stoffwechselleistungen, wachstum und differenzierung.swbplus.bsz-bw.de/bsz068713819kap.pdf ·...
TRANSCRIPT

17.1 Hormone als extrazelluläre Botenstoffe
! Hormone regulieren in vielzelligen OrganismenStoffwechselleistungen,Wachstum und Differenzierung.
Mit der Entstehung vielzelliger Organismen und der immer weiterge-henden Spezialisierung von Zellgruppen zu Organen und Gewebenmit spezifischen Funktionen ergab sich die Notwendigkeit der Signal-übermittlung von Zelle zu Zelle. Diese humorale Signalübermittlungbedient sich chemischer Verbindungen als materieller Signalüberträ-ger, die auch als Hormone oder allgemeiner als Signalstoffe bezeich-net werden. Die bei der Signalübertragung prinzipiell realisiertenMöglichkeiten sind in Abb. 17.1 zusammengestellt:
· Um parakrine Signalübermittlung handelt es sich, wenn der Sig-nalstoff durch Diffusion von der sezernierenden auf die benach-barte Zelle gelangt, die über einen spezifischen Rezeptor verfü-gen muû. Derartige Signalstoffe werden auch als Gewebshormo-ne bezeichnet.
· Um juxtakrine Signalübermittlung handelt es sich, wenn der Sig-nalstoff in der Plasmamembran der produzierenden Zelle ver-ankert ist und für die Wechselwirkung mit dem entsprechendenRezeptor auf der Zielzelle ein direkter Zell-Zell-Kontakt nötig ist.
17 Biochemie und Pathobiochemie des endokrinen Systems454
Hormon bzw. Signalstoff Rezeptor
b
a c
d
e
Abb. 17.1 a±e Möglichkeiten der interzellulären Signalübermittlung. a parakrin,b juxtakrin, c intern autokrin, d extern autokrin, e endokrin

· Um endokrine Signalübermittlung handelt es sich, wenn der Sig-nalstoff von einer endokrinen Drüse in die Blutbahn abgegebenwird, um seine Funktion an einer weiter entfernten Zelle auszu-üben. Man spricht in diesem Fall auch von Hormonen.
· Bei der autokrinen Signalübermittlung wirken die von einer se-zernierenden Zelle gebildeten Signalmoleküle auf diese Zelleselbst zurück.
! Hormone lassen sichnach funktionellen Gesichtspunkten einteilen.
Hormone oder extrazelluläre Signalstoffe lassen sich nach dem Typder sezernierenden Zelle einteilen:
· Glanduläre Hormone werden von spezifischen, in endokrinenDrüsen lokalisierten sekretorischen Zellen synthetisiert. Sie wir-ken i. allg. endokrin.
· Gewebshormone werden von in den verschiedensten Geweben lo-kalisierten endokrin aktiven Zellen gebildet und abgegeben. Siewirken i. allg. parakrin.
· Cytokine werden von den verschiedensten Zellen gebildet und re-gulieren meist als parakrine Faktoren Proliferation bzw. Diffe-renzierung und Funktion ihrer Zielzellen. Von besonderer Be-deutung sind die Cytokine, die Proliferation und Differenzierungder zellulären Bestandteile des Blutes regulieren (S. 463).
Alternativ zur Einteilung nach den produzierenden Zellen könnenHormone bzw. Signalstoffe auch nach funktionellen Aspekten einge-teilt werden (Abb. 17.2):
· In den ersten beiden Gruppen befinden sich diejenigen Hormo-ne, die Wachstum und Differenzierungsvorgänge beeinflussen.Es handelt sich um Cytokine sowie eine Reihe glandulärer Hor-mone.
· In der dritten Gruppe befinden sich Hormone, die für die schnel-le Umstellung des Stoffwechsels verantwortlich sind.
· In der vierten Gruppe finden sich die Hormone, die die Funktiondes Gastrointestinaltrakts regulieren.
· In der fünften Gruppe finden sich die Hormone, die in den Stoff-wechsel von Phosphat und Calcium eingreifen.
· In der sechsten Gruppe befinden sich die Hormone für die Regu-lation des Stoffwechsels von Wasser und Elektrolyten.
17.1 Hormone als extrazelluläre Botenstoffe 455

