untersuchungen zur eignung entomopathogener und zur ... · kosmopolit ist, der alle tropischen,...
TRANSCRIPT

Untersuchungen zur Eignung entomopathogener
und zur Identifizierung hyperparasitärer Pilze als Antagonisten der Honigbienenmilbe
Varroa destructor
Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften der Fakultät für Biologie und Biotechnologie an der Internationalen Graduiertenschule Biowissenschaften der
Ruhr-Universität Bochum
Arbeitsgruppe Verhaltensbiologie und Didaktik der Biologie
vorgelegt von
Markus Holt
aus
Gladbeck
Bochum
Februar 2010

Investigations into the suitability of
entomopathogenic fungi and the identification of hyperparasitic fungi as antagonists of the honeybee
mite Varroa destructor
Dissertation to obtain the degree Doctor rerum naturalium (Dr. rer. nat.) Faculty of Biology and Biotechnology
International Graduate School of Biosciences Ruhr-University Bochum
SG Behavioural Biology and Didactics of Biology
submitted by
Markus Holt
from
Gladbeck
Bochum
February 2010

Gutachter:
Prof. Dr. Wolfgang H. Kirchner
Prof. Dr. Günter A. Schaub

Inhaltsverzeichnis
Inhaltsverzeichnis
1 Einleitung .................................................................................................. 7
1.1 Die westliche Honigbiene Apis mellifera und ihr Parasit Varroa
destructor ................................................................................................. 7
1.2 Ansätze und Probleme bisheriger Bekämpfungsstrategien .................... 12
1.3 Entomopathogene Pilze als Antagonisten von Varroa destructor ........... 14
1.4 Zielsetzung ............................................................................................. 19
2 Material und Methoden ........................................................................... 21
2.1 Versuchstiere und verwendete Pilzstämme ............................................ 21
2.2 Medien und Puffer .................................................................................. 22
2.3 Bestimmung optimaler Kulturbedingungen ............................................. 23
2.4 Bestimmung einer ausreichenden Oberflächensterilisationsdauer
gesammelter Milben ............................................................................... 23
2.5 Ermittlung geeigneter Konservierungsmethoden gesammelter Milben ... 24
2.6 Erfassung der Mortalität mit Pilzkonidien behandelter Bienenlarven ...... 25
2.7 Bestimmung der Mortalität mit Pilzkonidien behandelter Adultbienen
und Milben .............................................................................................. 27
2.8 Erfassung des Pilzwachstums aus behandelten Milben ......................... 30
2.9 Halbfreilandversuch an Apis mellifera-Völkern zur Wirkung
entomopathogener Pilze auf Adultbienen, Bienenlarven und -puppen
sowie Varroa destructor .......................................................................... 30
2.10 Erfassung der subletalen Effekte entomopathogener Pilze auf
Adultbienen ............................................................................................ 33
2.11 Erfassung des Einflusses entomopathogener Pilze auf das
Immunsystem von Adultbienen .............................................................. 37
2.12 Screening ausgewählter Völker auf potentiell spezifische
Varroapathogene Pilze ........................................................................... 39
2.13 Identifizierung von Pilzen im Pilzscreening ............................................. 40
2.14 Statistik und Parameter der grafischen Darstellung ................................ 44
3 Ergebnisse ............................................................................................. 46
3.1 Voruntersuchungen ................................................................................ 46
3.1.1 Kulturbedingungen ................................................................................. 46

Inhaltsverzeichnis
3.1.2 Wirkung unterschiedlicher Oberflächensterilisationszeiten auf das
Pilzwachstum aus Milben ....................................................................... 48
3.1.3 Einfluss der Konservierungsmethode der Milben auf das
Pilzwachstum ......................................................................................... 50
3.2 Mortalität mit Pilzkonidien behandelter Larven ....................................... 51
3.3 Mortalität mit Pilzkonidien behandelter Adultbienen ............................... 52
3.3.1 Lecanicillium muscarium: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 52
3.3.2 Metarhizium anisopliae: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 54
3.3.3 Beauveria bassiana: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 56
3.3.4 Paecilomyces fumosoroseus: Mortalität/Versuchsgruppe und
kumulierte Mortalität ............................................................................... 58
3.4 Mortalität mit Pilzkonidien behandelter Milben ....................................... 60
3.4.1 Lecanicillium muscarium: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 60
3.4.2 Metarhizium anisopliae: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 62
3.4.3 Beauveria bassiana: Mortalität/Versuchsgruppe und kumulierte
Mortalität ................................................................................................. 64
3.4.4 Paecilomyces fumosoroseus: Mortalität/Versuchsgruppe und
kumulierte Mortalität ............................................................................... 66
3.5 Pilzwachstum aus behandelten Milben ................................................... 68
3.5.1 Lecanicillium muscarium ........................................................................ 68
3.5.2 Metarhizium anisopliae ........................................................................... 69
3.5.3 Beauveria bassiana ................................................................................ 70
3.5.4 Paecilomyces fumosoroseus .................................................................. 71
3.6 Wirkung entomopathogener Pilze auf Adultbienen, Bienenlarven
und -puppen sowie Varroa destructor im Halbfreilandversuch ............... 72
3.6.1 Einfluss von Lecanicillium muscarium auf die Anzahl der Bienen
pro Volk .................................................................................................. 73
3.6.2 Einfluss von Lecanicillium muscarium auf die Anzahl der Larven
pro Volk .................................................................................................. 74

Inhaltsverzeichnis
3.6.3 Einfluss von Lecanicillium muscarium auf die Anzahl der Puppen
pro Volk .................................................................................................. 75
3.6.4 Einfluss von Lecanicillium muscarium auf den Milbenfall pro Tag
und Volk ................................................................................................. 76
3.6.5 Einfluss von Lecanicillium muscarium auf den Milbenendbefall
adulter Bienen ........................................................................................ 77
3.6.6 Einfluss von Lecanicillium muscarium auf den Milbenendbefall
verdeckelter Brut .................................................................................... 78
3.7 Subletale Effekte entomopathogener Pilze auf Adultbienen ................... 79
3.7.1 Ernährungszustand im Langzeit-in-vitro-Assay ...................................... 79
3.7.2 Nahrungsaufnahme im Langzeit-in-vitro-Assay ...................................... 80
3.7.3 Lernleistung von Adultbienen im PER-Assay ......................................... 82
3.7.4 Diskriminationsfähigkeit von Adultbienen im Langzeit-PER-Assay......... 84
3.7.5 Einfluss entomopathogener Pilze auf das Immunsystem von
Adultbienen ............................................................................................ 86
3.8 Sreening ausgewählter Völker auf potentiell spezifische
Varroapathogene Pilze ........................................................................... 88
3.8.1 Populationsdynamik und Milbenbefall von Apis mellifera-Völkern .......... 89
3.8.2 Korrelation der Abundanz von Varroa destructor in Bienenvölkern
und des Pilzwachstums aus Milben ........................................................ 92
3.8.3 Spektrum identifizierter Spezies im Pilzscreening .................................. 95
3.8.4 Charakteristika von Meira geulakonigii ................................................. 101
3.8.5 Charakteristika von Simplicillium lamellicola ........................................ 104
4 Diskussion ............................................................................................ 109
5 Zusammenfassung ............................................................................... 126
6 Summary .............................................................................................. 128
7 Literatur ................................................................................................ 130

Einleitung
7
1 Einleitung
1.1 Die westliche Honigbiene Apis mellifera und ihr Parasit Varroa
destructor
Die westliche Honigbiene Apis mellifera LINNAEUS (Hymenoptera) ist eine von
neun Arten der Gattung Apis und unter ihnen die einzige, die außerhalb des
Sippenzentrums Asien verbreitet ist. Ursprünglich im nördlichen Afrika vorkommend,
dehnte sie postglazial in prähistorischer Zeit ihr Verbreitungsgebiet in den
eurasischen Raum aus (Whitfield et al. 2006). In historischer Zeit erfolgte eine
anthropogen bedingte weltweite Verbreitung, so dass Apis mellifera inzwischen ein
Kosmopolit ist, der alle tropischen, subtropischen und gemäßigten Klimate besiedelt
und nur in den Polregionen fehlt. Im Zusammenhang mit der weltweiten
Industrialisierung der Landwirtschaft nahm ihre ökonomische Bedeutung als
Bestäuber wichtiger menschlicher Nutzkulturen gegenüber der Bedeutung der
traditionell genutzten Produkte (Honig, Wachs) enorm zu. Beispielsweise ist allein die
kalifornische Mandelindustrie auf die Bestäubung durch eine Million Bienenvölker
angewiesen (Ratnieks et al. 2010). Inzwischen wird der weltwirtschaftliche Wert, der
durch die erbrachten Bestäubungsleistungen entsteht, auf 153 Milliarden Euro pro
Jahr (Angabe für 2005) geschätzt, entsprechend 9,5 Prozent des Wertes der
jährlichen Weltagrarproduktion an Lebensmitteln (Gallai et al. 2008). Auf Obst- und
Gemüsekulturen entfallen dabei jeweils 50 Milliarden Euro p.a.; die Bedeutung für
Kulturen essbarer Ölfrüchte wird auf 39 Milliarden Euro p.a. taxiert (Southwick et al.
1992; Gallai et al. 2008).
Ihrer wirtschaftlichen Bedeutung entsprechend stellt der Befall mit Parasiten
eine latente Gefahr für Bienenvölker in Kultur dar; dabei ist die intrakolonial hohe
Dichte verwandter Individuen sowie die in der Regel ebenfalls hohe Dichte von
Bienenvölkern in menschlicher Obhut ein die Ausbreitung von Pathogenen prinzipiell
begünstigender Faktor. Apis mellifera wird von einigen, inzwischen gut untersuchten
Parasiten befallen: Paenibacillus larvae ASH EMEND. GENERSCH ist ein ausschließlich
die Bienenbrut befallendes Pathogen, ebenso der Pilz Ascosphaera apis OLIVE AND
SPILTOIR, während Aspergillus flavus LINK neben Bienenbrut auch andere
Organismen, aber nur sehr selten adulte Bienen befällt. Das Grampositive Bakterium

Einleitung
8
Paenibacillus larvae beispielsweise ist in einer Dosis von nur etwa zehn Sporen
infektiös für junge Larven, die es mit dem Futter aufnehmen. Die Proliferation findet
im Mitteldarm der Larven ohne sichtbare Gewebszerstörungen statt, wie Yue mit
FISH (fluorescence-in-situ-hybridization)-Analysen zeigen konnte (Yue et al. 2008).
Während die Bakterien in diesem Stadium als Kommensalen im Wirt leben, werden
später Proteasen exprimiert, die lytisch auf interzelluläre Strukturen wirken und den
Bakterien nach Degradation des Darmepithels einen Übertritt ins Hämozoel
ermöglichen. In der Folge bildet sich durch Auflösung der Wirtsgewebe eine
fadenziehende Masse, die zu einem Schorf eintrocknet, der von Arbeiterinnen
aufgenommen wird, die so zur Verbreitung des Pathogens beitragen. Verflug
zwischen eng zusammenstehenden Völkern bedingt dann die schnelle Ausbreitung
der Infektion auch interkolonial. Während die Transmission von Paenibacillus larvae
horizontal stattfindet, verläuft sie bei allen anderen Parasitosen von Apis mellifera
vertikal – wie auch bei Melissococcus plutonius BAILEY AND COLLINS, einem
Pathogen, das eine vergleichbare Symptomatik zeigt, aber auf Kolonieebene in der
Regel nicht letal wirkt. Im Gegensatz zu den beiden vorgenannten Pathogenen
befallen die Sporidien Nosema apis ZANDER und Nosema ceranae FRIES nur adulte
Bienen; hier erfolgt die Verbreitung durch von infizierten Arbeiterinnen im Volk
abgegebenen Kot, der bei seiner Beseitigung durch bis dahin nicht betroffene
Imagines zu deren Ansteckung führt. Nosemainfektionen verlaufen nicht letal – dies
gilt auch für den Befall mit einem weiteren Vertreter der Protozoa, Malpighamoeba
mellificae PRELL.
Im Wesentlichen sind die Wirt-Parasit-Beziehungen zwischen Apis mellifera und
ihren Pathogenen stabil: Das Immunsystem des einzelnen Individuums ebenso wie
supraindividuelle Adaptionen gegenüber Pathogenen schließen Totalverluste auf
Volksebene in der Regel aus. Das gilt auch für den Befall mit Acarapis woodi RENNIE,
einer Milbe, die die thorakalen Tracheen adulter Bienen besiedelt und bei massivem
Befall durch Einschränkung der Respirationsfähigkeit zur Flugunfähigheit der Tiere
führt.
Mitte des 20. Jahrhunderts kam es zu ersten Beobachtungen eines bis dahin
auf Apis mellifera unbekannten Parasiten: 1952 wurde die Milbe Varroa destructor
ANDERSON AND TRUEMAN; (Mesostigmata: Varroidae) erstmals auf Apis mellifera in
den östlichen Küstenregionen der damaligen UdSSR entdeckt (bis 2000: Varroa
jacobsoni OUDEMANS (Anderson et al. 2000)). In der Folge kam es sowohl zu einer

Einleitung
9
natürlichen Migration Richtung Westen als auch zu anthropogen bedingten
Verschleppungen. Mitte der 50er Jahre des 20. Jahrhunderts wurde Varroa in
Pakistan gefunden, 1967 erreichte sie Bulgarien und 1977 Westdeutschland (Ritter
1982). Die Milbe ist inzwischen mit Ausnahme der Polregionen sowie Australiens
weltweit verbreitet.
Varroa destructor ist ein durchschnittlich 1 Millimeter langer und 1,5 Millimeter
breiter Vertreter der Mesostigmata und damit – gemessen an der Größe des Wirtes –
der größte bekannte Ektoparasit. Der gesamte Lebenszyklus ist an den Wirt
gebunden und gliedert sich in zwei Phasen: Während der phoretischen Phase kommt
es auf adulten Bienen zur Ausbreitung zwischen Völkern (Kuenen et al. 1997). Als
obligater Parasit ist Varroa hochadaptiert in Bezug auf die Ausbeutung von
Wirtssignalen: Kirchner konnte erstmals nachweisen, dass die Milben in der Lage
sind, Lichtreize und Vibrationen wahrzunehmen; derartige Vibrationen sind ein häufig
verwendetes Signal in Apis mellifera-Staaten (Kirchner 1987, 1993; Kirchner et al.
1986). In der phoretischen Phase halten sich die Milben auf den Bienen bevorzugt
zwischen den überlappenden Sterniten auf und ernähren sich von der Hämolymphe
des Wirtes (Sammataro et al. 2000). Während der reproduktiven Phase migrieren
adulte Weibchen in Brutzellen des Wirtes kurz vor deren Verdeckelung. Hierbei sind
olfaktorische Signale von zentraler Bedeutung. Die Milben sind vermutlich aufgrund
der distinkten Kohlenwasserstoffbouquets von Arbeiterinnen in der Lage, nach dem
Verlassen einer Brutzelle gezielt auf Bienen derjenigen Alterskohorte zu wechseln,
die sich innerhalb des Volkes hauptsächlich um die Brutpflege kümmert, so dass die
Wahrscheinlichkeit maximiert wird, in die unmittelbare Nähe von Brutzellen zu
gelangen (Kraus 1993; Chiroudi et al. 1997; Kuenen et al. 1997). Zellen, in denen
Drohnen aufgezogen werden, zeigen einen acht- bis zehnfach höheren Befall als
Arbeiterinnenzellen, was zum einen in der höheren Frequenz begründet liegt, mit der
Drohnenbrutzellen durch Arbeiterinnen aufgesucht werden, zum anderen in der
längeren Puppenruhe männlicher Bienen (Boot et al. 1995). Die Identifikation
geeigneter Brutzellen, i.e. Zellen, in denen sich eine Larve im fünften Larvenstadium
15 - 20 Stunden (Arbeiterinnen) bzw. 40 - 50 Stunden (Drohnen) vor der
Verdeckelung der Zelle befindet, wird ebenfalls durch das Differenzierungsvermögen
von Varroa gegenüber kutikulären Kohlenwasserstoffen des Wirtes ermöglicht.
Garrido konnte an Honigbienen erstmals für Insekten zeigen, dass es einen
kairomonalen Effekt polarer, kutikularer Fraktionen von Larven unmittelbar nach der

Einleitung
10
Verdeckelung auf die Initiation der Oogenese von Varroa gibt (Garrido et al. 2004).
Drei Tage nach Verdeckelung wird ein erstes Ei gelegt, aus dem sich innerhalb von
sechs Tagen ein adultes Männchen entwickelt (Steiner et al. 1994). Aus den
folgenden Eiern entwickeln sich Weibchen, die noch in der Zelle begattet werden.
Die Nymphenstadien ernähren sich während der Entwicklung von der Hämolymphe
der Bienenlarve, die einer „feeding zone“ genannten, vom adulten Weibchen
geschaffenen Öffnung der Puppenkutikula des Wirtes entnommen wird (Donzé et al.
1994). Ein bis zwei Tochtermilben gelangen zur Reife und verlassen mit der
Muttermilbe und der schlüpfenden Biene die Zelle. Martin ermittelte für
Arbeiterinnenbrutzellen eine Reproduktionsrate zwischen 1,3 und 1,5, für
Drohnenbrutzellen eine Rate zwischen 2,2 und 2,6 (Martin 1994a, 1994b).
Ursprünglicher Wirt der Milbe war die östliche Honigbiene Apis cerana
FABRICIUS. Ihr Verbreitungsgebiet umfasst den Indischen Subkontinent, Südostasien
sowie Teile Chinas und des östlichen Russlands. Neben Apis mellifera ist sie der
einzige von Menschen als Nutztier gehaltene Vertreter der Gattung. Die genauen
Umstände des Wirtswechsels sind nicht bekannt. Als wahrscheinlich wird
angenommen, dass es während einer sympatrischen Phase nach Verbringen von
Völkern der westlichen Honigbiene nach Ostasien und damit ins Verbreitungsgebiet
des ursprünglichen Wirtes in der ersten Hälfte des 20. Jahrhunderts, die zunächst für
einen unbekannten Zeitraum eine Koexistenz der beiden Apisarten ohne
Wirtswechsel beinhaltete, zu einem Wirtswechsel in diesem Gebiet kam (Oldroyd
1999).
Die Gattung Varroa umfasst vier Arten; innerhalb der Art Varroa destructor
werden sechs Haplotypen unterschieden, von denen vier Apis mellifera parasitieren.
Zwei dieser Haplotypen, der japanisch-thailändische und der koreanische sind
hochpathogen für Apis mellifera (De Guzman et al. 1999). Der Befall von Völkern der
westlichen Honigbiene mit diesen Haplotypen führt – mit Ausnahme von
Populationen in Südamerika und vermutlich einigen Regionen in Afrika – obligat
innerhalb von längstens drei bis vier Jahren zum Tod der Völker (Beetsma 1994).
Ursächlich hierfür sind Vitalitätsverluste durch den Verlust von Hämolymphe während
der Ontogenese, was zu einem reduzierten Gewicht der adulten Tiere führt; diese
Vitalitätsverluste bedingen zum Beispiel bei Drohnen eine reduzierte Flugleistung
(Duay et al. 2002). Parasitierte Adulti von Apis mellifera zeigen aber nicht nur
signifikante Unterschiede in Bezug auf die Quantität ihrer Hämolymphe, auch die

Einleitung
11
Proteinkonzentration in der Hämolymphe sinkt, und die Quotienten hoch- und
niedermolekularer Fraktionen differieren in Abhängigkeit vom Parasitierungsgrad
(Weinberg et al. 1985). Die durchschnittliche Lebensdauer von Arbeiterinnen
reduziert sich in Korrelation zum steigenden Brutbefallsgrad stark parasitierter
Kolonien im Spätsommer um bis zu 50 Prozent (Ritter et al. 1984; Kovac et al. 1988).
Untersuchungen an Arbeiterinnen zeigten eine Verminderung der assoziativen
Lernfähigkeit und eine reduzierte Rückkehrrate zum Volk (Kralj et al. 2007).
Unabhängig von den Schäden durch den Entzug von Hämolymphe kommt es zu
Schäden durch Viren, als deren Vektor Varroa fungiert, nachgewiesen exemplarisch
für das Kashmir bee virus, das Sacbrood virus, das Israeli acute paralysis virus und
das Deformed wing virus (Boecking et al. 2008). Letzteres beispielsweise führt
während der Ontogenese zur Entwicklung nichtfunktionaler Flügel, so dass adulte
Tiere für den Staat essentielle Aufgaben nicht wahrnehmen können. Dabei ist das
Deformed wing virus auch für Varroa infektiös, ohne allerdings klinische Symptome
zu verursachen, die bekannt geworden wären: Strangspezifische RT-PCR-Ansätze
konnten zeigen, dass virenreplikationsassoziierte DNA auch in den Milben auftrat
(Gisder et al. 2009). Für das Israeli acute paralysis virus demonstrierte Ball in
Untersuchungen an britischen Apis mellifera-Kolonien, dass persistierende, für die
Bienen subletale Virusvorkommen mit steigendem Befall durch Varroa „aktiviert“ und
damit letal werden können (Ball 1989).
Letale Schäden durch Varroa treten bei Apis cerana nicht auf – Wirt und Parasit
befinden sich im evolutiven Gleichgewicht. So vermehrt sich Varroa aus bisher
unbekannten Gründen nicht in Arbeiterinnenbrut, mehrfach befallene Drohnenbrut
kommt nicht zum Schlupf, so dass die in die Zellen migrierten Milben und ihre
Nachkommen mit den parasitierten Drohnen sterben, und adulte Bienen sind in der
Lage, Varroamilben auf anderen Bienen und in Brutzellen zu erkennen und diese
durch Allo- und Auto-„grooming“ zu entfernen, beziehungsweise auszuräumen (Peng
et al. 1987; Boecking et al. 1999; Boot et al. 1999). Auf Volksebene trägt ein
„absconding“ genanntes Verhalten, bei dem relativ stark befallene Nester verlassen
werden und damit der Anteil der Milben, der sich in Brutzellen befindet, zurückbleibt,
zu einer Reduzierung der Parasitenlast bei. Den gleichen Effekt hat (für die das Nest
verlassende Kohorte) das Schwärmen; dieses zentrale Element der Vermehrung
aller hoch eusozialen Bienenarten wird bei von Menschen gehaltenen Völkern von

Einleitung
12
Apis mellifera aber zumindest teilweise unterdrückt, da es aus ökonomischer
Perspektive unerwünscht ist.
1.2 Ansätze und Probleme bisheriger Bekämpfungsstrategien
Das obligate Sterben von Varroa befallener Bienenvölker macht – um
wirtschaftliche Einbußen bis zur maximalen Größe des wirtschaftlichen
Gesamtnutzens der westlichen Honigbiene zu vermeiden – weltweit den Einsatz von
Bekämpfungsmaßnahmen erforderlich. 177 von Menschen genutzte Kulturen sind
ganz oder teilweise von der Bestäubung durch Honigbienen abhängig (Crane 1990).
Für die USA schätzt Levin den wirtschaftlichen Nutzen auf 19 Milliarden Dollar pro
Jahr, während Southwick 1,6 - 5,7 Milliarden Dollar angibt (Levin 1984; Southwick et
al. 1992). Für Neuseeland werden die durch Varroa verursachten jährlichen Schäden
auf 267 - 602 Milllionen US-Dollar pro Jahr taxiert (McNeely et al. 2001).
Klassische Bekämpfungsmittel bis in die jüngste Zeit waren Akarizide auf der
Basis von Organophosphaten, Pyrethroiden und Amidinen. Allerdings reichern sich
diese fettlöslichen Substanzen im Bienenwachs und damit letztendlich auch im Honig
an, was aus lebensmittelhygienischer Sicht unerwünscht ist (Martel et al. 2007).
Darüber hinaus kam es Mitte der 1990er Jahre zum Auftreten von Resistenzen
(Milani 1999).
Standard in der Bekämpfung von Varroa destructor sind zurzeit organische
Säuren, namentlich Ameisensäure, Oxalsäure und Milchsäure (Calderone 1999;
Nanetti et al. 2003). Sie haben den Vorteil, ohnehin natürlicher Bestandteil des
Honigs zu sein. Zum richtigen Zeitpunkt appliziert, treten in den Bienenprodukten
keine Residuen auf, die oberhalb des natürlicherweise vorhandenen Levels liegen.
Ihr Nachteil besteht darin, dass sie ein hohes Anwenderwissen voraussetzen:
Ameisensäure wirkt varroazid auch in verdeckelten Brutzellen, die Applikation von
Oxal- und Milchsäure ist demgegenüber nur in brutfreien Phasen der
Volksentwicklung erfolgversprechend; darüber hinaus ist die therapeutische Wirkung
von Ameisensäure stark von abiotischen Faktoren wie der Umgebungstemperatur
und -feuchte abhängig. Der zielgenaue Einsatz aller drei Säuren setzt außerdem
voraus, dass genaue Informationen zum Befallsgrad eines Volkes mit Varroa

Einleitung
13
destructor vorliegen. Zudem ist ihr therapeutischer Index gering: Die
Schadensschwellen für Varroa destructor und Apis mellifera liegen dicht beieinander;
Überdosierung können zum Totalverlust des Bienenvolkes führen, Unterdosierungen
sind unwirksam.
In jüngerer Zeit wurde versucht, ursprünglich bei Apis cerana identifizierte
Varroatoleranzmerkmale, i.e. das Entfernen infizierter Brut und das Allo- und
Autogrooming von Arbeiterinnen, die in geringem Ausmaß auch bei Apis mellifera
beobachtet wurden, durch gezielte Selektion auf Populations- und individueller
Ebene zu selektieren. Versuche mit Völkern vermeintlich varroatoleranter Herkünfte
aus Ostasien zeigten allerdings, dass die Varianz im Befall auf milbenspezifische
Faktoren zurückzuführen war (Harris et al. 2003). Versuche, in denen die
Pathogenität des in Südamerika vorkommenden Milben-Haplotyps in Bezug auf
Bienen dortiger, varroatoleranter Herkünfte (sogenannte „afrikanisierte“ Bienen, eine
Hybride aus europäischen und afrikanischen Unterarten von Apis mellifera) und
europäischer Herkünfte getestet wurden, wiederum kamen zu dem Schluss, dass es
offenbar auch bienenspezifische Toleranzfaktoren gibt; allerdings scheinen sie nicht
unabhängig von weiteren Parametern aufzutreten: Varroatolerante Bienen aus dem
tropischen Brasilien zeigten diese Toleranz in gemäßigten Klimaten nicht mehr – ein
Effekt, der möglicherweise auf unter diesen Bedingungen veränderte
Individuenstärken des einzelnen Volkes und eine stärkere, saisonal klimabedingte
Populationsdynamik des Bienenvolkes zurückzuführen ist (Rosenkranz 1999;
Correa-Marques et al. 2002). Angenommen werden auch höhere Fitnesskosten
eines ausgeprägten Groomingverhaltens in gemäßigten Klimaten (Vandame et al.
2002). Dem entgegen stehen Berichte über langfristig ohne Behandlung
überlebende, nicht bewirtschaftete Populationen auch in Europa (Le Conte et al.
2007). Unklar ist, ob es auch in bewirtschafteten Bienenvölkern zu einem mutmaßlich
stabilen Wirt-Parasit-Verhältnis kommen könnte: Sich selbst überlassene Völker in
Schweden zeigten eine geringere Zahl aufgezogenen Nachwuchses und es traten
vermehrt Brutkrankheiten auf – diese Faktoren können zum einen den beobachteten
verringerten Parasitenbefall erklären, sind zum anderen in Populationen in Kultur
unerwünscht und unterliegen damit Manipulationsversuchen durch die Bienenhalter;
es erscheint fraglich, ob unter diesen Bedingungen der gleiche Selektionsdruck
existiert, wie in unbeeinflussten Völkern und ob es somit zu analogen adaptiven
Prozessen kommt (Rosenkranz et al. 2005). Für die Annahme, bienenspezifische

Einleitung
14
Faktoren seien nicht unerheblich, spricht auch die Beobachtung, dass L5-Stadien
europäischer Honigbienen sich im Vergleich zu denen afrikanisierter Honigbienen
doppelt so attraktiv für Varroa zeigten (Guzman-Novoa et al. 1999). Da Aumeier
diesen Effekt in in vitro-Wahlversuchen aber nicht nachweisen konnte, spielen
offenbar auch hier bisher unbekannte Effekte auf supraindividueller Ebene eine Rolle
(Aumeier et al. 2002). Trotzdem wird versucht, mit Ansätzen auf individueller Ebene
Honigbienenarbeiterinnen, die ein ausgeprägtes Ausräum- und Putzverhalten zeigen,
zu fertilisieren und mithilfe des aus den männlichen Nachkommen gewonnenen
Spermas Eier dieser Arbeiterinnen in vitro zu befruchten, um Königinnen zu erhalten,
die eine hohe Heritabilität der gewünschten Merkmale zeigen (Bienefeld et al. 2007;
Büchler et al. 2008). Das gleiche Ziel verfolgen Microarrayversuche, die differentielle
Expressionsmuster von Puppen aus nicht varroatoleranten und toleranten Völkern
vergleichen. Navajas fand hier eine Korrelation der Expression die neuronale
Entwicklung steuernder Gene und des Grooming- bzw. Hygieneverhaltens (Navajas
et al. 2008). Bisher liegen zu diesen Versuchen allerdings noch keine Ergebnisse
vor, die ein Überleben von Honigbienen ohne Behandlung erhoffen lassen. Versuche
mit Micrococcacaeen und Bacillacaeen als Kontrollagentien haben sich bisher
ausschließlich auf Laborassays beschränkt (Tsagou et al. 2004).
1.3 Entomopathogene Pilze als Antagonisten von Varroa
destructor
Angesichts der nach wie vor evidenten Herausforderung, eine nachhaltige und
effiziente Bekämpfungsstrategie gegen Varroa destructor zu entwickeln, sind etwa ab
dem Jahr 2000 zunehmend auch entomopathogene Pilze als potentielle
Antagonisten von Varroa ins Blickfeld der Forschung gerückt. Auch wenn die
Lebenszyklen entomopathogener Pilze komplex sind und zahlreiche, auf einzelne
Gattungen beschränkte Besonderheiten ausweisen, gleichen sie sich in wesentlichen
Aspekten ihrer Biologie häufig: Aus Dauersporen, die im Fall von Zygosporen aus
Hyphenfusionen, im Fall von Azygosporen durch Abschnürung terminaler
Hyphenenden hervorgegangen sind, entwickeln sich in Phasen, in denen die Wirte
aktiv sind, infektionsfähige Konidien. Konidien werden ohne das Zwischenstadium

Einleitung
15
Dauerspore ebenfalls gebildet, wenn entomopathogene Pilze in ihren Wirten zur
Reife gekommen sind und es zu direkten Infektionen weiterer Wirte kommen kann.
Der exakte Mechanismus der Differenzierung zwischen Ziel- und
Nichtzielorganismen ist in Bezug auf Insekten noch unklar. Aus Studien an Pflanzen
wird geschlussfolgert, dass spezifische Induktoren exprimiert werden, die mit
membrangebundenen Rezeptoren von Wirtszellen interagieren und zur Expression
eines Produktes führen, das seinerseits an Rezeptoren auf der Oberfläche des
Pathogens bindet und über Messengermoleküle die Expression für die Infektion
essentieller Enzyme initiiert (Bölker 2002). Bei Kontakt zu Wirtsoberflächen kommt es
zur Bildung eines Appressoriums, aus dem ein Keimschlauch austritt, der die
Kutikula des Wirtes mechanisch oder histolytisch durchdringt; daran beteiligt sein
können trypsinähnliche Proteasen, Carboxypeptidasen, Aminopeptidasen,
Esterasen, Lipasen, Chitinasen und weitere Enzyme, die überwiegend aus
Untersuchungen an Metarhizium anisopliae METSCHNIKOW und Beauveria bassiana
BALSAMO bekannt sind (Leger et al. 1991, 1993, 1996, 1997; Poinar et al. 1998). Die
Bildung von Appressorien kann aber bei manchen Arten auch in vitro auf
nichtartifiziellen Oberflächen ausgelöst werden. In einigen wenigen Fällen dringt der
Parasit auch ohne vorherige Ausbildung eines Appressoriums ein (Brobyn et al.
1977). Lytische Enzyme sowie Lektine können, zum Beispiel im Fall von Metarhizium
anisopliae, bereits auf der Oberfläche der Konidien vorhanden sein; zwingende
Voraussetzung für die Keimung ist zudem das Vorhandensein von Kohlenstoff- und
Stickstoffquellen auf der Wirtskutikula. Zwar können unspezifische Kohlenstoff-
beziehungsweise Stickstoffquellen in vitro auch eine Keimung initiieren, qualitative
und quantitative Differenzen hierbei scheinen jedoch ebenfalls eine wichtige Rolle bei
der Wirtserkennung zu spielen (Andrews et al. 1997). Innerhalb des Wirtes werden
dann zellwandlose Protoplasten gebildet, die mutmaßlich vom Immunsystem des
Wirts nicht oder nur schlecht detektiert werden können, oder es kommt zur Bildung
zönozytischer Hyphen oder einzelliger „hyphal bodies“ (Beauvais et al. 1989). Der
Tod des Wirtes tritt durch Faktoren wie den Verbrauch seiner Stoffwechselreserven,
mechanische Zerstörung von Geweben, die Freisetzung von Toxinen wie Pyridin-2,6-
Dicarbonsäure oder eine Kombination dieser Effekte ein (Claydon et al. 1982; Pell et
al. 2001).
Hawksworth listet nahezu 750 entomopathogene Pilzspezies aus 56 Gattungen
auf, die mit Arthropoden assoziiert sind (Hawksworth et al. 1995). Auf der Basis

Einleitung
16
dieses Pools sind mittlerweile über 20 Mykopestizide entwickelt worden,
überwiegend auf der Basis der Ascomyzeten Beauveria bassiana, Metarhizium
anisopliae, Lecanicillium muscarium ZIMMERMANN und Paecilomyces fumosoroseus
WIZE, die im Wesentlichen gegen Homopteren, Coleopteren, Lepidopteren, Dipteren
und Orthopteren zum Einsatz kommen (Shah et al. 1999; de Faria et al. 2007). Ein
kommerzielles Produkt, das aus einem Beauveria bassiana-Isolat entwickelt worden
ist, wird gegen die Milbe Tetranychus urticae KOCH in Gewächshauskulturen von
Rosen eingesetzt, Metarhizium anisopliae wurde erfolgreich gegen Ixodes ricinus
LINNAEUS sowie Boophilus annulatus SAY, Hyalomma excavatum KOCH und
Rhipicephalus sanguineus LATREILLE verwendet (Wright et al. 1996; Gindin et al.
2002; Strasser et al. 2007). Schon seit etwa 20 Jahren und damit sehr gut untersucht
sind ebenfalls die Wirt-Parasit-Interaktionen für Paecilomyces fumosoroseus,
Metarhizium anisopliae, Beauveria bassiana sowie insbesondere Lecanicillium
muscarium und die Weiße Fliege Trialeurodes vaporariorum WESTWOOD, die ein
bedeutender Schädling in Gewächshauskulturen ist; auf Basis dieser Erkenntnisse
sind gleichsam kommerzielle Produkte entwickelt worden, die seitdem erfolgreich
gegen Trialeurodes, aber auch andere wirtschaftlich bedeutende phytophage
Arthropoden appliziert werden (Wraight et al. 2000; de Faria et al. 2001, 2007;
Malsan et al. 2002). Die Pilze zeichnen sich durch leichte Kultivierbarkeit, ein
begrenztes Wirtsspektrum, eine hohe Pathogenität für verschiedene Stadien von
Trialeurodes vaporariorum und ihre Nichtpathogenität für Menschen aus (Ferron
1978; Mier et al. 1991; Ortiz-Caton et al. 1996).
In Assoziation zu Milben, die wie Tetranychus urticae Parasiten wirtschaftlich
bedeutender Kulturen oder Nutztiere sein können, sind 58 Pilzspezies bekannt, die in
der Lage sind, Wirte aller drei Ordnungen der Actinotrichida sowie innerhalb der
Anactinotrichida Vertreter der Ixodida und Gamasida zu infizieren: 24 Spezies sind
45 Wirten der Actinotrichida, 16 zehn Wirten der Ixodida und 18 insgesamt 14 Wirten
der Gamasida zugeordnet. Allein in Ixodes ricinus fand Kalsbeek sechs
Hyphomyceten; van der Geest beschreibt 30 Pilze für 16 Milbenfamilien (Kalsbeek et
al. 1995; van der Geest et al. 2000). Zwei dieser in Milben gefundenen Pilze –
Neozygites floridana WEISER AND MUMA und Hirsutella thompsonii FISHER – sind in
anderen Arthropodenklassen bisher nicht nachgewiesen worden (Chandler et al.
2000). Zwar ist Neozygites floridana als gamasidenspezifisch klassifiziert und könnte
daher ein potentieller Antagonist für Varroa destructor sein, ohne gleichzeitig Apis

Einleitung
17
mellifera negativ betreffende Seiteneffekte zu haben, allerdings konnte dieser Pilz
nicht in vitro kultiviert werden. Demgegenüber infiziert der zweite bisher nur Milben
zugeordnete Pilz Hirsutella thompsonii ausschließlich Vertreter der Actinedida und
wird erfolgreich gegen Phyllocoptruta oleivora ASHMEAD in Zitruskulturen und
Eriophyes guerreronis KEIFER auf Kokospalmen eingesetzt (McCoy 1981; Lampedro
et al. 1989; Allen et al. 1994).
Versuche zur Kontrolle von Vertretern der Ixodida konzentrierten sich daher auf
für diese Gruppe unspezifische Pilze, namentlich Beauveria bassiana, Metarhizium
anisopliae, Paecilomyces farinosus BROWN AND SMITH, Paecilomyces fumosoroseus
und Lecanicillium muscarium. Bittencourt konnte zeigen, dass Metarhizium, isoliert
aus anderen Arthropodenklassen, in der Lage war, Boophilus microplus CANESTRINI,
einen wirtschaftlich bedeutenden Parasiten auf Vieh, in vitro zu infizieren (Bittencourt
et al. 1994a, 1994b). Metarhizium anisopliae- und Beauveria bassiana-Isolate, die in
Ostafrika außerhalb der Acari gefunden wurden, befielen im Experiment
Rhipicephalus appendiculatus NEUMANN (Mwangi et al. 1995).
Anfang der letzten Dekade wurden erste Versuche unternommen, eine
potentiell antagonistische Wirkung unspezifischer entomopathogener Pilze auf
Varroa destructor und Apis mellifera zu testen. Dabei ist die Unterscheidung der
Pilze in (Labor-)Stämme für das Verständnis wichtig; die Pilze werden in der Folge
der Einfachheit halber aber nur unter ihrem Artnamen angesprochen – gemeint ist in
allen Fällen der der jeweiligen Untersuchung zugrundeliegende Stamm im Sinne
eines infraspezifischen Taxons respektive das Isolat einer Art.
Kanga gibt für zwei Isolate von Hirsutella thompsonii und Metarhizium
anisopliae, die in Laborversuchen an Milben auf Puppen und an Versuchsvölkern
(nur Hirsutella thompsonii) getestet wurden, bei denen eine konidienhaltige
Detergenslösung versprüht wurde, an, diese seien geeignet, Varroapopulationen
signifikant zu reduzieren, ohne Bienen zu schädigen; er bestimmte eine LT90 von vier
Tagen für Hirsutella thompsonii und von sechs Tagen für Metarhizium anisopliae
(Kanga et al. 2002). Für Bienen schloss er anhand vergleichbarer
Pilzwachstumsraten aus toten, nicht oberflächensterilisierten Bienen, sie würden
durch die Pilzstämme nicht negativ beeinflusst. Eine strenge Korrelation zwischen
der Zahl gefallener Milben und den aus diesen ohne Oberflächensterilisation
gewachsenen Pilzen sah er als Beleg für eine Infektion von Varroa destructor durch
Hirsutella thompsonii an. In einem weiteren Versuch an Völkern, die entweder mit

Einleitung
18
Konidienpulver oder dem pyrethroidhaltigen Tierarzneimittel Apistan behandelt
wurden, fand Kanga vergleichbare Mortalitätsraten und schlussfolgerte daraus auf
eine Eignung von Metarhizium als Kontrollagens (Kanga et al. 2003, 2005). In einem
Versuch, in dem 2006 Metarhizium-Konidien mit Hilfe von mit ihnen bedeckten
Kunststoffstreifen appliziert wurden, bestätigten sich die Ergebnisse (Kanga et al.
2006). Bei neun untersuchten Isolaten von Hirsutella thompsonii, die Peng
ausschließlich in vitro in Bezug auf ihre Wirkung auf Milben und Honigbienenpuppen
testete, indem sie Milben über eine reife Hirsutella-Kultur laufen ließ
beziehungsweise Puppen auf eine Hirsutella-Kultur legte, wurde für drei Isolate eine
Infektionsfähigkeit für Varroa destructor gefunden (Peng et al. 2002). Peng gibt für
diese drei Isolate eine LT50 von zwei, drei und vier Tagen an; eine negative Wirkung
auf Bienenpuppen wurde nicht festgestellt. Als Kontrollen wurden Gruppen
verwendet, die zu Medienplatten ohne Pilzkulturen Kontakt hatten. Shaw fand
demgegenüber für alle von ihr im Laborexperiment untersuchten Isolate eine
Infektionsfähigkeit für Varroa; alle wiesen darüber hinaus eine der Kontrollgruppe
vergleichbare oder höhere (bis zu 100 Prozent innerhalb des
Untersuchungszeitraums) Mortalität, bezogen auf adulte Bienen, auf (Shaw et al.
2002). In den Versuchen von Gerritsen an zuvor nicht vereinheitlichten Völkern mit
drei kommerziell erhältlichen Produkten auf der Basis von Metarhizium anisopliae-
und Lecanicillium muscarium-Stämmen zeigten sich keine signifikant erhöhten
Sterberaten für Varroa destructor und Apis mellifera (Gerritsen et al. 2006). 2006
untersuchte James den Metarhizium-Stamm, mit dem Kanga gearbeitet hatte, vor
dem Hintergrund der Fragestellung, welche Applikationsform die effektivste sei,
konnte dieses Mal aber keinerlei signifikante Effekte des Pilzes finden; zusätzlich
waren längstens nach 20 Tagen (Kanga 2003: 40 Tage) noch keimfähige Konidien
vorhanden (Kanga 2003; James et al. 2006). Meikle verglich drei Gruppen in vivo:
Eine wurde mit einem aus Varroa destructor isolierten Beauveria bassiana-Stamm
behandelt, eine weitere als Kontrolle mit einem als Matrix dienenden Pulver, während
die dritte Gruppe unbehandelt blieb (Meikle et al. 2007). Die Völker waren aus einer
zuvor nicht vereinheitlichten Grundgesamtheit gebildet worden und differierten in
Bezug auf die erfassten Parameter „Bienenmasse“ und „Fläche verdeckelter Brut“.
Völker der mit Konidien behandelten Gruppe wiesen einen signifikant erhöhten
Milbenfall am sechsten und achten Tag nach Applikation auf. Milben aus den mit
Konidien behandelten Völkern zeigten, ohne zuvor oberflächensterilisiert worden zu

Einleitung
19
sein, in allen Fällen ein signifikant höheres relatives Pilzwachstum. In einer
Repetition mit einer größeren Zahl nicht vereinheitlichter Völker und vier
verschiedenen Gruppen (Konidien und Carnaubawachsmatrix, Konidien und
Weizenmehlmatrix, Weizenmehl, keine Behandlung) trat ein verstärkter Milbenfall in
der Konidien-Carnaubawachsgruppe auf; dieser war allerdings nicht signifikant
erhöht (Meikle et al. 2008b). Der Milbenfall der Konidien-Weizenmehlgruppe lag
unterhalb des Falls der Weizenmehlgruppe und an drei Tagen auch unterhalb des
Falls der Kontrolle. Signifikant erhöhtes relatives Pilzwachstum aus einer
Konidiengruppe wertet Meikle als Beleg für die Eignung des Pilzes als
Varroaantagonist. Eine zweite Wiederholung erbrachte keinerlei Belege für eine
signifikante Reduktion der Parasitenlast der Versuchsvölker (Meikle et al. 2009).
Effekte entomopathogener Pilze auf Apis mellifera auf subletaler Ebene wurden
bisher in keiner Studie untersucht. Dabei könnte zum Beispiel eine Beeinträchtigung
des Lernvermögens und der Diskriminationsfähigkeit bei einem staatenbildenden
Insekt, das über komplexe Fähigkeiten zur räumlichen Orientierung und
olfaktorischen Differenzierung im Zusammenhang mit der Kommunikation innerhalb
des Staates verfügt, negative Konsequenzen haben. Gleiches gilt für eine
physiologische Belastung durch die pathogenbedingte Induktion einer Immunantwort
– auch dieser Aspekt ist bisher nicht untersucht worden. Dabei ist inzwischen auch
für die Honigbiene die Aktivierung einer Kaskade bestätigt, wie sie für andere
Insekten beschrieben ist (Evans et al. 2006; Müller et al. 2008; Randolt et al. 2008).
1.4 Zielsetzung
Entomopathogene Pilze erscheinen vor dem Hintergrund der Schwierigkeiten
der etablierten Behandlungsmethoden gegen Varroa destructor lohnend für eine
Erforschung als potentielle Antagonisten zu sein. Die bisherigen Untersuchungen
zeigen sehr variable respektive widersprüchliche Ergebnisse, die zum Teil
möglicherweise auf die Diversität dieser sehr pleomorphen Pilze zurückzuführen sind
– allerdings differierten die Resultate nicht nur zwischen den beteiligten Gruppen,
sondern auch bei Untersuchungen an identischen Isolaten innerhalb einer Gruppe.
Viele dieser Studien basieren auf unterschiedlichen methodischen Ansätzen, die

Einleitung
20
Interpretationen der Ergebnisse schwierig und ihre Vergleichbarkeit unmöglich
machen. Keine einzige Untersuchung beschäftigte sich mit der Frage einer
potentiellen Wirkung entomopathogener Pilze auf die Larvenstadien und das
Puppenstadium von Apis mellifera. Ebenso wenig standen mögliche subletale, aber
für die Bienen bedeutsame Effekte, wie eine Beeinträchtigung der Futteraufnahme,
des Lernvermögens oder des Immunsystems im Fokus von Untersuchungen. Und
schließlich lag keine Untersuchung vor, die sich mit dem Einfluss beschäftigt, den in
den Populationen vorhandene Pilze auf die Dynamik dieser Populationen und damit
auch auf die Varroapathologie von Apis mellifera haben könnten und die dabei
möglicherweise bisher in Varroa unentdeckte Pilze findet. Die vorliegende Arbeit
sollte anhand von vier Stämmen derjenigen Spezies entomopathogener Pilze, die
bisher hauptsächlich Gegenstand von Untersuchungen waren, die genannten Lücken
schließen, um eine valide, reproduzierbare Methodenbasis zur Untersuchung
entomopathogener Pilze in Bezug auf ihre Wirkung auf Varroa destructor und Apis
mellifera zu schaffen. Dies sollte eine bisher noch nicht existente Untersuchung einer
möglichen letalen Wirkung auf Larven und Puppen von Apis mellifera einschließen.
Darüber hinaus sollte untersucht werden, ob es eventuell subletale Effekte
entomopathogener Pilze auf Apis mellifera gibt, die in den bisherigen Versuchen
methodisch bedingt nicht hätten festgestellt werden können. Schließlich sollte
versucht werden, in vivo einen möglichen Zusammenhang zwischen der Abundanz
von Varroa destructor in Bienenvölkern und dem Vorkommen von Pilzen in Varroa
destructor zu untersuchen und im Rahmen eines Screenings varroaspezifische und
damit eventuell als Antagonisten geeignete Pilze zu erfassen.

Material und Methoden
21
2 Material und Methoden
2.1 Versuchstiere und verwendete Pilzstämme
Die Honigbienen (Apis mellifera) stammten aus Versuchsvölkern der AG
Verhaltensbiologie und Didaktik der Biologie der Ruhr-Universität Bochum, die im
Botanischen Garten der Ruhr-Universität Bochum sowie auf dem Versuchsgelände
der AG im Lottental, ebenfalls Bochum, untergebracht waren. Für die Unterbringung
wurden Holzkästen (im Folgenden „Magazine“ genannt) mit einer Grundfläche von
520 x 380 mm eingesetzt, die aus einzelnen, 220 mm hohen Modulen bestanden und
damit dem im Jahresverlauf schwankenden Raumbedarf eines Bienenvolkes
angepasst werden konnten. Die Magazine schlossen unten mit einem Rahmen (im
Folgenden „Gitterboden“ genannt) gleicher Grundfläche ab, der mit einem
Edelstahlgitter der Maschenweite 2 x 2 mm bespannt war, so dass von den im
Magazin befindlichen Bienen abfallende Organismen in einer unterhalb des Gitters
anbringbaren Polyethylenwanne gesammelt werden konnten, ohne durch Bienen
verschleppt werden zu können.
Die Völker wurden einheitlich nach anerkannten Praxisregeln geführt und
verfügten alle über eine Königin. Die Milben (Varroa destructor) stammten ebenfalls
aus Versuchsvölkern der AG. Völker, die Bestandteil des Screenings auf
entomopathogene Pilze waren, oder Donorvölker für Versuche an Milben wurden
entgegen der sonst üblichen Praxis im Spätsommer von der Applikation varroazider
Mittel ausgenommen, um die Ergebnisse nicht zu verfälschen, beziehungsweise eine
ausreichende Anzahl von Versuchsmilben zur Verfügung zu haben. Sämtliche
Versuche an Apis mellifera und Varroa destructor orientierten sich an den derzeit
gültigen Standards für Untersuchungen an Honigbienen (OEPP/EPPO 2001).
Die in den Untersuchungen zu Kulturbedingungen unspezifischer
entomopathogener Pilze, in den in vitro-Assays, im Versuch zur Duftdifferenzierung
und im Halbfreilandversuch untersuchten Pilze stammten vom
Landesgesundheitsamt Stuttgart (Metarhizium anisopliae (M.a. 153), Paecilomyces
fumosoroseus (P.fr. 9), Beauveria bassiana (B.b. 29)) beziehungsweise der Firma
Koppert BV, Niederlande (Lecanicillium muscarium („Mycotal“)).
Sämtliche Untersuchungen fanden in den Jahren 2005 - 2009 statt.

Material und Methoden
22
2.2 Medien und Puffer
Die Zusammensetzung von Medien und Puffern ist im Text erläutert;
komplexere Medien und Puffer sind nur namentlich erwähnt, ihre Zusammensetzung
ist Tabelle 2.1 zu entnehmen.
Medium/Puffer Bestandteil Bezugsquelle Malzextraktagar (MEA) 30 g/l Malzextrakt Roth
5 g/l Pepton Applichem 15 g/l Agar Applichem ddH2O add. 1 Liter
Sabouraud Dextrose Agar (SDA)
10 g/l Pepton Applichem 40 g/l Glucose-Monohydrat Baker 15 g/l Agar Applichem ddH2O add. 1 Liter
Yeast Pepton Dextrose Agar (YPDA)
40 g/l Glucose-Monohydrat Baker 10 g/l Pepton Applichem 10 g/l Hefeextrakt Applichem 15 g/l Agar Applichem ddH2O add. 1 Liter
Potato Dextrose Agar (PDA) 4 g/l Kartoffelextrakt Applichem, anwendungsfertige Mischung
20 g/l Glucose-Monohydrat 15 g/l Agar ddH2O add. 1 Liter
H2O-Agar 15 g/l Agar Applichem ddH2O add. 1 Liter
CM-Flüssigmedium 1,5 g/l KH2PO4 Fluka 0,5 g/l KCl Applichem 0,5 g/l MgSO4 Riedel-de Haën 3,7 g/l NH4Cl Roth 10 g/l Glucose-Monohydrat Baker 2 g/l Trypton Applichem 2 g/l Hefeextrakt Applichem 1ml/l 0,1% ZnSO4 Applichem 1 ml/l 0,1% FeCl2 Applichem 1ml/l 0,1% MnCl2 Applichem ddH2O add. 1 Liter
Lysepuffer (600 µl) 22 µl 1M Tris pH 8 Applichem 220 µl 250 mM EDTA Applichem 88 µl 5 M NaCl Roth 2,2 µl 20% SDS Roth ddH2O add. 600 µl
TE-Puffer 10 mM Tris pH 7,5 Applichem 1 mM EDTA Applichem
Tabelle 2.1: Komplexere Medien und Puffer, die im Text nur namentlich aufgeführt werden.

