vo einführung in die genetik und...
TRANSCRIPT

3.5.2011 Dienstag:13:30-15:0010.5.201117.5.2011 12:15-13:4524.5.201131.5.201114.6.201121.6.2011
VO Einführung in die Genetik und Zellbiologie
Mag.Dr. Selma Osmanagic-Myers

Zellbiologie-Lehrinhalte
Entstehung des Lebens. Aufbau der Pro- und Eukaryontenzelle. Endosymbiontentheorie. Aufbau der Zellmembranen. Membrantransport kleiner Moleküle und Ionen
Zellkompartimente und Proteinsortierung. Mitochondrien und Chloroplasten
Intrazellulärer Vesikeltransport
Das Cytoskelett. Zell-Zell und Zell-Substrat Interaktionen
Zellkommunikation. Spezialisierte Zelltypen und deren Eigenschaften (Motilität, Sekretion...), Aufbau von Geweben
Zellzyklus und dessen Kontrolle
Zellteilung. Krebsentstehung und Zellzykluskontrolle
Mag.Dr. Selma Osmanagic-Myers

Literatur
Kleinig / Sitte: Zellbiologie, Gustav Fischer
Molecular Cell Biology, 4e by Harvey Lodish, Arnold Berk, S. Lawrence Zipursky, Paul Matsudaira, David Baltimore, and James Darnellhttp://www.whfreeman.com/lodish/index.htm
Alberts / Bray /Lewis /Raff /Roberts /Watson: Molecular biology of the cell, Garland publishers
Powerpoint Folien unter:
Login mit unet ID und password
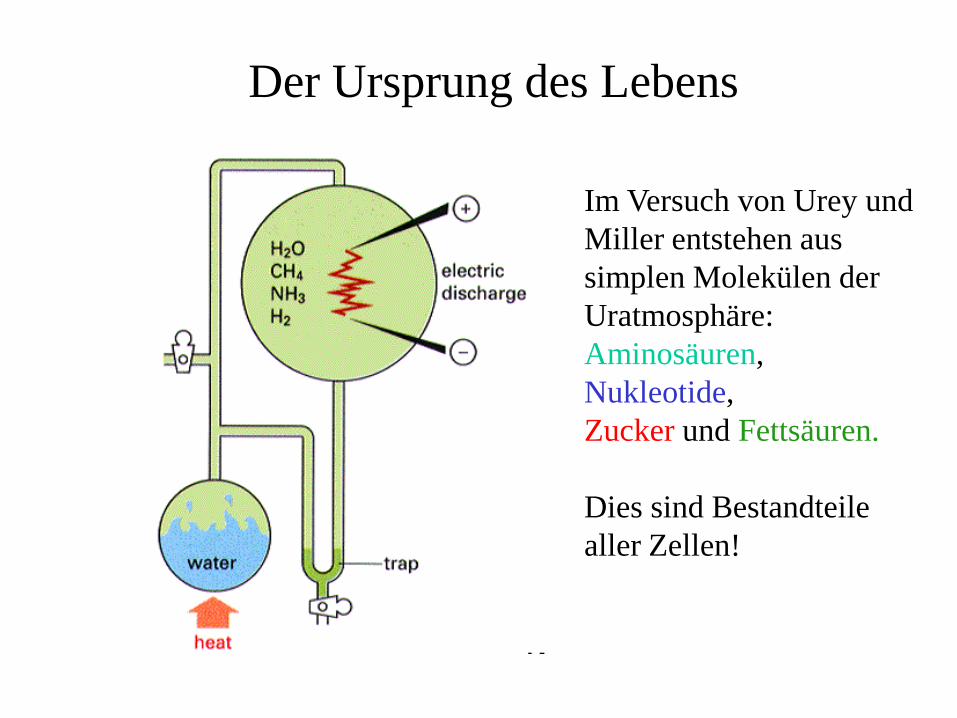
Der Ursprung des Lebens
Im Versuch von Urey und Miller entstehen aus simplen Molekülen der Uratmosphäre:Aminosäuren, Nukleotide,Zucker und Fettsäuren.
Dies sind Bestandteile aller Zellen!
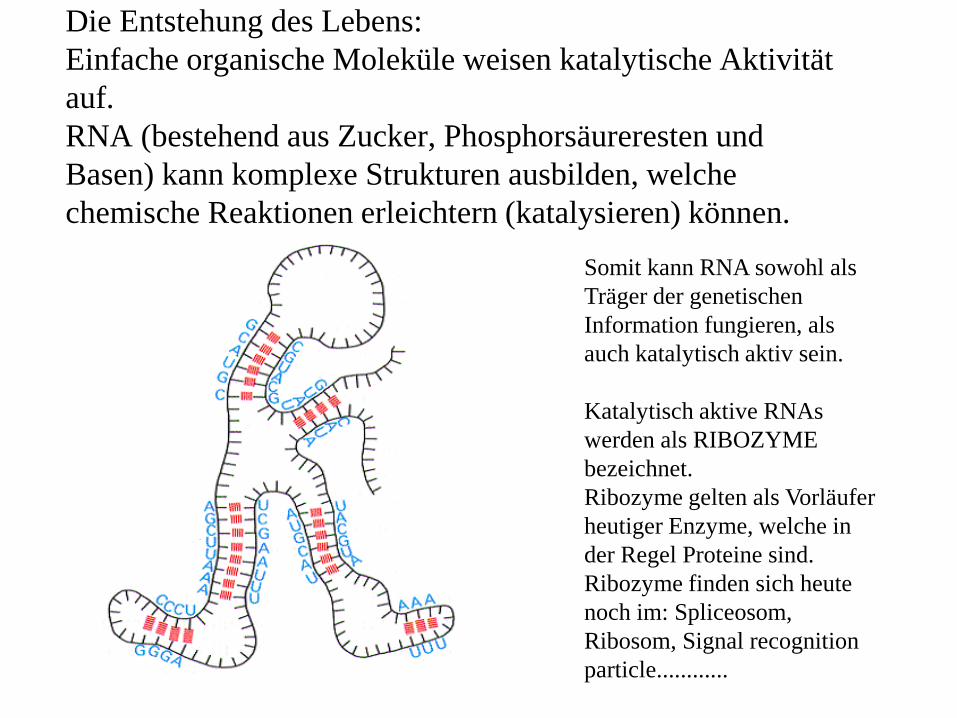
Die Entstehung des Lebens: Einfache organische Moleküle weisen katalytische Aktivität auf.RNA (bestehend aus Zucker, Phosphorsäureresten und Basen) kann komplexe Strukturen ausbilden, welche chemische Reaktionen erleichtern (katalysieren) können.
Somit kann RNA sowohl als Träger der genetischen Information fungieren, als auch katalytisch aktiv sein.
Katalytisch aktive RNAs werden als RIBOZYME bezeichnet.Ribozyme gelten als Vorläufer heutiger Enzyme, welche in der Regel Proteine sind.Ribozyme finden sich heute noch im: Spliceosom, Ribosom, Signal recognition particle............
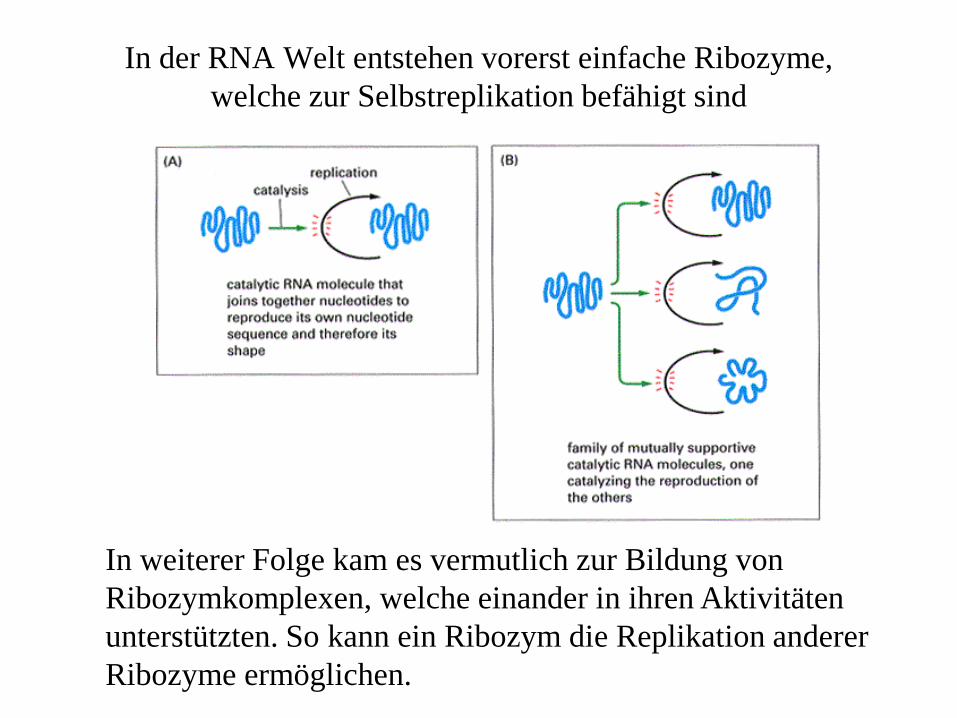
In der RNA Welt entstehen vorerst einfache Ribozyme, welche zur Selbstreplikation befähigt sind
In weiterer Folge kam es vermutlich zur Bildung von Ribozymkomplexen, welche einander in ihren Aktivitäten unterstützten. So kann ein Ribozym die Replikation anderer Ribozyme ermöglichen.
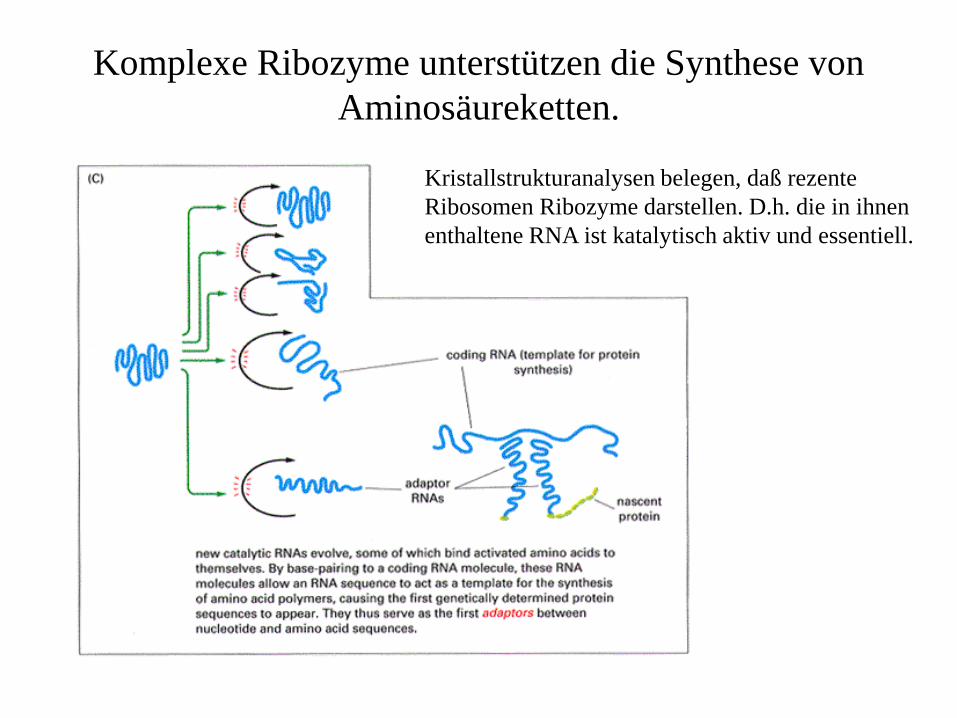
Komplexe Ribozyme unterstützen die Synthese von Aminosäureketten.
Kristallstrukturanalysen belegen, daß rezente Ribosomen Ribozyme darstellen. D.h. die in ihnen enthaltene RNA ist katalytisch aktiv und essentiell.
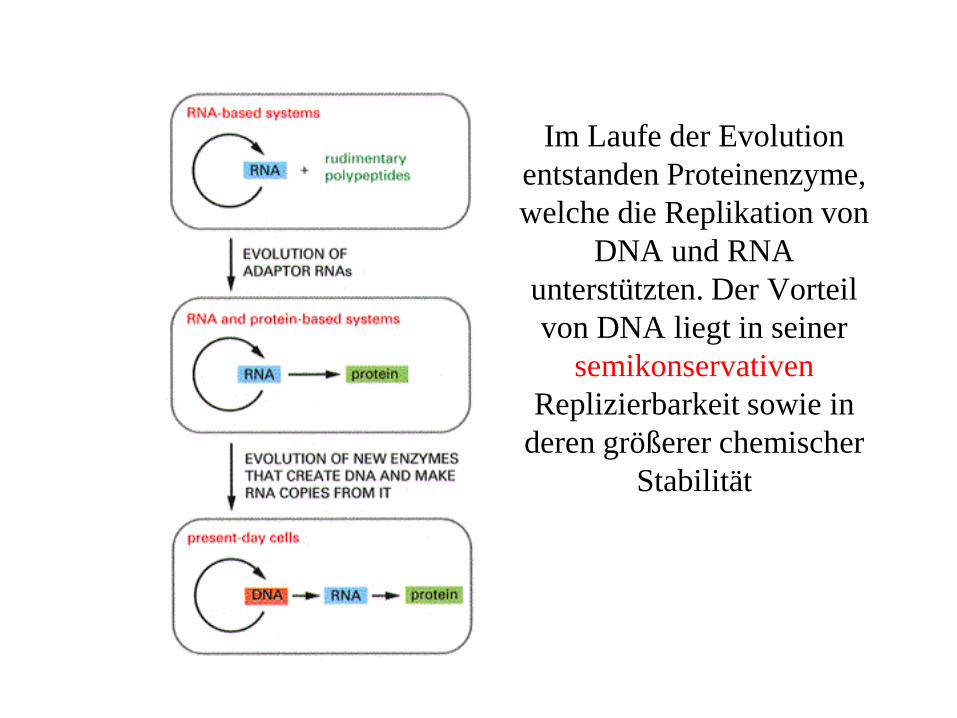
Im Laufe der Evolution entstanden Proteinenzyme, welche die Replikation von
DNA und RNA unterstützten. Der Vorteil von DNA liegt in seiner
semikonservativenReplizierbarkeit sowie in
deren größerer chemischer Stabilität

