse der Zellen auf der Probenoberfläche wird
die Vergrößerung am Mikroskop auf den Faktor 50 eingestellt. Mit diesem Pa-
rameter sind 128 Aufnahmen für die Abbildung der vollständigen Oberfläche
notwendig. Zur Reduzierung des damit verbundenen hohen Arbeitsaufwandes
ist die Verwendung einer Stichprobe der Zellaufnahmen denkbar. Tabelle 76
zeigt die Untersuchungsergebnisse der Abweichungen der durchschnittlichen
Zellanzahl verschiedener Stichprobenumfänge im Vergleich zum Referenzwert.
Unter dem Referenzwert ist hier die durchschnittliche Zellanzahl aller Proben-
ausschnitte zu verstehen, die keine Probenrandbereiche aufweisen.
Tabelle 76: Verschiedene Stichprobenumfänge mit der jeweiligen Abweichung zum Referenz-
wert. Für jeden Stichprobenumfang wurden 10 Versuche durchgeführt. Die zufälligen Stichpro-
ben umfassen 50, 30 und 10 Bilder, die gezielt ausgewählten Stichproben hingegen 15, 10 und
5 Bilder. Zeichenerklärung: Ø: Durchschnittliche Zellanzahl der Stichprobe; σ: Standardabwei-
chung der Stichprobe; ∆REF: relative Abweichung der durchschnittlichen Zellanzahl der Stichpro-
be zum Referenzwert; Abw.: Die maximale positive und negative Abweichung der durchschnitt-
lichen Zellzahl vom Referenzwert [%]
Stichprobenumfänge
zufällige Auswahl der Stichprobe
Nr. 80 Bilder 50 Bilder 30 Bilder 15 Bilder
Ø σ ∆REF Ø σ ∆REF Ø σ ∆REF Ø σ ∆REF
1 506,1 379,6 6,0 441,3 331,5 -7,5 439,1 257,0 -8,0 477,5 302,2 0,1
2 494,6 384,9 3,6 482,3 364,0 1,1 510,3 408,6 6,9 589,4 555,5 23,5
3 473,6 374,7 -0,8 496,6 412,1 4,1 394,1 217,1 -17,4 398,4 156,7 -16,5
4 491,6 380,2 3,0 471,4 328,0 -1,2 529,7 386,6 11,0 423,1 195,6 -11,3
5 472,0 356,5 -1,1 489,2 349,4 2,5 613,9 497,6 28,6 485,1 302,8 1,6
6 491,3 383,1 2,9 475,4 321,2 -0,4 487,7 412,9 2,2 384,1 73,1 -19,5
7 493,0 382,1 3,3 477,0 396,7 0,0 507,2 393,3 6,3 379,1 143,9 -20,6
8 457,3 319,9 -4,2 541,5 419,2 13,5 499,4 415,2 4,6 587,6 438,2 23,1
9 480,0 373,3 0,6 519,4 433,4 8,8 651,0 532,6 36,4 439,0 282,2 -8,0
10 474,6 373,3 -0,6 462,0 369,4 -3,2 497,2 359,2 4,2 478,1 316,4 0,2
Abw. -4,2% bis 6,0% -7,5% bis +13,5% -17,4% bis +36,4% -20,6% bis +23,5%
gezielte Auswahl der Stichprobe
Nr. 20 Bilder 15 Bilder 10 Bilder 5 Bilder
Ø σ ∆REF Ø σ ∆REF Ø σ ∆REF Ø σ ∆REF
1 463,2 66,7 -2,9 483,3 60,1 1,3 469,9 86,9 -1,5 429,0 51,6 -10,1
2 454,2 69,6 -4,8 475,9 77,1 -0,3 483,2 44,2 1,3 528,2 45,7 9,6
3 491,6 62,8 3,0 484,7 75,9 1,6 465,1 70,6 -2,5 487,6 47,3 2,2
4 499,2 45,1 4,6 476,2 65,7 -0,2 470,4 59,3 -1,5 426,2 70,1 -10,7
5 475,2 65,1 -0,4 481,5 70,5 0,9 507,9 104,1 6,4 415,8 38,1 -12,9
6 460,2 73,7 -3,6 461,3 65,7 -3,3 473,7 78,9 -0,7 444,0 44,4 -7,0

13. Anhang 198
7 477,9 59,8 0,1 487,9 62,4 2,2 459,6 69,6 -3,7 381,6 35,5 -20,0
8 477,4 60,2 0,0 487,4 89,2 2,0 417,3 41,7 -12,6 400,4 42,8 -16,1
9 484,9 61,6 1,6 475,6 61,5 -0,3 442,1 57 -7,3 498,8 71,9 4,5
10 464,6 47,3 -2,6 454,7 52,0 -4,7 420,2 84,6 -12,0 495,6 75,6 3,8
Abw. -4,8% bis +4,6% -4,7% bis +2,2% -12,6% bis +6,4% -20,0% bis 9,6%
13.15 Untersuchung zur manuellen Zellzählung in Suspension und
auf der Probenoberfläche
In der nachfolgenden Tabelle sind die Ergebnisse der manuellen Zellzählung in
Suspension mit Hilfe einer Fuchs-Rosenthal-Kammer für 6 Proben aufgeführt.