Material und Methoden
23
2.3 Bestimmung optimaler Kulturbedingungen
Untersucht wurden Malzextraktagar (MEA), Sabouraud Dextrose Agar (SDA),
Yeast Pepton Dextrose Agar (YPDA) und Potato Dextrose Agar (PDA).
Je Liter Kulturmedium wurden die angegebenen Mengen in ddH2O gelöst und
im Autoklaven bei 121°C sterilisiert. Anschließend wurden sterile 9 mm-Petrischalen
je etwa 3 mm hoch mit dem Medium gefüllt. Die Beimpfung erfolgte mit unmittelbar
zuvor geernteten Sporen aus Stammkulturen: Reife Sporen wurden mit 9 ml 0,85%
NaCl/0,1% Tween 80 (im Folgenden auch (Konidien-)Erntelösung genannt) pro
Petrischale durch Abkratzen mit einem sterilen Löffel suspendiert. Um ebenfalls
losgelöstes Myzel oder Kultursubstrat zu entfernen, wurde anschließend zweimal
durch sterile Filter (Sartorius 288) filtriert. Alle verwendeten Glasgeräte wurden zuvor
ebenfalls heißluftsterilisiert. Die Erfassung der Sporenkonzentration in der
Suspension erfolgte über Auszählung eines Aliquots in einer Neubauerzählkammer.
Anschließend wurde die Impfsuspension mit 0,85% NaCl/0,1% Tween 80 auf 107
Sporen/ml eingestellt. Zur Inokulation wurden je 5 µl mit einer sterilen Pipettenspitze
zentral auf die Platte gesetzt. Die Kultur erfolgte bei 25°C im Dauerdunkel für 14
Tage im Wärmeschrank (Memmert UNB 500). Die Erfassung der Parameter
„Koloniedurchmesser“ und „Konidiendichte“ wurde wie folgt durchgeführt: Zunächst
wurden die Petrischalen auf Millimeterpapier gesetzt, um den Koloniedurchmesser
zu erfassen. Anschließend wurde mit einem sterilen Spatel (Breite 2 cm) ein Quadrat
ausgestochen, das in eine neue Petrischale überführt wurde und dessen Konidien
wie oben beschrieben geerntet wurden. Die Auszählung erfolgte wiederum in einer
Neubauerzählkammer.
2.4 Bestimmung einer ausreichenden
Oberflächensterilisationsdauer gesammelter Milben
Unter den Gitterboden stark von Varroa destructor befallener Völker wurde zum
Auffangen der Milben eine Polyethylenwanne geschoben. Aus dem täglichen
Milbenfall der Versuchsvölker wurden die Milben manuell abgesammelt, gepoolt und
auf sieben Versuchsgruppen verteilt. Die Milben wurden in der Folge – mit

Material und Methoden
24
Ausnahme der Sterilisationszeit – gleich behandelt: Zunächst erfolgte durch
Eintauchen in 0,5% NaOCl die Oberflächensterilisation – je nach Gruppe für 10, 15,
20, 40, 60, 120 oder 240 Sekunden. Anschließend wurden die Milben in steriles
ddH2O überführt und für 30 Sekunden darin bewegt, um eventuell noch anhaftendes
NaOCl zu entfernen. Eine weitere Gruppe wurde keiner Oberflächensterilisation
unterzogen, sondern nur in sterilem ddH2O geschwenkt. Danach erfolgte mit sterilen
Holzstäbchen einzeln die Platzierung auf sterilen H2O-Agar-Platten und Kultur bei
25°C im Dauerdunkel. Einsetzendes Pilzwachstum wurde täglich erfasst, Milben, die
Pilzwachstum zeigten, wurden zur Vermeidung von Kreuzinfektionen unter sterilen
Bedingungen entfernt.
2.5 Ermittlung geeigneter Konservierungsmethoden gesammelter
Milben
Aus dem natürlichen Totenfall stark befallener Bienenvölker wurden die
gestorbenen Milben jeden Tag wie oben beschrieben gewonnen. Die Milben wurden
in der Folge gepoolt und in zwei Gruppen aufgeteilt: Eine Gruppe wurde bei
-26°C für sieben Tage eingefroren, die andere bei Raumtemperatur und
Raumfeuchte auf Filterpapier ebenfalls für sieben Tage getrocknet. Die Milben der
beiden Gruppen wurden im Fortgang gleich behandelt: Zunächst wurden sie durch
Eintauchen in 0,5% NaOCl für 120 Sekunden oberflächensterilisiert, eventuell noch
anhaftendes NaOCl dann durch Schwenken in sterilem ddH2O entfernt. Die Milben
wurden anschließend mit sterilen Holzstäbchen auf H2O-Agar-Platten überführt und
in der Folge bei 25°C und Dauerdunkel im Wärmeschrank kultiviert. Die
Überwachung eventuellen Pilzwachstums erfolgte täglich. Milben, die Pilzwachstum
zeigten, wurden inklusive des umgebenden Agars unter sterilen Bedingungen mit
einem sterilen Spatel ausgestochen, um Kreuzinfektionen zu vermeiden.

Material und Methoden
25
2.6 Erfassung der Mortalität mit Pilzkonidien behandelter
Bienenlarven
Die Methodik basiert auf einem von Peng entwickelten, aber modifizierten
Protokoll: Versuchsvölkern wurden Brutwaben mit Eiern kurz vor dem Schlupf der
Larven ohne aufsitzende Bienen entnommen, ins Labor gebracht und bis zur
Verwendung bei hoher Luftfeuchte gehalten (Peng et al. 1996). Im Labor wurden
sterile 24-well-Platten (Gibco) wie folgt vorbereitet: Zunächst wurde eine Larvendiät,
bestehend aus 20 ml Gelée royale und 10 ml einer Zuckerlösung (3,0 g Fruktose,
3,0 g Glukose (beide Baker) auf 33,3 ml steriles ddH2O), hergestellt, homogenisiert
und für mindestens 30 Minuten im Thermoblock auf eine Temperatur von 35°C
erwärmt. Dann wurden sechs Näpfe der 24-well-Platte mit Wasser gefüllt (jeweils
diagonal gegenüberliegend ein Endnapf jeder Reihe); anschließend erfolgte das
Einbringen von je 300 µl des Futters in je drei Näpfe pro Platte mit Hilfe einer
Omnifax-Tubukulin-Spritze. Den Waben wurden dann unter Kaltlicht frisch aus den
Eiern geschlüpfte Larven mit Hilfe einer Umlarvnadel entnommen und diese
zunächst auf vorgewärmte, mit einem angefeuchteten sterilen Papier (KimWipe)
ausgelegte Petrischalen gesetzt. In den Petrischalen erfolgte dann die Applikation
von entweder 1x106 Konidien/Larve, die wie unter 2.3 beschrieben gewonnen
wurden, oder eines entsprechenden Volumens Kontrolllösung durch Besprühen.
Anschließend wurden die Larven mittels der Umlarvnadel jeweils zu zehnt in einen
mit Futter ausgestatteten 24-well-Napf gesetzt (Abbildung 2.1). Die Hälterung
erfolgte in der Folge im Brutschrank bei 35°C und einer relativen Luftfeuchte von 90
Prozent. Alle 24 Stunden wurden die Larven mit Hilfe steriler, gebogener Laborspatel
passender Größe in neue, jeweils mit frisch zubereitetem und erwärmtem Futter
befüllte Näpfe umgesetzt und dabei schrittweise vereinzelt. Beim Umsetzen war
darauf zu achten, die Larven nicht um die Längsachse zu drehen, um ein allseitiges
Verkleben der Kutikula (und Verschluss der Stigmen beidseitig) zu vermeiden. Tote
und kranke Larven wurden entfernt und die jeweilige Zahl überlebender Larven
registriert; sofern eine Larve nicht eindeutig als krank oder tot klassifiziert werden
konnte, wurde sie in einem gesonderten Napf weiterkultiviert, bis Tod oder Überleben
feststanden – die Datierung im Fall des Todes erfolgte dann für den Tag der
Separation. Kurz vor der Verpuppung am sechsten oder siebten Tag des
Larvenstadiums (Mitteldarm öffnet sich, dunklere, kaudal gelegene Zone durch

Material und Methoden
26
Abgabe von Harnkristallen wird sichtbar, und die Larve streckt sich) wurden die
Larven in einen neuen, mit KimWipe-Papier ausgelegten Napf umgesetzt; zuvor
wurden die Larven vorsichtig auf KimWipe-Papier gerollt, um anhaftende
Futtersaftreste zu entfernen. Dieses Verfahren wurde in der Folge (nochmals 24h
später) wiederholt, bis kein Kot mehr abgegeben wurde. Danach wurden die Larven
unberührt im Brutschrank gehältert.
Abb. 2.1: Hälterung von Larven in vitro in 24-well-Platten. Je Reihe ist zur Erhöhung der
relativen Feuchte ein Gefäß mit ddH2O gefüllt. Larven vor der Verpuppung liegen auf
sterilem Papier.

Material und Methoden
27
2.7 Bestimmung der Mortalität mit Pilzkonidien behandelter
Adultbienen und Milben
Die Methodik zur Untersuchung der Mortalität mit Konidien unspezifischer
entomopathogener Pilze behandelter Adultbienen und aufsitzender Milben war für
alle untersuchten Pilze gleich und wird deshalb an dieser Stelle für beide Versuche
beschrieben.
Herstellung der Applikationssuspension entomopathogener Pilze:
Circa drei Wochen vor Versuchsbeginn wurden MEA-Platten aus den Pilz-
Stammkulturen frisch inokuliert und bei 25°C im Dauerdunkel gehalten, um zu
Versuchsbeginn eine möglichst hohe Anzahl Konidien ernten zu können. Die
Konidienernte erfolgte wie unter 2.3 beschrieben.
Bereitstellung adulter Bienen:
Adulte Bienen wurden Versuchsvölkern, die zuvor eine Behandlung mit 85%
Ameisensäure durchlaufen hatten und deshalb als „milbenfrei“
beziehungsweise „milbenarm“ klassifiziert werden konnten, durch Abschütteln
von Brutwaben durch einen Trichter in einen Eimer mit Gitterboden oder durch
Absaugen von Brutwaben mit Hilfe eines modifizierten Handstaubsaugers
entnommen. Anschließend erfolgte dreiminütiges Begasen der Tiere mit CO2
und nach erfolgreicher Betäubung eine Verteilung zunächst auf mit 30 radial
angebrachten Bohrungen versehene Polyethylenflaschen.
Bereitstellung von Milben:
Die zur Untersuchung benötigten Milben wurden unbehandelten
Versuchsvölkern, die in vorangegangenen Analysen des Milbenbefalls eine
hohe Milbenabundanz aufwiesen, wie folgt entnommen: Mehrere tausend den
Brutwaben aufsitzende Adultbienen wurden durch einen Trichter in einen mit
einem Gitter versehenen Eimer abgeschüttelt. Danach erfolgte ein Verschluss
der oberen Eimeröffnung durch ein Edelstahlsieb und durch dieses die
Applikation von jeweils 40 Gramm fein vermahlener Saccharose. Der Eimer
wurde anschließend manuell kreisend für etwa fünf Minuten bewegt und die von
den Bienen abfallenden Milben wurden in einer unter dem Eimer stehenden
Wanne aufgefangen. Diese Prozedur wurde mit jeweils fünf bis sechs
Donorvölkern wiederholt, um eine ausreichende Anzahl Milben zur Verfügung
zu haben. Die gesammelten Milben wurden anschließend zweimal mit ddH2O

Material und Methoden
28
gewaschen und der gelöste Zucker durch ein Feinsieb von den Milben getrennt.
Abschließend wurden die Milben auf feuchtem Laborpapier bis zu ihrer weiteren
Verwendung gehältert. Von Milben befreite Bienen wurden ohne weitere
Behandlung in ihre Herkunftsvölker zurückverbracht.
Besatz der Hälterungskäfige mit Bienen und Milben:
Die in Polyethylenflaschen bereitstehenden Bienen wurden erneut mit CO2
begast, den Flaschen entnommen und mit Milben besetzt: Je 50 Bienen
erhielten durch einzelnes Heranführen der Bienen an die Milben mit Hilfe einer
Pinzette einen Besatz von 25 Milben – das selbständige Aufsteigen auf die
Biene bei Annäherung derselben diente dabei als Vitalitätstest für die Milben.
Die Bienen mit den ihnen aufsitzenden Milben wurden dann in Edelstahl-
Versuchskäfige (50 x 75 x 40 mm) überführt. Die Käfige besaßen eine
durchlöcherte Bodenplatte und wurden an einer Seite mit einer Glasplatte zur
optischen Kontrolle verschlossen. Um Verschmutzungen durch Abgabe von Kot
zu reduzieren, wurden passende Papierstücke auf den Boden der Kästen
gelegt, die täglich gewechselt wurden. Im Weiteren wurden Bienen und Milben
in diesen in Brutschränken im Dauerdunkel bei 35°C und einer relativen
Luftfeuchte von 90 Prozent gehältert. Die Verabreichung von Futter (2M
Saccharoselösung) und Wasser erfolgte ad libitum. Die Hälterungskäfige
wurden einzeln auf Aluminiumprofile (50 x 30 x 30 mm) in Polyethylenwannen
(110 x 110 x 30 mm) gestellt, deren Rand innen 10 mm breit mit Vaseline
bestrichen wurde, um ein Entkommen abgefallener aber noch lebender Milben
zu verhindern (Abb. 2.2). Nach einem Tag wurden zunächst die durch das
Besatzverfahren eventuell letal geschädigten Tiere erfasst und entfernt.
Zur Applikation der Suspension wurden die Käfige einzeln entnommen, die
Glasscheibe durch ein Edelstahlgitter (Maschenweite 2 x 2 mm) ersetzt und die
je 50 Bienen und 25 Milben mit 0,544 ml der jeweiligen Konidiensuspension, die
mit Erntelösung (0,1% Tween 80, 0,5% NaCl) auf eine Konzentration 9 x 107
Konidien/ml eingestellt war, besprüht, so dass eine Konidiendosis von 1 x 106
Konidien/Biene erreicht wurde. Edelstahlgitter wurden nach Gruppen getrennt
verwendet und vor Wiederverwendung jeweils durch Ausglühen sterilisiert. Die
verwendeten 10-ml-Sprühflaschen waren zuvor durch Sprühtests, Auszählen
der Konidienkonzentration von Ausgangs- und versprühter Suspension in einer
Neubauerzählkammer und Ausplattieren einer Verdünnungsreihe versprühter

Material und Methoden
29
Suspension auf MEA-Platten auf Düsengängigkeit der Konidien und deren
Keimfähigkeit nach Düsendurchgang untersucht worden. Flaschen wurden
ebenfalls getrennt nach Gruppen verwendet und jeweils durch mehrmaliges
Spülen mit 70% Ethanol, gefolgt von Spülen mit ddH2O desinfiziert. Die
Kontrollgruppe wurde mit 0,544 ml 0,1% Tween 80, 0,5% NaCl pro 50 Bienen
besprüht, das als Emulgator für die Konidiensuspension diente.
Im Fortgang wurde der tägliche Bienen- und Milbentotenfall unter Rotlicht
erfasst, um die Bienen nicht zu stören. Tote Bienen wurden ebenso wie tote
Milben mit Pinzetten entfernt, die Bienen wurden nach sorgfältigem Absuchen
auf noch aufsitzende Milben verworfen, die Milben gesammelt und später auf
Pilzwachstum untersucht. Futter und Wasser wurden ergänzt und
gegebenenfalls verschmutzte Scheiben ausgewechselt.
Abb. 2.2: Aufbau des in vitro-Assays für Adultbienen. Unterbringung von je 50 Bienen
und 25 Milben in Hälterungskäfigen mit Zuckerlösung (Spritze) und Wasser (1,5 ml-
Reaktionsgefäß) ad libitum. Die Käfige stehen auf Aluminiumprofilen, die in Schalen,
deren Rand mit Vaseline bestrichen ist, untergebracht sind.

Material und Methoden
30
2.8 Erfassung des Pilzwachstums aus behandelten Milben
Zur Erfassung des anteiligen Pilzwachstums (i.e. der Anteil der gesammelten
Milben, aus denen Pilze wuchsen) aus dem Gesamtmilbenfall der in vitro-Assays an
Adultbienen und Milben (2.7) wurden die in diesen Versuchen gestorbenen Milben
täglich getrennt nach Pilz- und Versuchsgruppe gesammelt und zunächst durch 120-
sekündiges Eintauchen in 0,5% NaOCl gefolgt von Spülen in sterilem ddH2O für 30
Sekunden oberflächensterilisiert. Die Milben wurden dann mit sterilen Holzstäbchen
auf H2O-Agar-Platten gesetzt und bei 25°C im Dauerdunkel in Klimaschränken
kultiviert. Die Platten wurden täglich auf eventuelles Pilzwachstum kontrolliert;
Milben, die ein Pilzwachstum zeigten, wurden zu Vermeidung von Kreuzinfektionen
unter sterilen Bedingungen entfernt und verworfen.
2.9 Halbfreilandversuch an Apis mellifera-Völkern zur Wirkung
entomopathogener Pilze auf Adultbienen, Bienenlarven und
-puppen sowie Varroa destructor
Um in Bezug auf Verwandtschaftsgrad, Milbenbefall und Altersstruktur
vollkommen einheitliche Ausgangsbedingungen in jedem Versuchsvolk zu schaffen,
wurden insgesamt 21 Honigbienenvölkern die Königinnen entnommen und die
Bienen ohne Larvenstadien und Puppenstadium, aber mit aufsitzenden Milben in der
phoretischen Phase zunächst zu drei je etwa 20000 Bienen starken Großgruppen
vereinigt, die in zweiseitig vergitterten Zargen untergebracht waren und drei Tage bei
5°C im Dauerdunkel in einer Klimakammer gehältert wurden. Die zuvor
entnommenen Königinnen wurden in kleinen, geschlossenen Drahtkäfigen
zugehängt. Die Fütterung der Großgruppen erfolgte durch aufgelegten Futterteig
(Apifonda, Südzucker AG) und mit in umgedrehten Honiggläsern mit perforiertem
Deckel bereitgestellter 2M Saccharoselösung ad libitum.
Zu Beginn des eigentlichen Versuchs erfolgte unter Besprühen mit Wasser eine
Fusion der drei Großgruppen zu einer Supergruppe, die mechanisch durch
vorsichtiges Umrühren mit einer Schaufel und Schütteln in Bezug auf eine eventuell
vorhandene Nicht-Gleichverteilung der Einzelbienen vor dem Hintergrund der

Material und Methoden
31
Herkunft der Einzelbienen aus den Ausgangsgroßgruppen oder ihrer Alterstruktur
respektive ihres Befalls mit Milben, homogenisiert wurde. Die Verteilung der Bienen
erfolgte in gleich großen Quantitäten auf die insgesamt zehn Versuchsvölker, die in
Kästen gleicher Bauart auf leeren Waben untergebracht waren und je eine der zuvor
den Ausgangsvölkern entnommene Königin im Drahtkäfig unter Futterteigverschluss
erhielten, so dass die Königinnen innerhalb von 24 Stunden unter Verbrauch des
Futterteiges durch die Arbeiterinnen befreit wurden. Die Gabe von Wasser, Pollen
und Futter erfolgte ad libitum über eine Tränke pro Versuchsgruppe sowie den
Waben direkt aufgelegten Futterteig, der während des Versuchszeitraums laufend
ergänzt wurde, und zugehängte Futterwaben. Die Unterbringung der Kästen erfolgte
in zwei je 20 x 4 Meter großen Flugzelten am gleichen Standort und mit gleicher
Ausrichtung, so dass eventuelle Lageeffekte ausgeschlossen werden konnten
(Abbildung 2.3). Pilz- und Versuchsgruppe waren je in einem Zelt untergebracht, um
eine eventuelle Verschleppung von Lecanicillium-Konidien zu verhindern. Die
Fluglöcher der Völker in jedem Flugzelt wiesen in jeweils unterschiedliche
Himmelsrichtungen und waren farbig markiert, um ein Verfliegen von Bienen
zwischen den Völkern weitestgehend auszuschließen. Die Völker standen auf
Holzpaletten, um sie einerseits der Bodenfeuchte zu entziehen, und andererseits
durch Unterlegen von Holzkeilen exakt waagerecht ausrichten zu können. Zur
Erfassung des Milbenfalls wurde unter den Gitterboden jedes Volkes eine
passgenaue Polyethylenwanne geschoben, die etwa 5 Millimeter hoch mit Pflanzenöl
gefüllt wurde, um ein Verwehen gefallener Milben durch Wind, vor allem aber ein
Wegtragen durch fouragierende Ameisen zu verhindern. Nach dem Besatz wurde
sowohl den Pilz- als auch den Kontrollvölkern 24 Stunden Zeit gegeben, sich an die
Flugzelte und die zugesetzten Königinnen zu gewöhnen.
Die Vorbereitung der Konidiensuspension erfolgte zunächst nach
Herstellerspezifikation („Mycotal“, Koppert BV). Die additivhaltige Suspension wurde
dann aber gefiltert (Sartorius 288), um die Konidien von der Additivmatrix zu trennen
und die Konidiensuspension düsengängig zu machen. Anschließend wurde die
Konidienzahl pro Milliliter mit Hilfe einer Neubauerzählkammer bestimmt und die
Sporensuspension durch Verdünnen mit Wasser so eingestellt, dass mit vier
Pumpstößen pro Wabenseite aus einer 1,5-Liter-Sprühflasche eine single-bee-Dosis
von 1 x 106 Konidien appliziert werden konnte. Düsengängigkeit der Konidien und

Material und Methoden
32
deren Keimfähigkeit nach Düsendurchgang waren zuvor nach Sprühtests und
Ausplattieren einer Verdünnungsreihe auf MEA-Platten überprüft worden.
Die Pilzgruppe wurde einen Tag nach Besatz mit Lecanicillium muscarium-
Konidiensuspension besprüht, die Kontrollgruppe mit dem entsprechenden Volumen
Suspension, das analog aus konidienfreier Additivmatrix hergestellt wurde, die die
Firma Koppert zur Verfügung stellte.
In der Folge wurde der Milbenfall täglich durch Auszählen der ins Öl gefallenen
Milben erfasst, die Bienenzahl und die Zahl der Brutstadien wurden zu Beginn und
nach sieben, 14 und 17 Tagen durch Auflegen eines Schätzrahmens bestimmt. Nach
Ermittlung der Bienenzahl und Abschütteln der adulten Bienen in die Versuchsvölker
wurde mit Hilfe der gleichen Vorgehensweise die Zahl offener oder verdeckelter
Brutzellen erfasst. Die Zahl zum Zeitpunkt der Schätzung fouragierender
Arbeiterinnen wurde über die Erfassung innerhalb einer Minute an- beziehungsweise
abfliegender Bienen ermittelt. Der Kasteninnenwand ansitzende Bienen wurden
ebenfalls zur Gesamtzahl addiert.
Der relative Endbefall der Bienen wurde zum Versuchsende durch
Untersuchung der Arbeiterinnen ermittelt: Alle noch in den Völkern befindlichen
adulten Tiere wurden gesammelt, getötet und durch Waschen mit einer
detergenshaltigen Waschlösung von den aufsitzenden Milben befreit. Die abgelösten
Milben wurden anschließen in geeigneten Sieben aufgefangen und ebenso wie die
Bienen einzeln gezählt. Ebenfalls durch Auswaschen und Einzelauszählung wurde
der relative Milbenendbefall der verdeckelten Brut ermittelt: Zunächst wurden die
verdeckelten Brutzellen gezählt, nach Einfrieren mechanisch entdeckelt und die in
ihnen befindlichen Milben und Puppen mit einem scharfen Wasserstrahl ausgespült.
Nach Abschluss des Versuchs wurden alle Versuchstiere nach Autoklavieren
entsorgt.

Material und Methoden
33
2.10 Erfassung der subletalen Effekte entomopathogener Pilze auf
Adultbienen
Die Bienen für diesen Versuch stammten aus milbenarmen Versuchsvölkern
und wurden wie zuvor beschrieben gewonnen; ihnen standen 2M Saccharoselösung
und Wasser ad libitum zur Verfügung. Die Hälterung erfolgte analog zu den anderen
in vitro-Versuchen in Metallkäfigen bei 35°C und hoher Luftfeuchte im
Wärmeschrank. Zu Beginn des Versuchs wurden die Bienen mit einem den in
anderen Versuchen verwendeten Volumen äquivalenten Volumen
Konidienerntelösung besprüht. In der Folge wurden die Tiere keiner weiteren
Behandlung unterzogen, lediglich gestorbene Bienen wurden entfernt. Der Gruppe
Abb. 2.3: Aufstellung der Versuchsvölker in einem von zwei 20 x 4 Meter
großen Zelten auf waagerecht ausgerichteten Paletten mit versetzter
Ausrichtung der Fluglöcher.

Material und Methoden
34
wurden täglich Bienen entnommen, die zunächst auf Eis betäubt und denen dann
nach Fixierung des Thorax mit einer Pinzette durch Abziehen das Abdomen entfernt
wurde. Bei vorsichtiger Vorgehensweise wird dabei die am Oesophagus hängende
Honigblase nach kranial herausgezogen. Anschließend wurde die Honigblase mit
einer feinen Schere abgetrennt und auf einer Analysewaage (Sartorius) gewogen.
Der mögliche Einfluss entomopathogener Pilze auf das Lernverhalten wurde im
Rahmen einer Rüsselreflexdressur (auch PER-Dressur, „Poboscis Extension Reflex-
Dressur“) untersucht (Bittermann et al. 1983).
Für jede Versuchsreihe wurden den Versuchsvölkern 1000 Bienen durch
Abschütteln entnommen und nach CO2-Betäubung in Metallkästen (50 x 75 x 40
mm³) mit jeweils 50 Bienen überführt. Die Kästen verfügten über eine durchlöcherte
Bodenplatte und wurden an einer Seite mit einer Glasplatte verschlossen. Die
Hälterung der Bienen erfolgte bei 35°C und einer relativen Luftfeuchte von 90
Prozent im Dauerdunkel des Brutschrankes. Bis zum Dressurtag standen 2M
Saccharoselösung und Wasser ad libitum zur Verfügung. In der Pilzgruppe erfolgte
nach Besatz die Applikation von 0,544 ml Konidiensuspension des Pilzes
Lecanicillium muscarium einer Konzentration von 9 x 107 Konidien/ml, so dass eine
single-bee-Dosis von 1 x 106 Konidien/Biene erreicht wurde. Analog wurde die
Kontrollgruppe je Kasten mit 0,544 ml 0,1% Tween 80, 0,5% NaCl besprüht, das als
Emulgator für die Konidiensuspension diente.
Vor der Rüsselreflexdressur wurde den Bienen in Abhängigkeit vom erwarteten
Füllungsvolumen der Honigblase und dem ermittelten durchschnittlichen
Futterverbrauch Futter entzogen, so dass sie zu Beginn der Rüsselreflexdressur
hungrig waren. Die am Versuch teilnehmenden Bienen wurden aus den Kästen
entfernt und nach Betäubung durch Kühlung auf Eis einzeln in Reaktionsgefäße (1,5
ml, Eppendorf), deren Spitzen abgeschnitten waren, eingeführt. Die Tiere wurden am
Thorax so mit Klebeband fixiert, dass Vorderbeine, Rüssel und Antennen frei
beweglich blieben. Um eine Flucht aus den Reaktionsgefäßen zu verhindern, wurden
diese hinter dem Abdomen der Bienen mit Papierstopfen blockiert.
Etwa eine Stunde nach Einbringen der Bienen in die Reaktionsgefäße
(Abbildung 2.4) begann die PER-Testphase: Die Antennen der Bienen wurden alle
30 Minuten mit einem in 2M Saccharoselösung getauchten Schmelzpunkt-
Bestimmungsröhrchen (80 mm x 1 mm) berührt. Bienen, die auf die Gabe dieses
unbedingten Reizes nicht mit Herausstrecken des Rüssels reagierten, wurden für die

Material und Methoden
35
Rüsselreflexdressur verworfen. In der Testphase erfolgreiche Bienen wurden
anschließend in den PER-Versuchsaufbau (Abbildung 2.5) eingesetzt.
Für die Rüsselreflexdressur wurden Geraniol als Dressurduft und Citral als
Vergleichsduft (beide SIGMA-ALDRICH) verwendet. Dazu wurden jeweils 10 µl des
Duftstoffes auf ein Stück Filterpapier (Grade 288, 90 mm Durchmesser, 80 g/m²)
aufgetragen und das Filterpapier in fabrikneue Glaspasteurpipetten (Länge 150 mm)
eingebracht. Als Kontrolle diente unbehandeltes Filterpapier in einer
Glaspasteurpipette.
Die Pipetten führten den Bienen aus einer Druckluftflasche einen konstanten
Luftstrom von 50 ml/min zu, der durch ein Peltierelement auf 35°C aufgeheizt wurde.
Die Gabe des Duftreizes war über ein Magnetventil steuerbar, während ein zweiter
duftloser Luftstrom gleicher Durchflussmenge appliziert wurde, der eine Assoziation
der bloßen Luftbewegung mit dem unbedingten Reiz verhindern sollte. Bienen, die
sich auf dem drehbaren Karussell unmittelbar vor der Versuchsposition befanden,
wurden durch ein Holzbrett vom gerade applizierten Duft abgeschirmt. Zusätzlich war
über der Versuchsposition ein Ventilator angebracht, der für eine schnelle Abfuhr des
applizierten Duftes sorgte, um die anderen auf dem Karussell befindlichen Bienen
nicht zu beeinträchtigen.
Die Rüsselreflexdressur beinhaltete zunächst drei Lernakte, an die sich
abwechselnd acht Test- und Vergleichsakte anschlossen. Nach dem zweiten und
fünften Vergleichsakt wurde eine Kontrolle durchgeführt. Nach Positionierung der
jeweiligen Versuchsbiene unmittelbar vor den Pipettenspitzen und einer
Gewöhnungszeit von etwa fünf Sekunden wurde der Luftstrom für sechs Sekunden
mittels Magnetventilsteuerung von der Kontrollpipette auf die Pipette mit Dressurduft
umgeleitet. Nach etwa zwei Sekunden wurden die Antennen der Biene mit 2M
Saccharoselösung berührt und die Biene wurde nach Herausstrecken des Rüssels
damit belohnt. Bienen, die auf den neutralen Luftstrom mit einem Rüsselreflex oder
nicht auf den unbedingten Reiz reagierten, wurden aus dem Versuch entfernt. Auf
der horizontalen, drehbaren Scheibe des „Karussells“ befanden sich
Aufnahmebohrungen für 20 Reaktionsgefäße; jede Einzelbiene konnte etwa alle acht
bis zehn Minuten getestet werden.
Während der acht Testakte erfolgte keine Konditionierung mehr – die Tiere
wurden also nur bei einer positiven Antwort auf den bedingten Reflex durch Gabe

Material und Methoden
36
von Saccharoselösung belohnt. Antworten auf den Vergleichsduft oder Antworten
während der zwei Kontrollakte wurden in keinem Fall belohnt.
Die Klassifikation der Antworten erfolgte in den Kategorien „unbedingter
Reflex“, „bedingter Reflex“, „Reaktion auf Luft“ oder „keine Reaktion“.
Zur Untersuchung der Fragestellung, ob es einen Effekt der Applikation eines
entomopathogenen Pilzes auf die Futteraufnahme der Einzelbiene gibt, wurde der
Futterverbrauch je Einzelkasten alle 24 Stunden durch Wiegen der das Futter
enthaltenden Spritze und Differenzbildung zum 24 Stunden zuvor gemessenen Wert
erfasst; parallel wurde die Zahl der je Kasten noch verbliebenen Bienen ermittelt.
Abb. 2.4: Honigbienenarbeiterin in der Rüsselreflexdressur; das Tier wird während der Konditionierung
mit Saccharoselösung belohnt; links oben die Spitzen der Glaspasteurpipetten, durch die Dressur- und
Vergleichsduft appliziert werden.

Material und Methoden
37
2.11 Erfassung des Einflusses entomopathogener Pilze auf das
Immunsystem von Adultbienen
Versuchsvölkern wurden adulte Bienen durch Abschütteln entnommen und mit
CO2 betäubt. Anschließend erfolgte die Verteilung auf sieben Versuchsgruppen mit je
50 Bienen. Der Versuchsgruppe „amputiert“ wurde jeweils ein Bein amputiert, um
eine obligate Immunantwort auszulösen und eine Positivkontrolle zu erhalten. Die
anderen Gruppen wurden mit 0,544 ml einer Konidiensuspension besprüht, so dass
eine single-bee-Dosis von 1 x 106 Konidien/Biene erreicht wurde. Appliziert wurden
Konidien der entomopathogenen Pilze Lecanicillium muscarium, Metarhizium
anisopliae, Beauveria bassiana, Paecilomyces fumosoroseus, sowie Sporen von
Saccharomyces cerevisiae MEYEN. Die Negativkontrollgruppe erhielt eine äquivalente
Dosis konidienfreier Erntelösung. Analog zu allen anderen in vitro-Assays wurden die
Abb. 2.5: Versuchsapparatur zur Durchführung der Rüsselreflexdressur:
Glaspasteurpipetten mit Peltierelement, Ventilator, „Karussell“, Signalgeber.

Material und Methoden
38
Honigbienen anschließend in Metallkäfigen bei 35°C und hoher Luftfeuchtigkeit im
Dauerdunkel gehältert; Futter und Wasser standen ad libitum zur Verfügung. 24
Stunden später wurde den Bienen nach Betäubung auf Eis Hämolymphe
entnommen: Auf Thorax und Abdomen der Bienen wurde nach Abtrennen der Tibia
eines Mittelbeines leichter Druck ausgeübt, bis ein Hämolymphtropfen austrat, der
mit einer Eppendorfpipette aufgenommen wurde. Je 5 µl Hämolymphe wurden in ein
200 µl-Eppendorfgefäß überführt, das 1 µl einer Lösung Phenylthioharnstoff 0,1
mg/ml und des Proteaseinhibitors Aprotinin 0,1 mg/ml (beide SIGMA ALDRICH)
enthielt, um ein Melanisieren der Hämolymphe zu vermeiden. Die Lagerung erfolgte
bei -26°C im Gefrierschrank.
Als Teststämme für den Immunversuch wurden Escherichia coli (B) CASTELLANI
AND CHALMERS (DSM 613, DSMZ, Braunschweig) und Bacillus subtilis (168) COHN
(Mikrobielle Antibiotikaforschung, Ruhr-Universität Bochum) verwendet. Aus den
Stammkulturen wurden je 40 ml einer sterilen LB-Flüssigkultur (5 g/L Hefeextrakt, 10
g/L Trypton, 10 g/L NaCl, alle Applichem) angeimpft und über Nacht bei 37°C und
120 rpm inkubiert. Am nächsten Morgen wurden aus dieser Kultur erneut 5 ml LB
flüssig angeimpft; die Inkubation erfolgte bei 37°C und 120 rpm sowie stündlicher
Probennahme und Messung im Photometer (Shimadzu UVmini 1240) bis zu einer
OD650 von 0,5. Dem Ansatz wurde daraufhin ein Aliquot entnommen und mit LB
flüssig 1:10 verdünnt. 1 µl dieser Verdünnung wurden dann mit je 1,5 µl der
gewonnenen Hämolymphe vermischt; zwei weitere Ansätze enthielten keine
Hämolymphe, sondern nur 1 µl einer Lösung Phenylthioharnstoff 0,1 mg/ml und des
Proteaseinhibitors Aprotinin 0,1 mg/ml beziehungsweise sterilfiltriertes Streptomycin
100 µg/ml. Die Ansätze wurden kurz anzentrifugiert und anschließend für zwei
Stunden bei 37°C inkubiert. Jeder Ansatz wurde dann auf 10 µl aufgefüllt und es
wurde eine Verdünnungsreihe über 6 Potenzen erstellt (je 2 µl auf 18 µl LB-Medium).
2 µl je Ansatz wurden anschließen auf sterilen LB-Platten (5 g/L Hefeextrakt, 10 g/L
Trypton, 10 g/L NaCl, 15 g/L Agar, alle Applichem) ausplattiert und im Brutschrank
bei 37°C inkubiert. 24 Stunden später erfolgte die Auszählung und Ermittlung der
Zahl koloniebildender Einheiten (colony forming units, cfu) in Abhängigkeit von der
Verdünnungsstufe.

Material und Methoden
39
2.12 Screening ausgewählter Völker auf potentiell spezifische
varroapathogene Pilze
Die Erfassung der Bienenzahl erfolgte wie unter 2.9 beschrieben während des
jeweiligen Sommerhalbjahrs an insgesamt 215 Versuchsvölkern in vierwöchigen
Abständen fünfmal je Volk. Zur Ermittlung des täglichen Milbenfalls wurde unter das
Gitter des Bodens eines Volkes für je drei Tage eine passgenaue Polyethylenwanne
geschoben, die ein Verwehen gefallener Milben durch Wind verhinderte. Der
Milbenfall wurde für jedes Volk während des Untersuchungszeitraums an 21
Terminen durch genaues Auszählen ermittelt.
Zur Untersuchung einer eventuellen Korrelation von Milbenbefall und dem
Vorkommen von Pilzen wurden auf der Basis der Populationsschätzungen Völker
ausgewählt, deren Milbentotenfall in Bezug auf die Erzielbarkeit eines Pilzwachstums
in vitro untersucht wurde. Den zu untersuchenden Völkern wurde dazu eine
passgenaue Auffangwanne untergeschoben und der tägliche Milbenfall über einen
Zeitraum von bis zu zwei Wochen pro Volk gesammelt. Im Labor erfolgte dann die
Oberflächensterilisation durch Eintauchen der Milben in 0,5% NaOCl für 120
Sekunden und anschließendes Spülen in sterilem ddH2O. Danach wurden die Milben
– getrennt nach Herkunftsvölkern – mit sterilen Holzstäbchen auf H2O-Agar-Platten
(15 g/L Agar, Applichem) überführt und bei 25°C im Dauerdunkel in Brutschränken
kultiviert. Das aus diesen Milben erfolgte Pilzwachstum wurde registriert; Milben, die
ein Pilzwachstum zeigten, wurden entfernt und getrennt unter gleichen Bedingungen
weiterkultiviert, um eine Kreuzinfektion der auf den Agarplatten verbliebenen Milben
zu vermeiden. Der Einschub der Schubladen über bis zu zwei Wochen bedingte,
dass es zur Verschleppung von Milben durch fouragierende Ameisen kam. Aus
diesem Grund konnte nicht gleichzeitig der Befall des Volkes über den
Milbentotenfall in der Auffangwanne erfasst werden. Allen Völkern wurden deshalb
während der Phase des Milbensammelns Proben von etwa 1000 adulten Bienen
entnommen, die ausgezählt und deren aufsitzende Milben nach Abwaschen mit einer
Detergenslösung und Absieben ebenfalls erfasst wurden.

Material und Methoden
40
2.13 Identifizierung von Pilzen im Pilzscreening
Lactophenolblaufärbung
Um die isolierten Pilze nicht nur molekularbiologisch charakterisieren zu
können, wurden mikroskopische Präparate angelegt: Nach Beladen eines
Objektträgers mit 10 µl Lactophenolblau (Sigma Alrich) wurde ein transparenter,
einseitig klebender Kunststofffilmstreifen auf die Wuchszone im Randbereich des
Pilzes gedrückt, um Konidien und Myzel zu übertragen; der Streifen wurde
anschließend auf den Lactophenolblautropfen gelegt und nach Trocknen
mikroskopiert.
Flüssigkultur und DNA-Extraktion
Um Aussagen zur Spezieszugehörigkeit der im Screening aus den Milben
gewachsenen Pilze treffen zu können, wurden diese auf der Basis von
Sequenzdaten identifiziert: Zunächst erfolgte durch wiederholtes Überimpfen auf
MEA-Platten eine Isolation der gewachsenen Pilze, bis eine Kontamination durch
Fremdkulturen ausgeschlossen werden konnte. Die Kultur der isolierten Pilze erfolgte
bei 25°C im Dauerdunkel in Klimaschränken. Zur Gewinnung der DNA wurde je Pilz
eine Petrischale mit autoklaviertem CM-Flüssigmedium mit drei unter sterilen
Bedingungen ausgestochenen Myzelstücken beimpft und drei Tage bei 25°C im
Dauerdunkel kultiviert. Nach Einsetzen eines deutlichen Pilzwachstums in der
Flüssigkultur wurden die Agarstücke mit Hilfe steriler Pipettenspitzen entfernt, das
Myzel durch Zerreißen mit sterilen Pipettenspitzen grob zerkleinert und 2,0-ml-
Reaktionsgefäße (Eppendorf) zu einem Drittel mit Myzel gefüllt. Es folgte
Zentrifugation bei 16000 g für zwei Minuten und Abpipettieren des Überstandes. Je
Eppendorfgefäß wurden dann 600 µl Lysepuffer zugefügt. Anschließend folgten
sechs Homogenisier-Gefrier-Auftauzyklen: 1 Minute Homogenisierung im
Reagenzglasschüttler (Vortex), 30 Sekunden Gefrieren in flüssigem Stickstoff, 1
Minute Auftauen im Wasserbad (GFL) bei 70°C. Danach wurde der Gefäßinhalt bei
70°C komplett aufgetaut. Anschließend wurde unter dem Abzug 1 ml Phenol (Roth)
zugegeben, homogenisiert, und 15 Minuten bei 13500 g, 4°C zentrifugiert.
Der Überstand wurde in ein neues Gefäß überführt und 1 ml Phenol-Chloroform-

Material und Methoden
41
Isoamylalkohol zugegeben. Danach wurde wiederum homogenisiert und bei 13500 g
und 4°C für 10 Minuten zentrifugiert. Nach erneuter Überführung des wässrigen
Überstandes erfolgte die Zugabe von 0,5 Vol. 7,5M Ammoniumacetat und 1 Vol
Isopropanol. Nach vorsichtigem Invertieren und Aufbewahren bei -26°C über Nacht
wurde am nächsten Morgen bei 16000 g für 40 Minuten zentrifugiert, der Überstand
vorsichtig abgegossen und das Pellet mit 300 µl 70% Ethanol gewaschen. Nach
folgender Zentrifugation bei 16000 g für 10 Minuten wurde der Überstand verworfen
und das Pellet bei Raumtemperatur getrocknet. Abschließend erfolgte die Zugabe
von je 30 µl autoklaviertem ddH2O.
DNA-Aufreinigung
Die Proben wurden zunächst unter Vermeidung des Mitführens des nicht
resuspendierten Anteils in ein neues Eppendorfgefäß überführt, mit 4 µl RNase A (10
mg/ml, Fermentas) vermischt und 2 Stunden bei Raumtemperatur inkubiert.
Anschließend wurde die Lösung mit 1 Vol Phenol-Chloroform-Isoamylalkohol (Roth)
versetzt, ausgeschüttelt und bei 12300 g für 10 Minuten abzentrifugiert. Der wässrige
Überstand wurde unter Vermeidung des Mitführens der Interphase in ein neues
Eppendorfgefäß pipettiert, bevor der Vorgang wiederholt wurde. Anschließend
erfolgte ein Ausschütteln mit 1 Vol Chloroform, erneutes Ausschütteln und
Abzentrifugieren bei 12300 g für 10 Minuten und danach ein Versatz mit 0,6 Vol
Isopropanol. Nach Inkubation bei -26°C wurde bei 16000 g in 45 min quantitativ
abzentrifugiert, mit 70% Ethanol gewaschen, bei 45°C im Thermoblock getrocknet
und das Pellet danach erneut in 30 µl autoklaviertem ddH2O aufgenommen. Die
Überprüfung der Extraktion und Aufreinigung erfolgte über eine Roh-DNA-
Agarosegelelektrophorese auf einem 0,7% Agarosegel (NEEO Ultra, Roth) und
anschließende Färbung und Dokumentation unter einer UV-Bank (Biometra
BioDocAnalyze). Die aufgereinigten DNA-Proben wurden bis zur weiteren
Verwendung bei -26°C im Gefrierschrank gelagert.

Material und Methoden
42
Konzentrationsbestimmung
Die Bestimmung der Konzentration erfolgreicher Extraktionen erfolgte
photometrisch (Shimadzu UVmini 1240 Spectrophotometer): 5 µl DNA-Extrakt
wurden mit 95 µl TE-Puffer zu einer 1:10-Lösung verdünnt und in eine
Photometerküvette gegeben; der Nullabgleich erfolgte gegen TE-Puffer. Die OD-
Messung wurde dann mit jeweiligem Nullabgleich bei 260 und 280 nm durchgeführt.
PCR
Verwendet wurden je Ansatz 1µl 10x PCR-Puffer (Sigma Aldrich), 1µl dNTP-Mix
2mM (MWG) je 0,5 µl forward-/reverse-Primer, 0,2 µl RedTaq-Polymerase 5U/µl
(Sigma Aldrich) und ddH2O add. 10 µl. Grundsätzlich lief ein Ansatz als
Negativkontrolle ohne DNA mit.
Die verwendeten Primer (ITS1F (Bruns et al. 1993) und ITS4 (White et al.
1990), beide MWG) hatten folgende Sequenzen:
ITS1F 5´-CTTGGTCATTTAGAGGAAGTAA-3´ ITS4 5´-TCCTCCGCTTATTGATATGC-3´
Als Protokoll für die PCR (Thermocycler: Biometra UNO Thermoblock) wurden
nach Voruntersuchungen die in Tabelle 2.2 zusammengefassten Parameter
verwendet.
Schritt Temperatur (°C) Dauer (min) Initiale Denaturierung 94 3 Denaturierung 94 0,5 Annealing 55 0,5 Elongation 68 1 Finale Elongation 68 3 Ende 4 ∞
Die Kontrolle der PCR auf Erfolg und eventuelle Kontaminationen erfolgte auf
einem 1,7% Agarosegel.
30
Tabelle 2.2: Parameter der PCR der ITS-Region

Material und Methoden
43
PCR-Cleanup
Die Amplifikate wurden enzymatisch aufgereinigt. Verwendet wurden
Exonuclease I aus Escherichia coli und Shrimp Alkaline Phosphatase (SAP) (beide
Fermentas). 5 µl des PCR-Produkts wurden mit je 0,25 µl Exonuclease I und 0,5 µl
SAP versetzt und für 30 Minuten bei 37°C inkubiert. Anschließend erfolgte die
Inaktivierung bei 85°C für 15 Minuten (Thermomixer comfort, Eppendorf).
Sequenzierung
Die Sequenzierung erfolgte am Lehrstuhl für Biochemie I – Rezeptorbiochemie
der Ruhr-Universität Bochum auf einem Applied Biosystems 3130 xl Genetic
Analyzer nach den Spezifikationen des Lehrstuhls. Primer für die Sequenzierung war
ITS1F wie oben beschrieben.
Datenbankabfrage, Alignment und Stammbaumkonstruktion
Die Identifizierung der Pilze erfolgte durch Abgleich mit bereits abgelegten
Sequenzdaten in der EMBL Nucleotide Sequence Database
(http://www.ebi.ac.uk/Tools/fasta33/nucleotide.html, Stand Juni 2009). Als Parameter
wurden die in Tabelle 2.3 dargestellten Einstellungen verwendet.
Program FASTADatabases EMBL FungiMatrix noneGap open -14Gap extend -4KTUP 6Expectation upper value 10,0Expectation lower value defaultDNA strand bothSequence Range Start-EndDatabase Range Start-EndFilter noneStatistical estimates Regress
Tabelle 2.3: Übersicht der für die EMBL-Abfrage verwendeten Einstellungen

Material und Methoden
44
Das Alignment der Sequenzen erfolgte unter Verwendung des Programms
MEGA (Molecular Evolutionary Genetics Analysis) Version 4 auf einem PC unter
Windows XP (Tamura et al. 2007).
Die Alignierung wurde mit ClustalW durchgeführt. Anschließend wurden alle
Sites am Sequenzanfang und -ende, die nicht in allen Sequenzen ein Signal geliefert
hatten, manuell entfernt. Die Berechnung eines ungewurzelten Baumes auf der Basis
der Alignierung mit dem Neighbor-Joining-Verfahren wurde ebenfalls mit MEGA 4.0
durchgeführt (Saitou et al. 1987; Wägele 2001). Die Distanzwerte wurden nach dem
Maximum Likelihood-Verfahren ermittelt (Tamura et al. 2004).
Der Baum wurde außengruppenlos ohne orthologen Datensatz berechnet.
Als Parameter für die Baumberechnung wurden die in Tabelle 2.4 dargestellten
Einstellungen verwendet.
2.14 Statistik und Parameter der grafischen Darstellung
Die verwendeten statistischen Tests, der Stichprobenumfang und die
ermittelten Signifikanzniveaus sind zu den jeweiligen Ergebnissen angegeben.
Einzelheiten zur Durchführung der Tests können Backhaus, Sachs und Siegel
entnommen werden (Siegel et al. 1988; Backhaus 2008; Sachs et al. 2009).
Die Darstellung der Ergebnisse erfolgt in Balken-, Linien-, Streudiagrammen,
einer Streudiagrammmatrix oder Box- und Whiskerplots. Im Fall von Box- und
Whiskerplots ist als unterteilende Linie der Box der Median dargestellt. Im unteren
Bereich der Box wird das 25. Perzentil, im oberen Bereich der Box wird das 75.
Perzentil dargestellt. Die Länge der Whisker beträgt das 1,5-fache der Höhe der Box;
Data Type NucleotideAnalysis Phylogeny reconstructionMethod Neighbor-JoiningGaps/Missing Data Complete DeletionModel Nucleotide: Maximum Composite LikelihoodSubstitutions to Include d: Transitions + TransversionsPattern among Lineages Same (Homogeneous)Rates among sites Uniform rates
Tabelle 2.4: Übersicht der für die Baumberechnung verwendeten Einstellungen in MEGA 4.0

Material und Methoden
45
falls keine Fälle mit Werten in diesem Bereich vorhanden sind, wird die Länge durch
den maximalen beziehungsweise minimalen Wert festgelegt. Ausreißer sind als
Punkte dargestellt und umfassen Werte, die nicht innerhalb der Whisker liegen.
Extremwerte sind durch Sterne dargestellt und umfassen all jene Fälle, deren Werte
mehr als dreimal so groß sind wie die Höhe der Boxen. Im Fall von
Regressionsgeraden sind die Werte der quadratischen Regression jeweils
angegeben.