Die Entstehung des Lebens
Präbiotische (chemische) Evolution
Die RNA Welt (Ribozyme)
Der Progenot, Vorläufer aller Urzellen
Cenancestor, letzter gemeinsamer vorfahren von Archaea, Eubakteria und Eukaryota
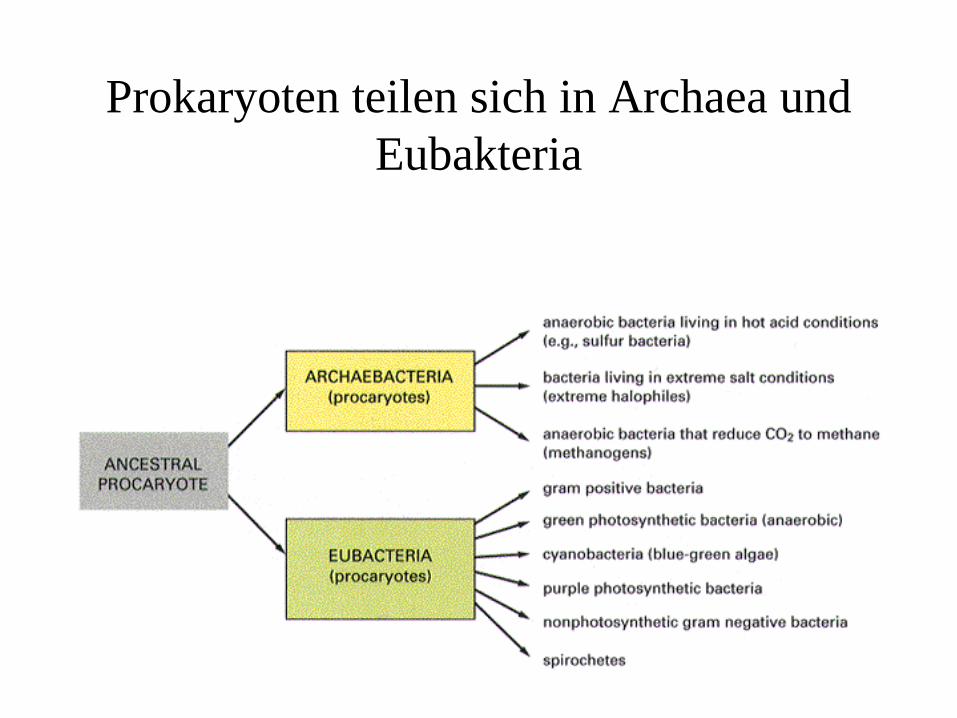
Prokaryoten teilen sich in Archaea und Eubakteria
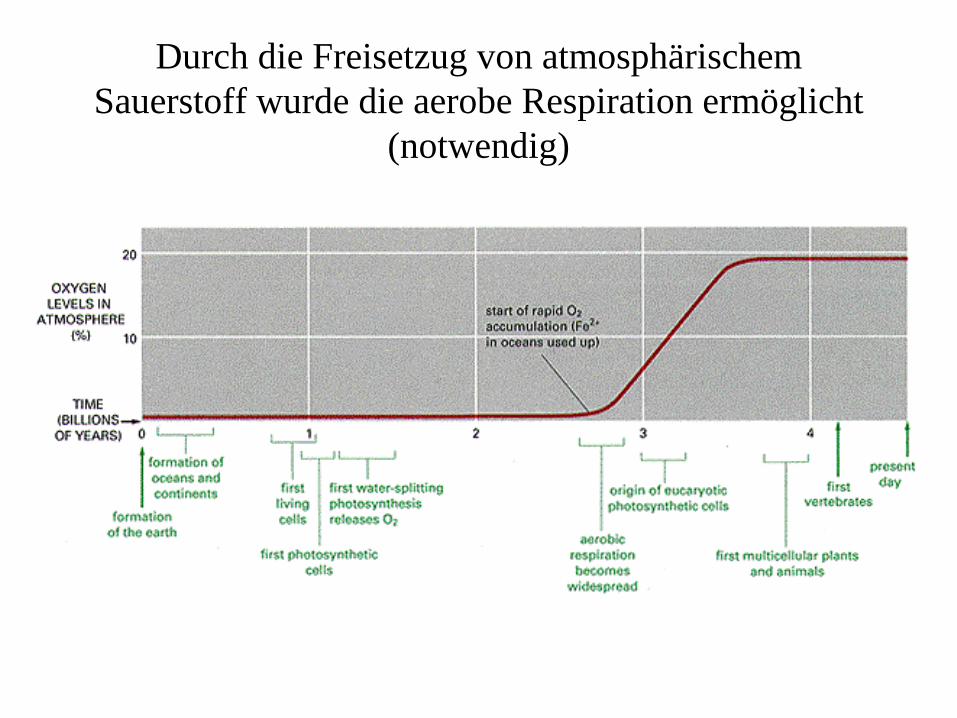
Durch die Freisetzug von atmosphärischem Sauerstoff wurde die aerobe Respiration ermöglicht
(notwendig)

Die Endosymbiontenhypothese
Plastiden (in Pflanzen) und Mitochondrien sind die Abkömmlinge ursprünglich frei lebender Prokaryoten, welche von der Eukaryotenzelle domestiziert wurden.
Plastiden sind die Abkömmlinge von Cyanobakterien
Mitochondrien sind die Abkömmlinge aerober Bakterien

Evidenz für die Endosymbiontenhypothese (1):
Endosymbionten werden auch heute noch gefunden (Cyanophora)
Mitochondrien und Plastiden sind zumindest von einer doppelten Memban umgeben: Die äußere Membran entspricht der Wirtsmembran, die innere der bakteriellen Membran
Die innere Membran von Mitochondrien und Plastiden weist auch heute noch viele bakterielle Merkmale auf
Die in Organellen vorkommenden Biosynthesewege entsprechen in verschiedener Hinsicht bakteriellen Biosynthesewegen

Evidenz für die Endosymbiontenhypothese (2):
Mitochondrien und Plastiden verfügen über ein eigenes genetisches System, welches in verschiedener Hinsicht jenem der Prokarionten entspricht. (Cirkuläres Genom, 70S Ribosomen, Translation)
Mitochondrien und Plastiden replizieren autonom und werden zufällig auf die Tochterzellen weiterverteilt

Jedoch:
Rezente Mitochondrien und Plastiden unterscheiden sich massgeblich von ihren prokaryotischen Vorläufern:
Das Genom ist stark reduziert, viele Proteine werden im Kern kodiert, im Zytoplama synthetisiert und in das jeweilige Organell importiert.
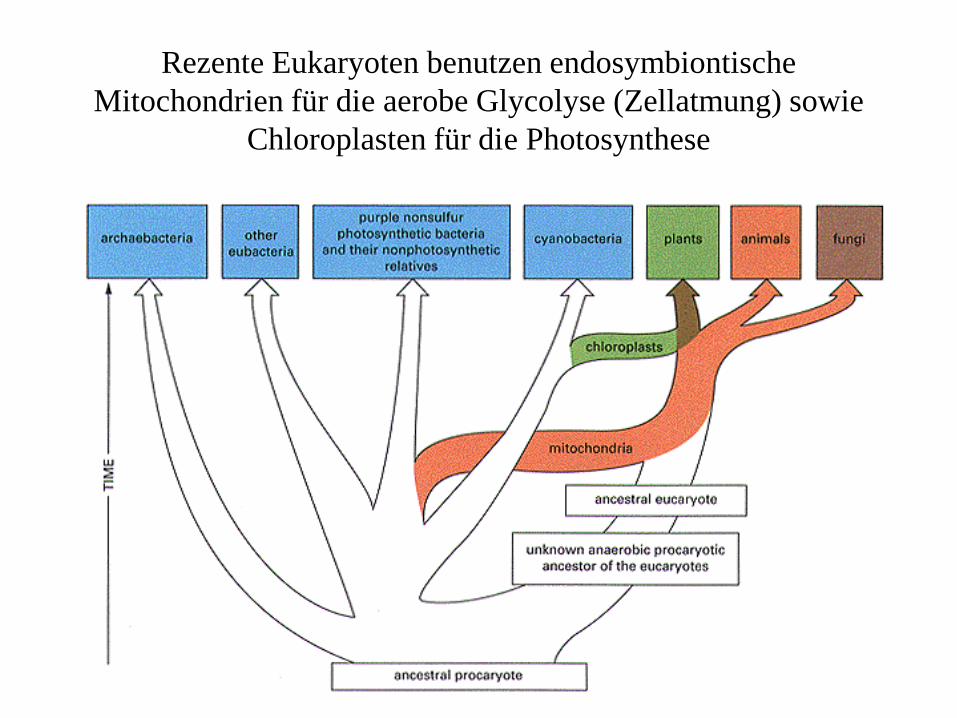
Rezente Eukaryoten benutzen endosymbiontische Mitochondrien für die aerobe Glycolyse (Zellatmung) sowie
Chloroplasten für die Photosynthese