! Der molekulare Mechanismus der Hormonwirkungberuht auf der Wechselwirkung des Hormonsmit einem Rezeptorprotein.
Ein schon zu Beginn des Jahrhunderts formuliertes Konzept über denMechanismus der Hormonwirkung postuliert, daû Hormone ihreFunktion nur in solchen Zellen erfüllen können, die über einen ent-sprechenden, spezifischen Hormonrezeptor verfügen. Man kann imPrinzip zwei Typen von Hormonrezeptoren unterscheiden:
· Die Wechselwirkung der intrazellulären Hormonrezeptoren mitdem Hormon setzt die Aufnahme des Hormons in die Zielzellevoraus.
· Bei den Plasmamembranrezeptoren erzeugt die Bindung des Hor-mons an den Rezeptor ein intrazelluläres Signal, das gelegentlichauch als zweiter Botenstoff (engl. 2nd messenger) bezeichnet wird.
! Intrazellulär lokalisierte Hormonrezeptoren sindligandenaktivierte Transcriptionsfaktoren.
Eine Reihe von Hormonen, v. a. die Steroidhormone, die Schilddrü-senhormone, die Vitamin D-Hormone, aber auch die Vitamin A-Säu-
17 Biochemie und Pathobiochemie des endokrinen Systems456
ICytokine
(z.B. PDGF, EGF,FGF, IGF-1 u.a.)
IIWachstumshormon
Schilddrüsen-hormone
SexualhormoneGlucocorticoide
IIIInsulin
AdrenalinGlucagon
VParathormonD-Hormone
Thyreocalcitonin
IVgastrointestinale
Hormone(Secretin, Gastrin u.a.)
VIVasopressin
MineralocorticoidenatriuretischesAtriumpeptid
Abb. 17.2 Einteilung der Hormone nach funktionellen Zusammenhängen. (Einzel-heiten s. Text)

re (Retinsäure) wirken durch ¾nderung der Transcription spezifi-scher Gene. Ihr Wirkungsmechanismus beruht generell darauf, daûsie Liganden und Aktivatoren spezifischer Transcriptionsfaktorensind. Die meisten von ihnen bilden die Familie der Steroidrezeptoren.Das Prinzip ihres Wirkungsmechanismus ist in Abb. 17.3 am Beispielder Aktivierung des Glucocorticoid-Rezeptors (S. 388) dargestellt:
· Glucocorticoide sind gut durch die Plasmamembran ihrer Ziel-zellen permeabel.
· Glucocorticoide binden im Cytosol der Zielzellen an den Gluco-corticoid-Rezeptor, der dort inaktiv im Komplex mit dem Hitze-schockprotein Hsp 90 und einem Protein p 59 vorliegt.
· Bindung des Glucocorticoids an den Rezeptor löst die Abdisso-ziation von Hsp 90 und p 59 sowie die Dimerisierung des Rezep-tors aus.
· Der dimere, durch das Glucocorticoid aktivierte Rezeptor wirdin den Zellkern transloziert und bindet in der Promotorregionentsprechend regulierter Gene an spezifische Basensequenzen,sogenannte Enhancer-Sequenzen.
· Dies führt zu ¾nderungen der Expression der entsprechend re-gulierten Gene.
17.1 Hormone als extrazelluläre Botenstoffe 457
Zellkern
Cytosol
Hsp 90
p 59Gr
Hsp 90
Abb. 17.3 Aktivierung intrazellulärer Hormonrezeptoren durch Liganden. Die Ab-bildung zeigt den bei Glucocorticoiden aufgedeckten Mechanismus. Gr Glucocor-ticoidrezeptor; Hsp 90 Hitzeschockprotein Hsp 90; p59 Protein p59. (Einzelheitens. Text)

In Tabelle 17.1 findet sich eine Zusammenstellung von Hormonen,die den geschilderten Wirkungsmechanismus benutzen.
! Bindung von Hormonen an Rezeptorenin der Plasmamembran löst die Bildungintrazellulärer Signalkaskaden aus.
Die Mehrzahl von Hormonen bzw. Signalstoffen wirkt über in derPlasmamembran lokalisierte Rezeptoren. Diese lassen sich in dieGruppen einteilen, die in Tabelle 17.2 dargestellt sind:
Ligandenaktivierte Ionenkanäle: Entsprechende Ionenkanäle werdendurch die jeweiligen Liganden (Hormone, Transmitter etc.) geöffnetoder geschlossen. Eine Signalweiterleitung in den intrazellulärenRaum ist nicht notwendig.
17 Biochemie und Pathobiochemie des endokrinen Systems458
Tabelle 17.1 Hormone als Aktivatoren von ligandenaktivierten Transcriptionsfak-toren (Auswahl)
Gruppe Hormone Seite
Steroidhormone GlucocorticoideMineralocorticoideProgesteronEstrogeneAndrogene
479500477477473
Schilddrüsenhormone Trijodthyronin 468
Vitamin D-Hormone 1,25-Dihydroxycholecalciferol 496
Retinoate all-trans-Retinoat9-cis-Retinoat
591591
Tabelle 17.2 Einteilung der Plasmamembranrezeptoren
Rezeptortyp Signaltrans-duktion
Beispiel Seite
LigandenaktivierteIonenkanäle
Keine Nicotinischer Acetylcholin-rezeptor
7-Transmembrando-mänen-Rezeptoren
HeterotrimereG-Proteine
b-adrenerge Rezeptorena1-adrenerger Rezeptor
484485
Tyrosinkinase-Rezeptoren
Proteine mitSH2-Domänen
InsulinrezeptorWachstumsfaktorrezeptoren
490460
Cytokinrezeptoren JAK's, STAT's Wachstumshormonrezeptoren 461
Guanylatcyclasen Keine ANF-Rezeptor 462