Ergebnisse
46
3 Ergebnisse
Um Aussagen darüber gewinnen zu können, ob als Kontrollagentien geeignet
erscheinende, entomopathogene Pilze eine negative Wirkung auf die Larvenstadien
beziehungsweise das Puppenstadium von Honigbienen respektive adulte
Honigbienen sowie adulte Milben haben und ob eine signifikant erhöhte
Pilzwachstumsrate bei Milben post mortem detektierbar war, wurden in den
Untersuchungen mit einer definierten Einzeldosis Konidien des jeweils untersuchten
Pilzes behandelte Individuen mit nur mit einer entsprechenden Menge der
Erntelösung behandelten Individuen in der Kontrollgruppe verglichen. Die dabei
verwendeten methodischen Ansätze unterschieden sich bei Honigbienen in
Abhängigkeit vom untersuchen Stadium des Tieres und differierten darüber hinaus
zwischen den Labor- und den Halbfreilanduntersuchungen. Im Folgenden werden
deshalb vor der Ergebnisbeschreibung zunächst noch einmal kurz die verwendeten
methodischen Ansätze vor dem Hintergrund der Zielsetzung des jeweiligen Versuchs
erläutert.
3.1 Voruntersuchungen
3.1.1 Kulturbedingungen
Um ein für die untersuchten entomopathogenen Pilze, vor allem aber für die im
Screening zu isolierenden Pilze mit unbekannten Kulturansprüchen geeignetes
Medium zu finden, das eine möglichst geringe Zahl von Arten durch Nichteignung
ausschließt, wurden die zur Verfügung stehenden entomopathogenen Pilze auf vier
unterschiedlichen Medien kultiviert und untersucht, inwiefern sich die Zahl erntbarer
Konidien pro Flächeneinheit sowie die Koloniedurchmesser unterschieden.
Die PDA- und YPDA-Nährböden wiesen bei jeweils einem untersuchen Pilz
(Beauveria bassiana beziehungsweise Paecilomyces fumosoroseus) deutlich
geringere erzielbare Konidiendichten und Koloniedurchmesser im Vergleich zur
Restgruppe auf, so dass diese beiden Medien mit Blick auf die Erfassung
undetektierte Pilze in phylogenetischer Nähe zu Beauveria bassiana und
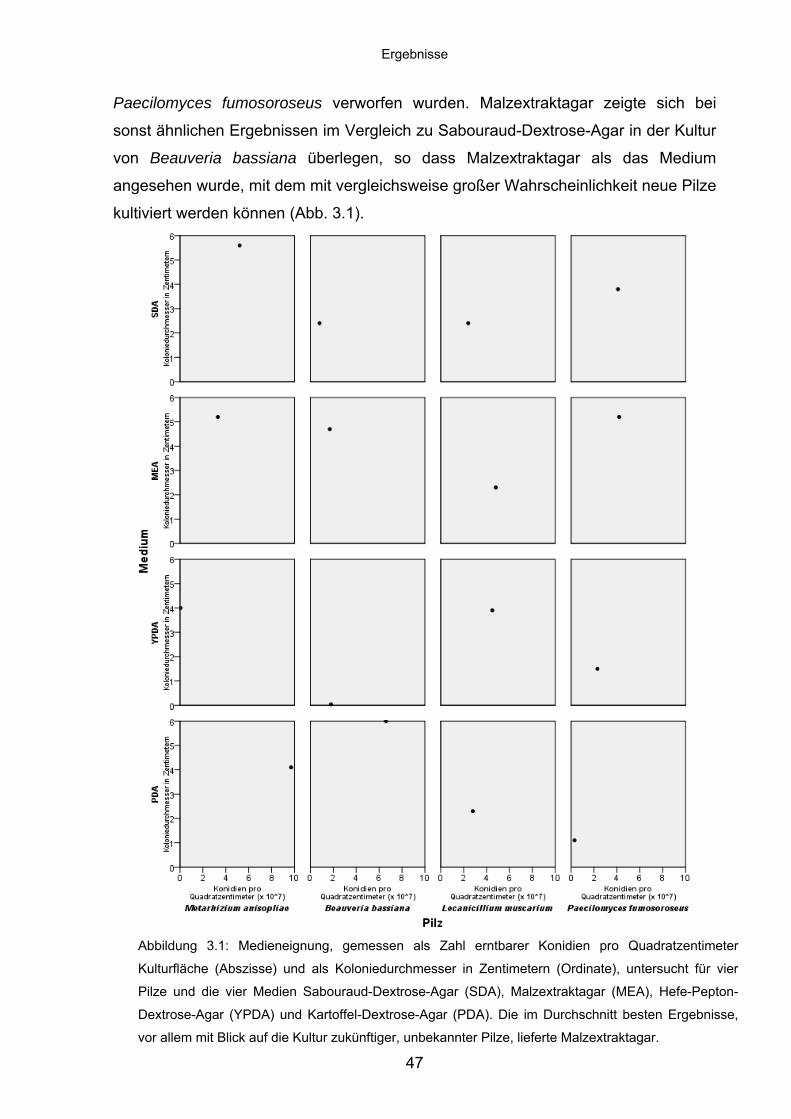
Ergebnisse
47
Paecilomyces fumosoroseus verworfen wurden. Malzextraktagar zeigte sich bei
sonst ähnlichen Ergebnissen im Vergleich zu Sabouraud-Dextrose-Agar in der Kultur
von Beauveria bassiana überlegen, so dass Malzextraktagar als das Medium
angesehen wurde, mit dem mit vergleichsweise großer Wahrscheinlichkeit neue Pilze
kultiviert werden können (Abb. 3.1).
Abbildung 3.1: Medieneignung, gemessen als Zahl erntbarer Konidien pro Quadratzentimeter
Kulturfläche (Abszisse) und als Koloniedurchmesser in Zentimetern (Ordinate), untersucht für vier
Pilze und die vier Medien Sabouraud-Dextrose-Agar (SDA), Malzextraktagar (MEA), Hefe-Pepton-
Dextrose-Agar (YPDA) und Kartoffel-Dextrose-Agar (PDA). Die im Durchschnitt besten Ergebnisse,
vor allem mit Blick auf die Kultur zukünftiger, unbekannter Pilze, lieferte Malzextraktagar.

Ergebnisse
48
3.1.2 Wirkung unterschiedlicher Oberflächensterilisationszeiten auf das
Pilzwachstum aus Milben
Die Beantwortung der Frage, ob ein Pilzwachstum nach Dosisapplikation
entomopathogener Pilze in den Versuchen im Labor- sowie im Halbfreilandmaßstab
nachweisbar war, setzte voraus, dass in der Kultur der Milben ein Wachstum der den
Milben äußerlich anhaftenden, opportunistischen Saprophyten oder anderer Pilze
weitestgehend ausgeschlossen werden konnte. Ebenso sollte im Screening auf
unbekannte Pilze ausschließlich eine Kultur interioren Ursprungs erfolgen.
Zu ermitteln war deshalb eine Mindeststerilisationszeit, die einen
weitestgehenden Ausschluss des Wachstums aus den Milben äußerlich anhaftenden
Diasporen sicherstellte. Gesammelte Milben wurden deshalb gepoolt und dem
Sterilans NaOCl für definierte Zeiträume ausgesetzt. Signifikante Reduktionen des
erzielbaren anteiligen Wachstums (Pilze pro 100 Milben) traten nach 20 und 120
Sekunden auf – in der Folge wurde deshalb eine Oberflächensterilisationszeit von
120 Sekunden verwendet (n = 212, p (a - b) < 0,01; p (a - c, b - c) < 0,001; χ2-Test,
zweiseitig; Abb. 3.2).
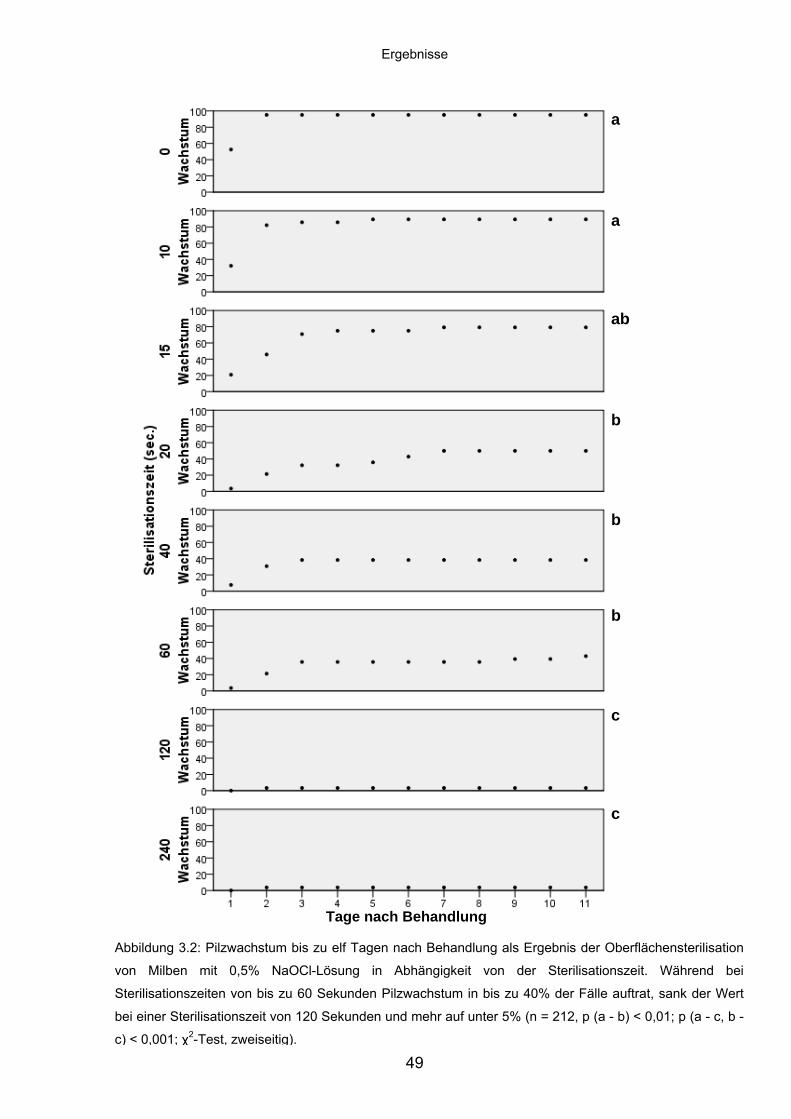
Ergebnisse
49
Abbildung 3.2: Pilzwachstum bis zu elf Tagen nach Behandlung als Ergebnis der Oberflächensterilisation
von Milben mit 0,5% NaOCl-Lösung in Abhängigkeit von der Sterilisationszeit. Während bei
Sterilisationszeiten von bis zu 60 Sekunden Pilzwachstum in bis zu 40% der Fälle auftrat, sank der Wert
bei einer Sterilisationszeit von 120 Sekunden und mehr auf unter 5% (n = 212, p (a - b) < 0,01; p (a - c, b -
c) < 0,001; χ2-Test, zweiseitig).
a
c
a
ab
b
b
b
c
Tage nach Behandlung
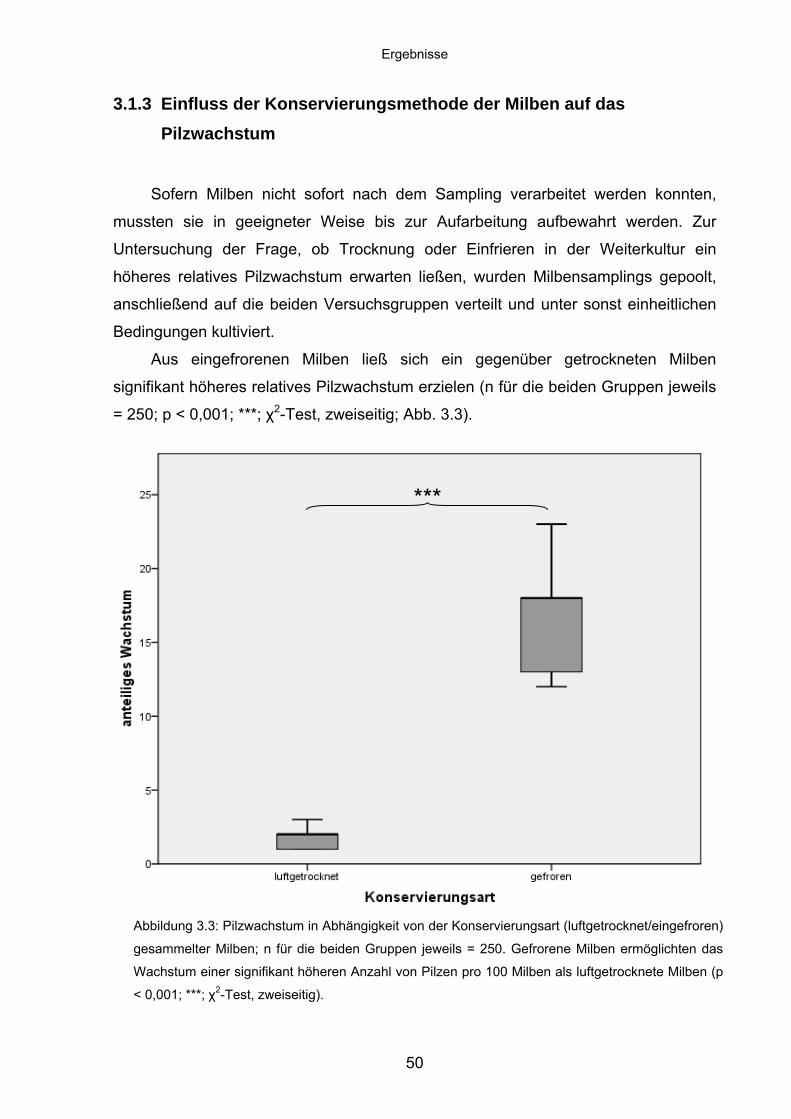
Ergebnisse
50
3.1.3 Einfluss der Konservierungsmethode der Milben auf das
Pilzwachstum
Sofern Milben nicht sofort nach dem Sampling verarbeitet werden konnten,
mussten sie in geeigneter Weise bis zur Aufarbeitung aufbewahrt werden. Zur
Untersuchung der Frage, ob Trocknung oder Einfrieren in der Weiterkultur ein
höheres relatives Pilzwachstum erwarten ließen, wurden Milbensamplings gepoolt,
anschließend auf die beiden Versuchsgruppen verteilt und unter sonst einheitlichen
Bedingungen kultiviert.
Aus eingefrorenen Milben ließ sich ein gegenüber getrockneten Milben
signifikant höheres relatives Pilzwachstum erzielen (n für die beiden Gruppen jeweils
= 250; p < 0,001; ***; χ2-Test, zweiseitig; Abb. 3.3).
***
Abbildung 3.3: Pilzwachstum in Abhängigkeit von der Konservierungsart (luftgetrocknet/eingefroren)
gesammelter Milben; n für die beiden Gruppen jeweils = 250. Gefrorene Milben ermöglichten das
Wachstum einer signifikant höheren Anzahl von Pilzen pro 100 Milben als luftgetrocknete Milben (p
< 0,001; ***; χ2-Test, zweiseitig).
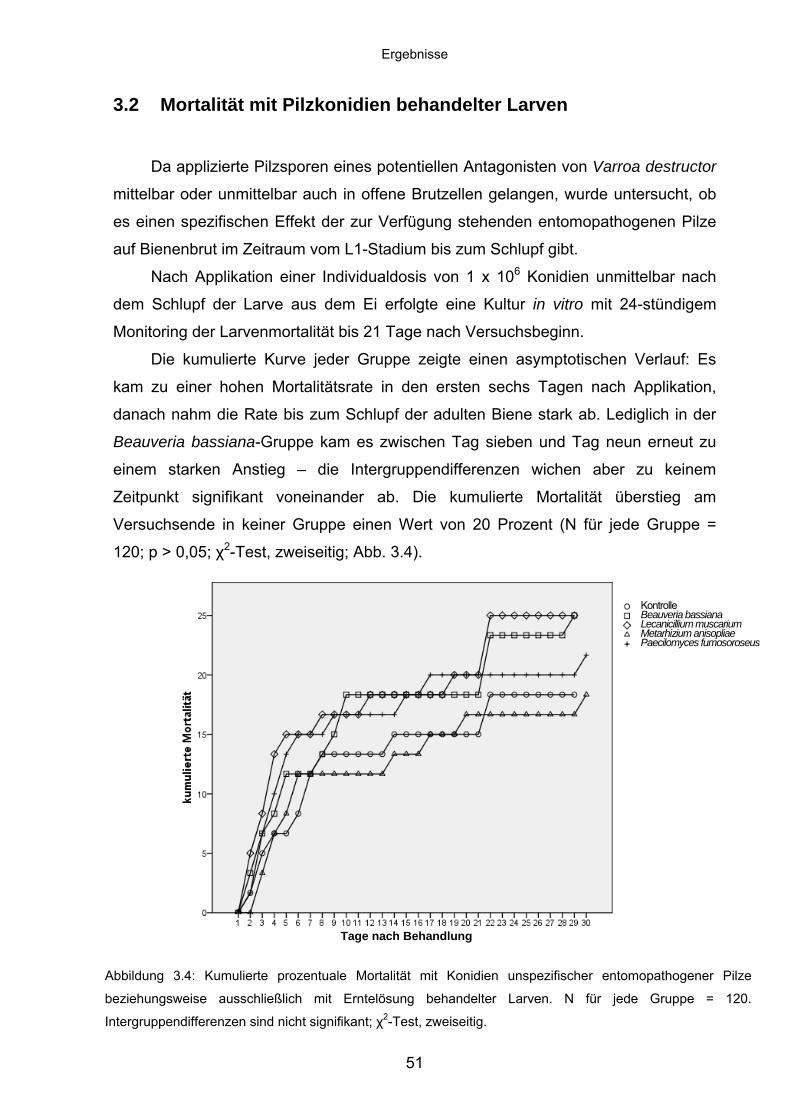
Ergebnisse
51
3.2 Mortalität mit Pilzkonidien behandelter Larven
Da applizierte Pilzsporen eines potentiellen Antagonisten von Varroa destructor
mittelbar oder unmittelbar auch in offene Brutzellen gelangen, wurde untersucht, ob
es einen spezifischen Effekt der zur Verfügung stehenden entomopathogenen Pilze
auf Bienenbrut im Zeitraum vom L1-Stadium bis zum Schlupf gibt.
Nach Applikation einer Individualdosis von 1 x 106 Konidien unmittelbar nach
dem Schlupf der Larve aus dem Ei erfolgte eine Kultur in vitro mit 24-stündigem
Monitoring der Larvenmortalität bis 21 Tage nach Versuchsbeginn.
Die kumulierte Kurve jeder Gruppe zeigte einen asymptotischen Verlauf: Es
kam zu einer hohen Mortalitätsrate in den ersten sechs Tagen nach Applikation,
danach nahm die Rate bis zum Schlupf der adulten Biene stark ab. Lediglich in der
Beauveria bassiana-Gruppe kam es zwischen Tag sieben und Tag neun erneut zu
einem starken Anstieg – die Intergruppendifferenzen wichen aber zu keinem
Zeitpunkt signifikant voneinander ab. Die kumulierte Mortalität überstieg am
Versuchsende in keiner Gruppe einen Wert von 20 Prozent (N für jede Gruppe =
120; p > 0,05; χ2-Test, zweiseitig; Abb. 3.4).
Abbildung 3.4: Kumulierte prozentuale Mortalität mit Konidien unspezifischer entomopathogener Pilze
beziehungsweise ausschließlich mit Erntelösung behandelter Larven. N für jede Gruppe = 120.
Intergruppendifferenzen sind nicht signifikant; χ2-Test, zweiseitig.
KontrolleBeauveria bassiana Lecanicillium muscarium Metarhizium anisopliae Paecilomyces fumosoroseus
Tage nach Behandlung

Ergebnisse
52
3.3 Mortalität mit Pilzkonidien behandelter Adultbienen
Im Fokus der bisherigen Untersuchungen zum Einsatz von Antagonisten gegen
Varroa destructor standen entomopathogene Pilze, die überwiegend aus anderen
Arthropodenklassen isoliert wurden. Um verlässliche Aussagen zu deren
Wirksamkeit in Bezug auf Apis mellifera treffen zu können, wurden die vier
verwendeten Pilzstämme in vitro unter exakt definierten Bedingungen getestet: Pro
Versuchsgruppe wurden je 50 Bienen nach Applikation einer single-bee-Dosis von
106 Konidien in Metallkäfigen im Wärmeschrank gehältert. Je Pilz wurden 60 Assays
angesetzt. Pollenteig, 2M-Zuckerlösung und Wasser standen ad libitum zur
Verfügung. Die Bienenmortalität wurde täglich erfasst. Die Kontrollgruppe wurde
analog behandelt, zu Beginn des Versuchs aber lediglich mit einem äquivalenten
Volumen Erntelösung ohne Konidien besprüht. Im Folgenden sind die Ergebnisse für
die vier untersuchten Pilze aufgelöst nach Einzeltagen und kumulativ dargestellt.
3.3.1 Lecanicillium muscarium: Mortalität/Versuchsgruppe und
kumulierte Mortalität
Bienen, die mit dem entomopathogenen Pilz Lecanicillium muscarium
behandelt wurden, zeigten im Vergleich zur Kontrollgruppe am vierten und siebten
Tag nach Applikation signifikant beziehungsweise höchstsignifikant erhöhte
Mortalitätsraten (N pro Versuchsgruppe = 50 Bienen; N gesamt = 60 Assays; p vier
Tage nach Behandlung < 0,05; *; p sieben Tage nach Behandlung < 0,001; ***; t-
Test, zweiseitig; Abb. 3.5). In der kumulierten prozentualen Mortalität unter dem
Einfluss von Lecanicillium muscarium zeigte sich nach analogem Verlauf der
Mortalitäten von Versuchsgruppe und Kontrollgruppe bis einschließlich des dritten
Tages nach Behandlung eine zunehmende Diskrepanz ab dem dritten Tag nach
Behandlung, in deren Folge die kumulierte Mortalität der Lecanicillium-Gruppe über
derjenigen der Kontrollgruppe lag. Diese Differenz führte über die gesamte
Versuchsdauer zu einer höchst signifikanten Differenz der kumulierten Raten (p <
0,001; χ2-Test, zweiseitig).
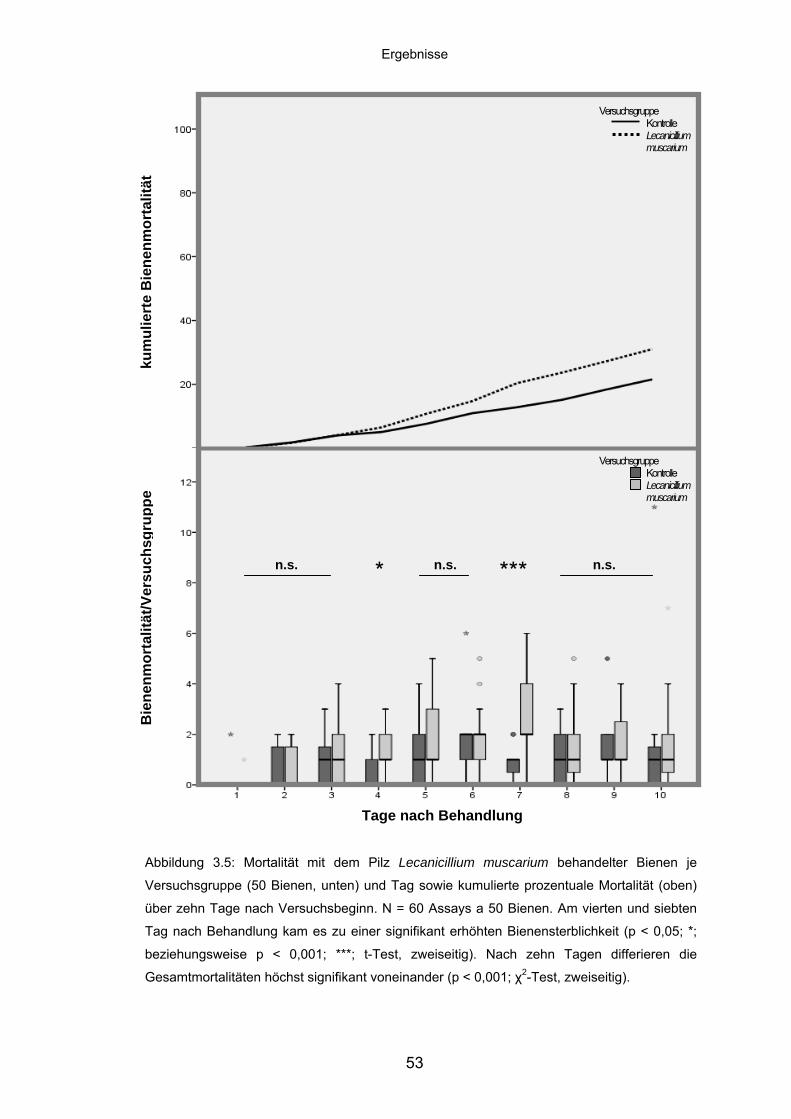
Ergebnisse
53
Tage nach Behandlung
Abbildung 3.5: Mortalität mit dem Pilz Lecanicillium muscarium behandelter Bienen je
Versuchsgruppe (50 Bienen, unten) und Tag sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 50 Bienen. Am vierten und siebten
Tag nach Behandlung kam es zu einer signifikant erhöhten Bienensterblichkeit (p < 0,05; *;
beziehungsweise p < 0,001; ***; t-Test, zweiseitig). Nach zehn Tagen differieren die
Gesamtmortalitäten höchst signifikant voneinander (p < 0,001; χ2-Test, zweiseitig).
Versuchsgruppe Kontrolle Lecanicillium muscarium
Versuchsgruppe Kontrolle Lecanicillium muscarium
* ***n.s. n.s. n.s.
kum
ulie
rte
Bie
nen
mo
rtal
ität
B
ien
enm
ort
alit
ät/V
ersu
chs
gru
pp
e

Ergebnisse
54
3.3.2 Metarhizium anisopliae: Mortalität/Versuchsgruppe und kumulierte
Mortalität
Nach Applikation der Versuchsdosis des Pilzes Metarhizium anisopliae kam es
nach einer sehr geringen Mortalität an den ersten beiden Tagen zu einem leichten
Anstieg des täglichen Totenfalls. Die Differenzen zwischen Kontrollgruppe und
Versuchsgruppe an den einzelnen Tagen waren aber zu keinem Zeitpunkt signifikant
(N pro Versuchsgruppe = 50 Bienen; N gesamt = 60 Assays; p > 0,05; t-Test,
zweiseitig; Abb. 3.6). Zwischen Kontrollgruppe und Versuchsgruppe traten unter dem
Einfluss von Metarhizium anisopliae in der kumulierten prozentualen Mortalität über
den gesamten Versuchsverlauf nur geringe Unterschiede auf. Der
Intergruppenvergleich zeigt keine signifikanten Differenzen (p > 0,05; χ2-Test,
zweiseitig).
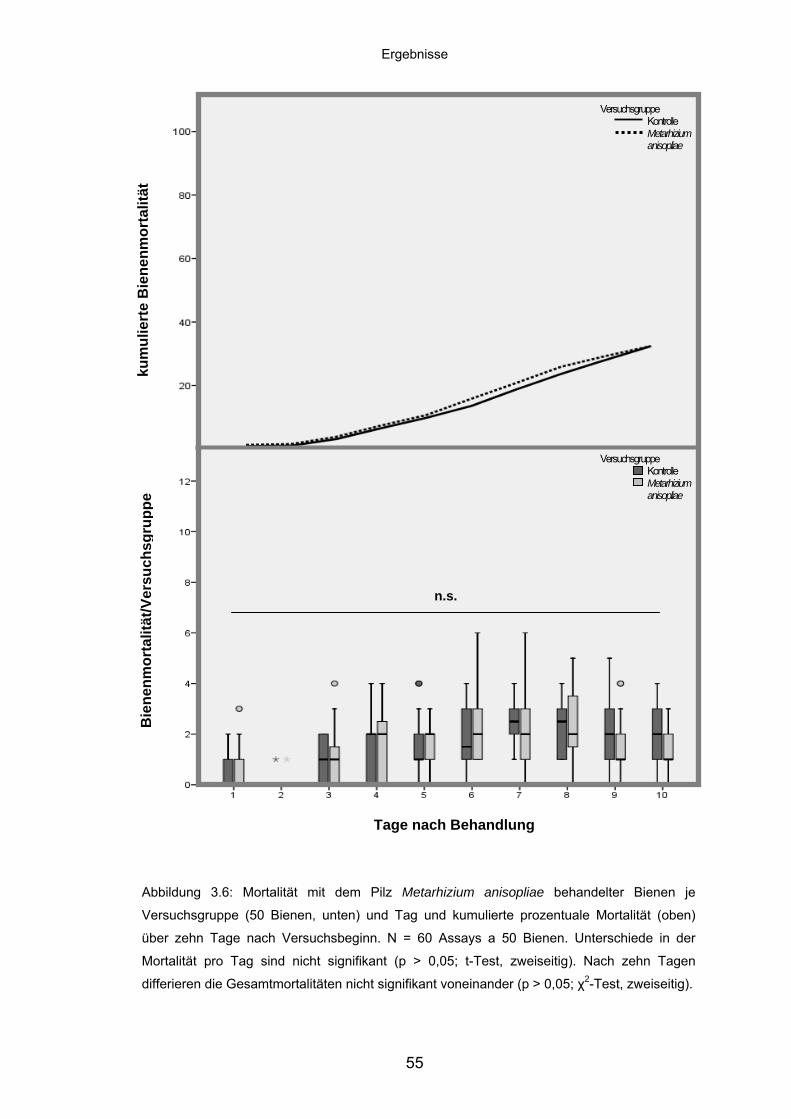
Ergebnisse
55
Abbildung 3.6: Mortalität mit dem Pilz Metarhizium anisopliae behandelter Bienen je
Versuchsgruppe (50 Bienen, unten) und Tag und kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 50 Bienen. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten nicht signifikant voneinander (p > 0,05; χ2-Test, zweiseitig).
days post treatment Tage nach Behandlung
kum
uli
erte
Bie
nen
mo
rtal
ität
B
ien
enm
ort
alit
ät/V
ersu
chs
gru
pp
e
Versuchsgruppe Kontrolle Metarhizium anisopliae
Versuchsgruppe Kontrolle Metarhizium anisopliae
n.s.

Ergebnisse
56
3.3.3 Beauveria bassiana: Mortalität/Versuchsgruppe und kumulierte
Mortalität
Unter dem Einfluss von Beauveria bassiana kam es nach einem sehr geringen
Bienentotenfall an den ersten beiden Tagen nach Dosisapplikation zu einer Zunahme
ab dem dritten Tag nach Behandlung. Signifikante Unterschiede zwischen den
beiden Versuchsgruppen traten am vierten und sechsten Tag nach Applikation auf:
An beiden Tagen kam es zu einem gegenüber der Kontrollgruppe erhöhten
Bienentotenfall in der mit Beauveria bassiana behandelten Gruppe (N pro
Versuchsgruppe = 50 Bienen; N gesamt = 60 Assays; p für Tag vier und Tag sechs
nach Behandlung < 0,05; *; t-Test, zweiseitig; Abb. 3.7). Der erhöhte Bienentotenfall
in der mit dem entomopathogenen Pilz Beauveria bassiana behandelten Gruppe an
den Tagen vier und sechs spiegelt sich in einer deutlichen Divergenz der kumulierten
prozentualen Mortalitäten an diesen Tagen wieder. Infolgedessen lag die
Gesamtmortalität der Pilzgruppe zum Versuchsende zehn Tage nach Applikation
höchst signifikant über derjenigen der Kontrollgruppe (p < 0,001; χ2-Test, zweiseitig).
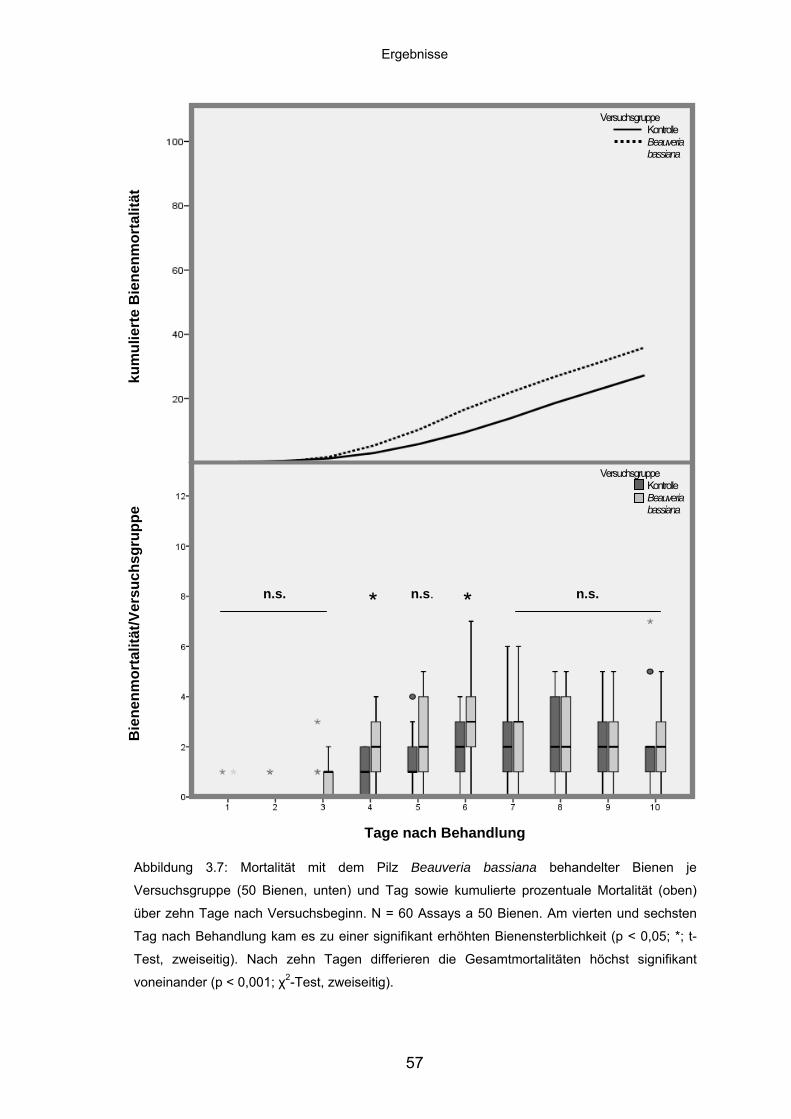
Ergebnisse
57
Abbildung 3.7: Mortalität mit dem Pilz Beauveria bassiana behandelter Bienen je
Versuchsgruppe (50 Bienen, unten) und Tag sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 50 Bienen. Am vierten und sechsten
Tag nach Behandlung kam es zu einer signifikant erhöhten Bienensterblichkeit (p < 0,05; *; t-
Test, zweiseitig). Nach zehn Tagen differieren die Gesamtmortalitäten höchst signifikant
voneinander (p < 0,001; χ2-Test, zweiseitig).
days post treatment
Tage nach Behandlung
kum
uli
erte
Bie
nen
mo
rtal
ität
B
ien
enm
ort
alit
ät/V
ersu
chsg
rup
pe
Versuchsgruppe Kontrolle Beauveria bassiana
Versuchsgruppe Kontrolle Beauveria bassiana
n.s. n.s. n.s. * *

Ergebnisse
58
3.3.4 Paecilomyces fumosoroseus: Mortalität/Versuchsgruppe und
kumulierte Mortalität
Nach Applikation der Versuchsdosis des Pilzes Paecilomyces fumosoroseus kam es
nach einer sehr geringen Mortalität an den ersten beiden Tagen zu einer leichten
Zunahme des täglichen Totenfalls bis Tag sieben nach Behandlung; danach sank die
Mortalitätsrate wieder leicht. Die Differenzen zwischen Kontrollgruppe und der mit
Paecilomyces fumosoroseus behandelten Versuchsgruppe an den einzelnen Tagen
waren zu keinem Zeitpunkt signifikant (N pro Versuchsgruppe = 50 Bienen; N gesamt
= 60 Assays; p > 0,05; t-Test, zweiseitig; Abb. 3.8). Ab Tag drei nach Applikation von
Konidiendosis beziehungsweise Kontrolllösung kam es zu einer leichten Divergenz
der kumulierten Mortalitäten zwischen Paecilomyces fumosoroseus- und
Kontrollgruppe; nach zehn Tagen wichen die kumulierten Mortalitäten höchst
signifikant voneinander ab (p < 0,001; χ2-Test, zweiseitig).
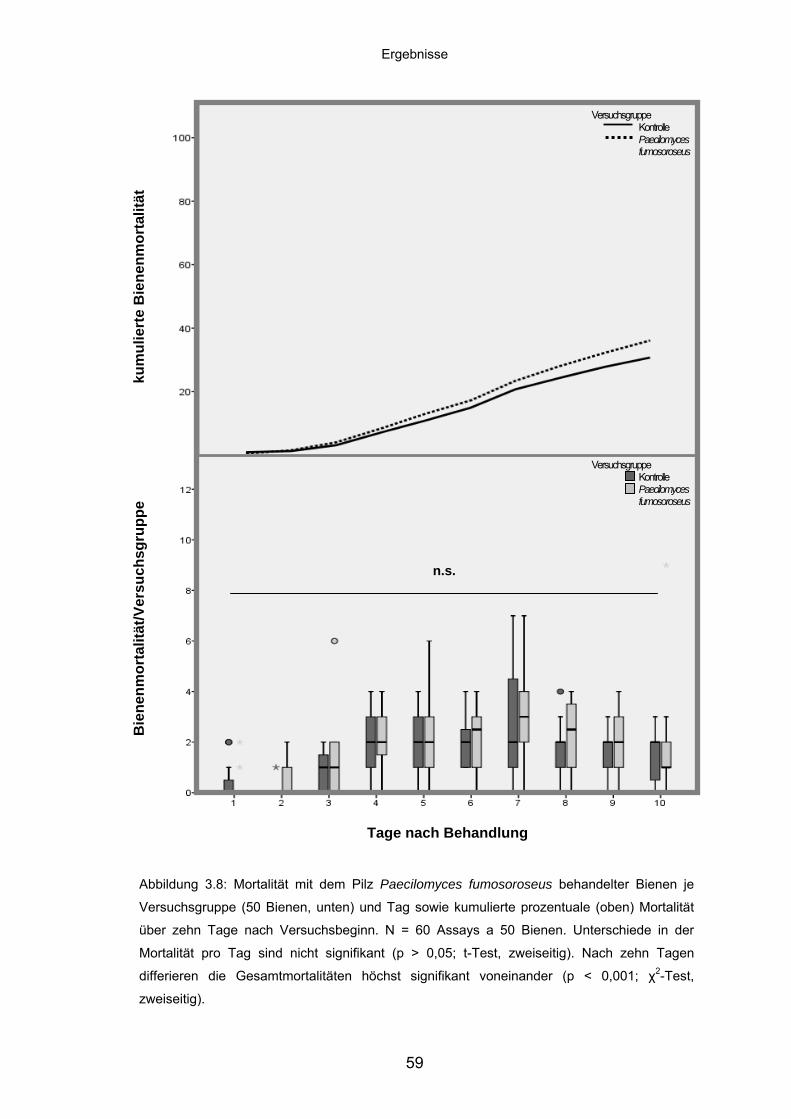
Ergebnisse
59
Abbildung 3.8: Mortalität mit dem Pilz Paecilomyces fumosoroseus behandelter Bienen je
Versuchsgruppe (50 Bienen, unten) und Tag sowie kumulierte prozentuale (oben) Mortalität
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 50 Bienen. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten höchst signifikant voneinander (p < 0,001; χ2-Test,
zweiseitig).
kum
ulie
rte
Bie
nen
mo
rtal
ität
B
ien
enm
ort
alit
ät/V
ersu
chsg
rup
pe
days post treatment
Versuchsgruppe Kontrolle Paecilomyces fumosoroseus
Versuchsgruppe Kontrolle Paecilomyces fumosoroseus
n.s.
Tage nach Behandlung

Ergebnisse
60
3.4 Mortalität mit Pilzkonidien behandelter Milben
Ebenfalls im in vitro-Assay wurde untersucht, ob die applizierten single-bee-
Dosen Auswirkungen auf die Überlebensrate adulter Weibchen von Varroa
destructor in der phoretischen Phase haben: Pro Versuchsgruppe wurden je 50
Bienen aus Völkern, die unmittelbar zuvor eine Behandlung mit Varroaziden
durchlaufen hatten, aber rückstandsfrei waren, mit genau 25 Milben besetzt, die
direkt vor Versuchsbeginn aus hochbefallenen Völkern gewonnen wurden. Nach
Applikation von Konidiensuspension beziehungsweise Kontrolllösung wurde die
Milbenmortalität täglich erfasst. Im Folgenden sind die Ergebnisse für die vier
untersuchten Pilze aufgelöst nach Einzeltagen und kumulativ dargestellt.
3.4.1 Lecanicillium muscarium: Mortalität/Versuchsgruppe und
kumulierte Mortalität
Nach Applikation der Versuchsdosis des Pilzes Lecanicillium muscarium stieg
die Mortalitätsrate zunächst an und erreichte Maxima an den Tagen drei und fünf
nach Behandlung mit einem Milbenfall von etwa vier Milben pro Assay und Tag.
Differenzen zwischen Kontrollgruppe und der mit Lecanicillium muscarium
behandelten Versuchsgruppe an den einzelnen Tagen waren zu keinem Zeitpunkt
signifikant (N pro Versuchsgruppe = 50 Bienen und 25 Milben; N gesamt = 60
Assays; p > 0,05; t-Test, zweiseitig; Abb. 3.9). Zwischen Kontrollgruppe und
Versuchsgruppe traten unter dem Einfluss von Lecanicillium muscarium in der
kumulierten prozentualen Mortalität über den gesamten Versuchsverlauf nur geringe
Unterschiede an den Tagen drei und sechs/sieben auf. Der Intergruppenvergleich
zeigt zum Versuchsende keine signifikanten Differenzen (p > 0,05; χ2-Test,
zweiseitig).
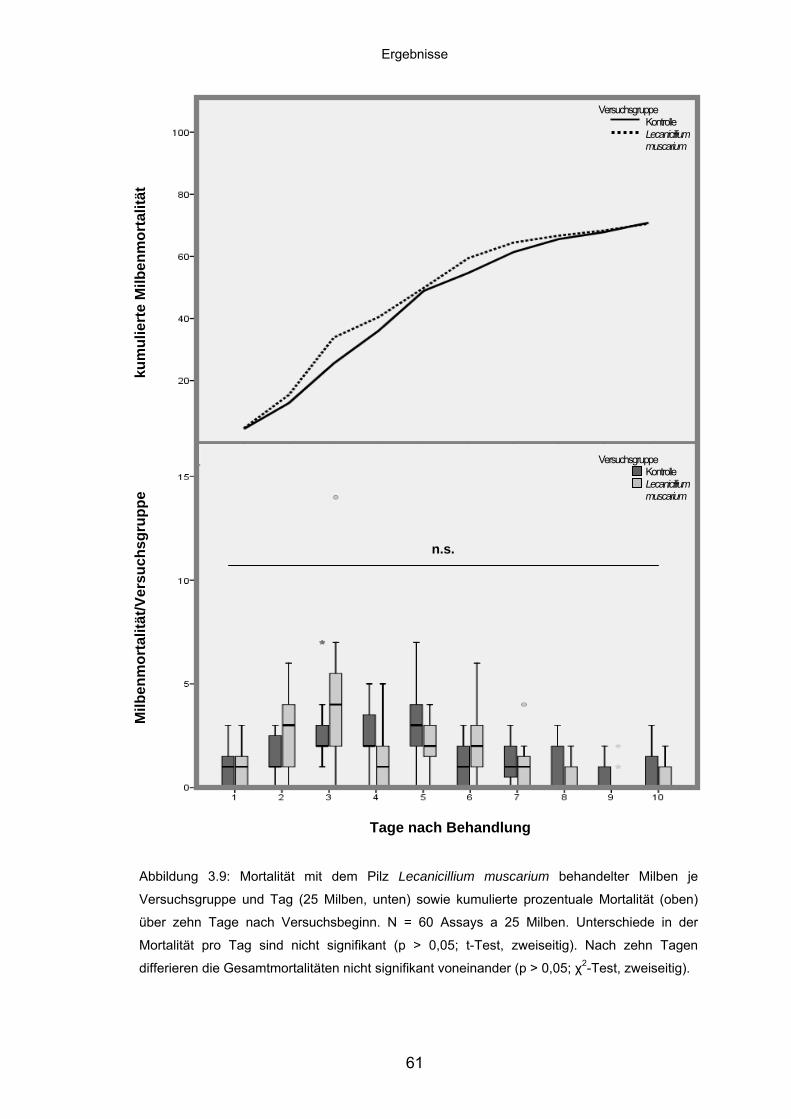
Ergebnisse
61
Abbildung 3.9: Mortalität mit dem Pilz Lecanicillium muscarium behandelter Milben je
Versuchsgruppe und Tag (25 Milben, unten) sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 25 Milben. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten nicht signifikant voneinander (p > 0,05; χ2-Test, zweiseitig).
kum
ulie
rte
Milb
enm
ort
alit
ät
Milb
enm
ort
alit
ät/V
ersu
chsg
rup
pe
days post treatment
n.s.
Versuchsgruppe Kontrolle Lecanicillium muscarium
Versuchsgruppe Kontrolle Lecanicillium muscarium
Tage nach Behandlung

Ergebnisse
62
3.4.2 Metarhizium anisopliae: Mortalität/Versuchsgruppe und kumulierte
Mortalität
Die Mortalitätsrate unter dem Einfluss von Metarhizium anisopliae erreichte ihre
Maxima an den Tagen drei, fünf und sechs, bevor sie bis zum Ende der
Untersuchung zehn Tage nach Applikation kontinuierlich sank. Deutliche Differenzen
zwischen Kontrollgruppe und Pilzgruppe traten nicht auf; die Abweichungen der
Verteilungen an den Einzeltagen waren zu keinem Zeitpunkt des untersuchten
Zeitraums signifikant (N pro Versuchsgruppe = 50 Bienen und 25 Milben; N gesamt =
60 Assays; p > 0,05; t-Test, zweiseitig; Abb. 3.10). Zu keinem Zeitpunkt des
Versuchs hatte der entomopathogene Pilz Metarhizium anisopliae einen Einfluss auf
die Mortalitätsrate der Pilzgruppe, der sich in einer deutlichen Divergenz der
kumulierten Mortalitäten widerspiegelte – zum Versuchsende traten keine
signifikanten Differenzen auf (p > 0,05; χ2-Test, zweiseitig).
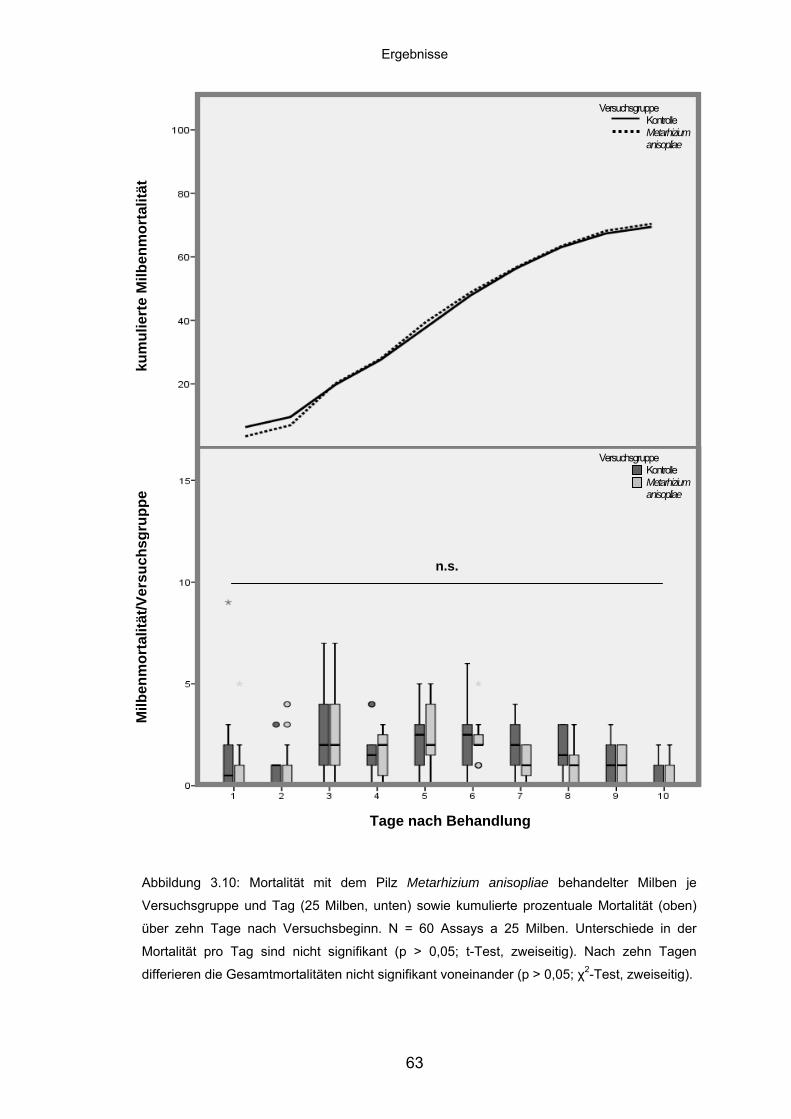
Ergebnisse
63
Abbildung 3.10: Mortalität mit dem Pilz Metarhizium anisopliae behandelter Milben je
Versuchsgruppe und Tag (25 Milben, unten) sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 25 Milben. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten nicht signifikant voneinander (p > 0,05; χ2-Test, zweiseitig).
kum
ulie
rte
Milb
enm
ort
alit
ät
Milb
enm
ort
alit
ät/V
ersu
chs g
rup
pe
days post treatment
n.s.
Versuchsgruppe Kontrolle Metarhizium anisopliae
Versuchsgruppe Kontrolle Metarhizium anisopliae
Tage nach Behandlung

Ergebnisse
64
3.4.3 Beauveria bassiana: Mortalität/Versuchsgruppe und kumulierte
Mortalität
Im Beauveria bassiana-Versuch stieg die Mortalitätsrate innerhalb der ersten
drei Versuchstage zunächst an, um dann wieder in etwa auf das Ausgangsniveau
abzusinken. An den Tagen fünf und sechs lag die Mortalitätsrate der Pilzgruppe über
der der Kontrollgruppe, an den Tagen sieben und zehn wies die Kontrollgruppe eine
erhöhte Mortalitätsrate auf – allerdings waren die Abweichungen der Verteilungen an
den Einzeltagen zu keinem Zeitpunkt des untersuchten Zeitraums signifikant (N pro
Versuchsgruppe = 50 Bienen und 25 Milben; N gesamt = 60 Assays; p > 0,05; t-Test,
zweiseitig; Abb. 3.11). Die voneinander abweichenden Verteilungen – auf nicht
signifikantem Niveau – der Mortalitätsraten von Pilz- und Kontrollgruppe in der
Betrachtung der Einzeltage des Beauveria bassiana-Versuchs führen zu einem
divergierenden Kurvenverlauf bei Auftragung der kumulierten Mortalitätsraten. Diese
wichen aber nach zehn Tagen nicht signifikant voneinander ab (p > 0,05; χ2-Test,
zweiseitig).
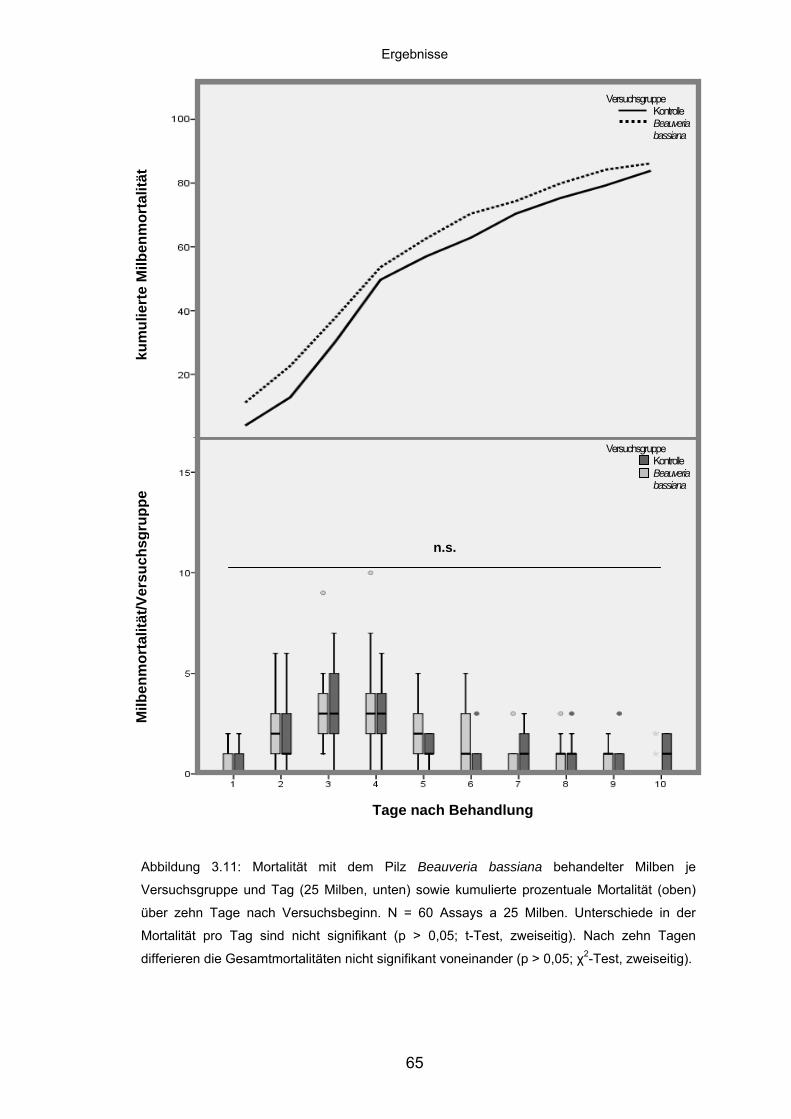
Ergebnisse
65
Abbildung 3.11: Mortalität mit dem Pilz Beauveria bassiana behandelter Milben je
Versuchsgruppe und Tag (25 Milben, unten) sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 25 Milben. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten nicht signifikant voneinander (p > 0,05; χ2-Test, zweiseitig).
kum
ulie
rte
Milb
enm
ort
alit
ät
Milb
enm
ort
alit
ät/V
ersu
chs g
rup
pe
days post treatment
n.s.
Versuchsgruppe Kontrolle Beauveria bassiana
Versuchsgruppe Kontrolle Beauveria bassiana
Tage nach Behandlung

Ergebnisse
66
3.4.4 Paecilomyces fumosoroseus: Mortalität/Versuchsgruppe und
kumulierte Mortalität
Zwischen Tag zwei und fünf lag die Mortalitätsrate im Paecilomyces
fumosoroseus-Versuch in beiden Gruppen bei etwa zwei Milben pro Tag;
Abweichungen der Verteilungen voneinander traten auf, waren aber zu keinem
Zeitpunkt des Versuchs signifikant (N pro Versuchsgruppe = 50 Bienen und 25
Milben; N gesamt = 60 Assays; p > 0,05; t-Test, zweiseitig; Abb. 3.12). Bis Tag
sieben nach Behandlung kam es in der Untersuchung des entomopathogenen Pilzes
Paecilomyces fumosoroseus zu einem weitestgehend ähnlichen Verlauf der
kumulierten Mortalitäten. In der Folge divergierten die Kurven zwar, so dass in der
Summe zum Versuchsende die Mortalität der Pilzgruppe über derjenigen der
Kontrollgruppe lag, diese Differenz war allerdings nicht signifikant (p > 0,05; χ2-Test,
zweiseitig).
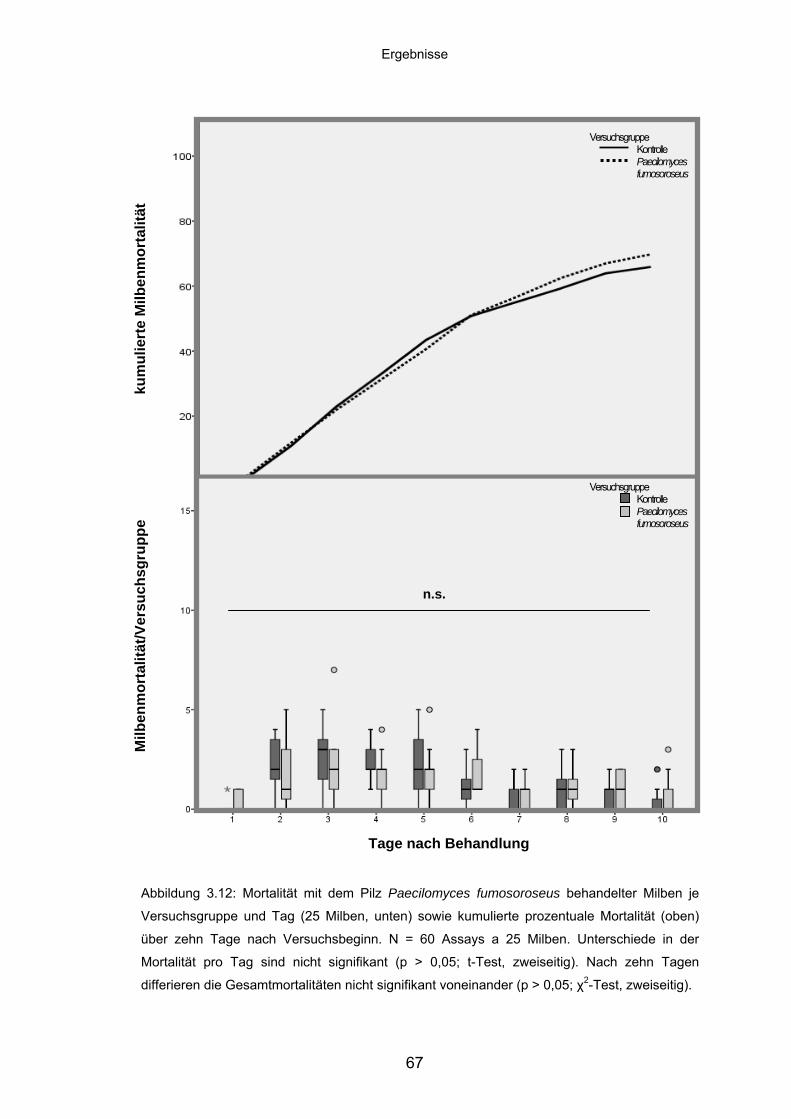
Ergebnisse
67
Abbildung 3.12: Mortalität mit dem Pilz Paecilomyces fumosoroseus behandelter Milben je
Versuchsgruppe und Tag (25 Milben, unten) sowie kumulierte prozentuale Mortalität (oben)
über zehn Tage nach Versuchsbeginn. N = 60 Assays a 25 Milben. Unterschiede in der
Mortalität pro Tag sind nicht signifikant (p > 0,05; t-Test, zweiseitig). Nach zehn Tagen
differieren die Gesamtmortalitäten nicht signifikant voneinander (p > 0,05; χ2-Test, zweiseitig).
kum
ulie
rte
Milb
enm
ort
alit
ät
Milb
enm
ort
alit
ät/V
ersu
chsg
rup
pe
days post treatment
Versuchsgruppe Kontrolle Paecilomyces fumosoroseus
Versuchsgruppe Kontrolle Paecilomyces fumosoroseus
Tage nach Behandlung
n.s.