Auf diese 6 Proben wurden im Vorfeld jeweils 10.000 L929-Zellen aufgebracht,
72 Stunden im Brutschrank inkubiert und danach für die Zählung in Suspension
abgelöst.
Tabelle 77: Manuell ermittelte Zellanzahl für 6 Proben in Suspension. Pro Probe wurden 10
Zählungen durchgeführt und der Mittelwert gebildet
Versuch-Nr. Ermittelte Zellanzahl Probe 1
1 47.813
2 52.813
3 44.375
4 51.250
5 50.625
6 51.250
7 44.063
8 45.625
9 47.188
10 47.813
Mittelwert 48.281
Standardabweichung 2.922
Probe 2
1 55.000
2 64.063
3 57.813
4 62.813
5 58.438
6 59.063
7 60.000
8 65.313
9 60.625
10 62.500
Mittelwert 60.563
Standardabweichung 2.987
Probe 3
1 49.063
2 50.625
3 53.438
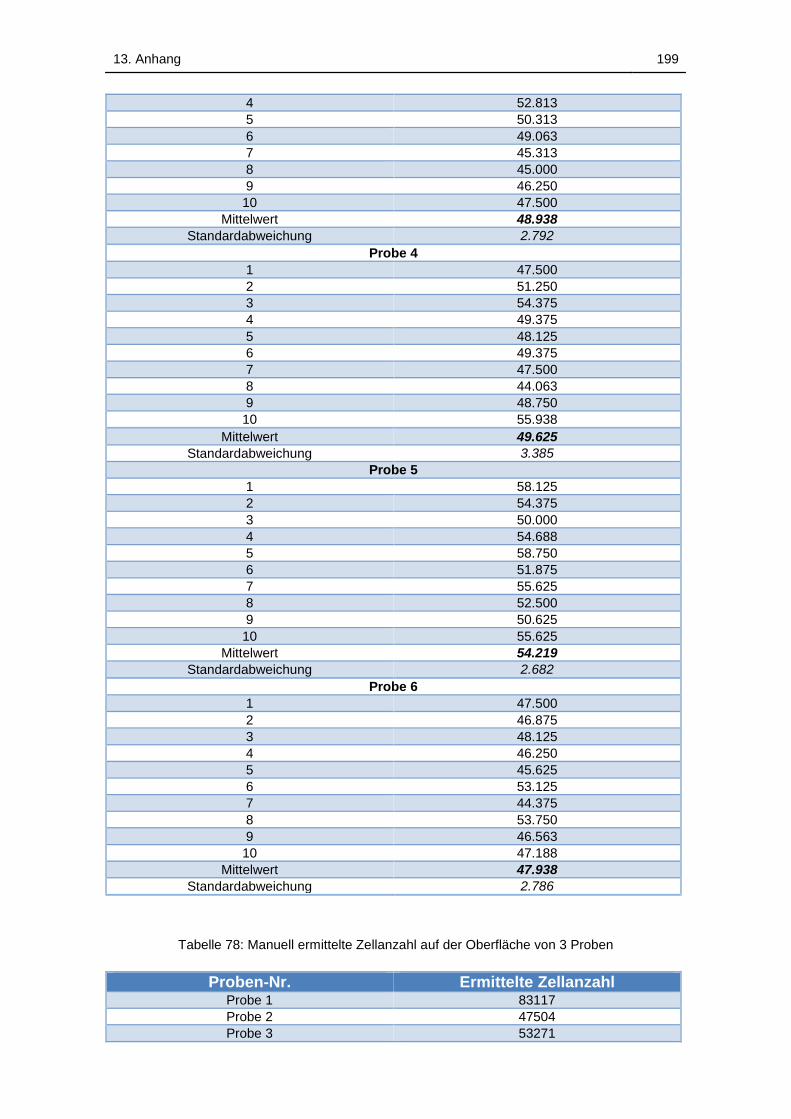
13. Anhang 199
4 52.813
5 50.313
6 49.063
7 45.313
8 45.000
9 46.250
10 47.500
Mittelwert 48.938
Standardabweichung 2.792
Probe 4
1 47.500
2 51.250
3 54.375
4 49.375
5 48.125
6 49.375
7 47.500
8 44.063
9 48.750
10 55.938
Mittelwert 49.625
Standardabweichung 3.385
Probe 5
1 58.125
2 54.375
3 50.000
4 54.688
5 58.750
6 51.875
7 55.625
8 52.500
9 50.625
10 55.625
Mittelwert 54.219
Standardabweichung 2.682
Probe 6
1 47.500
2 46.875
3 48.125
4 46.250
5 45.625
6 53.125
7 44.375
8 53.750
9 46.563
10 47.188
Mittelwert 47.938
Standardabweichung 2.786
Tabelle 78: Manuell ermittelte Zellanzahl auf der Oberfläche von 3 Proben
Proben-Nr. Ermittelte Zellanzahl Probe 1 83117
Probe 2 47504
Probe 3 53271
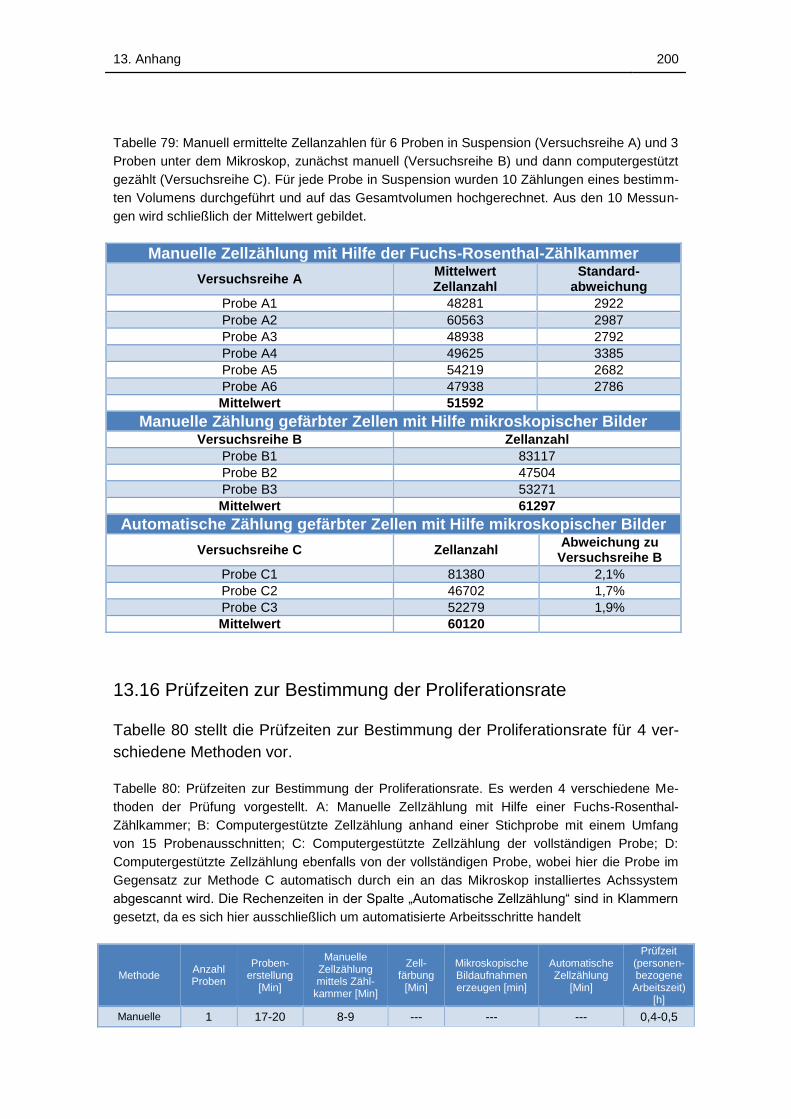
13. Anhang 200
Tabelle 79: Manuell ermittelte Zellanzahlen für 6 Proben in Suspension (Versuchsreihe A) und 3
Proben unter dem Mikroskop, zunächst manuell (Versuchsreihe B) und dann computergestützt