Eukaryota-Prokaryota, die wesentlichen Unterschiede
ProkaryotaNucleoidCirkuläres GenomKonjugationKein ZytoskelettFlagellenKeine OrganellenKein EndomembransystemKeine Endo/Exocytose
EukaryotaZellkernChromosomenMeiose / MitoseZytoskelettCillien und GeißelnOrganellenEndomembransystemEndo /Exozytose

Schema einer Prokaryotenzelle
• Einzelliger Organismus, kann jedoch Lebensgemeinschaften (Coenobien) ausbilden
• Zwei Haupttypen: Bacteria und Archaea• Einfache Struktur
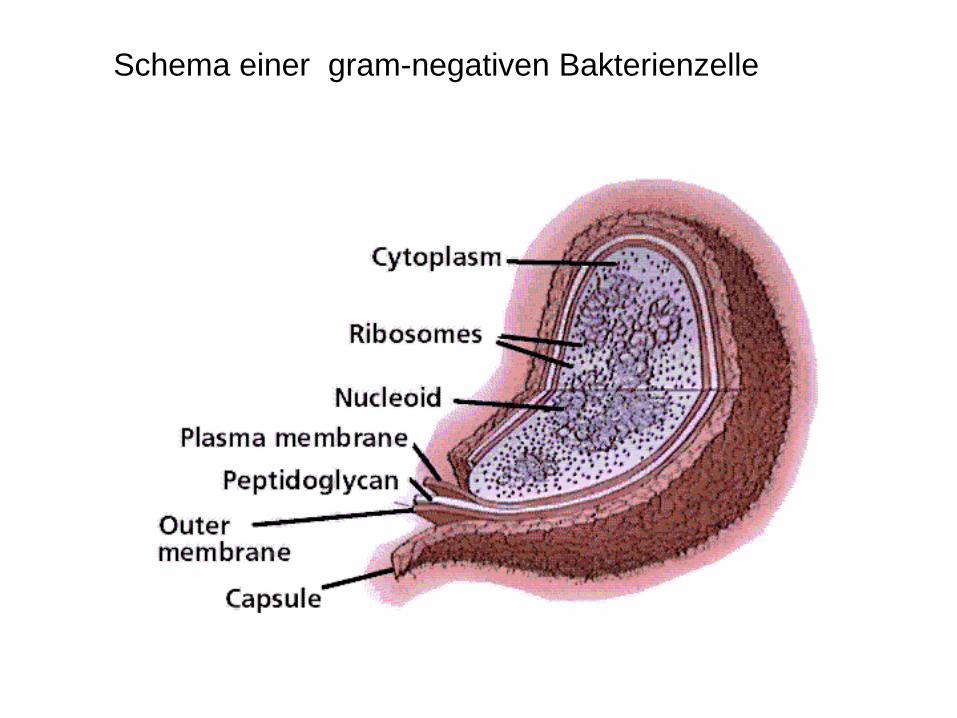
Schema einer gram-negativen Bakterienzelle
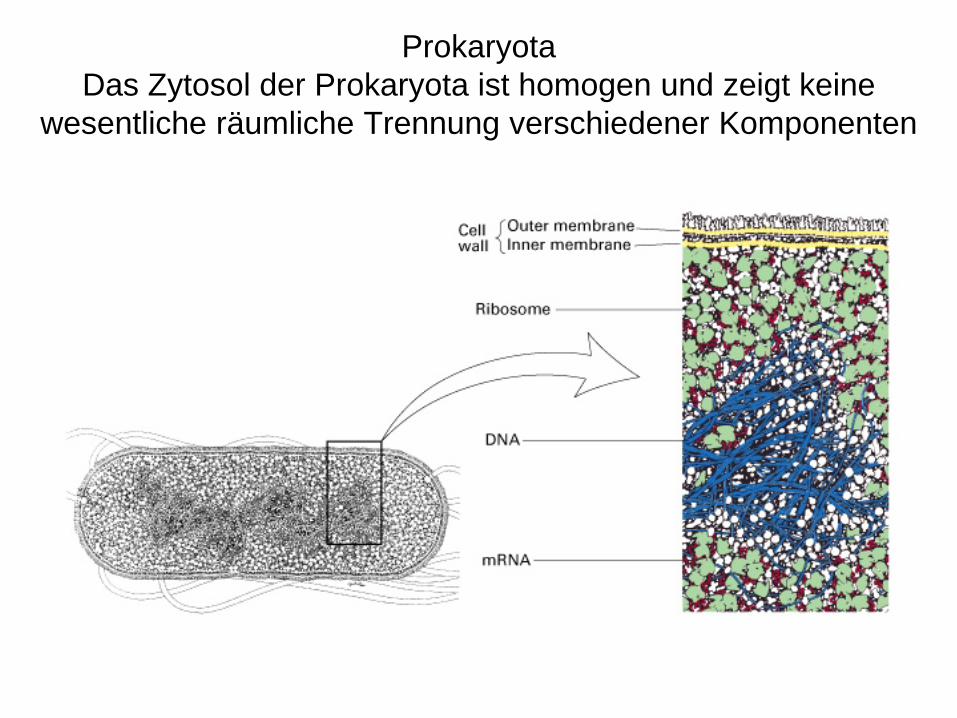
ProkaryotaDas Zytosol der Prokaryota ist homogen und zeigt keine
wesentliche räumliche Trennung verschiedener Komponenten
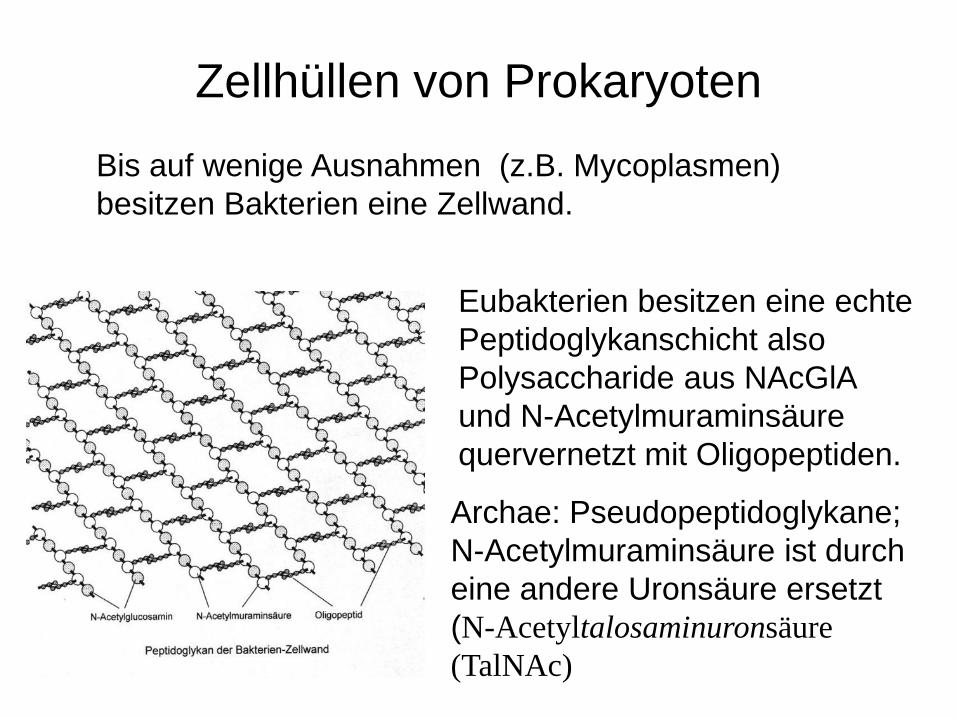
Eubakterien besitzen eine echte Peptidoglykanschicht also Polysaccharide aus NAcGlA und N-Acetylmuraminsäure quervernetzt mit Oligopeptiden.
Archae: Pseudopeptidoglykane; N-Acetylmuraminsäure ist durch eine andere Uronsäure ersetzt (N-Acetyltalosaminuronsäure (TalNAc)
Bis auf wenige Ausnahmen (z.B. Mycoplasmen) besitzen Bakterien eine Zellwand.
Zellhüllen von Prokaryoten
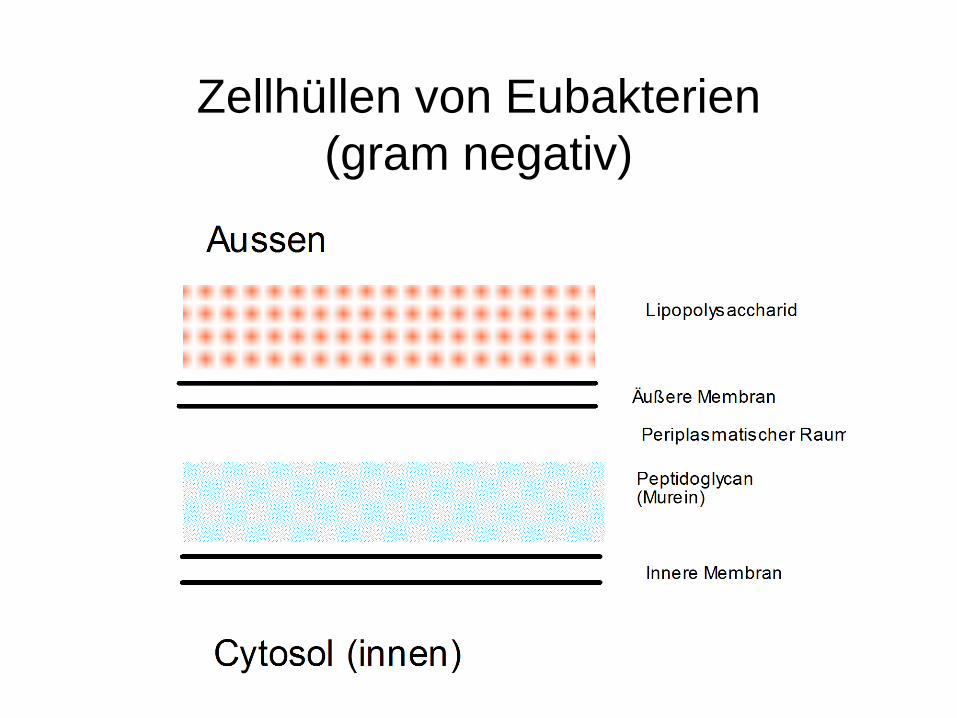
Zellhüllen von Eubakterien(gram negativ)
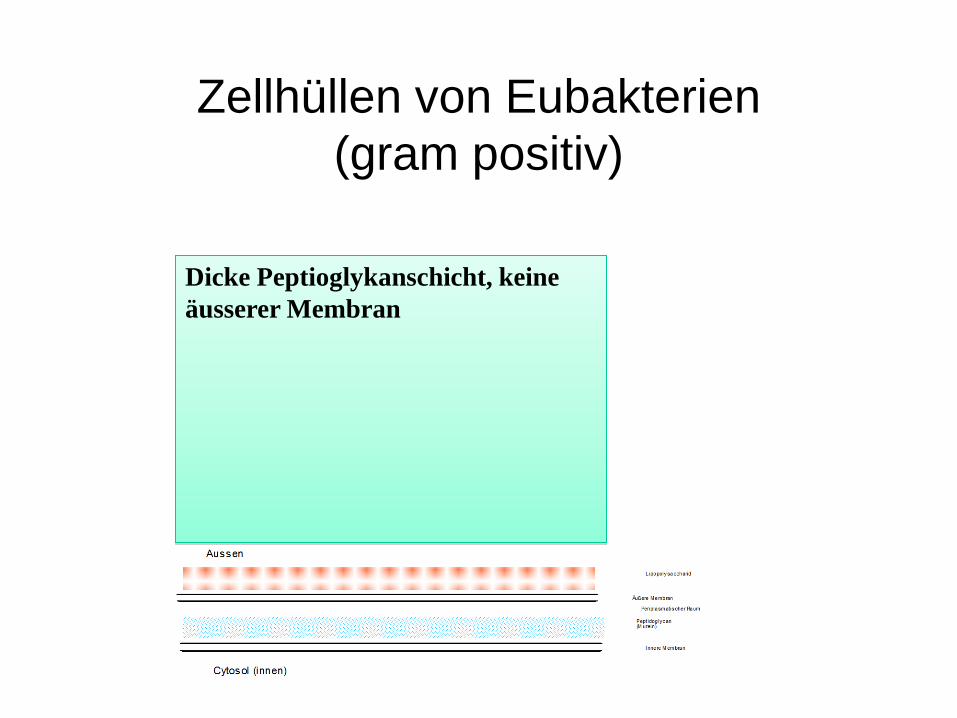
Zellhüllen von Eubakterien(gram positiv)
Dicke Peptioglykanschicht, keine äusserer Membran