Ergebnisse
68
3.5 Pilzwachstum aus behandelten Milben
Um einen signifikanten Einfluss entomopathogener Pilze auf die Mortalität von
Varroa destructor im Laborassay verifizieren zu können, wurde untersucht, welche
Pilzwachstumsrate (Anteil Milben, aus denen ein Pilz wuchs) sich aus den im Milben-
in vitro-Versuch gestorbenen Milben erzielen ließ: Die in diesem Versuch
gestorbenen Milben wurden täglich abgesammelt, gepoolt und ein Aliquant kultiviert.
Im anschließenden Monitoring bis Tag 13 nach Behandlung wurde das tägliche
Pilzwachstum aus Einzelmilben erfasst; Einzelmilben, die Pilzwachstum zeigten,
wurden zur Verhinderung von Kreuzinfektionen steril entfernt. Im Folgenden sind die
Ergebnisse von Pilz- und Kontrollgruppe für den jeweiligen untersuchten Pilz
einander gegenübergestellt. Auf der Ordinate aufgetragen ist das kumulierte,
prozentuale Pilzwachstum in Prozent an den jeweiligen Tagen nach
Behandlungsbeginn, auf der Abszisse sind die Tage des Samplings der Milben
aufgetragen. Unterschiedliche Kreisgrößen geben das prozentuale Wachstum in
Prozent in sechs Kategorien wieder (0 - 5, 5 - 10, 10 - 15, 15 - 20, 20 - 25). Der
verstrichene Zeitraum bis zu einer Zunahme im Wachstum der jeweiligen
Einzelgruppe ergibt sich aus der Differenz von Ordinaten- und Abszissenbetrag.
3.5.1 Lecanicillium muscarium
In der Kultur der während der Versuchsdauer des Lecanicillium muscarium-
Versuchs gesammelten Milben setzte ein detektierbares Pilzwachstum in den
jeweiligen Tagesgruppen etwa vier bis fünf Tage nach dem Tod der Milben ein.
Erhöhte Wachstumsraten traten in der Kontrolle bei den an den Tagen fünf, sieben
und acht nach Versuchsbeginn gestorbenen Milben auf, in der Pilzgruppe an Tag
acht. Der Intergruppenvergleich dieser Tage zeigte aber keine signifikanten
Differenzen (N Kontrolle = 128, N Lecanicillium muscarium = 146; p für alle
Vergleiche > 0,05, Fishers Exact Test, zweiseitig; Abb. 3.13).
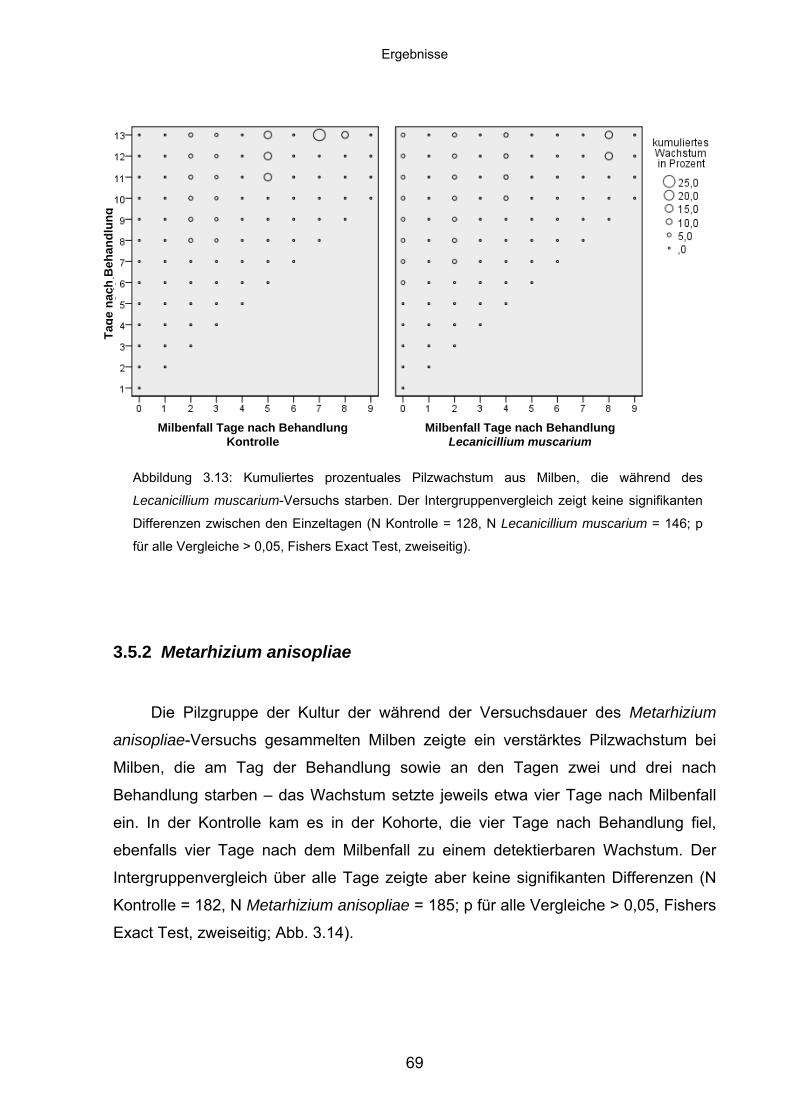
Ergebnisse
69
3.5.2 Metarhizium anisopliae
Die Pilzgruppe der Kultur der während der Versuchsdauer des Metarhizium
anisopliae-Versuchs gesammelten Milben zeigte ein verstärktes Pilzwachstum bei
Milben, die am Tag der Behandlung sowie an den Tagen zwei und drei nach
Behandlung starben – das Wachstum setzte jeweils etwa vier Tage nach Milbenfall
ein. In der Kontrolle kam es in der Kohorte, die vier Tage nach Behandlung fiel,
ebenfalls vier Tage nach dem Milbenfall zu einem detektierbaren Wachstum. Der
Intergruppenvergleich über alle Tage zeigte aber keine signifikanten Differenzen (N
Kontrolle = 182, N Metarhizium anisopliae = 185; p für alle Vergleiche > 0,05, Fishers
Exact Test, zweiseitig; Abb. 3.14).
Abbildung 3.13: Kumuliertes prozentuales Pilzwachstum aus Milben, die während des
Lecanicillium muscarium-Versuchs starben. Der Intergruppenvergleich zeigt keine signifikanten
Differenzen zwischen den Einzeltagen (N Kontrolle = 128, N Lecanicillium muscarium = 146; p
für alle Vergleiche > 0,05, Fishers Exact Test, zweiseitig).
Milbenfall days post treatment Kontrolle
Milbenfall days post treatment Lecanicillium muscarium
days
pos
t tre
atm
ent
Milbenfall Tage nach BehandlungKontrolle
Milbenfall Tage nach Behandlung Lecanicillium muscarium
Tag
e n
ach
Beh
and
lun
g
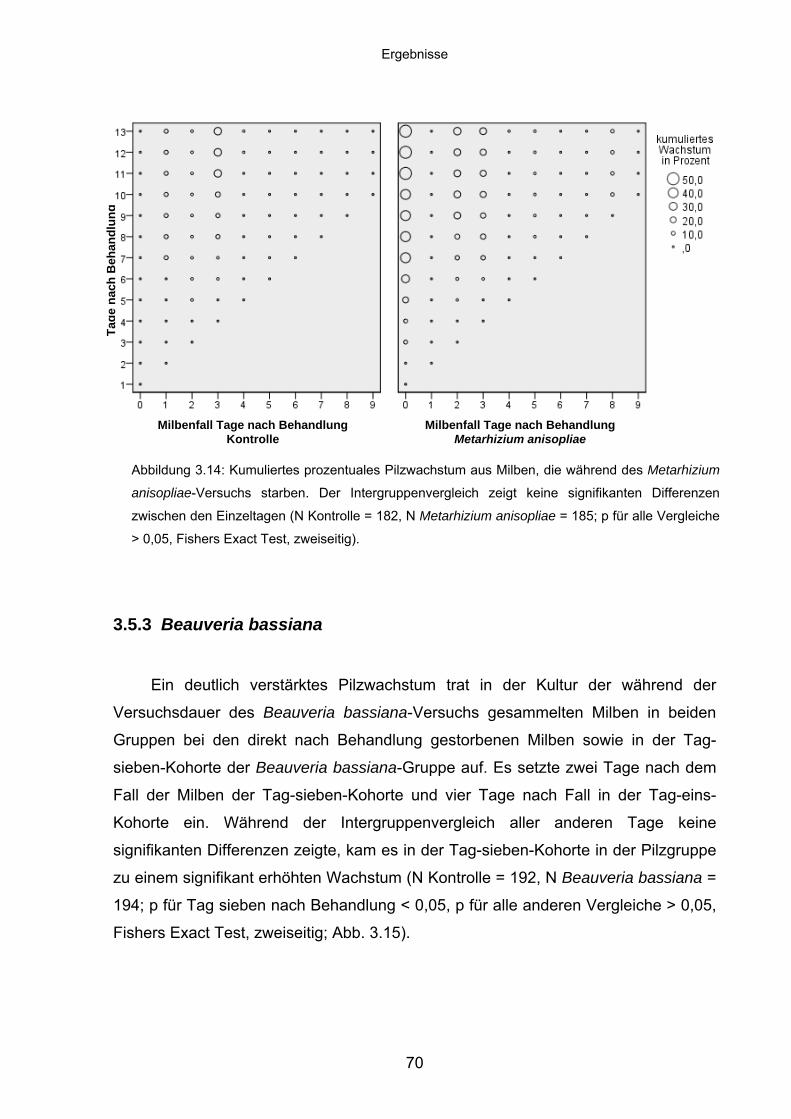
Ergebnisse
70
3.5.3 Beauveria bassiana
Ein deutlich verstärktes Pilzwachstum trat in der Kultur der während der
Versuchsdauer des Beauveria bassiana-Versuchs gesammelten Milben in beiden
Gruppen bei den direkt nach Behandlung gestorbenen Milben sowie in der Tag-
sieben-Kohorte der Beauveria bassiana-Gruppe auf. Es setzte zwei Tage nach dem
Fall der Milben der Tag-sieben-Kohorte und vier Tage nach Fall in der Tag-eins-
Kohorte ein. Während der Intergruppenvergleich aller anderen Tage keine
signifikanten Differenzen zeigte, kam es in der Tag-sieben-Kohorte in der Pilzgruppe
zu einem signifikant erhöhten Wachstum (N Kontrolle = 192, N Beauveria bassiana =
194; p für Tag sieben nach Behandlung < 0,05, p für alle anderen Vergleiche > 0,05,
Fishers Exact Test, zweiseitig; Abb. 3.15).
Abbildung 3.14: Kumuliertes prozentuales Pilzwachstum aus Milben, die während des Metarhizium
anisopliae-Versuchs starben. Der Intergruppenvergleich zeigt keine signifikanten Differenzen
zwischen den Einzeltagen (N Kontrolle = 182, N Metarhizium anisopliae = 185; p für alle Vergleiche
> 0,05, Fishers Exact Test, zweiseitig).
Milbenfall days post treatment Kontrolle
Milbenfall days post treatment Metarhizium anisopliae
days
pos
t tre
atm
ent
Milbenfall Tage nach BehandlungKontrolle
Milbenfall Tage nach Behandlung Metarhizium anisopliae
Tag
e n
ach
Beh
and
lun
g
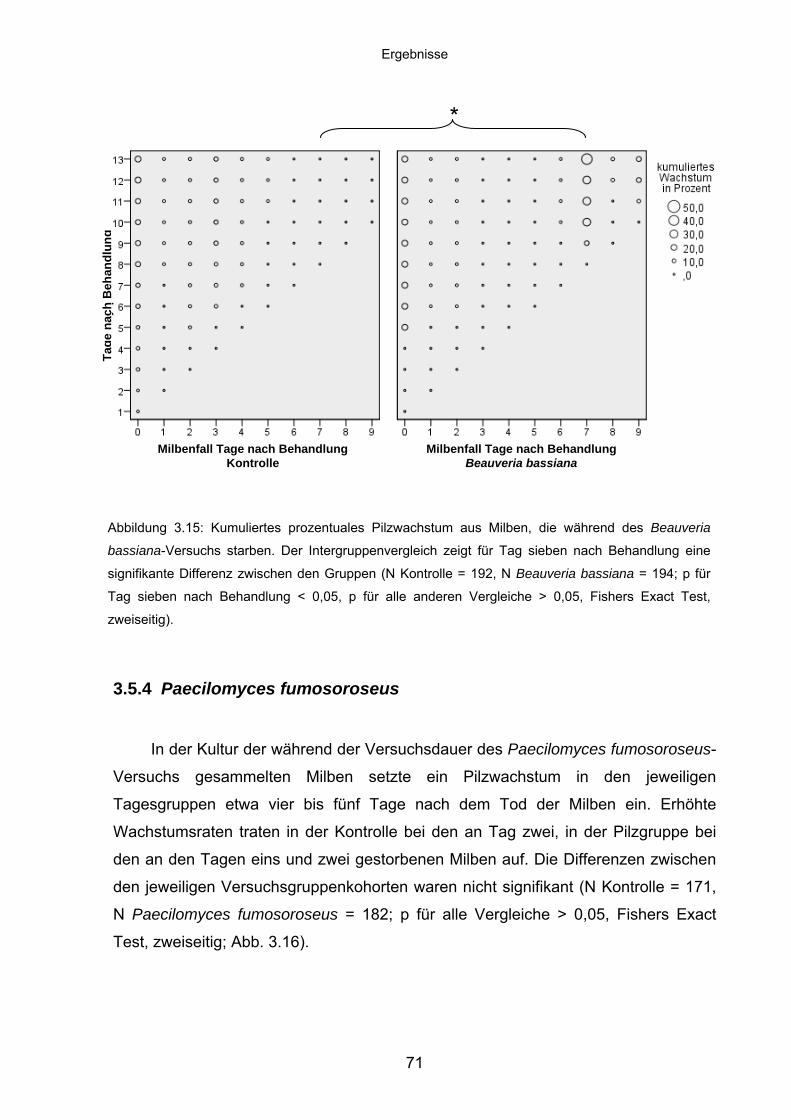
Ergebnisse
71
3.5.4 Paecilomyces fumosoroseus
In der Kultur der während der Versuchsdauer des Paecilomyces fumosoroseus-
Versuchs gesammelten Milben setzte ein Pilzwachstum in den jeweiligen
Tagesgruppen etwa vier bis fünf Tage nach dem Tod der Milben ein. Erhöhte
Wachstumsraten traten in der Kontrolle bei den an Tag zwei, in der Pilzgruppe bei
den an den Tagen eins und zwei gestorbenen Milben auf. Die Differenzen zwischen
den jeweiligen Versuchsgruppenkohorten waren nicht signifikant (N Kontrolle = 171,
N Paecilomyces fumosoroseus = 182; p für alle Vergleiche > 0,05, Fishers Exact
Test, zweiseitig; Abb. 3.16).
Abbildung 3.15: Kumuliertes prozentuales Pilzwachstum aus Milben, die während des Beauveria
bassiana-Versuchs starben. Der Intergruppenvergleich zeigt für Tag sieben nach Behandlung eine
signifikante Differenz zwischen den Gruppen (N Kontrolle = 192, N Beauveria bassiana = 194; p für
Tag sieben nach Behandlung < 0,05, p für alle anderen Vergleiche > 0,05, Fishers Exact Test,
zweiseitig).
Milbenfall days post treatment Kontrolle
Milbenfall days post treatment Beauveria bassiana
days
pos
t tre
atm
ent
*
Milbenfall Tage nach BehandlungKontrolle
Milbenfall Tage nach Behandlung Beauveria bassiana
Tag
e n
ach
Beh
and
lun
g

Ergebnisse
72
3.6 Wirkung entomopathogener Pilze auf Adultbienen,
Bienenlarven und -puppen sowie Varroa destructor im
Halbfreilandversuch
Insbesondere bei staatenbildenden Insekten sind in vitro-Versuche hoch
artifiziell – eine Übertragbarkeit ihrer Schlussfolgerungen auf die Verhältnisse, wie
sie unter Freilandbedingungen innerhalb eines Staates vorkommen, ist deshalb nicht
ohne Weiteres möglich. Im Halbfreilandversuch wurde deshalb ergänzend zu den
Laborversuchen exemplarisch die Wirkung eines entomopathogenen Pilzes –
Lecanicillium muscarium – auf die Zahl adulter Bienen, Larvenstadien und Puppen
sowie die Abundanz von Varroa destructor analysiert. Der Versuch wurde an zehn in
Bezug auf Verwandtschaftsgrad und Alterstruktur der Einzelbienen sowie ihren Befall
mit Varroa destructor vereinheitlichten, je etwa 5500 Individuen starken Völkern
vorgenommen. Der Milbenfall wurde täglich, Bienenzahl und Zahl der Brutstadien
wurden zu Beginn und nach sieben, 14 und 17 Tagen erfasst.
Abbildung 3.16: Kumuliertes prozentuales Pilzwachstum aus Milben, die während des Paecilomyces
fumosoroseus-Versuchs starben. Der Intergruppenvergleich zeigt keine signifikanten Differenzen
zwischen den Einzeltagen (N Kontrolle = 171, N Paecilomyces fumosoroseus = 182; p für alle
Vergleiche > 0,05, Fishers Exact Test, zweiseitig).
Milbenfall days post treatment Kontrolle
Milbenfall days post treatment Paecilomyces fumosoroseus
days
pos
t tre
atm
ent
Milbenfall Tage nach BehandlungKontrolle
Milbenfall Tage nach Behandlung Paecilomyces fumosoroseus
Tag
e n
ach
Beh
and
lun
g
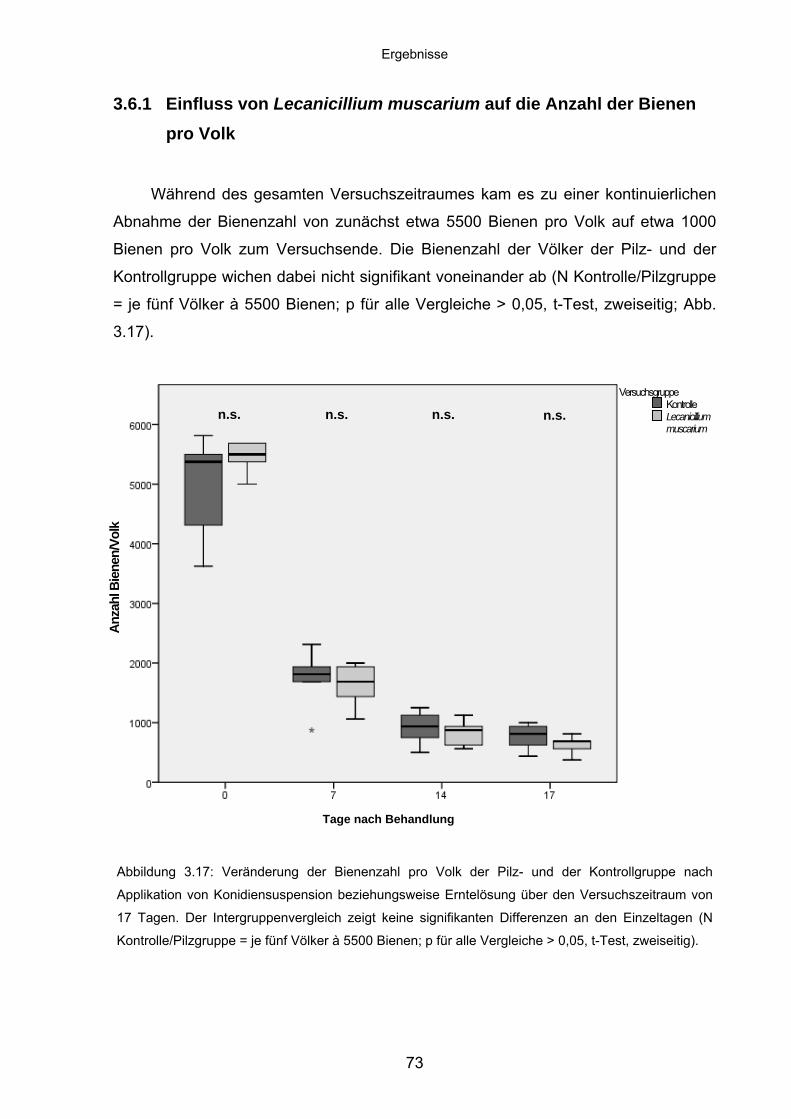
Ergebnisse
73
3.6.1 Einfluss von Lecanicillium muscarium auf die Anzahl der Bienen
pro Volk
Während des gesamten Versuchszeitraumes kam es zu einer kontinuierlichen
Abnahme der Bienenzahl von zunächst etwa 5500 Bienen pro Volk auf etwa 1000
Bienen pro Volk zum Versuchsende. Die Bienenzahl der Völker der Pilz- und der
Kontrollgruppe wichen dabei nicht signifikant voneinander ab (N Kontrolle/Pilzgruppe
= je fünf Völker à 5500 Bienen; p für alle Vergleiche > 0,05, t-Test, zweiseitig; Abb.
3.17).
Abbildung 3.17: Veränderung der Bienenzahl pro Volk der Pilz- und der Kontrollgruppe nach
Applikation von Konidiensuspension beziehungsweise Erntelösung über den Versuchszeitraum von
17 Tagen. Der Intergruppenvergleich zeigt keine signifikanten Differenzen an den Einzeltagen (N
Kontrolle/Pilzgruppe = je fünf Völker à 5500 Bienen; p für alle Vergleiche > 0,05, t-Test, zweiseitig).
n.s. n.s. n.s. n.s.
Versuchsgruppe Kontrolle Lecanicillium muscarium
An
zah
l Bie
nen
/Vo
lk
days post treatment Tage nach Behandlung
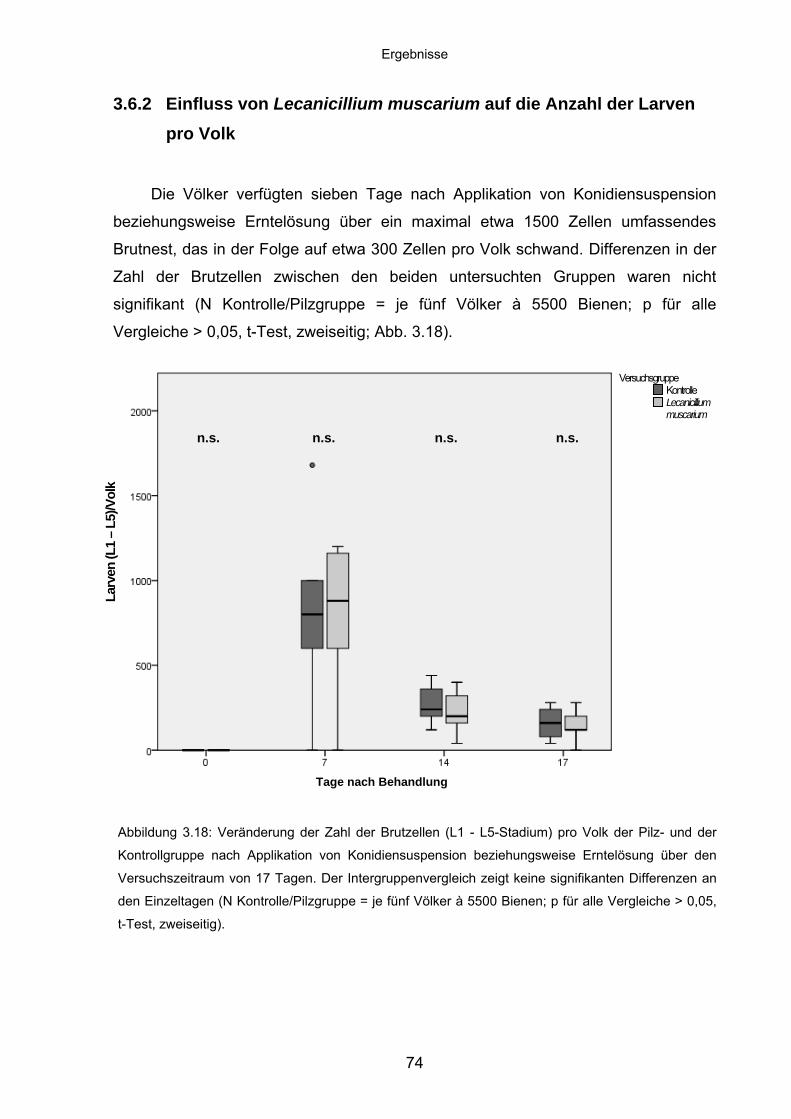
Ergebnisse
74
3.6.2 Einfluss von Lecanicillium muscarium auf die Anzahl der Larven
pro Volk
Die Völker verfügten sieben Tage nach Applikation von Konidiensuspension
beziehungsweise Erntelösung über ein maximal etwa 1500 Zellen umfassendes
Brutnest, das in der Folge auf etwa 300 Zellen pro Volk schwand. Differenzen in der
Zahl der Brutzellen zwischen den beiden untersuchten Gruppen waren nicht
signifikant (N Kontrolle/Pilzgruppe = je fünf Völker à 5500 Bienen; p für alle
Vergleiche > 0,05, t-Test, zweiseitig; Abb. 3.18).
Abbildung 3.18: Veränderung der Zahl der Brutzellen (L1 - L5-Stadium) pro Volk der Pilz- und der
Kontrollgruppe nach Applikation von Konidiensuspension beziehungsweise Erntelösung über den
Versuchszeitraum von 17 Tagen. Der Intergruppenvergleich zeigt keine signifikanten Differenzen an
den Einzeltagen (N Kontrolle/Pilzgruppe = je fünf Völker à 5500 Bienen; p für alle Vergleiche > 0,05,
t-Test, zweiseitig).
n.s. n.s. n.s. n.s.
Versuchsgruppe Kontrolle Lecanicillium muscarium
Lar
ven
(L1
– L5
)/Vo
lk
days post treatment Tage nach Behandlung
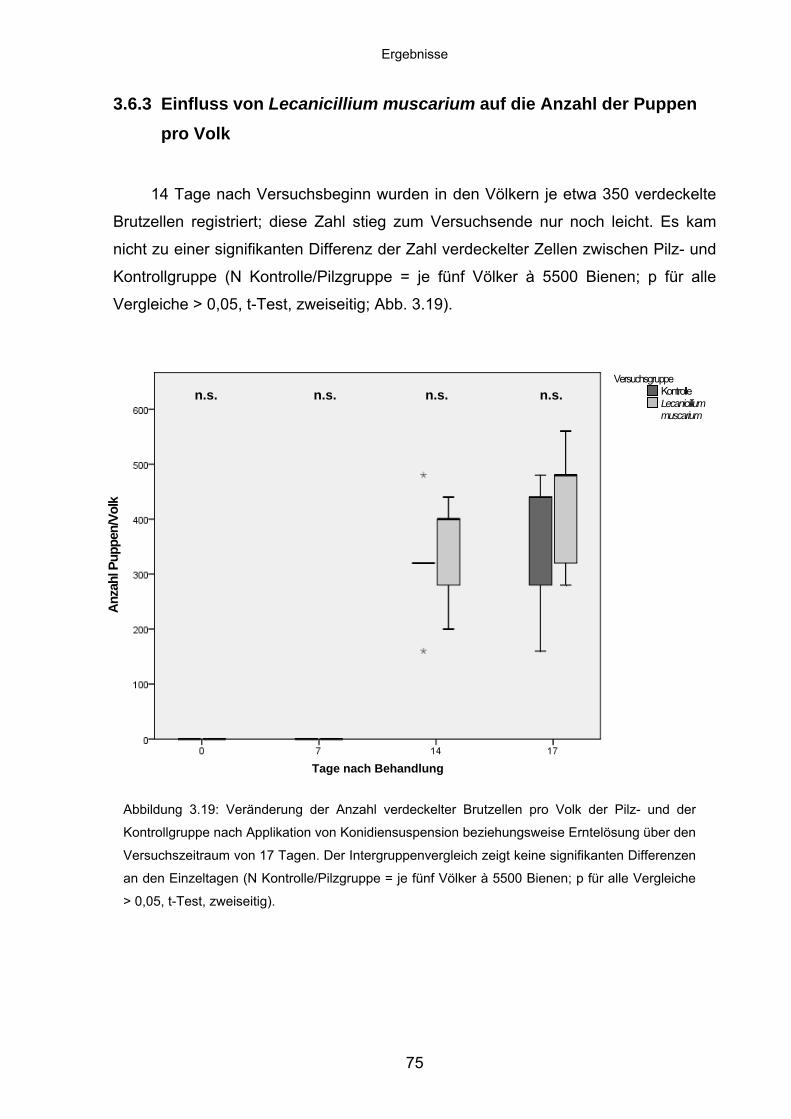
Ergebnisse
75
3.6.3 Einfluss von Lecanicillium muscarium auf die Anzahl der Puppen
pro Volk
14 Tage nach Versuchsbeginn wurden in den Völkern je etwa 350 verdeckelte
Brutzellen registriert; diese Zahl stieg zum Versuchsende nur noch leicht. Es kam
nicht zu einer signifikanten Differenz der Zahl verdeckelter Zellen zwischen Pilz- und
Kontrollgruppe (N Kontrolle/Pilzgruppe = je fünf Völker à 5500 Bienen; p für alle
Vergleiche > 0,05, t-Test, zweiseitig; Abb. 3.19).
Abbildung 3.19: Veränderung der Anzahl verdeckelter Brutzellen pro Volk der Pilz- und der
Kontrollgruppe nach Applikation von Konidiensuspension beziehungsweise Erntelösung über den
Versuchszeitraum von 17 Tagen. Der Intergruppenvergleich zeigt keine signifikanten Differenzen
an den Einzeltagen (N Kontrolle/Pilzgruppe = je fünf Völker à 5500 Bienen; p für alle Vergleiche
> 0,05, t-Test, zweiseitig).
n.s. n.s. n.s. n.s. Versuchsgruppe
Kontrolle Lecanicillium muscarium
Anz
ahl P
upp
en/V
olk
days post treatment Tage nach Behandlung
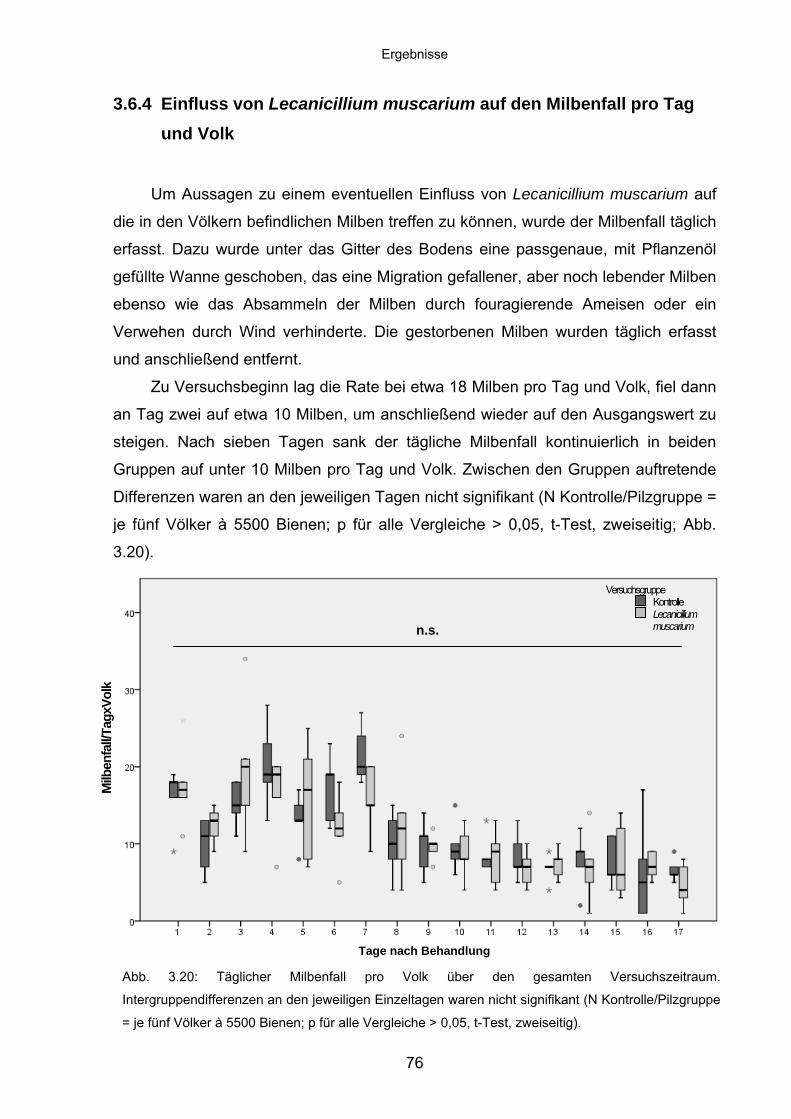
Ergebnisse
76
3.6.4 Einfluss von Lecanicillium muscarium auf den Milbenfall pro Tag
und Volk
Um Aussagen zu einem eventuellen Einfluss von Lecanicillium muscarium auf
die in den Völkern befindlichen Milben treffen zu können, wurde der Milbenfall täglich
erfasst. Dazu wurde unter das Gitter des Bodens eine passgenaue, mit Pflanzenöl
gefüllte Wanne geschoben, das eine Migration gefallener, aber noch lebender Milben
ebenso wie das Absammeln der Milben durch fouragierende Ameisen oder ein
Verwehen durch Wind verhinderte. Die gestorbenen Milben wurden täglich erfasst
und anschließend entfernt.
Zu Versuchsbeginn lag die Rate bei etwa 18 Milben pro Tag und Volk, fiel dann
an Tag zwei auf etwa 10 Milben, um anschließend wieder auf den Ausgangswert zu
steigen. Nach sieben Tagen sank der tägliche Milbenfall kontinuierlich in beiden
Gruppen auf unter 10 Milben pro Tag und Volk. Zwischen den Gruppen auftretende
Differenzen waren an den jeweiligen Tagen nicht signifikant (N Kontrolle/Pilzgruppe =
je fünf Völker à 5500 Bienen; p für alle Vergleiche > 0,05, t-Test, zweiseitig; Abb.
3.20).
Abb. 3.20: Täglicher Milbenfall pro Volk über den gesamten Versuchszeitraum.
Intergruppendifferenzen an den jeweiligen Einzeltagen waren nicht signifikant (N Kontrolle/Pilzgruppe
= je fünf Völker à 5500 Bienen; p für alle Vergleiche > 0,05, t-Test, zweiseitig).
n.s.
Versuchsgruppe Kontrolle Lecanicillium muscarium
Milb
enfa
ll/T
agxV
olk
days post treatment Tage nach Behandlung
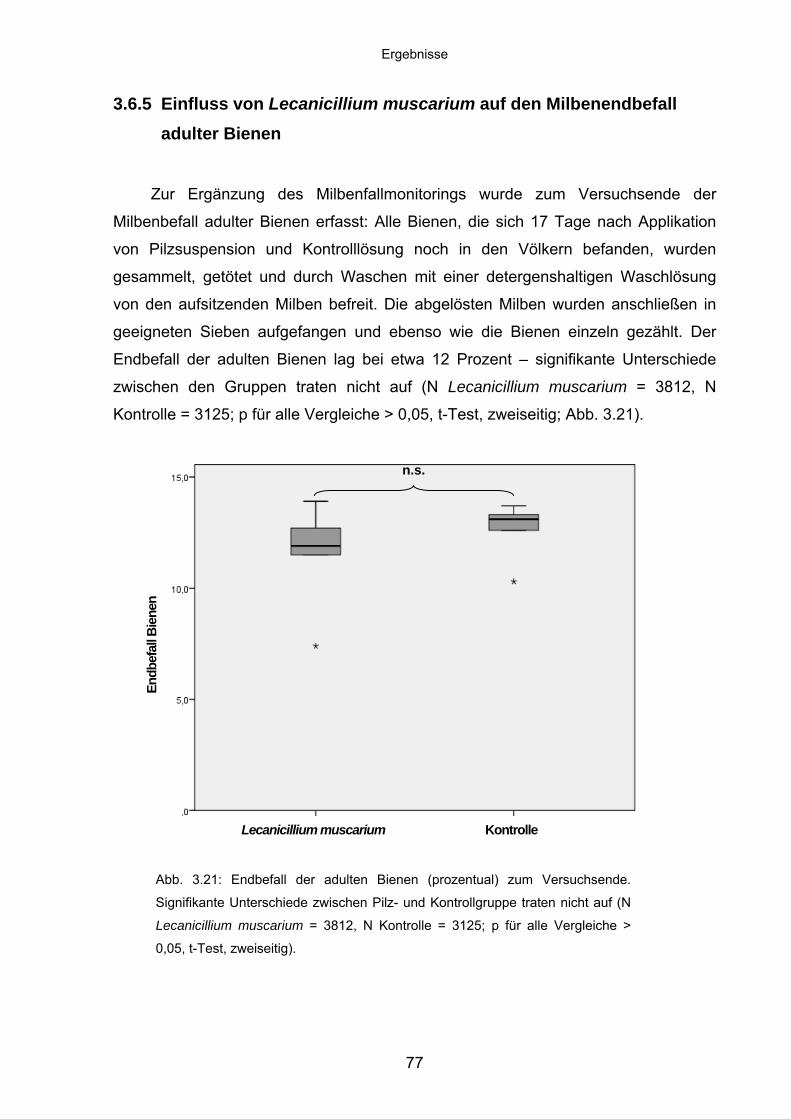
Ergebnisse
77
3.6.5 Einfluss von Lecanicillium muscarium auf den Milbenendbefall
adulter Bienen
Zur Ergänzung des Milbenfallmonitorings wurde zum Versuchsende der
Milbenbefall adulter Bienen erfasst: Alle Bienen, die sich 17 Tage nach Applikation
von Pilzsuspension und Kontrolllösung noch in den Völkern befanden, wurden
gesammelt, getötet und durch Waschen mit einer detergenshaltigen Waschlösung
von den aufsitzenden Milben befreit. Die abgelösten Milben wurden anschließen in
geeigneten Sieben aufgefangen und ebenso wie die Bienen einzeln gezählt. Der
Endbefall der adulten Bienen lag bei etwa 12 Prozent – signifikante Unterschiede
zwischen den Gruppen traten nicht auf (N Lecanicillium muscarium = 3812, N
Kontrolle = 3125; p für alle Vergleiche > 0,05, t-Test, zweiseitig; Abb. 3.21).
Abb. 3.21: Endbefall der adulten Bienen (prozentual) zum Versuchsende.
Signifikante Unterschiede zwischen Pilz- und Kontrollgruppe traten nicht auf (N
Lecanicillium muscarium = 3812, N Kontrolle = 3125; p für alle Vergleiche >
0,05, t-Test, zweiseitig).
n.s.
Lecanicillium muscarium Kontrolle
En
db
efal
l Bie
nen
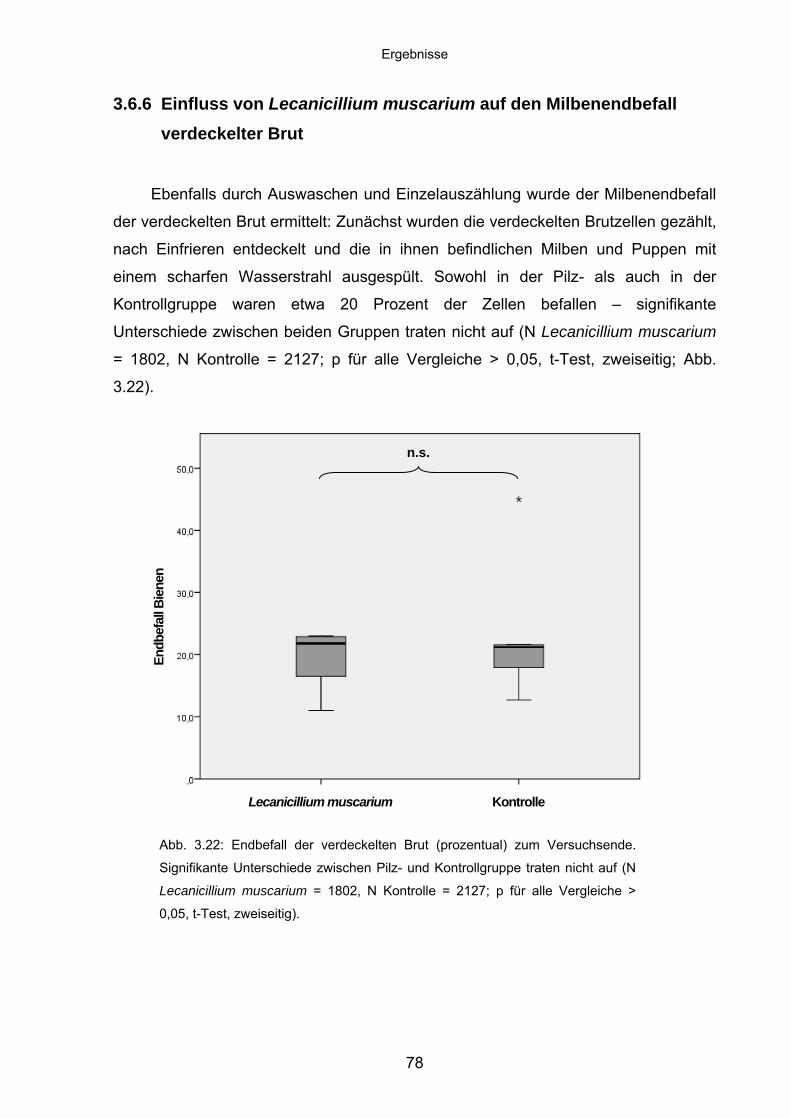
Ergebnisse
78
3.6.6 Einfluss von Lecanicillium muscarium auf den Milbenendbefall
verdeckelter Brut
Ebenfalls durch Auswaschen und Einzelauszählung wurde der Milbenendbefall
der verdeckelten Brut ermittelt: Zunächst wurden die verdeckelten Brutzellen gezählt,
nach Einfrieren entdeckelt und die in ihnen befindlichen Milben und Puppen mit
einem scharfen Wasserstrahl ausgespült. Sowohl in der Pilz- als auch in der
Kontrollgruppe waren etwa 20 Prozent der Zellen befallen – signifikante
Unterschiede zwischen beiden Gruppen traten nicht auf (N Lecanicillium muscarium
= 1802, N Kontrolle = 2127; p für alle Vergleiche > 0,05, t-Test, zweiseitig; Abb.
3.22).
Abb. 3.22: Endbefall der verdeckelten Brut (prozentual) zum Versuchsende.
Signifikante Unterschiede zwischen Pilz- und Kontrollgruppe traten nicht auf (N
Lecanicillium muscarium = 1802, N Kontrolle = 2127; p für alle Vergleiche >
0,05, t-Test, zweiseitig).
Lecanicillium muscarium Kontrolle
En
db
efal
l Bie
nen
n.s.