gezählt (Versuchsreihe C). Für jede Probe in Suspension wurden 10 Zählungen eines bestimm-
ten Volumens durchgeführt und auf das Gesamtvolumen hochgerechnet. Aus den 10 Messun-
gen wird schließlich der Mittelwert gebildet.
Manuelle Zellzählung mit Hilfe der Fuchs-Rosenthal-Zählkammer
Versuchsreihe A Mittelwert Zellanzahl
Standard-abweichung
Probe A1 48281 2922
Probe A2 60563 2987
Probe A3 48938 2792
Probe A4 49625 3385
Probe A5 54219 2682
Probe A6 47938 2786
Mittelwert 51592
Manuelle Zählung gefärbter Zellen mit Hilfe mikroskopischer Bilder
Versuchsreihe B
Zellanzahl
Probe B1 83117
Probe B2 47504
Probe B3 53271
Mittelwert 61297
Automatische Zählung gefärbter Zellen mit Hilfe mikroskopischer Bilder
Versuchsreihe C
Zellanzahl Abweichung zu
Versuchsreihe B
Probe C1 81380 2,1%
Probe C2 46702 1,7%
Probe C3 52279 1,9%
Mittelwert 60120
13.16 Prüfzeiten zur Bestimmung der Proliferationsrate
Tabelle 80 stellt die Prüfzeiten zur Bestimmung der Proliferationsrate für 4 ver-
schiedene Methoden vor.
Tabelle 80: Prüfzeiten zur Bestimmung der Proliferationsrate. Es werden 4 verschiedene Me-
thoden der Prüfung vorgestellt. A: Manuelle Zellzählung mit Hilfe einer Fuchs-Rosenthal-
Zählkammer; B: Computergestützte Zellzählung anhand einer Stichprobe mit einem Umfang
von 15 Probenausschnitten; C: Computergestützte Zellzählung der vollständigen Probe; D:
Computergestützte Zellzählung ebenfalls von der vollständigen Probe, wobei hier die Probe im
Gegensatz zur Methode C automatisch durch ein an das Mikroskop installiertes Achssystem
abgescannt wird. Die Rechenzeiten in der Spalte „Automatische Zellzählung“ sind in Klammern
gesetzt, da es sich hier ausschließlich um automatisierte Arbeitsschritte handelt