Rezeptoren mit sieben Transmembrandomänen: Derartige Rezepto-ren haben als Gemeinsamkeit, daû sie mit 7 Transmembrandomänenin der Plamamembran verankert sind. Für ihre Signaltransduktionbenötigen sie immer heterotrimere G-Proteine (Abb. 17.4):
· Der durch das Hormon aktivierte Rezeptor führt an dem aus ei-ner a-, b- und g-Untereinheit bestehenden G-Protein zum Aus-tausch von GDP gegen GTP an der a-Untereinheit sowie zur Dis-soziation.
· Die GTP-beladene a-Untereinheit kann, je nach ihrer Spezifitätentweder die Adenylatcyclase aktivieren oder hemmen oderaber die Phospholipase Cb aktivierten.
· Als intrazelluläre Signale (zweite Botenstoffe) entstehen dabeientweder cAMP oder durch Spaltung von Phosphatidylinositol-
17.1 Hormone als extrazelluläre Botenstoffe 459
Zellmembran
αGDP
β γ
αGTP
β γ
GDP
GTP
Pi
7-Trans-membran-Domänen-Rezeptor
Ligand
Aktivierung(Hemmung)
derAdenylat-
cyclase
Aktivierungder
PhospholipaseCβ
oder
Abb. 17.4 Molekularer Mechanismus der Signaltransduktion von 7 Transmem-brandomänen-Rezeptoren. (Einzelheiten s. Text)
Tabelle 17.3 Effekte von cAMP, Inositoltrisphosphat und Diacylglycerin
Effekt Folge
cAMP Aktivierung der Protein-kinase A
Phosphorylierung und Aktivie-rung/Inaktivierung regulierterEnzyme (z. B. Glycogenstoff-wechsel, Lipolyse etc.)
Inositol-trisphosphat
Aktivierung des liganden-aktivierten Calciumkanalsim ER
Erhöhung der cytosolischen Cal-ciumkonzentration, Aktivierungvon Zellen
Diacylglycerin Aktivierung der Protein-kinase C
Phosphorylierung von Rezepto-ren, Transcriptionsfaktoren u. a.
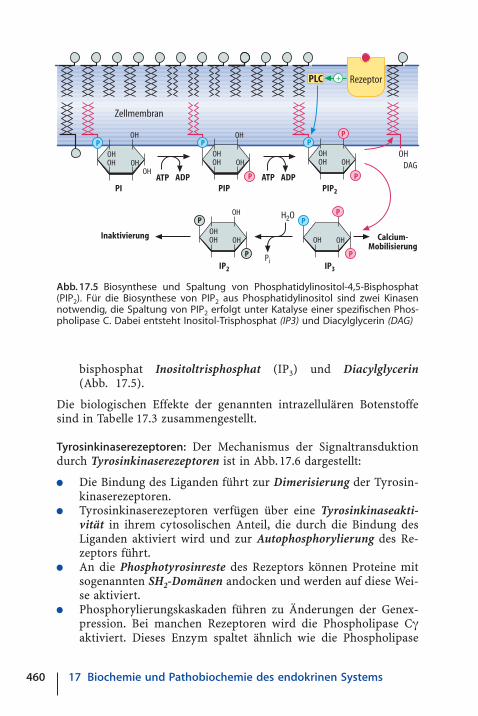
bisphosphat Inositoltrisphosphat (IP3) und Diacylglycerin(Abb. 17.5).
Die biologischen Effekte der genannten intrazellulären Botenstoffesind in Tabelle 17.3 zusammengestellt.
Tyrosinkinaserezeptoren: Der Mechanismus der Signaltransduktiondurch Tyrosinkinaserezeptoren ist in Abb. 17.6 dargestellt:
· Die Bindung des Liganden führt zur Dimerisierung der Tyrosin-kinaserezeptoren.
· Tyrosinkinaserezeptoren verfügen über eine Tyrosinkinaseakti-vität in ihrem cytosolischen Anteil, die durch die Bindung desLiganden aktiviert wird und zur Autophosphorylierung des Re-zeptors führt.
· An die Phosphotyrosinreste des Rezeptors können Proteine mitsogenannten SH2-Domänen andocken und werden auf diese Wei-se aktiviert.
· Phosphorylierungskaskaden führen zu ¾nderungen der Genex-pression. Bei manchen Rezeptoren wird die Phospholipase Cgaktiviert. Dieses Enzym spaltet ähnlich wie die Phospholipase
17 Biochemie und Pathobiochemie des endokrinen Systems460
PIADPATP
PIPADPATP
PIP2
IP3IP2
Inaktivierung Calcium-Mobilisierung
PLC
Zellmembran
DAGOHOH
OHOH
OH OHOH
OH OH
H2O
Pi
OHP P
P
PP
P
OH OH
Rezeptor
OH OHOH
OHP
P
OH OHOH
PP
P
Abb. 17.5 Biosynthese und Spaltung von Phosphatidylinositol-4,5-Bisphosphat(PIP2). Für die Biosynthese von PIP2 aus Phosphatidylinositol sind zwei Kinasennotwendig, die Spaltung von PIP2 erfolgt unter Katalyse einer spezifischen Phos-pholipase C. Dabei entsteht Inositol-Trisphosphat (IP3) und Diacylglycerin (DAG)