Ergebnisse
79
3.7 Subletale Effekte entomopathogener Pilze auf Adultbienen
Die in vitro-Versuche konnten erfassen, ob es zu einer erhöhten Mortalitätsrate
kam oder nicht – sie lieferten aber keine Erklärung, warum es zu einer solchen
Erhöhung kam und ob es eventuell weitere, subletale Effekte gab, die für eine
potentielle Anwendung von Bedeutung sind.
Um Hinweise zu erhalten, ob entomopathogene Pilze subletale Auswirkungen
auf Honigbienen haben könnten, wurden mit Pilzsporen beziehungsweise nur mit
einer Kontrolllösung behandelte Gruppen in Bezug auf ihre Futteraufnahme pro
Zeiteinheit als mögliches Indiz zum Beispiel einer Schädigung des Darmkanals,
sowie in Bezug auf ihr Lernvermögen als mögliches Indiz für eine Beeinträchtigung
wichtiger sozialer und räumlicher Orientierungskomponenten untersucht. Im
Folgenden werden die jeweiligen Ansätze kurz erläutert, bevor die Ergebnisse
beschrieben werden.
3.7.1 Ernährungszustand im Langzeit-in-vitro-Assay
In klassischen PER-Dressuren werden die Bienen den Völkern wenige Stunden
vor Versuchsbeginn entnommen – naturgemäß ist das in einem Versuch, der über
einen Zeitraum von 12 Tagen potentielle subletale Langzeiteffekte einer Behandlung
mit entomopathogenen Pilzen aufdecken soll, nicht möglich. Das sich daraus
ergebende Problem besteht in einem im Vergleich zur dem Volk direkt entnommenen
Biene a priori nicht kalkulierbaren Sättigungszustand der in vitro ad libitum mit Futter
versorgten Bienen, da Langzeit-PER-Versuche mit gekäfigten Bienen nie zuvor
etabliert worden sind. Das Hungern der Einzelbiene ist aber Voraussetzung für ein
Gelingen des Versuchs. In Vorserien hatte sich gezeigt, dass es starke
Schwankungen bezüglich der nötigen Dauer der Hungerphase gab: Beobachtet
wurden Hungerphasen von mehr als 12 Stunden, die nötig waren, um Bienen zur
Versuchsteilnahme zu bewegen, aber auch eine Mortalitätsrate von über 50 Prozent
nach einer Hungerphase von nur einer halben Stunde. Zunächst wurde deshalb
untersucht, ob über den Versuchszeitraum Veränderungen im Honigblasengewicht
und damit im Ernährungszustand auftraten, die eine Kalkulation der nötigen Dauer
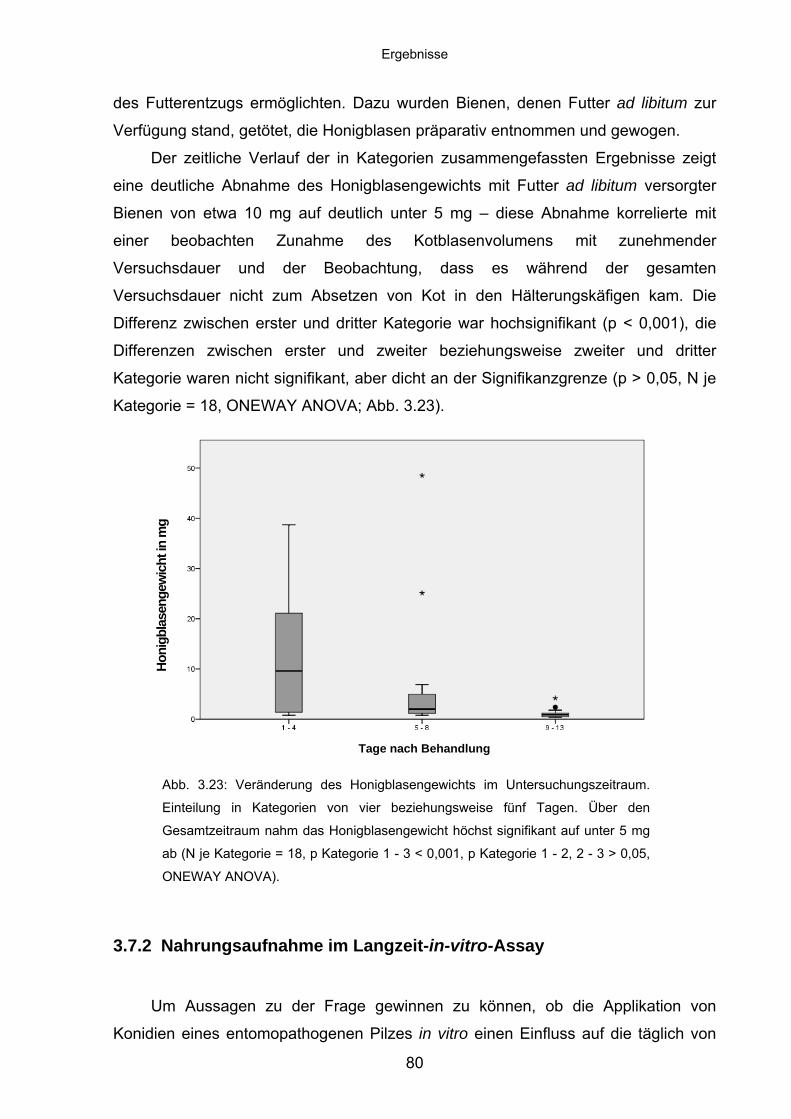
Ergebnisse
80
des Futterentzugs ermöglichten. Dazu wurden Bienen, denen Futter ad libitum zur
Verfügung stand, getötet, die Honigblasen präparativ entnommen und gewogen.
Der zeitliche Verlauf der in Kategorien zusammengefassten Ergebnisse zeigt
eine deutliche Abnahme des Honigblasengewichts mit Futter ad libitum versorgter
Bienen von etwa 10 mg auf deutlich unter 5 mg – diese Abnahme korrelierte mit
einer beobachten Zunahme des Kotblasenvolumens mit zunehmender
Versuchsdauer und der Beobachtung, dass es während der gesamten
Versuchsdauer nicht zum Absetzen von Kot in den Hälterungskäfigen kam. Die
Differenz zwischen erster und dritter Kategorie war hochsignifikant (p < 0,001), die
Differenzen zwischen erster und zweiter beziehungsweise zweiter und dritter
Kategorie waren nicht signifikant, aber dicht an der Signifikanzgrenze (p > 0,05, N je
Kategorie = 18, ONEWAY ANOVA; Abb. 3.23).
3.7.2 Nahrungsaufnahme im Langzeit-in-vitro-Assay
Um Aussagen zu der Frage gewinnen zu können, ob die Applikation von
Konidien eines entomopathogenen Pilzes in vitro einen Einfluss auf die täglich von
Abb. 3.23: Veränderung des Honigblasengewichts im Untersuchungszeitraum.
Einteilung in Kategorien von vier beziehungsweise fünf Tagen. Über den
Gesamtzeitraum nahm das Honigblasengewicht höchst signifikant auf unter 5 mg
ab (N je Kategorie = 18, p Kategorie 1 - 3 < 0,001, p Kategorie 1 - 2, 2 - 3 > 0,05,
ONEWAY ANOVA).
b ab a
days post treatment
Ho
nig
bla
sen
gew
ich
t in
mg
Tage nach Behandlung
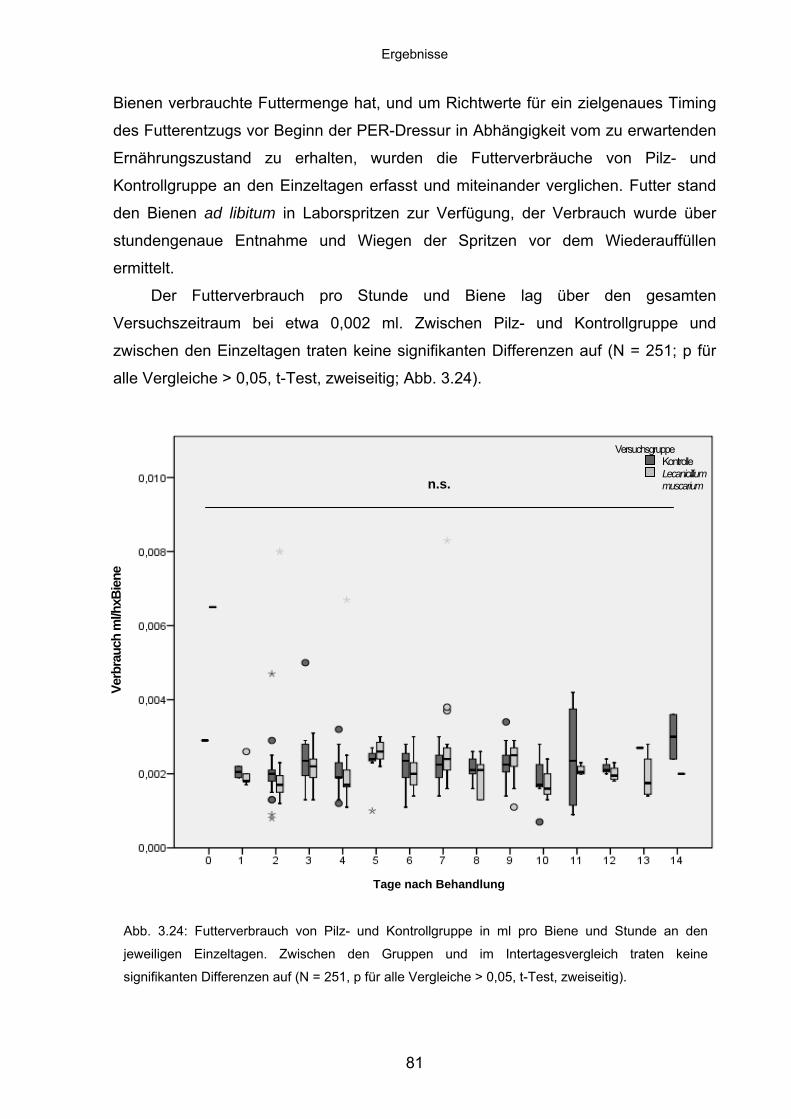
Ergebnisse
81
Bienen verbrauchte Futtermenge hat, und um Richtwerte für ein zielgenaues Timing
des Futterentzugs vor Beginn der PER-Dressur in Abhängigkeit vom zu erwartenden
Ernährungszustand zu erhalten, wurden die Futterverbräuche von Pilz- und
Kontrollgruppe an den Einzeltagen erfasst und miteinander verglichen. Futter stand
den Bienen ad libitum in Laborspritzen zur Verfügung, der Verbrauch wurde über
stundengenaue Entnahme und Wiegen der Spritzen vor dem Wiederauffüllen
ermittelt.
Der Futterverbrauch pro Stunde und Biene lag über den gesamten
Versuchszeitraum bei etwa 0,002 ml. Zwischen Pilz- und Kontrollgruppe und
zwischen den Einzeltagen traten keine signifikanten Differenzen auf (N = 251; p für
alle Vergleiche > 0,05, t-Test, zweiseitig; Abb. 3.24).
Abb. 3.24: Futterverbrauch von Pilz- und Kontrollgruppe in ml pro Biene und Stunde an den
jeweiligen Einzeltagen. Zwischen den Gruppen und im Intertagesvergleich traten keine
signifikanten Differenzen auf (N = 251, p für alle Vergleiche > 0,05, t-Test, zweiseitig).
days post treatment
Ver
bra
uch
ml/h
xBie
ne
n.s.
Versuchsgruppe Kontrolle Lecanicillium muscarium
Tage nach Behandlung

Ergebnisse
82
3.7.3 Lernleistung von Adultbienen im PER-Assay
Zur Analyse potentieller Effekte eines entomopathogenen Pilzes auf das
Lernvermögen von Honigbienen wurde ein Langzeit-PER-Assay verwendet. Das
Lernen von Honigbienen im PER-Assay folgt dabei den Regeln von klassischer und
instrumenteller Konditionierung. Die Versuchsbienen wurden mit einem Duft als
unkonditioniertem Reiz konfrontiert, dem ein Kontaktieren von 2M Zuckerlösung mit
den Antennen und anschließendes Trinkenlassen folgte. Während der
Konditionierungsphase wurde dann der ursprünglich neutrale Duftreiz mit dem
unbedingten Reiz assoziiert, so dass bei erfolgreichem Lernen auf die Duftgabe ein
Herausstrecken des Rüssels folgte.
Über die insgesamt vier Lernakte des Tests kam es zu einer linearen Zunahme
der positiven Antworten (in Prozent der Gesamtantworten) von null im ersten Lernakt
auf etwa 50 Prozent im vierten Lernakt beider Gruppen. Die Ergebnisse der
Einzelakte differierten signifikant bis höchst signifikant voneinander, während
zwischen den Versuchsgruppen keine signifikanten Unterschiede auftraten. (N
Kontrolle = 107, N Pilz = 110; p für alle Vergleiche innerhalb eines Lernaktes > 0,05,
p b - c < 0.05, p c - d < 0,01, p b - d < 0,001, p a - (b/c/d) < 0,001, χ2-Test, zweiseitig;
Abb. 3.25).
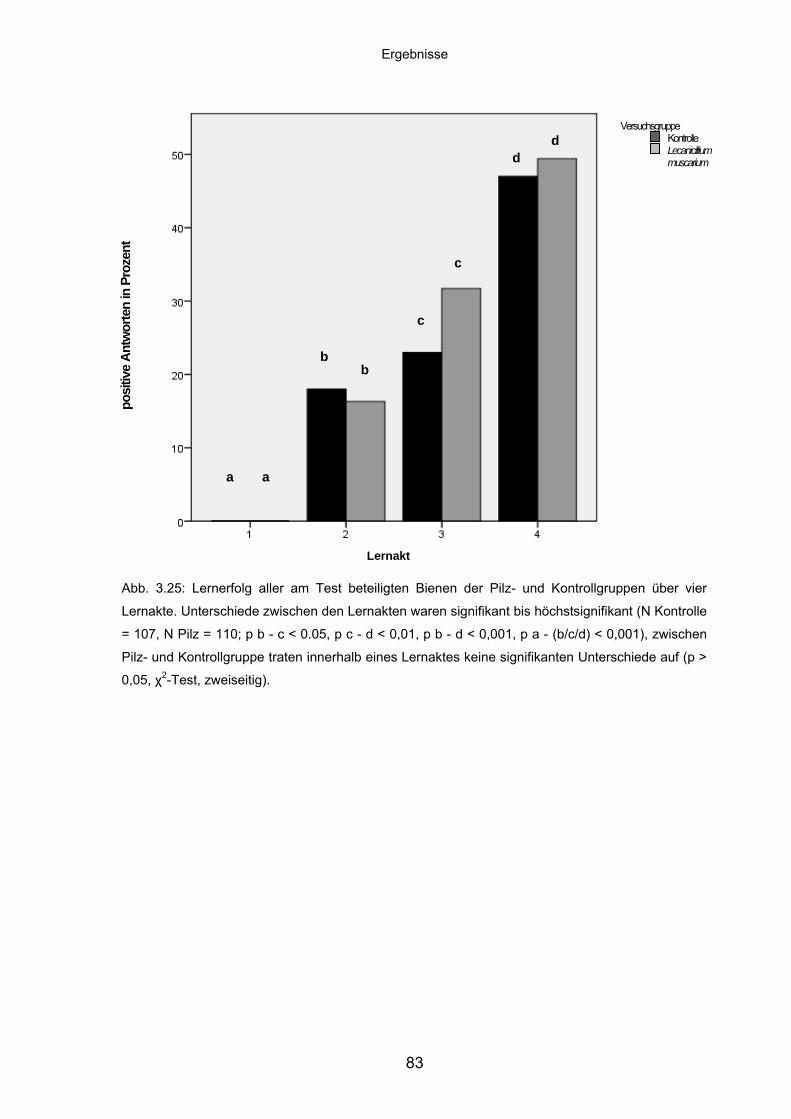
Ergebnisse
83
Abb. 3.25: Lernerfolg aller am Test beteiligten Bienen der Pilz- und Kontrollgruppen über vier
Lernakte. Unterschiede zwischen den Lernakten waren signifikant bis höchstsignifikant (N Kontrolle
= 107, N Pilz = 110; p b - c < 0.05, p c - d < 0,01, p b - d < 0,001, p a - (b/c/d) < 0,001), zwischen
Pilz- und Kontrollgruppe traten innerhalb eines Lernaktes keine signifikanten Unterschiede auf (p >
0,05, χ2-Test, zweiseitig).
Versuchsgruppe Kontrolle Lecanicillium muscarium
Lernakt
pos
itive
Ant
wo
rten
in P
roze
nt
a a
b b
c
c
d d
Lernakt

Ergebnisse
84
3.7.4 Diskriminationsfähigkeit von Adultbienen im Langzeit-PER-Assay
In der Langzeit-PER-Dressur erfolgte die Untersuchung eines möglichen
Einflusses von Lecanicillium muscarium auf das Lernvermögen adulter Honigbienen
über insgesamt 20 Tests je Einzelbiene: Je achtmal wurde die Reaktion auf den
konditionierten Reiz (Geraniol) im Wechsel mit einem Vergleichsduft (Citral),
unterbrochen von zwei Kontrolldurchgängen nach den Durchgängen zwei und fünf, in
denen eine potentielle Reaktion auf zugeströmte Luft ohne Test- oder
Vergleichssubstanz getestet wurde, analysiert. Verglichen wurden Bienen der Pilz-
und der Kontrollgruppe, getestet wurde über einen Zeitraum von 12 Tagen in drei
Wiederholungen. Bienen, die keine Reaktionen mehr zeigten oder auf Luft
reagierten, wurden ausgeschlossen.
Im Folgenden sind – als Maßstab der Diskriminationsfähigkeit – die
prozentualen Antwortdifferenzen von Pilz- und Kontrollgruppe einander
gegenübergestellt. Unter prozentualer Antwortdifferenz wird die Differenz zwischen
gegebenen positiven Antworten auf die Testsubstanz in Prozent und gegebenen
positiven Antworten auf die Vergleichssubstanz in Prozent verstanden.
Die Antwortdifferenzen schwankten in der Pilzgruppe zwischen 16,7 Prozent
und 37,5 Prozent, in der Kontrollgruppe zwischen 25,0 Prozent und 39,2 Prozent.
Von einem Versuchsintervall abgesehen (Tage acht und neun nach Applikation von
Lecanicillium muscarium beziehungsweise Kontrolllösung) lag die Antwortdifferenz
der Kontrollgruppe immer über derjenigen der Pilzgruppe. In den Intervallen vier,
sechs, acht, zehn und zwölf waren die Intergruppendifferenzen nicht signifikant
voneinander verschieden (p > 0,05, χ2-Test, zweiseitig), im Intervall eins allerdings
kam es zu einem hoch signifikanten Unterschied zwischen den beiden Gruppen, i.e.,
unter dem Einfluss von Lecanicillium muscarium lernten Honigbienen in der PER-
Dressur signifikant schlechter, zwischen Test- und Vergleichssubstanz zu
unterscheiden (N Kontrolle = 107, N Pilz = 110; p Intervalle 4 - 12 > 0,05, p Intervall 2
< 0,01, χ2-Test, zweiseitig; Abb. 3.26).
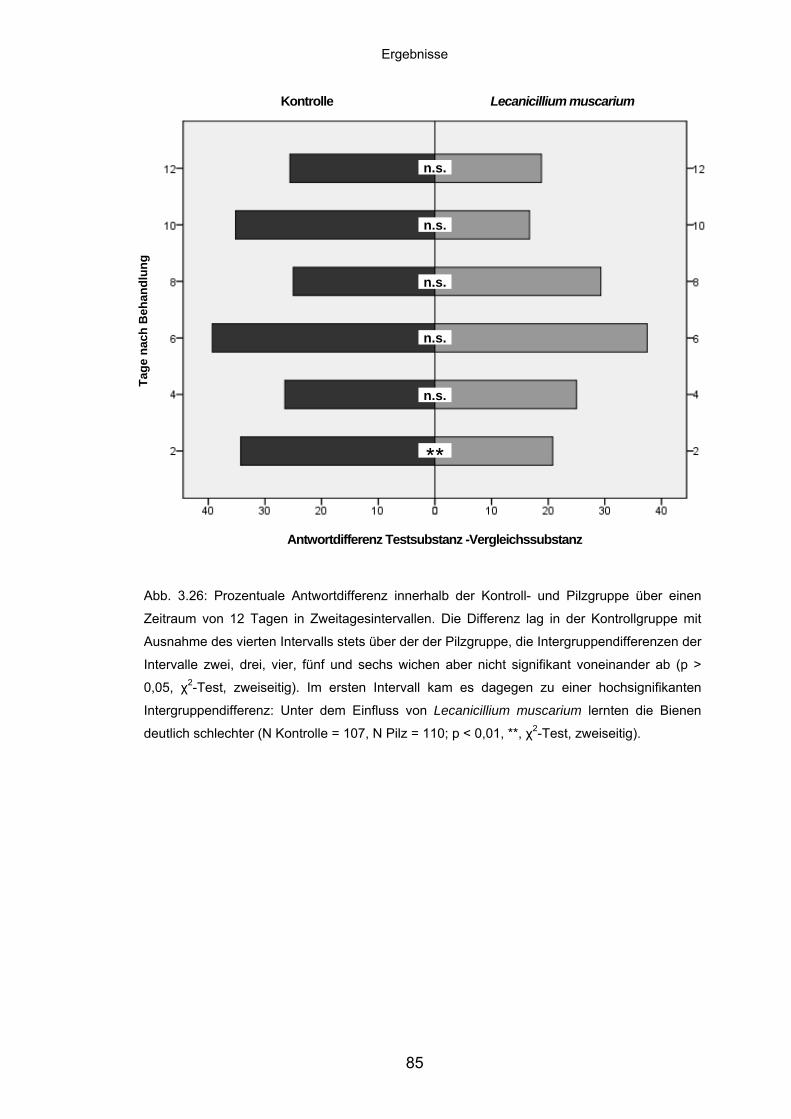
Ergebnisse
85
Abb. 3.26: Prozentuale Antwortdifferenz innerhalb der Kontroll- und Pilzgruppe über einen
Zeitraum von 12 Tagen in Zweitagesintervallen. Die Differenz lag in der Kontrollgruppe mit
Ausnahme des vierten Intervalls stets über der der Pilzgruppe, die Intergruppendifferenzen der
Intervalle zwei, drei, vier, fünf und sechs wichen aber nicht signifikant voneinander ab (p >
0,05, χ2-Test, zweiseitig). Im ersten Intervall kam es dagegen zu einer hochsignifikanten
Intergruppendifferenz: Unter dem Einfluss von Lecanicillium muscarium lernten die Bienen
deutlich schlechter (N Kontrolle = 107, N Pilz = 110; p < 0,01, **, χ2-Test, zweiseitig).
day
s p
ost
trea
tmen
t
Antwortdifferenz Testsubstanz -Vergleichssubstanz
Kontrolle Lecanicillium muscarium
**
n.s.
n.s.
n.s.
n.s.
n.s.
Tag
e n
ach
Beh
and
lun
g

Ergebnisse
86
3.7.5 Einfluss entomopathogener Pilze auf das Immunsystem von
Adultbienen
Für entomopathogene Pilze kann angenommen werden, dass sie unter
Umständen auch Nichtzielorganismen in einer Weise schädigen, die über den
Nachweis der Induktion einer Immunantwort belegbar ist. Eine solche Immunantwort
wiederum könnte zum Beispiel mit einem reduzierten Kognitionsvermögen
korrelieren. Um Hinweise auf diesen denkbaren Zusammenhang zu erhalten, wurden
immunisierte beziehungsweise Kontrollbienen einem sensitiven Immunassay
unterzogen, der Aufschlüsse über eine potentielle Immunantwort geben konnte. Der
Nachweis einer Aktivierung der IMD- oder Toll-Kaskade erfolgt dabei über eine
Suppression des Wachstums mit Hämolymphe behandelter Bienen inkubierter
Bakterien (Müller et al. 2008). Es wurden Versuchsgruppen gebildet, die entweder
mit Konidien von Lecanicillium muscarium, Metarhizium anisopliae, Beauveria
bassiana, Paecilomyces fumosoroseus, beziehungsweise als Kontrolle mit Sporen
von Saccharomyces cerevisiae oder konidienfreier Erntelösung besprüht wurden.
Den Bienen einer weiteren Gruppe wurde zu Versuchsbeginn jeweils ein Bein
amputiert, um eine obligate Immunantwort zu initiieren. 24 Stunden später wurde den
Bienen nach Betäubung Hämolymphe entnommen, die mit je einer Grampositiven
und einer Gramnegativen Bakterienkultur in der log-Phase in einer
Verdünnungsreihe über sieben Potenzen inkubiert und dann auf Nährmedium
ausplattiert wurde. Nach weiteren 24 Stunden wurde die Zahl kolonieformender
Einheiten (colony forming units, cfu) als Maßstab für eine eventuell induzierte
Immunantwort erfasst.
Die Ergebnisse der Gruppe amputierter Bienen und der Bakterienkulturen, die –
ohne Hämolymphzusatz – nur mit einer Aprotinin-Phenylthioharnstoff-Lösung
beziehungsweise Streptomycinlösung versetzt wurden, validierten die Methodik der
Untersuchung.
Im vergleichenden Versuch mit dem Grampositiven Bakterium Bacillus subtilis
zeigte sich eine gegenüber Saccharomyces cerevisiae und der Erntelösung um
nahezu 90 Prozent verringerte Zahl kolonieformender Einheiten im Lecanicillium
muscarium-, Metarhizium anisopliae-, und Beauveria bassiana-Ansatz. Im
Paecilomyces fumosoroseus-Ansatz kam es zu keiner Reduktion (Abb. 3.27).
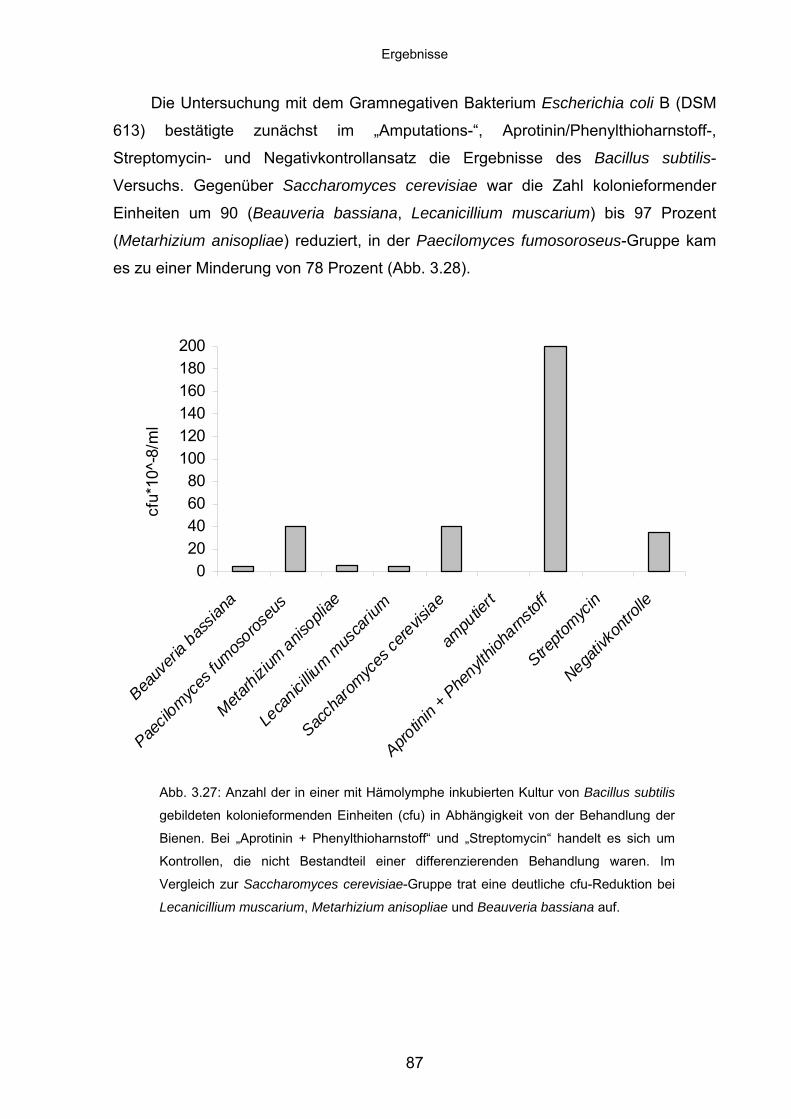
Ergebnisse
87
020406080
100120140160180200
Beauv
eria b
assia
na
Paecil
omyc
es fu
mos
orose
us
Met
arhizi
um a
nisopli
ae
Leca
nicilliu
m mus
cariu
m
Sacch
arom
yces
cere
visia
e
ampu
tiert
Aprotin
in + P
henylt
hioha
rnsto
ff
Strept
omyc
in
Negativ
kont
rolle
cfu*
10^-
8/m
l
Die Untersuchung mit dem Gramnegativen Bakterium Escherichia coli B (DSM
613) bestätigte zunächst im „Amputations-“, Aprotinin/Phenylthioharnstoff-,
Streptomycin- und Negativkontrollansatz die Ergebnisse des Bacillus subtilis-
Versuchs. Gegenüber Saccharomyces cerevisiae war die Zahl kolonieformender
Einheiten um 90 (Beauveria bassiana, Lecanicillium muscarium) bis 97 Prozent
(Metarhizium anisopliae) reduziert, in der Paecilomyces fumosoroseus-Gruppe kam
es zu einer Minderung von 78 Prozent (Abb. 3.28).
Abb. 3.27: Anzahl der in einer mit Hämolymphe inkubierten Kultur von Bacillus subtilis
gebildeten kolonieformenden Einheiten (cfu) in Abhängigkeit von der Behandlung der
Bienen. Bei „Aprotinin + Phenylthioharnstoff“ und „Streptomycin“ handelt es sich um
Kontrollen, die nicht Bestandteil einer differenzierenden Behandlung waren. Im
Vergleich zur Saccharomyces cerevisiae-Gruppe trat eine deutliche cfu-Reduktion bei
Lecanicillium muscarium, Metarhizium anisopliae und Beauveria bassiana auf.
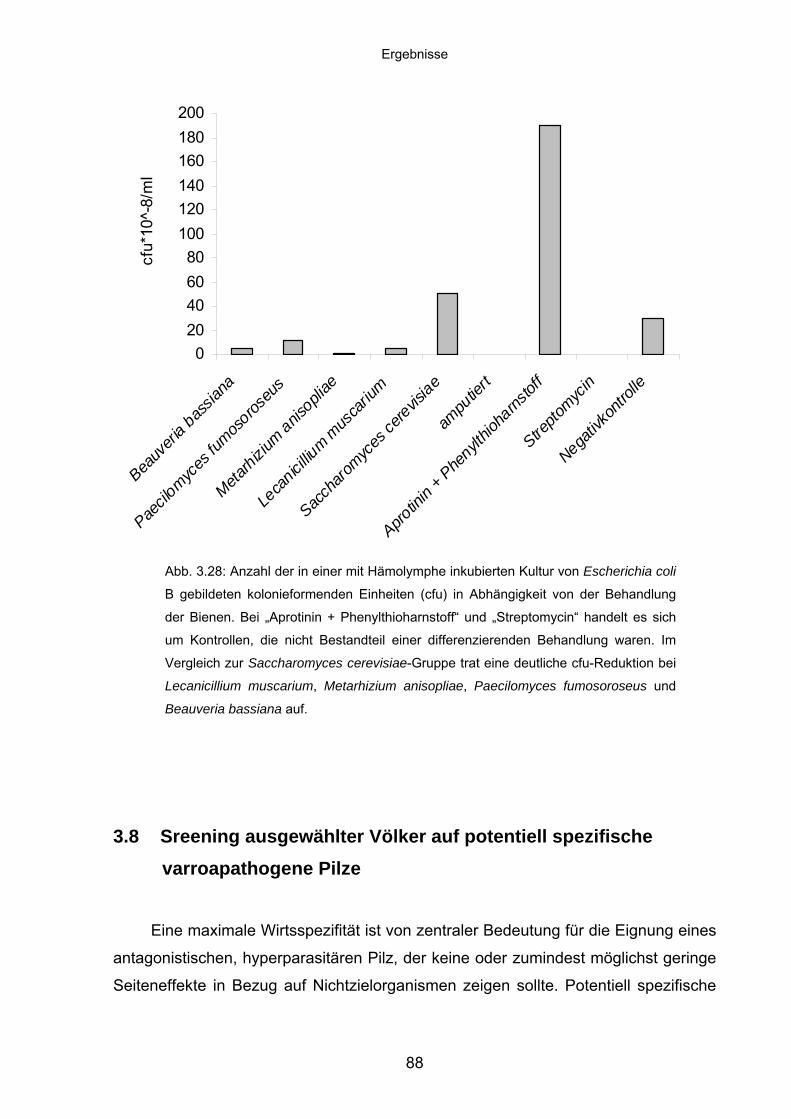
Ergebnisse
88
3.8 Sreening ausgewählter Völker auf potentiell spezifische
varroapathogene Pilze
Eine maximale Wirtsspezifität ist von zentraler Bedeutung für die Eignung eines
antagonistischen, hyperparasitären Pilz, der keine oder zumindest möglichst geringe
Seiteneffekte in Bezug auf Nichtzielorganismen zeigen sollte. Potentiell spezifische
Abb. 3.28: Anzahl der in einer mit Hämolymphe inkubierten Kultur von Escherichia coli
B gebildeten kolonieformenden Einheiten (cfu) in Abhängigkeit von der Behandlung
der Bienen. Bei „Aprotinin + Phenylthioharnstoff“ und „Streptomycin“ handelt es sich
um Kontrollen, die nicht Bestandteil einer differenzierenden Behandlung waren. Im
Vergleich zur Saccharomyces cerevisiae-Gruppe trat eine deutliche cfu-Reduktion bei
Lecanicillium muscarium, Metarhizium anisopliae, Paecilomyces fumosoroseus und
Beauveria bassiana auf.
0
20
40
60
80
100
120
140
160
180
200
Beauv
eria b
assia
na
Paecil
omyc
es fu
mos
orose
us
Met
arhizi
um a
nisopli
ae
Leca
nicilliu
m mus
cariu
m
Sacch
arom
yces
cere
visia
e
ampu
tiert
Aprotin
in + P
henylt
hioha
rnsto
ff
Strept
omyc
in
Negativ
kont
rolle
cfu*
10^-
8/m
l

Ergebnisse
89
Hyperparasiten zu finden, die möglicherweise als Antagonisten von Varroa destructor
geeignet sind, war deshalb eines der Hauptziele der vorliegenden Untersuchung.
Dazu wurden über insgesamt vier Jahre (2005 - 2008) sämtliche
Versuchsvölker einem Monitoring von Populationsdynamik und Varroabefall
unterzogen. Repräsentative Völker dieses Monitorings (i.e. Völker, die einen sehr
hohen, sehr niedrigen oder durchschnittlichen Milbenbefall aufwiesen) wurden
daraufhin ausgewählt und zusätzlich auf das aus gestorbenen Milben aus diesen
Völkern erfolgende Pilzwachstum untersucht, um mögliche Korrelationen zwischen
Milbenbefall und anteiligem Pilzwachstum zu finden, ohne zunächst die Art beteiligter
Pilze zu berücksichtigen.
Im Anschluss daran wurden im Screening gefundene Pilze isoliert, in Reinkultur
genommen, mikroskopisch typisiert und auf der Basis ihrer Sequenzdaten
identifiziert. Im Folgenden werden die jeweiligen Ansätze kurz erläutert, bevor die
Ergebnisse beschrieben werden.
3.8.1 Populationsdynamik und Milbenbefall von Apis mellifera-Völkern
Die Auswahl von Kandidatenvölkern erfolgte von 2005 - 2008 auf der Basis von
Populationsschätzungsdaten und der Erfassung des Milbenfalls als repräsentativen
Maßstabes für den Befall eines Volkes mit Milben. 2005 wurden 12, 2006 55, 2007
81 und 2008 67 Völker in die Untersuchung einbezogen. Die Bienenzahl wurde
während des jeweiligen Sommerhalbjahres in vierwöchigen Abständen insgesamt
fünfmal erfasst. Der Milbenfall wurde für jedes Volk während des
Untersuchungszeitraums an 21 Terminen mit einer untergeschobenen
Kunststoffwanne ermittelt. Die Abbildungen 3.29 und 3.30 zeigen exemplarisch den
Verlauf der Populationsdynamik und des Milbenfalls für sechs untersuchte Völker.
Trotz ähnlicher Entwicklungen in Bezug auf die Bienenzahl kam es bei ansonsten
gleichen Ausgangsbedingungen und gleicher Behandlung der Völker im
Jahresverlauf zu sehr unterschiedlichen Befallsgraden mit Varroa destructor, die vor
dem Hintergrund der Annahme einer Beteiligung pathogener Pilze an der Mortalität
von Milben als Hinweis darauf gedeutet wurden.
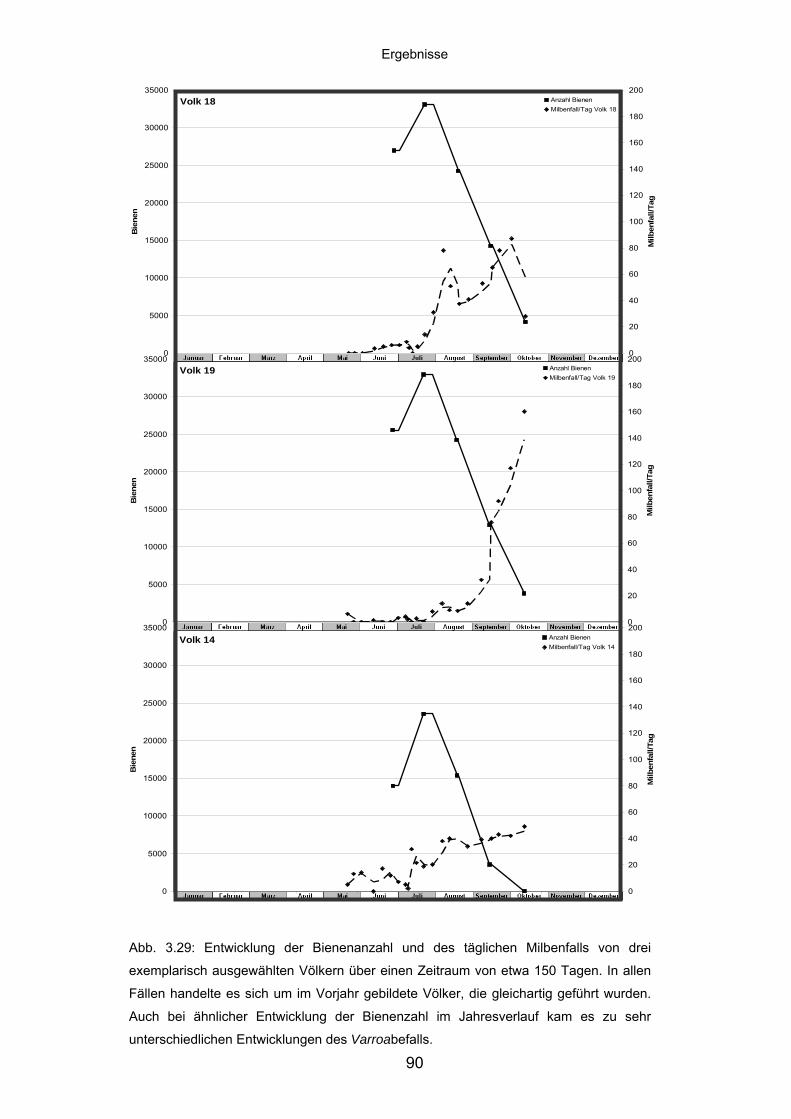
Ergebnisse
90
Volk 19
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milbenfa
ll/Tag
Anzahl Bienen
Milbenfall/Tag Volk 19
Volk 18
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milbenfa
ll/T
ag
Anzahl Bienen
Milbenfall/Tag Volk 18
Volk 14
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milbenfa
ll/T
ag
Anzahl Bienen
Milbenfall/Tag Volk 14
Abb. 3.29: Entwicklung der Bienenanzahl und des täglichen Milbenfalls von drei
exemplarisch ausgewählten Völkern über einen Zeitraum von etwa 150 Tagen. In allen
Fällen handelte es sich um im Vorjahr gebildete Völker, die gleichartig geführt wurden.
Auch bei ähnlicher Entwicklung der Bienenzahl im Jahresverlauf kam es zu sehr
unterschiedlichen Entwicklungen des Varroabefalls.
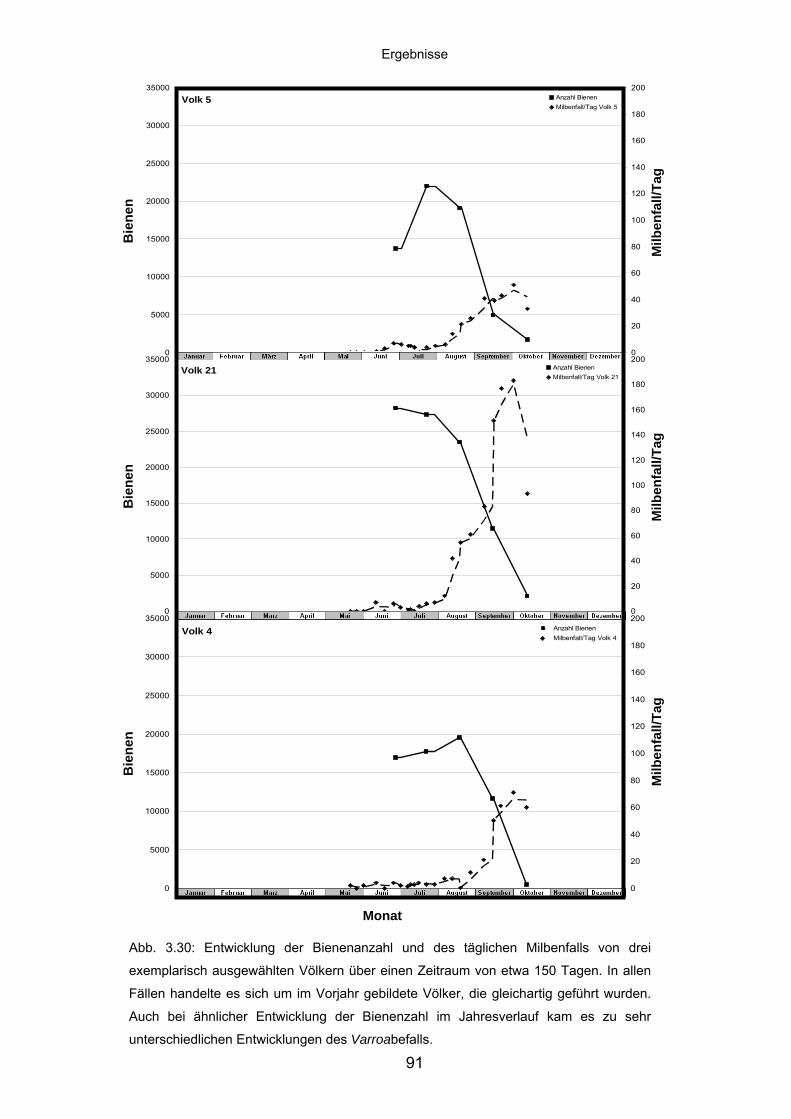
Ergebnisse
91
Abb. 3.30: Entwicklung der Bienenanzahl und des täglichen Milbenfalls von drei
exemplarisch ausgewählten Völkern über einen Zeitraum von etwa 150 Tagen. In allen
Fällen handelte es sich um im Vorjahr gebildete Völker, die gleichartig geführt wurden.
Auch bei ähnlicher Entwicklung der Bienenzahl im Jahresverlauf kam es zu sehr
unterschiedlichen Entwicklungen des Varroabefalls.
Volk 21
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milben
fall/T
ag
Anzahl Bienen
Milbenfall/Tag Volk 21
Volk 5
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milb
enfa
ll/Tag
Anzahl Bienen
Milbenfall/Tag Volk 5
Volk 4
0
5000
10000
15000
20000
25000
30000
35000
Bie
nen
0
20
40
60
80
100
120
140
160
180
200
Milb
enfa
ll/Tag
Anzahl Bienen
Milbenfall/Tag Volk 4
Milb
enfa
ll/T
ag
Milb
enfa
ll/T
ag
Milb
enfa
ll/T
ag
Bie
nen
B
ien
en
Bie
nen
Monat