Die Gram-Färbung unterscheidet die Dicke der Peptidoglykanschicht
Eine dicke Mureinschicht lässt sich schwer entfärben: Gram positivEine dünne Mureinschicht läßt sich leicht entfärben: Gram negativ

Für die Quervernetzung der Oligopeptide in der Mureinschicht sind Transpeptidasen verantwortlich.Diese Vernetzen eine endständige Aminogruppe von Glycin mit einem nicht-endständigen Alanin Rest.
Antibiotika der Penicilline und Cephalosporine inhibieren die Transpeptidasen und verhindern somit die Zellwandbildung
Lysozym (im Hühnereiweiss, Tränen, Schweiss, Speichel enthalten). Spaltet die glycosidische Bindung zwischen einer Acetylmuraminsäure und Acetylglucosamin und dient somit als Schutz gegen bakterielle Infektionen.
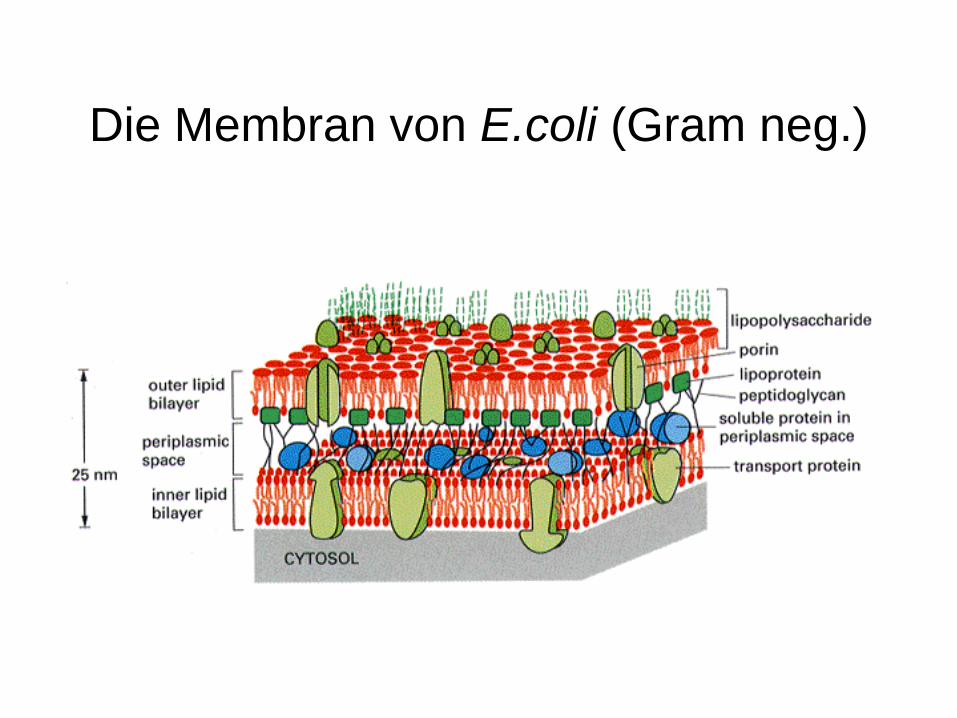
Die Membran von E.coli (Gram neg.)
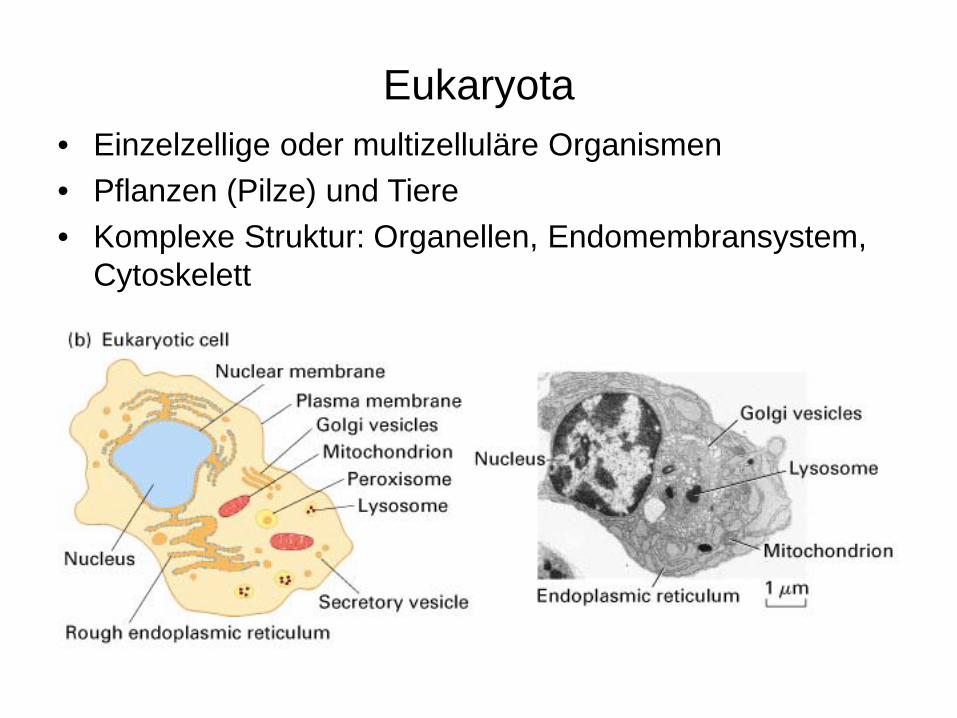
Eukaryota• Einzelzellige oder multizelluläre Organismen• Pflanzen (Pilze) und Tiere • Komplexe Struktur: Organellen, Endomembransystem,
Cytoskelett

Kompartimente der Eukarya
•Plasmamembran•Cytoplasma•Zellkern•Endoplasmatisches Retikulum•Golgi Apparat•Lytisches Kompartiment (Lysosomen)•Peroxisomen•Mitochondrien•Plastiden (Pflanzen)•Zellwände (Pilze und Pflanzen)