Cb Phosphatidylinositol-Bisphosphat und erzeugt damit Inosi-toltrisphosphat und Diacylglycerin (S. 460).
Cytokinrezeptoren: Cytokinrezeptoren bilden eine Familie von Hor-monrezeptoren, die gewisse strukturelle ¾hnlichkeiten mit den Tyro-sinkinaserezeptoren haben, sich jedoch von diesen durch das Fehleneiner Tyrosinkinaseaktivität in ihrem cytoplasmatischen Anteil un-terscheiden. Abbildung 17.7 gibt den für sämtliche Rezeptoren dieserGruppe gültigen Signaltransduktionsmechanismus wieder:
· Die Bindung des Liganden löst die Dimerisierung des Rezeptorsaus.
· Der dimerisierte Rezeptor kann tyrosinspezifische Proteinkina-sen, sog. Jak's binden.
· Jak's phosphorylieren spezifische Tyrosylreste der Rezeptoren,an die anschlieûend über SH2-Domänen sogenannte STAT's(engl. signal transducers and activators of transcription) binden.
17.1 Hormone als extrazelluläre Botenstoffe 461
PM
RAF
SOS
MAPK
MAPKK
Ras-GDP
Kern
PMRas
DAGPIP2
IP3
PI-3-P2
PIPP
MAPKK
GTPRAF
KernÄnderung der Transcription
PMAPKPLCγ
PI3-K PI3-K
PLCγGAP
GAPGRB2
GRB2
PP
P
SOS
P
a b
L
Abb. 17.6 a, b Intrazelluläre Signalkette nach ligandeninduzierter Phosphorylie-rung von Tyrosylresten der Tyrosinkinaserezeptoren. a Ohne Liganden liegendie Rezeptoren in monomerer Form vor, im Cytosol befinden sich eine Reihevon Proteinen, die nach Aktivierung mit dem Rezeptor interagieren können.b Die Bindung des Liganden löst die Dimerisierung des Rezeptors aus. Über soge-nannte SH 2-Domänen können eine Reihe von Proteinen an Phosphotyrosylrestedes Rezeptors andocken. Eines der Adapterproteine in GRB2 bindet ein Guaninnu-cleotid releasing Protein, SOS, welches das kleine G-Protein Ras aktiviert. Hieranschlieût sich eine Kaskade von Proteinkinasen an, die eine ¾nderung der Genex-pression und die Auslösung von Proliferation zur Folge hat. MAPKK Map-Kinase-Kinase; MAPK MAP-Kinase; MAP mitogenaktivierte Proteinkinase