Methode Anzahl Proben
Proben-erstellung
[Min]
Manuelle Zellzählung mittels Zähl-
kammer [Min]
Zell-färbung
[Min]
Mikroskopische Bildaufnahmen erzeugen [min]
Automatische Zellzählung
[Min]
Prüfzeit (personen-bezogene
Arbeitszeit) [h]
Manuelle 1 17-20 8-9 --- --- --- 0,4-0,5

13. Anhang 201
Zellzählung [A]
5 37-52 17-19 --- --- --- 0,9-1,2
10 62-92 33-38 --- --- --- 1,6-2,2
25 137-212 81-94 --- --- --- 3,6-5,1
50 262-412 163-188 --- --- --- 7,1-10,0
Automatische Zellzählung (Stichprobe
mit 15 Bildern)
[B]
1 17-20 --- 11 4 (1) 0,5-0,6
5 37-52 --- 13 20 (5) 1,2-1,4
10 62-92 --- 15 40 (10) 2,0-2,5
25 137-212 --- 20 100 (25) 4,3-5,5
50 262-412 --- 29 200 (50) 8,2-10,7
Automatische Zellzählung (vollständige Zellzählung)
[C]
1 17-20 --- 11 36 (8,5) 1,1
5 37-52 --- 13 180 (3h) (42,5) 3,8-4,1
10 62-92 --- 15 360 (6h) (85) 7,3-7,8
25 137-212 --- 20 900 (15h) (212,5) 17,6-18,9
50 262-412 --- 29 1800 (30h) (425) 34,9-37,4
Automatischer Probenscan
und computer-gestützte
Zellzählung [D]
1 17-20 --- 11 (8,5) 0,5
5 37-52 --- 13 (42,5) 0,8-1,1
10 62-92 --- 15 (85) 1,3-1,8
25 137-212 --- 20 (212,5) 2,6-3,9
50 262-412 --- 29 (425) 4,9-7,4
13.17. Elimination von Ausreißern mit Hilfe des Grubbs-Tests
Die Ergebnisse der Zellzählung der Versuchsreihen A, B und C sind in Kap.
13.15, Tabelle 79 gezeigt. Dabei weichen teilweise Werte deutlich vom Mittel-
wert ab. Der Grund liegt darin, dass mit der Methode zum Aufbringen der Zellen
auf die Probenoberfläche (Suspensionsmethode) nicht reproduzierbar die glei-
che Anzahl Zellen auf unterschiedliche Oberflächen aufgebracht werden kön-
nen. Um eine zuverlässigere Aussage über die Biokompatibilität des zugrunde