Ergebnisse
92
3.8.2 Korrelation der Abundanz von Varroa destructor in Bienenvölkern
und des Pilzwachstums aus Milben
Auf der Basis des jährlichen Monitorings der Versuchsvölker wurden zur
Untersuchung einer eventuellen Korrelation von Milbenbefall und Vorkommen von
Pilzen Völker ausgewählt, deren Milbentotenfall in Bezug auf die Erzielbarkeit eines
Pilzwachstums in vitro untersucht wurde. Dazu wurden täglich über einen Zeitraum
von bis zu zwei Wochen pro Volk die in die Auffangwanne gefallenen Milben
gesammelt, oberflächensterilisiert und unter sterilen Bedingungen auf H2O-Agar
kultiviert. In der Folge wurde das aus diesen Milben erfolgte Pilzwachstum registriert.
Da der Einschub der Schubladen über bis zu zwei Wochen bedingte, dass es zur
Verschleppung von Milben durch fouragierende Ameisen kam, konnte nicht
gleichzeitig der Befall des Volkes über den Milbentotenfall in der Auffangwanne
erfasst werden. Allen Völkern wurden deshalb Proben von etwa 1000 adulten Bienen
entnommen, die exakt ausgezählt und deren aufsitzende Milben nach Abwaschen
mit einer Detergenslösung und Absieben ebenfalls erfasst wurden.
Im Folgenden sind die Ergebnisse für die Zusammenhänge zwischen
Milbenbefall pro Biene und anteiligem Pilzwachstum zunächst getrennt für die Jahre
2005 (Abb. 3.31, N = fünf Völker), 2006 (Abb. 3.32, N = 19 Völker), 2007 (Abb. 3.33,
N = 16 Völker) und 2008 (Abb. 3.34, N = 17 Völker), im Anschluss daran in einer
Übersicht (Abb. 3.35, N Gesamt = 57 Völker) dargestellt. Das Quadrat des
Korrelationskoeffizienten nach Pearson ist in der jeweiligen Grafik angegeben.
Für 2005, 2006 und 2008 zeigt sich ein Zusammenhang zwischen Milbenbefall
und Pilzwachstum – im Jahr 2007 gibt es einen solchen Zusammenhang nicht. Für
die Jahre 2005 und 2008 ist die Korrelation – für 2005 allerdings vor dem
Hintergrund eines geringen Stichprobenumfangs – mit mittlerer Güte belegt, in 2006
zeigte sich eine sehr starke Korrelation der beiden Variablen.
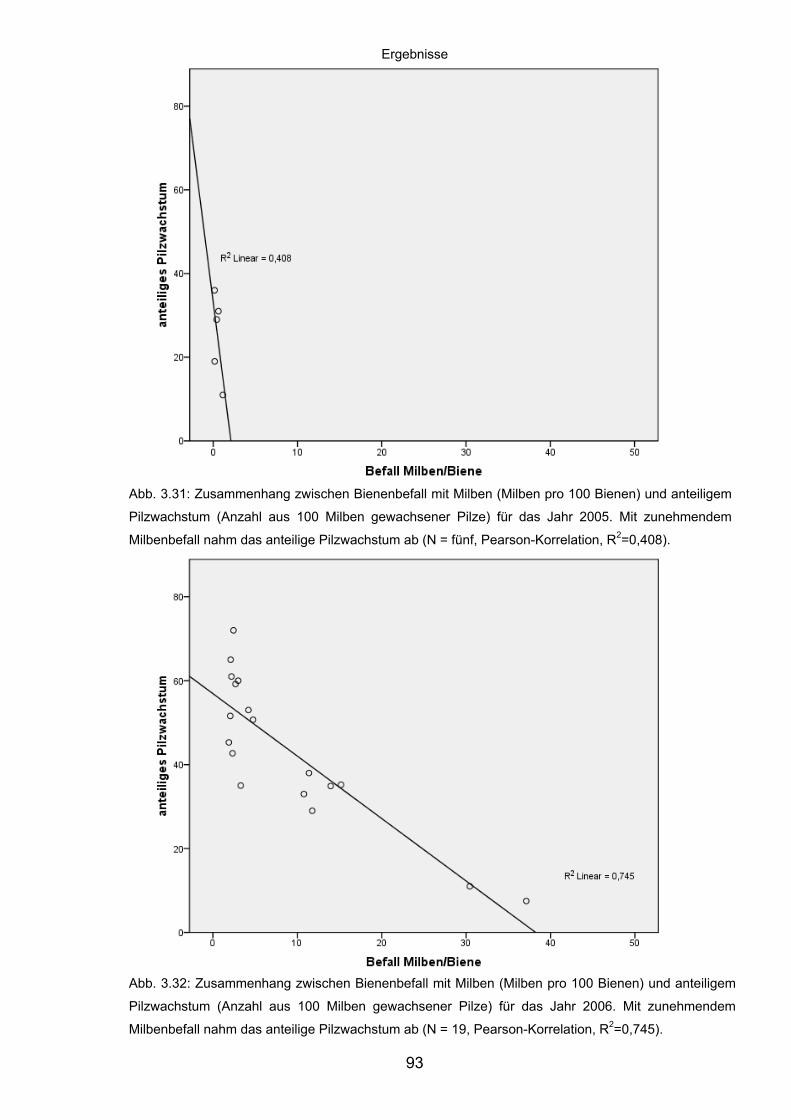
Ergebnisse
93
Abb. 3.32: Zusammenhang zwischen Bienenbefall mit Milben (Milben pro 100 Bienen) und anteiligem
Pilzwachstum (Anzahl aus 100 Milben gewachsener Pilze) für das Jahr 2006. Mit zunehmendem
Milbenbefall nahm das anteilige Pilzwachstum ab (N = 19, Pearson-Korrelation, R2=0,745).
Abb. 3.31: Zusammenhang zwischen Bienenbefall mit Milben (Milben pro 100 Bienen) und anteiligem
Pilzwachstum (Anzahl aus 100 Milben gewachsener Pilze) für das Jahr 2005. Mit zunehmendem
Milbenbefall nahm das anteilige Pilzwachstum ab (N = fünf, Pearson-Korrelation, R2=0,408).
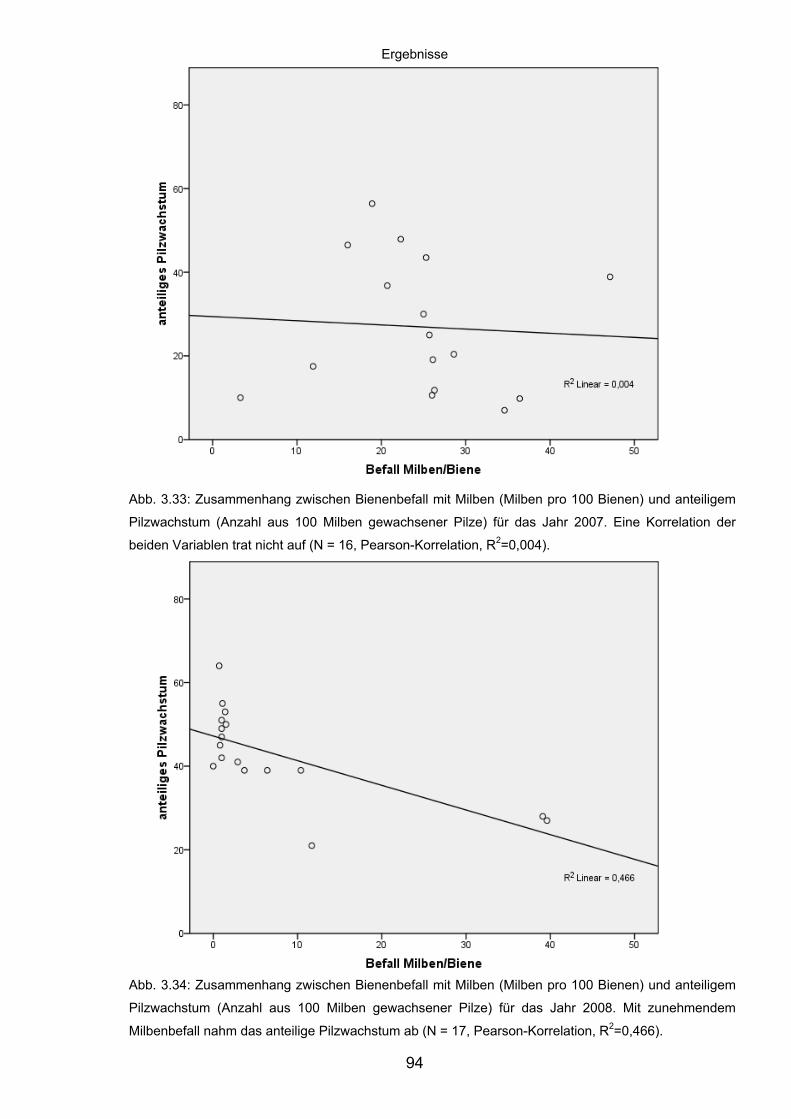
Ergebnisse
94
Abb. 3.33: Zusammenhang zwischen Bienenbefall mit Milben (Milben pro 100 Bienen) und anteiligem
Pilzwachstum (Anzahl aus 100 Milben gewachsener Pilze) für das Jahr 2007. Eine Korrelation der
beiden Variablen trat nicht auf (N = 16, Pearson-Korrelation, R2=0,004).
Abb. 3.34: Zusammenhang zwischen Bienenbefall mit Milben (Milben pro 100 Bienen) und anteiligem
Pilzwachstum (Anzahl aus 100 Milben gewachsener Pilze) für das Jahr 2008. Mit zunehmendem
Milbenbefall nahm das anteilige Pilzwachstum ab (N = 17, Pearson-Korrelation, R2=0,466).
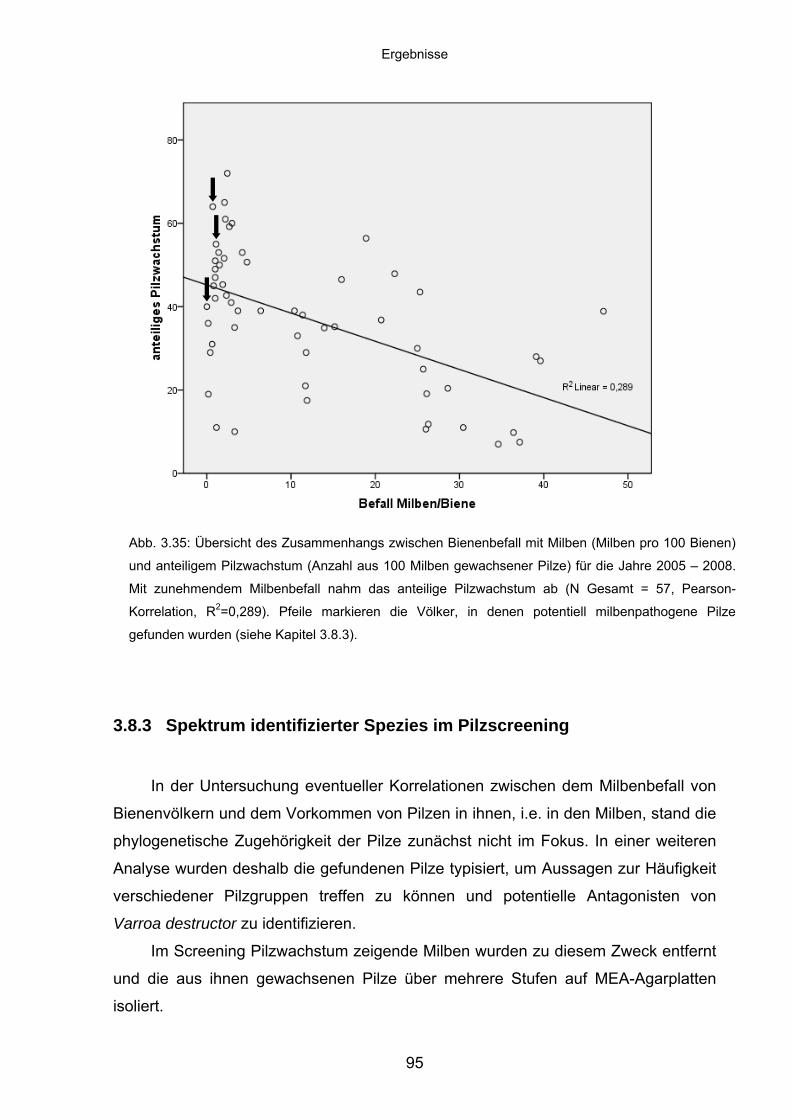
Ergebnisse
95
3.8.3 Spektrum identifizierter Spezies im Pilzscreening
In der Untersuchung eventueller Korrelationen zwischen dem Milbenbefall von
Bienenvölkern und dem Vorkommen von Pilzen in ihnen, i.e. in den Milben, stand die
phylogenetische Zugehörigkeit der Pilze zunächst nicht im Fokus. In einer weiteren
Analyse wurden deshalb die gefundenen Pilze typisiert, um Aussagen zur Häufigkeit
verschiedener Pilzgruppen treffen zu können und potentielle Antagonisten von
Varroa destructor zu identifizieren.
Im Screening Pilzwachstum zeigende Milben wurden zu diesem Zweck entfernt
und die aus ihnen gewachsenen Pilze über mehrere Stufen auf MEA-Agarplatten
isoliert.
Abb. 3.35: Übersicht des Zusammenhangs zwischen Bienenbefall mit Milben (Milben pro 100 Bienen)
und anteiligem Pilzwachstum (Anzahl aus 100 Milben gewachsener Pilze) für die Jahre 2005 – 2008.
Mit zunehmendem Milbenbefall nahm das anteilige Pilzwachstum ab (N Gesamt = 57, Pearson-
Korrelation, R2=0,289). Pfeile markieren die Völker, in denen potentiell milbenpathogene Pilze
gefunden wurden (siehe Kapitel 3.8.3).
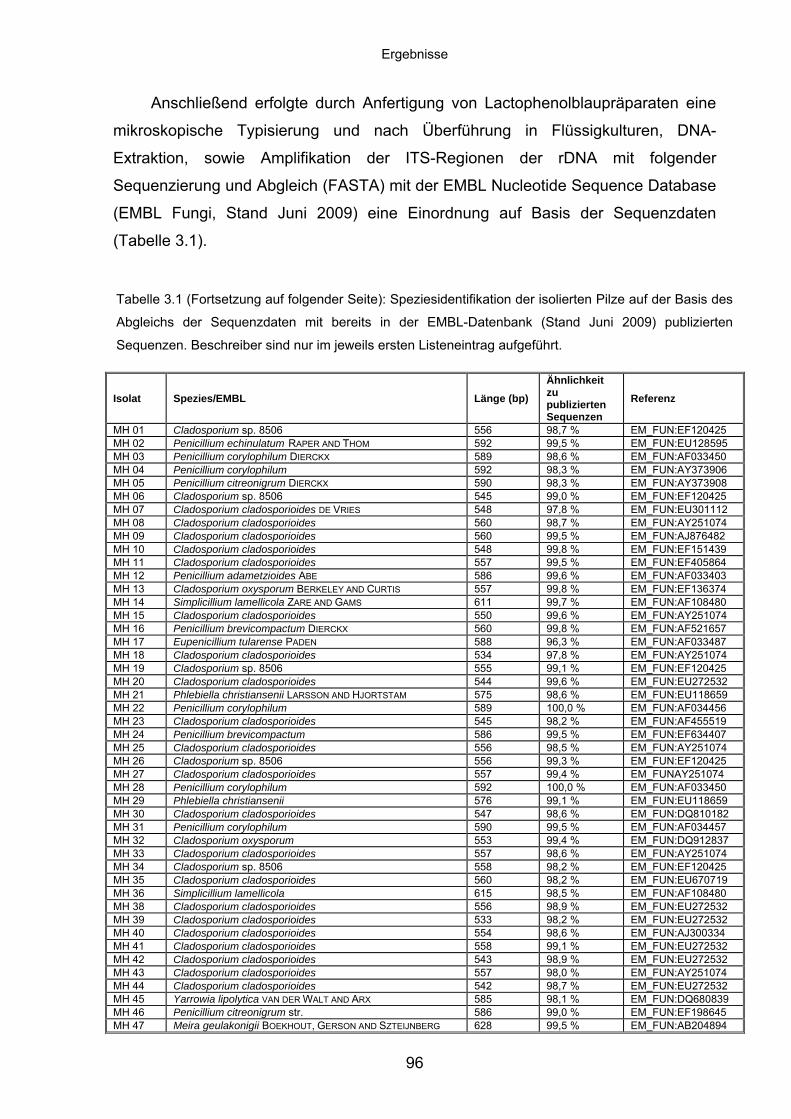
Ergebnisse
96
Anschließend erfolgte durch Anfertigung von Lactophenolblaupräparaten eine
mikroskopische Typisierung und nach Überführung in Flüssigkulturen, DNA-
Extraktion, sowie Amplifikation der ITS-Regionen der rDNA mit folgender
Sequenzierung und Abgleich (FASTA) mit der EMBL Nucleotide Sequence Database
(EMBL Fungi, Stand Juni 2009) eine Einordnung auf Basis der Sequenzdaten
(Tabelle 3.1).
Isolat Spezies/EMBL Länge (bp)
Ähnlichkeit zu publizierten Sequenzen
Referenz
MH 01 Cladosporium sp. 8506 556 98,7 % EM_FUN:EF120425 MH 02 Penicillium echinulatum RAPER AND THOM 592 99,5 % EM_FUN:EU128595 MH 03 Penicillium corylophilum DIERCKX 589 98,6 % EM_FUN:AF033450 MH 04 Penicillium corylophilum 592 98,3 % EM_FUN:AY373906 MH 05 Penicillium citreonigrum DIERCKX 590 98,3 % EM_FUN:AY373908 MH 06 Cladosporium sp. 8506 545 99,0 % EM_FUN:EF120425 MH 07 Cladosporium cladosporioides DE VRIES 548 97,8 % EM_FUN:EU301112 MH 08 Cladosporium cladosporioides 560 98,7 % EM_FUN:AY251074 MH 09 Cladosporium cladosporioides 560 99,5 % EM_FUN:AJ876482 MH 10 Cladosporium cladosporioides 548 99,8 % EM_FUN:EF151439 MH 11 Cladosporium cladosporioides 557 99,5 % EM_FUN:EF405864 MH 12 Penicillium adametzioides ABE 586 99,6 % EM_FUN:AF033403 MH 13 Cladosporium oxysporum BERKELEY AND CURTIS 557 99,8 % EM_FUN:EF136374 MH 14 Simplicillium lamellicola ZARE AND GAMS 611 99,7 % EM_FUN:AF108480 MH 15 Cladosporium cladosporioides 550 99,6 % EM_FUN:AY251074 MH 16 Penicillium brevicompactum DIERCKX 560 99,8 % EM_FUN:AF521657 MH 17 Eupenicillium tularense PADEN 588 96,3 % EM_FUN:AF033487 MH 18 Cladosporium cladosporioides 534 97,8 % EM_FUN:AY251074 MH 19 Cladosporium sp. 8506 555 99,1 % EM_FUN:EF120425 MH 20 Cladosporium cladosporioides 544 99,6 % EM_FUN:EU272532 MH 21 Phlebiella christiansenii LARSSON AND HJORTSTAM 575 98,6 % EM_FUN:EU118659 MH 22 Penicillium corylophilum 589 100,0 % EM_FUN:AF034456 MH 23 Cladosporium cladosporioides 545 98,2 % EM_FUN:AF455519 MH 24 Penicillium brevicompactum 586 99,5 % EM_FUN:EF634407 MH 25 Cladosporium cladosporioides 556 98,5 % EM_FUN:AY251074 MH 26 Cladosporium sp. 8506 556 99,3 % EM_FUN:EF120425 MH 27 Cladosporium cladosporioides 557 99,4 % EM_FUNAY251074 MH 28 Penicillium corylophilum 592 100,0 % EM_FUN:AF033450 MH 29 Phlebiella christiansenii 576 99,1 % EM_FUN:EU118659 MH 30 Cladosporium cladosporioides 547 98,6 % EM_FUN:DQ810182 MH 31 Penicillium corylophilum 590 99,5 % EM_FUN:AF034457 MH 32 Cladosporium oxysporum 553 99,4 % EM_FUN:DQ912837 MH 33 Cladosporium cladosporioides 557 98,6 % EM_FUN:AY251074 MH 34 Cladosporium sp. 8506 558 98,2 % EM_FUN:EF120425 MH 35 Cladosporium cladosporioides 560 98,2 % EM_FUN:EU670719 MH 36 Simplicillium lamellicola 615 98,5 % EM_FUN:AF108480 MH 38 Cladosporium cladosporioides 556 98,9 % EM_FUN:EU272532 MH 39 Cladosporium cladosporioides 533 98,2 % EM_FUN:EU272532 MH 40 Cladosporium cladosporioides 554 98,6 % EM_FUN:AJ300334 MH 41 Cladosporium cladosporioides 558 99,1 % EM_FUN:EU272532 MH 42 Cladosporium cladosporioides 543 98,9 % EM_FUN:EU272532 MH 43 Cladosporium cladosporioides 557 98,0 % EM_FUN:AY251074 MH 44 Cladosporium cladosporioides 542 98,7 % EM_FUN:EU272532 MH 45 Yarrowia lipolytica VAN DER WALT AND ARX 585 98,1 % EM_FUN:DQ680839 MH 46 Penicillium citreonigrum str. 586 99,0 % EM_FUN:EF198645 MH 47 Meira geulakonigii BOEKHOUT, GERSON AND SZTEIJNBERG 628 99,5 % EM_FUN:AB204894
Tabelle 3.1 (Fortsetzung auf folgender Seite): Speziesidentifikation der isolierten Pilze auf der Basis des
Abgleichs der Sequenzdaten mit bereits in der EMBL-Datenbank (Stand Juni 2009) publizierten
Sequenzen. Beschreiber sind nur im jeweils ersten Listeneintrag aufgeführt.
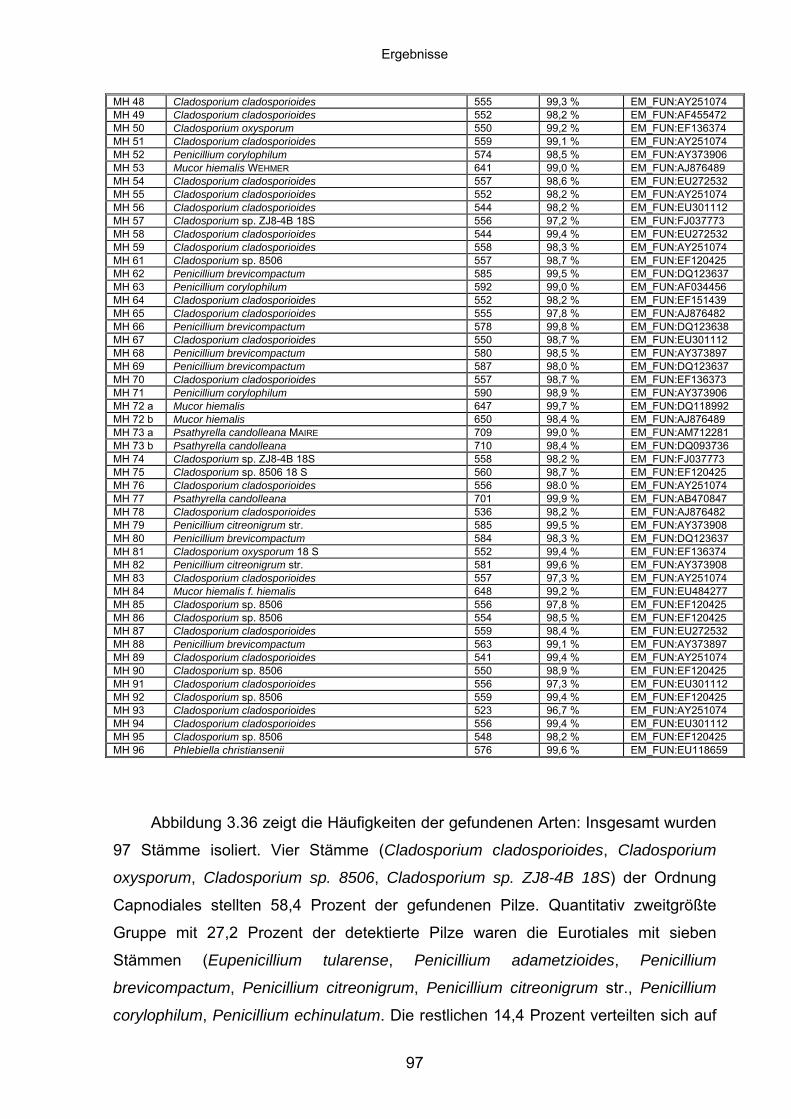
Ergebnisse
97
Abbildung 3.36 zeigt die Häufigkeiten der gefundenen Arten: Insgesamt wurden
97 Stämme isoliert. Vier Stämme (Cladosporium cladosporioides, Cladosporium
oxysporum, Cladosporium sp. 8506, Cladosporium sp. ZJ8-4B 18S) der Ordnung
Capnodiales stellten 58,4 Prozent der gefundenen Pilze. Quantitativ zweitgrößte
Gruppe mit 27,2 Prozent der detektierte Pilze waren die Eurotiales mit sieben
Stämmen (Eupenicillium tularense, Penicillium adametzioides, Penicillium
brevicompactum, Penicillium citreonigrum, Penicillium citreonigrum str., Penicillium
corylophilum, Penicillium echinulatum. Die restlichen 14,4 Prozent verteilten sich auf
MH 48 Cladosporium cladosporioides 555 99,3 % EM_FUN:AY251074 MH 49 Cladosporium cladosporioides 552 98,2 % EM_FUN:AF455472 MH 50 Cladosporium oxysporum 550 99,2 % EM_FUN:EF136374 MH 51 Cladosporium cladosporioides 559 99,1 % EM_FUN:AY251074 MH 52 Penicillium corylophilum 574 98,5 % EM_FUN:AY373906 MH 53 Mucor hiemalis WEHMER 641 99,0 % EM_FUN:AJ876489 MH 54 Cladosporium cladosporioides 557 98,6 % EM_FUN:EU272532 MH 55 Cladosporium cladosporioides 552 98,2 % EM_FUN:AY251074 MH 56 Cladosporium cladosporioides 544 98,2 % EM_FUN:EU301112 MH 57 Cladosporium sp. ZJ8-4B 18S 556 97,2 % EM_FUN:FJ037773 MH 58 Cladosporium cladosporioides 544 99,4 % EM_FUN:EU272532 MH 59 Cladosporium cladosporioides 558 98,3 % EM_FUN:AY251074 MH 61 Cladosporium sp. 8506 557 98,7 % EM_FUN:EF120425 MH 62 Penicillium brevicompactum 585 99,5 % EM_FUN:DQ123637 MH 63 Penicillium corylophilum 592 99,0 % EM_FUN:AF034456 MH 64 Cladosporium cladosporioides 552 98,2 % EM_FUN:EF151439 MH 65 Cladosporium cladosporioides 555 97,8 % EM_FUN:AJ876482 MH 66 Penicillium brevicompactum 578 99,8 % EM_FUN:DQ123638 MH 67 Cladosporium cladosporioides 550 98,7 % EM_FUN:EU301112 MH 68 Penicillium brevicompactum 580 98,5 % EM_FUN:AY373897 MH 69 Penicillium brevicompactum 587 98,0 % EM_FUN:DQ123637 MH 70 Cladosporium cladosporioides 557 98,7 % EM_FUN:EF136373 MH 71 Penicillium corylophilum 590 98,9 % EM_FUN:AY373906 MH 72 a Mucor hiemalis 647 99,7 % EM_FUN:DQ118992 MH 72 b Mucor hiemalis 650 98,4 % EM_FUN:AJ876489 MH 73 a Psathyrella candolleana MAIRE 709 99,0 % EM_FUN:AM712281 MH 73 b Psathyrella candolleana 710 98,4 % EM_FUN:DQ093736 MH 74 Cladosporium sp. ZJ8-4B 18S 558 98,2 % EM_FUN:FJ037773 MH 75 Cladosporium sp. 8506 18 S 560 98,7 % EM_FUN:EF120425 MH 76 Cladosporium cladosporioides 556 98.0 % EM_FUN:AY251074 MH 77 Psathyrella candolleana 701 99,9 % EM_FUN:AB470847 MH 78 Cladosporium cladosporioides 536 98,2 % EM_FUN:AJ876482 MH 79 Penicillium citreonigrum str. 585 99,5 % EM_FUN:AY373908 MH 80 Penicillium brevicompactum 584 98,3 % EM_FUN:DQ123637 MH 81 Cladosporium oxysporum 18 S 552 99,4 % EM_FUN:EF136374 MH 82 Penicillium citreonigrum str. 581 99,6 % EM_FUN:AY373908 MH 83 Cladosporium cladosporioides 557 97,3 % EM_FUN:AY251074 MH 84 Mucor hiemalis f. hiemalis 648 99,2 % EM_FUN:EU484277 MH 85 Cladosporium sp. 8506 556 97,8 % EM_FUN:EF120425 MH 86 Cladosporium sp. 8506 554 98,5 % EM_FUN:EF120425 MH 87 Cladosporium cladosporioides 559 98,4 % EM_FUN:EU272532 MH 88 Penicillium brevicompactum 563 99,1 % EM_FUN:AY373897 MH 89 Cladosporium cladosporioides 541 99,4 % EM_FUN:AY251074 MH 90 Cladosporium sp. 8506 550 98,9 % EM_FUN:EF120425 MH 91 Cladosporium cladosporioides 556 97,3 % EM_FUN:EU301112 MH 92 Cladosporium sp. 8506 559 99,4 % EM_FUN:EF120425 MH 93 Cladosporium cladosporioides 523 96,7 % EM_FUN:AY251074 MH 94 Cladosporium cladosporioides 556 99,4 % EM_FUN:EU301112 MH 95 Cladosporium sp. 8506 548 98,2 % EM_FUN:EF120425 MH 96 Phlebiella christiansenii 576 99,6 % EM_FUN:EU118659
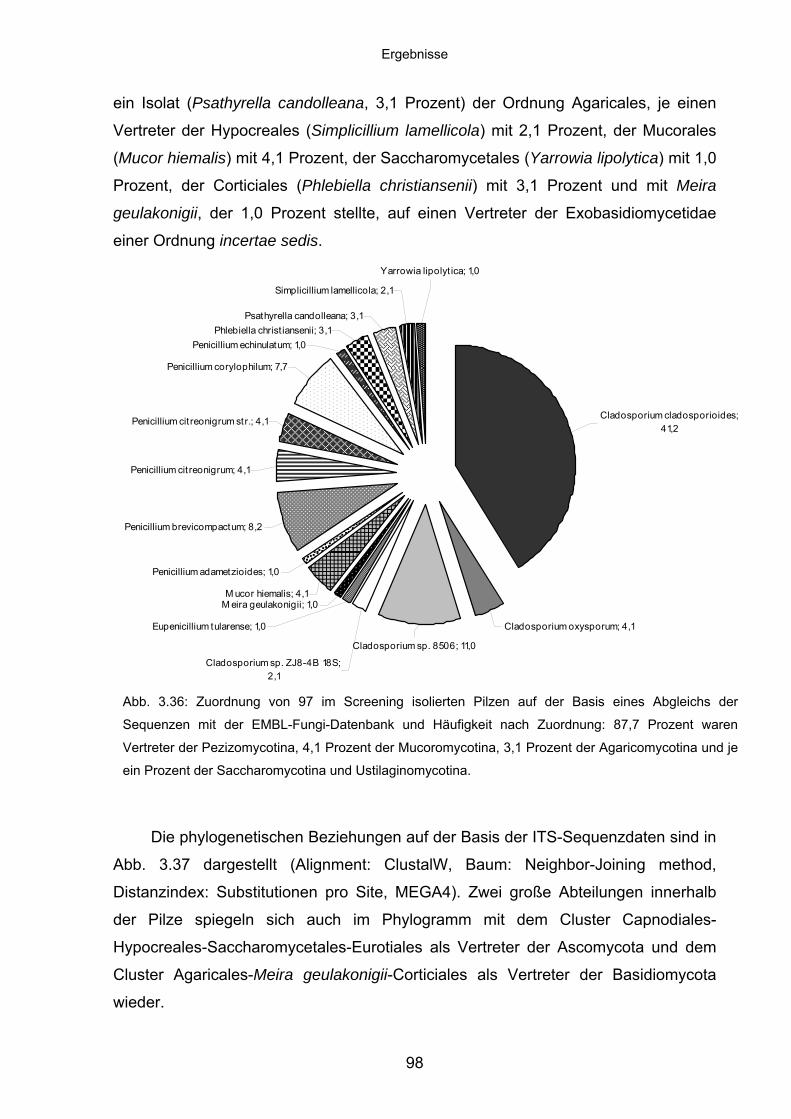
Ergebnisse
98
Penicillium cit reonigrum str.; 4,1
Penicillium cit reonigrum; 4,1
Penicillium brevicompactum; 8,2
Penicillium adametzioides; 1,0
M ucor hiemalis; 4,1
Eupenicillium tularense; 1,0
M eira geulakonigii; 1,0
Cladosporium sp. ZJ8-4B 18S; 2,1
Cladosporium sp. 8506; 11,0
Cladosporium oxysporum; 4,1
Cladosporium cladosporioides; 41,2
Penicillium corylophilum; 7,7
Phlebiella christ iansenii; 3,1
Penicillium echinulatum; 1,0
Psathyrella candolleana; 3,1
Simplicillium lamellicola; 2,1
Yarrowia lipolyt ica; 1,0
ein Isolat (Psathyrella candolleana, 3,1 Prozent) der Ordnung Agaricales, je einen
Vertreter der Hypocreales (Simplicillium lamellicola) mit 2,1 Prozent, der Mucorales
(Mucor hiemalis) mit 4,1 Prozent, der Saccharomycetales (Yarrowia lipolytica) mit 1,0
Prozent, der Corticiales (Phlebiella christiansenii) mit 3,1 Prozent und mit Meira
geulakonigii, der 1,0 Prozent stellte, auf einen Vertreter der Exobasidiomycetidae
einer Ordnung incertae sedis.
Die phylogenetischen Beziehungen auf der Basis der ITS-Sequenzdaten sind in
Abb. 3.37 dargestellt (Alignment: ClustalW, Baum: Neighbor-Joining method,
Distanzindex: Substitutionen pro Site, MEGA4). Zwei große Abteilungen innerhalb
der Pilze spiegeln sich auch im Phylogramm mit dem Cluster Capnodiales-
Hypocreales-Saccharomycetales-Eurotiales als Vertreter der Ascomycota und dem
Cluster Agaricales-Meira geulakonigii-Corticiales als Vertreter der Basidiomycota
wieder.
Abb. 3.36: Zuordnung von 97 im Screening isolierten Pilzen auf der Basis eines Abgleichs der
Sequenzen mit der EMBL-Fungi-Datenbank und Häufigkeit nach Zuordnung: 87,7 Prozent waren
Vertreter der Pezizomycotina, 4,1 Prozent der Mucoromycotina, 3,1 Prozent der Agaricomycotina und je
ein Prozent der Saccharomycotina und Ustilaginomycotina.

Ergebnisse
99
Die Mucorales als Vertreter der Zygomycota sind in der Analyse ohne direkte
Verwandte und clustern, bedingt durch die untersuche DNA-Region, als Artefakt
innerhalb der Basidiomycota. Nahezu alle Vertreter der beiden Cluster werden von
opportunistischen, euryöken Saprobionten gestellt, in beiden Clustern tritt aber auch
je ein Pilz auf, der Arten zugeordnet ist, die für Arthropoden pathogen sein können:
Innerhalb der Basidiomycota handelt es sich um Meira geulakonigii (Isolat MH47-
ITS1F), im Ascomycota-Cluster um Simplicillium lamellicola (Isolate MH 14-ITS1F
und MH 36-ITS1F).
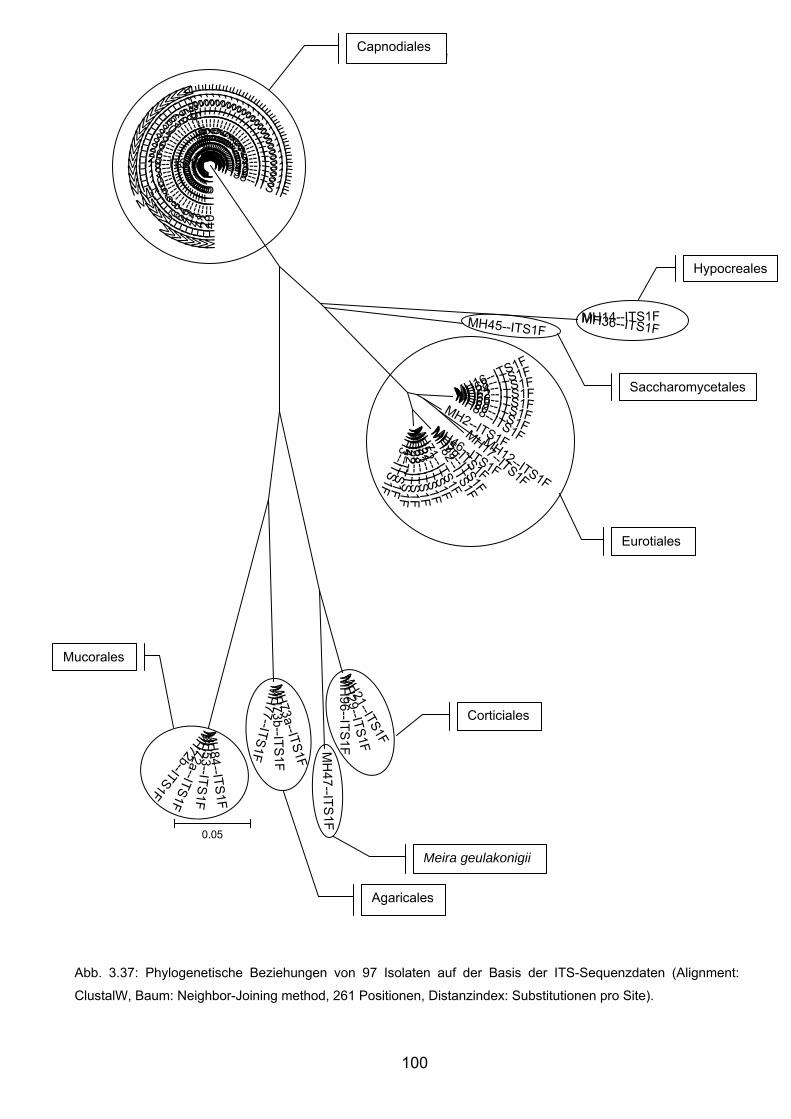
Ergebnisse
100
Capnodiales
Corticiales
Meira geulakonigii
Mucorales
Agaricales
Eurotiales
Hypocreales
Saccharomycetales
MH
40--
ITS
1F
MH
23--
ITS
1FM
H41
--IT
S1F
MH
43--
ITS
1F
MH
44--
ITS
1F
MH
49--
ITS
1F
MH
64--
ITS
1F
MH
67--I
TS1F
MH
8--IT
S1F
MH1-
-ITS1F
MH6--ITS1F
MH7--ITS1F
MH15--ITS1F
MH9--ITS1F
MH10--ITS1FMH11--ITS1FMH13--ITS1FMH18--ITS1FMH19--ITS1FMH25--ITS1FMH26--ITS1F
MH27--ITS1F
MH30--ITS1F
MH32--ITS1F
MH33--ITS1F
MH34--ITS1F
MH50--ITS1F
MH61--ITS1F
MH
65--ITS1F
MH
70--ITS1F
MH
58--ITS1F
MH
20--ITS
1F
MH
35--ITS
1F
MH
95--
ITS
1FM
H94
--IT
S1F
MH
93-
-IT
S1F
MH
92-
-IT
S1F
MH
91-
-IT
S1F
MH
90--
ITS
1FM
H89
--IT
S1F
MH
87--
ITS
1F
MH
86--
ITS
1F
MH
85--
ITS
1F
MH
83--I
TS1F
MH81
--ITS
1F
MH78
--ITS1F
MH76--ITS1F
MH75--ITS1F
MH74--ITS1F
MH59--ITS1F
MH57--ITS1F
MH56--ITS1FMH55--ITS1FMH54--ITS1FMH51--ITS1FMH48--ITS1FMH42--ITS1FMH39--ITS1F
MH38--ITS1F
MH14--ITS1FMH36--ITS1FMH45--ITS1F
MH16--ITS1F
MH68--ITS1F
MH24--ITS1FMH62--ITS1FMH66--ITS1FMH69--ITS1F
MH80--ITS1F
MH88--ITS1F
MH2--ITS1FMH12--ITS1F
MH17--ITS1F
MH46--ITS1F
MH5--ITS1F
MH
79--ITS1F
MH
82--ITS1F
MH
71--ITS1F
MH
52--ITS
1F
MH
31--ITS
1F
MH
63--ITS
1FM
H2
8--ITS
1F
MH
22--ITS
1FM
H4--IT
S1F
MH
3--ITS1F
MH
21--ITS1F
MH
29--ITS
1F
MH
96 --IT
S1
FMH
47--I T
S1F
MH
73a--ITS
1F
MH
73b
--ITS
1FM
H77--IT
S1F
MH
84--ITS
1FM
H53--IT
S1F
MH
72a--ITS
1F
MH
72b--ITS1F
0.05
Abb. 3.37: Phylogenetische Beziehungen von 97 Isolaten auf der Basis der ITS-Sequenzdaten (Alignment:
ClustalW, Baum: Neighbor-Joining method, 261 Positionen, Distanzindex: Substitutionen pro Site).

Ergebnisse
101
3.8.4 Charakteristika von Meira geulakonigii
Die Abbildungen 3.38 (eigene Daten rot markiert) und 3.39 verdeutlichen die
Beziehungen des gefundenen Isolates mit bereits publizierten Sequenzen, die der
Gattung Meira zugeordnet sind: Größte Homologie (Abb. 3.38, erste und zweite
Reihe) besteht zu den Isolaten PFS 014, das Yasuda publiziert hat, und AS 004, das
von Boekhout stammt und Typuskultur der Erstbeschreibung ist (Boekhout et al.
2003; Yasuda et al. 2007). Yasuda isolierte den Pilz aus Pyrus pyrifolia NAKAI-
Kulturen in Japan, Boekhout aus der phytopathogenen Milbe Phyllocoptruta oleivora
ASHMEAD auf Citrus grandis OSBECK-Kulturen in Israel. Das in der vorliegenden
Untersuchung gefundene Isolat aus Varroa destructor weist, verglichen mit den
beiden oben beschriebenen Isolaten, zwar nur geringe Sequenzdifferenzen auf, kann
aber von diesen abgegrenzt werden; Tabelle 3.2 gibt die paarweisen Distanzen zu
insgesamt elf, Arten der Gattung Meira zugeordneten Sequenzen an (Maximum
Composite Likelihood-Modell, 484 Sites).
1 2 3 4 5 6 7 8 9 10 11
1 Meira geulakonigii PFS014
2 Meira geulakonigii AS004
0,000
3 Meira argovae AS005 0,043 0,043
4 Meira argovae AS003 0,043 0,043 0,000
5 MH47-ITS1F 0,002 0,002 0,043 0,043
6 Meira sp. Vega E1-92 0,021 0,021 0,043 0,043 0,023
7 Meira argovae AS006 0,043 0,043 0,000 0,000 0,043 0,043
8 Meira argovae AS002 0,043 0,043 0,000 0,000 0,043 0,043 0,000
9 Meira argovae MAFF 240320
0,047 0,047 0,008 0,008 0,047 0,043 0,008 0,008
10 Meira sp. Vega E2-32 0,047 0,047 0,008 0,008 0,047 0,043 0,008 0,008 0,004
11 Meira nashicola PFS 002
0,058 0,058 0,067 0,067 0,060 0,056 0,067 0,067 0,072 0,072
12 Meira sp. IZ-1189 0,058 0,058 0,065 0,065 0,060 0,051 0,065 0,065 0,069 0,069 0,004
Tabelle 3.2: Distanzmatrix für elf der Gattung Meira zugeordnete, publizierte Sequenzen sowie für das
eigene Isolat. Die geringsten Distanzen bestehen zu den zwei als Meira geulakonigii publizierten
Isolaten. Berechnung auf der Basis von 484 Sites, Maximum Composite Likelihood-Modell.

Ergebnisse
102
Abb. 3.38: Alignment für zehn der Gattung Meira zugeordnete, publizierte Sequenzen sowie für das
eigene Isolat (jeweils erste Zeile, gerahmt). Höchste Homologie (erste Reihe) besteht zu einem Isolat,
das aus der Milbe Phyllocoptruta oleivora isoliert wurden; EMBL-FASTA, Juni 2009.
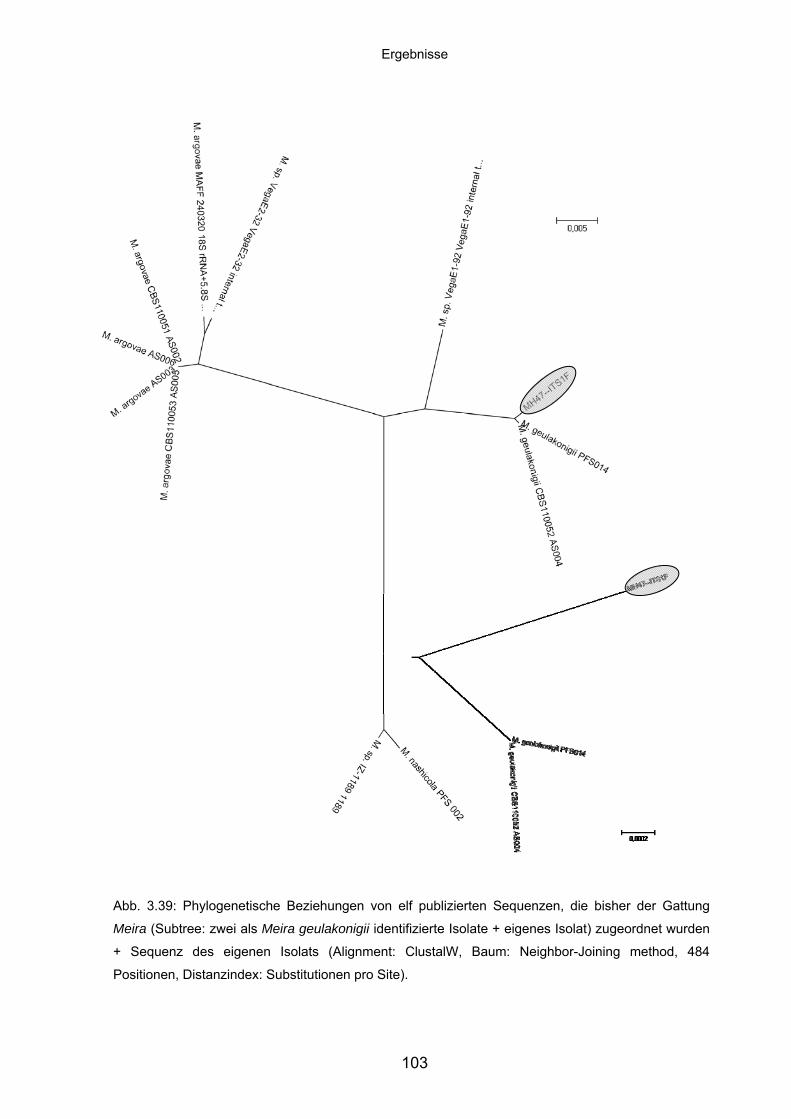
Ergebnisse
103
Abb. 3.39: Phylogenetische Beziehungen von elf publizierten Sequenzen, die bisher der Gattung
Meira (Subtree: zwei als Meira geulakonigii identifizierte Isolate + eigenes Isolat) zugeordnet wurden
+ Sequenz des eigenen Isolats (Alignment: ClustalW, Baum: Neighbor-Joining method, 484
Positionen, Distanzindex: Substitutionen pro Site).
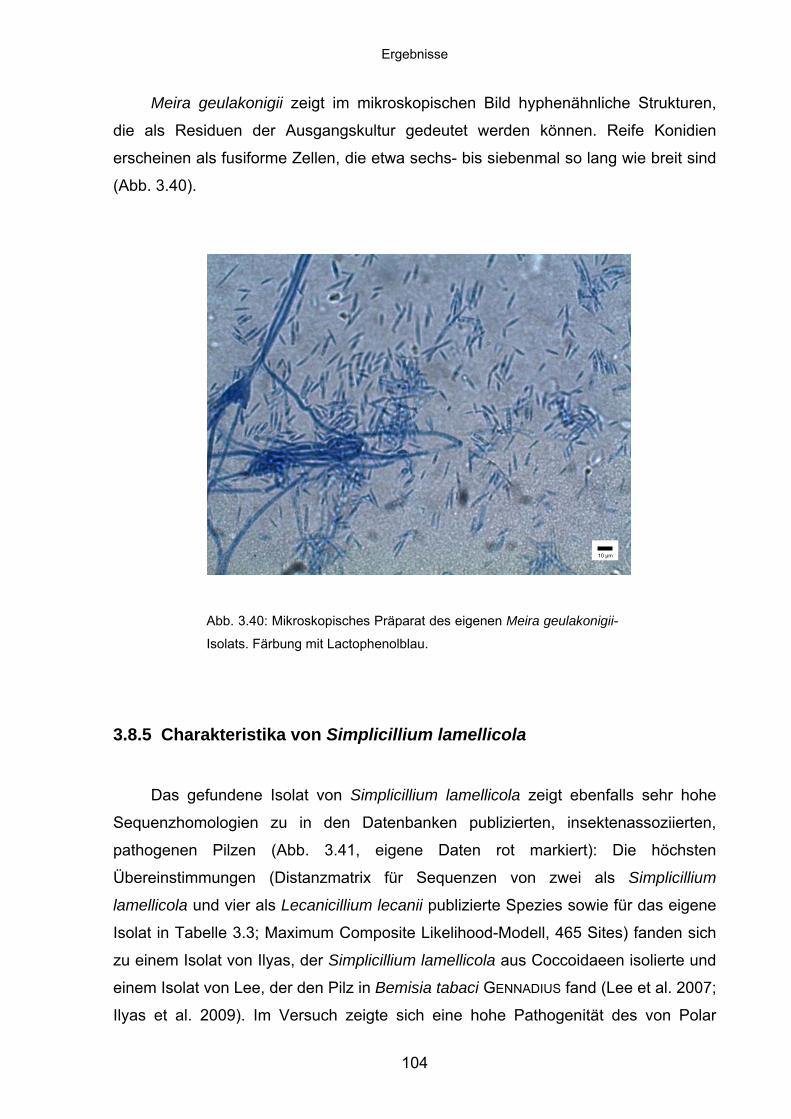
Ergebnisse
104
Meira geulakonigii zeigt im mikroskopischen Bild hyphenähnliche Strukturen,
die als Residuen der Ausgangskultur gedeutet werden können. Reife Konidien
erscheinen als fusiforme Zellen, die etwa sechs- bis siebenmal so lang wie breit sind
(Abb. 3.40).
3.8.5 Charakteristika von Simplicillium lamellicola
Das gefundene Isolat von Simplicillium lamellicola zeigt ebenfalls sehr hohe
Sequenzhomologien zu in den Datenbanken publizierten, insektenassoziierten,
pathogenen Pilzen (Abb. 3.41, eigene Daten rot markiert): Die höchsten
Übereinstimmungen (Distanzmatrix für Sequenzen von zwei als Simplicillium
lamellicola und vier als Lecanicillium lecanii publizierte Spezies sowie für das eigene
Isolat in Tabelle 3.3; Maximum Composite Likelihood-Modell, 465 Sites) fanden sich
zu einem Isolat von Ilyas, der Simplicillium lamellicola aus Coccoidaeen isolierte und
einem Isolat von Lee, der den Pilz in Bemisia tabaci GENNADIUS fand (Lee et al. 2007;
Ilyas et al. 2009). Im Versuch zeigte sich eine hohe Pathogenität des von Polar
Abb. 3.40: Mikroskopisches Präparat des eigenen Meira geulakonigii-
Isolats. Färbung mit Lactophenolblau.
10 µm
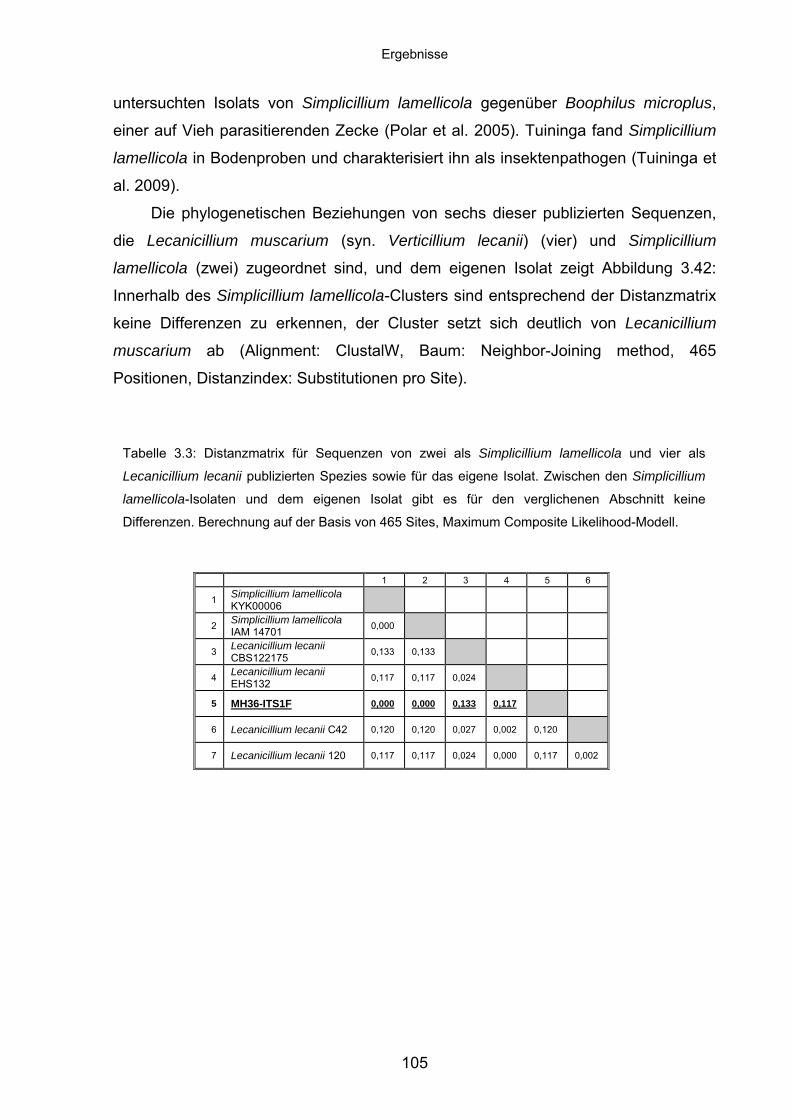
Ergebnisse
105
untersuchten Isolats von Simplicillium lamellicola gegenüber Boophilus microplus,
einer auf Vieh parasitierenden Zecke (Polar et al. 2005). Tuininga fand Simplicillium
lamellicola in Bodenproben und charakterisiert ihn als insektenpathogen (Tuininga et
al. 2009).
Die phylogenetischen Beziehungen von sechs dieser publizierten Sequenzen,
die Lecanicillium muscarium (syn. Verticillium lecanii) (vier) und Simplicillium
lamellicola (zwei) zugeordnet sind, und dem eigenen Isolat zeigt Abbildung 3.42:
Innerhalb des Simplicillium lamellicola-Clusters sind entsprechend der Distanzmatrix
keine Differenzen zu erkennen, der Cluster setzt sich deutlich von Lecanicillium
muscarium ab (Alignment: ClustalW, Baum: Neighbor-Joining method, 465
Positionen, Distanzindex: Substitutionen pro Site).
1 2 3 4 5 6
1 Simplicillium lamellicola KYK00006
2 Simplicillium lamellicola IAM 14701
0,000
3 Lecanicillium lecanii CBS122175
0,133 0,133
4 Lecanicillium lecanii EHS132
0,117 0,117 0,024
5 MH36-ITS1F 0,000 0,000 0,133 0,117
6 Lecanicillium lecanii C42 0,120 0,120 0,027 0,002 0,120
7 Lecanicillium lecanii 120 0,117 0,117 0,024 0,000 0,117 0,002
Tabelle 3.3: Distanzmatrix für Sequenzen von zwei als Simplicillium lamellicola und vier als
Lecanicillium lecanii publizierten Spezies sowie für das eigene Isolat. Zwischen den Simplicillium
lamellicola-Isolaten und dem eigenen Isolat gibt es für den verglichenen Abschnitt keine
Differenzen. Berechnung auf der Basis von 465 Sites, Maximum Composite Likelihood-Modell.
Ergebnisse
106
Abb. 3.41: Alignment für 10 den Spezies Simplicillium lamellicola (Reihen eins bis sechs, acht bis zehn) und
Guignardia vaccinii (Reihe sieben) zugeordnete, publizierte Sequenzen sowie für das eigene Isolat (jeweils
erste Zeile, gerahmt). Höchste Homologie besteht zu einem Isolat aus unbekannter Quelle (erste Reihe,
unpubliziert) und zu einem Isolat, das aus Coccidaeen isoliert wurde (zweite Reihe); EMBL-FASTA, Juni 2009.
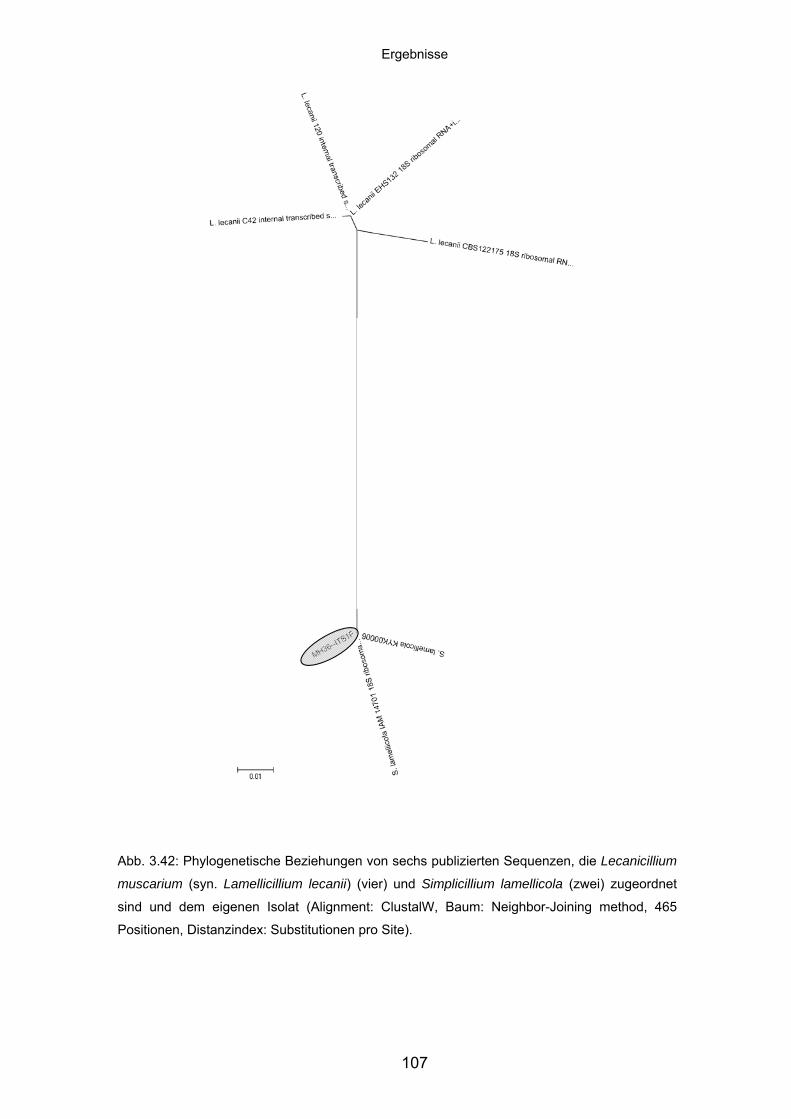
Ergebnisse
107
Abb. 3.42: Phylogenetische Beziehungen von sechs publizierten Sequenzen, die Lecanicillium
muscarium (syn. Lamellicillium lecanii) (vier) und Simplicillium lamellicola (zwei) zugeordnet
sind und dem eigenen Isolat (Alignment: ClustalW, Baum: Neighbor-Joining method, 465
Positionen, Distanzindex: Substitutionen pro Site).