· Nach Phosphorylierung dimerisieren die STAT's und werden inden Kern transloziert, wo sie die Transcription spezifischerGene verändern.
Membrangebundene Guanylatcyclase: Die membrangebundene Gua-nylatcyclase ist ein monomeres Transmembranprotein, auf dessen ex-trazellulärer Seite die Bindungsstelle für aktivierende Liganden liegt.Nur wenn diese gebunden haben, erfolgt eine cGMP-Produktion ausGTP durch das Enzym. Liganden, die die membrangebundene Guany-latcyclase aktivieren können, sind die sog. Guanyline. Zu ihnen gehörtu. a. das natriuretische Atriumpeptid (S. 503). cGMP aktiviert cGMP-abhängige Proteinkinasen und cGMP-abhängige Natriumkanäle.
Auûer der membrangebundenen Guanylatcyclase kommt in vie-len Zellen eine lösliche Guanylatcyclase vor. Dieses dimere Enzymwird durch Nitroxid (NO) aktiviert. Nitroxid ist ein erst in der jüng-sten Zeit entdecktes intra- und interzelluläres Signalmolekül. Es ent-steht enzymatisch unter Katalyse der Nitroxidsynthase aus Arginin.
!WICHTIGHormone regulieren in vielzelligen Organismen Stoffwechsel-leistungen, Wachstum und Differenzierung. Glanduläre Hor-mone werden in spezifischen endokrinen Drüsen gebildet, Cy-tokine und Gewebshormone von endokrin aktiven, in ver-schiedenen Geweben verteilten Zellen.
17 Biochemie und Pathobiochemie des endokrinen Systems462
Jak
STAT
Jak Jak
L
PP
Jak Jak
P
STAT
P
STAT
L
Iak Iak
PP
Jak Jak
P
STAT
P
STAT
STAT
L
Phosphorylierungdes Rezeptors
Anlagerung undPhosphorylierung
der STATs
Dimerisierungder STATs
Translokationin den Kern
DimerisierungAnlagerung der
Jak's
Abb. 17.7 Signaltransduktionsmechanismus der Cytokinrezeptoren. (Einzelheitens. Text)

Hormone stellen extrazelluläre Signale dar, die eine intrazellu-läre Antwort auslösen. Sie müssen deswegen von Zellen durchspezifische Rezeptoren erkannt und gebunden werden. DerartigeRezeptoren können intrazellulär lokalisiert sein und sind dannmeist durch Hormone (Liganden) aktivierte Transcriptionsfakto-ren. Die gröûere Gruppe von Hormonrezeptoren sind Proteineder Plasmamembran. Sie lassen sich in Liganden-aktivierte Io-nenkanäle, Rezeptoren mit 7 Transmembrandomänen, Tyrosinki-naserezeptoren und Cytokinrezeptoren einteilen. Jedem dieserHormontypen kommt ein spezifischer Signaltransduktionswegzu.
17.2 Regulation von Wachstumund Differenzierung
! Cytokine regulieren als lokal wirkende FaktorenWachstum und Differenzierung spezifischer Zellenund Gewebe.
Unter dem Begriff Cytokine faût man eine groûe Zahl von Wachs-tums- und Differenzierungsfaktoren (bis heute weit über 100) zusam-men, die von den verschiedensten Zellen freigesetzt werden könnenund überwiegend, allerdings nicht ausschlieûlich, als lokal wirkendeFaktoren Wachstum und Differenzierung spezifischer Zellen und Ge-webe regulieren (Tabelle 17.4):
· Eine Reihe sogenannter Wachstumsfaktoren (bFGF, EGF, PDGF)wirken als mitogene Faktoren auf viele Zellarten, besonders je-doch auf mesenchymale Zellen. Sie sind u. a. wichtig für Wund-heilung.
· Die groûe Gruppe der Interleukine hat mitogene und differenzie-rende Wirkung, vor allem an Lymphocyten und Makrophagenund spielen bei der Regulation der Immunantwort (S. 547) einebedeutende Rolle.
· Erythropoietin wird von Nieren und Hepatocyten abgegeben undstimuliert die Erythropoese im Knochenmark.
· TGFb ist ein Wachstumsinhibitor für viele Zellen.
· TNFa hat eine cytotoxische Wirkung auf manche Tumorzellen,ist jedoch für andere Zellen ein Mitogen
17.2 Regulation von Wachstum und Differenzierung 463