Membrane (1)Kompartimentierungsregel:
Biologische Membrane trennen in der Regel eine plasmatische Phase von einer nicht-plasmatischen.
Cytoplasma, Karyoplasma,Mitoplasma (Matrix), Plastoplasma(Stroma). Um von einem Zellplasma in das nächste zu gelangen, müssen immer 2 Membranen überwunden werden. Zwischen den 2 Membranen liegt eine nicht plasmatische Phase.
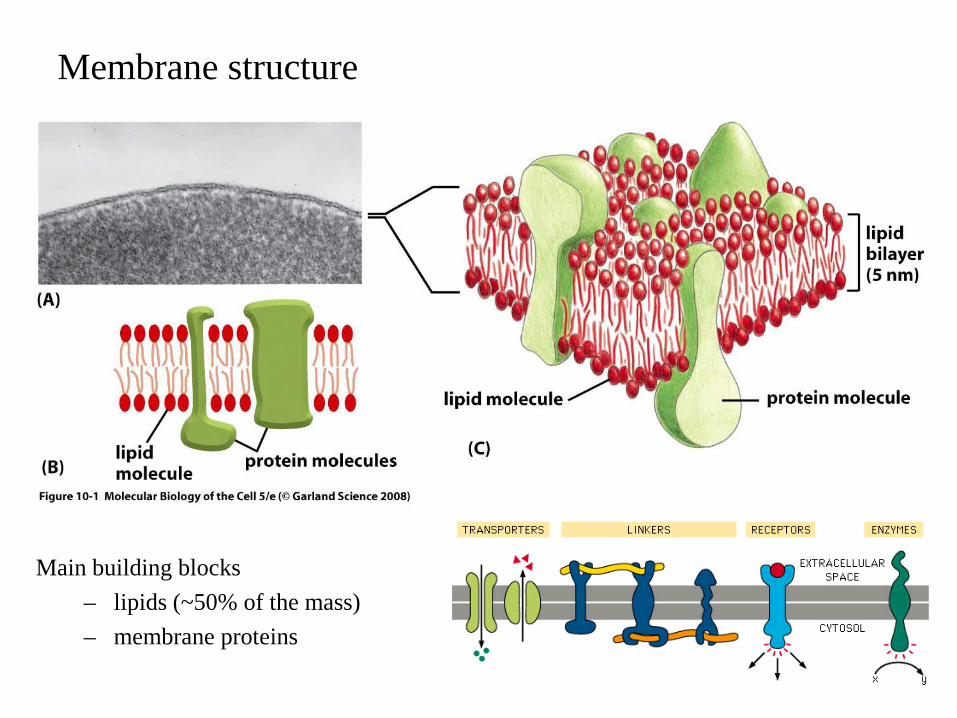
Membrane structure
Main building blocks– lipids (~50% of the mass)– membrane proteins
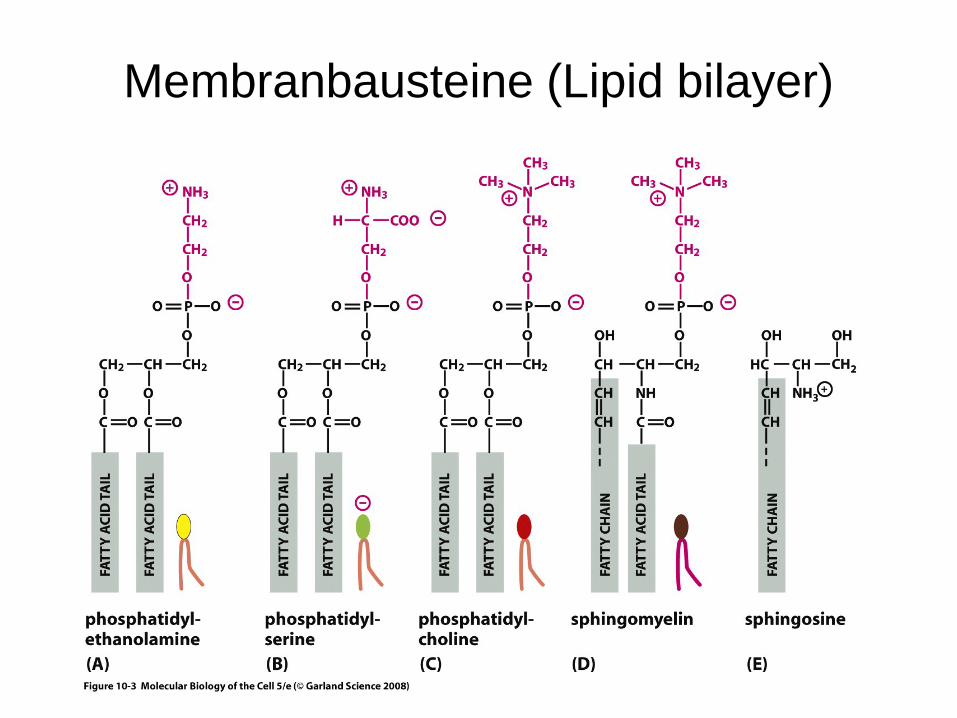
Membranbausteine (Lipid bilayer)
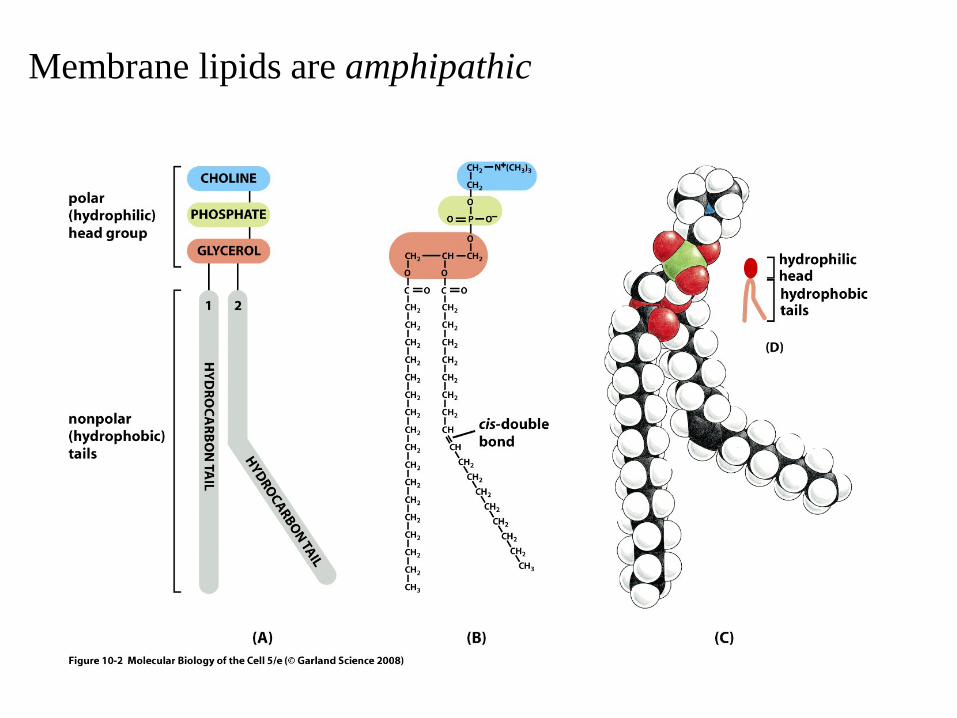
Membrane lipids are amphipathic
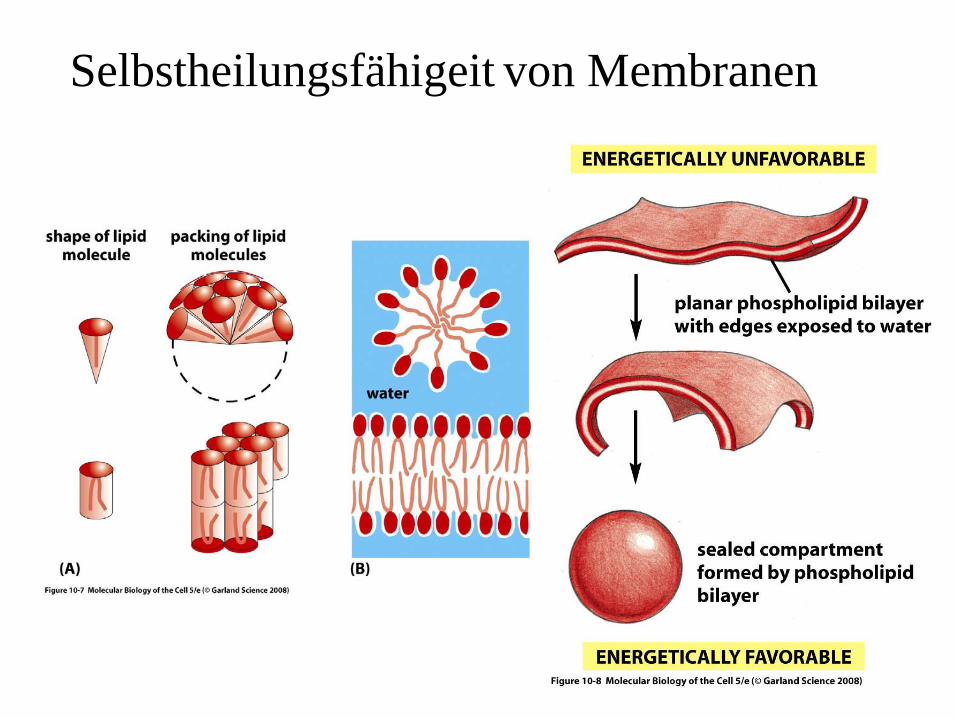
Selbstheilungsfähigeit von Membranen
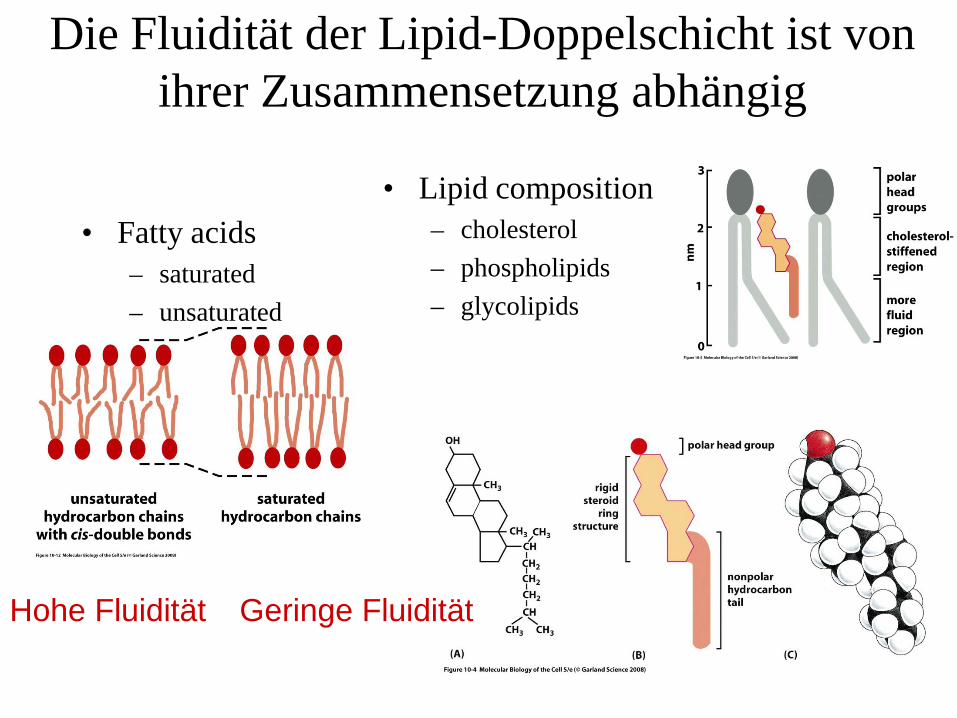
Die Fluidität der Lipid-Doppelschicht ist von ihrer Zusammensetzung abhängig
• Fatty acids– saturated– unsaturated
• Lipid composition– cholesterol– phospholipids– glycolipids
Hohe Fluidität Geringe Fluidität
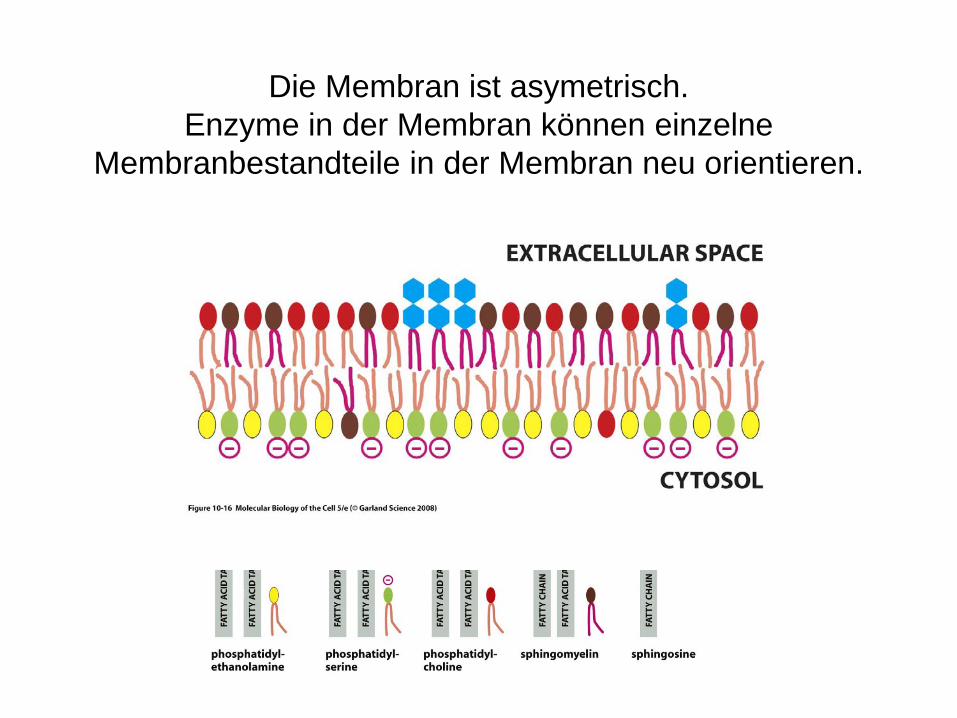
Die Membran ist asymetrisch.Enzyme in der Membran können einzelne
Membranbestandteile in der Membran neu orientieren.