liegenden Materials zu gewährleisten, ist die Bestimmung der Ausreißer der
Versuchsreihen sinnvoll. Anschließend wird der Mittelwert der Zellanzahl für die
Versuchsreihen neu bestimmt. Auf diese Weise ist eine genauere Bestimmung
der Proliferationsrate gewährleistet. Eine Methode zur Bestimmung der Ausrei-
ßer für die Versuchsreihen A, B und C ist der Grubbs-Test (84) nach folgender
Vorschrift:
max( )in
x xT
s
mit (Gl. 28)
xi: Zellanzahl auf der i-ten Probe
x : Mittelwert der Zellzahlen aller Proben

13. Anhang 202
s: Standardabweichung aller Zellzahlen
Für die Versuchsreihe A ergibt sich nach Gl. 28 der Testwert TA = 1,984, für die
Versuchsreihe B TB = 1,398 und für die Versuchsreihe C TC = 1,398 (Tabelle
81).
Aus der Grubbs-Tabelle (84) entnimmt man unter Beachtung eines geeigneten
Signifikanzniveaus, hier z.B. α = 5% den festgelegten Schwellwert, der ebenso
von der Anzahl der aufgenommenen Messwerte abhängt. Für Versuchsreihe A
erhält man den Schwellwert SA = 1,82 und für die Versuchsreihen B und C je-
weils den Schwellwert SB = SC = 1,15. Ist der Testwert TA > SA so handelt es
sich bei dem entsprechenden Messwert um einen Ausreißer. Der Vorgang wird
anschließend für die verbleibenden Messwerte wiederholt bis kein weiterer Aus-
reißer ermittelt wurde. Für die Versuchsreihen A, B und C ergeben sich in die
Tabelle 81 aufgeführten Ausreißer.
Tabelle 81: Testwerte des Grubbs-Tests für die Versuchsreihen A, B und C zur Identifikation
von Ausreißern. Als Signifikanzniveau wurde α = 5% gewählt. Aus der Grubbs-Tabelle (84)
erhält man für n = 6 Messwerte und α=5% einen Schwellwert SA = 1,82. Für n = 3 Messwerte
und ebenfalls α=5% resultiert ein Schwellwert SB = SC = 1,15.