! Für Wachstum und Differenzierung verantwortlicheHormone werden durch ein hierarchisches Systemglandotroper Hormone reguliert.
Für die eigentlichen, von endokrinen Drüsen gebildeten und Wachs-tum und Differenzierung regulierenden Hormone spielt das inAbb. 17.8 dargestellte Regulationssystem aus Hypothalamus und Hy-pophysenvorderlappen eine bedeutende Rolle:
· Aus dem Hypothalamus werden unter dem Einfluû neuraler oderhumoraler Stimuli Freisetzungshormone (Releasing Hormones,Liberine) abgegeben.
17 Biochemie und Pathobiochemie des endokrinen Systems464
Tabelle 17.4 Cytokine (Auswahl)
Bezeichnung Produziert von Rezeptorfamilie Wirkung
bFGF(basic fibroblastgrowth factor)
Mesodermale undneuroektodermaleZellen, Endothel-zellen
Tyrosinkinase Mitogen für vieleZellen, Modulatorder Zelldifferen-zierung
EGF(epidermalgrowth factor)
Hirn, Niere,Speicheldrüse
Tyrosinkinase Mitogen für vieleZellarten
EPO(erythropoietin)
Nieren,Hepatocyten
Cytokinrezeptor Stimulierung derErythropoese
IL 1±15(Interleukine1±15)
VerschiedeneKlassen vonLeukocyten
Cytokinrezeptor Mitogene unddifferenzierendeWirkung v. a. aufLymphocyten undMakrophagen
PDGF(platelet derivedgrowth factor
Megakaryocyten,Makrophagen,Endothelzellen
Tyrosinkinase Mitogene Wirkungfür mesenchymaleZellen
TGFb(transforminggrowth factor b)
Megakaryocyten,Makrophagen,Lymphocyten
eigene Familie Wachstumsinhibi-tor für viele Zellen
TNF a(tumor necrosisfactor a)
Makrophagen,Fibroblasten,glatte Muskelzellen
Cytokinrezeptor Mitogen für Fibro-blasten, Cytolysevon Tumorzellen invitro, chemotakti-sche Wirkung
"Abb. 17.8 Prinzip der Regulation des hypothalamisch-hypophysären Systems undseiner peripheren Zielgewebe

matostatin notwendig (Abb. 20.21). Die Sekretion von Pepsinogendurch die Hauptzellen des Magenfundus wird durch cholinerge Reizesowie ebenfalls durch Gastrin stimuliert. Die Mucinproduktion durchdie Mucinzellen des Magens wird durch cholinerge Reize, Sekretinund Prostaglandin E1 stimuliert sowie durch Glucocorticoide ge-hemmt.
20 Ernährung, Verdauung, Resorption610
Tabelle 20.13 Gastrointestinale Peptidhormone und Neurotransmitter (Auswahl)
Bezeichnung Aminosäure-reste
Vorkommen Wichtigste Funktion
HormoneGastrin 17 bzw. 34 Antrum des
Magens, oberesDuodenum
Stimulierung derHCl-Sekretion
Sekretin 27 Duodenum,Jejunum
Stimulierung derpankreatischenHCO�
3-SekretionCholecystokinin/Pankreozymina
33 Duodenum,Jejunum
Stimulierung derpankreatischenEnzymsekretionKontraktion derGallenblase
Gastroinhibitori-sches Peptid (GIP)
43 Duodenum bisoberes Jejunum
Stimulierung derInsulinsekretion
Motilin 22 Oberes Jejunum,Duodenum
Stimulierung derMotilität von Magenund Dünndarm
Neurotensin 13 Unterer Dünn-darm, Colon
Stimulierung derSekretion von Insulin,Glucagon, Gastrin
Enteroglucagon ~ 70 Ileum und Colon Trophischer Faktor fürEpithelzellen desIntestinaltraktes
Somatostatina 14 GesamterIntestinaltrakt,Pankreas
Hemmung sekretori-scher Vorgänge
NeurotransmitterVasoaktives inte-stinales Peptida
14/28 Neurone undNervenfasern desIntestinaltrakts
Vasodilatation, Rela-xation der glattenMuskulatur
Substanz Pa 11 GesamterIntestinaltrakt
Kontraktion der glat-ten Muskulatur
Bombesina 14 Magen,Duodenum,Jejunum
Pankreassekretion?
Enkephalina 5 GesamterIntestinaltrakt
?
a Vorkommen im Zentralnervensystem gesichert.