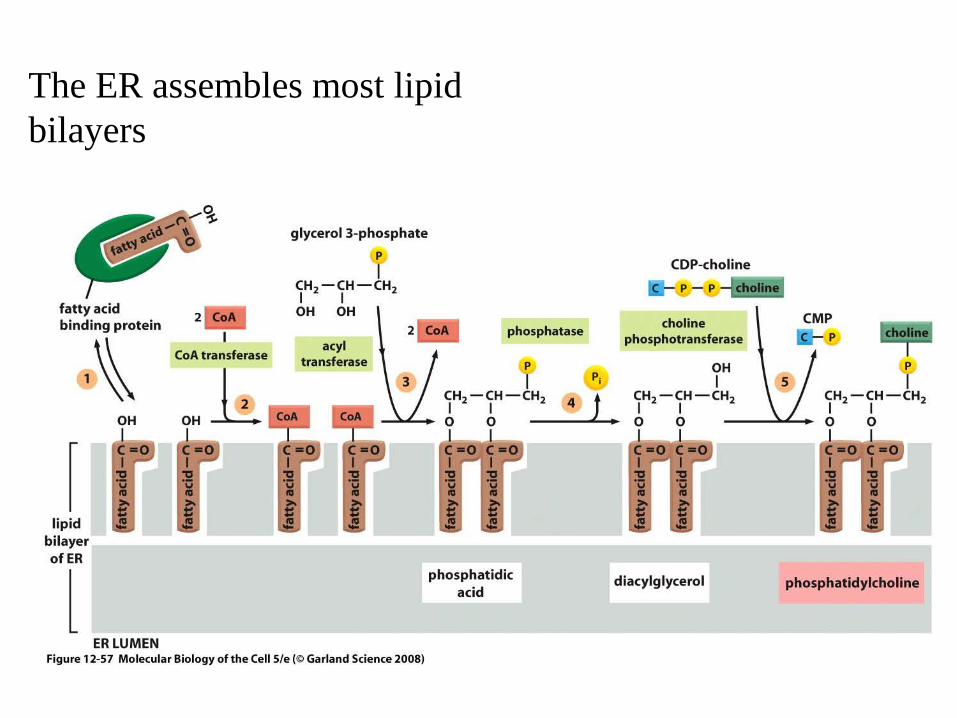
The ER assembles most lipid bilayers
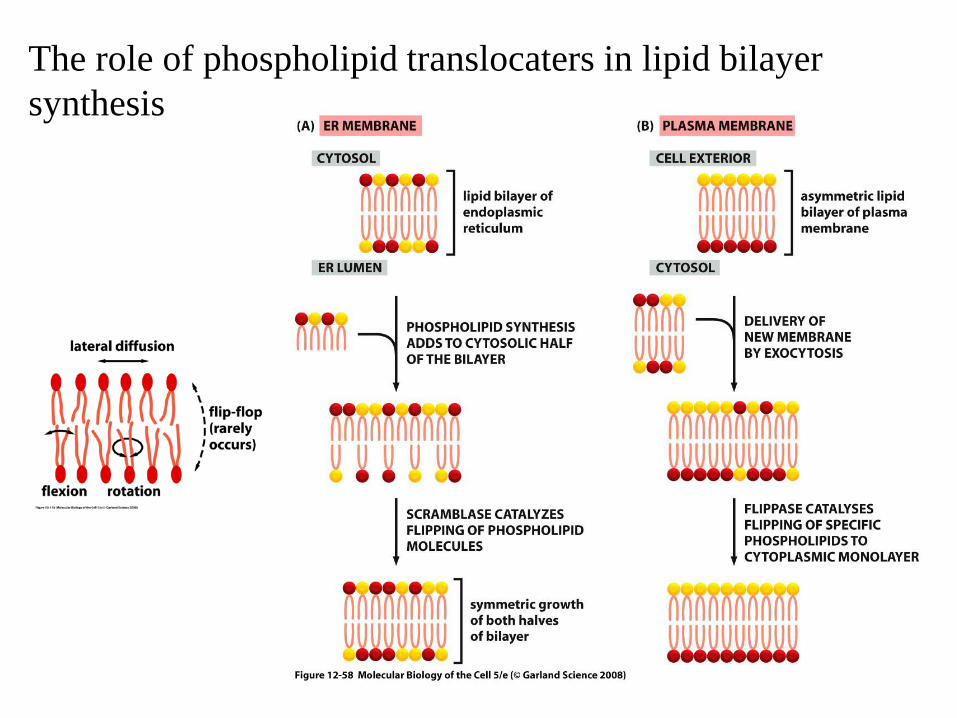
The role of phospholipid translocaters in lipid bilayer synthesis
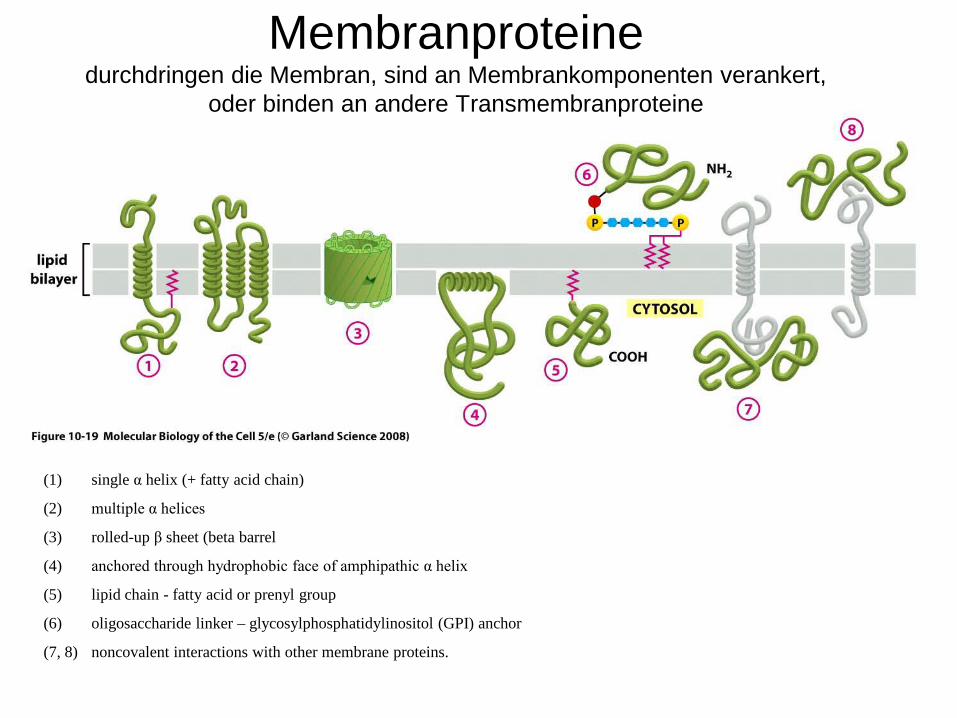
(1) single α helix (+ fatty acid chain)
(2) multiple α helices
(3) rolled-up β sheet (beta barrel
(4) anchored through hydrophobic face of amphipathic α helix
(5) lipid chain - fatty acid or prenyl group
(6) oligosaccharide linker – glycosylphosphatidylinositol (GPI) anchor
(7, 8) noncovalent interactions with other membrane proteins.
Membranproteinedurchdringen die Membran, sind an Membrankomponenten verankert,
oder binden an andere Transmembranproteine
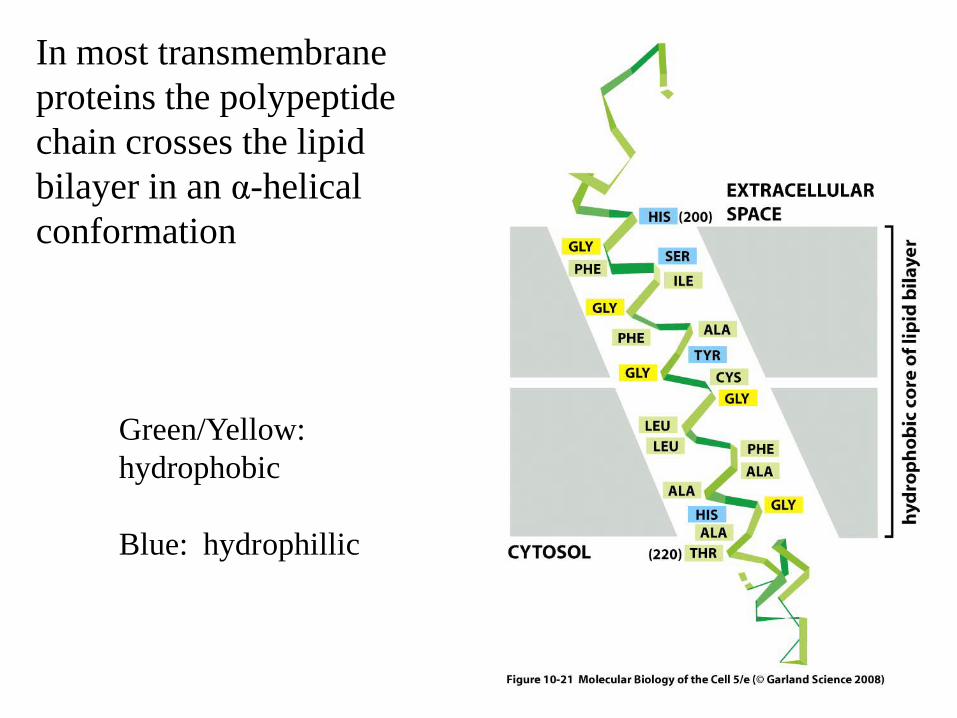
In most transmembrane proteins the polypeptide chain crosses the lipid bilayer in an α-helical conformation
Green/Yellow: hydrophobic
Blue: hydrophillic
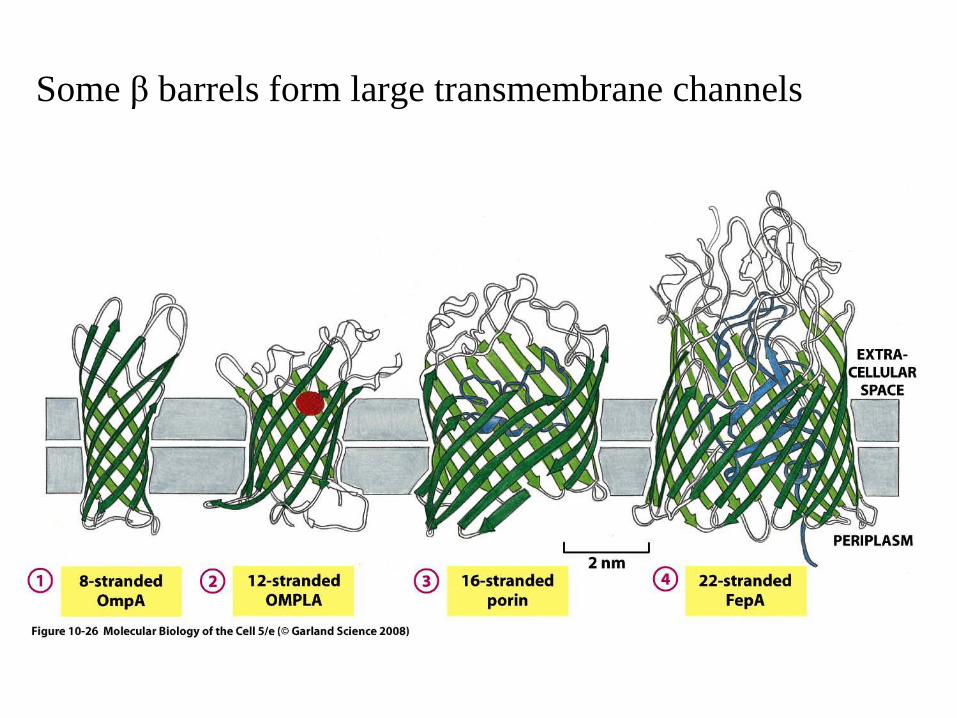
Some β barrels form large transmembrane channels
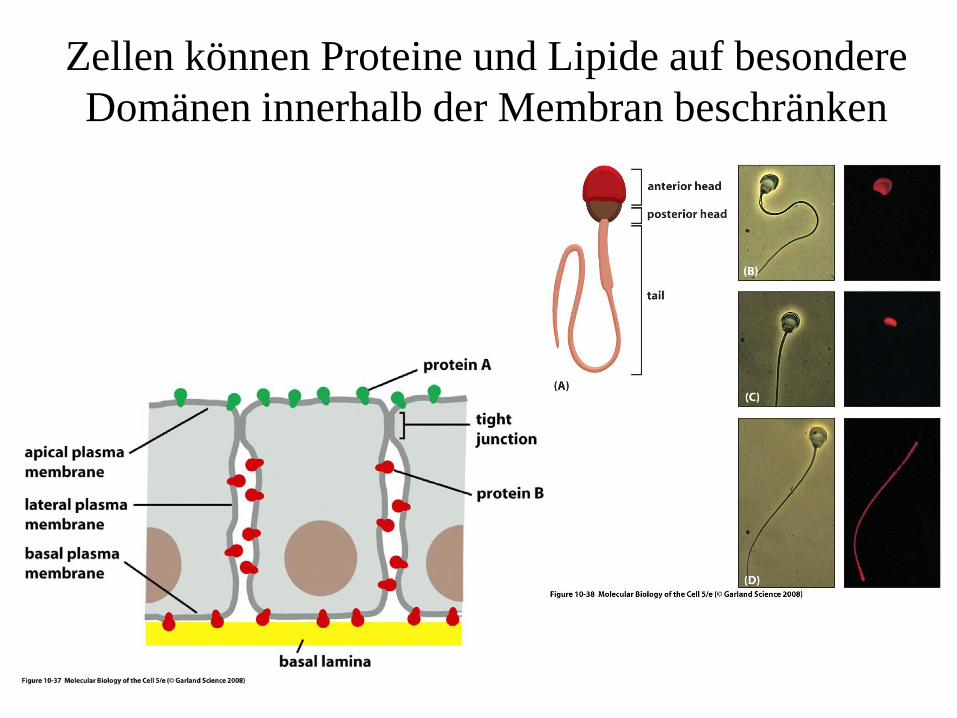
Zellen können Proteine und Lipide auf besondere Domänen innerhalb der Membran beschränken