Manuelle Zellzählung mit Hilfe der Fuchs-Rosenthal-Zählkammer
Versuchsreihe A Zellanzahl Grubbs-
Testwert VA Ausreißer
Probe A1 48281 0,732 Nein
Probe A2 60563 1,984 Ja
Probe A3 48938 0,587 Nein
Probe A4 49625 0,435 Nein
Probe A5 54219 0,581 Nein
Probe A6 47928 0,810 Nein
Mittelwert mit Ausreißer 51592
Standardabweichung 4953,4
Mittelwert ohne Ausreißer 49798
Manuelle Zählung gefärbter Zellen mit Hilfe mikroskopischer Bilder
Versuchsreihe B
Zellanzahl Grubbs-
Testwert VB Ausreißer
Probe B1 83117 1,398 Ja
Probe B2 47504 0,884 Nein
Probe B3 53271 0,514 Nein
Mittelwert mit Ausreißer 61297
Standardabweichung 15607,4
Mittelwert ohne Ausreißer 50388
Automatische Zählung gefärbter Zellen mit Hilfe mikroskopischer Bilder
Versuchsreihe C
Zellanzahl Grubbs-
Testwert VC Ausreißer
Probe C1 81380 1,398 Ja
Probe C2 46702 0,883 Nein
Probe C3 52279 0,516 Nein
Mittelwert mit Ausreißer 60120
Standardabweichung 15204,3
Mittelwert ohne Ausreißer 49491

203
Tabelle 81 zeigt, nachdem die Ausreißer ermittelt und die Mittelwerte ohne Aus-
reißer neu berechnet werden, resultieren mit 49798 Zellen für Versuchsreihe A,
50388 Zellen für Versuchsreihe B und 49491 Zellen für Versuchsreihe C ähnli-
che Werte.

Publikationsliste 204
Publikationsliste
S. Buhl, B. Neumann, S.C. Schäfer, “Context-Based Separation of Cell Clusters
for the Automatic Biocompatibility Testing of Implant Materials”, Journal of
Computational Medicine, 2014, Article ID 542521 doi:10.1155/2014/542521
S. Buhl, B. Neumann, S.C. Schäfer, A. Severing, “Automatic cell segmentation
in strongly agglomerated cell networks for different cell types”, International
Journal of Computational Biology and Drug Design, 2013, Bd. 6, 1, S. 200 - 220
S. Buhl, B. Neumann, M. Schneider, U. Lehmann, E. Eisenbarth, "Automatische
Segmentierung von Zellkernen und den dazugehörigen Zellen in Zellclustern für
die Biokompatibilitätsprüfung", 20. Workshop Computational Intelligence des
VDI/GMA-FA 5.14 "Computational Intelligence" und der GI-FG "Fuzzy-Systeme
und Soft-Computing" in Bommerholz bei Dortmund, 1.-3. Dez. 2010
S. Buhl, B. Neumann, E. Eisenbarth, "Segmentation Of Cytological Stained Cell
Areas And Generation Of Cell Boundaries In Complex Shaded Cell Clusters",
55.IWK - International Scientific Colloquium, Isle Publisher, Ilmenau, 2010, pp.
511-514
S. Buhl, B. Neumann, A. Grothe, "Bestimmung der Kraftausübung einer opti-
schen Pinzette auf Partikel in flüssiger Umgebung mit Methoden der digitalen
Bildverarbeitung", DGaO-Proceedings 2010 - http://www.dgao-proceedings.de -
ISSN: 1614-8436
S. Buhl, B. Neumann, A. Grothe, "Bewertung verschiedener Methoden zur Un-
tersuchung mikroskopischer Zellbilder mit CV und CI für die Bioverträglichkeit
von Implantatwerkstoffen", DGaO-Proceedings 2010 - http://www.dgao procee-
dings.de - ISSN: 1614-8436
A. Grothe, B. Neumann, S. Buhl, "Smart Illumination für die Detektion von Arte-
fakten auf glänzenden gekrümmten Oberflächen mit digitaler Bildverarbeitung"
DGaO-Proceedings 2010 - http://www.dgao-proceedings.de - ISSN: 1614-8436
S. Buhl, B. Neumann, M. Schneider, U. Lehmann, E. Eisenbarth, T. Weißbach,
"Automatische Segmentierung von mikroskopischen Zellbildern für die Biokom-
patibilitätsprüfung mit Hilfe eines Künstlich Neuronalen Netzes", 19. Workshop

205
Computational Intelligence des VDI/GMA-FA 5.14 "Computational Intelligence"