Regulation der Pankreassekretion: Für die Bildung des Pankreassekre-tes sind nervale und endokrine Faktoren verantwortlich (Abb. 20.22):
· Sekretin und das vasoaktive intestinale Peptid (VIP) stimulierenin den Gangzellen des Pankreas die Wasser- und Hydrogencarbo-natsekretion.
· Die Enzym- und Proenzymsekretion durch die Acinuszellen desPankreas wird auûer durch Acetylcholin (muscarinischer Acetyl-cholinrezeptor) durch Cholecystokinin/Pankreozymin, Sekretinsowie VIP stimuliert. Das erstere wirkt über eine Erhöhung dercytosolischen Calciumkonzentration durch Stimulierung desPhosphatidylinositol-Cyclus (S. 460), die beiden letzteren durchAktivierung des Adenylatcyclasesystems.
Regulation der Gallensekretion: Die Sekretion der Gallenflüssigkeitwird auf zweifache Weise reguliert:
· Substanzen, die die Gallensekretion durch Hepatocyten stimulie-ren, werden als Choleretica bezeichnet. Die wichtigsten Cholere-tica sind die Gallensäuren.
20.3 Verdauung und Resorption 611
PeptideAminosäuren
G-Zelle
Acetyl-Cholin GRP
Gastrin
Histamin
GRP
Somatostatin
Acetyl-CholinBelegzelleH+
H+
ECL-Zelle
D-Zelle
Abb. 20.21 Regulation der Salzsäureproduktion durch die Belegzellen. Für die Re-gulation der Salzsäureproduktion ist das Zusammenspiel cholinerger und pepti-derger Impulse mit dem aus den Gastrinzellen stammenden Gastrin, dem ausden ECL-Zellen stammenden Histamin, sowie den durch die D-Zellen gebildetenSomatostatin notwendig. GRP gastrin releasing peptide

· Cholecystokinin/Pankreozymin führt dagegen zu einer Kontrak-tion der Gallenblase mit Entleerung von Gallenflüssigkeit in dasDuodenum.
! Die Resorption der Kohlenhydrateerfolgt durch sekundär aktive, natriumabhängigeTransportsysteme.
Die Nahrungskohlenhydrate sind überwiegend Stärke, daneben Di-saccharide wie Saccharose und Lactose. Die Stärkespaltung erfolgthauptsächlich im Duodenum unter der Einwirkung der Pankreas-amylase, das Reaktionsprodukt ist ein Gemisch aus Maltotriose undMaltose. Für deren Spaltung sowie für die Spaltung von Saccharoseund Lactose sind eine Reihe von Disaccharidasen (Isomaltase, Malta-se, Lactase, Saccharase) verantwortlich, die im Bürstensaum der Mu-cosazelle lokalisiert sind.
In unmittelbarer Nachbarschaft zum Ort der Disaccharidspal-tung im Bürstensaum der Mucosazellen befinden sich die für die Mo-nosaccharidresorption zuständigen Transportsysteme. Die Einzelhei-
20 Ernährung, Verdauung, Resorption612
Lumen
EnzymeProenzyme
cAMPcAMP Ca 2+
AcinuszelleGangzelle
HCO3–
H2O
SekretinVIP
Cholecystokinin-Pankreozymin
Acetylcholin
EnzymeProenzyme
HCO3–
H2O
Abb. 20.22 Enzym-, Wasser- und Hydrogencarbonatsekretion durch Acinus- bzw.Gangzellen des Pankreas. VIP vasoaktives intestinales Peptid. (Einzelheiten s. Text)

ten der Monosaccharidaufnahme sind in Abb. 20.23 am Beispiel derGlucoseresorption zusammengestellt:
Auf der luminalen Seite findet sich ein Transportsystem, welchesGlucose im Symport mit Natriumionen in das Cytosol der Mucosa-zelle transportiert. Die Triebkraft für den dabei häufig notwendigenBergauf-Transport der Glucose gegen ein Konzentrationsgefälle istder steile Natriumgradient zwischen Darmlumen und dem cytosoli-schen Raum der Mucosazellen. Dieser wird durch die Aktivität derNa/K-ATPase aufrecht erhalten, die auf der Serosaseite der Mucosa-zellen lokalisiert ist. Für den Transport der Glucose aus dem Innerender Mucosazelle in den Extrazellulärraum genügt der Glucosetrans-porter GLUT 1 aus der GLUT-Familie (S. 125).
! Lipide werden nach Aufnahme in die Mucosazellenzu Chylomikronen verpacktund in die intestinalen Lymphbahnen sezerniert.
Unter Einwirkung der Pankreaslipase und einem als Colipase bezeich-neten Hilfsprotein werden die Triacylglycerine der Nahrung in ein Ge-misch von Monoacylglycerinen, Fettsäuren und Glycerin zerlegt. Mo-noacylglycerine, Fettsäuren und Gallensäuren bilden anschlieûend ge-mischte Mizellen, in die u. a. Cholesterin und fettlösliche Vitamineeingelagert werden. Am Bürstensaum der Mucosazellen zerfallen der-artige Mizellen und die Mizellen bildenden Lipide werden in die Mu-cosazelle aufgenommen. Dort erfolgt eine Resynthese von Triacylgly-cerinen (Abb. 20.24). Anschlieûend lagern sich die Triacylglycerine
20.3 Verdauung und Resorption 613
Na+
Nas+ Na i
+
Ks+
NaL+
Glus
GluGlu
ATP
ADP+ Pi
Na-Pumpe
GluL
Serosaseite Mucosazelle DarmlumenBürstensaum
Abb. 20.23 Natriumabhängiger Transportmechanismus für die Glucoseresorption.GluL bzw. Na+
L Glucose bzw. Na+ im Lumen; GluI bzw. Na+I Glucose bzw. Na+ intra-
zellulär; GluS bzw. Na+S bzw. K+
S Glucose, Na+ bzw. K+ auf der Serosaseite