Ways of restricting the lateral mobility of specific plasma membrane proteins
(A) self-assembly into large aggregates (e.g. Bacteriorhodopsin crystals); interactions with assemblies of macromolecules outside (B) or inside (C) the cell; (D) interaction of proteins on the surface of different cells
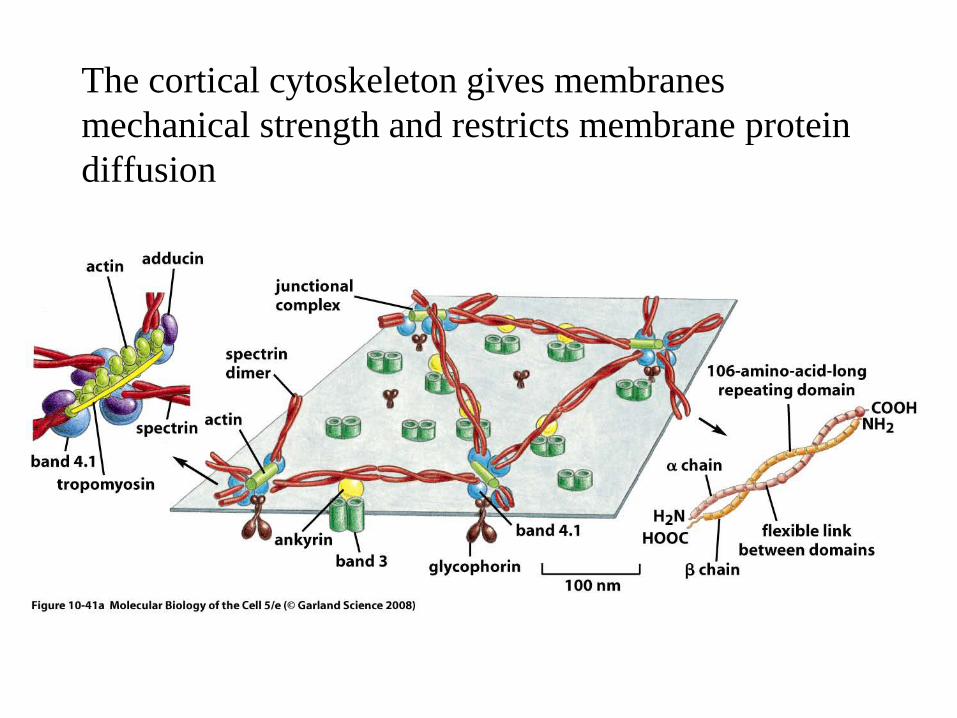
The cortical cytoskeleton gives membranes mechanical strength and restricts membrane protein diffusion
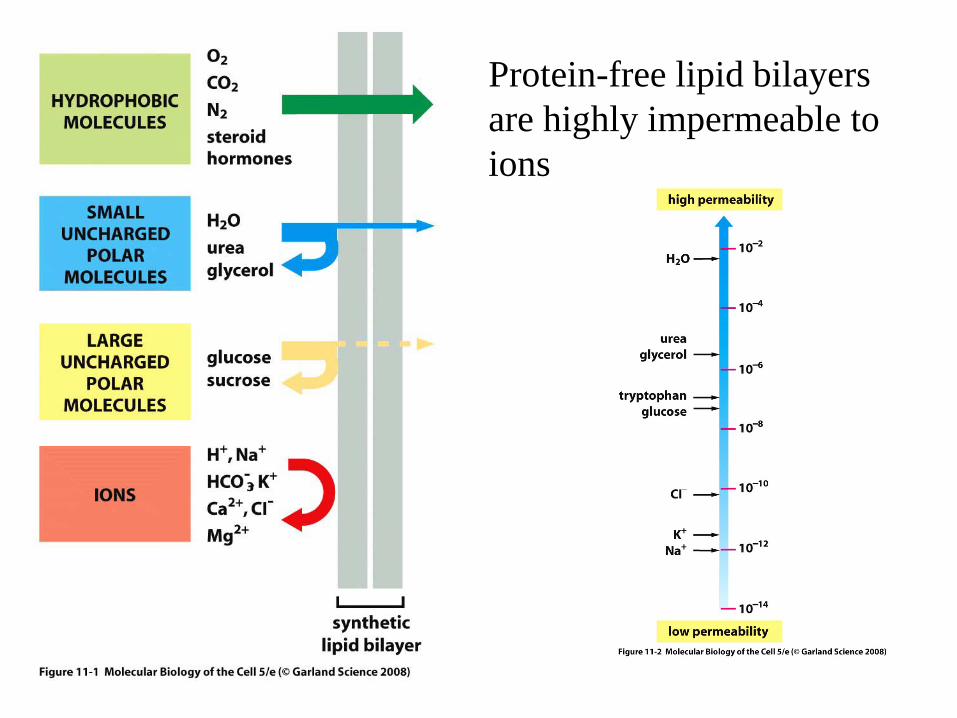
Protein-free lipid bilayers are highly impermeable to ions
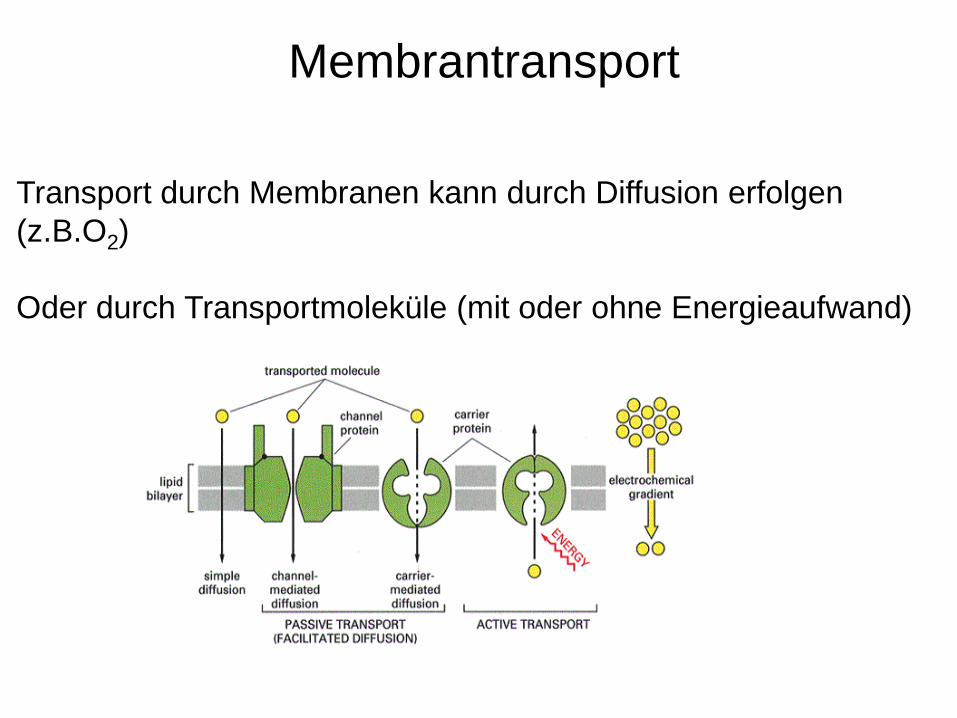
Membrantransport
Transport durch Membranen kann durch Diffusion erfolgen (z.B.O2)
Oder durch Transportmoleküle (mit oder ohne Energieaufwand)
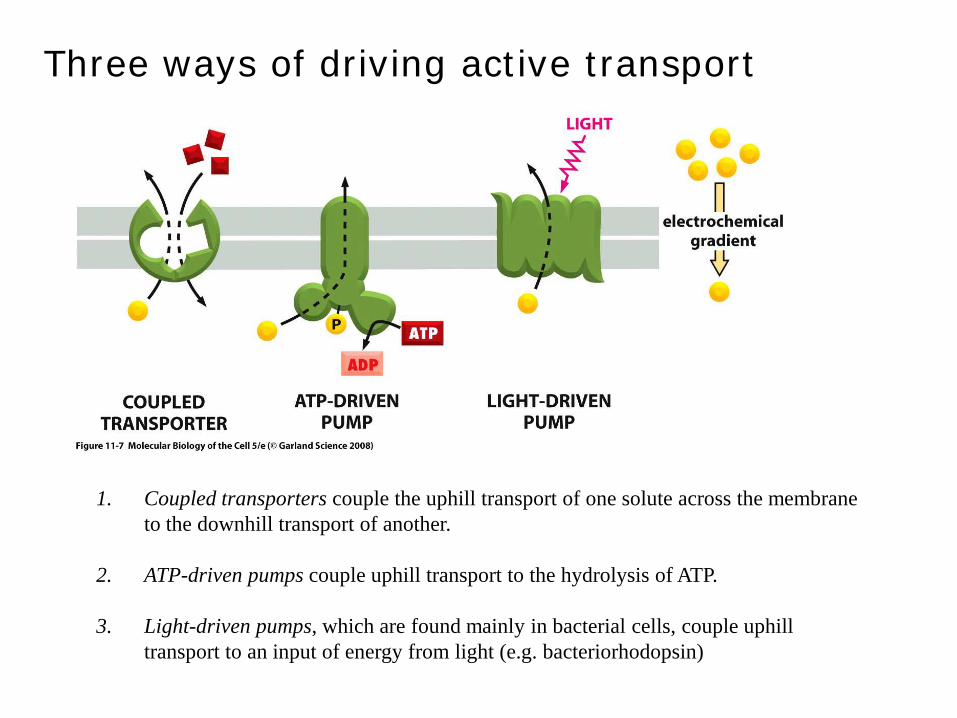
Three ways of driving active transport
1. Coupled transporters couple the uphill transport of one solute across the membrane to the downhill transport of another.