und der GI-FG "Fuzzy-Systeme und Soft-Computing" in Bommerholz bei Dort-
mund, 2.-4. Dez. 2009
V. Gottfried, U. Lehmann, U. Klug, B. Neumann, M. Schneider, J. Brenig, S.
Buhl, "Simulation von Neuronalen Netzen für E-Learning", 19. Workshop Com-
putational Intelligence des VDI/GMA-FA 5.14 "Computational Intelligence" und
der GI-FG "Fuzzy-Systeme und Soft-Computing" in Bommerholz bei Dortmund,
2.-4. Dez. 2009

Lebenslauf 206
Lebenslauf
Persönliche Daten
Sven Buhl
Alter Schloßweg 29
58119 Hagen
E-Mail: [email protected]
geboren am 11.09.1981 in Menden
Berufliche Erfahrungen
01.06.2012 – heute Infineon Technologies AG, Process Engineer
Projektmanagement von AOI- und AXI-Themen (Automatic Optical Inspection and Automatic Xray Inspection)
Entwicklung von Bildverarbeitungs-Softwaretools
01.2006 – 31.05.2012 Fachhochschule Südwestfalen, wissenschaftlicher
Mitarbeiter
95% Drittmittelprojekt: Koordination des Softwa-reteams, Softwareentwicklung, Algorithmenent-wicklung
5% Arbeit in der Lehre
2002 – 2004 studentische Aushilfskraft bei der LMB GmbH
Einrichten und Bedienen von Laseranlagen zum Schweißen, Schneiden und Beschriften verschiedener Objekte
Studium
2008 – 2015 Fernuniversität in Hagen
Promotion an der Fakultät für Mathematik und In-
formatik bei Prof. Dr. J. Jahns im Bereich Computer
Vision mit dem Thema: Prüfung der Biokompatibilität
von Implantatwerkstoffen mit Methoden der digitalen
Bildverarbeitung
2006 – 2008 Fachhochschule Südwestfalen
Master of Science im Studiengang Computer Vision
and Computational Intelligence (CV&CI)
Masterarbeit: Konzeption und Realisierung einer
modularen Software auf .NET-Basis für die Anwen-
dung im Robot-Vision Bereich

Lebenslauf 207
2002 – 2005 Fachhochschule Südwestfalen
Diplomstudiengang Physikalische Technik
Diplomarbeit: Stabilisierung der Prozesssicherheit
bei der Laserbeschriftung unter Verwendung indust-
rieller Bildverarbeitung
2002 6 Monate Grund- und Fachpraktikum für den Studi-
engang „Physikalische Technik“ an der Fachhoch-
schule Südwestfalen bei der LMB GmbH und dem
Ingenieurbüro Düputell in Iserlohn
Schulbildung
1988 Einschulung in die Grundschule Wesselbach in
Hagen
1992 – 2001 Abitur an dem städt. Gymnasium Hohenlimburg in
Hagen

Eidesstattliche Erklärung 208
Eidesstattliche Erklärung
Ich erkläre hiermit, dass ich diese Dissertation selbstständig ohne Hilfe Dritter
und ohne Benutzung anderer als der angegebenen Quellen und Hilfsmittel ver-
fasst habe. Alle den benutzten Quellen wörtlich oder sinngemäß entnommenen
Stellen sind als solche einzeln kenntlich gemacht.
Diese Arbeit ist bislang keiner anderen Prüfungsbehörde vorgelegt worden und
auch nicht veröffentlicht worden.
Ich bin mir bewusst, dass eine falsche Erklärung rechtliche Folgen haben wird.
_______________________________________________________________
Ort, Datum, Unterschrift