zusammen mit Cholesterin und anderen resorbierten Lipiden mit demApolipoprotein B 48 zusammen, die dabei entstehenden Chylomikro-nen (S. 207) werden durch Exocytose in die Lymphbahn abgegeben.
! Proteine werden bis auf die Stufe von Aminosäurenund Oligopeptiden gespalten und die dabeientstehenden Bruchstücke mit Hilfe eines sekundäraktiven Transportsystems aufgenommen.
Bei der Proteinverdauung werden durch die verschiedenen Protea-sen der Verdauungssekrete Oligopeptide und freie Aminosäuren er-zeugt.
20 Ernährung, Verdauung, Resorption614
Lumen Lymph-bahn
R
R
R
O COH2C
HC O CO
O COH2C
R
OHH2C
OCOH2C
OHH2C
β-Monoacyl-glycerin
α-Monoacyl-glycerin
R
OHH2C
OCOH2C
OHHC
Glycerin
OHH2C
OHH2C
OHHC
3 R COO–
R
OHH2C
OCOH2C
OHHC
OHH2C
OHH2C
OHHC
R
OHH2C
OCO
OHH2C
HC
Acyl-CoA
Glycerin
OHH2C
OHH2C
OHHC
R
H2C
OCO
H2C
HC
ROCO
ROCO
Triacylglycerin
α-Glycerophosphat
Pi
ApolipoproteinB48
ATP
ADP
H2C
OH
H2C
HC
OPO3– –
OH
Triacyl-glycerin
B48
B48
B48
Triacyl-glycerin
B48
B48
B48
Pankreas-lipase
Isomerase
Pankreas-lipase
22%
5%
72%
Abb. 20.24 Intestinale Spaltung und Resynthese von Triacylglycerinen. (Einzelhei-ten s. Text)

Oligopeptide werden durch sekundär aktiven protonenabhängi-gen Transport in die Mucosazellen aufgenommen. Der hierfür not-wendige Protonengradient wird durch einen Na-H+-Austauscheraufrecht erhalten, der an die serosaseitig gelegene Na/K-ATPasegekoppelt ist. Intrazellulär werden Peptide auf die Stufe von Ami-nosäuren zerlegt und durch ein entsprechendes Transportsystemauf der Serosaseite wieder aus der Mucosazelle ausgeschleust(Abb. 20.25).
¾hnlich wie für Monosaccharide kommen auch für Aminosäurenspezifische Transportsysteme in der Mucosazelle vor, die einen se-kundär aktiven energieabhängigen Aminosäuretransport in die Mu-cosazelle ermöglichen. Ihr Transportmechanismus hat ¾hnlichkeitmit dem für Monosaccharide beschriebenen.
! Die Rückresorption von Wasserund Elektrolyten erfolgt zum gröûten Teilim Jejunum und im Colon.
Die Wasserrückresorption erfolgt gröûtenteils im Jejunum und Co-lon. Im Jejunum wird sie durch den Export von Natriumionen aufSerosaseite der Mucosazellen aufrecht erhalten, dessen Ausmaû vonder Resorption von Monosacchariden bzw. Aminosäuren abhängt(s. o.). Dies führt zum Entstehen eines osmotischen Gradienten zwi-schen intra- und extrazellulärem Raum, der dafür verantwortlich
20.3 Verdauung und Resorption 615
Lumen
H+ H+
Na+ Na+
ADP; Pi
ADPK+ K+
Na+
Peptid
Aminosäuren
Peptid
Amino-säuren
Abb. 20.25 Mechanismus der Aufnahme von Peptiden oder Peptidantibiotikadurch Mucosazellen. Der Transport erfolgt gegen ein Konzentrationsgefälle alsProtonencotransport. (Einzelheiten s. Text)