2. ATP-driven pumps couple uphill transport to the hydrolysis of ATP.
3. Light-driven pumps, which are found mainly in bacterial cells, couple uphill transport to an input of energy from light (e.g. bacteriorhodopsin)
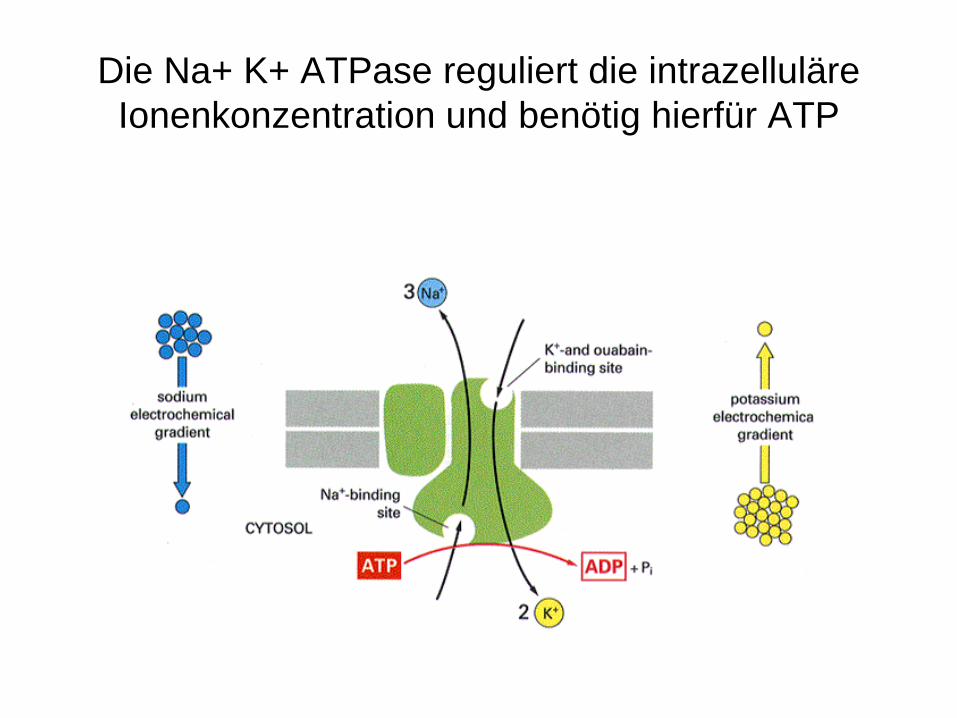
Die Na+ K+ ATPase reguliert die intrazelluläre Ionenkonzentration und benötig hierfür ATP
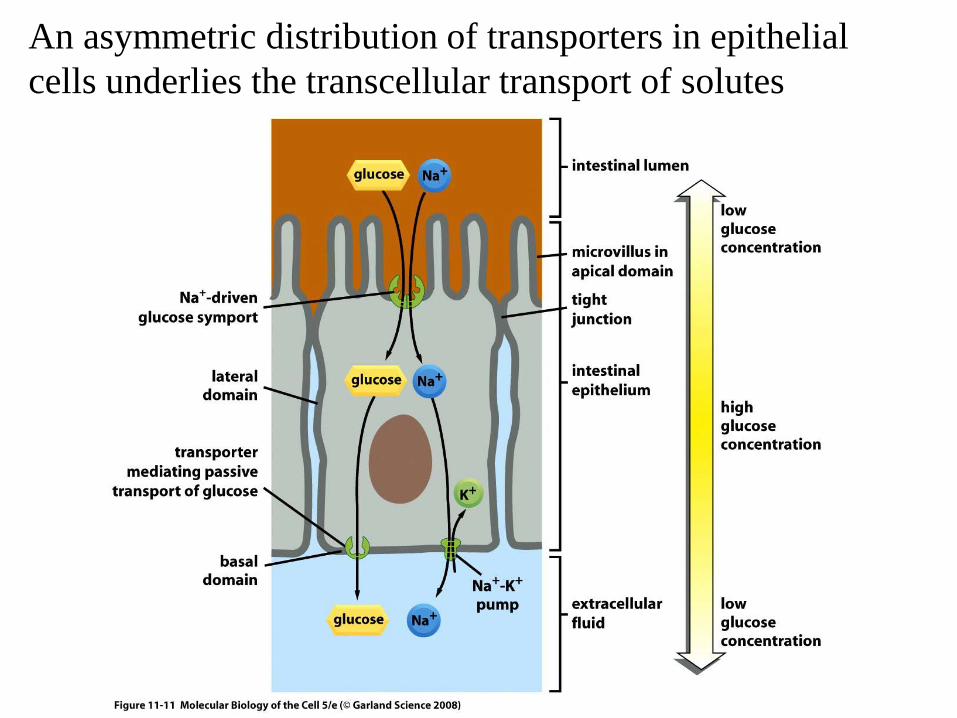
An asymmetric distribution of transporters in epithelial cells underlies the transcellular transport of solutes
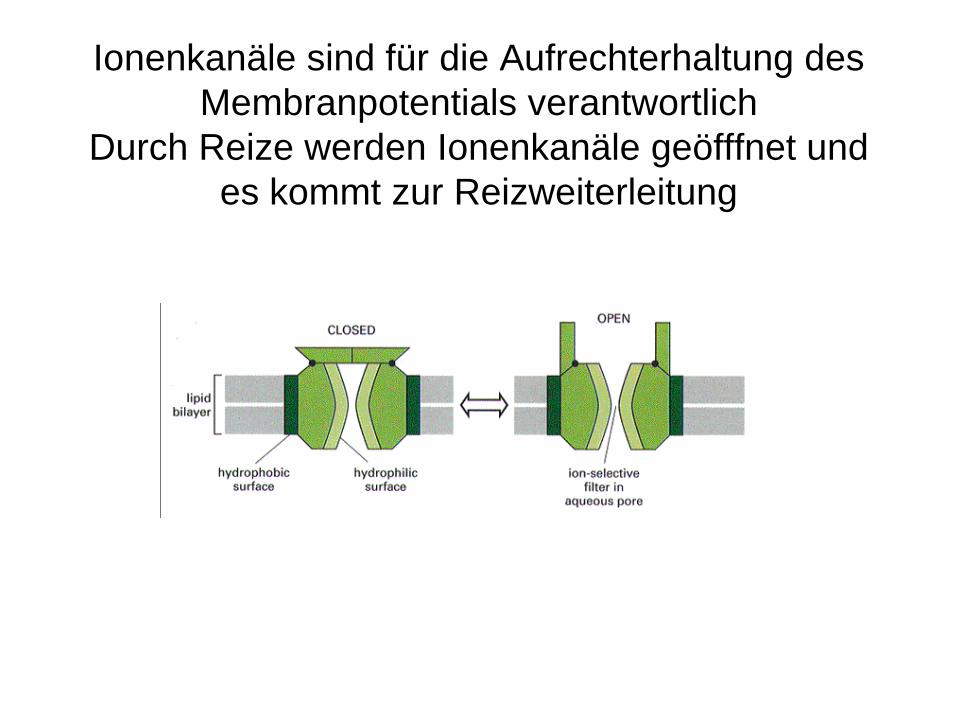
Ionenkanäle sind für die Aufrechterhaltung des Membranpotentials verantwortlich
Durch Reize werden Ionenkanäle geöfffnet und es kommt zur Reizweiterleitung
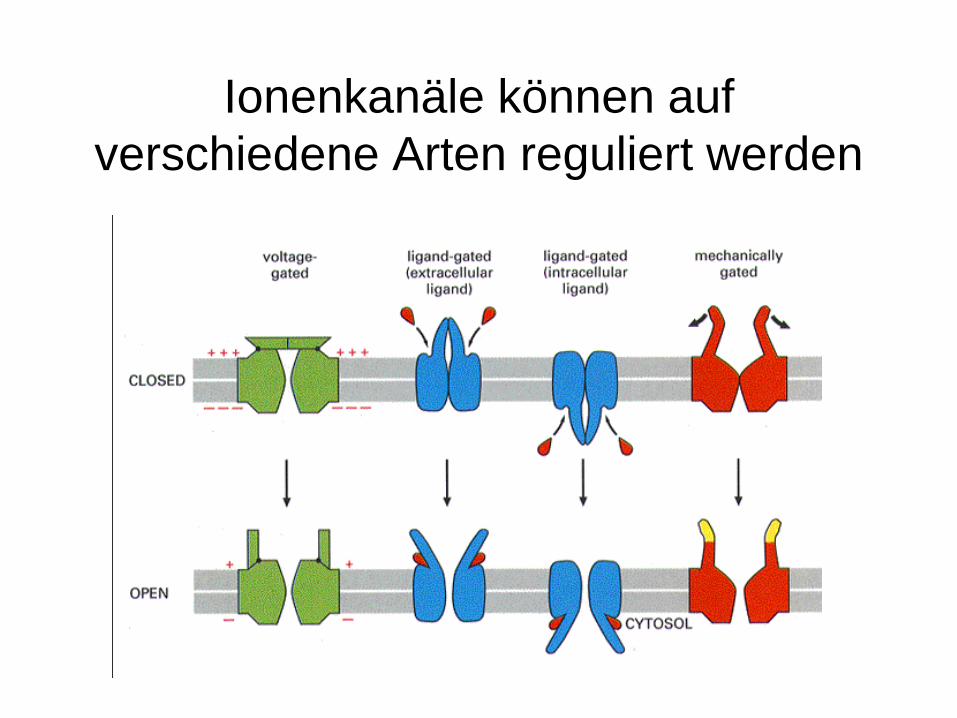
Ionenkanäle können auf verschiedene Arten reguliert werden
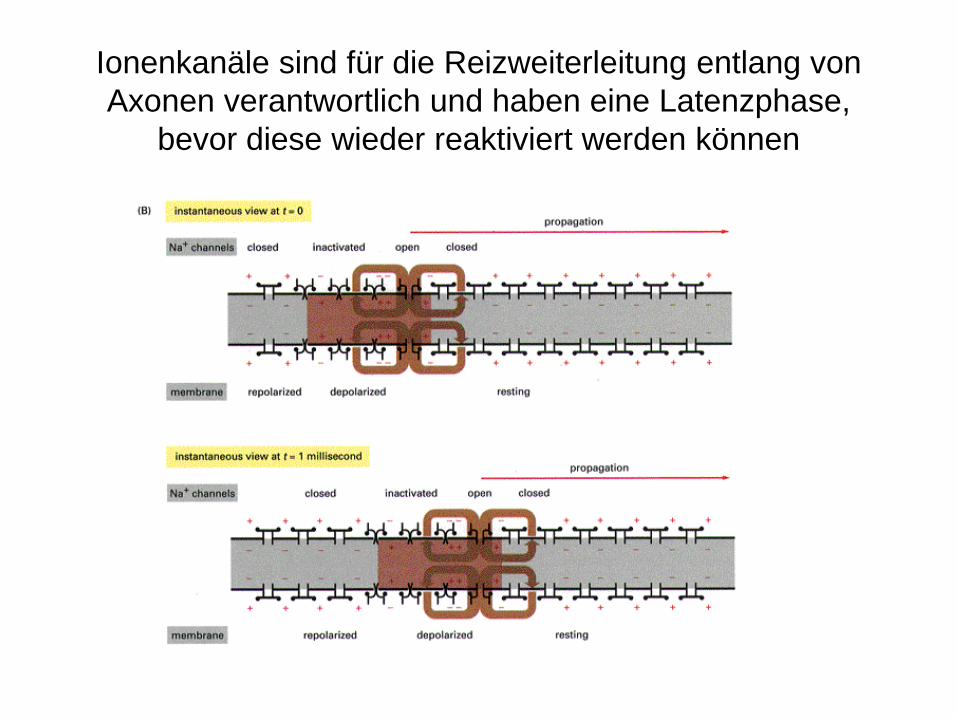
Ionenkanäle sind für die Reizweiterleitung entlang von Axonen verantwortlich und haben eine Latenzphase,
bevor diese wieder reaktiviert werden können