Ergebnisse
108
Im Gegensatz zu den zugespitzten Sporen von Meira geulakonigii zeigen sich
die Konidien im mikroskopischen Bild von Simplicillium lamellicola länglich
abgerundet; sie sind etwa fünfmal so lang wie breit; ihr Verbund löst sich nach
Abschnürung und bildet keine zusammenhängenden Ketten (Abb. 3.42).
Abb. 3.43: Mikroskopisches Präparat des eigenen Simplicillium
lamellicola-Isolats. Färbung mit Lactophenolblau.
10 µm

Diskussion
109
4 Diskussion
Entomopathogene Pilze als potentielle Antagonisten von Varroa destructor
werden als aussichtsreichste mittelfristige Lösung gesehen, um Apis mellifera-Völker
in Kultur halten zu können, ohne diesen Parasiten mit einer komplexen,
zeitaufwändigen Methodik bekämpfen zu müssen, da Erfolge in Versuchen, durch
markergestützte Selektion oder die gezielte Fertilisation und folgende Rückkreuzung
von Honigbienenarbeiterinnen, die ein Varroaantagonistisches Verhalten zeigen,
eine nachhaltige Varroaresistenz oder zumindest -toleranz zu erzielen, die
menschliche Eingriffe obsolet macht, bisher ausblieben (Büchler et al. 2008). Kurz
nach der letzten Jahrtausendwende wurde begonnen, verschiedene unspezifische
entomopathogene Pilze in Bezug auf ihre Wirkung auf Varroa destructor und Apis
mellifera zu testen. Auch wenn es in der Folge wiederholt zu Untersuchungen kam,
die eine antagonistische Wirkung entomopathogener Pilze konstatierten, liegen
bisher keine Berichte zu einem Durchbruch in Bezug auf einen Einsatz dieser Pilze
gegen Varroa vor.
Die in dieser Arbeit untersuchten Stämme von vier Pilzen – Beauveria
bassiana, Metarhizium anisopliae, Lecanicillium muscarium und Paecilomyces
fumosoroseus – zeigten in vitro kein Potential, den Befall von Honigbienen mit dem
Ektoparasiten Varroa destructor signifikant zu senken – in allen Versuchen gab es
keine signifikanten Differenzen zwischen den mit Konidien behandelten Gruppen und
den Kontrollen. Maxima des täglichen Milbenfalls traten sowohl in den Pilzgruppen
als auch in den Kontrollen etwa an den Tagen zwei bis fünf auf, was als Effekt der
spezifischen Bedingungen, wie der Gewinnungsmethode oder dem Besprühen mit
der der Ernte der Konidien dienenden Lösung gedeutet werden kann. Geringe
Differenzen in der Mortalität zwischen den Kontrollen der einzelnen Versuche
wiederum sind der Methodik der Gewinnung der Versuchstiere geschuldet: Die große
Zahl benötigter Milben lässt sich nur aus jeweils unterschiedlichen Völkern gewinnen,
zudem sind derart umfangreiche Versuche nur seriell durchzuführen; damit ist davon
auszugehen, dass sich die Kohorten der verwendeten Milben (und auch Bienen) in
den in vitro-Versuchen zum Beispiel in Bezug auf ihre Altersstruktur nicht gleichen.
Die Ergebnisse der vorliegenden Untersuchungen decken sich nicht mit den
Ergebnissen der beiden anderen Studien an Milben in vitro, die sich mit den gleichen
Spezies beschäftigten (Kanga et al. 2002; Shaw et al. 2002). Shaw gibt für

Diskussion
110
behandelte Milben eine Mortalität von 63 bis 100 Prozent für den siebten Tag nach
Behandlung, für Milben der Kontrollgruppe eine zu vernachlässigende Mortalität an
und schließt die Kontrollgruppe in der Folge von der Analyse aus. In der
vorliegenden Arbeit lagen die Mortalitäten der Versuchs- und Kontrollgruppen in allen
Untersuchungen zwischen 70 und 80 Prozent, ohne signifikant voneinander
abzuweichen. Allerdings unterscheiden sich die Untersuchungen grundsätzlich in
ihrer Methodik: Im Gegensatz zur in dieser Arbeit beschriebenen Vorgehensweise
applizierte Shaw die Konidiensupension durch Eintauchen der Milben und hälterte
diese anschließend in Fünfergruppen auf Bienenpuppen, die in 1,5 ml-
Eppendorftubes untergebracht waren, bei 25 oder 30 Grad, also bei artifiziellen
Bedingungen, die sehr stark von den Verhältnissen im Brutnest von Honigbienen
abweichen: Die Mortalitäten in vitro gehälterter Larvenstadien und Puppen und damit
auch der Milben liegen über denen im Volk, die gewählten Temperaturen weichen
erheblich von den Brutnesttemperaturen ab und eine fünffache Parasitierung von
Puppen kommt allenfalls in Bienenvölkern unmittelbar vor deren Zusammenbruch vor
– ein Überleben von derart stark parasitierten Puppen und damit der Milben bis zur
Imaginalhäutung beziehungsweise bis zum Schlupf der Bienen erscheint äußerst
fraglich. Zur Behandlung der Kontrolle macht Shaw im Übrigen keine Angaben, was
die Vergleichbarkeit der Ergebnisse zusätzlich erschwert.
Bestätigt werden die vorliegenden in vitro-Ergebnisse für Varroa destructor
durch die Ergebnisse des Halbfreilandversuchs: Abgesehen von der Beschränkung
des Flugraums gab es in Bezug auf die Haltungsbedingungen keine artifiziellen
Bedingungen, die Effekte auf Milben erklären könnten; auch hier konnte eine
Wirkung auf Varroa destructor nicht beobachtet werden
Kanga testete einen Stamm von Metarhizium anisopliae in Laborversuchen an
Milben auf Puppen, bei denen eine konidienhaltige Detergenslösung versprüht
wurde, und beobachtete eine signifikante Reduktion der Varroapopulationen, ohne
dass die Bienen geschädigt wurden; er bestimmte eine LT90 von sechs Tagen für
Metarhizium anisopliae (Kanga et al. 2002). In einem weiteren Versuch an Völkern,
die entweder mit Konidienpulver oder dem pyrethroidhaltigen Tierarzneimittel Apistan
behandelt wurden, fand Kanga vergleichbare Mortalitätsraten und schlussfolgerte
daraus auf eine Eignung von Metarhizium als Kontrollagens, obwohl die kumulierten
Mortalitätsraten zu Versuchsende teilweise unter denen der Kontrollgruppe lagen
(Kanga et al. 2003). In einem Versuch in 2006, in dem Metarhiziumkonidien mit Hilfe

Diskussion
111
von mit ihnen bedeckten Kunststoffstreifen appliziert wurden, bestätigten sich die
Ergebnisse (Kanga et al. 2006). 2006 untersuchte James den Metarhiziumstamm,
mit dem Kanga gearbeitet hatte, vor dem Hintergrund der Fragestellung, welche
Applikationsform die effektivste sei, konnte dabei aber keinerlei signifikante Effekte
des Pilzes finden; zusätzlich waren längstens nach 20 Tagen (Kanga 2003: 40 Tage)
noch keimfähige Konidien vorhanden (Kanga et al. 2003; James et al. 2006). In den
Freilandversuchen von Gerritsen mit drei kommerziell erhältlichen Produkten auf der
Basis von Metarhizium anisopliae- und Lecanicillium muscarium-Stämmen zeigten
sich signifikant erhöhte Sterberaten weder für Varroa destructor noch für Apis
mellifera (Gerritsen et al. 2006). Diese widersprüchlichen Ergebnisse können zum
einen darin begründet liegen, dass es sich bei den untersuchten Pilzen um
ausgesprochen pleomorphe Spezies handelt, was eine Vergleichbarkeit der
Untersuchungen, sofern sie nicht an identischen Stämmen vorgenommen wurden,
schwierig macht. So hat beispielweise das Konidienkeimverhalten verschiedener
Stämme Einfluss auf die Virulenz gegenüber Wirten: Für Beauveria bassiana wurde
von Talaei-Hassanloui gezeigt, dass unidirektional keimende Konidien
appressorienartige Strukturen ausbildeten und hochvirulent waren, während sich bi-
und multipolar keimende Konidien mit hyphenartigem Wachstum als kaum bzw. nicht
virulent herausstellten (Talaei-Hassanloui et al. 2007). Ähnliche Ergebnisse fand
Altre für Paecilomyces fumosoroseus als Pathogen von Plutella xylostella LINNAEUS:
Hier korrelierte die Virulenz hochsignifikant mit der Sporenlänge und der
Keimgeschwindigkeit (Altre et al. 1999). Darüber hinaus ergab beispielsweise eine
Revision von Verticillium lecanii auf rDNA-Basis, dass es sich um einen Komplex aus
den fünf Spezies Lecanicillium attenuatum ZARE AND GAMS, Lecanicillium lecanii ZARE
AND GAMS, Lecanicillium longisporum ZARE AND GAMS, Lecanicillium muscarium und
Lecanicillium nodulosum ZARE AND GAMS handelt (Zare et al. 2000; Gams et al. 2001;
Zare et al. 2001). Da einige Autoren die Revision ohne eigene Identifizierung des
ihnen vorliegenden Isolats durch einfachen Ersatz des Gattungsnamens
nachvollzogen haben, handelt es sich möglicherweise auch bei den im
Zusammenhang mit ihrer Rolle als potentielle Varroaantagonisten aktuell diskutierten
Spezies um unterschiedliche Arten (Sugimoto et al. 2003; Koike et al. 2007). Weil
deren Wirtsspektren erheblich divergieren und von Lepidopteren über Homopteren
bis zu Vertretern der Acari und der Annelida reichen, könnten unterschiedliche
Ergebnisse zur „gleichen“ Art hierin begründet liegen. Zum anderen differieren wie

Diskussion
112
auch in den in vitro-Versuchen die verwendeten Methodiken erheblich: Die in der
vorliegenden Untersuchung im Halbfreilandversuch verwendeten Völker wurden
durch Poolen aus Donorvölkern gebildet, so dass sie zwingend homogen in Bezug
auf potentielle Toleranz- oder Resistenzfaktoren von Bienenarbeiterinnen gegenüber
Varroa destructor, aber auch in Bezug auf den Verwandtschaftsgrad der
Arbeiterinnen untereinander und ihre Altersstruktur waren. Ebenso stellte diese
Vorgehensweise sicher, dass die Abundanz von Varroa in allen Versuchsvölkern zu
Beginn exakt gleich hoch war und die Völker mit gleichen Individuenzahlen adulter
Bienen sowie ohne Brutstadien, die während des Versuchs Einfluss auf die Zahl
adulter Bienen haben, starteten. Diese Vorgehensweise wurde lediglich von Kanga in
seinen Versuchen an Bienenvölkern gewählt (Kanga et al. 2003). Auch die
Applikationstechnik der Konidien wurde unterschiedlich gehandhabt: Die Konidien
der eigenen Versuche wurden als Suspension versprüht, Kanga brachte die Konidien
als Pulver in die Völker ein, Shaw ließ in ihren Laborversuchen Milben über reife
Pilzkulturen laufen (Shaw et al. 2002; Kanga et al. 2003). Nicht untersucht wurde in
beiden Ansätzen dabei der mögliche Einfluss eines nichttoxischen, mechanischen
Effekts der Applikation, vergleichbar dem nach Einbringen fein vermahlener
Saccharose in Bienenvölker; mit dieser Technik wurden in der vorliegenden
Untersuchung Milben für die in vitro-Versuche gewonnen (Aliano et al. 2005; Ellis et
al. 2009).
Für einen aus Varroa destructor isolierten Beauveria bassiana-Stamm
beschreibt Meikle nach Untersuchungen an je zwei Völkern – ausgehend von einer
zuvor nicht vereinheitlichten Grundgesamtheit – einer mit Konidien behandelten,
einer nur mit einem als Matrix dienenden Pulver behandelten und einer nicht
behandelten Gruppe einen signifikant erhöhten Milbenfall der mit Konidien
behandelten Gruppe am sechsten und achten Tag nach Applikation; die Völker
differierten zuvor in Bezug auf die erfassten Parameter „Bienenmasse“ und „Fläche
verdeckelter Brut“ (Meikle et al. 2007). Milben aus den mit Konidien behandelten
Völkern zeigten, ohne zuvor oberflächensterilisiert worden zu sein, in allen Fällen ein
signifikant höheres relatives Pilzwachstum. In einer Repetition mit einer größeren
Zahl nicht vereinheitlichter Völker und vier verschiedenen Gruppen (Konidien und
Carnaubawachsmatrix, Konidien und Weizenmehlmatrix, Weizenmehl, keine
Behandlung) trat ein erhöhter Milbenfall in der Konidien-Carnaubawachsgruppe auf,
dieser war allerdings nicht signifikant erhöht (Meikle et al. 2008b). Der Milbenfall der

Diskussion
113
Konidien-Weizenmehlgruppe lag unterhalb des Falls der Weizenmehlgruppe und an
drei Tagen auch unterhalb des Falls der Kontrolle. Signifikant erhöhtes relatives
Milbenwachstum aus einer Konidiengruppe wertet Meikle aber als Beleg für die
Eignung des Pilzes als Varroaantagonist. Eine zweite Wiederholung erbrachte
keinerlei Belege für eine signifikante Reduktion der Parasitenlast der Versuchsvölker,
was als Hinweis auf die Notwendigkeit einer homogenen Ausgangsbasis gewertet
werden kann (Meikle et al. 2009).
Die Daten der vorliegenden Arbeit belegen für drei der vier untersuchten Pilze
(Lecanicillium muscarium, Beauveria bassiana und Paecilomyces fumosoroseus)
signifikant eine negative Wirkung auf Bienen, die für Paecilomyces in der Kumulation
über den gesamten Untersuchungszeitraum sichtbar wurde, bei Lecanicillium und
Beauveria sogar signifikante Differenzen an Einzeltagen (vier und sieben respektive
vier und sechs) zeigte. Die Mortalitätsraten der Bienen lagen in der vorliegenden
Untersuchung nach zehn Tagen zwischen 30 und 40 Prozent in den Pilzgruppen und
20 bis 30 Prozent in den Kontrollen, was – bezogen auf die Kontrollen – die
Ergebnisse von Shaw stützt, deren Methodik zur Untersuchung an Honigbienen
ähnlich war (Shaw et al. 2002). In den Pilzgruppen traten in ihrer Untersuchungen
Mortalitäten von bis zu 100 Prozent 14 Tage nach Behandlung auf; von wenigen
Fällen abgesehen lagen sie immer über denen der Kontrollen. Für Bienen schloss
Kanga in seinem Versuch aus vergleichbaren Pilzwachstumsraten aus toten Bienen,
sie würden durch die Pilzstämme nicht negativ beeinflusst – allerdings wurden die
Oberflächen der Bienen nicht sterilisiert, was mit Blick auf opportunistische
Saprobionten problematisch sein dürfte (Kanga et al. 2003). Der Befund
unterschiedlicher Ergebnisse für Bienen bestätigt die großen Unterschiede von
Pathogenität und Virulenz in Abhängigkeit vom untersuchten Isolat und legt, da in
den vorliegenden und den in vitro-Untersuchungen von Shaw die Mortalität der
Bienen unter dem Einfluss der Pilze erhöht war, nahe, dass diese möglicherweise
isolatunabhängig nicht spezifisch genug sind, um als Kontrollagens geeignet zu sein.
Im Halbfreilandversuch war eine erhöhte Mortalität der Bienen ebenso wie der
Larvenstadien und der Puppen nicht detektierbar – das widerspricht nicht zwingend
den Ergebnissen des in vitro-Assays: In in vitro-Versuchen an gekäfigten Bienen
kommt es zu Eingriffen wie zum Beispiel dem regelmäßigen Entfernen toter Bienen,
das auch bei vorsichtiger Handhabung unter Rotlicht zu einer stark zunehmenden
Aktivität der Bienen führt; derartige Eingriffe können damit immer als Stressoren

Diskussion
114
wirken. Bienen, die solchen Stressoren ausgesetzt sind, zeigten in anderen
Versuchen eine erhöhte Mortalität, so dass denkbar ist, dass unter diesem Einfluss
durch Immunsuppression ein Agens pathogen werden kann, das unter natürlichen
Bedingungen keine Effekte auslöst (Dustmann et al. 1988; Decourtye et al. 2003).
Die Nichtpathogenität der in dieser Untersuchung getesteten Pilze in Bezug auf
Varroa bestätigte sich in der Infektionsanalyse der im Versuchszeitraum gestorbenen
Milben: Nur für Beauveria bassiana (sieben Tage nach Behandlung) wurde ein
signifikanter Unterschied (p < 0,05) im erzielbaren relativen Pilzwachstum aus Milben
der Pilz- und Kontrollgruppe gefunden – in allen anderen Gruppen zeigten sich keine
Differenzen. Shaw fand in ihrer Untersuchung bei bis zu 100 Prozent der
gestorbenen Milben nach sieben Tagen Mykosen, gibt aber, da die Kontrollgruppe zu
Versuchsbeginn einen zu geringen Milbenfall aufwies, keinen Wert für die Kontrolle
an (Shaw et al. 2002). Das ist insofern problematisch, als vor der Kultur der Milben
keine Oberflächensterilisation stattfand. Die vorliegenden Untersuchungen zeigen
aber die zentrale Bedeutung einer ausreichenden Sterilisationsdauer für den
Ausschluss oder zumindest die weitestgehende Einschränkung des Wachstums der
Oberfläche anhaftender Konidien der applizierten Spezies oder opportunistischer
Saprobionten: Selbst nach einer sehr langen Sterilisationszeit von 240 Sekunden
kam es noch zu einem (geringen) Pilzwachstum aus nicht mit Pilzen behandelten
Milben, was an ingestierten oder intersegmental überdauerten Stadien vorhandener
Pilze liegen dürfte. Ausgeschlossen werden kann deshalb auch nicht, dass die
signifikante Differenz auf dem 95 Prozent-Niveau auf Residuen zurückzuführen ist.
Zwar unterzieht Kanga die in seinem Versuch gestorbenen Milben einer
Sterilisation, korreliert aber die Absolutbeträge der Zahl gefallener Milben und der
aus ihnen gewachsenen Pilze (Kanga et al. 2003). Meikle findet in seinen
Freilandversuchen in Abhängigkeit von der Versuchsdauer zwischen 100 Prozent
(am Tag der Behandlung) und 30 Prozent (50 Tage nach Behandlung) infizierte
Milben, verzichtet aber wie Shaw auf eine Sterilisation (Shaw et al. 2002; Meikle et
al. 2007, 2008a).
Die vorliegende Untersuchung an zehn Bienenvölkern im Halbfreilandversuch
mit Lecanicillium muscarium bestätigte die eigenen Ergebnisse aus dem Laborassay
für Milben auch im Detail: Über den gesamten Untersuchungszeitraum zeigte sich
keine signifikante Schädigung der Milbenpopulation im Vergleich zu Kontrollvölkern,
was sich nicht nur an der Zahl gestorbener Milben, analog zur Erfassung in den in

Diskussion
115
vitro-Assays feststellen lässt; auch der relative Endbefall adulter Bienen und
verdeckelter Brutzellen belegt keine Wirkung des Pilzes. Im Gegensatz zum
Laborassay wurde auch kein negativer Seiteneffekt gefunden – weder auf die Zahl
adulter Bienen, noch auf die Zahl der Larvenstadien oder verdeckelter Brutzellen.
Der Befund einer Nichtwirksamkeit gegenüber Milben und Bienen wird gestützt durch
die Untersuchungen von Gerritsen, die für den gleichen Lecanicillium muscarium-
Stamm sowie für einen Metarhizium anisopliae-Stamm im Freilandversuch an
allerdings zuvor nicht vereinheitlichten Völkern ebenfalls keinen Effekt belegen
(Gerritsen et al. 2006). Meikle konnte eine Wirkung des von ihm untersuchten
Beauveria bassiana-Stamms, der nach seinen früheren Ergebnissen letal auf Varroa
wirkte, in seiner jüngsten Untersuchung nicht mehr bestätigen (Meikle et al. 2006;
2007; 2008a; 2008b; 2009). Meikle arbeitete im Gegensatz zur vorliegenden Arbeit
ebenso wie Gerritsen mit einem nicht über eine Supergruppenbildung in Bezug auf
Altersstruktur, Verwandtschaft und Varroabefall homogenisierten Bienenmaterial,
was angesichts einer exponentiellen Vermehrung des Parasiten und einem
Generationenabstand von nur 30 Tagen eine erhebliche Varianz zwischen den
Jahren erklären kann. Zudem waren die Völker beider Gruppen nicht in Flugzelten
untergebracht, was die Gefahr signifikanter Änderungen des Befallsgrades birgt
(Greatti et al. 1992; Goodwin et al. 2006).
Noch nie zuvor wurde untersucht, ob es letale Auswirkungen
entomopathogener Pilze auf Larven von Honigbienen bereits ab dem L1-Stadium
gibt. Für die untersuchten Stämme im Larven-in-vitro-Assay kann ein solcher Einfluss
der Pilze auf Larven des L1- bis L5-Stadiums und auf Puppen von Apis mellifera
verneint werden. In allen Gruppen kam es innerhalb der ersten sechs bis neun Tage
zu einem Anstieg der kumulierten Mortalität auf etwa 15 Prozent, danach bis zum
Versuchsende nach 21 Tagen nur noch zu einer leichten Zunahme auf etwa 20
Prozent. Diese Rate dürfte den artifiziellen Bedingungen eines in vitro-Versuchs
geschuldet sein und deckt sich in der Kontrolle mit den beobachteten Mortalitätsraten
ähnlicher Versuche an Paenibacillus larvae (Genersch, mdl.). Unter den
nichtartifiziellen Bedingungen des Halbfreilandversuchs an Lecanicillium muscarium
kam es ebenfalls nicht zu Auffälligkeiten – weder für die Larvenstadien noch für das
Puppenstadium gab es signifikante Differenzen zwischen Pilz- und Kontrollgruppe.

Diskussion
116
Auch subletale Effekte entomopathogener Pilze, die unter Umständen
Ausschlusskriterium für ihren Einsatz sein können, sind in bisherigen
Untersuchungen noch nie berücksichtigt worden. Zur Klärung der Frage, ob es nicht-
letale Effekte auf adulte Bienen gab, die der in vitro-Versuch nicht erfassen konnte,
wurden mit Pilzsporen beziehungsweise nur mit einer Kontrolllösung behandelte
Gruppen in Bezug auf ihre Futteraufnahme pro Zeiteinheit als mögliches Indiz zum
Beispiel einer Schädigung des Darmkanals untersucht; in diesem Zusammenhang
wurde auch analysiert, ob das Gewicht der Honigblase über den
Gesamtversuchszeitraum Schwankungen unterworfen war. Klassische PER-
Versuche müssen darauf keine Rücksicht nehmen: Hier werden die Bienen den
Völkern wenige Stunden vor Versuchsbeginn entnommen und sind nach kurzer Zeit
hungrig, was Voraussetzung für eine erfolgreiche Teilnahme der Individuen am
Versuch ist (Bittermann et al. 1983). Bei über einen Versuchszeitraum von 12 Tagen
je Durchgang gekäfigten Bienen aber ist der Sättigungszustand der ad libitum mit
Futter versorgten Tiere nicht kalkulierbar; Langzeit-PER-Versuche mit gekäfigten
Bienen wurden nie zuvor etabliert und in Vorserien zeigte sich, dass es starke
Schwankungen bezüglich der nötigen Dauer der Hungerphase gab: Beobachtet
wurden Hungerphasen von mehr als 12 Stunden, die nötig waren, um Bienen zur
Versuchsteilnahme zu bewegen, aber auch eine Mortalitätsrate von über 50 Prozent
nach einer Hungerphase von nur einer halben Stunde.
Die Ergebnisse zeigen deutlich das signifikant abnehmende
Honigblasengewicht von etwa 10 mg auf etwa 2 mg während des
Versuchszeitraums. Dieses Ergebnis korreliert mit der Beobachtung, dass gekäfigte
Bienen zumindest während des Versuchszeitraums nicht defäkieren und die Menge
retenierter Fäzes permanent zunimmt – offensichtlich auf Kosten des zur Verfügung
stehenden Honigblasenvolumens. Die Replizierbarkeit der Ergebnisse machte in
Kombination mit den Ergebnissen der Messung des Futterverbrauchs ein
passgenaues Timing des Beginns des Futterentzugs möglich, so dass eine
ausreichend große Anzahl teilnahmefähiger Bienen ohne zu große
Präversuchsverluste zur Verfügung stand, und eignet sich damit als Grundlage für
zukünftige PER-Versuche an über lange Zeiträume gekäfigten Bienen.
Eine signifikante Änderung der Futteraufnahme konnte nicht festgestellt
werden: Sie lag über den gesamten Untersuchungszeitraum konstant bei etwa 0,002

Diskussion
117
ml pro Stunde und Biene – dies kann als Indiz dafür gewertet werden, dass es durch
die Applikation der Pilzsporen nicht zu Schädigungen des Verdauungstraktes kam.
Der PER-Versuch sollte klären, ob es zu einer Beeinträchtigung des
Lernvermögens durch Lecanicillium muscarium kam; dieser Aspekt ist gerade bei
Honigbienen besonders wichtig: Arbeiterinnen orientieren sich während des
Fouragierens in einem Umkreis von im Extremfall bis zu zehn Kilometern an
himmlischen und terrestrischen Merkmalen, sie sind in diesem Zusammenhang zu
einer hochdifferenzierten Mustererkennung fähig, verfügen über eine präzise
endogene Chronometrie und eine sensible olfaktorische Wahrnehmung, die auch
zentrales Element der Verwandtenerkennung ist (Lindauer 1963; Dyer et al. 1981;
Arnold et al. 1996; Breed 1998; Moore et al. 1998; Dyer et al. 2008). Eine
Beeinträchtigung dieser Fähigkeit durch ein exogenes Agens könnte einen
erheblichen Einfluss auf das Funktionieren der Summe der Einzeltiere in der
komplexen Sozialstruktur eines Bienenvolkes haben. Angesichts der zahlreichen, am
assoziativen Duftlernen von Honigbienen beteiligten Strukturen kann bereits die
Beeinträchtigung einer Komponente gravierende Folgen haben (Erber et al. 1980;
Hammer et al. 1998). So ist für die systemischen Insektizide Imidacloprid und Fipronil
nachgewiesen worden, dass sie auch weit unterhalb der letalen Konzentration die
Besuchsraten an Futterquellen reduzieren; für Imidacloprid konnte in PER-
Versuchen gezeigt werden, dass es die Fähigkeit zur Duftunterscheidung
beeinträchtigt (Colin et al. 2004; Decourtye et al. 2004). Es bindet nach
Untersuchungen von Nauen an nikotinische Acetylcholinrezeptoren der antennalen
Loben; da nACh-Rezeptoren mit Hilfe von Immunassays auch für andere Regionen
des Bienengehirns, die zentral für Lern- und Erinnerungsprozesse sind, wie die
Alphaloben und die Pilzkörper nachgewiesen wurden, sind prinzipiell auch hier
Beeinträchtigungen zu erwarten (Kreissl et al. 1989; Bicker 1999; Nauen et al. 2001).
Die Wirkung entomopathogener Pilze beruht wesentlich auf Toxinen wie den
nichtpeptidischen Agentien Oosporin und Bassianin, linearen Peptiden wie
Leuconistin und Efrapeptin sowie zyklischen Peptiden, namentlich Beauvericin und
Destruxin, die für diverse Spezies nachgewiesen worden sind (Khachatourians
1996). Zyklische Enniatine (C31-36H53-63N3O9) aus Fusarium spp. interagierten in
Versuchen mit GABAA-Rezeptoren in Rattengehirnen; für diesen Rezeptor wurden
Gemeinsamkeiten mit GABA-Rezeptoren von Invertebraten beschrieben, so dass
eine mögliche Wirkung pilzlicher Enniatine auch auf Insekten vermutet werden kann

Diskussion
118
(Anthony et al. 1993; Anke et al. 2002). Zunächst unbenommen der Beantwortung
der Frage, ob eine Beeinträchtigung des Lernvermögens durch Lecanicillium
muscarium auf der Wirkung ingestierter Pilztoxine, der Beeinflussung durch erst
innerhalb des Bienenkörpers generierte Metaboliten des Pilzstoffwechsels oder der
Induktion einer Immunantwort beruht, respektive, ob nur einer oder mehrere dieser
Faktoren einen Einfluss haben und welcher physiologische Mechanismus ihm
zugrunde liegt, zeigen die Ergebnisse des PER-Versuchs, dass es einen Einfluss
von Lecanicillium muscarium auf die Fähigkeit zur Duftunterscheidung bei Apis
mellifera gibt. Während die Lernkurve keine Auffälligkeiten zeigte, kam es innerhalb
der ersten zwei Tage nach Behandlung zu einer hochsignifikanten Beeinträchtigung
der Unterscheidungsfähigkeit bei Bienen, die mit Konidien von Lecanicillium
muscarium in Kontakt kamen. Die daraus resultierende Annahme, der Kontakt zu
Konidien könne eine subletale, negative Wirkung haben, wird durch den Befund des
Immunassays gestützt: Lecanicillium muscarium induzierte sowohl in Tests mit
Gramnegativen als auch in Tests mit Grampositiven Organismen eine neunfach
erhöhte Antwort des Immunsystems; das galt mit einer Einschränkung (Paecilomyces
fumosoroseus – Bacillus subtilis) ebenfalls für alle anderen untersuchten
entomopathogenen Pilze. Damit wird die Annahme einer durch Induktion einer
Immunantwort bedingten Reduktion des Lernvermögens des Individuums in den
ersten beiden Tagen nach Kontakt mit dem untersuchten Stamm von Lecanicillium
muscarium als bestätigt angesehen. Ob dieser Effekt tatsächlich Auswirkungen auf
der Ebene des Staats hat, ließe sich nur in Versuchen an einer sehr großen Anzahl
zuvor weitestgehend vereinheitlichter Völker untersuchen: Zu groß sind die
interkolonialen Varianzen zum Beispiel in Bezug auf die Sammelleistung, als dass
diese Fragestellung mit Versuchen an nur wenigen Völkern geklärt werden könnte.
Völlig unabhängig von der Beobachtung einer Beeinträchtigung des Lernvermögens
können natürlich zusätzlich auch pilzassoziierte Effekte, die ohne direkten Einfluss
auf die Sozialstruktur sind und zunächst nur zu einer physiologischen Schädigung
der Einzelbiene führen, durch Verringerung ihrer Vitalität, ohne ihren Tod zu
verursachen, negative Konsequenzen für den Staat haben. Immunassays als Test
für die Induktion einer Immunantwort nach Kontakt zu pathogenen Organismen sind
für Honigbienen etabliert; so wies Randolt nach Injektion von Escherichia coli 682
über einen ähnlichen Ansatz differierende Immunantworten von Bienenlarven und
Adultbienen nach (Randolt et al. 2008). Pilze initiieren nach Untersuchungen von

Diskussion
119
Hultmark und Stenbak an Drosophila melanogaster MEIGEN eine Immunantwort über
die Toll-Kaskade (Hultmark 2003; Stenbak et al. 2004). Für Honigbienen nimmt
Evans Gleiches an, nachdem seit der vollständigen Publikation des Genoms von
Apis mellifera etliche Homologe zu beteiligten Drosophilagenen gefunden wurden
(Evans et al. 2006; Weinstock et al. 2006). Inzwischen sind mit Abaecin, Apidaecin,
zwei Defensinen, Hymenoptaecin und einem Lysozym sechs antimikrobielle
Effektoren identifiziert, die bei Honigbienen nach Stimulation des Immunsystems
exprimiert werden.
Im Zusammenhang mit der Frage einer eventuellen Schädigung des
Lernvermögens nach Induktion einer Immunantwort durch entomopathogene Pilze ist
ein Versuch aber im Rahmen der vorliegenden Arbeit zum ersten Mal verwendet
worden und konnte zusammen mit den Ergebnissen der PER-Dressur zeigen, das
erst die inklusive Untersuchung subletaler Effekte mit der vorgestellten Methodik eine
Aussage über die potentielle Eignung entomopathogener Pilze, wie sie Kanga, Shaw
und Meikle aufgrund ihrer Versuche treffen, zulässt (Kanga et al. 2002, 2003, 2006;
Meikle et al. 2006, 2007, 2008a, 2008b, 2009; Shaw et al. 2002).
Die Ergebnisse des vierjährigen Monitorings an 215 Versuchsvölkern
verdeutlichen eine große Varianz des Parasitenbefalls zwischen den Völkern. Zwar
haben zahlreiche Faktoren einen Einfluss auf die Populationsdynamik von
Bienenvölkern und damit auch auf ihre Parasiten, was in Bezug auf unter
Freilandbedingungen kultivierte Völker insbesondere für die Verfügbarkeit von
Nahrungsquellen, die genetische Diversität innerhalb eines Volkes, aber auch die
mikroklimatischen Gegebenheiten am Standort gilt, trotzdem gestattet diese
Beobachtung die Vermutung, bisher unbekannte, interkolonial differierende
intrakoloniale Faktoren wie zum Beispiel die Abundanz von Hyperparasiten, könnten
einen nicht unerheblichen Einfluss auf die Abundanz und Pathogenität auch von
Varroa destructor haben. Die Korrelationsanalyse des Befalls von Apis mellifera-
Völkern mit Varroa destructor und dem Quantum der aus diesen Milben isolierbaren
Pilze ohne Berücksichtigung ihrer Gattungszugehörigkeit für die Jahre 2005 bis 2008
stützt diese These: Es treten sowohl Jahre mit einer vergleichsweise starken
Korrelation als auch praktisch beziehungslose Jahre auf. Ursächlich könnten zum
Beispiel die erwähnten mikroklimatischen beziehungsweise klimatischen
Bedingungen sein: Entomopathogene Pilze sind stenök in Bezug auf die relative
Luftfeuchte. Verticillium lecanii, Beauveria bassiana und Metarhizium anisopliae

Diskussion
120
beispielsweise zeigten in Experimenten von Ekbom und von Riba nur bei sehr hohen
Feuchtewerten eine Infektiösität in Bezug auf ihre Wirte (Ekbom 1981; Riba et al.
1984). Eine Reduktion der relativen Feuchte um etwa 20 Prozent für 80 Stunden
reduzierte in Versuchen von Drummond die Mortalität des vierten Larvenstadiums
von Trialeurodes vaporariorum unter dem Einfluss von Verticillium signifikant
(Drummond et al. 1987). Für die Sporulation entomopathogener Pilze ist eine hohe
Luftfeuchte sogar essentiell und Mycar, ein kommerzielles Produkt auf der Basis von
Hirsutella thompsonii, scheiterte am Markt, weil unter Feldbedingungen die relative
Feuchte zu gering war, um eine Keimung zu ermöglichen (McCoy 1986; Osborne et
al. 1992). Demgegenüber sinkt der Fortpflanzungserfolg von Varroa mit steigender
relativer Luftfeuchte: Eine Zunahme von 59 bis 68 Prozent auf 79 bis 85 Prozent
reduzierte in Versuchen von Kraus die Reproduktionsrate signifikant von 53 auf 2
Prozent (Kraus et al. 1997). Arbeiterinnen von Apis mellifera zeigen zwar gegenüber
zu hohen Feuchtewerten im Brutnest eine geringere Toleranz im Sinn der Induktion
einer regulierenden Antwort im Vergleich zu einer zu niedrigen Luftfeuchte,
gemessen am präferierten Wert, der bevorzugte Bereich liegt mit 90 - 95 Prozent
aber deutlich über dem Level, bei dem Varroa erfolgreich reproduziert (Doull 1976;
Ellis et al. 2008). Da Honigbienen aber auch schon bei einer relativen Luftfeuchte
von 55 Prozent im Brutnest in der Lage sind, erfolgreich zu reproduzieren, ergeben
sich – basierend auf der Annahme, die relative Luftfeuchte sei bestimmender Faktor
– zunächst zwei Erklärungsmuster für die Varianz zwischen den Jahren: Das Klima
des jeweiligen Jahres könnte unmittelbar die Abundanz der Milben und der Pilze
beeinflussen, ohne dass es einen ursächlichen Zusammenhang zwischen beiden
Größen gibt, oder dies mittelbar tun, indem in Jahren mit überdurchschnittlichen
relativen Feuchtewerten die Virulenz der Pilze positiv beeinflusst wird und damit die
Abundanz der Milben sinkt. Weitere, nicht offensichtliche Parameter könnten
stattdessen oder ergänzend natürlich ebenfalls eine Rolle spielen: Da unter den
gefundenen Arten ein großer Anteil saprober Arten auftrat, ist zum Beispiel auch
denkbar, dass diese Pilze von einer durch unbekannte Faktoren verursachten
erhöhten Mortalität innerhalb der Varroapopulationen als Opportunisten profitierten.
Dem steht allerdings entgegen, dass die Funde mutmaßlich milbenpathogener Pilze
in einem Jahr geringen Varroabefalls und sämtlichst in Völkern mit einer individuell
geringen Parasitenlast erfolgten.

Diskussion
121
Die Funde von Simplicillium lamellicola und Meira geulakonigii sind nach den
Funden von Beauveria bassiana in Varroa durch Meikle, Steenberg und Garcia-
Fernández weltweit die bisher einzigen Nachweise pathogener Pilze in Varroa
(Meikle et al. 2006; Garcia-Fernández et al. 2008; Steenberg et al. 2010). Meikle
allerdings fand Beauveria bassiana auf nicht oberflächensterilisierten Milben, Garcia-
Fernández macht dazu keine Angaben – da Beauveria bassiana verbreitet auch aus
Bodenproben isoliert wird, ist die endogene Herkunft möglicherweise nicht sicher.
Die großen Sequenzhomologien des eigenen Isolats von Simplicillium
lamellicola zu den Isolaten von Ilyas und von Lee, insbesondere aber der Versuch
von Polar deuten auf eine mögliche Pathogenität für Varroa hin (Polar et al. 2005;
Lee et al. 2007; Ilyas et al. 2009). Polar fand für Simplicillium lamellicola, der aus
einer unidentifizierten Milbe isoliert wurde, in Tests gegen Boophilus microplus eine
Reduktion der durchschnittlichen Lebensdauer des Wirts um 40 Prozent; Larven von
Boophilus blieben durch den Pilz unbeeinflusst.
Das in dieser Arbeit gefundene Isolat von Meira geulakonigii ist der weltweit
zweite Fund dieses Pilzes in Arthropoden und der dritte nach der Erstbeschreibung
(Boekhout et al. 2003). Die genaue Stellung der Art ist unklar; sie wird von Hibbett
den Exobasidiomycetes zugeordnet (Hibbett et al. 2007). Der Großteil der Vertreter
der Exobasidiomycetes kommt in einer saproben, haploiden Phase und einer
dikaryotischen, parasitischen Phase vor (Begerow et al. 2006). In mehreren
molekularen Analysen konnte gezeigt werden, dass die Exobasidiomyceten einen
Teil der Ustilaginomyceten darstellen (Begerow ebd.). Die große Mehrheit der Arten
der Ustilaginomycotina sind Parasiten höherer Pflanzen, lediglich Malassezia
parasitiert auf Warmblütern. Arten der Gattung Meira nähmen als
Arthropodenparasiten damit eine Sonderstellung innerhalb der Ustilaginomycotina
ein, auch wenn Tanaka Meira argovae BOEKHOUT, GERSON AND SZTEIJNBERG auf
Bambusschösslingen fand und Meira nach phylogenetischen Analysen neben das
Schwestertaxon Dicellomyces stellt, das mit Dicellomyces gloeosporus OLIVE
ebenfalls einen Parasiten auf Bambusblättern enthält (Tanaka et al. 2008).
Begerow ordnet Meira innerhalb der Exobasidiomycetes den Exobasidiales
und dort der Familie Brachybasidiaceae mit den Gattungen
Brachybasidium
Dicellomyces
Exobasidiellum

Diskussion
122
Kordyana
Meira und
Proliferobasidium
zu (Begerow et al. 2006). Allerdings gilt auch für die Brachybasidiaceae, was bereits
für die Exobasidiomycetes galt: In Abhängigkeit von der Auswahl der in die
Untersuchung einbezogenen Spezies und der verwendeten Topologieanalyse
werden sie entweder als Mono- oder als Paraphylum angesprochen. Der Fund des
eigenen Isolats von Meira geulakonigii könnte aufgrund der spezifischen
Besonderheiten der Spezies damit einen wichtigen Beitrag zur Klärung der
phylogenetischen Beziehungen innerhalb der Ustilaginomycotina liefern.
Bisher untersuchte Vertreter der Brachybasidiaceae sind Phytoparasiten auf
Monokotyledonen – auch in dieser Hinsicht nähme Meira eine Sonderstellung ein,
fand Boekhout sein Isolat doch auf Citrus grandis (Boekhout et al. 2003).
In Studien zur möglichen evolutionären Entwicklung der Exobasidiales zeigten
Vertreter der Brachybasidiaceae, dass die komplexen Strukturen, mit deren Hilfe der
Wirt parasitiert wird, nicht innerhalb von Haustorien lokalisiert sind, sondern von
interzellulären Hyphen gebildet werden, die Wirtszellen anliegen (Begerow et al.
2002). Die Sporulation findet auf der Oberfläche der Wirte statt. Die hilaren
Appendizes der Basidiosporen sind bei Brachybasidium und Dicellomyces adaxial
orientiert; probasidiale Verdickungen sind immer vorhanden. Die phylogenetischen
Beziehungen der Subtaxa der Exobasidiales korrelieren mit denen ihrer Wirte, was
als Hinweis auf Cospeziation verstanden wird (Begerow, ebd.).
Die bisher untersuchten Vertreter der Ordnung sind dimorphisch und zeigen zyklisch
ein saprobiontisches, hefeähnliches Stadium sowie parasitisch lebende Myzele
(Bauer et al. 2000). Dieser Umstand könnte sowohl den Fund von Meira geulakonigii
in Varroa klären, als auch das Vorkommen von hyphenartigen Strukturen im
mikroskopischen Bild: Die saprobiontischen Phasen der Schwestertaxa
metabolisieren in diesem Stadium überwiegend Mono- und Disaccharide und damit
einfache Zucker, die zahlreich im Nektar von von Bienen aufgesuchten Blüten und in
der Folge im Honig vorkommen, so dass die saprobe Phase von Meira
möglicherweise in einigen Blüten oder im Stock persistiert (Begerow, mdl.). Die
hyphenähnlichen Strukturen könnten demnach als Bestandteil der parasitischen
Phase angesprochen werden, die sich als Residuen der Ausgangskultur in der
Reinkultur finden; eine Vergleichsmöglichkeit dazu ist aus anderen Quellen allerdings

Diskussion
123
nicht bekannt – bisher ist nur die Anamorphe von Meira identifiziert. Der Fund der
Teleomorphen könnte – wenn sich ein Parasitieren auf Varroa destructor bestätigte –
ein „missing link“ innerhalb der Ustilaginomycotina sein: Innerhalb dieser
Unterabteilung der Basidiomycota mit rund 1800 Arten gibt es bisher mit Malassezia
nur eine einzige Gattung, deren Anamorphe nicht auf Pflanzen sondern auf
homoiothermen Vertebraten parasitiert. Eine Beschreibung auch der Teleomorphen
eines Arthropodenparasiten, der sowohl auf Pflanzen als auch auf Tieren gefunden
wird, könnte demnach einen wichtigen Beitrag zur Klärung der Frage leisten, wie
innerhalb der Subdivisio ein Interregnumsprung in Bezug auf den Wirt möglich war.
Boekhout fand den Pilz ebenfalls in einer Milbe (Boekhout et al. 2003). Das von
Boekhout gefundene Isolat wurde von Paz gegen Eutetranychus orientalis KLEIN,
Panonychus citri MCGREGOR, Tetranychus cinnabarinus BOISDUVAL und Tetranychus
urticae (alle Tetranychidae), Phyllocoptruta oleivora (Eriophyidae) sowie den Nicht-
Zielorganismus Rhizoglyphus robini CLAPAREDE (Acaridae), und die Raubmilben
Phytoseiulus persimilis ATHIAS-HENRIOT, Iphiseius degenerans BERLESE und
Neoseiulus californicus MCGREGOR (alle Phytoseiidae), getestet; die Applikation von
Konidien reduzierte die Milbenpopulation aller Phytophagen mit Ausnahme von
Rhizoglyphus robini um 60 Prozent, die Applikation pilzlicher Metabolite um 86
Prozent, verglichen mit den Kontrollapplikationen (Paz et al. 2007a). Die Raubmilben
wurden, abgesehen von Iphiseius degenerans, nicht beeinträchtigt, was für eine
hohe Selektivität der untersuchten Spezies spricht. Paz´ Befund (ebd.), dass es mit
konventionellen mykologischen Methoden nicht möglich war, Meira-Isolate von
Citrus-Oberflächen zu gewinnen, legte den Verdacht nahe, es handele sich bei Meira
nicht um einen Epi- sondern um einen Endophyten. In Anlehnung an Stone wies Paz
daraufhin Meira mit klassischen Methoden, sowie rasterelektronenmikroskopisch und
im Rahmen einer quantitativen Real-Time-PCR im Endokarp der Früchte nach
(Stone et al. 2000). In einer weiteren Studie mit ähnlichen Ergebnissen konnte Paz
beobachten, dass Meira nicht in den Körper der Milben eingedrungen war, die ohne
physischen Kontakt zum Pilz gestorben waren; gleichwohl war Meira in der Lage, auf
den Milbenkadavern zu wachsen (Paz et al. 2007b). Sztejnberg untersuchte
ebenfalls Boekhouts Isolat und fand eine hochsignifikante antagonistische Wirkung
gegenüber Phyllocoptruta oleivora, Panonychus citri, Tetranychus cinnabarinus und
Sphaerothecafusca spec. (Sztejnberg et al. 2004). In Bezug auf Sphaerothecafusca
spec. allerdings könne es sich Sztejnberg zufolge lediglich um einen Sekundäreffekt

Diskussion
124
handeln, da die Milben mutmaßlich als Vektor fungieren. Gerson bestätigte diese
Effekte: Meira geulakonigii tötete in Versuchen 80 – 90 Prozent der Individuen aus
mehreren Spinnmilbenarten; auch die Beobachtung von Paz, dass die Applikation
pilzfreier Extrakte des Kulturmediums signifikant auf die Milbenpopulation wirkte, wird
bei Gerson bestätigt und stützt die Annahme, dass zumindest ein Teil der
pathogenen Wirkung auf Pilztoxine zurückzuführen ist (Gerson et al. 2008). Darüber
hinaus identifizierten quantitative RT-PCR-Untersuchungen Meira geulakonigii in
Blüten und Fruchtschalen von Citrus grandis – ebenda fand Yasuda Meira
geulakonigii und schloss, der Pilz sei verantwortlich für Fruchtschäden (Yasuda et al.
2007). Diese Annahme wird von Gerson nicht bestätigt, sondern im Gegenteil
aufgrund der Assoziation des Pilzes mit Phyllocoptruta oleivora geschlussfolgert, der
Pilz fungiere als „endophytischer Bodyguard“ auf Citrus grandis und möglicherweise
anderen Pflanzen – demnach stünde die Identifikation des eigentlichen Wirtes auf
Pyrus pyrifolia, wo Yasuda den Pilz fand, noch aus. Würde er gefunden, verwunderte
auch der Zusammenhang zwischen der Abundanz von Meira und den beobachteten
Fruchtschäden auf Pyrus nicht, die Yasuda Meira zuschrieb. Die Annahme, der Pilz
sei primär milbenassoziiert und damit als Hyperparasit nur sekundär auf der
Wirtspflanze zu finden, wird durch die vorliegenden Ergebnisse sehr gut gestützt,
fehlt im Fall des Systems Apis mellifera – Varroa destructor – Meira geulakonigii der
pflanzliche Wirt doch völlig. Zumindest theoretisch kann aber auch eine tatsächlich
endophytische Herkunft des Isolats von Yasuda nicht ausgeschlossen werden. Bei
Meira handelt es sich, solange keine Einigung zur phylogenetischen Stellung
besteht, um eine Ordnung incertae sedis; für andere arthropodenpathogene Pilze,
unter anderem Metarhizium anisopliae und drei Verticillium-Spezies, konnte
Spatafora in einer Studie an sechs locis von insgesamt 69 Vertretern hauptsächlich
der Clavicipitaceae zeigen, dass es in mindestens fünf bis acht Fällen zu
Wirtswechseln auf Ebene des Regnums kam (Spatafora et al. 2007). Ausgehend von
einem basalen, tierpathogenen Vorläufer fanden drei bis fünf Wechsel zu Pilzen, ein
bis zwei Wechsel zu anderen Tieren und ein Wechsel zu Pflanzen statt. Die völlige
Übereinstimmung der publizierten Sequenzdaten von Boekhout und Yasuda spricht
allerdings dagegen (Boekhout et al. 2003; Yasuda et al. 2007). Da die Klassifizierung
des eigenen Isolats auf der Basis von Sequenzdaten eines loci erfolgte, dieses Isolat
sich aber von Boekhouts und Yasudas Isolaten – geringfügig – distinkt zeigt, wäre zu
klären, inwiefern ein Vergleich von Daten weiterer locorum sowie physiologischer

Diskussion
125
und morphologischer Daten eine genauere Abgrenzung des eigenen Isolats zuließe.
Denkbar ist, dass es sich beim eigenen Isolat um einen möglicherweise nicht nur
milben- sondern sogar Varroaspezifischen Pilz handeln könnte, der damit ein sehr
aussichtsreicher Kandidat für weitergehende Untersuchungen zu seiner
Einsetzbarkeit als Varroaantagonist wäre. Mit der in der vorliegenden Untersuchung
etablierten Methodik ist dafür eine Grundlage geschaffen worden. Zu prüfen wäre
zunächst in vitro die Hypothese, dass Meira geulakonigii ursächlich für den geringen
Milbenbefall der Völker ist, in denen der Pilz vorkommt, respektive, ob Meira keinen
oder einen vernachlässigbar geringen negativen Einfluss auf alle Stadien von Apis
mellifera hat. Als nächster Schritt schlösse sich eine Untersuchung im
Halbfreilandversuch an, um die artifiziellen Bedingungen von in vitro-Versuchen an
Bienen als Einflussfaktoren auszuschließen. Nicht zuletzt wäre ein Test des
Einflusses auf Nicht-Zielorganismen obligat: Der weitestgehend unkontrollierte Ex-
und Import von Honigbienen, und damit auch von Varroa, bedingt immer auch die
Gefahr der Verschleppung anderer Organismen; in Europa könnte Meira allochthon
sein, so dass eine Abschätzung des Risikopotentials bei einer Freisetzung
angebracht wäre. Vorausgesetzt, Meira erwiese sich als geeignetes Kontrollagens
und ungefährlich für andere Organismen, stünde, wenn der Pilz im technischen
Maßstab kultivierbar ist, ein Antagonist zur Verfügung, der zumindest einen Teil der
derzeitigen Probleme in der Kontrolle von Varroa destructor lösen würde.

Zusammenfassung
126
5 Zusammenfassung
Vor etwa 60 Jahren fand in Ostasien ein Wirtswechsel der ektoparasitischen
Milbe Varroa destructor von der östlichen Honigbiene Apis cerana auf die
ursprünglich nicht sympatrisch vorkommende westliche Honigbiene Apis mellifera
nach zuvor erfolgter weltweiter, anthropogen bedingter Verbreitung von Apis
mellifera als Nutztier statt. Unter dem Einfluss des präadaptierten Parasiten kommt
es nahezu im gesamten Verbreitungsgebiet von Apis mellifera angesichts
unbefriedigender verfügbarer Bekämpfungsstrategien zu massiven Verlusten von
Bienenvölkern und damit zu gravierenden wirtschaftlichen Schäden durch Ausfall
dieses wichtigen Bestäubers in landwirtschaftlichen Kulturen entomophiler
Nutzpflanzen. Seit der Jahrtausendwende wurde deshalb auch versucht,
Hyperparasiten, und zwar vor allem Pilze, als einsetzbare Antagonisten von Varroa
zu finden. Die bisher vorliegenden Studien konzentrierten sich vor allem auf die
bereits gegen andere Arthropoden erfolgreich eingesetzten Pilze Lecanicillium
muscarium, Metarhizium anisopliae, Beauveria bassiana und Paecilomyces
fumosoroseus. Während einige Untersuchungen die getesteten Stämme als geeignet
klassifizierten, fanden andere, zum Teil auch vom gleichen Autor und mit demselben
Stamm durchgeführt, keine Effekte. Ursächlich dafür ist zum einen die große Varianz
in der Pathogenität der sehr pleomorphen Spezies, die die Vergleichbarkeit der an
verschiedenen Stämmen gewonnenen Ergebnisse erschwert, zum anderen die
Heterogenität der zum Einsatz gekommenen und zum Teil diskutierbaren
Untersuchungsmethodiken.
Die vorliegende Arbeit schafft eine valide, reproduzierbare Basis, um in Frage
kommende Pilze unter vergleichbaren Bedingungen zu testen. Sie führt mit in vitro-
Untersuchungen an Larven ab dem L1-Stadium, PER-Tests zur Identifikation
subletaler Effekte und einem Immunassay drei neue, bisher in diesem
Zusammenhang nicht zum Einsatz gekommene Methoden ein, die die Beantwortung
von mit bisherigen Analysen nicht zugänglichen offenen Fragen erlauben.
Die in der vorliegenden Arbeit untersuchten Stämme von Lecanicillium
muscarium, Metarhizium anisopliae, Beauveria bassiana und Paecilomyces
fumosoroseus zeigten sich sowohl in vitro als auch im Halbfreilandversuch
gegenüber Varroa destructor wirkungslos. Larvenstadien und Puppenstadium von
Apis mellifera wurden nicht beeinträchtigt; Lecanicillium muscarium und Beauveria

Zusammenfassung
127
bassiana schädigten in vitro allerdings adulte Bienen signifikant. Zudem wurden für
Lecanicillium muscarium negative subletale Effekte auf Apis mellifera nachgewiesen:
Der Pilz reduzierte die Fähigkeit zu assoziativem Lernen an den ersten beiden Tagen
nach Kontakt mit ihm. Diese Beobachtung korreliert mit dem Nachweis einer
Induktion des Immunsystems durch einen sensitiven Test an Bacillus subtilis und
Escherichia coli: Mit nur einer Ausnahme (Paecilomyces fumosoroseus/Bacillus
subtilis) kam es zu einer starken Antwort, die als Beleg einer physiologischen
Belastung durch diese Pilze angesehen wird.
Im Rahmen der vorliegenden Arbeit sollten auch spezifische, acaripathogene
Pilze gesucht werden, die als Antagonisten einsetzbar sein könnten. Die dabei zum
Einsatz gekommene Methodik ist ebenfalls erstmals in diesem Zusammenhang
angewandt worden und führte mit dem Fund von Simplicillium lamellicola und Meira
geulakonigii zum Erfolg. Die Pilze sind nach den Funden des unspezifischen Pilzes
Beauveria bassiana durch Meikle, der Beauveria aber in unsterilisierten Milben fand,
und Steenberg die ersten Funde entomo- beziehungsweise potentiell
acaripathogener Pilze in Varroa (Meikle et al. 2006; Steenberg et al. 2010). Das in
der vorliegenden Untersuchung gefundene Isolat von Meira geulakonigii ist darüber
hinaus der weltweit zweite Fund dieses Pilzes in Arthropoden und der dritte nach der
Erstbeschreibung. Bei Meira handelt es sich um eine Ordnung incertae sedis. Als
Arthropodenparasit wäre Meira geulakonigii eine große Ausnahme innerhalb der
Ustilaginomycotina und könnte einen wichtigen Beitrag zum Verständnis von
Cospeziationsprozessen beziehungsweise Wirtswechseln zwischen höheren Taxa
liefern. Möglich ist, dass Meira geulakonigii in einem saprobiontischen, hefeähnlichen
Stadium in Blüten oder in Bienenvölkern vorkommt, in welchem einfache Zucker
metabolisiert werden, und in einem hyphenähnlichen Stadium als Milbenparasit – die
Typusbeschreibung stammt von einem Isolat aus Phyllocoptruta oleivora, das später
auch als Endophyt in deren Wirt Citrus grandis gefunden wurde. Für die Möglichkeit
einer spezifischen Acaripathogenität des eigenen Isolats spricht, dass es in einem
Apis mellifera-Volk gefunden wurde, das zu einem Cluster von Völkern gehörte, die
in Korrelationsuntersuchungen der Varroaabundanz und der Zahl der aus Varroa
isolierbaren Pilze einen deutlichen Zusammenhang von geringem Varroabefall und
einem starken Pilzvorkommen zeigten. Ein aussichtsreicher Kandidat für einen
einsetzbaren Varroaantagonisten könnte mit dem in der vorliegenden Arbeit
beschriebenen Isolat von Meira geulakonigii damit gefunden sein.

Summary
128
6 Summary
60 years ago the ectoparasitic mite Varroa destructor originating from the
eastern honey bee Apis cerana began to infest colonies of the western honey bee
Apis mellifera after its global spread by man. Preadapted, Varroa caused serious
economic damage, since Apis mellifera is an important pollinator of many crops.
Currently available counterstrategies are not satisfying: synthetic acaricides
lead to residues in honey and wax and triggered resistance in Varroa while
biotechnical methods are time-intensive. The use of organic acids is of limited
success if not applied in an exact manner. Great efforts have been made to select
Varroa-resistent honeybees but no success has been reported to date.
Some years ago one started to test already known unspecific
entomopathogenic fungi against Varroa, namely Lecanicillium muscarium,
Metarhizium anisopliae, Beauveria bassiana and Paecilomyces fumosoroseus.
Results of studies are contradictory: while some reported a reduction of mites, others
did not find any effect. On the one hand this may be caused by the use of different
strains of pleomorphic fungi, on the other hand studies differ in methods used and
sometimes lack essential techniques like standardization of bee material.
The examinations presented here create a reproducible basis for further
analyses. Three techniques not yet used in honey bee-fungus interaction research
are introduced to investigate potential fungus-induced effects on honey bee larvae
and pupae, learning ability of adult bees and induction of immune system response
of adult bees by contact with fungi.
The strains analyzed in this study did not have any effect on Varroa, neither in
vitro nor in semi-field studies, but harmed adult bees in two cases (Lecanicillium
muscarium, Beauveria bassiana). Furthermore, Lecanicillium muscarium turned out
to reduce learning ability of adult bees significantly within the first two days after
application and induced a strong reaction of the immune system.
A second aim of this study was to find specific acaripathogenic fungi, again
using a new approach. With Simplicillium lamellicola and Meira geulakonigii two new
fungi never found before in Varroa have been isolated. The own isolation of Meira
geulakonigii is the third worldwide after Boekhout´s first description of Meira found in
Israel (Boekhout et al. 2003). Still classified as an order incertae sedis, Meira as a
member of Ustilaginomycotina is assumed to live in a saprobiontic, yeast-like phase

Summary
129
und a parasitic phase with hyphae. While Simplicillium lamellicola is known to be
entomopathogenic, Meira geulakonigii has only been found in mites or in plant
tissues and therefore maybe is solely acaripathogenic. This hypothesis is supported
by correlation analyses between 2005 and 2008: For four years dependence is
shown between occurrence of fungi and frequency of mites in bee hives. Both
Simplicillium and Meira have only been found in a year with such a dependence and
in hives with many fungi while bees and cells are less parasitized by mites.
Therefore, the present results suggest that Meira geulakonigii has the potential to be
the longed for biocontrol agent to control the honey bee devastating mite Varroa
destructor.

Literatur
130
7 Literatur Aliano, N.P. and Ellis, M.D. (2005). A strategy for using powdered sugar to reduce
Varroa populations in honey bee colonies. Journal of Apicultural Research 44: 54-57.
Allen, J.C., Yang, Y., Knapp, J.L., Stansley, P.A., Rosen, D., Bennet, F.D. and
Capinera, J.L. (1994). The citrus rust mite story: A modelling approach to a fruit-mite-pathogen system. In: Rosen, D., Bennett, F.D. and Capinera, J.L. (ed) Pest management in the subtropics: Biological control - a Florida perspective. Andover, Intercept Limited: 619-639.
Altre, J.A., Vandenberg, J.D. and Cantone, F.A. (1999). Pathogenicity of
Paecilomyces fumosoroseus isolates to diamondback moth, Plutella xylostella: Correlation with spore size, germination speed, and attachment to cuticle. Journal of Invertebrate Pathology 73: 332-338.
Anderson, D.L. and Trueman, J.W. (2000). Varroa jacobsoni (Acari: Varroidae) is
more than one species. Experimental and Applied Acarology 24: 165-189. Andrews, J.H. and Harris, R.F. (1997). Dormancy, germination, growth, sporulation,
and dispersal. In: Esser, K. and Lemke, P.A. (ed) The Mycota IV. Environmental and microbial relationships. New York, Springer-Verlag: 3-13.
Anke, H. and Sterner, O. (2002). Insecticidal and nematicidal metabolites from fungi.
In: Esser, K. and Bennet, J.W. (ed) The Mycota X. Industrial applications. New York, Springer-Verlag: 110-127.
Anthony, N.M., Harrison, J.B. and Sattelle, D.B. (1993). GABA receptor molecules
of insects. Experientia Supplementum: Comparative Molecular Neurobiology 63: 172-209.
Arnold, G., Quenet, B., Cornuet, J.M., Masson, C., De Schepper, B., Estoup, A.
and Gasqui, P. (1996). Kin recognition in honeybees. Nature 379: 498. Aumeier, P., Rosenkranz, P. and Francke, W. (2002). Cuticular volatiles, attractivity
of worker larvae and invasion of brood cells by Varroa mites. A comparison of africanized and european honey bees. Chemoecology 12: 65-75.
Backhaus, K. (2008). Multivariate Analysemethoden. Berlin, Springer Verlag. Ball, B.V. (1989). Varroa jacobsoni as a virus vector. In: Cavalloro, R. (ed) Present
status of Varroatosis in Europe and progress in the Varroa mite control. Commission of the european communities, Luxembourg: 241-244.
Bauer, R., Begerow, D., Oberwinkler, F., Piepenbrink, M., Berbee, M.L. (2000).
Ustilaginomycetes. In: Esser, K. and Lemke, P.A. (ed) The Mycota VII, Part B. Systematics and evolution. New York, Springer-Verlag: 57-83.

Literatur
131
Beauvais, A., Latgé, J.P., Vey, A. and Prevost, M.C. (1989). The role of surface components of the entomopathogenic fungus Entomophaga aulicae in the cellular immune response of Galleria mellonella (Lepidoptera). Journal of General Microbiology 135: 489-498.
Beetsma, J. (1994). The Varroa mite, a devasting parasite of western honeybees
and an economic threat to beekeeping. Outlook on Agriculture 23: 169-175. Begerow, D., Bauer, R. and Oberwinkler, F. (2002). The Exobasidiales: An
evolutionary hypothesis. Mycological Progress 1: 187-199. Begerow, D., Stoll, M. and Bauer, R. (2006). A phylogenetic hypothesis of
Ustilaginomycotina based on multiple gene analyses and morphological data. Mycologia 98: 906-916.
Bicker, G. (1999). Histochemistry of classical neurotransmitters in antennal lobes
and mushroom bodies of the honeybee. Microscopy Research and Technique 45: 174-183.
Bienefeld, K. and Arnold, G. (2007). Evidence for significant intra-colonial variation
in uncapping of Varroa-infested brood cells in Apis mellifera. Proceedings of the 40th Apimondia International Congress, Melbourne: 144-145.
Bittencourt, V.R.E.P., Massard, C.L. and De Lima, A.F. (1994a). Action of the
fungus Metarhizium anisopliae on eggs and larvae of the tick Boophilus microplus. Revista Universidade Rural. Serie Ciencias da Vida 16: 41-47.
Bittencourt, V.R.E.P., Massard, C.L. and De Lima, A.F. (1994b). Action of the
fungus Metarhizium anisopliae on the free living phase of the life cycle of Boophilus microplus. Revista Universidade Rural. Serie Ciencias da Vida 16: 49-55.
Bittermann, A.E., Menzel, R., Fietz, A. and Schäfer, S. (1983). Classical
conditioning of proboscis extension in honeybees (Apis mellifera). Journal of Comparative Psychology 97: 107-119.
Boecking, O. and Genersch, E. (2008). Varroosis – the ongoing crisis in bee
keeping. Journal für Verbraucherschutz und Lebensmittelsicherheit 3: 221-228.
Boecking, O. and Spivak, M. (1999). Behavioral defenses of honey bees against
Varroa jacobsoni Oud. Apidologie 30: 141-185. Boekhout, T., Theelen, B., Houbraken, J., Robert, V., Scorzetti, G., Gafni, A.,
Gerson, U. and Sztejnberg, A. (2003). Novel anamorphic mite-associated fungi belonging to the Ustilaginomycetes: Meira geulakonigii gen. nov., sp. nov., Meira argovae sp. nov. and Acaromyces ingoldii gen. nov., sp. nov.. International Journal of Systematic and Evolutionary Microbiology 53: 1655-1664.

Literatur
132
Bölker, M. (2002). Signal transduction pathways in phytopathogenic fungi. In: Esser, K. and Bennett, J.W. (ed) The Mycota XI. Agricultural applications. New York, Springer-Verlag: 273-288.
Boot, W.J., Schoenmaker, J., Calis, J.N.M. and Beetsma, J. (1995). Invasion of
Varroa jacobsoni into drone brood cells of the honey bee, Apis mellifera. Apidologie 26: 109-118.
Boot, W.J., Calis, J.N.M., Beetsma, J., Hai, D.M., Lan, N.K., Toan, T.V., Trung,
L.Q. and Minh, N.H. (1999). Natural selection of Varroa jacobsoni explains the different reproductive strategies in colonies of Apis cerana and Apis mellifera. Experimental and Applied Acarology 23: 133-144.
Breed, M. D. (1998). Recognition pheromones of the honey bee. The chemistry of
nestmate recognition. BioScience 48: 463-470. Brobyn, P.J. and Wilding, N. (1977). Invasive and developmental processes of
Entomophthora species infecting aphids. Transactions of the British Mycological Society 69: 349-366.
Bruns, T.D. and Gardes, M. (1993). ITS primers with enhanced specificity for
Basidiomycetes - application to the identification of mycorrhizae and rusts. Molecular Ecology 2: 113-118.
Büchler, R., Garrido, C., Ehrhardt, K. and Bienefeld, K. (2008). Selection for
Varroa tolerance: Concept and results of a long-term selection project. Apidologie 39: 598.
Calderone, N. W. (1999). Evaluation of formic acid and a thymol-based blend of
natural products for the fall control of Varroa jacobsoni (Acari: Varroidae) in colonies of Apis mellifera (Hymenoptera: Apidae). Journal of Economic Entomology 92: 253-260.
Chandler, D., Davidson, G., Pell, J.K., Ball, B.V., Shaw, K. and Sunderland, K.D.
(2000). Fungal biocontrol of acari. Biocontrol Science and Technology 10: 357-384.
Chiroudi, E., Rosenkranz, P. and Aumeier, P. (1997). Altersabhängige und
rassenspezifische Duftstoffmuster bei Arbeiterinnen der Honigbiene (Apis mellifera). Mitteilungen der deutschen Gesellschaft für Allgemeine und Angewandte Entomologie 11: 179-182.
Claydon, N. and Grove, J.F. (1982). Insecticidal secondary metabolic products from
the entomogenous fungus Verticillium lecanii. Journal of Invertebrate Pathology 40: 413-418.
Colin, M.E., Bonmatin, J.M., Moineau, I., Gaimon, C., Brun, S. and Vermandere,
J.P. (2004). A method to quantify and analyze the foraging activity of honey bees: Relevance to the sublethal effects induced by systemic insecticides. Archives of Environmental Contamination and Toxicology 47: 387-395.

Literatur
133
Correa-Marques, M.H., De Jong, D., Rosenkranz, P. and Goncalves, L.S. (2002). Varroa-tolerant italian honey bees introduced from Brazil were not more efficient in defending themselves against the mite Varroa destructor than carniolan bees in Germany. Genetics and Molecular Research 2: 1-6.
Crane, E. (1990). Bees and beekeeping. Science, practice and world resources. New
York, Cornell University Press. de Faria, M.R. and Wraight, S.P. (2001). Biological control of Bemisia tabaci with
fungi. Crop Protection 20: 767-778. de Faria, M.R. and Wraight, S.P. (2007). Mycoinsecticides und mycoacaricides: A
comprehensive list with worldwide coverage and international classification of formulation types. Biological Control 43: 237-256.
De Guzman, L.I. and Rinderer, T.E. (1999). Identification and comparison of Varroa
species infesting honey bees. Apidologie 30: 85-95. Decourtye, A., Lacassie, E. and Pham-Delegue, M.H. (2003). Learning
performances of honeybees (Apis mellifera L.) are differentially affected by imidacloprid according to the season. Journal of Chemical Technology and Biotechnology 59: 269-278.
Decourtye, A., Armengaud, C., Renou, M., Devillers, J., Cluzeau, S., Gauthier, M.
and Pham-Delegue, M.H. (2004). Imidacloprid impairs memory and brain metabolism in the honeybee (Apis mellifera L.). Pesticide Biochemistry and Physiology 78: 83-92.
Donzé, G. and Guerin, P.M. (1994). Behavioral attributes and parental care of
Varroa mites parasitizing honeybee brood. Behavioural Ecology and Sociobiology 34: 305-319.
Doull, K.M. (1976). The effect of different humidities on the hatching of eggs of
honeybees. Apidologie 7: 61-66. Drummond, J., Heale, J.B. and Gillespie, A.T. (1987). Germination and effect of
reduced humidity on expression of pathogenicity in Verticillium lecanii against the glasshouse whitefly Trialeurodes vaporariorum. Annals of Applied Biology 111: 193-201.
Duay, P., De Jong, D. and Engels, W. (2002). Decreased flight performance and
sperm production in drones of the honey bee (Apis mellifera) slightly infested by Varroa destructor mites during pupal development. Genetics and Molecular Research 1: 227-232.
Dustmann, J.H. and von der Ohe, W. (1988). Einfluss von Kälteeinbrüchen auf die
Frühjahrsentwicklung von Bienenvölkern (Apis mellifera L.). Apidologie 19: 245-254.

Literatur
134
Dyer, A.G., Rosa, M.G. and Reser, D.H. (2008). Honeybees can recognise images of complex natural scenes for use as potential landmarks. Journal of Experimental Biology 211: 1180-1186.
Dyer, F. and Gould, J. (1981). Honey bee orientation: A backup system for cloudy
days. Science 214: 1041-1042. Ekbom, B.S. (1981). Humidity requirements and storage of the entomopathogenic
fungus (Verticillium lecanii) for use in greenhouses. Annals Entomologici Fennici 47: 61-62.
Ellis, A.M., Hayes, G.W. and Ellis, J.D. (2009). The efficacy of dusting honey bee
colonies with powdered sugar to reduce Varroa mite populations. Journal of Apicultural Research 48: 72-76.
Ellis, M.B., Nicolson, S.W., Crewe, R. and Dietemann, V. (2008). Hygropreference
and brood care in the honeybee (Apis mellifera). Journal of Insect Physiology 54: 1516-1521.
Erber, J., Masuhr, T. and Menzel, R. (1980). Localization of short-term memory in
the brain of the bee Apis mellifera. Physiological Entomology 5: 343-358. Evans, J.D., Aronstein, K., Chen, Y.P., Hetru, C., Imler, J.L., Jiang, H., Kanost,
M., Thompson, G.J., Zou, Z. and Hultmark, D. (2006). Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Molecular Biology 15: 645-656.
Ferron, P. (1978). Biological control of insect pests by entomogenous fungi. Annual
Review of Entomology 23: 409-442. Gallai, N., Salles, J.N., Settele, J. and Vaissière, B.E. (2008). Economic valuation
of the vulnerability of world agriculture confronted with pollinator decline. Ecological Economics 68: 810-821.
Garcia-Fernández, P., Santiago-Alvarez, C. and Quesada-Moraga, E. (2008).
Pathogenicity and thermal biology of mitosporic fungi as potential control agents of Varroa destructor (Acari: Mesostigmata), an ectoparasitic mite of the honey bee, Apis mellifera (Hymenoptera: Apidae). Apidologie 39: 662-673.
Gams, W. and Zare, R. (2001). A revision of Verticillium section Prostrata. III:
Genetic classification. Nova Hedwigia 72: 329-337. Garrido, C. and Rosenkranz, P. (2004). Volatiles of the honey bee larva initiate
oogenesis in the parasitic mite Varroa destructor. Chemoecology 14: 193-197. Gerritsen, L. and Cornelissen, B. (2006). Biological control of Varroa destructor by
fungi. Proceedings of the Section Experimental and Applied Entomology of the Netherlands Entomological Society 17: 125-132.

Literatur
135
Gerson, U., Gafni, A., Paz, Z. and Sztejnberg, A. (2008). A tale of three acaropathogenic fungi in israel: Hirsutella, Meira and Acaromyces. Experimental and Applied Acarology 46: 183-194.
Gindin, G., Samish, M., Zangi, G., Mishoutchenko, A. and Glazer, I. (2002). The
susceptibility of different species and stages of ticks to entomopathogenic fungi. Experimental and Applied Acarology 28: 283-288.
Gisder, S., Aumeier, P. and Genersch, E. (2009). Deformed wing virus: Replication
and viral load in mites (Varroa destructor). Journal of General Virology 90: 463-467.
Goodwin, R.M., Taylor, M.A., McBrydie, H.M. and Cox, H.M. (2006). Drift of Varroa
destructor-infested worker honey bees to neighbouring colonies. Journal of Apicultural Research 45: 155-156.
Greatti, M., Milani, N. and Nazzi, F. (1992). Reinfestation of an acaricide-treated
apiary by Varroa jacobsoni Oud. Experimental and Applied Acarology 16: 279-286.
Guzman-Novoa, E., Vandame, R. and Arechavaleta, M.E. (1999). Susceptibility of
european and africanized honeybees (Apis mellifera L.) to Varroa jacobsoni Oud. in Mexico. Apidologie 30: 173-182.
Hammer, M. and Menzel, R. (1998). Multiple sites of associative odor learning as
revealed by local brain microinjectons of octopamine in honeybees. Learning and Memory 5: 146-156.
Harris, J.W., Harbo, J.R., Villa, J.D. and Danka, R.G. (2003). Variable population
growth of Varroa destructor (Mesostigmata: Varroidae) in colonies of honey bees (Hymenoptera: Apidae) during a 10-year period. Environmental Entomology 32: 1305-1312.
Hawksworth, D.L., Kirk, P.M., Sutton, B.C. and Pegeler, D.N. (1995). Dictionary of
the fungi. Wallingford, UK, CAB International. Hibbett, D.S., Binder, M., Bischoff, J.F., Blackwell, M., Cannon, P.F., Eriksson,
O.E. and Begerow, D. (2007). A higher-level phylogenetic classification of the fungi. Mycological research 111: 509-547.
Hultmark, D. (2003). Drosophila immunity: Paths and patterns. Current Opinion in
Immunology 15: 12-19. Ilyas, M., Kurihara, Y., Sukarno, N., Yuniarti, E., Mangunwardoyo, W.,
Sawaswati, R., Park, J.Y., Sjamsuridzal, W., Inaba, S., Widyastuti, Y., Ando, K. and Harayama, S. (2009) "Simplicillium species from Indonesia and Japan." EMBL Nucleotide database: http://srs.ebi.ac.uk/srsbin/cgi-bin/wgetz?-e+[EMBL:AB378533]+-newId.

Literatur
136
James, R.R., Hayes, G. and Leland, J.E. (2006). Field trials on the microbial control of Varroa with the fungus Metarhizium anisopliae. American Bee Journal 146: 968-972.
Kalsbeek, V., Frandsen, F. and Steenberg, T. (1995). Entomopathogenic fungi
associated with Ixodes ricinus ticks. Experimental and Applied Acarology 19: 45-51.
Kanga, L.H.B., James, R.R. and Boucias, D.G. (2002). Hirsutella thompsonii and
Metarhizium anisopliae as potential microbial control agents of Varroa destructor (Acari: Varroidae), a honey bee parasite. Journal of Invertebrate Pathology 81: 175-184.
Kanga, L.H.B., Jones, W.A. and James, R.R. (2003). Field trials using the fungal
pathogen, Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) to control the ectoparasitic mite, Varroa destructor (Acari: Varroidae) in honey bee, Apis mellifera (Hymenoptera: Apidae) colonies. Journal of Economic Entomology 96: 1091-1099.
Kanga, L.H.B., Jones, W.A. and James, R.R. (2005). Enlisting fungi to protect the
honey bee. Biologist 52: 88-94. Kanga, L.H.B., Jones, W.A. and Gracia, C. (2006). Efficacy of strips coated with
Metarhizium anisopliae for control of Varroa destructor (Acari: Varroidae) in honey bee colonies in Texas and Florida. Experimental and Applied Acarology 40: 249-258.
Khachatourians, G.G. (1996). Biochemistry and molecular biology of
entomopathogenic fungi. In: Esser, K. and Lemke, P.A. (ed) The Mycota VI: Human and animal relationships. New York, Springer-Verlag: 331-363.
Kirchner, W.H. (1987). Tradition im Bienenstaat - Kommunikation zwischen den
Imagines und der Brut der Honigbiene durch Vibrationssignale. Dissertation. Universität Würzburg, Fakultät für Biologie.
Kirchner, W.H. (1993). Vibrational signals in the tremble dance of the honeybee Apis
mellifera. Behavioral Ecology and Sociobiology 33: 169-172. Kirchner, W.H., Lindauer, M. and Michelsen, A. (1986). Vibrationssignale in der
Sprache der Bienen. Verhandlungen der Deutschen Zoologischen Gesellschaft 79: 411-412.
Koike, M., Sugimoto, M., Aiuchi, D., Nagao, H., Shinya, R., Tani, M. and
Kuramochi, K. (2007). Reclassification of japanese isolate of Verticillium lecanii to Lecanicillium spp.. Japan Journal of Applied Entomology 51: 234-237.
Kovac, H. and Crailsheim, K. (1988). Lifespan of Apis mellifera carnica Pollm.
infested by Varroa jacobsoni Oud. in relation to season and extent of infestation. Journal of Apicultural Research 27: 230-238.

Literatur
137
Kralj, J., Brockmann, A., Fuchs, S. and Tautz, J. (2007). The parasitic mite Varroa destructor affects non-associative learning in honey bee foragers, Apis mellifera L. Journal of comparative physiology. A: Neuroethology, sensory, neural, and behavioral physiology 193: 363-370.
Kraus, B. (1993). Preferences of Varroa jacobsoni for honey bees Apis mellifera L.
of different ages. Journal of Apicultural Research 32: 57-64. Kraus, B. and Velthuis, H.H.W. (1997). High humidity in the honey bee (Apis
mellifera L.) brood nest limits reproduction of the parasitic mite Varroa jacobsoni Oud.. Naturwissenschaften 84: 217-218.
Kreissl, S. and Bicker, G. (1989). Histochemistry of acetylcholinesterase and
immunocytochemistry of an acetylcholine receptor-like antigen in the brain of the honeybee. Journal of Comparative Neurology 286: 71-84.
Kuenen, L.P.S. and Calderone, N.W. (1997). Transfers of Varroa mites from newly
emerged bees: Preferences for age- and function-specific adult bees (Hymenoptera: Apidae). Journal of Insect Behavior 10: 213-228.
Lampedro, L. and Luis-Rosas, J. (1989). Selection of Hirsutella thompsonii (Fisher)
strains to fight the coconut mite Eriophyes guerreronis (Keifer). Revista Mexicana de Micologia 5: 225-231.
Le Conte, Y., de Vaublanc, G., Crauser, D., Jeanne, F., Rousselle, J.C. and
Bécard, J.M. (2007). Honey bee colonies that have survived Varroa destructor. Apidologie 38: 566-572.
Lee, M. and Park, H. (2007) "Pathogenicity of entomopathogenic fungi on Bemisia
tabaci." EMBL Nucleotide database: http://srs.ebi.ac.uk/srsbin/cgi-bin/wgetz?-e+[EMBL:EU284714]+-newId.
Leger, R.J., Goettel, M., Roberts, D.W. and Staples, R.C. (1991). Preprenetration
events during infection of host cuticle by Metarhizium anisopliae. Journal of Invertebrate Pathology 58: 168-179.
Leger, R.J., Cooper, R.M. and Charnley, A.K. (1993). Analysis of aminopeptidase
and dipeptidase from the entomopathogenic fungus Metarhizium anisopliae. Journal of Genetic Microbiology 139: 237-243.
Leger, R.J., Joshi, L., Bidochka, M.J. and Roberts, D.W. (1996). Characterisation
of chitinases from Metarhizium anisopliae, Metarhizium flavoviride and Beauveria bassiana, and ultrastructural localization of chitinase production during invasion of insect cuticle. Applied Environmental Microbiology 62: 907-912.
Leger, R.J., Joshi, L. and Roberts, D.W. (1997). Adaption of proteases and
carbohydrases of saprophytic, phytopathogenic and entomopathogenic fungi to the requirements of their ecological niches. Microbiology 143: 1983-1992.

Literatur
138
Levin, M.D. (1984). Value of bee pollination to United States agriculture. American Bee Journal 124: 184-186.
Lindauer, M. (1963). Time sense of bees. In: Moore, J.A. (ed) Proceedings of the
XVI International Congress of Zoology Washington: 351-359. Malsan, O., Kilian, M., Oerke, E. and Dehne, H.W. (2002). Oils for increased
efficacy of Metarhizium anisopliae to control whiteflies. Biocontrol Science and Technology 12: 337-348.
Martel, A.C., Zeggane, S., Aurieres, C., Drajnudel, P., Faucon, J.P. and Aubert,
M. (2007). Acaricide residues in honey and wax after treatment of honey bee colonies with Apivar or Asuntol 50. Apidologie 38: 534-544.
Martin, S.J. (1994a). Ontogenesis of the mite Varroa jacobsoni Oud. in worker brood
of the honey bee Apis mellifera L. under natural conditions. Experimental and Applied Acarology 18: 87-100.
Martin, S.J. (1994b). Ontogenesis of the mite Varroa jacobsoni Oud. in drone brood
of the honeybee Apis mellifera L. under natural conditions. Experimental and Applied Acarology 19: 199-210.
McCoy, C.W. (1981). Pest control by the fungus Hirsutella thompsonii. Microbial
control of pests and plant diseases 1970-1980. Burges. London, Academic Press: 499-512.
McCoy, C.W. (1986). Factors governing the efficacy of Hirsutella thompsonii in the
field. In: Samson, R.A., Vlak, J.M. and Peters, D. (ed) Fundamental and applied aspects of invertebrate pathology. Foundation of the IVth International Colloquium of Invertebrate Pathology, Wageningen: 171-174.
McNeely, J.A., Mooney, H.A., Neville, L.E., Schel, P. and Waage, J.K. (2001). A
global strategy on invasive alien species. Gland, Cambridge, IUCN. Meikle, W.G., Mercadier, G., Girod, V., Derouane, F. and Jones, W.A. (2006).
Evaluation of Beauveria bassiana (Balsamo) Vuillemin (Deuteromycota: Hyphomycetes) strains isolated from Varroa mites in southern France. Journal of Apicultural Research 45: 219-220.
Meikle, W.G., Mercadier, G., Holst, N., Nansen, C. and Girod, V. (2007). Duration
and spread of an entomopathogenic fungus, Beauveria bassiana (Deuteromycota: Hyphomycetes), used to treat Varroa mites (Acari: Varroidae) in honey bee (Hymenoptera: Apidae) hives. Journal of Economic Entomology 100: 1-10.
Meikle, W.G., Mercadier, G., Holst, N. and Girod, V. (2008a). Impact of two
treatments of a formulation of Beauveria bassiana (Deuteromycota: Hyphomycetes) conidia on Varroa mites (Acari: Varroidae) and on honeybee (Hymenoptera: Apidae) colony health. Experimental and Applied Acarology 46: 105-117.

Literatur
139
Meikle, W.G., Mercadier, G., Holst, N., Nansen, C. and Girod, V. (2008b). Impact of a treatment of Beauveria bassiana (Deuteromycota: Hyphomycetes) on honeybee (Apis mellifera) colony health and on Varroa destructor mites (Acari: Varroidae). Apidologie 39: 247-259.
Meikle, W.G., Mercadier, G., Annas, F. and Holst, N. (2009). Effects of multiple
applications of a Beauveria based biopesticide on Varroa destructor (Acari: Varroidae) densities in honey bee (Hymenoptera: Apidae) colonies. Journal of Apicultural Research 48: 220-222.
Mier, T., River, F., Bermudez, J.C., Dominguez, Y., Benavides, C. and Ulloa, M.
(1991). Primer reporte en Mexico del aislamiento de Verticillium lecanii a partir de la mosquita blanca y pruebas de patogenicidad in vitro sobre este insecto. Revista Mexicana de Micologia 7: 149-156.
Milani, N. (1999). The resistance of Varroa jacobsoni Oud. to acaricides. Apidologie
30: 229-234. Moore, D., Angel, J., Cheeseman, I., Fahrbach, S. and Robinson, G. (1998).
Timekeeping in the honey bee colony: Integration of circadian rhythms and division of labor. Behavioral Ecology and Sociobiology 43: 147-160.
Müller, U., Vogel, P., Alber, G. and Schaub, G.A. (2008). The innate immune
system of mammals and insects. Contributions to Microbiology 15: 21-44. Mwangi, E.N., Kaaya, G.P. and Essuman, S. (1995). Experimental infections ot the
tick Rhipicephalus appendiculatus with entomopathogenic fungi, Beauveria bassiana and Metarhizium anisopliae, and natural infections of some ticks with bacteria and fungi. Journal of African Zoology 109: 151-160.
Nanetti, A., Büchler, R., Charriere, J.D., Fries, I., Helland, S., Imdorf, A., Korpela,
S. and Kristiansen, P. (2003). Oxalic acid treatments for Varroa control (review). Apiacta 38: 81-87.
Nauen, R., Ebbinghaus-Kintscher, U. and Schmuck, R. (2001). Toxicity and
nicotinic acetylcholine receptor interaction of imidacloprid and its metabolites in Apis mellifera (Hymenoptera: Apidae). Pest Management Science 57: 577-586.
Navajas, M., Migeon, A., Alaux, C., Martin-Magniette, M.L., Robinson, G.E.,
Evans, J.D., Cros-Arteil, S., Crauser, D. and Le Conte, Y. (2008). Differential gene expression of the honey bee Apis mellifera associated with Varroa destructor infection. BMC Genomics 9: 301.
OEPP/EPPO (2001). Hazards of pesticides to bees. Guidelines for the efficacy
evaluation of plant protection products. Side-effects on honeybees. E. a. M. P. P. Organization. Paris, l´institut national de la recherche agronomique: 279-286.
Oldroyd, B.P. (1999). Coevolution while you wait: Varroa jacobsoni, a new parasite
of western honeybees. Trends in Ecology and Evolution 14: 312-315.

Literatur
140
Ortiz-Caton, M. and Alatorre-Rosas, R. (1996). Factores que influyen sobre el impacto de los hongos entomopatogenos en el control de la mosquita blanca. In: Arredondo-Bernal, H.C., Jones, W.A. and Alatorre-Rosas, R. (ed) Simposio Biológico de Mosquita Blanca. XIX Congreso Nacional de Control Biológico. Sociedad Mexicana de Control Biológico. Culiacán, Sinaloa, México: 34-39.
Osborne, L.S. and Landa, Z. (1992). Biological control of whiteflies with
entomopathogenic fungi. Florida Entomologist 75: 456-471. Paz, Z., Burdman, S., Gerson, U. and Sztejnberg, A. (2007a). Antagonistic effects
of the endophytic fungus Meira geulakonigii on the citrus rust mite Phyllocoptruta oleivora. Journal of Applied Microbiology 103: 2570-2579.
Paz, Z., Gerson, U. and Sztejnberg, A. (2007b). Assaying three new fungi against
citrus mites in the laboratory and a field trial. Biocontrol 52: 855-862. Pell, J.K., Eilenberg, J., Hajek, A.E. and Steinkraus, D.C. (2001). Biology, ecology
and pest management potential of Entomophthorales. In: Butt, T.M., Jackson, C.W. and Magan, N. (ed) Fungi as biocontrol agents - progress, problems and potential. CABI Publishing, Wallingford: 71-154.
Peng, C.Y.S., Fang, Y., Xu, S. and Ge, L. (1987). The resistance mechanism of the
asian honey bee Apis cerana Fabr. to an ectoparasitic mite, Varroa jacobsoni Oudemanns. Journal of Invertebrate Pathology 49: 54-60.
Peng, C.Y.S., Mussen, E., Fong, A., Cheng, P., Wong, G. and Montague, M.A.
(1996). Laboratory and field studies on the effects of the antibiotic tylosin on honey bee Apis mellifera L. (Hymenoptera: Apidae) development and prevention of american foulbrood disease. Journal of Invertebrate Pathology 67: 65-71.
Peng, C.Y.S., Zhou, X. and Kaya, H.K. (2002). Virulence and site of infection of the
fungus Hirsutella thompsonii to the honey bee ectoparasitic mite, Varroa destructor. Journal of Invertebrate Pathology 81: 185-195.
Poinar, R. and Poinar, G. (1998). Parasites and pathogens of mites. Annual Review
of Entomology 43: 449-469. Polar, P., Kairo, M.T., Peterkin, D., Moore, D., Pegram, R. and John, S.A. (2005).
Assessment of fungal isolates for development of a myco-acaricide for cattle tick control. Vector Borne and Zoonotic Diseases 5: 276-284.
Randolt, K., Gimple, O., Geissendorfer, J., Reinders, J., Prusko, C., Mueller, M.
J., Albert, S., Tautz, J. and Beier, H. (2008). Immune-related proteins induced in the hemolymph after aseptic and septic injury differ in honey bee worker larvae and adults. Archives of Insect Biochemistry and Physiology 69: 155-167.
Ratnieks, F.L.W. and Carreck, N.L. (2010). Clarity on honey bee collapse? Science
327: 152-153.

Literatur
141
Riba, G. and Marcandier, S. (1984). Influence de l´humidité relative sur l´agressivité et la variabilité des souches de Beauveria bassiana Vuillemin et de Metarhizium anisopliae Sorokin, hyphomycètes pathogènes de la pyrale du mais Ostrinia nubilalis Hübner. Agronomie 4: 189-194.
Ritter, W. (1982). Varroatose: Eine neue Krankheit der Honigbiene Apis mellifera.
Tierärztliche Umschau 37: 344-354. Ritter, W., Leclercq, E. and Koch, W. (1984). Observations des populations
d'abeilles et de Varroa dans les colonies à différents niveaux d'infestation. Apidologie 15: 389-400.
Rosenkranz, P. (1999). Honey bee (Apis mellifera L.) tolerance to Varroa jacobsoni
Oud. in South America. Apidologie 39: 159-172. Rosenkranz, P. and Fries, I. (2005). Does a four year selection in a closed honey
bee population lead to Varroa tolerance? Test of queens deriving from surviving colonies. IUSSI, Halle.
Sachs, L. and Hedderich, J. (2009). Angewandte Statistik. Berlin, Springer Verlag. Saitou, N. and Nei, M. (1987). The neighbor-joining method: A new method for
reconstructing phylogenetic trees. Molecular Biology and Evolution 4: 406-425.
Sammataro, D., Gerson, U. and Needham, G. (2000). Parasitic mites of honey
bees: Life history, implications, and impact. Annual Review of Entomology 45: 519-548.
Shah, P.A. and Goettel, M.S. (1999). Directory of microbial control products and
services. Society for Invertebrate Pathology, Microbial Control Division. Shaw, K.E., Davidson, G., Clark, S., Ball, B.V., Pell, J.K., Chandler, D. and
Sunderland, K.D. (2002). Laboratory bioassays to assess the pathogenicity of mitosporic fungi to Varroa destructor (Acari: Mesostigmata), an ectoparasitic mite of the honeybee, Apis mellifera. Biological Control 24: 266-276.
Siegel, S. and Castellan, N.J. (1988). Nonparametric statistics for the behavioral
sciences. Boston, McGraw-Hill. Southwick, E.E. and Southwick, L. (1992). Estimating the economic value of honey
bees (Hymenoptera: Apidae) as agricultural pollinators in the United States. Journal of Economic Entomology 85: 621-633.
Spatafora, J.W., Sung, G.H., Sung, J.M., Hywel-Jones, N.L. and White, J.F.
(2007). Phylogenetic evidence for an animal pathogen origin of ergot and the grass endophytes. Molecular Ecology 16: 1701-1711.
Steenberg, T., Kryger, P. and Holst, N. (2010). A scientific note on the fungus
Beauveria bassiana infecting Varroa destructor in worker brood cells in honey bee hives. Apidologie 41: 127-128.

Literatur
142
Steiner, J., Dittmann, F., Rosenkranz, P. and Engels, W. (1994). The first gonocycle of the parasitc mite (Varroa jacobsoni) in relation to preimaginal development of its host, the honey bee (Apis mellifera carnica). Invertebrate Reproduction and Development 25: 175-183.
Stenbak, C.R., Ryu, J.H., Leulier, F., Pili-Floury, S., Parquet, C., Herve, M.,
Chaput, C., Boneca, I. G., Lee, W. J., Lemaitre, B. and Mengin-Lecreulx, D. (2004). Peptidoglycan molecular requirements allowing detection by the Drosophila immune deficiency pathway. Journal of Immunology 173: 7339-7348.
Stone, J.K., Bacon, C.W. and White, J.F. (2000). An overview of endophytic
microbes: Endophytism defined. In: Stone, J.K., Bacon, C.W. and White, J.F. (ed) Microbial endophytes. New York, Dekker: 3-29.
Strasser, H., Vanas, V., Hutwimmer, S. and Zelger, R. (2007). Biologische
Kontrolle von Zecken (Ixodes ricinus) durch den insektentötenden Pilz Metarhizium anisopliae (Metch). INTERREG IIIA-Programm Österreich-Italien 2000-2006, Land Tirol und Bundesministerium für Land- und Forstwirtschaft, Umwelt und Wasserwirtschaft Österreich.
Sugimoto, M., Koike, M., Hiyama, N. and Nagao, H. (2003). Genetic,
morphological, and virulence characterization of the entomopathogenic fungus Verticillium lecanii. Journal of Invertebrate Pathology 82: 176-187.
Sztejnberg, A., Paz, Z., Boekhout, T., Gafni, A. and Gerson, U. (2004). A new
fungus with dual biocontrol capabilities: Reducing the numbers of phytophagous mites and powdery mildew disease damage. Crop Protection 23: 1125-1129.
Talaei-Hassanloui, R., Kharazi-Pakdel, A., Goettel, M.S., Little, S. and Mozaffari,
J. (2007). Germination polarity of Beauveria bassiana conidia and its possible correlation with virulence. Journal of Invertebrate Pathology 94: 102-107.
Tamura, K., Nei, M. and Kumar, S. (2004). Prospects for inferring very large
phylogenies by using the neighbor-joining method. Proceedings of the National Academy of Science of the USA 101: 11030-11035.
Tamura, K., Dudley, J., Nei, M. and Kumar, S. (2007). MEGA4: Molecular
evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution 24: 1596-1599.
Tanaka, E.S., Kiminori, I., Yumi, Y., Tanaka, C., Shimizu, K., Imanishi, Y.,
Yasuda, F. and Tanaka, C. (2008). Isolation of basidiomycetous anamorphic yeast-like fungus Meira argovae found on japanese bamboo. Mycoscience 49: 329-333.
Tsagou, V., Lianou, A., Lazarakis, D., Emmanouel, N. and Aggelis, G. (2004).
Newly isolated strains belonging to Bacillaceae (Bacillus sp.) and Micrococcaceae accelerate death of honey bee mite, Varroa destructor (V. jacobsoni), in laboratory assays. Biotechnology Letters 26: 529-532.

Literatur
143
Tuininga, A.R., Miller, J.L., Morath, S.U., Daniels, T.J., Falco, R.C., Marchese, M., Sahabi, S., Rosa, D. and Stafford, K. C. (2009). Isolation of entomopathogenic fungi from soils and Ixodes scapularis (Acari: Ixodidae) ticks: Prevalence and methods. Journal of Medical Entomology 46: 557-565.
van der Geest, L.P.S., Elliot, S.L., Breeuwer, J.A.J. and Beerling, E.A.M. (2000).
Diseases of mites. Experimental and Applied Acarology 24: 497-560. Vandame, R., Morand, S., Colin, M.E. and Belzunces, L.P. (2002). Parasitism in
the social bee Apis mellifera: Quantifying costs and benefits of behavioral resistance to Varroa destructor mites. Apidologie 33: 433-445.
Wägele, J. W. (2001). Grundlagen der phylogenetischen Systematik. Verlag Dr.
Friedrich Pfeil, München. Weinberg, K.P. and Madel, G. (1985). The influence of the mite Varroa jacobsoni
Oud. on the protein concentration and the haemolymph volume of the brood of worker bees and drones of the honey bee Apis mellifera L. Apidologie 16: 421-436.
Weinstock, G.M., Robinson, G.E., Gibbs, R.A., Worley, K., Evans, J.D.,
Maleszka, R., Robertson, H.M., Weaver, D.B. and Beye, M. (2006). Insights into social insects from the genome of the honeybee Apis mellifera. Nature 443: 931-949.
White, T.J., Bruns, T., Lee, S. and Taylor, J. (1990). Amplification and direct
sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols: A guide to methods and applications. San Diego, Academic Press: 315-322.
Whitfield, C.W., Behura, S.K., Berlocher, S.H., Clark, A.G., Johnston, J.S.,
Sheppard, W.S., Smith, D.R., Suarez, A.V., Weaver, D. and Tsutsui, N.D. (2006). Thrice out of Africa: Ancient and recent expansions of the honey bee, Apis mellifera. Science 314: 642-645.
Wraight, S.P., Carruthers, R.I., Jaronski, S.T., Bradley, C.J., Garca, C.J. and
Galaini-Wraight, S. (2000). Evaluation of the entomopathogenic fungi Beauveria bassiana and Paecilomyces fumosoroseus for microbial control of the silverleaf whitefly, Bemisia argentifolii. Biological Control 17: 203-217.
Wright, J.E. and Kennedy, F.G. (1996). A new biological product for control of major
greenhouse pests. In: Proceedings of the Brighton Crop Protection Conference - Pests and Diseases, British Crop Protection Council 3: 885-892.
Yasuda, F., Yamagishi, D., Izawa, H., Kodama, M. and Otani, H. (2007). Fruit stain
of japanese pear caused by basidiomycetous, yeastlike fungi Meira geulakonigii and Pseudozyma aphidis. Japanese Journal of Phytopathology 73: 166-171.

Literatur
144
Yue, D., Nordhoff, M., Wieler, L.H. and Genersch, E. (2008). Fluorescence in situ hybridization (FISH) analysis of the interactions between honeybee larvae and Paenibacillus larvae, the causative agent of american foulbrood of honeybees (Apis mellifera). Environmental Microbiology 10: 1612-1620.
Zare, R. and Gams, W. (2001). A revision of Verticillium section Prostrata. IV: The
genera Lecanicillium and Simplicillium. Nova Hedwigia 73: 1-50. Zare, R., Gams, W. and Culham, A. (2000). A revision of Verticillium section
Prostrata. I: Phylogenetic studies using ITS sequences. Nova Hedwigia 71: 465-480.

145
Erklärung
Hiermit erkläre ich, dass ich die Arbeit selbständig verfasst und bei keiner anderen
Fakultät eingereicht und dass ich keine anderen als die angegebenen Hilfsmittel
verwendet habe. Es handelt sich bei der heute von mir eingereichten Dissertation um
sechs in Wort und Bild völlig übereinstimmende Exemplare.
Weiterhin erkläre ich, dass digitale Abbildungen nur die originalen Daten enthalten
und in keinem Fall inhaltsverändernde Bildbearbeitung vorgenommen wurde.
Bochum, den 26. Februar 2010
(Markus Holt)

146
Markus Holt,
geboren am 27. April 1971
Curriculum vitae
seit 04.2005 Promotion in der Internationalen Graduiertenschule
Biowissenschaften der Ruhr-Universität Bochum
Gutachter: Prof. Dr. Wolfgang H. Kirchner
Prof. Dr. Günter A. Schaub
10.1994 – 11.2004 Studium der Biologie an der Universität Bochum
Abschluss: Diplom
Diplomarbeit in der AG Verhaltensbiologie und Didaktik der
Biologie: „Mikrosatelliten-basierte Untersuchungen zur
Soziogenetik des Europäischen Luchses (Lynx lynx (L.))“
08.1981 – 06.1990 Heisenberg-Gymnasium in Gladbeck
Abschluss: Allgemeine Hochschulreife, Note 1,4
08.1977 – 06.1981 Käthe-Kollwitz-Grundschule in Gladbeck
seit 10.2009 Studienkoordinator des Fachbereichs Biologie der
Westfälischen Wilhelms-Universität Münster
02.2008 – 09.2009 Leiter des Ausbildungslabors für Biologielaboranten der
Ruhrkohle AG Bildung
03.2007 – 07.2007 Selbständiger Dozent der Ruhrkohle AG Bildung
01.2005 – 09.2006 Wissenschaftlicher Mitarbeiter der AG Verhaltensbiologie und
Didaktik der Biologie
01.2002 – 03.2003 Mitarbeiter des Instituts für Management und Organisation
(IMO) Bochum
05.1995 – 04.2009 Mitarbeiter des Westfälischen Industriemuseums
Henrichshütte Hattingen, Entwicklung und Durchführung des
Programms „Ökologie der Industriebrache“
12.2006 – 01.2008 Organisation des Doktorandenforums der Internationalen
Graduiertenschule Biowissenschaften der Ruhr-Universität
Bochum

147
Danksagung
Mein Dank gilt zuvorderst Prof. Dr. Wolfgang H. Kirchner für die Entwicklung des
Themas der Arbeit, vor allem aber für seinen immer zielführenden Rat und für sein
Vertrauen, das unabdingbare Basis einer selbständigen Arbeit war.
Für die freundliche Übernahme des Zweitgutachtens, für konstruktive Diskussionen
und die Kooperation im Rahmen des Immunassays möchte ich mich bei Prof. Dr.
Günter A. Schaub bedanken – seiner Mitarbeiterin Dipl.-Biol. Jennifer Pausch sei
herzlich für ihre praktische Unterstützung in der Durchführung des Assays gedankt.
Wissenschaftliche Arbeit profitiert immer von einem Austausch zwischen einzelnen
Disziplinen – für durchweg hilfreiche Hinweise möchte ich mich stellvertretend für alle
bei Jun. Prof. Dr. Julia Bandow, Prof. Dr. Dominik Begerow, PD Dr. Elke Genersch
(Länderinstitut für Bienenkunde Hohen Neuendorf), PD Dr. Minou Nowrousian und
PD Dr. Christine Struck (Universität Rostock) bedanken.
Ein ganz besonderer Dank gilt Dr. Pia Aumeier: Zum einen wären die Labor- und
Halbfreilandversuche an Bienen und Milben genauso wie das Pilzscreening aufgrund
des sehr erheblichen Versuchstierbedarfs ohne den von ihr exzellent geführten,
großen Bestand an Versuchsvölkern schlicht unmöglich gewesen, zum anderen
stand sie immer für einen freundschaftlichen und kompetenten Rat zur Verfügung.
Dem Landesgesundheitsamt Baden-Württemberg und der Firma Koppert Biological
Systems danke ich für die Zurverfügungstellung von Pilzstämmen für die Labor- und
Halbfreilandversuche.
Meinen Codoktorandinnen Dr. Julia Eggermann, Dr. Christina Heimken und Dr. Nina
Minkley danke ich dafür, dass sie wesentlich zu einem guten Arbeitsklima
beigetragen haben: Nullius boni sine socio iucunda possessio est (Seneca).
Nicht zuletzt möchte ich mich bei meinen Arbeitgebern der Jahre 2007 – 2010, der
Ruhrkohle AG und der Westfälischen Wilhelms-Universität Münster, bedanken: Für
ihr Entgegenkommen, das insbesondere in der Bereitschaft bestand, mir bis an die
Grenzen des ihnen Möglichen Flexibilität einzuräumen, um neben der Arbeit die
Promotion fortführen zu können.
Finis coronat opus (Ovid)